
Intuitive Theories of Information: Beliefs about the Value of Redundancy
Upload
independentCategory
view
3download
0
Reciprocal activation of HSF1 and HSF3 in brain and blood tissues:
Is redundancy developmentally related?
Ariel Shabtay1,2 and Zeev Arad1,*
1Department of Biology, Technion-Israel Institute of Technology,
Haifa 32000, Israel
2Department of Cattle and Genetic Sciences, Institute of Animal Science,
Agricultural Research Organization, Newe-Ya’ar Research Center, P.O.B
1021, Ramat Yishay 30095, Israel.
*Corresponding author: Zeev Arad, Department of Biology, Technion, Haifa 32000,
Israel. Tel: 972-4-8293416; Fax: 972-4-8225153; e-mail: [email protected]
Running head: Reciprocal Activation of Heat Shock Factors
Copyright © 2006 by the American Physiological Society.

2
Abstract
Transcriptional induction of heat shock genes in response to temperature elevation
and other stresses is mediated by heat shock transcription factors (HSFs). Avian
cells express two redundant heat-shock responsive factors, HSF1 and HSF3, which
differ in their activation kinetics and threshold induction temperature. Unlike the
ubiquitous activation of HSF1, the DNA-binding activity of HSF3 is restricted to
undifferentiated avian cells and embryonic tissues. Herein we report a reciprocal
activation of HSF1 and HSF3 in vivo. Whereas HSF1 mediates transcriptional
activity only in the brain upon severe heat shock, HSF3 is exclusively activated in
blood cells upon light, moderate and severe heat shock, promoting induction of heat
shock genes. Although not activated, HSF1 is expressed in blood cells nuclei in a
granular appearance, suggesting regulation of genes other than heat shock genes.
Intraspecific comparison of heat sensitive and heat resistant fowl strains indicates
that the unique activation pattern of HSF3 in blood tissue is a general phenomenon,
non-related to thermal history. Taken together, HSF1 and HSF3 mediate
transcriptional activity of adult tissues and differentiated cells in a non-redundant
manner. Instead, an exclusive, tissue-specific activation is observed, implying that
redundancy may be developmentally related. The physiological and developmental
implications are discussed.
Keywords: HSF, reciprocal activation, redundancy

3
Introduction
The heat shock response is characterized by induced synthesis of heat shock proteins
(HSPs), the accumulation of which confers cellular thermotolerance (15). The expression
of HSPs is primarily regulated at the level of transcription by a family of heat shock
transcription factors (37). To date, four members of the HSF gene family have been
isolated and characterized in vertebrates (42, 44, 46, 47, 51). The existence of multiple
family members suggests that different HSFs mediate the responses to various forms of
physiological and environmental stimuli (45). Accordingly, HSF1 acts as a classical
stress-responsive factor, inducing transcriptional activity of heat shock genes in response
to diverse forms of stress. HSF2 is not activated in response to classical stress stimuli, but
is rather active under developmentally related conditions (14, 54-55) and mediates
bookmarking of inducible HSPs (61). The unique avian-specific factor HSF3 (19), like
HSF1, is a heat responsive transcription factor but is considered as a high temperature
activator (43, 56). The mammalian HSF, HSF4, is restricted to certain tissues, and has the
properties of a negative regulator of heat shock gene expression (44, 58). Among the
three HSF genes that are expressed in avian cells, HSF1 and HSF3 are considered
redundant heat shock responsive factors (42-43). Consistent with this, HSF3 exhibits a
delayed response compared to the relatively rapid activation of HSF1, and is detected
predominantly upon severe heat shock (43, 56).
The activation of HSF1, the predominant stress-responsive factor, involves multiple
steps, including translocation to the nuclear compartment, oligomerization from the latent
monomer to a trimer, acquisition of DNA-binding activity, inducible serine
phosphorylation, and subsequent transcriptional activity (9, 11, 18, 27, 48, 59). Non-

4
active HSF3 is a dimer which is exclusively cytosolic (43). Upon heat stress, HSF3 is
translocated into the nucleus, using the nuclear localization signal to be converted to an
active trimer (40, 43).
In contrast to the ubiquitous activation of HSF1 in vertebrates (48), acquisition of HSF3
DNA-binding activity in response to heat shock appears to be a cell-specific event, and
active HSF3 has been detected only in avian undifferentiated cells and embryonic tissues
(28, 43, 56). Most interestingly, disruption of HSF3 in HSF1-expressing chicken
B-lymphoblast cells results in impaired heat shock response and in loss of
thermotolerance (57). In light of the above reported findings we sought to gain insight
into the relative contribution of HSF1 and HSF3 to the transcriptional induction of heat
shock genes in mature tissues and differentiated cells, in vivo. For this purpose we chose
brain and blood of mature hen, two unique tissues that are differentially confronted with
thermal challenges. Our results show that in each tissue, only one transcription factor is
activated upon heat stress. Whereas HSF1 mediates transcriptional activity in the brain,
HSF3 is exclusively activated in blood cells upon moderate and severe heat shock to
induce the expression of heat shock genes.
Materials and Methods
Real time in vivo measurements of the heat shock response
Two different groups of mature chickens, Leghorn and Bedouin (n=3 in each group), that
differ genetically in their resistance to heat were examined in this setup. The setup was
recently described in details (52). Briefly, Twenty hours prior to experiment, a
polyethylene cannula was implanted in the brachial vein of mature fowl under local

