
Progressive development of the rat osteoblast phenotype in vitro: reciprocal …
11
JOURNAL OF CELLULAR PHYSIOLOGY 143:420-430 (1990) Progressive Development of the Rat Osteoblast Phenotype In Vitro: Reciprocal Relationships in Expression of Genes Associated With Osteoblast Proliferation and Differentiation During Formation of the Bone Extracellular Matrix THOMAS A. OWEN, MICHAEL ARONOW, VICTORIA SHALHOUB, LEESA M. BARONE, LAURENS WILMING, MELISSA S. TASSINARI, MARY BETH KENNEDY, SHlRWlN POCKWINSE, JANE B. LIAN, AND GARY S. STEIN Department of Cell Biology, University of Massachusetts Medical Center, Worcester, Massachusetts 0 1655 The relationship of cell proliferation to the temporal expression of genes charac- terizing a developmental sequence associated with bone cell differentiation was examined in primary diploid cultures of fetal calvarial derived osteoblasts by the combined use of autoradiography, histochemistry, biochemistry, and mRNA as- says of osteoblast cell growth and phenotypic genes. Modifications in gene ex- pression define a developmental sequence that has 1) three principle periods- proliferation, extracellular matrix maturation, and mineralization-and 2) two restriction points to which the cells can progress but cannot pass without further signals-the first when proliferation is down-regulated and gene expression as- sociated with extracellular matrix maturation is induced, and the second when mineralization occurs. Initially, actively proliferating cells, expressing cell cycle- and cell growth-regulated genes, produce a fibronectinhype I collagen extracel- lular matrix. A reciprocal and functionally coupled relationship between the decline in proliferative activity and the subsequent induction of genes associated with matrix maturation and mineralization is supported by 1) a temporal se- quence of events in which there is an enhanced expression of alkaline phos- phatase immediately following the proliferative period, and later, an increased expression of osteocalcin and osteopontin at the onset of mineralization; 2) in- creased expression of a specific subset of osteoblast phenotype markers, alkaline phosphatase and osteopontin, when proliferation is inhibited by hydroxyurea; and 3) enhanced levels of expression of the osteoblast markers as a function of ascorbic acid-induced collagen deposition, suggesting that the extracellular ma- trix contributes to both the shutdown of proliferation and the development of the osteoblast phenotype. Osteoblasts isolated from the calvaria o f 21 day fetal rat pups differentiate in culture as reflected by the production and deposition of an organized collagen ex- tracellular matrix and the appearance of nodules con- sisting of multiple layers of cells within a mineralized extracellular matrix (Bellows et al., 1986; Bhargava et al., 1988; Escarot-Charrier et al., 1983, 1988). Culture conditions that support high levels of production of os- teoblast markers such as type I collagen and osteocal- cin have been demonstrated, and, under such condi- tions, a temporal sequence of expression of type I collagen, alkaline phosphatase, osteocalcin, and min- eral deposition has been observed (Aronow et al., 1990; Gerstenfeld et al., 1987; Stein et al., 1989a), reflecting progressive osteoblast differentiation. The question arises o f how modifications in gene expression mediate these biochemical parameters. Equally important is determination of the signalling mechanisms for the temporal expression of genes encoding the osteoblast phenotype and the relationships between genes ex- pressed during the developmental sequence of osteo- blast differentiation. In the present studies, we examined the selective expression of various cell growth and differentiation associated genes as reflected by mRNA levels, which serve as markers for cells at different stages of matu- ration throughout the osteoblast developmental se- Received December 19, 1989; accepted February 9, 1990. @> 1990 WILEY-LISS, INC
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Progressive development of the rat osteoblast phenotype in vitro: reciprocal …

JOURNAL OF CELLULAR PHYSIOLOGY 143:420-430 (1990)
Progressive Development of the Rat Osteoblast Phenotype In Vitro: Reciprocal
Relationships in Expression of Genes Associated With Osteoblast Proliferation and
Differentiation During Formation of the Bone Extracellular Matrix
THOMAS A. OWEN, MICHAEL ARONOW, VICTORIA SHALHOUB, LEESA M. BARONE, LAURENS WILMING, MELISSA S. TASSINARI, MARY BETH KENNEDY,
SHlRWlN POCKWINSE, JANE B. LIAN, AND GARY S. STEIN Department of Cell Biology, University of Massachusetts Medical Center, Worcester,
Massachusetts 0 1655
The relationship of cell proliferation to the temporal expression of genes charac- terizing a developmental sequence associated with bone cell differentiation was examined in primary diploid cultures of fetal calvarial derived osteoblasts by the combined use of autoradiography, histochemistry, biochemistry, and mRNA as- says of osteoblast cell growth and phenotypic genes. Modifications in gene ex- pression define a developmental sequence that has 1 ) three principle periods- proliferation, extracellular matrix maturation, and mineralization-and 2) two restriction points to which the cells can progress but cannot pass without further signals-the first when proliferation is down-regulated and gene expression as- sociated with extracellular matrix maturation is induced, and the second when mineralization occurs. Initially, actively proliferating cells, expressing cell cycle- and cell growth-regulated genes, produce a fibronectinhype I collagen extracel- lular matrix. A reciprocal and functionally coupled relationship between the decline in proliferative activity and the subsequent induction of genes associated with matrix maturation and mineralization is supported by 1) a temporal se- quence of events in which there i s an enhanced expression of alkaline phos- phatase immediately following the proliferative period, and later, an increased expression of osteocalcin and osteopontin at the onset of mineralization; 2) in- creased expression of a specific subset of osteoblast phenotype markers, alkaline phosphatase and osteopontin, when proliferation i s inhibited by hydroxyurea; and 3) enhanced levels of expression of the osteoblast markers as a function of ascorbic acid-induced collagen deposition, suggesting that the extracellular ma- trix contributes to both the shutdown of proliferation and the development of the osteoblast phenotype.
Osteoblasts isolated f r o m t h e ca lvar ia o f 21 day fe ta l rat pups di f ferent iate in cu l tu re as ref lected by the product ion and deposit ion of an organized col lagen ex- t racel lu lar matrix and the appearance of nodules con- s is t ing of mul t i p le layers of cells within a mineral ized extracel lu lar matrix (Bellows e t al., 1986; Bhargava e t al., 1988; Escarot-Charrier e t al., 1983, 1988). Cu l tu re condit ions that support high levels of product ion of os- teoblast marke rs such as type I col lagen and osteocal- c i n have been demonstrated, and, under such condi- tions, a tempora l sequence of expression o f type I collagen, a l ka l i ne phosphatase, osteocalcin, and min- e ra l deposition has been observed (Aronow e t al., 1990; Gerstenfeld e t al., 1987; S te in e t al., 1989a), re f l ec t i ng progressive osteoblast dif ferentiat ion. The quest ion
arises o f h o w modif icat ions in gene expression mediate these biochemical parameters. Equally impor tan t i s de terminat ion o f t h e s igna l l ing mechanisms for t he temporal expression of genes encoding t h e osteoblast phenotype and the relat ionships between genes ex- pressed during t h e developmental sequence o f osteo- b las t d i f ferent iat ion.
