
Recent developments in anti-inflammatory natural products
54
Recent Developments in Anti- In£ammatory Natural Products Raju Gautam and Sanjay M. Jachak Department of Natural Products, National Institute of Pharmaceutical Education and Research (NIPER), Sector 67, S.A.S Nagar, Mohali-160 062, Punjab, India Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/med.20156 . Abstract: Many of the inflammatory diseases are becoming common in aging society throughout the world. The clinically used anti-inflammatory drugs suffer from the disadvantage of side effects and high cost of treatment (in case of biologics). Alternative to these drugs are traditional medicines and natural products, which offer a great hope in the identification of bioactive lead compounds and their development into drugs for treating inflammatory diseases. Since ancient times traditional medicines and phytopharmaceuticals are being used for the treatment of inflammatory and other disorders. The present review article describes anti- inflammatory natural products derived from plants and marine sources reported during last decade. The compounds described belong to different chemical classes such as alkaloids, steroids, terpenoids, poly- phenolics, phenylpropanoids, fatty acids and lipids, and various miscellaneous compounds. The attempt is also being made to enumerate the possible leads, e.g. curcumin, resveratrol, baicalein, boswellic acid, betulinic acid, ursolic acid, and oleanolic acid, for further development with the help of structure–activity relationship (SAR) studies and their current status. In addition SAR studies carried out on the anti- inflammatory activity of flavonoid compounds and clinical studies performed on anti-inflammatory natural products are also discussed. & 2009 Wiley Periodicals, Inc. Med Res Rev Key words: natural products; anti-inflammatory; alkaloids; terpenoids; polyphenolics 1. INTRODUCTION Inflammation is caused by a variety of stimuli including physical damage, ultra violet irra- diation, microbial invasion, and immune reactions. The classical key features of inflamma- tion are redness, warmth, swelling, and pain. Inflammation cascades can lead to the development of diseases such as chronic asthma, rheumatoid arthritis, multiple sclerosis, inflammatory bowel disease, and psoriasis. Many of these diseases are debilitating and are becoming increasingly common in our aging society. Rheumatoid arthritis and osteoarthritis are the major inflammatory diseases affecting people worldwide. Rheumatoid arthritis is an inflammatory condition that usually affects multiple joints. It affects 0.3–1.0% of the general Correspondence to: Sanjay M. Jachak, Department of Natural Products, NIPER, Sector-67, S.A.S Nagar, Mohali-160 062, Punjab, India, E-mail: [email protected] Medicinal Research Reviews & 2009 Wiley Periodicals, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Recent developments in anti-inflammatory natural products

Recent Developments in Anti-In£ammatoryNatural Products
Raju Gautam and Sanjay M. Jachak
Department of Natural Products, National Institute of Pharmaceutical Education and Research (NIPER), Sector
67, S.A.S Nagar, Mohali-160 062, Punjab, India
Published online in Wiley InterScience (www.interscience.wiley.com).
DOI 10.1002/med.20156
.
Abstract: Many of the inflammatory diseases are becoming common in aging society throughout the
world. The clinically used anti-inflammatory drugs suffer from the disadvantage of side effects and high cost
of treatment (in case of biologics). Alternative to these drugs are traditional medicines and natural products,
which offer a great hope in the identification of bioactive lead compounds and their development into drugs
for treating inflammatory diseases. Since ancient times traditional medicines and phytopharmaceuticals are
being used for the treatment of inflammatory and other disorders. The present review article describes anti-
inflammatory natural products derived from plants and marine sources reported during last decade. The
compounds described belong to different chemical classes such as alkaloids, steroids, terpenoids, poly-
phenolics, phenylpropanoids, fatty acids and lipids, and various miscellaneous compounds. The attempt
is also being made to enumerate the possible leads, e.g. curcumin, resveratrol, baicalein, boswellic acid,
betulinic acid, ursolic acid, and oleanolic acid, for further development with the help of structure–activity
relationship (SAR) studies and their current status. In addition SAR studies carried out on the anti-
inflammatory activity of flavonoid compounds and clinical studies performed on anti-inflammatory natural
products are also discussed. & 2009 Wiley Periodicals, Inc. Med Res Rev
Key words: natural products; anti-inflammatory; alkaloids; terpenoids; polyphenolics
1. INTRODUCTION
Inflammation is caused by a variety of stimuli including physical damage, ultra violet irra-diation, microbial invasion, and immune reactions. The classical key features of inflamma-tion are redness, warmth, swelling, and pain. Inflammation cascades can lead to thedevelopment of diseases such as chronic asthma, rheumatoid arthritis, multiple sclerosis,inflammatory bowel disease, and psoriasis. Many of these diseases are debilitating and arebecoming increasingly common in our aging society. Rheumatoid arthritis and osteoarthritisare the major inflammatory diseases affecting people worldwide. Rheumatoid arthritis is aninflammatory condition that usually affects multiple joints. It affects 0.3–1.0% of the general
Correspondence to: Sanjay M. Jachak, Department of Natural Products, NIPER, Sector-67, S.A.S Nagar, Mohali-160 062, Punjab,
India, E-mail: [email protected]
Medicinal Research Reviews
& 2009 Wiley Periodicals, Inc.

population and is more prevalent among women in developed countries. Persistent in-flammation leads to joint destruction, but the disease can be controlled with drugs. Os-teoarthritis, which is characterized by loss of joint cartilage that leads to pain and loss offunction primarily in the knees and hips, affects 9.6% of men and 18% of women aged morethan 60 years. Increases in life expectancy and aging populations are expected to makeosteoarthritis the fourth leading cause of disability by the year 2020.1,2
Chronic inflammation leads to cancer development and in recent times experimental andclinical studies have supported this hypothesis, which is now globally accepted. The epide-miological studies have identified chronic infections and inflammation as major risk factorsfor various types of cancer. It has been estimated that the underlying infections and in-flammatory reactions are linked to 15–20% of all cancer deaths.3 Further studies havesuggested that prolonged use of nonsteroidal anti-inflammatory drugs (NSAIDs), such asaspirin or of selective cyclooxygenase (COX)-2 inhibitors, delays the development of pre-malignant tumors and reduces the incidence of different forms of neoplasia. The cancerassociated inflammation includes the presence of several mediators such as cytokines, che-mokines, growth factors, lipid messengers, matrix degrading enzymes, and leukocytes in-filtrating the tumor microenviornment.4
Several classes of drugs, such as corticosteroids, NSAIDs, and biologics, are used to treatthe inflammatory disorders. All these drugs possess several adverse effects and biologics areexpensive to be used. Corticosteroids have long been used for the management of rheumatoidarthritis, but they suffer from some serious adverse effects, such as Cushing’s habitus (ap-pearance with rounded face, narrow mouth, supraclavicular hump, obesity of the trunk withrelatively thin limbs), hypertension, hyperglycemia, muscular weakness, increased suscept-ibility to infection, osteoporosis, glaucoma, psychiatric disturbances, growth arrest, etc.5
Similarily, the side effects associated with the use of NSAIDs, such as gastrointestinal ul-ceration and bleeding, and platelet dysfunction, are due to the inhibition of COX-1-derivedprostanoids, whereas inhibition of COX-2-dependent prostaglandin (PG) biosynthesis ac-counts for the anti-inflammatory, analgesic, and antipyretic effects. Thus, the selective COX-2 inhibitors (coxibs) were developed with better therapeutic activity and safety profile, duringthe last decade. The coxibs also exhibited cardiovascular side effects due to inhibiton ofprostacyclin formation in the infarcted heart, tipping the balance of prostacyclin/throm-boxane, coupled with a diminution in prostacyclin in heart muscle.6 The use of biologicaltherapies represented the difficulty of predicting in vivo adverse responses based on in vitroor ex vivo assays. The patients who received biologic therapies are more likely at risk for thedevelopment of cardiac complications.7
The inflammation cascades are the source of molecular targets that when antagonized,block the out put of the cascade or pathway. The major anti-inflammatory targets (Fig. 1)include enzymes, COX-1, COX-2, inosine monophosphate dehydrogenase; cytokines andcytokine receptors, tumor necrosis factor (TNF)-a and TNF-RII, interleukin (IL)-1b and IL-1RA, IL-2 and IL-2R,8 interferon (IFN)-a2, IFN-b1, and IFN-g; G-protein-coupled re-ceptors, histamine 1 and cysteinyl leukotriene 1; nuclear hormone receptors; cell interactionmolecules such as leukocyte function-associated antigen (LFA)-1 and cluster of differentia-tion molecules (CD)-11a, LFA-3 and CD-2, very late activation antigen (VLA)-4 andCD49d, and cytotoxic T lymphocyte antigen-4 immunoglobulin (CTLA-4-Ig). Over the past20 years, there has been a significant increase in knowledge about immunology both in termsof molecular targets and molecular mechanisms. For example 339 leukocyte surface antigens,80 cytokines and their receptors, 20 chemokines and their G-protein coupled receptors havebeen characterized with potential roles in inflammation described.9,10 Apart from these thetranscription factor, nuclear factor (NF)-kB, mitogen-activated protein kinases (MAPKs),c-Jun-N-terminal kinase (JNK), and p38 kinases have also been described as molecular
2 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

targets of inflammation.11,12 NF-kB is involved in the regulation of expression of numerousgenes in setting the inflammatory response. Several in vitro and in vivo studies havedemonstrated the contribution of components of the NF-kB signaling pathways to thepathogenesis of rheumatic diseases such as osteoarthritis and rheumatoid arthritis.13
Natural products or natural product-derived compounds represent great structural di-versity, which is not commonly seen in synthetic compounds. Of the 1184 new chemicalentities reported during 01/1981 to 06/2006, 60% are derived from or based on naturalproducts. Thus, natural products play a dominant role in the discovery of leads for thedevelopment of drugs for treating human diseases.14 Natural products (and traditionalmedicines) offer great hope in the identification of bioactive compounds and their develop-ment into drugs for the treatment of inflammatory diseases. Plants have been the basis ofmany traditional medicine systems throughout the world for thousands of years and continueto provide mankind with new remedies. The plant-based medicines initially dispensed in theform of crude drugs such as tinctures, teas, poultices, powders, and other herbal formula-tions, now serve as the basis of novel drug discovery.15 There are several records in tradi-tional medicine focusing on relief from pain and inflammation. Since ancient times manypeople suffering from inflammation were treated with phytochemicals, which is evident fromthe discovery of first anti-inflammatory, analgesic drug aspirin. The discovery of aspirin wasbased on the known analgesic and antipyretic properties of the bark of willow-tree since 400BC by the Greeks and Romans. In 1899 acetylsalicylic acid (aspirin) was introduced as thefirst potent drug to treat rheumatic diseases.16
Several reviews have been published recently on natural anti-inflammatory compoundsbut they have been restricted to either a particular target like lipoxygenases (LOXs),17,18
arachidonic acid (AA) pathway,19 nitric oxide (NO),19 NF-kB,19,20 TNF-a,21 cytokines,chemokines, and adhesion molecules22 or a particular class of compounds like polyphenols.23
Figure 1. The various drug targets involved in the inflammation cascade.
ANTI-INFLAMMATORYNATURALPRODUCTS K 3
Medicinal Research Reviews DOI 10.1002/med

All of the above reviews have described either some of natural products for a particularinflammation target or a very few selected compounds acting on different targets. None ofthe reviews included compounds from marine or microbial sources.
We are engaged in discovering COX-2 inhibitors of natural, semi-synthetic, or syntheticorigin24–29 and recently published a review on current status of natural COX inhibitors,describing 90 compounds belonging to different chemical classes.6 The present review articleaims at describing anti-inflammatory compounds from plants and marine sources acting onvarious targets mentioned above. The literature is reviewed from January 2000 to March2008 and 241 references to 254 compounds that have shown significant activity are beingcited. These compounds represents a wide structural diversity as they belong to differentchemical classes such as alkaloids, steroids, terpenoids, polyphenolics, phenylpropanoids,fatty acids and lipids, and various miscellaneous compounds. The attempt is also being madeto enumerate the possible leads e.g. curcumin, resveratrol, baicalein, boswellic acid (BA),betulinic acid, ursolic acid, and oleanolic acid for further development with the help of SARstudies and their current status. Additionally SAR studies carried out on the anti-in-flammatory activity of flavonoid compounds and clinical studies performed on anti-in-flammatory natural products are also included in the article.
2. Alkaloids
Tryptanthrin (1), an indoloquinazoline alkaloid, isolated from Isatis tinctoria leaves, wasresponsible for COX-2 inhibitory activity of dichloromethane crude extract and was found tobe less potent than the preferential COX-2 inhibitors nimesulide (IC50 39 nM) and NS-398(IC50 2 nM)30–32 (Table I). Tryptanthrin inhibited NO, PGE2, and inducible Nitric Oxidesynthase (iNOS) expression in IFN-g and lipopolysaccharide (LPS)-stimulated RAW 264.7cells, whereas it had no effect on COX-2 protein level. It was revealed that the inhibitoryeffect on NO was mediated by inhibiting the iNOS expression.33 Tryptanthrin showedtherapeutic effects in dextran sodium sulfate-induced colitis in mice by reducing the colondamage.34 The tryptanthrin and three different extracts of I. tinctoria leaves were evaluatedfor anti-inflammatory efficacy upon topical application in healthy human volunteers. Thetreatment with extracts showed significant effects than tryptanthrin in sodium lauryl sulfate(SLS)-induced irritant contact dermatitis in the eliciting phase as compared to the untreatedvolunteers.35 Fruits of Evodia rutaecarpa have long been used for inflammatory disorders andcontain the active constituents such as rutaecarpine (2), dehydroevodiamine (3), and evo-diamine (4) (Fig. 2). Rutaecarpine36 and evodiamine are reported to inhibit PGE2 in LPS-treated RAW 264.7 cells, while evodiamine and dehydroevodiamine have down-regulated theCOX-2 expression by inhibiting NF-kB activation and rutaecarpine did not show any effect.The PGE2 inhibitory effects of evodiamine and dehydroevodiamine could be due to down-regulation of COX-2 expression.37,38 Compounds 2–4 suppressed NO as well as iNOS ex-pression in microglial cells.39 Rutaecarpine inhibited the COX-1 and COX-2 catalyzed PGD2
in bone marrow cells and COX-2-catalyzed PGE2 in HEK293 cells, whereas it neither in-hibited the COX-1 nor COX-2 proteins nor mRNA expression in bone marrow cells, in-dicating its inhibitory effect on COX-2 enzyme directly (Table I). Furthermore, rutaecarpineshowed in vivo anti-inflammatory activity in carrageenan-induced rat paw edema.40 Ru-taecarpine also exhibited the immunosuppressive activity through inhibition of cytokineproduction and cell cycle arrest in G01G1 phase.41 Evodiamine and rutaecarpine inhibitedTNF-a and IL-4 protein expressions in RBL-2H3 cells, revealing that these compounds maybe effective for allergic diseases such as atopic dermatitis and rhinitis.42 Two quinolonealkaloids 5 and 6, isolated from E. rutaecarpa exhibited inhibitory effect on leukotriene (LT)biosynthesis in human granulocytes and their effects were comparable to the positive control
4 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