5
anaesthesia, and a 5cm-long, custom-made polyethylene cannula was implanted dorsal to
the rectum and fastened to the skin. Experiments were carried out within a temperature-
controlled room (±0.3oC; 12L:12D light cycle, lights on at 6 am). The birds had free
access to food and water and could freely move in their individual cages. Each
experiment started at 8-9 am at an ambient temperature (Ta) of 24oC (RH=50±5%) which
is within the thermoneutral zone of chickens (5). A copper-constantan thermocouple was
introduced into the rectal cannula, locked at a pre-determined depth of 5cm and
connected to a digital thermometer (±0.1oC). All temperature probes were calibrated
against a mercury-in-glass thermometer having accuracy (0.1 oC) traceable to the U.S.
Bureau of Standards. After approximately 10min, body temperature (Tb) stabilized
around 41oC (the normal resting Tb for this species), after which blood was remotely
sampled through extended tubing. Ta was then elevated to 38oC (RH=50±5%). Body
temperature was followed continuously and blood samples were taken at each 1oC
increase in Tb up to 45oC. At this time, ambient temperature was lowered back to 24oC
and blood samples taken at each 1oC decrease in Tb down to 41oC. These samples were
analyzed for HSF DNA-binding activity, transcriptional and translational activity and
HSF translocation. After each blood sample the cannula was flushed with heparinized
saline.
Protein extraction
For the purpose of protein extraction, brain tissues were sampled from Tb=45oC heat
shocked mature hens. The dissected tissues were homogenized in ice cold buffer,
containing 0.1M NaCl, 20mM Tris pH 7.4, 0.2mM EDTA, 20% glycerol (v/v), 0.5mM

6
dithiothreitol (DTT), 15µg/ml leupeptin, 1mM phenylmethylsulfonylfluoride (PMSF).
Samples were centrifuged for 30 min (4oC, 12000 rpm) and supernatants were collected,
frozen in liquid nitrogen and stored at -70oC. To extract proteins from blood cells, blood
samples were washed once with ice cold PBS and centrifuged for 2 min (4oC, 1500 rpm).
The cells were re-suspended in ice cold TMP buffer (containing 10mM Tris pH 7.4, 1mM
EDTA, 5mM MgCl2, 0.5mM DTT, 15µg/ml leupeptin, 1mM PMSF), and were frozen
(liquid nitrogen) and thawed (37oC) in 4 cycles. Samples were centrifuged for 30 min
(4oC, 12000 rpm) and supernatants were collected, frozen in liquid nitrogen and stored at
-70oC. Blood cell nuclear proteins were extracted as previously described by Dyer and
Herzog (13).
SDS PAGE and Western Blot
Blood cell nuclear proteins lysates were boiled in sample application buffer containing
2-mercaptoethanol. Equal amounts of proteins (in each lane) were loaded and separated
by SDS-polyacrylamide gel (10%), and transferred onto nitrocellulose membrane
(Schleicher&Schuell). The membranes were probed with polyclonal anti HSF1β (a
generous gift from Professor Akira Nakai), followed by appropriate secondary antibodies.
The proteins were visualized by enhanced chemiluminescence. Quantification of HSF1
expression was carried out by a densitometry software (TINA).

7
Electromobility shift assay (EMSA)
Electromobility shift assay was performed as previously described (38). Briefly, equal
amounts of cellular proteins (20µg for brain extraction and 5µg for blood nuclei proteins)
were incubated with a 32P-labeled double-stranded oligonucleotide
(5’- CTAGAAGCTTCTAGAAGCTTCTAG-3’). The protein-bound and free
oligonucleotides were electrophoretically separated by 4% native polyacrylamide gels.
The gels were dried and auto-radiographed.
Supershift experiments
The antibody supershift experiments were performed by incubating 2µl of 1:10 diluted
antisera (unless otherwise stated in the figure legend) of anti HSF1γ and anti HSF3 γ (a
kind gift from Professor Akira Nakai) in PBS, with 1–2 µl of cell lysates in a total
volume of 10 µl. These antibodies recognize the carboxyl-terminal region of each HSF.
After incubation on ice for 20 min, a nucleotide probe mixture containing 0.1ng of 32P-
labeled self-complementary ideal HSE oligonucleotides and 0.5µg of poly(dI-dC)
(Pharmacia Fine Chemicals, Piscataway, NJ) in 10mM Tris-HCl (pH 7.8) and 1 mM
EDTA was added to a final volume of 25µl. The binding reactions were incubated on ice
for 20 min, mixed with 2µl of a dye solution (0.2% bromphenol blue, 0.2% xylene
cyanol, and 50% glycerol) and then separated on 4% native polyacrylamide gels. Gels
were run in 0.5xTBE buffer at room temperature for 1.7 h at 140 V, dried, and exposed to
x-ray film at -80°C with intensifying screens.

8
RNA isolation and Northern blot
Total RNA was isolated from blood cells by TRI REAGENT-BD (MRC) according to
the manufacturer's instructions. RNA (5µg) was separated in formaldehyde-agarose gel
and transferred onto nylon membrane (Zeta-Probe, Bio-Rad). Chicken HSP90α cDNA
(a kind gift from Professor Akira Nakai) was labeled by the extension priming method
using [α-32P]dATP. Membrane was hybridized for 16 hours at 55oC, washed with
0.1%SDS in 1 X SSC at 45oC, 50oC and 55oC, and exposed to x-ray film (Kodak BioMax
MS) at –70oC in the presence of intensifying screen.
Immunocytochemistry
Blood samples from control and Tb=45oC heat shocked mature hens were prepared as
smears. The smears were air dried and fixed with 4% paraformaldehyde (in PBS). After
blocking with 10% normal goat serum in PBS, cells were incubated with 1:500 diluted
polyclonal αHSF1 (a kind gift from Professor Akira Nakai). The HSF1 signal was
detected by a secondary goat anti-rabbit antibody conjugated to FITC. Confocal
reconstructions were made by using an MRC-1024, laser confocal scanning microscope
(Bio-Rad, Hempstead, UK) with the objective Nikon Plan Apo 603/1.40. DNA was
counterstained with 250ng/ml propidium iodide (PI).