In t h e present studies, w e examined the selective expression o f var ious ce l l g r o w t h and di f ferent iat ion associated genes as ref lected by mRNA levels, wh ich serve as marke rs for cells at di f ferent stages o f matu- r a t i o n th roughout the osteoblast developmental se-
Received December 19, 1989; accepted February 9, 1990.
@> 1990 WILEY-LISS, INC

GENE EXPRESSION DURING OSTEOBLAST DIFFERENTIATION 42 1
quence. Our studies show not only that a temporal se- quence of expression of the genes encoding osteoblast phenotypic markers occurs in culture but also that the pattern of their expression defines three distinct peri- ods separated by two transition points. Initially, there is a period of active proliferation during which cell growth-related genes are actively expressed, and max- imal levels of type I collagen mRNA are observed. Fol- lowing the down-regulation of proliferation, a period of matrix maturation occurs, when the alkaline phos- phatase gene is maximally expressed and the extracel- lular matrix is rendered competent for mineralization, the third period of the developmental sequence. Evi- dence is provided for two transitions between the de- velopmental periods representing restriction points to which cells can proceed but cannot pass without other signals. The first transition occurs with completion of the proliferative period, and the second is reached when the expression of genes such as osteocalcin and osteopontin become significantly elevated, with the on- set of mineralization. In addition, our results support two concepts related to the progressive development of the osteoblast phenotype. First, the proliferative period is requisite for collagen gene expression and initial pro- duction of a collagen matrix that subsequently sup- ports maturation of the osteoblast phenotype. Second, there exists a reciprocal and functionally coupled rela- tionship between the down-regulation of proliferation and initiation of expression of osteoblast phenotypic markers such as alkaline phosphatase and osteopontin.
MATERIALS AND METHODS
Cell culture Calvaria from fetal rats of 21 days gestation were
isolated and subjected to sequential digestions of 20, 40, and 90 min at 37°C in 2 mg/ml collagenase A (Boeh- ringer-Mannheim, Indianapolis, IN)/0.25% trypsin (Gibco, Grand Island, NY) (Aronow et al., in press). The cells of the first two digests were discarded, and those released from the third digestion were plated in mini- mal essential medium (MEM; Gibco) supplemented with 10% fetal calf serum (FCS) in 100 mm dishes or six-well dishes (Corning, Corning, NY) a t a density of 5 x lo5 or 3.6 x lo5 cells/dish, respectively. At con- fluence (day 7), the time course of mineralization was accelerated in some cultures by the addition of BGJb medium (Gibco) supplemented with 10% FCS, 50 pg/ml ascorbic acid, and 10 mM P-glycerol phosphate. In other cultures the concentration of ascorbic acid was varied, and in yet other cultures no P-glycerol phos- phate was added. In some experiments, proliferation was inhibited in actively proliferating cells (day 5 ) by the addition of 5 mM hydroxyurea directly to the cul- ture medium. Cell number was determined for points after plating by extensively incubating three indepen- dent wells of cells with 0.25% trypsin and counting the cells in a hemocytometer. When calvarial osteoblasts were isolated from 21 day fetal rats, the time of max- imal expression of the principle phenotype markers of the 35 day developmental sequence did not vary by more than 48 h r in independent experiments using a standardized lot of fetal calf serum (Aronow et al., 1990).
RNA isolation and analysis For each time point and experimental condition an-
alyzed, cells were scraped and pooled from 3 x 100 mm dishes in phosphate-buffered saline (PBS) and pelleted by centrifugation. The PBS was decanted, and the cell pellet was immediately frozen in liquid nitrogen and stored a t - 70°C until the experiment was completed. Total cellular RNA was isolated from each cell pellet by the SDS-proteinase K method as described by Plumb et al. (1983). RNA preparations were quanti- tated by absorbance a t 260 nm and intactness assessed by ethidium bromide staining following separation in 6.6% formaldehyde-1% agarose gels. RNA fractionated in such gels was transferred to Zeta-Probe membrane (BioRad, Richmond, CA) in 20 x SSC by the capillary method of Thomas (1983). In other cases, RNA samples were bound to Zeta-Probe blotting membrane by slot blot under conditions described by the apparatus manufacturer (Schleicher and Schuell, Keene, NH). DNA probes used for hybridization were the following: rat H4 histone (Grimes et al., 1987), human c-fos (Curran et al., 19831, mouse c-myc (Toscani et al., 1988), rat type I collagen (Genovese et al., 19841, ra t type I1 collagen (Kohno et al., 1984), rat fibronectin (Schwarzbauer et al., 1983), human transforming growth factor p (TGF-p) (Derynck et al., 1985), ra t al- kaline phosphatase (Noda et al., 19871, rat osteopontin (Oldberg et al., 1986), and rat osteocalcin (Lian et al., 1989). All DNA probes were labeled with 32P-dCTP by the random primer method (Feinberg and Vogelstein, 1983 1. All prehybridizations and hybridizations were performed in 50% formamide; 5 x SSC; 5 x Denhardts; 50 mM PO, buffer, pH 6.5; 1% SDS; and 250 bg/ml salmon sgerm DNA at 42°C for 18 hr. For hybridiza- tions, 10 cpmiml probe and 10% dextran sulfate were added (Maniatis et al., 1982). Following hybridization, blots were washed twice a t room temperature and once at 65°C in 2 x SSC/O.l% SDS (15 min per wash) and then twice a t 65°C in 1 x SSC/O.l% SDS for nonhomol- ogous probes or in 0.1 x SSC/O.l% SDS for homologous probes (30 min per wash). The resulting autoradio- graphs were quantitated by scanning laser densitome- try. Each time point represents the average of tripli- cate assays from a t least two independent experiments.
Transmission electron microscopy Cultured fetal rat osteoblasts growing on glass cov-
erslips and isolated intact 21 day gestation fetal rat calvaria were fixed in 3% glutaraldehyde in 0.1 M so- dium cacodylate buffer, pH 7.4, with 3 mM CaC1, for 5 hr at room temperature. Intact rat calvaria were de- mineralized by incubation in 10% EDTA, pH 7.4, for 5 days a t room temperature. Specimens were postfixed in 1% osmium tetroxide in 0.1 M sodium cacodylate buffer, pH 7.4; dehydrated; and embedded in Poly/Bed 812 (Polysciences, Warrington, PA). After sectioning, specimens were stained with 4% uranyl acetate and Reynolds’ lead citrate and examined in a Jeol 100s transmission electron microscope.
Histochemical analysis Cells growing on 22 mm glass coverslips were rinsed
twice with ice-cold PBS and fixed for 10 min in absolute methanol ( -20°C). Alkaline phosphatase activity was

422 OWEN ET AL.
100 -
80 -
60 -
40 -
20 -
visualized by incubating the cells for 30 min at room temperature with shaking in 10 mM Tris HC1, pH 8.4, containing 20 mg/ml disodium napthol AS-MX phos- phate and 40 mgiml fast red TR salt (Sigma Chemical Co., St. Louis, MO). Mineral deposition was assessed by von Kossa staining of the cultures (30 min in 3% AgN03) (Clark, 1981).