zileuton (IC50 10.4 mM).43 Yarosh et al.44 have observed that a standardized extract ofE. rutaecarpa fruits exhibited a significant topical anti-inflammatory activity in methyl nicoti-nate-induced erythema in human skin. Sinomenine (7), isolated from a Chinese plant Sino-menium acutum, inhibited the proliferation of rat fibroblasts and lymphocytes stimulatedwith transforming growth factor (TGF)-b2 or IL-1b and NF-kB mediated expressions ofTNF-a and IL-1b proteins in adjuvant arthritis. Positive correlations were found betweenchanges of NF-kB activity and expression of TNF-a and IL-1b.45 The anti-arthritic activityof sinomenine might be due to the suppression of both T-helper cells (Th)1 and Th2 immuneresponses and TGF-b2 may at least in part contributes to these responses.46 Sinomenineattenuated 2,4,6-trinitrobenzene sulfonic acid-induced colitis in mice through the suppression
Table I. The Effect of Alkaloids on Various Mechanisms of Inflammation
Compounds Targets, cells IC50 (mM) Ref.
Tryptanthrin (1) COX-2, Mono Mac 6 64 nM 30
6-keto-PGF1a, RAW 264.7 and BAECs – 32
12-HHT, BAECs and bovine thrombocytes – 32
TXB2, HEL – 32
LTB4, human neutrophils – 32
NO and PGE2, RAW 264.7 – 33
Rutaecarpine (2) PGE2, RAW 264.7 1.4 36
NO and iNOS, microglial cells – 39
COX-1 and -2, BMMC 8.7, 0.28 40
COX-2, HEK293 – 40
TNF-a and IL-4, RBL-2H3 – 41
Dehydroevodiamine (3) PGE2, COX-2 gene and NF-kB, RAW 264.7 – 38
NO1 and iNOS1,2, microglial1 and RAW 264.72 – 38
Evodiamine (4) PGE2, RAW 264.7 3.7 37
NF-kB, RAW 264.7 1.6 37
NO and iNOS, microglial – 39
5 LT, human granulocyte 10.0 43
6 LT, human granulocyte 10.1 43
Sinomenine (7) PGE3 and LTC4, macrophages – 45
NO and TNF-a, macrophages – 45
Fangchinoline (8) IL-5, human 3.7 54
Tetrandrine (9) IL-5, human 5.7 54
IL-5, murine 9.9 54
NO and iNOS, human monocytes – 56
PGE2 and COX-2 gene, human monocytes – 56
JNK, ERK, AP-1 – 57
Berberine (10) TNF-a and IL-6, 3T3-L1 adipocytes cells – 60
NO and iNOS, RAW 264.7 – 61
IL-1b and TNF-a, A-549 epithelial cells – 64
IL-1b and TNF-a, HFL1 fibroblasts cells – 64
IL-1b and TNF-a, U-937 lymphoma cells – 64
13-methylberberine (11) NO, RAW 264.7 11.6 66
iNOS, TNF-a and COX-2 gene, RAW 264.7 – 66
13-ethylberberine (12) NO, RAW 264.7 9.3 66
iNOS, TNF-a and COX-2 gene, RAW 264.7 – 66
Piperlactam S (13) TNF-a1 and IL-1b2, RAW 264.7 4.31 and 8.72 67
16 iNOS1 and PGE22, RAW 264.7 2.01 and 0.392 70
17 iNOS1 and PGE22, RAW 264.7 2.81 and 1.392 70
ANTI-INFLAMMATORYNATURALPRODUCTS K 5
Medicinal Research Reviews DOI 10.1002/med

of up-regulated colonic TNF-a and IFN-g productions.47 In dendritic cells, sinomenineimpeded the IL-2 and IFN-g secretions in co-cultured CD41T cells and modulated theirdifferentiation, maturation, and functionality, showing its relevance to immunomodulatoryeffects in controlling specific immune responses in autoimmune diseases and transplanta-tions.48,49 It showed therapeutic intervention in multiple sclerosis through inhibition oflymphocyte proliferation, chemokines and NF-kB activation50,51 and inflammation-mediatedneurodegenerative diseases.52 Tang et al.53 synthesized some sinomenine analogs mainlymodified on the ring C, and evaluated their in vivo biological activity against xylene-inducedear edema in mice. But none of these analogs showed the profound activity, which couldfurther be used for lead optimization. Bisbenzyl isoquinoline alkaloids, fangchinoline (8) andtetrandrine (9), isolated from Stephania tetrandrae roots, exhibited anti-inflammatory effectson croton oil-induced mouse ear edema. Fangchinoline showed a potent inhibitory activityon human IL-5, while it had no effect on murine IL-5 (Table I).54 These alkaloids showedinhibitory effects on N-formyl-methionyl-leucyl-phenylalanine (fMLP)- and LTB4-inducedneutrophil adhesion and transmigration in human polymorphonuclear leukocytes (PMNL)55
and reduction in the levels of IL-1b and TNF-a in stimulated human peripheral blood
N
N
O
O
1
NH
N
N
O
NH
N
N
O
4
2
NH
N
N
O
3
N
O
R
CH3
5, R =
6, R =OCH3
O
HO
H3CO
N CH3
7
NOR
OCH3
O
H3CO
N
OOCH3
CH3H3C
8, R = H9, R = CH3
N
OO
OCH3
OCH3
R
10, R = H11, R = CH3
12, R = C2H5
NOCH3
HO
H3CO
O
13
14
NH
N
OCH3
R
16, R = OCH3
17, R = H
+H3C
H3C
H
H3CO
H3CO
N
+
N
O
H
H
O
H
H3CO
H3CO
N
N
O
H
H
O
H
15O
Figure 2. Chemical structures of the alkaloids.
6 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

mononuclear cells (PBMC).22 Tetrandrine is reported to inhibit NO, PGE2, proinflammatorycytokines (TNF-a, IL-4, and IL-8),55 and NF-kB DNA-binding activity induced by CD28co-stimulation, IkBa kinase (IKK) degradation, and activation of MAP kinases.57
Berberine (10), a protoberberine alkaloid of the genera Berberis and Coptis, effectivelyinhibited COX-2 transcriptional activity in human colon cancer cells58,59 but not in oralcancer cells. It inhibited the levels of TNF-a, IL-1b, IL-6, C-reactive protein, and hepa-toglobin.60,61 Berberine is reported to inhibit NO production and iNOS gene expression, butnot the enzyme activity.62 IL-12 plays a pivotal role in the development of Th-1 immuneresponses and berberine has been found to induce the IL-12 p40 production and p38 MAPkinase activation, suggesting its immunotherapeutic potential in induction of IL-12 pro-duction.63 Berberine also exhibited the suppressive effects on receptor activator of NF-kBligand (RANKL)-induced osteoclastogenesis by inhibiting NF-kB activation-mediatedphosphorylation at the activation loop of IkBa kinase b, phosphorylation and degradation ofIkBa, and NF-kB p65 nuclear translocation.64 Berberine showed in vivo anti-inflammatoryactivity against serotonin-induced hind paw edema and acetic acid-induced increase in vas-cular permeability.65 The berberine derivatives, 13-methylberberine (11) and 13-ethylber-berine (12) (Fig. 2), exhibited the inhibitory effect on iNOS, TNF-a, and COX-2 expressionsin RAW 264.7 cells. They increased the IL-12 production in LPS-treated macrophages andindicated the posttranscriptional regulatory mechanism of iNOS gene expression and in-hibition of COX-2 activity.66 Piperlactam S (13), isolated from Piper kadsura, suppressed thecomplement 5a-induced migration across a fibrinogen-coated barrier and elevated the levelsof TNF-a and IL-1b67 and also exerted anti-inflammatory effects on cell migration, F-actinpolymerization, filopodia formation, and/or CDc42 activation.68 Brucine (14) and brucine N-oxide (15) of Strychnos nux-vomica seeds showed significant anti-inflammatory activity incarrageenan-induced rat paw edema assay and reduced the contents of PGE2 in paws. Bothalkaloids exhibited a weak effect on peritoneal inflammation produced by acetic acid inmice.69 The b-carboline alkaloids 16 and 17, isolated from Melia azedarach, exhibited in-hibitory effect in citrulline formation assay for iNOS activity (Table I). They also inhibitedthe PGE2, COX-2 expression, and DNA binding of NF-kB.70
3. Fatty Acids
Four fatty acids viz. a-linolenic acid (18), docosahexaenoic acid (19), eicosapentaenoic acid(20), and 5-thia-8,11,14,17-eicosatetraenoic acid (21) (Fig. 3) have been reported to exhibitpotential inhibitory effects on COX-2 and COX-1-catalyzed prostaglandin biosynthesis. Allthe four compounds were found to be more potent inhibitors of the COX-2 enzyme ascompared to the COX-2 selective NS-398 (53 mM) (Table II). Compound 18 was less potentinhibitor than 19 and 20, but among the compounds tested, it was the most COX-2 selective,with a COX-2/COX-1 ratio of 0.1. Of the semi-synthetic fatty acids, 21 was the most potentCOX-2 inhibitor with a COX-2/COX-1 ratio of 0.2. It inhibited the COX-2 enzyme morethan its precursor, 20. When the chain lengths of fatty acids were compared, those with morethan 20 atoms were inhibitors and those with less than 16 atoms were inactive. Among thefatty acids with 18 atoms, some were active and some were not, which might correlate withless unsaturation or with an ether function. The active fatty acids with more than 18 atomsare probably accepted as a substrate by the enzymes.71 Su et al.72 reported that mixture offatty acids (linoleic, oleic, and stearic) showed significant activity against COX-2 (IC50
6.7 mM) and was inactive against COX-1 (IC504350 mM). The pure linoleic and oleic acidswere found to be active selectively against COX-2, while stearic acid was inactive. Two fattyacids, 13-(S)-hydroxy-9Z,11E-octadecadienoic acid [(S)-coriolic acid] (22) and (7)-glycerol-1-monolinolate of Hernandia ovigera exhibited potent inhibitory effect on COX-2 enzyme
ANTI-INFLAMMATORYNATURALPRODUCTS K 7
Medicinal Research Reviews DOI 10.1002/med

than the standard trans-resveratrol (IC50 0.30 mM).73 A novel fatty acid 23 derived fromlinoleic acid, markedly inhibited the expression of proinflammatory genes in LPS-stimulatedmacrophages via blockade of NF-kB and activator protein-1 (AP-1) pathways. At higherconcentrations, (7)-10-oxo-trans-11-octadecen-13-olide, (7)-13-hydroxy-10-oxo-trans-11-octadecenoic acid (13-HOA) and 9-HOA exhibited profound suppressive effects on the ex-pression of iNOS, COX-2, IL-6, and TNF-a. The ability of 13-HOA to attenuate the geneexpression of proinflammatory proteins such as COX-2, iNOS, TNF-a, and IL-6 makes it animportant lead for further optimization and development (Fig. 3).21
Fatty acid amide hydrolase (FAAH) is a hydrolytic enzyme that plays a central role inmodulating endogenous N-acylethanolamine (NAE) levels in vertebrates, and, in part,constitutes an ‘‘endocannabinoid’’ signaling pathway that regulates diverse physiological andbehavioral processes in animals. As the endocannabinoids produced by tissues during pa-thological conditions often have protective roles, FAAH inhibitors have been proposed astherapeutic drugs.74,75 NAEs are fatty acid amides that are derived from an N-acylatedphoshatidylethanolamine presursor, a minor membrane lipid constituent of plant and animalcells. NAEs have been recognized with anti-inflammatory activity in mammals for almost 50years. The various biological roles of this family of compounds began to be explored in more
COOH
COOH
COOH
S COOH
18
19
20
21
COOH
O OH23
COOH
OH22
HOH
OH
OOCH3
O
H
H
HOH
OOCH3
O
H
HHO
H
OH
OOH
O
H
HOH
26
27
OHHO
O
H
OH
29
O
H O
H
30 O
H
31H H
OHO
28OH
H H
H H
HN
O
OH24
HN
O
OH 25
Figure 3. Chemical structures of the fatty acids and steroids.
8 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

detail with the discovery that several NAEs and related lipids accumulated in damagedtissues. NAEs 12:0, 14:0, 16:0, 18:0, 18:1, 18:2, and 18:3 are the representative examples,NAE 16:0 (24) and NAE 18:2 (25) (Fig. 3) being the most abundant, reported in the seeds ofthe Leguminosae family, to name a few includes Bauhinia congesta, Caesalpinia gilliesii,Mimosa borealis, Arachis hypogaea, Medicago sativa, Pisum sativum, Vigna unguiculata,Glycine max, etc. NAE 16:0 acts as an anti-inflammatory and analgesic agent. It also appearsto modulate the endocannabinoid activity by down-regulating the FAAH enzyme. NAE 16:0has also been shown to accumulate in induced cerebral ischemia and to be neuroprotec-tive.76,77 Recently, peroxisome proliferator-activated receptor-a was identified as the mole-cular target responsible for the anti-inflammatory properties of NAE 16:0.76 NAE 18:2 hasbeen shown to similarly potentiate endocannabinoid activity by competing with endogenousligands for FAAH enzyme.78
4. Steroids
Three new ergostanes, methyl antcinate L (26), antcin M (27), and methyl antcinate K (25),isolated from Antrodia salmonea, exerted anti-oxidative effect on the production of reactiveoxygen species (ROS) and NO in neutrophils and microglial cells, respectively. All com-pounds significantly suppressed LPS- and IFN-g-induced NO production and were morepotent than L-NAME, a nonspecific NOS inhibitor (Table II).79 The anti-inflammatoryeffect of Commiphora mukul gum was studied in PBMC and bioassay-guided fractionationled to the isolation of guggulsterol (29). Guggulsterol showed an inhibitory effect on pro-liferative response of PBMC in mitogenic lymphocyte proliferation and mixed lymphocytereaction assays. Further studies on inflammatory mediators such as IFN-g, IL-12, TNF-a,IL-1b, NO, and p38 MAPKs showed down-regulation, whereas no inhibition was observedin the case of anti-inflammatory cytokine IL-10.80 Singh et al.81 conducted both preclinical
Table II. Anti-inflammatory Actions of Fatty Acids and Steroids
Compounds Targets, cells IC50 (mM) Ref.
Fatty acids
a-Linolenic acid (18) COX-2, free enzyme 121 and 1.92 711, 722
Docosahexaenoic acid (19) COX-2, free enzyme 9.8 71
Eicosapentaenoic acid (20) COX-2, free enzyme 7.1 71
5-thia-8,11,14,17-eicosatetra-enoic
acid (21)
COX-2, free enzyme 3.9 71
Linoleic acid COX-2, free enzyme 1.9 72
Oleic acid COX-2, free enzyme 0.7 72
(S)-coriolic acid (22) COX-2, free enzyme 0.14 73
(7)-glycerol 1-monolinolate COX-2, free enzyme 0.18 73
23 iNOS, COX-2, IL-6 and TNF-a – 21
24 FAAH, PPAR-a – 77,78
25 FAAH – 77
Steroids
Methyl antcinate L (26) NO, neutrophils1 and microglials2 2.71 and 1.72 79
Antcin M (27) NO, neutrophils1 and microglials2 16.51 and 12.12 79
Methyl antcinate K (28) NO, neutrophils1 and microglials2 2.51 and 0.92 79
Guggulsterol (29) IFN-g, IL-12, TNF-a, IL-1b and NO – 80
Guggulsterone (30) COX-2, MMP-9, NF-kB – 82
Neridienone A (31) ICAM-1, A549 7.0 83
ANTI-INFLAMMATORYNATURALPRODUCTS K 9
Medicinal Research Reviews DOI 10.1002/med