9
Results
HSF3 is exclusively activated in chicken blood tissue
We have recently described the in vivo, real-time heat shock response of fowl from
strains that differ in their resistance to heat. Our data revealed a differential DNA-binding
kinetics of total HSF in each of the groups that was followed by transcriptional and
translational induction of heat shock genes (52). To elucidate the redundancy in the
activation of both, HSF1 and HSF3, in response to elevated temperature, blood of mature
hen was sampled, at 1oC increments of Tb. We determined the relative levels of HSF1 and
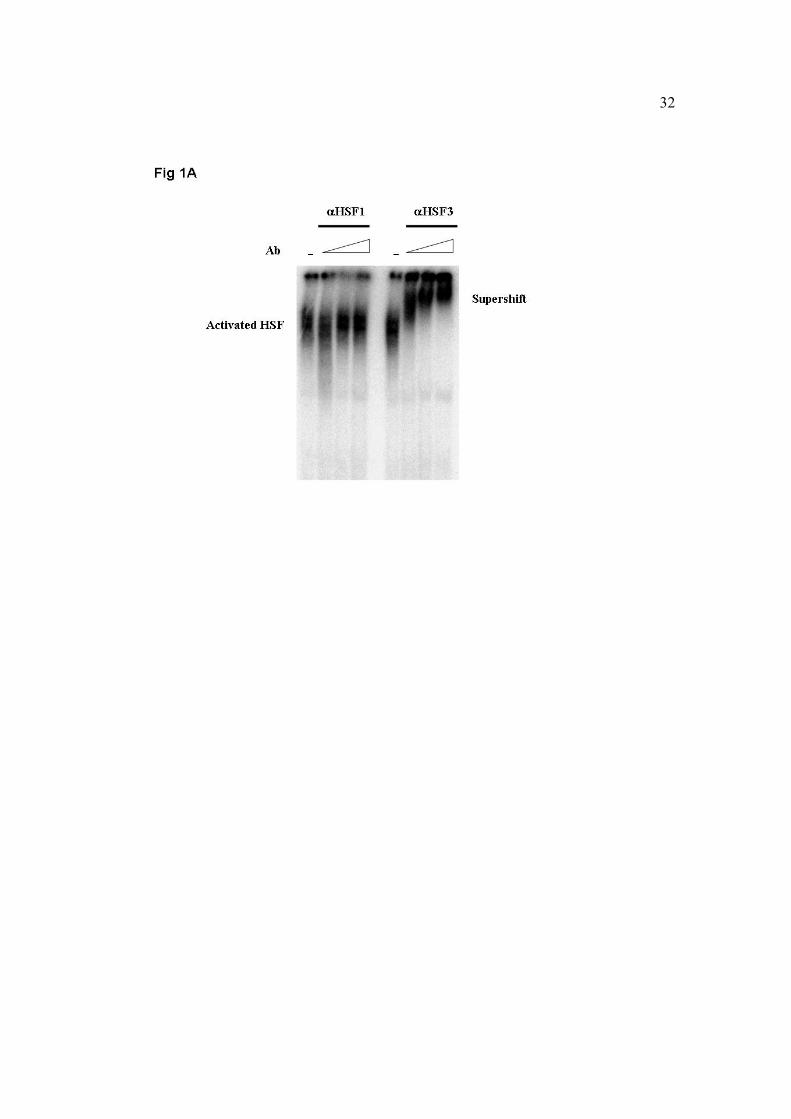
HSF3 DNA-binding activity by supershift experiments. At Tb = 45oC, HSF3-bound DNA
was entirely supershifted by each antisera concentration, while the DNA-binding activity
of HSF1 was undetected (Fig. 1A). It has been previously reported that HSF3 is the
dominant heat shock transcription mediator during severe heat shock conditions. It was
thus reasonable to assume that the activation of HSF1 may be abolished at Tb = 45oC, but
is likely to occur at lower body temperatures. To check the activation pattern of both
HSFs at moderate heat shock conditions, we performed supershift experiments of blood
sampled at Tb = 43oC, a well-documented temperature for the activation of the ubiquitous
factor HSF1. In this experiment we used the same HSF1 antisera concentrations, but 10-
fold more diluted concentrations of HSF3 antisera, to allow the examination of the anti-
HSF3 antibody specificity. As seen in figure 1B, HSF3 was the exclusive DNA-bound
heat shock transcription factor, also upon moderate heat shock conditions.

10
HSF1 and HSF3 DNA-binding states are not affected by intraspecific variations in
thermal resistance
Our previous comparisons of the Leghorn chicken with the desert-origin, genetically heat
resistant Bedouin fowl identified various thermoregulatory mechanisms that contribute to
the superiority of the Bedouin fowl with respect to heat resistance (2, 4, 32). This
superiority was accompanied by a considerable delayed HSP response, both in relation to
time scale and to Tb level (52). In light of the above-mentioned observations it was
intriguing to examine the DNA-binding state of both HSFs in heat sensitive and heat
resistant strains of the same species. Consistent with the findings shown in Figure 1,
HSF3 was solely in a DNA-binding form in both fowl strains, and along the entire range
of heat shock conditions (Tb = 42oC to Tb = 45oC), without any redundant acquisition of
HSF1 DNA-binding activity in response to heat shock (Fig. 2).
The HSF3 DNA-binding activity in both the Leghorn and the Bedouin strains is slightly
detected in control, non-stress conditions. However, the intraspecific differences in the
cellular heat shock response are manifested at heat shock conditions, with Leghorn
showing a profound HSF3 activation at Tb = 42oC, in comparison with Tb = 44oC of the
Bedouin strain (Fig. 2).
HSF1 is expressed in blood cells and forms nuclear granules, but is not activated
Following the surprising findings that HSF1 may not participate in mediating in vivo the
cellular heat shock response of avian blood tissue, we next examined the expression of
HSF1 in chicken blood cells. We extracted nuclear proteins from blood samples of hens
that were exposed to heat shock and allowed to recover, and analyzed the expression of