DNA synthesis and autoradiography For determination of the rate of DNA synthesis at
each point indicated, 3H-thymidine (20 Ci/mmol; Am- ersham, Arlington Heights, IL) was added to the me- dium of cells growing in three wells of a six-well dish to 10 p.Ci/ml for 1 hr. Cells were rinsed with PBS and incubated with 5% (w/v) trichloroacetic acid (TCA) for 5 min. This TCA incubation was repeated, each cell layer solubilized in 0.5 ml 10% SDS, and counted in a scintillation counter following addition of 4 ml fluor.
For in situ autoradiography, 3H-thymidine was added to the medium of cells growing on 22 mm glass coverslips to 1 yCi/ml for 24 hr. Cells were then rinsed twice in ice-cold PBS and fixed in absolute methanol (-20°C). Coverslips were then air dried overnight or first stained for alkaline phosphatase as described above and then dried. Autoradiography was performed using Ilford K-5 emulsion as described by Baserga and Malamud (1969). Exposures were for 5-7 days at 4°C.
Biochemical determinations For protein, collagen, and mineral content, cell lay-
ers were hydrolyzed in 6 N HC1 under vacuum at 110°C for 24 hr. Aliquots of the hydrolysates were diluted in 0.5% lanthium chloride for determination of calcium concentration by atomic absorption spectroscopy in a Perkin-Elmer model 2001 spectrograph. Other aliquots were assayed for total amino acid composition using a Beckman 121M autoanalyzer and collagen content was calculated from the hydroxyproline concentration. Other cell layers were harvested by scraping in appro- priate buffers for each individual assay. Total DNA was assayed by a fluorometric procedure using diami- nobenzoic acid reagent as described by Vysatek (1982). Alkaline phosphatase activity was determined spectro- photometrically as described by measuring the amount of p-nitrophenol formed at 37°C after 30 min (Lowry et al., 1954). The medium from all these cell layers was simultaneously analyzed for osteocalcin by radioimmu- noassay as previously described (Gundberg et al., 1984). For all biochemical determinations, the value represents the mean of three independent samples.
RESULTS AND DISCUSSION Temporal sequence of gene expression during
the osteoblast developmental sequence During a 35 day period, primary cultures of calva-
rial-derived osteoblasts progressively develop a bone tissue-like organization consisting of multilayered nod- ules of cells in an ordered, mineralized collagen extra- cellular matrix. To address the extent to which genes associated with the develoDment of the bone cell Dhe-
A .
P z ln ln W
X W
E
9 E 9 I- z W V W a a
B.
0 z v) v) W U 0. X W
s z 5
z X
I-
W 0.
C.
11.
a a
z v) v) W
X W
9 R ; c z W 0 W 0. a
I I
04 0 1 0 2 0 3 0 4 0
0 1 0 20 30 J C
DAYS IN CULTURE
Fig. 1. Temporal expression of cell growth-, extracellular matrix-, and osteoblast phenotype-related genes during the development of the osteoblast phenotype in vitro. Primary isolated cells were cultured after confluence in BGJb medium supplemented with 10% FCS, 50 Fg/ml ascorbic acid, and 10 mM B-glycerol phosphate. Cellular RNA was isolated at the times indicated (3, 5, 7, 10, 12, 14, 16, 20, 28, and 35 days) during the differentiation time course and assayed for the steady state levels of various transcripts by Northern blot or slot blot analysis. The resulting blots were quantitated by scanning densitom- etry and the results plotted relative to the maximal expression of each transcript. A DNA synthesis (0) and cell growth-related genes: H4 histone (0); c-myc (w); c-fos (0). B Extracellular matrix-associated genes: type I collagen (0); fibronectin (0); TGF-B (0). C: Osteoblast phenotype marker genes: osteopontin (0); osteocalcin (0); alkaline phosphatase (0). Note the induction of alkaline phosphatase at the end of proliferation and the induction of osteocalcin and osteopontin with the onset of calcium deposition (day 12). Calcium accumulation
notype in vitro are expresskd, cellular levels of mRNA for cell growth and tissue-specific genes were deter- mined at selected time points throughout a 35 day cul- ture period. Figure l shows the results of Northern blot is represented in Figure 2

GENE EXPRESSION DURING OSTEOBLAST DIFFERENTIATION 423
analysis of RNA transcribed from three different classes of genes. Immediately following isolation of the cells from calvaria, the protooncogene c-fos was ex- pressed at its highest level, and c-myc was maximally expressed 1 day later (Fig. 1A). The expression of the cell growth-associated H4 histone gene (Fig. 1A) re- flects its well-known coupling to DNA synthesis (Plumb et al., 1983), and showed that the isolated rat osteoblasts undergo an initial period of proliferation during the first 10-12 days in culture. The down-reg- ulation of the expression of H4 histone and the protoon- cogenes c-fos and c-myc marks the end of the prolifer- ative period, with their mRNA levels less than 20% of maximum by day 15.
Figure 1B shows that several genes associated with formation of the extracellular matrix, type I collagen, fibronectin, and TGF-P were maximally expressed dur- ing the period of proliferation and were gradually down-regulated during the subsequent stages of the cultures. Type I1 collagen mRNA, a phenotypic marker for cartilage (Kohno et al., 19841, was not detectable a t any time in the osteoblast cultures, indicating the ab- sence of chondrocytes either coisolated with the osteo- blasts from calvaria or chondrocytes derived from po- tential progenitor cells during the 35 day culture period.
The expression of several genes associated with the bone cell phenotype is shown in Figure 1C. The expres- sion of alkaline phosphatase mRNA increases greater than ten-fold immediately following the down-regula- tion of proliferation during the period from 12 to 18 days. Then, as the cultures progressed into the miner- alization stage, cellular levels of alkaline phosphatase mRNA declined. Two other bone-related genes, osteo- pontin and osteocalcin, exhibited a different pattern of expression. Osteocalcin was not detectable prior to day 12 in culture and did not reach a significant level of expression until 16-20 days after isolation when its expression rose coordinately with the increase in total mineral accumulation in the cultures. Osteopontin similarly reached its peak expression during the min- eralization period (days 16-20). It was also expressed during the period of active proliferation (25% of maxi- mal levels), at days 5 and 7 (Fig. 10.