and clinical investigations of guggul in patients with osteoarthritis and observed a significantimprovement in patients for the reduction of pain, stiffness, improved function and toler-ability Therefore, guggul appears to be a relatively safe and effective supplement to reducethe symptoms of osteoarthritis. Another steroid, (E)-guggulsterone (30) (Fig. 3), of this plantinhibited NF-kB activation and down-regulated the expression of inflammatory gene pro-ducts such as COX-2 and matrix metalloproteinase (MMP)-9 (Table II), which are majorplayers in the development of arthritis. Guggulsterone also suppressed the osteoclastogenesisinduced by RANKL.82 A pregnane steroid neridienone A (31), obtained from Neriumoleander exhibited a potent inhibitory activity against intercellular adhesion molecule(ICAM)-1 in human cultured cell line A549; however, showed a weak inhibitory activityagainst cell growth of A549.83
5. Terpenoids
Four ent-akurane diterpenoids 32–3584 of Croton tonkinensis leaves and five ent-akuranediterpenoids 36–4085 (Fig. 4) of Isodon excisus aerial parts strongly inhibited the NF-kBactivation and NO production in LPS-stimulated RAW 264.7 cells. The inhibitory effects ofthese compounds were very much potent than the positive control, parthenolide (IC50 2.34and 2.01 mM, for NF-kB activation and NO production, respectively) and aminoguanidine(IC50 32.17 mM for NO production) (Table III). The potent NF-kB inhibitory activity ofthese compounds could be accounted by the presence of reactive centers, such as an exo-methylene group conjugated to a carbonyl group in the cyclopentanone ring. This functionalgroup can react with biological nucleophiles, especially the sulfhydryl group of the cysteineresidue in the DNA-binding domain of the NF-kB subunit.84,85 Taraxasteryl acetate (41)isolated from Pluchea sagittalis showed activity against H2O2-, phorbol 12-myristate 13-acetate (PMA)- and fMLP-induced ROS, sodium nitroprusside-induced reactive nitrogenspecies, and heat shock protein72 production in human neutrophils; and exhibited morepotent topical anti-inflammatory activity than indomethacin in AA-, croton oil- and PMA-induced mice ear edema.86 Agnuside (42), an iridoid glycoside, isolated from Vitex ped-uncularis showed a preferential inhibition of COX-2 greater than aspirin (IC50 0.058mg/mL),a nonselective COX-2 inhibitor.87 A new lanostane triterpene glycoside 43, isolated fromFomitopsis pinicola fruits exhibited more potent inhibitory effect on COX-2 enzyme thanindomethacin (IC50 0.60 mM) and a weak inhibitory effect on COX-1.88 Sugiol (44), a di-terpene from Calocedrus formosana bark was reported to inhibit ROS, pro-IL-1b, IL-1b, andTNF-a production and suppressed the activation of MAPKs, including extracellular signal-regulated kinase (ERK), JNK, and p38 in stimulated J774A.1 murine macrophages.89
Triptolide (45) and tripdiolide (46), diterpene epoxides isolated from Tripterigium wilfordii,down-regulated the COX-2, iNOS, and IL-1b gene expression in LPS-stimulated macro-phages.90 Clinical studies have been performed on the extract of T. wilfordii and substantialwork has been reported on triptolide and celastrol (47) (Fig. 4, Table III); the two majorbioactive compounds of this plant. Wu and Guo91 studied the clinical effects of triptolidetablets in patients with psoriasis vulgaris and observed 75% effective rate. Celastrol, a noveltriterpene showed promise as an anti-inflammatory agent in animal models of arthritis,lupus, amyotrophic lateral sclerosis, and Alzheimer’s disease.92 Celastrol inhibited the TNF-a-induced activation of IkBa kinase, phosphorylation, degradation; p65 nuclear transloca-tion and phosphorylation and NF-kB-mediated reporter gene expression and suggested thatcelastrol potentiates TNF-a-induced apoptosis and inhibits invasion through suppression ofthe NF-kB pathway.93 Tao et al.94 conducted a prospective, double-blind, placebo-controlledstudy to assess the efficacy and tolerability of ethanol/ethyl acetate extracts of T. wilfordii
10 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

roots in patients with rheumatoid arthritis and found the therapeutic benefits in 60% patientswith well tolerability at the tested doses.
Sesquiterpene lactones (SLs) are active constituents of medicinal plants belonging toAsteraceae family, which have been used for centuries to treat inflammation and arthritis infolk medicine.95 Parthenolide (48), a predominant SL found in the medicinal plant, feverfew(Tanacetum parthenium), is responsible for its anti-inflammatory effects.95 The IkB kinase b(IKK b) subunit is known to play a critical role in cytokine-mediated signaling. Parthenolidehas been shown to bind directly to and inhibit IKK b and mutation of cysteine179 in theactivation loop of IKK b abolished sensitivity toward parthenolide.96 Parthenolideshowed change in the gene expression profile in TNF-a stimulated human 293 cells using acDNA microarray analysis.97 Costunolide (49) and parthenolide, isolated from Magnolia
R1
R2
OHOR3
32, R1 = R2 = H, R3 = OH33, R1 = H, R2 = OAc, R3 = H34, R1 = OAc, R2 = H, R3 = OH35, R1 = H, R2 = OAc, R3 = OH
OHO
R1
AcO
R2
H
R3
36, R1 = OH, R2 = H, R3 = OH37, R1 = OH, R2 = H, R3 = OAc38, R1 = OH, R2 = = O, R3 = OAc39, R1 = H, R2 = R3 = OH40, R1 = R2 = OH, R3 = OAc
AcO 41
O
HO
H
HH
OCH2CO OH
HO OH
OH
OH42
43AcOH
OH
C
OH
OH
44
O
O
O
OH
H
R
O
O
45, R = H46, R = OH
O
HO
O
HO
COOH
H
47
OO
O48O
O49, R = H50, R = OH
O
O
O
H
HO51
H O
OOH
O
O
O
O
54
O
HO O
O
O
O
53HO
OH
OO52
R
O
OOHO
HOHO
Figure 4. Chemical structures of the terpenoids.
ANTI-INFLAMMATORYNATURALPRODUCTS K 11
Medicinal Research Reviews DOI 10.1002/med

grandiflora, inhibited NO production by down-regulating the iNOS expression, which in turnthrough the inhibition of IkB phosphorylation and degradation, essential for the NF-kBactivation. The inhibitory effects of costunolide were higher than that of parthenolide.98
Castro et al.99 reported that costunolide and 7-hydroxycostunolide (50) completely inhibitedthe transcription factor NF-kB at the concentration of 50 and 10 mM, respectively. Costu-nolide also showed anti-cancer activity by exerting antiproliferative effects in MCF-7, humanbreast cancer cells by interacting with microtubule proteins, and by inducing apoptosis inHL-60 cells, by suppressing Bcl-2 protein expression and by activating caspase-3.100 AnotherSL, helenalin (51) isolated from Arnica montana exhibited anti-inflammatory activity. LyXet al.101 reported that helenalin selectively inhibited the activation of the transcription factorNF-kB, but later on the same group described the mechanism of its action as the selectivemodification of the p65 subunit of the NF-kB, most probably by bifunctional alkylationof two specific cysteinyl residues in the DNA-binding region, thereby inhibiting DNA
Table III. The Effect of Terpenoids on Various Mechanisms of Inflammation
Compounds Targets, cells IC50 (mM) Ref.
32 NF-kB1 and NO2, RAW 264.7 0.111 and 0.262 84
33 NF-kB1 and NO2, RAW 264.7 0.101 and 0.212 84
34 NF-kB1 and NO2, RAW 264.7 0.421 and 0.472 84
35 NF-kB1 and NO2, RAW 264.7 0.071 and 0.152 84
36 NF-kB1 and NO2, RAW 264.7 0.261 and 0.672 85
37 NF-kB1 and NO2, RAW 264.7 0.241 and 0.482 85
38 NF-kB1 and NO2, RAW 264.7 0.231 and 0.692 85
39 NF-kB1 and NO2, RAW 264.7 0.401 and 0.632 85
40 NF-kB1 and NO2, RAW 264.7 0.441 and 0.942 85
Taraxasteryl acetate (41) ROS and RNS, human neutrophils – 86
Agnuside (42) COX-2, free enzyme 20 mg/mL 87
43 COX-2, free enzyme 0.15 88
Sugiol (44) pro-IL-1b, IL-1b and TNF-a,J774A.1 cells
– 89
Triptolide (45) COX-21, iNOS2 and IL-1b3,macrophages
0.031, 0.042
and 0.013
90
Tripdiolide (46) COX-21, iNOS2 and IL-1b3,macrophages
0.031,3, 0.052 90
Celastrol (47) IkBa, IKK, TAK-1, NF-kB – 93
Parthenolide (48) NO, RAW 264.7 0.56 98
PAF1 and fMLP2-induced human
neutrophils
4.161, 5.112 105
Costunolide (49) NO, RAW 264.7 0.43 98
NF-kB, EMSA 50� 99
7-hydroxy-costunolide (50) NF-kB, EMSA 10� 99
Helenalin (51) NF-kB, EMSA – 101,102
5-LOX1 and LTC42, Human platelets 91, 122 103
NF-kB, EMSA 10� 106
52 NO, J774 0.8 104
Molephantin (53) PAF1 and fMLP2-induced human
neutrophils
2.561, 2.922 105
54 PAF1 and fMLP2-induced human
neutrophils
2.901,2 105
NF-kB, EMSA 10� 106
�Concentration (mM), which completely inhibit the transcription factor NF-kB.
12 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

recognition and binding.102 Tornhamre et al.103 reported that helenalin also acts by inhibiting5-LOX and LCT4 synthase in human blood cells. A new SL 52, isolated from Laurus nobilisleaves, potently inhibited the NO release from LPS-activated murine macrophages with anIC50 value of 0.8 mM.104 Siedle et al.105 studied the effect of various SLs on the elastase releasein human neutrophils and on isolated elastase and found that molephantin (53) fromElephantopus mollis, 4b,15-Epoxy-miller-9E-enolide (54) from Milleria quinqueflora andparthenolide exhibited the highest inhibitory effect on the PAF- and fMLP-induced elastaserelease from the human neutrophils. Parthenolide and 4b,15-Epoxy-miller-9E-enolide alsocompletely inhibited NF-kB by selectively alkylating the p65 subunit probably by reactingwith cysteine residues.106
1. Oleanolic and ursolic acids and their analogsOleanolic acid (55) and ursolic acid (56) are ubiquitous triterpenoids in the plant kingdomand are an integral part of the human diet.107 Ursolic acid was isolated from Plantago majorfollowing bioactivity-directed fractionation using in vitro COX-2 catalyzed prostaglandinbiosynthesis assay. It showed a significant inhibitory effect on COX-2 enzyme with COX-2/COX-1 selectivity ratio of 0.6. The structural isomer, oleanolic acid, was less active thanursolic acid but showed a similar activity ratio (0.8) (Table IV).6 Ursolic acid also inhibitedCOX-2 in PMA-treated human mammary and oral epithelial cells.108 The in vivo anti-inflammatory activity of ursolic acid was found to be greater than oleanolic acid and
Table IV. The Inhibiory Effects of Oleanolic Acid and Ursolic Acid Analogues
Compounds Targets, cells IC50 (mM) Ref.
Oleanolic acid (55) COX-2, free enzyme 130 6
NO, mouse macrophages 401, 4402,3,4 1131, 1142, 1153, 1164
Ursolic acid (56) COX-2, free enzyme 295 6
NO, mouse macrophages a, b 1131, 1162, 1173
Oleanonic acid (57) LTB4, rat peritoneal
leukocytes
17 112
NO, mouse macrophages 37 117
58 NO, mouse macrophages 0.91, 0.172 1131, 1142
59 NO, mouse macrophages 1.8 113
CDDO (60) NO, mouse macrophages 0.41, 0.22,3,
0.444 nM
1141, 1152, 1163, 1184
61 NO, mouse macrophages 0.6 114
62 NO, mouse macrophages 0.81,2, 9.553 nM 1151, 1162, 1183
CDDO methyl
ester (63)
NO, mouse macrophages 0.11,2, 0.113 nM 1151, 1162, 1183
64 NO, mouse macrophages 20 nM 115
65 NO, mouse macrophages 40 nM 115
66 NO, mouse macrophages 70 nM 115
67 NO, mouse macrophages 70 nM 115
68 NO, mouse macrophages 5.6 117
69 NO, mouse macrophages 70 nM 117
70 NO, mouse macrophages 3.5 pM 118
71 NO, mouse macrophages 70 pM 118
72 NO, mouse macrophages 98 pM 118
73 NO, mouse macrophages 14 pM 118
aStrongly toxic to primarymousemacrophages (toxic above 5--10mM).
bToxic to cells above10mMandwas not active below10mM.
ANTI-INFLAMMATORYNATURALPRODUCTS K 13
Medicinal Research Reviews DOI 10.1002/med

indomethacin109,110 and oleanolic acid exhibited inhibitory effects on carageenan-induced ratpaw edema111 and formalin-induced arthritis. Oleanonic acid (57), a 3-oxo-analog of olea-nolic acid from Pistacia terebinthus galls, showed inhibitory activity against inflammationinduced by 12-deoxyphorbol-13-phenylacetate (DPP), 12-O-tetradecanoylphorbol-13-acetate(TPA), bradykinin and phospholipase (PL) A2. It inhibited the LTB4 production in ratperitoneal leukocytes with an IC50 value of 17 mM.112
Honda et al.113–118 have synthesized a variety of oleanane and ursane type triterpeneanalogs (Fig. 5) and studied their effects on NO production induced by IFN-g in mousemacrophages. Among the new enone derivatives, molecule 58 showed the highest activityfollowed by 59.113 The derivatives with electron-withdrawing groups at C-2 position in ringA, an analog 2-cyano-3,12-dioxoolean-1,9(11)-dien-28-oic acid (CDDO, 60) showed thehighest activity (IC50 0.4 nM)114; among the oleanane triterpenes with modified ring A and C,methyl 2-carboxy-3,12-dioxooleana-l,9(11)-dien-28-oate (62) showed similar activity as thatof CDDO and CDDO methyl ester (63), followed by 64–67 (Table IV).115 Among the novel
HOH
H
COOH
55HO
H
H
COOH
56 H
H
COOH
O57
H
COOH
O
O
58 59H
COOH
O
O
H H
COOH
O
R
61, R = CN68, R = H69, R = CO2H
H
H
CO2R2
O
O
R1
60, R1 = CN, R2 = H 62, R1 = CO2H, R2 = Me 63, R1 = CN, R2 = Me
H
COOR
O
NC
O
H
64, R = Me 65, R = H H
COOH
O
NC
O
H
66
H
COOH
O
HOOC
H
67
H
R2
O
O
R1
70, R1 = R2 = CN 72, R1 = CN, R2 = CO-D-Glu(OAc)4
73, R1 = CN, R2 = CONH2
HO
O
NCC ON
N71
Figure 5. Chemical structures ofoleanolic acid, ursolic acid, and their analogs.
14 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