11
HSF1 by Western blot analysis, using polyclonal anti HSF1 antibody. HSF1 was revealed
in the nuclear fraction of the blood cells in control, during heat shock and during recovery
from heat shock. However, the amount of expressed HSF1 was not uniform in all
samples; its concentration tended to decrease around the peak of heat shock (Tb = 45oC;
Tb = 44oC, at recovery. These values were 2 and 3.3 fold lower, respectively, than the
control values) and tended to increase during recovery from heat shock while Tb was
decreasing (Fig. 3A). Despite its expression in blood cells nuclei in non-stress conditions
(Fig. 3A), immunohistochemical preparates from control and from heat shocked cells
(Tb = 44oC) indicate that HSF1 tranlocates from a cytosolic distribution in control to a
nuclear granular distribution upon heat shock (Fig. 3B).
Activated HSF1 exclusively mediated transcriptional activity of HSPs in chicken
brain tissue
Is the exclusive HSF3 DNA-binding pattern unique to the blood tissue, or may it reflect a
general in vivo phenomenon in adult fowl? To address this issue we conducted supershift
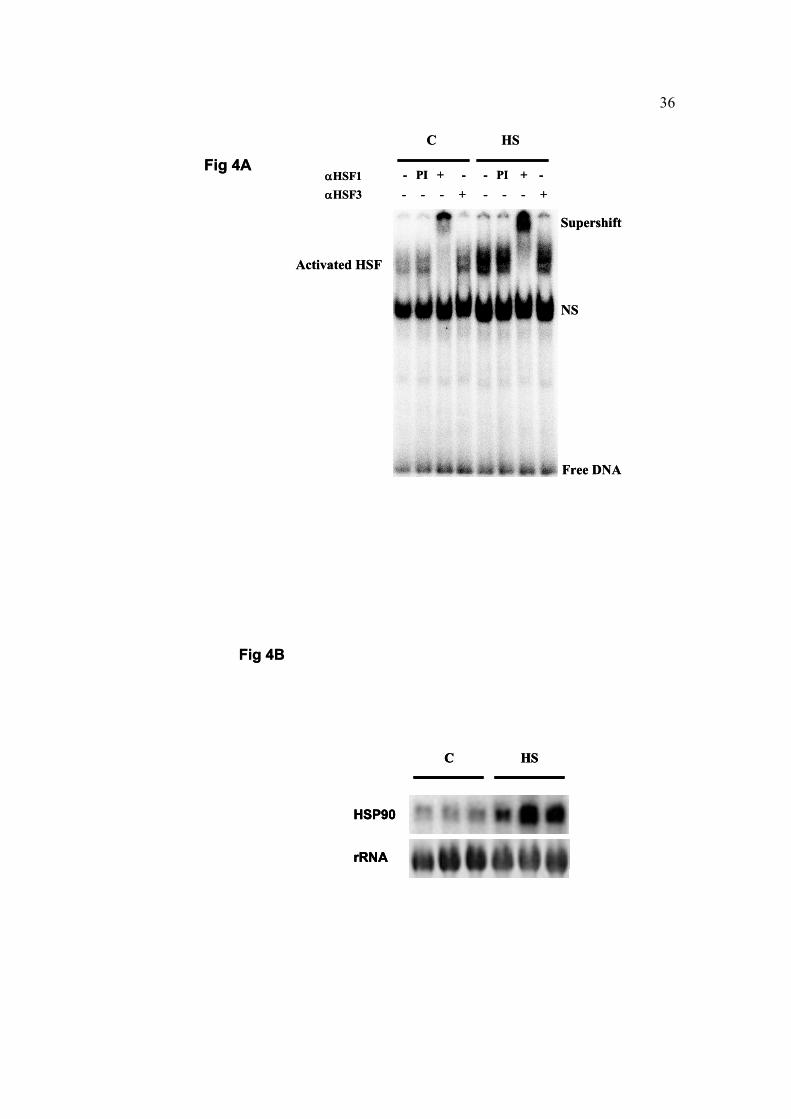
experiments in brain extracts from heat shocked (Tb = 45oC) mature hen, using polyclonal
anti HSF1 and anti HSF3 antibodies. A reciprocal mode of HSF activation to that of the
blood tissue was revealed in the brain, indicating that HSF1 was exclusively in a DNA-
binding state (Fig. 4A).
To elucidate whether the DNA-binding state of HSF1 in the brain induces the activation
of heat shock genes, we performed a Northern blot analysis with the entire chicken
HSP90α cDNA as a probe. A burst induction of HSP90 was observed in response to

12
Tb = 45oC heat shock, implying that HSF1 is capable of mediating the transcriptional heat
shock response in the brain in a non-HSF3 redundant manner (Fig. 4B).
Discussion
Stress-induced transcription of heat shock genes in avian cells is mediated by HSF1 and
HSF3 (39). The co-expression and co-activation of these factors has led to the discovery
of the HSF functional redundancy (37). Indeed, when undifferentiated avian cells and
avian embryonic tissues are subjected to thermal challenges, HSF1 and HSF3 are
activated with different kinetic properties and threshold temperatures, to induce
transcription of heat shock genes (28, 43, 56). Surprisingly, as demonstrated in the
present study, circulating blood cells lose their functional redundancy and activate only
HSF3, also upon light and moderate heat shocks. These results are of developmental and
physiological significance, since they imply that during maturation of erythroblasts,
HSF1 activity is degenerated. As maturating erythroblasts go through morphological,
physiological and biochemical changes (26), loss of redundancy due to diminished
expression of HSF1 may be a reasonable explanation. Supported by the nuclear
expression pattern of HSF1 in control and heat shocked blood cells, it is more likely that
this degenerative event has occurred along the pathway that regulates DNA binding
activity. Transcriptional induction of heat shock genes may be a multiple step process.
Under normal conditions of cell growth, the non-DNA binding state of HSF1 is regulated
by two distinct mechanisms, involving both negative regulatory domains and constitutive
phosphorylation at serine residues (16, 29-30, 53). To induce transcriptional activity of
heat shock genes, HSF1 must acquire DNA binding activity, preceded by oligomerization
and nuclear localization (48). However, the trimer state of HSF1 does not assure binding

13
interactions with the DNA, as interactions of the HSF1 oligomerization domain with
regulatory proteins may negatively affect HSF1 DNA-binding activity (50). Consistent
with the findings that DNA binding and transcriptional activities are uncoupled processes
(27, 31) HSF1 must be inducibly phosphorylated to promote transcription of heat shock
genes (11, 18).
In our current study, wing blood cells immunohistochemistry, show that apart from its
nuclear expression in control and heat shock conditions, HSF1 translocates from a
cytosolic distribution at control conditions to a nuclear granular distribution at Tb = 44oC.
Nuclear stress granules have been described as intra-nuclear sites at which HSF1 is
concentrating in response to stress conditions (48). The kinetics of appearance of HSF1
granules parallels the activation of HSF1 and the transient transcriptional activity of heat
shock genes (12). It has been suggested that these nuclear foci are locations at which
HSF1 is stored and regulated in its different activation states (22), thereby coordinating
the regulation of heat shock gene expression (23). As HSF1 stress granules are also
present in heat shocked mitotic cells that are devoid of transcription, it is alternatively
hypothesized that they are involved in chromatin organization and thus have a structural
role in protecting hypersensitive or fragile sites of the genome (23-25). The expression
levels of HSF1 in the present study tend to decrease as Tb reaches its experimental peak
and return to control levels upon recovery. This observation is in accordance with a
previous study (56), which reported, in vitro, the movement of HSF1 to the insoluble
fraction at high ambient temperature.
Nearly four decades ago the unique thermoregulatory capacities of the desert-origin
Bedouin fowl were described (1-2, 4-8, 32-33). Recently, we have monitored in real-time