The expression of alkaline phosphatase mRNA and the onset of enzyme activity (one of the bone phenotype markers), preceded any significant mineralization of the osteoblast cultures, as measured either by mineral accumulation or by the increase in osteocalcin levels. These results suggest that alkaline phosphatase may be involved in preparation of the extracellular matrix for the ordered deposition of mineral and that the co- expression of other genes such as osteocalcin and os- teopontin may support the onset and progression of ex- tracellular matrix mineralization. Alternatively, the induction of expression of these genes may reflect an acquisition of osteoblast properties associated with sig- nalling in bone turnover. The expression of at least two of these genes follows the pattern of expression of their proteins, which were determined biochemically. Alka- line phosphatase enzyme activity, measured both by histochemical staining and spectrophotometrically, closely followed the expression of its mRNA (compare Fig. 1C and Fig. 2). Similarly, the increase in osteocal- cin mRNA paralleled the levels of osteocalcin detect-
able by radioimmunoassay. For example, osteocalcin synthesis on day 12 measured 0.8 nglyg DNA, in- creased to 3.7 nglyg DNA by day 16, and to 9.5 nglpg DNA by day 20. These increases occur within the pe- riod of mineral accumulation in these cultures (see Fig. 2 for calcium deposition). Taken together, the patterns of expression of these genes demonstrate that a tempo- ral sequence of gene expression exists during the cul- ture period associated with development of the extra- cellular matrix and reflects maturation of the osteoblast phenotype in vitro.
It is of interest to note that osteopontin mRNA ex- hibits a biphasic pattern of expression during the os- teoblast developmental sequence, with mRNA levels during the period of cell proliferation that are about 25% those observed during the period of mineraliza- tion. This is not an unexpected result for two reasons. First, osteopontin expression during the proliferative period is consistent with its increased level of expres- sion during the prereplicative phase of the cell cycle following serum stimulation of quiescent fibroblasts (Nomura et al., 1988). Second, its induced expression coincident with mineralization may be related to phys- ical properties of the protein. Osteopontin is a 60 kd acidic glycoprotein (Oldberg et al., 1986) containing o-phosphoserine, thereby possessing several putative calcium binding sites, which are known to be impor- tant both for cell proliferation and also for mineraliza- tion of the extracellular matrix in bone (Glimcher, 1985). It therefore appears that expression of the osteo- pontin gene early and late in the osteoblast develop- mental sequence may be mediated by alternative reg- ulatory mechanisms. The possibility should also be considered that the lower levels of osteopontin mRNA observed during the proliferative period may in part reflect mRNA transcribed in vivo in osteoblasts under- going matrix mineralization prior to their isolation from the fetal calvaria.
Role of the extracellular matrix in the shutdown of proliferation and expression of the
osteoblast phenotype Although the temporal patterns of expression of
these representative cell growth- and osteoblast- related genes define discrete periods within a sequence of events that supports or promotes development of the osteoblast phenotype in vitro, they do not provide a direct indication of specific mechanisms by which ex- pression of these genes mediates progressive bone cell phenotype development. Here, a particularly relevant question is whether these temporal events are func- tionally coupled or causally related. Functional rela- tionships between expression of genes and events asso- ciated with osteoblast phenotype development are suggested by their patterns of expression. It can be inferred from the high levels of type I collagen mRNA expression during osteoblast proliferation that the pro- liferative period supports synthesis and deposition of collagen required for subsequent events that are nec- essary to render the collagen matrix competent for the ordered deposition of mineral.
Experimental results that are consistent with a func- tional as well as a temporal relationship between pro- liferation and collagen synthesis are shown in Figure 2. The effect of various ascorbic acid concentrations (0,
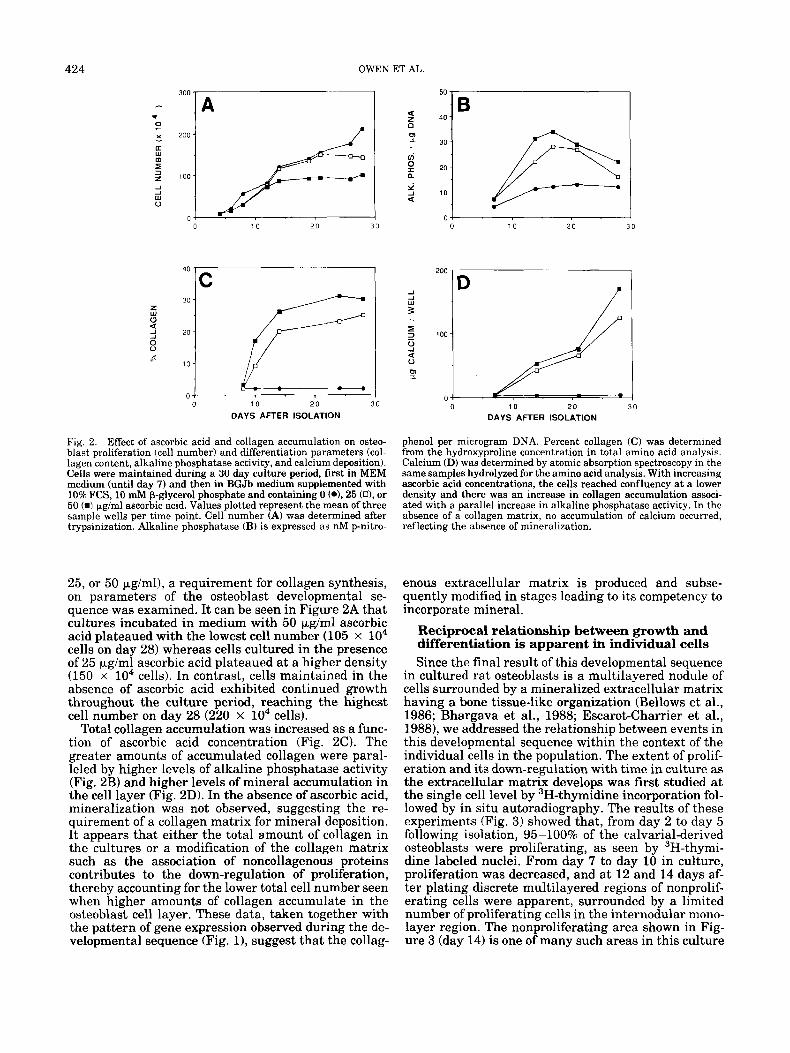
424
20 30: 10
OWEN ET AL.
gls U 0
Y
7
I
a m w
5 z 1 1 UI
U
z W 0 4 8 -1
c
300] A
0 1 0 20 3 0
C
p::::::i-l 0 10 20 30
DAYS AFTER ISOLATION
Fig. 2. Effect of ascorbic acid and collagen accumulation on osteo- blast proliferation (cell number) and differentiation parameters (col- lagen content, alkaline phosphatase activity, and calcium deposition). Cells were maintained during a 30 day culture period, first in MEM medium (until day 7) and then in BGJb medium supplemented with 10% FCS, 10 mM p-glycerol phosphate and containing 0 (*L 25 (n), or 50 (W pg/ml ascorbic acid. Values plotted represent the mean of three sample wells per time point. Cell number (A) was determined after trypsinization. Alkaline phosphatase (B) is expressed as nM p-nitro-
25, or 50 pglml), a requirement for collagen synthesis, on parameters of the osteoblast developmental se- quence was examined. It can be seen in Figure 2A that cultures incubated in medium with 50 pglml ascorbic acid plateaued with the lowest cell number (105 x lo4 cells on day 28) whereas cells cultured in the presence of 25 p,g/ml ascorbic acid plateaued at a higher density (150 x lo4 cells). In contrast, cells maintained in the absence of ascorbic acid exhibited continued growth throughout the culture period, reaching the highest cell number on day 28 (220 x lo4 cells).