olean- and urs-1-en-3-one derivatives with nitrile and carboxyl groups at C-2 in ring A andwith 9(11)-en-12-one and 12-en-11-one functionalities in ring C, compounds 60, 62, and 63
showed extremely high potency (IC50 0.1 nM level) and this combination increased the po-tency by about 10,000 times as compared with the lead compound 68 (IC50 5.6 mM).116 Thestudy involving the synthesis of novel olean- and urs-12-ene triterpenes with a 1-en-3-onefunctionality in ring A, 3-oxooleana-1,12-diene derivatives with carboxyl, methoxycarbonyl,and nitrile groups showed higher activity than the lead compound 68, whereas 69 exhibitedthe highest activity (IC50 70 nM).117 Among the effects of various groups at C-17 position ofCDDO and CDDO methyl ester, an analog 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oni-trile (70) showed extremely high inhibitory activity (IC50 1 pM level) followed by compounds71–73118 (Table IV) and were about 100 times and 30 times more potent than that of CDDOand dexamethasone, respectively. The most potent compounds exhibited potency similar todexamethasone and hydrocortisone, although they do not act through the glucocorticoidreceptor. The further developments revealed that CDDO has completed Phase I clinical trialstudy by NCI in July 2008 on 40 patients for the treatment of metastatic or unresectable solidtumors.119
Based on several studies on oleanolic acid and ursolic acid analogs, carried out by Hondaet al., a concise and informative SAR (Fig. 5) can be formulated as: (1) the oleanane skeletonis more active than the ursane. (2) Modifications in ring A; (i) although 1-en-3-one structuralunit without a substituent is important for activity, a methoxycarbonyl, carboxyl, and nitrilegroups at C-2 enhanced the activity. (ii) Hydroxyl, aminocarbonyl, methoxy, chloride, andbromide groups decreased the activity. (iii) A formyl group did not show activity, onlytoxicity.113–115 In conclusion, an electron-withdrawing group at C-2 increase the potency,whereas electron-donating group decreases. (3) Modifications in ring C; (i) a carbonyl groupat C-11 and/or C-12 is important. An insertion of a double bond at the cis-trans position ofC-11 and/or C-12 ketone enhanced the activity. (ii) Bis-enones with 1-en-3-one and 9-en-12-one structural units showed the highest activity. The introduction of 12-en-11-one and 13-en-11-one functionalities also enhanced the activity. (iii) The nitrile and carboxyl groups at C-2in bis-enones provided extremely active compounds. (iv) The saturated ring C, 11-13(18)-diene, and 9,11-epoxide were less potent than the original 12-ene. (v) The amide and formylgroups at C-2 did not enhance potency. (vi) 23,24-dimethyl groups are important for sig-nificant activity.115–117 (4) Modifications at position C-17; (i) A nitrile group enhanced thepotency, whereas an ester moiety decreased. (ii) Acetoxymethyl, hydroxymethyl, and formylgroups decreased the potency. (iii) An amide group decreased the potency, and the less polarthe amide, the less is its potency. (iv) A carbonyl imidazole moiety was more reactive than theother moieties with a nucleophile. (v) The role of methoxycarbonyl and carboxyl groups atC-17 is ambiguous. In some analogs, the carboxyl group is more potent than the methox-ycarbonyl group, while in others the carboxyl and methoxycarbonyl groups showed similarpotency.116–118
2. Betulinic acid and its analogsBetulinic acid (74), a pentacyclic lupane-type triterpene is widely distributed throughout theplant kingdom. A number of biological activities have been reported for betulinic acid viz.anti-cancer, anti-HIV, and antimalarial. Betulinic acid and its derivatives have especiallyshowed anti-HIV and cytotoxicity against a variety of tumor cell lines comparable to someclinically used drugs. However, betulinic acid also displayed considerable anti-inflammatoryactivity in various in vitro and in vivo model systems.120–122 Betulinic acid inhibited PLA2,
123
bovine PG synthase120 and suppressed the NF-kB activation induced by TNF-a, PMA,cigarette smoke, okadaic acid, IL-1, and H2O2. The NF-kB-dependent gene expressions ofCOX-2 and MMP-9 were also suppressed.124 It exhibited the selectivity toward inhibition of
ANTI-INFLAMMATORYNATURALPRODUCTS K 15
Medicinal Research Reviews DOI 10.1002/med

COX-2 as compared with COX-1.72 Betulinic acid also reduced the level of cytokine-inducedneutrophil chemoattractant-1 in stimulated rat macrophages, IL-1b-stimulated rat fibroblastcells,120 NO in stimulated RAW 264.7 cells.125 In various in vivo models, betulinic acid wasfound to be effective against skin inflammation and ear edema induced by mezerein, 12-deoxyphorbol-13-tetradecanoate (DPT), DPP, bradykinin, or TPA in mice.126
Honda et al.127 synthesized betulinic acid analogs (Fig. 6) for NO inhibitory activity inIFN-g-stimulated RAW cells. The analogs 75–84 with cyanoenone functionality in ring Awere highly active, with a potency similar to that of CDDO (IC50 0.02 mM), whereas be-tulinic acid was found to be inactive. The analogs 85–90 with carboxyl or methoxycarbonylenone functionality in ring A were less active. Most importantly, analog 82 was found to besignificantly more potent in vivo than both betulinic acid and CDDO. It caused a significantinduction of the anti-inflammatory and cytoprotective enzyme, heme oxygenase-1. The SARstudy revealed that for NO inhibition: cyano enone functionality in ring A is necessary, anacyl imidazole increased the in vivo potency and enone functionality in ring C is not ne-cessary. Modifications at C-17 did not affect the potency in the RAW cell assay.
3. Boswellic acidsBoswellia species have been traditionally used in folk medicine to treat various chronicinflammatory diseases. The experimental data from animal models and studies with humansubjects confirmed the potential of Boswellia species extracts for the treatment of not only
HOH
H
H COOH
74
OH
H
H R
75, R = CO2Me (IC50 0.03 µM)127
76, R = CO2H (IC50 0.2 µM)127
77, R = CO2Et (IC50 0.02 µM)127
78, R = CONH2 (IC50 0.03 µM)127
79, R = CONHMe (IC50 0.05 µM)127
80, R = CONHEt (IC50 0.07 µM)127
81, R = CONMe2 (IC50 0.03 µM)127
OH
H
H
N
O
NR
82, R = H (IC50 0.03 µM)127
83, R = Me (IC50 0.05 µM)127
84, R = Et (IC50 0.03 µM)127
OH
H
H CO2Me
R2
85, R1 = CO2Me, R2 = H (IC50 0.6 µM)127
86, R1 = CO2Me, R2 = OH (IC50 1 µM)127
87, R1 = CO2Me, R2 = F (IC50 0.3 µM)127
88, R1 = CO2H, R2 = H (IC50 0.3 µM)127
89, R1 = CO2H, R2 = OH (IC50 0.9 µM)127
90, R1 = CO2H, R2 = F (IC50 0.3 µM)127
NC
NC
R1
Figure 6. Chemical structures of betulinic acid and its analogs.
16 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

inflammation but also of cancer. Analysis of the constituents of the extracts revealed thatpentacyclic triterpenes, BAs possess biological activities and appear to be responsible for therespective pharmacological actions. BAs were assumed to be the anti-inflammatory principlesof Boswellia species. Extensive research in the past 30 years identified the active componentsof the extracts as BA (91) and its derivatives [3-O-acetyl-b-boswellic acid (92), 11-keto-b-boswellic acid (93), and 3-O-acetyl-11-keto-b-boswellic acid (AKBA, 94)]82,128,129 (Fig. 7).Initially, it was reported that BAs inhibit LT biosynthesis and 5-LOX enzyme, whereassuppression of PG synthesis and inhibition of COXs was excluded. Subsequent studiesconfirmed AKBA as the most active constituent and inhibited LT biosynthesis. Accordingly,AKBA was proposed as a novel specific nonredox-type inhibitor of 5-LOX.130 The SARstudy by Sailer et al.131 revealed that a pentacyclic triterpene ring system is crucial for bindingto the highly selective effector site, whereas functional groups (especially the 11-keto functionwith a hydrophilic group at C4 of ring A) are essential for 5-LOX inhibitory activity. Theremoval of the acetoxy group or reduction of the carboxyl function to alcohol slightlydiminished the potency of AKBA. The AKBA potently suppressed the formation of 12-LOXproducts in intact human platelets with higher potency for p12-LOX in cell-free assays ascompared to crude 5-LOX.132 Recently, Siemoneit et al.133 reported that AKBA potentlyinhibits the activity of COX-1 enzyme in intact human platelets as well as in cell free assays,whereas COX-2 enzyme was inhibited less efficiently.
In animal models of inflammation, BA was reported to be an effective adjuvant miti-gating bovine serum albumin-induced arthritis and osteoarthritis. The anti-arthritic potentialof BAs is a result of its anti-inflammatory activity mediated through inhibition of NF-kB,COX-2, and 5-LOX.73 A preclinical study carried by Singh et al.134 revealed the synergisticeffect between BAs mixture and glucosamine for anti-inflammatory activity in acute carra-geenan-induced rat paw edema and anti-arthritic activity inMycobacterium-induced arthritis.In a randomized, double-blind, placebo-controlled crossover study, Kimmatkar et al.135
studied safety, efficacy, and tolerability of a standardized Boswellia extract (WokVelTM,Pharmanza, India) containing 40% total BAs in patients with osteoarthritis in the knee andobserved the significant decreased knee pain, increased knee flexion, increased walking dis-tance, and ability to climb stairs. The symptoms returned after withdrawal of treatment. Inulcerative colitis patients, the alcoholic extract of B. serrata oleogum resin showed significant
HOHHOOC
H
H
91O
HHOOC
H
H
H3C
O
92
HOHHOOC
H
H
93
O
OHHOOC
H
H
H3C
O
O
94
Figure 7. Chemical structures of the boswellic acids.
ANTI-INFLAMMATORYNATURALPRODUCTS K 17
Medicinal Research Reviews DOI 10.1002/med

improvement and the remission rate of the treatment (82%) was better than the sulphasa-lazine (75%).129 The similar clinical effects were also observed by Gupta et al.136 in a studyincluding patients of chronic colitis.
6. POLYPHENOLICS
A. Flavonoids
Flavonoids, a major class of polyphenolic compounds widely distributed throughout theplant kingdom, possess interesting anti-inflammatory actions. Luteolin (95) and quercetin(96) inhibited the LPS-stimulated TNF-a and IL-6 release in RAW 264.7 cells. Luteolininhibited the in vivo TNF-a production, reduced the ear edema (induced by PMA andoxazolone) and ICAM-1 expression in LPS-treated mice.22 Luteolin, kaempferol (97), andquercetin exhibited a significant inhibitory effect on MMP-9 and MMP-2.137 Luteolin alsoinhibited the protein tyrosine phosphorylation, NF-kB-mediated gene expression, and pro-inflammatory cytokine production in murine macrophages.21 Luteolin significantly sup-pressed the rat paw edema induced by carrageenan and a similar tendency was observed inthe cotton pellet granuloma test. It markedly reduced the number of infiltered leukocytes andthe level of 6-keto-PGF1a in the exudates in air pouch test. The results of RT-PCR studyrevealed that luteolin is a selective inhibitor of COX-2 and down-regulated the mRNAexpression of COX-2.138 Baicalin (98) and wogonin (99) (Fig. 8) were isolated from Scu-tellaria baicalensis roots. Baicalin was found to reduce the fMLP- and PMA-induced accu-mulation of reactive oxygen intermediates and surface expression of the integrin MAC-1(CD11b/CD18) and MAC-1-dependent neutrophil adhesion in human neutrophils andmonocytes.22 Wogonin is reported to inhibit the NO and PGE2 production and induction ofiNOS and COX-2 in LPS-activated RAW cells.6 Jang et al.139 synthesized some wogoninanalogs and evaluated their activity against COX-2 catalyzed PGE2 production. Most of theanalogs were less active than wogonin. The alkylation of 5- and 7-hydroxyl groups andsubstitution on ring B of wogonin caused a reduction in the activity. An isoflavonoid, gen-istein (100) is reported to inhibit the TPA-stimulated PGE2 production in rat peritonealmacrophages140 and LPS-stimulated TNF-a and IL-6 release in RAW 264.7 cells, productionof thromboxane (TX) and LT, and release of IL-5.22,141,142 Among the flavanones, nar-ingenin (101) was reported to exhibit a moderate TNF-a inhibitory activity, whereas erio-dictyol (102) and hesperetin inhibited the TNF-a release.21 Panduratin A (104), a chalconeisolated from Kaempferia pandurata, strongly inhibited the NO and PGE2 production fromLPS-stimulated RAW 264.7 cells and suppressed the expression of iNOS and COX-2 enzymewithout any appreciable cytotoxic effect. It also suppressed the NF-kB-mediated phos-phorylation and degradation of IkBk and inhibited the transcriptional activity of NF-kB.Panduratin A exhibited better anti-inflammatory activity in TPA-induced ear edema assaythan diclofenac.143,144 Gemichalcone A (105) and B (106), isolated from Hypericum gemi-niflorum, inhibited the release of b-glucuronidase and histamine, and b-glucuronidase andlysozyme from compound 48/80 and fMLP-stimulated mast cells of rats.145 Apigenin (107)inhibited the production of NO and PGE2 by suppressing the expression of iNOS and COX-2proteins, respectively, in BV-2 cells. Moreover, apigenin suppressed p38 MAPK, JNKphosphorylation without affecting the activity of ERK.146 Apigenin profoundly reduced theTNF-a-induced adhesion of monocytes to human umbilical vein endothelial cells (HUVEC)monolayer, up-regulation of vascular cell adhesion molecule (VCAM)-1, ICAM-1, andE-selectin-mRNA to the basal levels. Apigenin also inhibited the production ofpro-inflammatory cytokines IL-1b, IL-8, and TNF-a; and transcriptional activity of NF-kBin LPS-stimulated human monocytes and mouse macrophages. These findings suggested that
18 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

apigenin may be useful for therapeutic management of inflammatory diseases.147,148 Theeffect of various other flavonoids like diosmetin (108), tetra-O-methyl luteolin (109), hexa-O-methyl myricetin (110), fisetin (111), and pilloin (112) are described at the cellular mechan-isms of inflammation149 (Table V).
1. SAR of flavonoidsIn the several studies,150–155 the SARs of flavonoids have been discussed using differenttargets of inflammation. The analysis of various studies helped us to formulate an in-formative SAR: (1) A planar ring system is essential in the flavonoid molecules to exhibit theactivity. (2) The presence of a double bond between C2 and C3, and hydroxyl groups at5- and 7- position of A-ring are necessary. (3) The lack of hydroxyl groups on B-ring virtuallyabolished the inhibitory actions. (4) The flavones and flavonols having hydroxyl group at40- position of B-ring showed higher activity than those lacking the hydroxyl group onB-ring. (5) The flavonols having hydroxyl groups at 30- and 40-positions (catechol type)exhibited higher activity than those having 30,40,50-trihydroxyl groups (pyrogallol type). (6)The presence of a methoxyl group at 8-position in A-ring and hydroxyl groups at 40- or30,40-positions (ortho) in B-ring affected inhibitory activity favorably, while hydroxyl groupsat 20 and 40-positions (meta) abolished the activity. (7) The methylation of the hydroxylgroups at 3, 5, or 40-positions enhanced the activity. (8) The methylation of the 3-hydroxylgroup reduced the cytotoxicity. (9) Flavones exhibited higher activities than the corre-sponding isoflavones, flavonols, and flavanones. (10) The introduction of a glycoside moietyreduced the activity. The possible reason could be that flavonoid glycosides may not pene-trate the cell membrane due to their hydrophilicity, or there might be steric hindrance due tobulky glycosyl residues.
In a study, Yamaki et al.140 tested the effect of isoflavones and their glycosides on PGE2
production in TPA-stimulated rat peritoneal macrophages and observed that the isoflavones113–116 were more active as compared to their corresponding glycosides 117–121 (Fig. 8),suggesting that 6-methoxylation and 5-hydroxylation increases the potency, whereas 7-O-glycosylation decreases. Sophoricoside (122), an isoflavonone glycoside isolated from So-phora japonica and its derivatives were tested for inhibitory activity against IL-5. Among thetested derivatives, 5-benzyloxy-3-(4-hydroxyphenyl) chromen-4-one (123) showed the mostpotent activity comparable to sophoricoside. The important structural requirements of theseisoflavonone analogs exhibiting the inhibitory activity against IL-5 were recognized as (1)planarity of chromen-4-one ring, (2) existence of hydroxyl at 4-position of B-ring, and (3)introduction of benzyloxy at 5-position, which may act as a bulky group for occupyinghydrophobic pocket in putative binding site. However, the glucopyranosyl moiety of so-phoricoside is not an essential motif for the activity.156 Sadik et al.157 studied the effects offlavonoids on rabbit reticulocyte 15-LOX-1 and soybean LOX-1 using linoleic acid as asubstrate and found that: (1) the presence of a hydroxyl group at 3-position of A-ring provedto be not essential but rather interfering. (2) The substitution of a catechol moiety in either A-ring or B-ring increased the inhibitory potency but was not essential for it. (3) In the presenceof a catechol moiety, the inhibitory potency inversely correlates with the number of hydroxylgroups. Martinez et al.158 found that isoflavones and isoflavanones were selective inhibitorsof 12-hLOX, whereas isoflavans were selective inhibitors of 15-hLOX-1 and a few flavonoidstarget 15-hLOX-2. Ishikawa et al.159 investigated the inhibitory effects of flavonoids on LPS-induced PGE2 production and found that reduction of the double bond between C2 and C3
resulted in a loss of activity; presence of hydroxyl groups at 5- and 7-positions of A-ringcorrelates with higher activity than those flavonoids having hydroxyl group at 7-positiononly or without hydroxyl groups at 5- and 7-positions. The isoflavones with hydroxyl groupat 5-position are more effective than those without hydroxyl group at 5-position. The fla-
ANTI-INFLAMMATORYNATURALPRODUCTS K 19
Medicinal Research Reviews DOI 10.1002/med