14
in vivo the heat shock response of this heat resistant fowl strain, and showed that its heat
resistance superiority was characterized by a considerable delayed activation of HSF,
compared with a heat sensitive, commercial strain (52). In the present study we
demonstrate that HSF3 is solely in a DNA-binding form in both fowl strains and along
the entire range of heat shock conditions, and that no redundant activity of HSF1 is taking
place. This intraspecific comparison of two evolutionary distinct strains, with respect to
thermoregulatory strategies, suggests that the unique activation pattern of HSF3 in the
blood tissue is a general phenomenon, non-related to thermal history. Together with the
above mentioned findings, these results imply that loss of HSF1 DNA-binding activity is
not caused by decreased levels of HSF1 message, nor because HSF1 had become
insoluble.
In chicken embryonic tissues, including red blood cells and brain tissue, HSF3 was the
dominant heat shock responsive factor in certain developmental stages, whereas in other
stages HSF1 and HSF3 showed functional redundancy (28). As opposed to the blood, in
which paralleled activation of HSF3 in embryos (28) and adult chicken (present study) is
sustained, the present findings in the brain tissue revealed an unexpected loss of HSF3
activation and the introduction of HSF1 as an exclusive heat shock responsive factor.
Chicken HSF3 remains as an inactive dimmer, exclusively in the cytoplasm of normally
growing cells and is translocated into the nucleus and converted to an active trimer upon
heat stress. Its DNA-binding properties, however, are regulated in a cell-specific manner
(43). As HSF3 activation in embryonic brain tissue is evident (28), and as HSF3 is
expressed in adult chicken brain tissue (present study, data not shown), we believe that
the unique regulation of avian brain temperature could provide a reasonable physiological

15
explanation for the non DNA-binding state of HSF3 in this tissue. Equipped with a
unique heat-exchange vasculature, the ophthalmic rete (35), the avian brain temperature
is maintained around 1oC below body temperature in a wide range of ambient
temperatures (see review by Arad; 3) and hydration conditions (8, 21, 36). In the context
of the present study this means that brain temperature probably did not exceed 44oC
during heat shock. Thus, although HSF3 has been previously shown to be activated at
44oC (56), we cannot exclude that the brain was not confronted with temperatures above
the threshold for HSF3 activation. It may alternatively be claimed that as the brain is
protected from reaching such high temperatures, it has lost part of its temperature induced
DNA-binding activity to promote transcription of heat shock genes, during ontogeny.
While chicken body temperature that exceeds 45oC is considered harmful and non-
reversible, this issue must be solved by a method of acclimation that would allow us
reach a body temperature of 46oC.
In mammals, where HSF1 is believed to be the only factor that mediates heat shock
signals (59), its disruption may not affect adulthood attainment, but may result in
multiple defective phenotypes, including “maternal effect” mutation (10, 60). Moreover,
mammalian HSF1-deficient cells are incapable of stress-induced transcription of heat
shock genes, and of acquirement of thermotolerance (34). Compared to the mammalian
HSF1, avian HSF1 has a little potency of activating heat shock genes in cells subjected to
heat stress (19). It is becoming clearer, however, that similar to the mammalian HSF1,
avian HSF1 also possesses thermal-protective traits, independent of heat shock genes
induction (19-20, 41). Also similar to HSF1 in mammalian cells, HSF3 is essential for the
heat shock response in avian cells, and its disruption decreases markedly the induction of

16
heat shock genes (57). This is further emphasized during chicken embryonic
development, where HSF3 that has nearly identical levels among most examined tissues
is the major heat shock responsive factor (28). These data are in accordance with the
“blood picture” but fail to explain the “brain picture” in the present study. The
physiological nature of both tissues in respect to thermoregulation may shed light on the
reciprocal activation pattern of both HSFs. In marked difference from the brain that
"escapes" the actual body temperature (review by Arad, 3), the blood tissue, among its
other roles, is responsible for heat transfer from the body core to the surface, and as such
is routinely confronted with temperature changes, and should have the ability to respond
to a broad range of temperatures. This function is shown here to be entirely fulfilled by
HSF3. In addition to the biochemical characteristics of HSF1 and HSF3 in regard to the
heat shock response (56), and as HSF3 lacks (so far, as we know) the characteristics of
non-stress responsive genes regulator, these physiological challenges make HSF3 a
reasonable candidate for taking over the transcriptional mediation of heat shock genes.
Indeed, HSF3 might be sufficient to activate transcription in chicken blood tissue, since
red blood cells of chicken embryos reveal a heat shock response in the absence of HSF1
(28). As to the functional redundancy observed in non-differentiated cells such as
erythroblasts, these cells are progenitors to circulating cells and are considered as the
bank of the differentiated cells. It is thus logical to have them protected from
physiological and environmental damage by a redundant system. In the brain, loss of
HSFs’ functional redundancy during ontogeny may be explained by the different thermal
states of the embryo and the mature chicken. The embryo, which is considered
ectothermic, undergoes a transition from ectothermy to endothermy, which is completed,

17
at the earliest, by hatching (17). While ectotherms are prone to changes in body
temperature in correlation with ambient temperature variations, the internal physiological
milieu of endotherms remains relatively stable despite acute external thermal
fluctuations. We have recently shown that embryos are the most heat sensitive stage in
the chicken's life cycle, and are tightly dependent on HSP synthesis for survival. On the
other hand, mature hen master additional mechanisms for heat dissipation and Tb
regulation during ontogeny (52). In light of the above, we suggest that functional
redundancy of HSFs in the brain is required in the thermal state that is most susceptible to
heat stress (i.e. embryos), but may be lost as the individual is capable of maintaining
homeostasis (i.e. adult).
It has been previously shown that the temperature set-point for HSF activation varies
among tissues of a single organism (49). Herein we present data that may confirm also
the exsitance of a tissue-specific activated HSF.
Taken together, the functional HSFs’ redundancy, previously described in avian
embryonic tissues and in undifferentiated cells, may be lost during ontogeny. As a result,
an exclusive reciprocal activation of HSF1 and HSF3 is observed in vivo in the brain and
in blood tissues, respectively. HSFs’ functional redundancy may be a mechanism unique
to undifferentiated embryonic cells, and is lost due to physiological alterations that
accompany maturation.