Total collagen accumulation was increased as a func- tion of ascorbic acid concentration (Fig. 2 0 . The greater amounts of accumulated collagen were paral- leled by higher levels of alkaline phosphatase activity (Fig. 2B) and higher levels of mineral accumulation in the cell layer (Fig. 2D). In the absence of ascorbic acid, mineralization was not observed, suggesting the re- quirement of a collagen matrix for mineral deposition. It appears that either the total amount of collagen in the cultures or a modification of the collagen matrix such as the association of noncollagenous proteins contributes to the down-regulation of proliferation, thereby accounting for the lower total cell number seen when higher amounts of collagen accumulate in the osteoblast cell layer. These data, taken together with the pattern of gene expression observed during the de- velopmental sequence (Fig. l), suggest that the collag-
0 1 0 2 0 3 0
200
100
0
7 D
0 10 2 0 30
DAYS AFTER ISOLATION
phenol per microgram DNA. Percent collagen (C) was determined from the hydroxyproline concentration in total amino acid analysis. Calcium (D) was determined by atomic absorption spectroscopy in the same samples hydrolyzed for the amino acid analysis. With increasing ascorbic acid concentrations, the cells reached confluency at a lower density and there was an increase in collagen accumulation associ- ated with a parallel increase in alkaline phosphatase activity. In the absence of a collagen matrix, no accumulation of calcium occurred, reflecting the absence of mineralization.
enous extracellular matrix is produced and subse- quently modified in stages leading to its competency to incorporate mineral.
Reciprocal relationship between growth and differentiation is apparent in individual cells Since the final result of this developmental sequence
in cultured rat osteoblasts is a multilayered nodule of cells surrounded by a mineralized extracellular matrix having a bone tissue-like organization (Bellows et al., 1986; Bhargava et al., 1988; Escarot-Charrier et al., 19881, we addressed the relationship between events in this developmental sequence within the context of the individual cells in the population. The extent of prolif- eration and its down-regulation with time in culture as the extracellular matrix develops was first studied at the single cell level by 3H-thymidine incorporation fol- lowed by in situ autoradiography. The results of these experiments (Fig. 3) showed that, from day 2 to day 5 following isolation, 95-100% of the calvarial-derived osteoblasts were proliferating, as seen by 3H-thymi- dine labeled nuclei. From day 7 to day 10 in culture, proliferation was decreased, and at 12 and 14 days af- ter plating discrete multilayered regions of nonprolif- erating cells were apparent, surrounded by a limited number of proliferating cells in the internodular mono- layer region. The nonproliferating area shown in Fig- ure 3 (day 14) is one of many such areas in this culture

GENE EXPRESSION DURING OSTEOBLAST DIFFERENTIATION 425
Fig. 3. Down-regulation of proliferation during the initial period of the development of the osteoblast phenotype in vitro reveals the ini- tial cessation of proliferation in multilayered regions of the cultures. Primary cultures of osteoblasts growing on glass coverslips in MEM supplemented with 10% FCS and 50 pg/ml ascorbic acid (added at each feeding after day 7) were incubated with 'H-thymidine, fixed, and autoradiographed in situ as described in Materials and Methods. The days after initial plating are indicated under each photomicro- graph. ~100.
and demonstrates that, as proliferation is down-regu- lated, the extent of multilayering of cells increases. By day 21, the entire culture was multilayered, with little evidence of proliferative activity (Fig. 1).
The temporal relationship between proliferation and the formation of multilayered regions within the cul- tures reflects the down-regulation of the cell growth genes and the concurrent accumulation of an extracel- lular matrix (Figs. 1, 3). As indicated in Figure 1, al- kaline phosphatase mRNA was not induced until com- pletion of the proliferative period. To identify at the single cell level the relationship between proliferation and initiation of alkaline phosphatase expression, we combined the techniques of 3H-thymidine labeling and autoradiography with alkaline phosphatase histo- chemistry. Figure 4 directly demonstrates that it is the nonproliferating multilayered regions (seen in Fig. 3, day 12) that initially express alkaline phosphatase. The results shown in Figure 4 also confirm on an indi- vidual cell basis that a temporal sequence of gene ex- pression occurs and that at least some events (prolifer- ation and alkaline phosphatase expression) appear to be sequential, mutually exclusive events in the same cell. That is, proliferation must be down-regulated prior to the expression of alkaline phosphatase. Figure
Fig. 4. Histochemical changes during the rat osteoblast developmen- tal sequence. A The combination of alkaline phosphatase histochem- istry and autoradiography following 3H-thymidine incorporation shows that it is in multilayered regions of the cultures that prolifer- ation is first down-regulated, and i t is these regions that first become intensely alkaline phosphatase positive. Day 12. x 100. B: The entire culture then becomes alkaline phosphatase positive. Day 16. x 10. C: An ordered deposition of mineral in nodules as extracellular matrix develops within the multilayered regions of cells in the cultures. Day 35. x50.
4B shows that, following the initial expression of alka- line phosphatase in the heavily multilayered nodules where proliferation was first down-regulated, by day 21 all cells in the culture have ceased proliferation and exhibit alkaline phosphatase activity. This transition from a proliferating cell to one that can express an

426 OWEN ET AL.
Fig. 5. Transmission electron micrographs comparing cross sections of a 21 day fetal rat calvarium (column A) with those of a mineralized nodule formed in a 35 day osteoblast culture (column B). The top micrographs in each column show that the overall patterns of miner- alization of bone extracellular matrix in vivo and in vitro are indis- tinguishable, with mineralized extracellular matrix enveloping the cells. The second row of micrographs shows early stages of mineral- ization around the fringes of regions of heavy mineral deposition. Note the mineral deposition associated with collagen fibrils and the absence of intracellular calcification. The lower micrographs demon- strate identical orthogonal organization of the extracellular collagen matrix in both the intact calvarium and in the mineralized nodule formed in vitro. The lower micrograph in column A was made follow- ing sectioning of a demineralized calvarium. Bars in the lower micro- graphs represent 2.5 pm; others represent 5 pm.
early marker of the osteoblast phenotype (alkaline phosphatase) represents a restriction point where ces- sation of proliferation appears to be required for initi- ation of tissue-specific gene expression.
The completed developmental sequence is shown in Figure 4C, which demonstrates intense von Kossa staining of the mineralized nodules reflecting hydroxy- apatite deposition. The development of in uiuo-like bone-tissue organization in these cultures is further shown by comparison of the ultrastructure of the min- eralized regions of the culture with sections through an intact 21 day fetal rat calvarium (Fig. 5). Note the similar type of ordered deposition of crystals within and between the orthogonally organized bundles of col- lagen fibrils. No evidence for cell necrosis or intracel- lular calcification is indicated in the cultures, particu- larly where mineralized matrix has enveloped the osteoblasts.