vones and flavonols without hydroxyl group on B-ring are more active than those having at30- and 40-positions.
Chen et al.160 synthesized C–C biflavones, which have flavone–flavone subunit linkagesat A ring–A ring, A ring–B ring, B ring–B ring, and B ring–C ring and investigated their
O
OR1
R2
R3
R4
95, R1 = R2 = R3 = R4 = OH109, R1 = R2 = R3 = R4 = OCH3
O
OOH
HO
OH
R
OH
96, R = OH97, R = H
O
OOH
O
HO
98
O
OOH
HO
OCH3
99
O
OOH
HO
OH100
O
OOH
HO
R2
R1
101, R1 = H, R2 = OH102, R1 = R2 = OH103, R1 = OH, R2 = OCH3
O
OH
OH
H3CO
104
O
R
HO
O
CH3
HO
OH O
OH
105, R = OCH3
106, R = H
O
OOH
R1
107R1 = R3 = OH, R2 = H108, R1 = R2 = OH, R3 = OCH3
112, R1 = R3 = OCH3, R2 = OH
R2
R3
O
OOCH3
OCH3
OCH3
H3COOCH3
OCH3
110
O
O
OH
OH
HO
OH
111O
OR4
R3
R1
R2
113, R1 = R3 = R4 = OH, R2 = OCH3
114, R1 = R2 = H, R3 = R4 = OH115, R1 = H, R2 = OCH3, R3 = R4 = OH116, R1 = R3 = OH, R2 = R4 = OCH3
O
OR3
O
R1
R2
117, R1 = R3 = OH, R2 = OCH3
118, R1 = R2 = H, R3 = OH119, R1 = H, R2 = OCH3, R3 = OH121, R1 = R3 = OH, R2 = H
H
OHO
HO
OH
OH
OHOHO
OH
OH
O
Figure 8. Chemical structures of the flavonoids.
20 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

inhibitory activity against PLA2. The biflavone 124 (having 40-40-linkage) showed inhibitoryactivity comparable to a natural biflavonoid, ochnaflavone (125, IC50 3.5 mM), and seven-fold stronger activity than amentoflavone (126, IC50 23.8 mM). The biflavones 127 and 128
(Fig. 8) showed inhibitory activities similar to amentoflavone. Agli et al.161 studied the effectof (7)-gallocatechin-3-gallate (GCG, 129), selectively deprived of hydroxyl groups, onMMP-9 activity, transcription, and secretion; and observed that (7)-GCG and (7)-ca-techin-3-gallate (130) exhibited the direct inhibition of MMP-9 activity. Conversely, whentranscription was the target, (7)-trans-3-flavanol-3-benzoate (131, IC50 4.2 mM), lacking allthe hydroxyl groups, was most effective in lowering MMP-9 promoter activity, secretion, andNF-kB-mediated transcription. Sartor et al.162 studied the effects of various flavonoid onleukocyte release and gelatinase inhibitory activity and observed the presence of the galloylgroup, number and position of hydroxyl groups, and stereochemistry of the C ring in theflavan skeleton as the essential features.
O
O
O
O
124
O
OH
O
O
OH
OH
HO
O
O
125
O
OHOH
O
OH
HO
OH
O
HO
O
126
O
O
O
O127
O
O
O
O
128
O
O
R1
R2
R3
R4
R5
R6
R7
R8
O
129, R1 = R2 = R3 = R4 = R5 = R6 = R7= R8 = OH130, R3 = H, R1 = R2= R4 = R5 = R6 = R7 = R8 = OH131, R1 = R2 = R3 = R4 = R5 = R6 = R7= R8 = H
OH
O
OOR3
R2
OR1
122, R1 = H, R2 = OH, R3 = Glc123, R1 = benzyl, R2 = R3 = H
O
OOCH3
O
OH
H3CO
120
OHOHO
OH
O
O
HOHO
HO
Figure 8. Continued
ANTI-INFLAMMATORYNATURALPRODUCTS K 21
Medicinal Research Reviews DOI 10.1002/med

Table V. The Inhibiory Effects of Flavonoids on Various Inflammatory Targets
Compounds Targets, cells IC50 (mM) Ref.
Luteolin (95) TNF-a, RAW 264.71, RBL-2H32 o11, 5.82 221, 1522
MMP-21 and -92, free enzyme 8.61, 8.42 137
NO, RAW 264.7 271, 202 1501, 1512
IL-41 and b-hexosaminidase2,
RBL-2H3
3.71, 32 152
15-LOX-1, rabbit reticulocytes 0.6 157
LOX L1, soyabean 3 157
Quercetin (96) TNF-a, RAW 264.71,2 and
RBL-2H33o51, 12, 5.63 211, 222, 1523
MMP-21 and -92, free enzyme 11.71, 12.42 137
NO, RAW 264.7 36 151
IL-41 and b-hexosaminidase2,
RBL-2H3
7.51, 4.62 152
15-LOX-1, rabbit reticulocytes 4.0 157
LOX L1, soyabean 4 157
PGE2, rat macrophages 13.9 159
Kaempferol (97) MMP-21 and -92, free enzyme 131, 15.22 137
NO, RAW 264.7 29 151
b-hexosaminidase, RBL-2H3 14 152
15-LOX-1, rabbit reticulocytes 15 157
LOX L1, soyabean 50 157
PGE2, rat macrophages 13.1 159
Baicalin (98) ROS, MAC-1, CD11b/CD18,
neutrophils and monocytes
– 22
NO, iNOS and TNF-a, RAW 264.7 – 22
Wogonin (99) NO1,2 and iNOS, RAW 264.7 311, 172 61, 1502
PGE21 and iCOX-2, RAW 264.7 0.31 6
IL-6 and -8, human epithelial cells – 22
b-hexosaminidase, RBL-2H3 29 152
Genistein (100) TNF-a, J774.11 and RAW 264.72 52 21, 222
PGE2, macrophages 15 140
5-LOX 80 141
NO, RAW 264.7 26 151
15-LOX-1, rabbit reticulocytes 18 157
PGE2, rat macrophages 7.2 159
Naringenin (101) TNF-a, J774.1 – 21
PGE2, rat macrophages 7.9 159
Eriodictyol (102) TNF-a, J774.1 – 21
PGE2, rat macrophages 7.2 159
Hesperetin (103) TNF-a, J774.1 – 21
PGE2, rat macrophages 8.4 159
Panduratin A (104) NO1 and iNOS, RAW 264.7 0.1751 6
PGE21 and iCOX-2, RAW 264.7 0.0191 6
Gemichalcone A (105) b-glucuronidase and histamine, mast 145
b-glucuronidase and lysozyme, mast 145
Gemichalcone B (106) b-glucuronidase and histamine, mast 145
b-glucuronidase and lysozyme, mast 145
Apigenin (107) MMP-21 and -92, free enzyme 7.51, 12.92 137
NO, RAW 264.7 231, 7.72 1501, 1512
IL-41 and TNF-a2, RBL-2H3 3.61, 5.32 152
22 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

2. Baicalein and its analogsBaicalein (132) inhibited the LPS-induced IL-12 production in macrophages and RAW 264.7cells. It also inhibited the activation of IL-12 gene promoter in transfected RAW 264.7 cellswith IL-12 gene promoter/luciferase constructs.163 Deschamps et al.164 reported that baica-lein is not selective against 12-hLOX vs. human reticulocyte 15-LOX-1 (15-hLOX-1) (15-hLOX-1/12-hLOX5 1.3). However, in the presence of detergents (Triton X-100), the se-lectivity of baicalein increased toward 12-LOX (15-hLOX-1/12-hLOX5 7). The molecularmodeling study showed that the mechanism of inhibition of 15-hLOX-1 is reduction, which isthrough direct binding of the catechol moiety of baicalein to the iron.
Huang et al.165 synthesized baicalein analogs (Fig. 9) and evaluated their anti-in-flammatory and anti-aggregatory activities. The chlorinated analogs 133 and 134 showed themost potent inhibition of NO production in LPS-stimulated RAW246.7 cells than baicaleinand positive control, L-NAME (IC504300 mM). The inhibitory effects of the analogs mightbe due to the inhibition of iNOS expression. The analogs 133 and 135–137 (Table VI) showedstronger inhibition of AA- and collagen-induced platelet aggregation in rabbit washed pla-telets than baicalein and aspirin. The SAR studies revealed that the introduction of anelectron-withdrawing group such as 20-Cl, 40-Cl, or 40-phenyl in the B-ring increased theactivity toward inhibition of NO and platelet aggregation. The co-planar feature of theflavones framework was essential for the pharmacological activity. The intermolecular H-bonding interactions in A-ring of the baicalein involving three hydroxyl groups (C-5, C-6,and C-7), reinforces the co-planarity. Surprisingly, 5,6,7-O-trimethyl baicalein (138) wascompletely devoid of activity on NO production and platelet aggregation.
3. Chrysin and its analogsChrysin (139) showed the preventing effect in COX-2 suppression and in inhibiting theplatelet aggregation.19 The various chrysin analogs modulated the activity, as well as theexpression of COX-2 and iNOS enzymes. The NO production was suppressed by chrysin andits analogs 140 and 141 (Fig. 10). Interestingly, the COX-2 enzyme was strongly inhibited by141 (IC50 2.7 mM), but not by 140. The molecular modeling study revealed that 141 fits well
Table V. Continued
Compounds Targets, cells IC50 (mM) Ref.
b-hexosaminidase, RBL-2H3 6.1 152
12-LOX, human platelets 81 164
15-LOX-1, human reticulocytes 3.4 164
Diosmetin (108) MMP-21 and -92, free enzyme 37.61, 11.62 137
NO, RAW 264.7 8.9 151
IL-41 and TNF-a2, RBL-2H3 31, 5.32 152
b-hexosaminidase, RBL-2H3 2.1 152
Tetra-O-
methylluteolin (109)
NO, RAW 264.7 2.4 151
Hexa-O-methyl-
myricetin (110)
NO, RAW 264.7 7.4 151
Fisetin (111) IL-41 and TNF-a2, RBL-2H3 3.81, 4.22 152
b-hexosaminidase, RBL-2H3 3.0 152
15-LOX-1, rabbit reticulocytes 1.8 157
LOX L1, soyabean 3.5 157
Pilloin (112) NO, RAW 264.7 11 151
b-hexosaminidase, RBL-2H3 28 152
ANTI-INFLAMMATORYNATURALPRODUCTS K 23
Medicinal Research Reviews DOI 10.1002/med

into the binding pocket of COX-2 and a hydrogen bond exists between the oxygen of theacetyl group at the 7-position of 141 and the hydroxyl group of Tyr355. The docking ofanalog 141 into the V523I mutant of COX-2 showed that Ile523 of COX-1 might contributeto the selectivity toward COX-2 over COX-1. The analog 141 showed no effect on iNOSactivity, whereas 140 and chrysin exhibited the weaker inhibition of iNOS in hemoglobinassay.166
O
OOH
HO
HO
R1
R2
R3
133, R1 = Cl, R2 = R3 = H 134, R1 = R2 = H, R3 = Cl135, R1 = H, R2 = R3 = Cl
O
OOH
HO
HO
132
O
OOCH3
H3CO
H3CO
138
O
OOH
HO
HOBr
F
O
OOH
HO
HO
136
137
Figure 9. Chemical structures of baicalein and its analogs.
Table VI. The Inhibitory Effects of Baicalein and its Analogues
Compounds Targets, cells IC50 (mM) Ref.
Baicalein (132) IL-12, macrophages1 and RAW 264.72 43.71, 17.42 162
IL-6 and -8, human epithelial cells – 22
IL-1b, IL-6, TNF-a, IFN-g and MIP-1a/b, PMNL – 22
b-hexosaminidase, RBL-2H3 16 152
15-LOX-1, rabbit1 and human2 reticulocytes 1.01, 1.62 1571, 1642
12-LOX, human platelets 0.64 164
NO, RAW 264.7 66.4 165
Platelet aggregation by AA1 and collagen2, RAW 264.7 1811, 36.52 165
133 NO, RAW 264.7 53.5 165
Platelet aggregation by AA1 and collagen2, RAW 264.7 24.81, 10.32 165
134 NO, RAW 264.7 46.1 165
Platelet aggregation by AA1 and collagen2, RAW 264.7 1091, 80.52 165
135 NO, RAW 264.7 85.4 165
Platelet aggregation by AA1 and collagen2, RAW 264.7 1021, 342 165
136 NO, RAW 264.7 87.0 165
Platelet aggregation by AA1 and collagen2, RAW 264.7 78.51, 62.42 165
137 NO, RAW 264.7 4100 165
Platelet aggregation by AA1 and collagen2, RAW 264.7 581, 7.72 165
138 NO, Platelet aggregation by AA and collagen, RAW 264.7 Inactive 165
24 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