18
Refferences
1. Arad Z. Effect of water deprivation and heat exposure on body weight loss and
drinking capacity in four breeds of fowl (Gallus domesticus). Comp. Biochem. Physiol.
73A: 297-299, 1982.
2. Arad Z. Thermoregulation and acid-base status in the panting dehydrated fowl. J.
Appl. Physiol. 54: 234-243, 1983.
3. Arad Z. Avian brain cooling - a review. J. Basic & Clin. Physiol. Pharmacol. 1: 241-
254, 1990.
4. Arad Z and Marder J. Strain differences in heat resistance to acute heat stress
between the Bedouin desert fowl, the White Leghorn and their crossbreeds. Comp.
Biochem. Physiol. 72A: 191-193, 1982a.
5. Arad Z and Marder J. Comparative thermoregulation of four breeds of fowl (Gallus
domesticus), exposed to gradual increase of ambient temperatures. Comp. Biochem.
Physiol. 72A: 179-184, 1982b.
6. Arad Z and Marder J. Effect of long-term heat acclimation on the thermoregulatory
mechanisms of the desert Sinai fowl and the White Leghorn (Gallus domesticus). Comp.
Biochem. Physiol. 72A: 185-190 (1982c)

19
7. Arad Z and Marder J. Acid-base regulation during thermal panting in the fowl
(Gallus domesticus): Comparison between breeds. Comp. Biochem. Physiol. 74A: 125-
130, 1983.
8. Arad Z, Marder J, and Eylath U. Serum electrolyte and enzyme responses to heat
stress and dehydration in the fowl (Gallus domesticus). Comp. Biochem. Physiol. 74A:
449-453, 1983.
9. Baler R, Dahl G, and Voellmy R. Activation of human heat shock genes is
accompanied by oligomerization, modification, and rapid translocation of heat shock
transcription factor HSF1. Mol. Cell. Biol. 13: 2486–2496, 1993.
10. Christians E, Davis AA, Thomas SD, and Benjamin IJ. Maternal effect of Hsf1 on
reproductive success. Nature 407: 693-694, 2000.
11. Cotto JJ, Kline M, and Morimoto RI. Activation of heat shock factor 1 DNA
binding precedes stress-induced serine phosphorylation. Evidence for a multistep
pathway of regulation. J Biol Chem. 271: 3355-3358, 1996.
12. Cotto J, Fox S, and Morimoto RI. HSF1 granules: a novel stress-induced nuclear
compartment of human cells. J. Cell Sci. 110: 2925-2934, 1997.

20
13. Dyer RB and Herzog NK. Isolation of intact nuclei for nuclear extract preparation
from a fragile B-lymphocyte cell line. Biotechniques 19: 192-195, 1995.
14. Eriksson M, Jokinen E, Sistonen L, and Leppa S. Heat shock factor 2 is activated
during mouse heart development. Int J Dev Biol. 44: 471-477, 2000.
15. Feder ME, and Hofmann GE. Heat-shock proteins, molecular chaperones, and the
stress response: evolutionary and ecological physiology. Annu. Rev. Physiol. 61: 243-
282, 1999.
16. Green M, Schuetz TJ, Sullivan EK, and Kingston RE. A heat shock-responsive
domain of human HSF1 that regulates transcription activation domain function. Mol.
Cell. Biol. 5: 3354-3362, 1995.
17. Hohtola E and Visser GH. Avian Growth and Development Evolution within the
Altricial-Precocial Spectrum. Oxford: Oxford University Press, 1998, p. 157-173.
18. Holmberg CI, Hietakangas V, Mikhailov A, Rantanen JO, Kallio M, Meinander
A, Hellman J, Morrice N, MacKintosh C, Morimoto RI, Eriksson JE, and Sistonen
L. Phosphorylation of serine 230 promotes inducible transcriptional activity of heat shock
factor 1. EMBO J. 20: 3800-3810, 2001.

21
19. Inouye S, Katsuki K, Izu H, Fujimoto M, Sugahara K, Yamada S, Shinkai Y,
Oka Y, Katoh Y, and Nakai A. Activation of heat shock genes is not necessary for
protection by heat shock transcription factor 1 against cell death due to a single exposure
to high temperatures. Mol Cell Biol. 23: 5882-5895, 2003.
20. Izu H, Inouye S, Fujimoto M, Shiraishi K, Naito K, and Nakai A. Heat shock
transcription factor 1 is involved in quality-control mechanisms in male germ cells. Biol.
Reprod. 70: 18-24, 2004.
21. Itsaky-Gluklich S and Arad Z. The effect of dehydration on brain temperature
regulation of the Japanese quail (Coturnix coturnix japonica). Comp. Biochem. Physiol.
101A: 583-588, 1992.
22. Jolly C, Morimoto R, Robert-Nicoud M, and Vourc'h C. HSF1 transcription factor
concentrates in nuclear foci during heat shock: relationship with transcription sites. J.
Cell Sci. 110: 2935-2941, 1997.
23. Jolly C, Usson Y, and Morimoto RI. Rapid and reversible relocalization of heat
shock factor 1 within seconds to nuclear stress granules. Proc Natl Acad Sci U S A. 96:
6769-6774, 1999.