Functional coupling of the stages of osteoblast development
Support for the concept that proliferation must be down-regulated prior to the onset of events associated
with osteoblast differentiation was also found at the molecular level. A specific change in proteidDNA in- teractions in the proximal promoter of a cell growth- regulated gene, H4 histone, has previously been shown both in this rat osteoblast differentiation model system (Owen et al., in press; Holthuis et al., in press) and following the induction of differentiation of HL60 pro- myelocytic leukemia cells by phorbol esters (Stein et al., 1989b). The down-regulation of proliferation is ac- companied first by a loss of H4 histone mRNA followed by a down-regulation of transcription and concomitant loss of the specific interaction of a nuclear promoter binding factor, HiNF-D, with a region of the H4 histone gene proximal promoter that influences both specific- ity and level of transcription of the gene. It i s this loss of H4 histone gene transcription and of HiNF-D bind- ing that accompanies differentiation of the cells. Other molecular markers for the transition from proliferation to the onset of differentiation are expression of a cell cycle-independent histone gene that encodes a high- molecular-weight poly-A + RNA (Shalhoub et al., 1989) and specific modifications in the composition of nuclear matrix proteins (Dworetzky, et al., in press).
A more direct demonstration that the down-regula- tion of proliferation results in the induction of expres- sion of some genes that normally are expressed later in the osteoblast developmental sequence is derived from experiments in which DNA synthesis was inhibited in actively proliferating (subconfluent) osteoblasts by hy- droxyurea treatment (5 mM). As demonstrated by a >90% decrease in DNA synthesis and H4 histone mRNA levels by 1 hr after hydroxyurea treatment (Fig. 61, cell proliferation is rapidly and selectively in- hibited under these experimental conditions. Simi- larly, by 1 hr following inhibition of DNA synthesis, a fourfold increase in alkaline phosphatase mRNA levels was observed, indicating that the down-regulation of proliferation prematurely induced the expression of an early marker for the extracellular matrix maturation period of the osteoblast developmental sequence. With inhibition of DNA synthesis by hydroxyurea, the levels of osteopontin mRNA increased to levels approximat- ing those present at the late stage (day 22) of the min- eralization period of the cultured osteoblasts (Fig. 6 ) . These data suggest a direct functional coupling of the down-regulation of proliferation and the preferential expression of genes whose induction is normally asso- ciated with the appearance of the mature osteoblast phenotype in a mineralized matrix. The absence of os- teocalcin induction is consistent with the concept that there is at least a second set of genes whose expression is coupled not directly to the down-regulation of prolif- eration but rather to development of the more differ- entiated osteoblast in a mineralized matrix. These ex- periments in which premature differentiation has been promoted by hydroxyurea inhibition of proliferation re- veal a second transition point in the developmental sequence of osteoblast differentiation, since inhibition of proliferation supports progression of the develop- mental sequence only to the stage where mineraliza- tion is initiated. Mineral deposition may be required to signal expression of a subset of osteoblast phenotype genes, such as osteocalcin. Other genes expressed in relation to the developing extracellular matrix (which are not induced by hydroxyurea inhibition of prolifer-

GENE EXPRESSION DURING OSTEOBLAST DIFFERENTlATION 427
z 0 cn cn W
P X W w >
- a
2 4 w U
@%-z-d " C . 1 4 M
b I M
D.
Fig. 6. Coupling of alkaline phosphatase and osteopontin but not osteocalcin expression to the down-regulation of proliferation. Prolif- eration was inhibited in actively growing osteoblasts (day 5) by addi- tion of 5 mM hydroxyurea (HU). Following HU addition, cells were harvested at 1 and 4 hr, examined for DNA synthesis (A), and cellular RNA prepared and analyzed for H4 histone (A), alkaline phosphatase (B), and osteopontin (C). The Northern blots of these transcripts and osteocalcin is also shown (D). Note that osteocalcin, which is not present in 5 day cultures (0, is also not induced following inhibition of proliferation by HU. For comparison, the relative expression of these genes in non-HU-treated control ( C ) and in mineralized (MI cultures (30 days after plating) is also shown.
ation) may be required to render the matrix competent for mineralization.
To address further the relationship between miner- alization and genes expressed during the osteoblast
developmental sequence, we modified the onset and extent of mineral deposition in the cultures. To ac- complish this, the cultures were maintained in medi- um containing ascorbic acid after day 7 but lacking the organic phosphate source, p-glycerol phosphate (pGP). Figure 7A demonstrates that, under accelerated min- eralizing conditions (in the presence of pGP), osteocal- cin mRNA and synthesis increased steadily beginning at day 15 in parallel with calcium accumulation in the cell layer. In contrast, when cultures were maintained in the absence of PGP, calcium did not begin to accu- mulate in the cell layer until approximately day 25. Notably, osteocalcin was not detectable until this point, and its low level of synthesis reflects the slower rate of mineralization. It is important to note that the presence or absence of PGP had no effect on the prolif- erative period (as measured by rate of DNA synthesis) or on the onset of alkaline phosphatase expression. These experiments provide additional evidence to support the existence of the second transition point. The cells can progress through the maturation se- quence to the onset of mineralization but cannot ini- tiate expression of genes related to the mineralization stage unless mineral accumulation occurs. It appears that genes such as osteocalcin are not only temporally expressed but also "coupled" to deposition of hydroxy- apatite.
The in vitro osteoblast developmental sequence is a reinitiation of events occurring in vivo
Since our primary cultures of osteoblasts are derived from developing fetal bone, a key question is whether we are observing in culture the initiation of develop- ment of the osteoblast phenotype from progenitor cells or whether the osteoblast developmental sequence observed in vitro is a reinitiation and reoccurrence of the fetal developmental sequence in vivo. Therefore, we examined the representation of mRNA transcripts from osteoblast phenotype marker genes in cells of the third trypsin-collagenase digestion used to establish the cultures. High levels of osteocalcin and osteopontin mRNA were present at isolation and were then down-regulated very rapidly. Osteocalcin mRNA was not detectable, and osteopontin levels were lower in cells cultured for 2 days. Since active expression of these genes is not restored until formation and maturation of the extracellular matrix (Figs. 1, 31, it appears that the in vitro temporal sequence is a recapitulation of events that occur in vivo. Indeed, the biological relevance of the developmental sequence we observe in vitro is supported by a similar sequential expression of genes in fetal rat calvaria during gestation (Yoon et al., 1987). In light of the high levels of osteocalcin and osteopontin transcripts found in the third digest cells and the removal of the osteoprogeni- tor cells due to scraping of the perichondrium and discarding of the first and second enzymatic digests, we believe that the cells expressing the developmental sequence in vitro are the ones that have expressed the osteoblast phenotype in vivo. The similarity of the levels of bone phenotype-associated transcripts in vivo and in vitro and the appearance of mineral organiza- tion in the electron micrographs support the physiolog- ical significance of this model system as an appropri-

428
4w-
3w -
OWEN ET AL.