B. Lignans
Two lignans, woorenoside IV (142)167 and 143,168 isolated from rhizomes of Coptis japonicasuppressed TNF-a and NO production, as well as lymphocyte proliferation triggered by in-flammatory signals such as various mitogens. Both the compounds strongly inhibited the mi-togenic response by activated macrophages and lymphocytes. Their effects on TNF-aproduction from LPS-stimulated RAW 264.7 cells were more potent than the positive control,pentoxifylline (IC50 247mM) and less potent than prednisolone (IC50 25.4mM). The effect ofwoorenoside IV was more pronounced than 143 on NO production. Sauchinone (144) obtainedfrom Saururus chinensis roots inhibited the LPS-stimulated NO production in RAW 264.7 cellsand prevented the NF-kB activation, which is known to play a crucial role in iNOS expression.Sauchinone exerted inhibitory effects on iNOS, TNF-a, and COX-2 gene expression and on theactivation of NF-kB, AP-1, c-AMP-response element-binding protein, and CCAAT/enhancer-binding protein in RAW 264.7 cells.169 Diphyllin acetylapioside (145), isolated from Haplo-phyllum hispanicum exhibited 5-LOX inhibitory activity (Table VII). It fulfils the main structuralrequirement for an aryl naphthalene lignan to inhibit the 5-LOX activity, namely the presence ofan ‘‘up’’ lactone and an acetylglycoside residue increase its polarity and therefore its solubility inaqueous solvents.170 Manassantin A (146) and B (147) (Fig. 11), isolated from S. chinensis,showed a potent inhibitory activity against TNF-a-induced NF-kB activation in HeLa cells,which were transiently transfected with the plasmid containing NF-kB-directed luciferase ex-pression system. The presence of an phenylpropanoid moiety attached to C-4 and/or C-40 wasfound to be important for the inhibition of NF-kB activation.171 Sibyllenone (148), a noveldiastereomer of ocobullenone, isolated from stem bark of Ocotea bullata, showed potent in-hibition of 5-LOX enzyme with IC50 value of 18.6mM.172 Arctigenin (149) inhibited thephosphorylation of MAP kinases ERK1/2, p38 kinase, and JNK, and their activities in LPS-stimulated RAW 264.7 cells. Arctigenin also blocked the TNF-a production and NF-kB acti-vation, and suppressed the TNF-a mRNA level and nuclear translocation of p65 accompaniedby inhibition of IkBa phosphorylation in the cells exposed to LPS.169
C. Phloroglucinols
Garcinielliptone M (150), a new phloroglucinol isolated from Garcinia subelliptica seedsshowed comparable inhibitory effects on p-methoxy-N-methylphenethylamine (compound48/80)-stimulated release of b-glucuronidase and histamine from rat peritoneal mast cells incomparison to mepacrine (IC50 13.7 and 23.3 mM, respectively).173 Arzanol (151) (Fig. 11),isolated from Helichrysum italicum ssp. microphyllum showed potent inhibition on the pro-duction of IL-1b and TNF-a, and to a lesser extent of IL-6, IL-8, and PGE2 in LPS-stimulated human peripheral monocytes (Table VII).174
D. Quinones
Hypericin (152), a napthodianthrone isolated from H. perforatum inhibited the PMA- andTNF-a-induced activation of NF-kB in HeLa and TC10 cells, respectively.22 Hypericin
O
OH
HO
O
O
OH
H3CO
O
O
O
O
OC
C
O
H3C
O
H3C
139 141140
Figure 10. Chemical structures of chrysin and its analogs.
ANTI-INFLAMMATORYNATURALPRODUCTS K 25
Medicinal Research Reviews DOI 10.1002/med
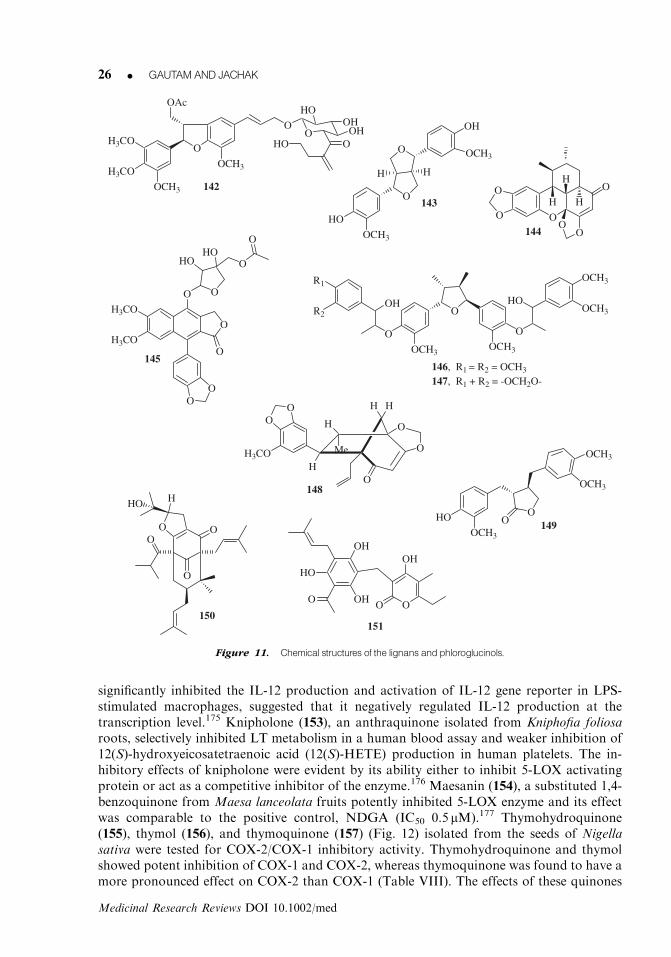
significantly inhibited the IL-12 production and activation of IL-12 gene reporter in LPS-stimulated macrophages, suggested that it negatively regulated IL-12 production at thetranscription level.175 Knipholone (153), an anthraquinone isolated from Kniphofia foliosaroots, selectively inhibited LT metabolism in a human blood assay and weaker inhibition of12(S)-hydroxyeicosatetraenoic acid (12(S)-HETE) production in human platelets. The in-hibitory effects of knipholone were evident by its ability either to inhibit 5-LOX activatingprotein or act as a competitive inhibitor of the enzyme.176 Maesanin (154), a substituted 1,4-benzoquinone from Maesa lanceolata fruits potently inhibited 5-LOX enzyme and its effectwas comparable to the positive control, NDGA (IC50 0.5 mM).177 Thymohydroquinone(155), thymol (156), and thymoquinone (157) (Fig. 12) isolated from the seeds of Nigellasativa were tested for COX-2/COX-1 inhibitory activity. Thymohydroquinone and thymolshowed potent inhibition of COX-1 and COX-2, whereas thymoquinone was found to have amore pronounced effect on COX-2 than COX-1 (Table VIII). The effects of these quinones
O
OCH3
H3CO
H3CO
OCH3
O
OAc
O
O
HH
OH
OCH3
OCH3
HO
142
143O
O O
H
O
H
OO
144
O
OO
O O
HOHO O
O
H3CO
H3CO
145
R1
R2
O
O
O
OCH3
OCH3HO
OCH3OCH3
OH
146, R1 = R2 = OCH3
147, R1 + R2 = -OCH2O-
OO
H3CO O
O
O
H
H
H H
148
OO
OCH3
OCH3
OCH3
HO149O
OO
HO
O
H
150O
OHOH
HO
OOHO
151
Me
O
H
OOH
OHO
HO
HO
Figure 11. Chemical structures of the lignans and phloroglucinols.
26 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

were comparable to indomethacin.178 An anthraquinone, emodin (158) isolated from Aloevera, inhibited the TNF-a-induced NF-kB activation, IkB degradation, DNA binding andexpression of cell surface adhesion proteins in human vascular endothelial cells(HVEC).179,180 Emodin inhibited the invasiveness of human cancer cells by suppressingMMP-9 expression through inhibiting AP-1 and NF-kB signaling pathways. Emodin sup-pressed the phosphorylation of two MAPKs, ERK and JNK, but not p38 kinase, leading toreduced c-Jun phosphorylation and AP-1 DNA binding. Moreover, emodin inhibited TPA-induced degradation of IkBa, nuclear translocation of p65, and NF-kB DNA-binding ac-tivity.181
E. Phenylpropanoids
Phenylpropanoids 159 and 160 (Fig. 12) isolated from plants of the Illicium species sig-nificantly inhibited A23187-stimulated histamine release from RBL-2H3 cells. Furthermore,compounds 159 and 160 (Table VIII) caused a reduction in TNF-a level. Compound 159
exhibited a more pronounced inhibitory effect on TNF-a level than 160. Both phenylpro-panoids contain a prenyl moiety on the aromatic ring, and O-prenylated compound, 159showed a greater inhibition of TNF-a activity than C-prenylated compound, 160. The pre-sence of an O- or C-prenyl moiety in the molecules seems to influence the inhibitory activitytoward TNF-a production and/or secretion from RBL-2H3 cells.182 Acteoside (161), aphenylpropanoid glycoside isolated from Buddleja officinalis showed a significant inhibitionon release of NO in LPS-stimulated RAW 264.7 cells.183
F. Stilbenes
Resveratrol (3,5,40-trihydroxystilbene) first isolated in 1940 from the roots of white helleboreVeratrum grandiflorum exists in two isoforms: trans-resveratrol (162) and cis-resveratrol (163)(Fig. 13) where, the trans-isomer is the more stable than the cis-isomer. While trans to cis-isomerization is facilitated by ultra violet light and high pH, the cis to trans conversion isfacilitated by visible light, high temperature, or low pH. Resveratrol has been of great scientific
Table VII. The Inhibitory Effects of Various Lignans and Phloroglucinols on Inflammatory
Mechanisms
Compounds Targets, cells IC50 (mM) Ref.
Lignans
Woorenoside IV (142) TNF-a1 and NO2, RAW 264.7 46.61, 14.42 167
143 TNF-a, RAW 264.7 39.3 168
Sauchinone (144) NO, iNOS, TNF-a and COX-2, RAW 264.7 – 169
Diphyllin acetylapioside (145) LTB41, 5-HETE2 and LT3 0.61, 0.72, 0.53 170
Manassantin A (146) NF-kB, HeLa 2.5 171
Manassantin B (147) NF-kB, HeLa 2.7 171
Sibyllenone (148) 5-LOX, free enzyme 18.6 172
Arctigenin (149) ERK, p38 and JNK, RAW 264.7 – 169
NO, iNOS, TNF-a and NF-kB, RAW 264.7 – 169
Phloroglucinols
Garcinielliptone M (150) b-glucoronidase1 and histamine2, mast cells 13.61, 19.02 173
Arzanol (151) HIV-1-LTR, T cells 5.0 174
IL-1b1 and TNF-a2, human monocytes 5.61, 9.22 174
IL-61, -82 and PGE23, human monocytes 13.31, 21.82, 18.73 174
ANTI-INFLAMMATORYNATURALPRODUCTS K 27
Medicinal Research Reviews DOI 10.1002/med

interest over the years.23,82,184 Resveratrol inhibited COX-1 and COX-2 enzyme greater thanaspirin, and exhibited anti-inflammatory activity better than phenylbutazone and similar toindomethacin against carrageenan-induced paw edema in rats.27 It significantly inhibited theLPS-stimulated release of TNF-a and NO from rat cortical microglia and N9 microglial cells.It appeared to suppress the LPS-induced degradation of IkBa, expression of iNOS and phos-phorylation of p38 MAPK in N9 microglial cells,185 suppressed NF-kB activation and down-regulation of inflammatory gene products such as COX-2, 5-LOX, IL-1b, and IL-6.23,186,187
Resveratrol is thought to contribute to the protective effects of red wine by potentiating theaction of endogenous antiplatelet substances—PGI2 and PGE1.
188 Elmali et al.189,190 investigatedthe in vivo effects of intra-articular injections of resveratrol on cartilage and synovium in anexperimental osteoarthritis model in rabbits and found that resveratrol significantly reducedcartilage tissue destruction. Oxyresveratrol (164), obtained from Morus alba showed aninhibitory effect against FeSO4/H2O2-induced lipid peroxidation in rat microsomes and reducedthe carageenan-induced rat paw edema significantly. Oxyresveratrol inhibited the nitrite accu-mulation and suppressed the iNOS expression in LPS-stimulated RAW 264.7 cells, but showed alittle effect on iNOS enzyme activity (Table IX), suggesting the inhibitory effect was mainly dueto the inhibition of iNOS expression rather than the enzyme activity.191 A prenylated derivative165 of resveratrol, purified from fungally infected peanuts, reduced the LPS-induced COX-2protein expression better than resveratrol in J774 cells. Resveratrol and 165 suppressed theCOX-2 mRNA level by 35 and 53%, respectively, at 30mM.192 a-Viniferin (166), a trimer of
O
O
OH OH
OH OH
CH3
CH3
HO
HO
152
O
O
OH
CH3
HO OH
OCH3
OH
O
153
O
O
OH
H3CO (CH2)9CH=CH(CH2)3CH3
154
OH
OH
O
OOH
157156
155
O
O
OHOH
CH3HO
158
OCH3
O
H3CO
OCH3
OH
H3CO
159
160
O
O
HO
HO
O OH
OHOH
OH
O
OH3CHO
OHOH
161
Figure 12. Chemical structures of the quinones andphenylpropanoids.
28 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

resveratrol isolated from Caragana chamlagu, showed a more potent inhibitory activity onCOX-2 than COX-1 enzyme. a-Viniferin inhibited the synthesis of the COX-2 and the iNOStranscript in LPS-activated RAW 264.7 cells. a-Viniferin also exhibited significant anti-inflammatory activity in carrageenan-induced paw edema in mice193 and adjuvant-inducedarthritic inflammation and bony change in rats.194
1. Resveratrol and its analogsIn order to find more selective COX-2 inhibitors, some hydroxylated and methoxylatedanalogs of resveratrol were synthesized by Murias et al.195 All the hydroxylated and meth-oxylated analogs (Fig. 13) exhibited selectivity indices of COX-1/COX-2 more than that ofresveratrol. The hydroxylated analogs 167–171 (Table X) showed a high rate of COX-2inhibition and more selectivity toward COX-2 than celecoxib (IC50, COX-1: 19.026 mM;COX-2: 0.03482 mM), a selective COX-2 inhibitor already established in the market. Amongthe hydroxylated analogs, 171 was most potent with selectivity indices (COX-1/COX-2)higher than celecoxib (COX-1/COX-25 546). The methoxylated analogs 172–174 alsoshowed selectivity to COX-2, but exhibited weaker inhibition of COX-1 and selectivity indexof 3–31. Hydroxylated analogs therefore represent a novel class of highly selective COX-2inhibitors and promising candidates for in vivo studies.195 Handler et al.196 synthesized someresveratrol analogs in which stilbenes are bridged by a methylene or ethylene group andusually named as 1,2-dihydronaphthalene derivatives and 1H-indene derivatives. The bridgedstilbenes showed a high rate of COX-1 inhibition than COX-2. Among the bridged analogs,polymethoxylated molecules 175–181 (Table X) exhibited potent and selective COX-1 in-hibition, and 177 and 180 were most potent. Cho et al.197 studied the effect of variousresveratrol-related hydroxylstilbenes on NO production in LPS-stimulated RAW 264.7 cells.It was observed that compounds 182–185 (Fig. 13) were potent inhibitors with order of IC50
Table VIII. The Inhibitory Effects of Various Quinones and Phenylpropanoids on Inflammatory
Mechanisms
Compounds Targets, cells IC50 (mM) Ref.
Quinones
Hypericin (152) NF-kB, HeLa and TC10 cells – 22
IL-12, macrophages 1.45mg/ml
175
Knipholone (153) LT, human blood 4.2 176
12(S)-HETE, human platelets – 176
Maesanin (154) 5-LOX, free enzyme 0.7 177
Thymohydroquinone
(155)
COX-11 and -22, free enzyme 0.11, 1.02 178
Thymol (156) COX-11 and -22, free enzyme 0.61, 0.22 178
Thymoquinone (157) COX-2, free enzyme 0.3 178
Emodin (158) NF-kB and IkB, HVEC – 179–181
MMP-9, ERK, JNK, DNA-binding
Phenylpropanoids
159 Histamine, RBL-2H3 96.9� 182
TNF-a, RBL-2H3 – 182
160 Histamine, RBL-2H3 84.3� 182
TNF-a, RBL-2H3 – 182
Acteoside (161) NO, iNOS and AP-1, RAW 264.7 – 183
�Represent the percent inhibition at 50mM.
ANTI-INFLAMMATORYNATURALPRODUCTS K 29
Medicinal Research Reviews DOI 10.1002/med

values as 182418441854183. SAR study revealed that the structural balance betweenoxygen functional groups on the benzene rings is important for their activity.
G. Diarylheptanoids
Curcuma longa popularly known as turmeric, has been used for centuries as a spice in theIndian subcontinent. An extract of C. longa rhizomes contain curcumin (186), demethox-ycurcumin (187), and bisdemethoxycurcumin (188) (Fig. 14). Curcumin is well documentedin Ayurveda as an anti-inflammatory agent23,82 and several reviews described its variousmechanisms of anti-inflammatory action.198–201 Curcumin presents high interest as a lead
HO
HO
OH
162
HO
OH OH
163
HO
HO
OH
OH165
HO
OHOH
OH164
O
O
O
HO
OH
OH
HH
HH
H H
OH
OH
HO
166
173, R = H174, R = OCH3
H3CO
H3CO
R
OCH3
OCH3
H3CO
O
O179
H3CO
R2
R1
180, R1 = R2 = H181, R1 = R2 = OCH3
HO
HOOCH3
OH
OCH3
OCH3
R
175, R = OCH3
176, R = H
OCH3
OCH3
RH3CO
177, R = H178, R = OCH3
HO
HOOR
H3CO
H3COOCH3
182, R = H183, R = CH3
184
185
HO
HO
OHHO
HO
HO
R1
OH
R2
167
168, R1 = H, R2 = OH 169, R1 = OH, R2 = H
HO
HO
R
OH
OHHO
170, R = H171, R = OH
H3CO
H3CO
OCH3
OCH3
172
Figure 13. Chemical structures of stilbenes and various resveratrol analogs.
30 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