22
24. Jolly C, Konecny L, Grady DL, Kutskova YA, Cotto JJ., Morimoto RI, and
Vourc'h C. In vivo binding of active heat shock transcription factor 1 to human
chromosome 9 heterochromatin during stress. J Cell Biol. 156: 775-781, 2002.
25. Jolly C, Metz A, Govin J, Vigneron M, Turner BM, Khochbin S, and Vourc'h C.
Stress-induced transcription of satellite III repeats. J Cell Biol. 164: 25-33, 2004.
26. Junqueira LC, Carneiro J, and Kelley RO. Hematopoiesis. In: Basic Histology; a
Lange medical book, edited by Appleton and Lange, 1989, p. 245-250.
27. Jurivich DA, Sistonen L, Kroes RA, and Morimoto RI. Effect of sodium salicylate
on the human heat shock response. Science. 255: 1243-1245, 1992.
28. Kawazoe Y, Tanabe M, Sasai N, Nagata K, and Nakai A. HSF3 is a major heat
shock responsive factor during chicken embryonic development. Eur. J. Biochem. 65:
688-697, 1999.
29. Kline MP and Morimoto RI. Repression of the heat shock factor 1 transcriptional
activation domain is modulated by constitutive phosphorylation. Mol. Cell. Biol. 17:
2107-2115, 1997.

23
30. Knauf U, Newton EM, Kyriakis J, and Kingston RE. Repression of human heat
shock factor 1 activity at control temperature by phosphorylation. Genes Dev. 10: 2782-
2793, 1996.
31. Lee BS, Chen J, Angelidis C, Jurivich DA, and Morimoto RI. Pharmacological
modulation of heat shock factor 1 by antiinflammatory drugs results in protection against
stress-induced cellular damage. Proc Natl Acad Sci U S A. 92: 7207-7211 1995.
32. Marder J, Arad Z, and Gafni M. The effect of high ambient temperatures on acid-
base balance of the panting Bedouin fowl (Gallus domesticus). Physiol. Zool. 47: 180-
189, 1974.
33. Marder, J. and Arad, Z. Panting and acid-base regulation in heat stressed birds -
minireview. Comp. Biochem. Physiol. 94A: 395-400, 1989.
34. McMillan DR, Xiao X, Shao L, Graves K, and Benjamin IJ. Targeted disruption
of heat shock transcription factor 1 abolishes thermotolerance and protection against
heat-inducible apoptosis J. Biol. Chem. 273: 7523–7528, 1998.
35. Midtgard U. Scaling of the brain and eye cooling system in birds: a morphometric
analysis of the rete ophthalmicum. J. Exp. Zool. 225: 197-207, 1983.

24
36. Midtgard U, Arad Z, and Skadhauge E. The rete ophthalmicum and the relation of
its size to the body-to-brain temperature differences in the fowl (Gallus domesticus). J.
Comp. Physiol. B. 153: 241-246, 1983.
37. Morimoto RI. Regulation of the heat shock transcriptional response: cross talk
between a family of heat shock factors, molecular chaperones, and negative regulators.
Genes & Dev. 12: 3788-3796, 1998.
38. Mosser DD, Theodorakis NG, and Morimoto RI. Coordinate changes in heat shock
element-binding activity and HSP70 gene transcription rates in human cells. Mol. Cell.
Biol. 8: 4736-4744, 1988.
39. Nakai A. New aspects in the vertebrate heat shock factor system: HSF3 and HSF4.
Cell Stress & Chaperones 4: 86-93, 1999.
40. Nakai A, and Ishikawa T. A nuclear localization signal is essential for stress-
induced dimer-to-trimer transition of heat shock transcription factor 3. J. Biol. Chem.
275: 34665-34671, 2000.
41. Nakai A and Ishikawa T. Cell cycle transition under stress conditions controlled by
vertebrate heat shock factors. EMBO J. 20: 2885-2895, 2001.

25
42. Nakai A and Morimoto RI. Characterization of a novel chicken heat shock
transcription factor, HSF3, suggests a new regulatory pathway. Mol. Cell. Biol. 13: 1983-
1997, 1993.
43. Nakai A, Kawazoe Y, Tanabe M, Nagata K, and Morimoto RI The DNA-binding
properties of two heat shock factors, HSF1 and HSF3, are induced in the avian
erythroblast cell line HD6. Mol. Cell. Biol. 15: 5268-5278, 1995.
44. Nakai A, Tanabe M, Kawazoe Y, Inazawa J, Morimoto RI, and Nagata K. HSF4
a new member of the human heat shock factor family which lacks properties of a
transcriptional activator. Mol. Cell. Biol. 17: 469-481, 1997.
45. Pirkkala L, Nykanen P, and Sistonen L. Roles of the heat shock transcription
factors in regulation of the heat shock response and beyond. FASEB J. 15: 1118-1131,
2001.
46. Rabindran SK, Giorgi G, Clos J, and Wu C. Molecular cloning and expression of a
human heat shock factor, HSF1. Proc. Nat. Acad. Sci. USA 88: 6906-6910, 1991.
47. Sarge KD, Zimarino V, Holm K, Wu C, and Morimoto RI. Cloning and
characterization of two mouse heat shock factors with distinct inducible and constitutive
DNA-binding ability. Genes Dev. 5: 1902-1911, 1991.

26
48. Sarge, K. D., Murphy, S. P., and Morimoto, R. I. Activation of heat shock gene
transcription by heat shock factor 1 involves oligomerization, acquisition of DNA
binding activity, and nuclear localization and can occur in the absence of stress. .Mol.
Cell. Biol. 13: 1392–1407, 1993.
49. Sarge KD, Bray AE, and Goodson ML. Altered stress response in testis. Nature
374: 126, 1995.
50. Satyal SH, Chen D, Fox SG, Kramer JM, and Morimoto RI. Negative regulation
of the heat shock transcriptional response by HSBP1. Genes Dev. 12: 1962–1974, 1998.
51. Schuetz TJ, Gallo, GJ, Sheldon L, Tempst P, and Kingston RE. Isolation of a
cDNA for HSF2: evidence for two heat shock factors genes in humans. Proc. Nat. Acad.
Sci. USA. 88: 6911-6915, 1991.
52. Shabtay A and Arad Z. Ectothermy and Endothermy: Evolutionary perspectives of
thermoprotection by HSPs. J. Exp. Biol. 208: 2773-2781, 2005.
53. Shi Y, Kroeger PE, and Morimoto RI. The carboxyl-terminal transactivation
domain of heat shock factor 1 is negatively regulated and stress responsive. Mol. Cell.
Biol. 15: 4309-4318, 1995.