B. A. 1 4 0
DAYS AFTER ISOLATION
Fig. 7. Conditions that delay mineralization of the osteoblast cul- tures do not effect the earlier events of proliferation and extracellular matrix maturation. Cultures maintained in BGJb medium supple- mented with 50 pg/ml ascorbic acid and 10 mM p-glycerol phosphate (A) show an earlier accumulation of calcium in the extracellular ma- trix and earlier osteocalcin expression than cultures maintained in
ate one in which to study molecular mechanisms asso- ciated with bone formation.
Consistent with a model in which the extracellular matrix surrounding the cells controls the differentia- tion process, enzymatic removal of the in vivo synthe- sized extracellular matrix when the primary calvarial- derived osteoblast cultures are established supports the reinitiation of the developmental sequence of pro- liferation, extracellular matrix maturation, and min- eralization. These cultures can be maintained for up to 120 days and possibly longer, with the cells retaining bone-like features. The reinitiation of proliferation- and differentiation-related events also occurs in sub- cultivated cells. Trypsinization and replating of ma- ture mineralized cultures of osteoblasts (after day 35) results in a rapid down-regulation of osteocalcin and osteopontin mRNAs. As is shown in Figure 8, cells pas- saged four times were capable of reinitiating expres- sion of osteoblast phenotype markers and mineralizing under the appropriate culture conditions. However, al- though subcultivated cells remain competent to ex- press the developmental sequence, we observed a vari- able lengthening of the time course required to develop a mineralized extracellular matrix. Whether cells in vivo are capable of reinitiating a similar developmen- tal sequence under the appropriate circumstances re- mains to be established, but the ability of the calvarial cells, once freed of the extracellular matrix, to differ- entiate in culture certainly suggests that this is possi- ble.
400
300
200 lo 4 0 100
200 4 :j ._k 0 0 10 20 30
40
30 20
10 lo 40
DAYS AFTER ISOLATION
the same medium without p-glycerol phosphate (B). However, cul- tures maintained under either culture regimen had essentially iden- tical periods of proliferation ("H-thymidine incorporation) and extra- cellular matrix development (alkaline phosphatase activity) (top panels in A and B). DNA synthesis (01, alkaline phosphatase activity (m), calcium (n), osteocalcin (m).
Fig. 8. Mineralization of osteoblast cultures following four passages of the cells. Osteoblasts isolated as described were allowed to progress through the in vitro developmental sequence by addition of BGJb medium supplemented with 50 pglml ascorbic acid and 10 mM p- glycerol phosphate and then passaged by trypsinization after devel- opment of the mineralized extracellular matrix. This sequence was repeated four times. The cells shown developed mineralized nodules 66 days after plating following the fourth passage and were stained by von Kossa. x 10.
Conclusions and model of osteoblast differentiation in vitro
The results we have presented based on dekrmina- tions of molecular, biochemical, and histochemical pa-

GENE EXPRESSION DURING OSTEOBLAST DIFFERENTIATION 429
CGUMITMENT PERIODS AND RESTRICTION POINTS
A, , DAYS
1 1 I
Fig. 9. Model of the relationship between proliferation and differen- tiation during the rat osteoblast developmental sequence. This rela- tionship is schematically illustrated within the context of modifica- tions in expression of cell cycle- and cell growth-regulated genes as well as genes associated with the maturation, development, and min- eralization of the osteoblast extracellular matrix. The three principle periods of the osteoblast developmental sequence are designated within broken vertical lines (proliferation, matrix development and maturation, mineralization). Commitment periods and restriction points, which are indicated in the lower portion of the figure, were experimentally established. A functional relationship between the
rameters are consistent with a reciprocal and func- tional relationship between proliferation and the se- quential development of the osteoblast phenotype, which is schematically shown in Figure 9. The progres- sive and interdependent series of biochemical events that characterizes the osteoblast developmental se- quence reflects the selective expression initially of cell growth and subsequently of a series of tissue-specific genes. Such modifications in gene expression define a developmental sequence with three distinct periods: proliferation, extracellular matrix maturation, and mineralization. Our experimental results indicate two principle restriction points in the osteoblast develop- mental sequence to which the cells can progress but cannot pass without further appropriate signals, the first when proliferation is down-regulated and gene ex- pression associated with extracellular matrix matura- tion is initiated and the second when mineralization occurs.
Our working hypothesis is that genes involved in the production and deposition of the extracellular matrix must be expressed during the proliferative period for the onset and progression of differentiation to occur. We postulate that proliferation is functionally related to the synthesis of an organized bone-specific extracel- lular matrix and that the maturation and organization of the extracellular matrix contributes to the shutdown of proliferation, which then renders the matrix compe- tent for the mineralization that is essential for com- plete expression of the osteoblast phenotype. This
I
down-regulation of proliferation and the initiation of extracellular matrix maturation and development is based on stimulation of alka- line phosphatase and osteopontin gene expression when proliferation is inhibited, but the developmental sequence is induced only to the second transition point. Growth of the osteoblast under conditions that do not support mineralization confirms the day 20 restriction point since the developmental sequence proceeds through the prolif- eration and the extracellular matrix development/maturation peri- ods, but not further. DNA, DNA synthesis; col I, collagen Type I; AP, alkaline phosphatase; OP, osteopontin; OC; osteocalcin; mineral, total accumulated calcium.
working model provides the basis for addressing whether particular stages of osteoblast differentiation exhibit selective responsiveness to actions of hormones and other physiological factors that influence osteo- blast activity and other such questions related to the molecular mechanisms associated with bone formation.
ACKNOWLEDGMENTS We thank Marie Giorgio for photographic assistance.
These studies were supported by grants from the Na- tional Institutes of Health (GM32010, GM32381, AR35166, AR39122, HD22400), the National Science Foundation (DCB88-96116), the March of Dimes Birth Defects Foundation (1-813), and the International Life Sciences Institute (Washington, DC).
LITERATURE CITED Aronow, M.A., Gerstenfeld, L.C., Owen, T.A., Tassinari, M.S., Stein,
G.S., and Lian, J.B. Factors that promote progressive development of the osteoblast phenotype in cultured fetal rat calvaria cells. J. Cell. Physiol., 143:213-221.
Baserga, R., and Malamud, D. (1969) Autoradiography. Hoeber, New York.
Bellows, C.G., Aubin, J.E., Heersche, J.N.M., and Antosa, M.E. (1986) Mineralized bone nodules formed in vitro from enzymatically re- leased rat calvaria populations. Calcif. Tissue Int., 38:143-154.
Bhargava U., Bar-Lev, M., Bellows, C.G., and Aubin, V. (1988) U1- trastructural analysis of bone nodules formed in vitro by isolated fetal rat calvarial cells. Bone, 9:155-163.
Clark, G. (1981) Miscellaneous stains. In Staining Procedures, 4th Ed. G. Clark, ed. Williams and Wilkins, Baltimore, p. 187.

430 OWEN ET AL.
Curran, T., MacConnell, W.P., van Straaten, F., and Verma, I.M. (1983) Structure of the FBJ Murine Osteosarcoma Virus genome: Molecular cloning of its associated helper virus and the cellular homolog of the v-fos gene from mouse and human cells. Mol. Cell. Biol., 37914-921.