molecule to develop new clinically relevant anti-inflammatory drugs. Curcumin blocked theIL-12 mediated T cells proliferation and Th1 differentiation and down-regulated the TNF-a-induced increase in MMP-13 mRNA and protein expression in human chondrocytes andSW1353 cells by inhibiting the NF-kB activation and JNK.202,203 Curcumin significantlyinhibited the increase of IL-1b and TNF-a in chronic inflammation in rats.204 The potentialuse of curcumin in therapeutic settings has raised the obvious issues of toxicity and tolerance.At least three different phase I clinical trials indicated that curcumin is well tolerated whentaken at doses as high as 12 g/day. In the clinical trial of curcumin’s efficacy as an anti-rheumatic agent, investigators compared its activity with that of phenylbutazone in a short-term, double-blind, crossover study involving 18 relatively young patients (age range, 22–48
Table IX. The Inhibitory Effects of Stilbenes on Inflammatory Mechanisms
Compounds Targets, cells IC50 (mM) Ref.
Resveratrol (162) COX-11 and COX-22, free enzyme 151, 852 6
TNF-a, cortical microglial – 185
NO, iNOS and p38, N9 microglial – 185
COX-2, 5-LOX, IL-1b and IL-6 – 186,187
PGE2, human PBMC 3.2 188
PGI2 and PGE1, Endogenous platelets – 188
COX-2 expression, J774 cells 35� 192
Oxyresveratrol (164) NO and iNOS, RAW 264.7 – 192
165 COX-2 expression, J774 cells 53� 192
a-Viniferin (166) COX-2, free enzyme 4.9 193
NO1 and iNOS2, RAW 264.7 2.71, 4.72 193
�Represent the percent inhibition at 30mM.
Table X. The Inhibitory Effects of Resveratrol and its Analogues on COX-1 and COX-2
IC50 (mM)
Compounds COX-1 COX-2
COX-1/COX-2
selectivity
COX-2/COX-1
selectivity Ref.
Resveratrol (162) 0.535 0.996 0.54 1.86 195,196
167 2.072 0.04537 45.67 – 195
168 0.00998 0.00171 5.83 – 195
169 4.713 0.0113 417.08 – 195
170 0.01027 0.00138 7.44 – 195
171 0.748 0.00104 719.23 – 195
172 2.834 0.796 3.56 – 195
173 7.247 0.514 14.11 – 195
174 11.348 0.355 31.97 – 195
175 0.48 0.66 – 1.38 196
176 61.1 4100 – 41.63 196
177 0.38 0.03 – 0.079 196
178 0.015 4100 – 46666 196
179 12.8 4100 – 47.8 196
180 0.046 62.3 – 135 196
181 0.03 1.33 – 44.3 196
ANTI-INFLAMMATORYNATURALPRODUCTS K 31
Medicinal Research Reviews DOI 10.1002/med

R1
HO
R2
OH
O O
186, R1 = R2 = OCH3
187, R1 = H, R2 = OCH3
188, R1 = R2 = H
MeO
HO
OMe
OH
N X
189, X = NH190, X = O
MeO
HO
OMe
OH
HNR
192, R = Me193, R = i-Bu
MeO
HO
OMe
OH
O O
194
MeO
HO
OMe
OH
O R
195, R = OH197, R = H
MeO
HO
OMe
OH
OH OH
196 HO OH
O O
F F
198
HO
O
F FOH
199
R2
R3
R2
R3
O OR1 R1
200, R1 = R2 = R3 = OCH3
201, R1 = R2 = H, R3 = OCOCH3
MeO
HO
OMe
OH
O
191
OO
HO
HO
HO
OR1
R2
R3
R4
R1
R2
R3
R4
206, R1 = R4 = H, R2 = OCH3, R3 = OH207, R1 = R4 = H, R2 = OCH3, R3 =
208, R1 = R2 = R4 = H, R3 = O(CH2)3N(CH3)2
209, R1 = R2 = R4 = H, R3 = N(CH3)2
O O
OR1
R2
R3
R4
R1
R2
R3
R4
210, R1 = R4 = H, R2 = OCH3, R3 = OH211, R1 = R2 = R4 = H, R3 =
212, R1 = R2 = R4 = H, R3 = OCH2CH=CH2
O O
OR1
R2
R3
R4
R1
R2
R3
R4
202, R1 = R4 = H, R2 = OCH3, R3 = OH203, R1 = R4 = H, R2 = OCH3, R3 =
204, R1 = R2 = R4 = H, R3 = OCH2CH=CH2
O O
O O BrBr
O
205
(a)
(b)
Figure 14. Chemical structures of diarylheptanoids.
32 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

years) receiving a daily dose of either curcumin (1200mg) or phenylbutazone (300mg) for 2weeks. At the dose used, curcumin was well tolerated, had no side effects, and exerted anantirheumatic activity comparable to that of phenylbutazone.198 Curcumin appears to im-prove the clinical outcomes of patients suffering from chronic pancreatitis, an intenselypainful inflammatory condition induced by oxidative stress, by reversing lipid peroxidation.As shown in a randomized, placebo-controlled pilot study involving 20 patients with tropicalpancreatitis, an oral combination of curcumin 500mg and piperine 5mg provided effectivepain relief and beneficially modulated a pair of markers of oxidative stress (i.e. significantlyreduced malonyldialdehyde levels and increased glutathione levels in erythrocytes).205
We have reported the synthesis and evaluation of eight pyrazole and isoxazole analogs ofcurcumin for COX-1/COX-2 inhibition and antioxidant activity. The SAR study revealedthat the pyrazole analog (189) of curcumin increased the COX-1 inhibition slightly(80.5–87.0%), whereas the COX-2 inhibition was increased twofold (35–61.0%). Thus,pyrazole analog showed a significant increased in the selectivity toward COX-2 enzyme(COX-2/COX-15 0.70) compared to curcumin (COX-2/COX-15 0.43). Similarly, the iso-xazole analog (190) showed the selectivity toward COX-2 inhibition with a COX-2/COX-1ratio of 0.72. The replacement of b-diketo fragment of curcumin by a pyrazole ring sig-nificantly enhanced COX-2/COX-1 selectivity.28 Lee et al.206 synthesized some diarylhepty-lamines 192 and 193, and diarylheptanoid analogs 194–197 (Fig. 14) similar to a natural anti-inflammatory agent oregonin207 (191) and evaluated their effects against LPS-induced ex-pression of iNOS and COX-2. The analogs showed stronger potency than oregonin againstCOX-2-derived PGE2 formation and 195 was most potent with an IC50 value of 0.7 mM. Incontrast to moderate inhibition of NO, these analogs showed significant inhibition of PGE2
and have potential to develop as anti-inflammatory agents. Some curcumin analogs weresynthesized and evaluated for COX-2 inhibition and chemopreventive potential.208 Curcumin(IC50 15.9 mM) and its analogs 198 (IC50 23.7 mM) and 199 (IC50 5.5 mM) were the mostpotent inhibitors of COX-2, whereas the other analogs were less active than curcumin.Analog 198 showed a weaker inhibition of iNOS expression and had no effect on the ex-pression of COX-2. The SAR study revealed that modifications in the b-diketone functionin curcumin, which led to a shorter chain and/or a more rigid molecule, could enhancethe activity. An electron-withdrawing group (fluorine atom/methoxy group) ortho tothe OH-group of p-hydroxyphenyl moiety enhanced the activity and selectivity toward COX-2, whereas an electron-donating group is important for the effective mitochondrial perme-ability transition pore. The bulkier groups on the aromatic moiety lead to complete loss ofactivity.
Recently it is reported that COX-1 plays an important role in inflammation and carci-nogenesis; therefore, the selective COX-1 inhibitors could find their potential as anti-canceragents. Handler et al.209 synthesized some novel curcumin analogs and evaluated their effectson COX-1 and COX-2 catalyzed PGE2 production. All the analogs showed higher rate ofCOX-1 inhibition and the most potent analogs were 200 (COX-1: IC50 0.06mM, COX-2: IC50
100mM, COX-2/COX-15>1666) and 201 (COX-1: IC50 0.05mM, COX-2: IC504100mM,COX-2/COX-15>2000). It was observed that the introduction of nonpolar (methoxy group)and polar (methyl ester) substituents on the aromatic ring improved the selectivity towardCOX-1. The molecular docking study revealed that the curcumin analogs showed a good vander Waals interaction and hydrogen bonding toward COX-1 enzyme. Recently, Liang et al.210
synthesized some mono-carbonyl analogs of curcumin and tested their activity against LPS-induced TNF-a and IL-6 production. The cycohexanone-derived analogs 202–205 were moreactive than cyclopentanone-derived 206–209 and acetone-derived 210–212 analogs (Fig. 14).The analogs 206, 207, 210–212, and 202–205 showed more potent inhibition of TNF-a ex-pression than curcumin, whereas 210, 202, and 208 exhibited better inhibitory effects on IL-6
ANTI-INFLAMMATORYNATURALPRODUCTS K 33
Medicinal Research Reviews DOI 10.1002/med

expression. The SAR study revealed that a 3-methoxy group is critical and allyloxy-substitutedanalogs showed stronger inhibition than dimethylamino- and heterocyclo-substituted analogs.
7. Marine Natural Products
Ascidiathiazone B (213), a tricyclic thiazine-containing quinolinequinone alkaloid, isolatedfrom a New Zealand ascidian Alpidium species, inhibited the PMA-stimulated superoxideproduction from human neutrophils in vitro as well as in vivo in a gouty model of arthritis atoral dose of 25.6 mM/kg.211 Myltayl-4(12)-ene-2-caffeate (214), isolated from the Malagasyliverwort Bazzania nitida, inhibited the NO production in LPS-activated RAW 264.7 cells.212
Fucoside B (215), isolated from Caribbean gorgonian Eunicea fusca, inhibited the transfor-mation of AA to LTB4 by inhibiting 5-LOX.210 Didemnilactones A (216) and B (217) (TableXI), isolated from tunicate Didemnum moseleyi, showed inhibition against 5-LOX in humanPMNL with IC50 values of 9.4 and 8.5 mM, respectively.213 Ambrein (218) and its derivatives
Table XI. The Inhibitory Effects of Marine Natural Products
Compounds Targets, cells
IC50
(mM) Ref.
Ascidiathiazone B (213) Superoxide, neutrophils 0.44 211
Myltayl-4(12)-ene-2-caffeate (214) NO, RAW 264.7 6.3 212
Fucoside B (215) 5-LOX, free enzyme 18.0 213
Didemnilactones A (216) 5-LOX, PMNL 9.4 213
Didemnilactones B (217) 5-LOX, PMNL 8.5 213
Ambrein (218) Elastase, human neutrophils 4.6 214
219 Elastase, human neutrophils 95.1� 214
220 Elastase, human neutrophils 92.2� 214
Sodium 1-(12-hydroxy)-octadecanyl
sulfate (221)
MMP-2, free enzyme 9.0 mg/ml
215
Manoalide (222) PLA2, free enzyme 1.7 213
PLA2, human synovial 3.9 218
PLA2, bee venom 7.5 218
223 PLA2, human synovial 0.9 218
PLA2, bee venom 0.5 218
Secomanoalide (224) PLA2, free enzyme – 213
Dysidine (225) PLA2, free enzyme 2.0 219
Cacospongionolides B (226) PLA2, human – 213
iNOS, COX-2 and TNF-a – 213
Cacospongionolides E (227) PLA2, synovial 1.4 213
228 PLA2, human synovial 4.2 218
PLA2, bee venom 1.7 218
Scalaradial (229) PLA2, venom 0.07 180
sPLA2, human recombinant 5.4 180
PLA2, U937 cells 28 180
Astaxanthin (230) NO, PGE2, TNF-a, IL-1b and NOS, mouse
macrophage
– 223
Epitaondiol (231) PLA2 and LTB4, human neutrophils – 224
TXB2, human neutrophils 3.8 224
Bolinaquinone (232) PLA2, iNOS, LTB4 and PGE2 – 225,226
Scytonemin (233) polo-like kinase 11 and PKC b12 2.31 227
5.42
�Represent the percent inhibition at10mg/ml.
34 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

219 and 220 (Fig. 15), isolated from Physeter macrocephalus, inhibited the elastase releasefrom human neutrophils. Ambrein showed the most potent effect, whereas 219 and 220
exhibited 95.1 and 92.2% inhibition, respectively, at 10 mg/ml and their effects were betterthan the positive control, genistein (51.6% inhibition).214 Sodium 1-(12-hydroxy) octadeca-nyl sulfate (221) was isolated from a marine tunicate of the family Polyclinidae as a MMP-2inhibitor.215 Manoalide (222), isolated from the sponge Luffariella variabilis, reported toirreversibly inhibit PLA2 by modifying a specific number of its lysine residues. The me-chanism of its action was proposed that the rings of the pyranofuranone moiety open togenerate a,b-unsaturated carbonyl compound, which could react in different ways with lysineresidue of PLA2. It was proposed that the aldehyde group arising from the hemiacetalfunction in the dihydropyran ring was essential to irreversibly inactivate PLA2.
216 Faulkner’sgroup demonstrated that manoalide could be obtained in fair yield from this sponge, andtherefore it was considered to be a good candidate for drug development. It advanced toPhase II clinical trials as a topical antipsoriatic, but work on the natural product stopped assufficient quantities of the compound would not pass through the skin using the formulationsdeveloped for the trials.217 De Rosa et al.218 synthesized some manoalide analogs, of whichan analog containing a farnesyl residue linked to the pyranofuranone (FMA, 223), was foundto be the most potent PLA2 inhibitor than manoalide. It inhibited human synovial and beevenom PLA2 with IC50 values of 0.9 and 0.5 mM, respectively. FMA was tested in the mousecarrageenan paw edema at the oral dose of 10mg/kg and showed an activity comparable toindomethacin. Three congeners of manoalide were also obtained from the same sponge, ofwhich secomanoalide (224) was more potent against bovine pancreatic PLA2.
213 Dysidine(225), isolated from a sponge Dysidea species, significantly inhibited the human synovialPLA2 and exerted a higher potency and selectivity toward this enzyme than manoalide. Italso showed the inhibition of secretory PLA2 belonging to Naja naja venom, porcine pan-creatic enzyme, bee venom, cPLA2 from 264.7 cells. In addition, dysidine modulated theother human leukocyte functions such as degranulation and superoxide production.219 Ca-cospongionolides B (226) and E (227), isolated from the Mediterranean sponge Fasciospongiacavernosa, exhibited specific inhibition against human and synovial PLA2, respectively (TableXI). A synthetic analog of cacospongionolide B containing a farnesyl residue (FCA, 228),inhibited human synovial and bee venom PLA2 with IC50 values of 4.2 and 1.7 mM, re-spectively.218 Scalaradial (229), isolated from marine sponge Cacospongia species, selectivelyinhibited sPLA2. Scalaradial efficiently inhibited the bee venom PLA2 (IC50 0.07 mM) andpurified human recombinant type II sPLA2 activity in vitro (IC50 5.4 mM) but displayed aweak inhibition of cytosol 85 kDa PLA2 from U937 cells (IC50 28 mM).180 Due to its peculiardialdehyde structural feature, it has been proposed that scalaradial exerts PLA2 enzymaticinactivation by means of an irreversible covalent modification of its target.220 Scalaradial alsoexerted an inhibitory effect on HL-60 cells proliferation, and inhibited the epidermal growthfactor receptor (EGFR)-mediated Akt phosphorylation, and this effect was found to beindependent of sPLA2.
221 Scalaradial showed an interesting finding in BEL-7402 cells byexhibiting the inhibition of ERK1/2 phosphorylation in early stage and stimulation ofERK1/2 phosphorylation in the later stage of its treatment, which displayed its biphasicregulatory responses.222 Astaxanthin (230) (Fig. 15), a red–orange carotenoid pigment, canprevent inflammatory processes by blocking the expression of pro-inflammatory genes as aconsequence of suppressing the NF-kB activation. It inhibited the production of NO andPGE2 and the pro-inflammatory cytokines TNF-a and IL-1b. Astaxanthin exhibited a dose-dependent ocular anti-inflammatory effect by suppression of NO, PGE2, and TNF-a pro-duction by direct blockade of NOS activity. In PBMC from asthmatic patients, astaxanthinalone or in combination with gingkolide B, can suppress T-cell activation comparable tocetrizine dihydrochloride and azelastine, two commonly used antihistamines.223 A terpenoid,
ANTI-INFLAMMATORYNATURALPRODUCTS K 35
Medicinal Research Reviews DOI 10.1002/med