27
54. Sistonen L, Sarge K D, Phillips B, Abravaya K, and Morimoto RI. Activation of
heat shock factor 2 during hemin-induced differentiation of human erythroleukemia cells.
Mol. Cell. Biol. 12: 4104-4111, 1992.
55. Sistonen L, Sarge KD, and Morimoto RI. Human heat shock factors 1 and 2 are
differentially activated and can synergistically induce hsp70 gene transcription. Mol.
Cell. Biol. 14: 2087-2099, 1994.
56. Tanabe M, Nakai A, Kawazoe Y, and Nagata K. Different thresholds in the
responses of two heat shock transcription factors, HSF1 and HSF3. J. Biol. Chem. 272:
15389-15395, 1997.
57. Tanabe M, Kawazoe Y, Takeda S, Morimoto RI, Nagata K, and Nakai A.
Disruption of the HSF3 gene results in the severe reduction of heat shock gene expression
and loss of thermotolerance. EMBO J. 17: 1750-1758, 1998.
58. Tanabe M, Sasai N, Nagata K, Liu X-D, Liu PCC, Thiele DJ, and Nakai A. The
mammalian HSF4 gene generates both an activator and a repressor of heat shock genes
by alternative splicing. J. Biol. Chem. 274: 27845–27856, 1999.
59. Wu C. Heat shock transcription factors: structure and regulation. Annu Rev Cell Dev
Biol. 11: 441-469, 1995.

28
60. Xiao X, Zuo X, Davis AA, McMillan DR, Curry BB, Richardson JA, and
Benjamin IJ. HSF1 is required for extra-embryonic development, postnatal growth and
protection during inflammatory responses in mice. EMBO J. 18: 5943-5952, 1999.
61. Xing H, Wilkerson DC, Mayhew CN, Lubert EJ, Skaggs HS, Goodson ML,
Hong Y, Park-Sarge O-K, and Sarge KD. Mechanism of hsp70i Gene Bookmarking.
Science 307: 421-423, 2005.

29
Acknowledgements
We thank Professor Akira Nakai for kindly providing the anti-HSF1 and anti-HSF3
antibodies and the HSP90α cDNA. The technical assistance of Ms. Shoshana Goldenberg
and Mr. Udi Ron is deeply acknowledged. This research was supported by the Israel
Science Foundation. This study was approved by the Technion Review Board of Animal
Experimentation (No. B-93-6).

30
Figure legends
Fig. 1. The effect of heat exposure on HSF1 and HSF3 activation in blood cells from
mature hen at Tb=45oC (A) or at Tb=43oC (B). Samples were prepared from blood cell’
nuclei extracts for electro mobility shift assay (EMSA). After incubation of nuclei lysates
(5µg) with αHSF1γ or αHSF3γ (0.5, 1, 2µl of 1:10 diluted serum in PBS, panel A or 0.5,
1, 2µl of 1:10 diluted serum in PBS for αHSF1 and 0.5, 1, 2µl of 1:100 diluted serum in
PBS for αHSF3, panel B) at room temperature for 15 min, [32P]oligonucleotide,
containing four inverted nGAAn repeats (HSE) was added. The mixtures were loaded on
4% native polyacrylamide gel.
Fig. 2. The effect of heat exposure on HSF3 activation in blood cells from mature hens of
thermosensitive (Leghorn) and thermoresistant (Bedouin) fowl. Samples were prepared
from blood cell’ nuclei extracts for EMSA. After incubation of nuclei lysates (5µg) with
αHSF3γ (1µl of 1:10 diluted serum in PBS) at room temperature for 15 min,
[32P]oligonucleotide, containing four inverted nGAAn repeats (HSE) was added. The
mixtures were loaded on 4% native polyacrylamide gel.
Fig. 3. The effect of heat exposure on the nuclear expression (A) and translocation (B) of
HSF1 in blood cells. Blood was sampled from brachial vein of mature hen at Tb=45oC
and prepared as a smear for microscopic observation. The smear was fixed, treated with
RNase and stained with PI for DNA visualization (red) and with polyclonal αHSF1β. The
HSF1 signal was detected by a secondary antibody conjugated to FITC (green). The

31
yellow appearance is a result of merge between green and red and indicates the
translocation of HSF1 into the nucleus. Images were obtained by confocal microscopy.
Fig. 4. A. The effect of heat exposure at Tb=45oC on the activation of HSF1 and HSF3
(A) and on the transcription of HSP90 (B) in the brain tissue of mature hens.
A. Samples were prepared from whole brain extracts for electro mobility shift assay
(EMSA). After incubation of lysates (10 µg) with αHSF1γ or αHSF3γ (1 µl of 1:10
diluted serum in PBS) at room temperature for 15 minutes, [32P]oligonucleotide
containing four inverted nGAAn repeats (HSE) was added. The mixtures were loaded on
4% native polyacrylamide gel. NS – non-specific binding, PI – pre-immune serum. B.
HSP90 mRNA levels were detected by chicken HSP90α cDNA as a probe. Each lane
represents a different individual.
32

33

34

35
HSF1
Tb(oC) C 42 43 44 45 44r 43r 42r 41r
Fig 3A
HSF1
Tb(oC) C 42 43 44 45 44r 43r 42r 41r
Fig 3A
Control HS
Fig 3B
Control HS
Fig 3B
36
HSP90
rRNA
C HS
Fig 4B
HSP90
rRNA
C HS
HSP90
rRNA
C HS
Fig 4B
Activated HSF
NS
Supershift
Free DNA
- PI + - - PI + -- - - + - - - +
C HS
Fig 4AαHSF1αHSF3
Activated HSF
NS
Supershift
Free DNA
- PI + - - PI + -- - - + - - - +
C HS
Fig 4AαHSF1αHSF3
Copyright © 2022 FDOKUMEN




