Derynck, R., Jarrett, J.A., Chen, E.Y., Eaton, D.H., Bell, J.R., As- soian. R.K.. Roberts. A.B.. SDorn. M.B.. and Goeddel. D.V. (1985) Human transforming growih factor+ ' complementary DNA se- quence and expression in normal and transformed cells. Nature, 316:701-705.
Dworetzky, S.I., Fey, E.G., Penman, S., Stein, J.L., Lian, J.B., and Stein, G.S. Progressive changes in the protein composition of the nuclear matrix during rat ostoeblast differentiation. Proc. Natl. Acad. Sci. USA, in press.
Escarot-Charrier, B., Glorieux, F.H., van der Rest, M., and Pereira, G. (1983) Osteoblasts isolated from mouse calvaria initate matrix min- eralization in culture. J . Cell Biol., 96:639-643.
Escarot-Charrier, B., Shepard, N., Charette, G., Grynpas, M., and Glorieux, F.H. (1988) Mineralization in osteoblast cultures: a light and electron microscopic study. Bone, 9:147-154.
Feinberg, A.P., and Vogelstein, B. (1983) A technique for radiolabel- ing DNA restriction endonuclease fragments to a high specific ac- tivity. Anal. Biochem., 132:6-13.
Genovese, C., Rowe, D., and Kream, B. (1984) Construction of DNA sequences complementary to rat a1 and a 2 collagen mRNA and their use in studying the regulation of type I collagen synthesis by 1,25-dihydroxy D. Biochemistry, 23:6210-6216.
Gerstenfeld, L.C., Chipman, S.D., Glowacki, J., and Lian, J.B. (1987) Expression of differentiated function by mineralizing cultures of chicken osteoblasts. Dev. Biol., 122:49-60.
Glimcher, M.J. (1985) Role of collagen and phosphoproteins in the calcification of bone and other collagenous tissues. In Calcium in Biological Systems. R.P. Rubin, G.B. Weiss, and J.W. Putney, ed. Plenum Press, New York, pp. 607-616.
Grimes, S., Weisz-Carrington, P., Daum, H. 111, Smith, J., Green, L., Wright, K., Stein, G., and Stein, J. (1987) A rat histone H4 gene closely associated with the testis-specific H l t gene. Exp. Cell Res., 173534445 ,
Gundbera. C.M., Hauschka, P.V., Lian, J.B., and Gallop, P.M. (1984) Osteocalcin isolation, characterization, and detection: Methods En- zymol., 107:516-544.
Holthuis, J., Owen, T.A., van Wijnen, A.J., Wright, K.L., Ramsey- Ewing, A., Kennedy, M.B., Cosenza, S.C., Carter, R., Soprano, K.J., Lian, J.B., Stein, J.L., and Stein, G.S. Tumor cells exhibit deregu- lation of the cell cycle histone gene promoter factor HiNF-D. Sci- ence, in press.
Kohno, K., Martin, G.R., and Yamada, K. (1984) Isolation and char- acterization of a cDNA clone for the amino-terminal portion of the pro-al(I1) chain of cartilage collagen. J . Biol. Chem., 259.13668- 13673.
Lian, J., Stewart, C., Puchaz, E., Mackowiak, S., Shalhoub, V., Col- lart, D., Zambetti, G., and Stein, G. (1989) Structure of the ra t osteocalcin gene and regulation of vitamin D-dependent expression. Proc. Natl. Acad. Sci. USA. 86.1143-1147.
Lowry, O.H., Roberts, N.R., Wu, M., Hixen, W.S., and Crawford, D. (1954) The quantitative histochemistry of brain 11. Enzyme mea- surements. J. Biol. Chem., 207:13-19.
Maniatis, T., Fritsch, E., and Sambrook, J . (1982) Molecular Cloning, A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Noda, M., Yoon, K., Thiede, M., Buenga, R., Weiss, M., Henthorn, P., Harris, H., and Rodan, G. (1987) cDNA cloning of alkaline phos- phatase from rat osteosarcoma (ROS 17/2.8) cells. J. Bone Mineral Res., 2:161-164.
Nomura, S., Wills, A.J., Edwards, D.R., Heath, J.K., and Hogan, B.L.M. (1988) Developmental expression of 2ar (osteopontin) and SPARC (osteonectin) RNA as revealed by in situ hybridization. J. Cell Biol., 106:441-450.
Oldberg, A,, Franze, A,, and Heinegard, D. (1986) Cloning and se- quence analysis of rat bone sialoprotein (osteopontin) cDNA reveals an Arg-Gly-Asp cell-binding sequence. Proc. Natl. Acad. Sci. USA, 83:8819-8823.
Owen, T.A., Holthuis, J., Markose, E., van Wijnen, A.J., Lian, J.B., and Stein, G.S. Modifications of protein-DNA interactions in the proximal promoter a t a cell growth regulated histone gene during the onset and progression of osteoblast differentiation. Proc. Natl. Acad. Sci. USA, in press.
Plumb, M., Stein, J., and Stein, G. (1983) Coordinate regulation of multiple histone mRNAs during the cell cycle in HeLa cells. Nu- cleic Acids Res., 11:2391-2410.
Sehwarzbauer, J.E., Tamkin, J.W., Lemischka, I.R. and Hynes, R.O. (1983) Three different fibronectin mRNAs arise by alternative splicing within the coding region. Cell, 35.421-431.
Shalhouh, V., Gerstenfeld, L.C., Collart, D., Lian, J.B., and Stein, G.S. (1989) Downregulation of cell growth and cell cycle regulated genes during chick osteoblast differentiation with the reciprocal expres- sion of histone gene variants. Biochemistry, 28.5318-5322.
Stein, G.S., Lian, J.B., Gerstenfeld, L., Shalhoub, V., Aronow, M., Owen, T., and Markose, E. (1989a) The onset and progression of osteoblast differentiation is functionally related to cellular prolif- eration. Connective Tissue Res., 20.3-13.
Stein, G., Lian, J . , Stein, J., Briggs, R., Shalhoub, V., Wright, K., Pauli, U., and van Wijnen, A (1989b) Altered binding of human histone gene transcription factors during the shutdown of prolifer- ation and onset of differentiation in HL60 cells. Proc. Natl. Acad. Sci. USA, 86:1865-1869.
Thomas, P.S. (1983) Hybridization of denatured RNA transferred or dotted to nitrocellulose paper. Methods Enzymol., 100:255-266.
Toscani, A., Soprano, D.R., and Soprano, K.J. (1988) Molecular anal- ysis of sodium butyrate induced growth arrest. Oncogene Res., 3: 223-238.
Vysatek, R. (1982) A sensitive fluorometric assay for the determina- tion of DNA. Anal. Biochem., 48:243-248.
Yoon, K., Buenaga, R., and Rodan, G.A. (1987) Tissue specificity and developmental expression of osteopontin. Biochem. Biophys. Res. Commun., 148:1129-1136.




