S
NH
N
O
COOH
OO O
213HH
O
O
OH
OH
214
OH
218
OHO
219
O
220
O
H H
O
HO HOO
O
OH
O
O
OH
215 216
217
OSO3Na
OH
221
O O
O
OH
HO
222
O
O
OH
HO
OH224
O
O
HO
NH
SO3HH
225
O OH
OH
O
226
OHO
OHO
O223
OH
O
HO
OHH
231
O
O
OH
OCH3
H
232
OH
N
H
O
N
H
O
233OH
O
HO
O
OH
230HH
H
OO
O
O
229
O OH
OH
O
227
O
O
O
OH228
(a)
(b)
Figure 15. Chemical structures of marine natural products.
36 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

epitaondiol (231) isolated from Stypopodium flabelliforme was tested in vitro on sPLA2, onear mice edema and on other functions of human neutrophils. The IC50 value measuredfor the decrease of TXB2 production was 3.8 mM, whereas the effect on LTB4 inhibitionwas lower. It blocked hsPLA2 activity and showed topical anti-inflammatory effects onTPA-induced mouse ear edema and was more potent than indomethacin.224 Bolinaquinone(232), a sesquiterpenoid isolated from a Dysidea spp. significantly inhibited cytokinerelease, iNOS expression and eicosanoid (LTB4, PGE2) generation in vitro and in vivo inseveral models of inflammation through secretory PLA2 inhibition.
225,226 Scytonemin (233),a yellow-pigment isolated from marine cyanobacteria, inhibited both polo-like kinase 1(IC50 2.3 mM) and protein kinase C b1 (IC50 5.4 mM) in vitro, while topical applicationof this novel pharmacophore reduced phorbol–ester-induced mouse ear edema(IC50 10.9 mg/ear).
227
8. Miscellaneous
Capsaicin (234), the major pungent principle in Capsicum was isolated in 1846 and Capsicumextracts were proposed as instant relief for toothache in the mid of the 19th Century. The Ca21
influx caused by capsaicin releases neuropeptides such as substance P and calcitonin gene-related peptides, which are thought to be mainly involved in the resulting neurogenic in-flammation. It is used as topical ointment formulation for neuropathic and osteoarthritic pain.ALGRX-4975 is a high-concentration injectable formulation of capsaicin, which is beingdeveloped by AlgoRX pharmaceuticals. This formulation is intended to be used in manage-ment of pain associated with osteoarthritis, tendonitis, and postsurgical conditions. Phase IIclinical trials were underway in 2004. Capsaicin has poor oral bioavailability, is highly
Table XII. The Inhibitory Effects of Miscellaneous Compounds
Compounds Targets, cells IC50 (mM) Ref.
236 COX-2, free enzyme 6.9 229
237 COX-2, free enzyme 10.1 229
Antrocamphin (238) Superoxide, neutrophils 9.33 230
Antcin A (239) Superoxide, neutrophils 8.55 230
Antcin B (240) Superoxide, neutrophils 9.82 230
241 NO 15.3 231
242 NO 19.2 231
243 NO 13.4 231
244 LOX, COX, PAF – 232
245 ROS, NO and NF-kB, RAW 264.7 – 233
iNOS, IL-6, TNF-a and COX-2, RAW 264.7 – 233
246 COX-11 and -22, free enzyme 0.0011, 2.22 234
Acetylbritannilatone (247) NO, PGE2, iNOS and COX-2, RAW 264.7 – 235
Anemonin (248) NO, ET-1 and ICAM-1, RIMEC – 237
Magnolol (249) NO, RAW 264.7 16.80 237
TNF-a, IL-8, COX-2 and NF-kB, THP-1 – 238
Honokiol (250) NO, RAW 264.7 6.44 239
TNF-a, IL-8, COX-2 and NF-kB, THP-1 – 238
D9-THC (251) CB1 and CB2 receptors, CNS – 228
252 CB2 receptors, CNS – 228
(-)-Cannabidiol (253) PGE2, NO, iNOS – 228
254 TNF-a, IFN-g – 228,240
ANTI-INFLAMMATORYNATURALPRODUCTS K 37
Medicinal Research Reviews DOI 10.1002/med

pungent, and relatively toxic upon systemic administration. Therefore, there has been con-siderable interest in the synthesis of improved analogs. Capsavanil (235), an amide ofhomovanillic acid, is in Phase II clinical trials as topical analgesic.228 Two new compounds236 and 237, isolated from Bauhinia purpurea roots, inhibited the COX-2 enzyme comparableto aspirin (IC50 19.8mM).229A new compound antrocamphin (238), and known compoundsantcin A (239) and antcin B (240) (Table XII) isolated from Antrodia camphorata showedbetter inhibition against fMLP-induced superoxide production than ibuprofen.230 The bi-benzyl derivatives 241–243 (Fig. 16), isolated from Dendrobium nobile stems displayed higherantioxidant activity and showed a potent inhibitory effects on LPS-stimulated NO produc-tion.231 (E)-1-(3,4-dimethoxyphenyl) butadiene (DMPBD, 244), isolated from Zingiber cas-sumunar, inhibited rat ear edema induced by ethyl phenylpropiolate (EPP), AA, and TPA, andit was more potent than the standard drugs oxyphenbutazone, phenidone, and diclofenac. Italso inhibited the rat paw edema induced by carrageenan. The in vitro study showed thatDMPBD inhibited collagen-, adenosine diphosphate-, AA-, and platelet activating factor-induced platelet aggregation. It was revealed that DMPBD exhibited more prominent in-hibitory effects on the LOX than COX as a mechanism of inflammation.232 Compound 245,isolated from Periostracum cicadae, inhibited the ROS and NO production, NF-kB activationas well as the expression of pro-inflammatory molecules such as iNOS, IL-6, TNF-a, andCOX-2 in LPS-induced RAW 264.7 cells.233 A new phorbol derivative 12-O-[(2R)-N,N-di-methyl-3-methylbutanoyl]-4-deoxyphorbol 13-acetate (246), isolated from Croton cilia-toglandulifer aerial parts, exhibited a potent inhibitory activity against COX-1 and -2 thanindomethacin (IC50, COX-1: 1.4mM; COX-2: 5.8mM) (Table XII).234 Acetylbritannilatone(ABL, 247), isolated from Inula britannica, inhibited the LPS/IFN-g-activated NO and PGE2
production, iNOS and COX-2 expression in RAW 264.7 cells. It effectively inhibited the NF-kB activation, essential for the expression of iNOS and COX-2, with its binding motif in thepromoter of target genes.235 A dilactone of cyclobutane-1,2-diol-1,2-diacrylic acid (anemonin,248), isolated from Pulsatilla chinensis roots, significantly inhibited the LPS-induced NOproduction, endothelin (ET)-1 and down-regulated the ICAM-1 expression in cultures of ratintestinal microvascular endothelial cells. These observations suggested that anemonin ex-hibited anti-inflammatory activity by inhibiting NO production, ET-1, and ICAM-1, thuspreventing intestinal microvascular dysfunction (Fig. 16).236 Two biphenyls, magnolol (249)and honokiol (250), isolated from stem bark of Magnolia obovata, inhibited NO, TNF-a, IL-8, COX-2. Honokiol suppressed NF-kB activation and NF-kB-regulated gene expressionthrough the inhibition of IKKs. It also inhibited the production of NF-kB-regulated in-flammatory and carcinogenic gene products, including MMP-9, IL-8, ICAM-1, and monocytechemotactic protein-1.237–239 THC (D9-tetrahydrocannabinol, 251) has been identified as apsychotropic principle in Cannabis sativa, which acts through the specific cannabinoid re-ceptor (CB). CB1 receptors are most abundant in the CNS and also occur on peripheralnervous system and CB1 agonists could have possible clinical use in chronic inflammatory andneuropathic pain, whereas CB2 receptors are restricted to immune tissues and cells and CB2
agonists may be useful in inflammatory and neuropathic pain. THC is a nonspecific agonist atCB1 and CB2 receptors, whereas its analog 252 is a selective CB2 agonist. (-)-Cannabidiol(253) possesses strong anti-inflammatory activity through inhibition of PGE2, NO, lipidperoxide, and endothelial NOS but no affinity to CB receptors and its derivative 254 showedincreased anti-inflammatory activity and very low affinity to CB1 and CB2 receptors. Can-nabidiol was evaluated in a number of in vitro and in vivo models and showed anti-arthriticproperties in collagen-induced arthritis in mice, down-regulated the TNF-a and IFN-grelease,and suppressed proliferative resonse of lymph node cells.228,240 An endogenous cannabimi-metic compound N-palmitoyl-ethanolamine exhibited potent anti-inflammatory and analgesiceffects in various experimental models.241
38 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

9. CONCLUSION
Natural products and traditional medicines are rich source of leads for pharmaceutical in-dustry. The compounds derived from medicinal extracts are often stereochemically complex,multi- or macrocyclic molecules and tend to have interesting biological properties. Despite theseadvantages, the path from traditional medicines/natural products to western pharmaceuticalsposes with challenges along the key steps of development such as western discovery, isolation orsynthesis of active component, elucidation of molecular mechanism, and pharmaceutical
O
H3CO
OH
OCH3
HO
O
O
OCH3
OHO236 237
OCH3
OCH3H3CO
238
H3CO
HO
H3CO
OCH3
OH
OH
O
H3CO
H3CO
HO OH
OCH3
OH
OCH3
241
242
OCH3
H3CO 244
O
O
NH
O
HN
O
OH
OH
245
O
H
H
H
O
OO
H
N
OH
OH246
O
O
O
O
248
OO
CH2
H
HO
O 247
O
O
RO239, R = H240, R = O
O
HO
OH
R2 R1
243, R1 = OH R2 = OCH3
H3CO
HO
NH
O
H3CO
O
HN
H2N O 532432
OH OH
OH
OHO
OH
251
H3CO
OCH3
HO
252
HO
OH
253
HO
OH
254
COOH
249
250
Figure 16. Chemical structures of the miscellaneous compounds.
ANTI-INFLAMMATORYNATURALPRODUCTS K 39
Medicinal Research Reviews DOI 10.1002/med

development. Another important limitation with natural products is insufficient amounts, re-quired for development and clinical use. Alternatives could be chemical synthesis using cost-effective method or develop cell cultures or tran sgenesis to enable scale-up and production.Despite continued advances in synthetic chemistry, the inherent complexicity of natural pro-ducts that is responsible for their biological function can make production difficult. The hugeamount of interdisciplinary research efforts is necessary to utilize the pharmaceutical potentialof natural products. In the light of these considerations, the expectations are high from naturalproducts as a source of effective anti-inflammatory agents, as the current clinically used drugsfor inflammatory disorders are mostly biologic in nature and expensive to be used.
In the present review a variety of natural product compounds derived from plants andmarine organisms reported to possess anti-inflammatory activity during last decade, arebeing described. The major categories of compounds are polyphenolics that modulate theinflammatory pathways. Several polyphenolic compounds such as flavonoids, lignans,phloroglucinols, quinones, stilbenes, phenylpropanoids, and diarylheptanoids exhibited sig-nificant anti-inflammatory activity in vivo as well as in vitro. Among the several flavonoids,luteolin (95), quercetin (96), wogonin (99), apigenin (107), fisetin (111), and baicalein (132)showed anti-inflammatory activity in vitro through various molecular mechanisms and thesewere also reported to possess significant anti-inflammatory activity in animal models ofinflammation. Several SAR studies performed on naturally occurring as well as syntheticflavonoids confirmed these findings. Thus, the polyphenolic compound serves as markers/biomarkers to prepare standardized extracts and dietary supplements, which in turn may beused as promising therapeutic agents for treating inflammatory disorders.
Another prominent category of compounds is terpenoids that have shown marked anti-inflammatory activity. Triptolide (45), a diterpene epoxide inhibited IL-1b gene expression inLPS-stimulated macrophages at nanomolar concentration. Clinical trial on the standardizedextract of Trypterigium wilfordii containing triptolide was conducted in the US for rheu-matoid arthritis, suggesting its potential as an anti-inflammatory agent. Triterpenoids,CDDO (60) and CDDO methyl ester (63) (Oleanolic acid analogs) inhibited NO productionin mouse macrophages at nanomolar concentrations. BA (91) and its analogs present in theclinically used herbal drug Boswellia serrata exhibited potent 5-LOX inhibitory activity.Alkaloids such as rutaecarpine (2), sinomenine (7), and tetrandine (9) identified as an activeconstituents from Chinese medicinal plants emerged as promising anti-inflammatory agentsacting on various molecular targets of inflammation. Resveratrol (162), a specific COX-2inhibitor and effective cancer chemopreventive agent and its hydroxylated derivatives 169
and 171 have been reported as a novel class of COX-2 inhibitors and serves as promisingcandidates for further preclinical and clinical development as anti-inflammatory agents. Overthe last decade curcumin (186) has emerged as a lead compound for development of noveland effective anti-inflammatory and anti-cancer agents as its molecular mechanism/(s) ofaction became better understood through a number of studies. Curcumin suffers from adrawback of poor oral bioavailability; therefore, several SAR studies underway to preparecurcumin analogs with better anti-inflammatory and bioavailability profile. One such SARstudy carried out recently resulted in the identification of a few novel curcumin analogs 195,200, and 201 with enhanced anti-inflammatory activity for further drug development. Ourstudy on curcumin also resulted in the identification of hydrazinocurcumin (189) and theisoxazole analog (190) as lead compounds for further development. Other most activecompounds reported include fatty acid, (S)-coriolic acid (22); panduratin A (104), a chalconecompound; diphyllin acetylapioside (145), a lignan glycoside; quinone compounds maesanin(154), thymohydroquinone (155), and thymoquinone (157); marine natural products man-oalide (222) and its analog 223, cacospongionolides B (226) and scalaradial (229). All thesecompounds exhibited anti-inflammatory activity through different molecular mechanisms at
40 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

sub-micromolar concentrations. Although, these active compounds are not drugs per se, theydeserve further investigation as potential candidates for anti-inflammatory and anti-cancerdrug development through preclinical and clinical studies.
10. ABBREVIATIONS
AA arachidonic acidAP-1 activator protein-1cAMP cyclic adenosine monophosphateCD cluster of differentiation moleculeCOX cyclooxygenaseCTLA cytotoxic T lymphocyte antigenDPP 12-deoxyphorbol-13-phenylacetateDPT 12-deoxyphorbol-13-tetradecanoateEGFR epidermal growth factor receptorEPP ethyl phenylpropiolateERK extracellular signal-regulated kinaseET-1 endothelin-1fMLP N-formyl-methionyl-leucyl phenylalanineHVEC human vascular endothelial cellsICAM-1 intercellular adhesion molecule-1IFN interferonIg immunoglobulinIL interleukiniNOS inducible nitric oxide synthaseJNK c-Jun-N-terminal kinaseLFA-1 leukocyte function-associated antigen-1LOX lipoxygenaseLPS lipopolysaccharideLT leukotrieneMAPK mitogen-activated protein kinaseMMP matrix metalloproteinaseNF-kB nuclear factor-kBNO nitric oxidePBMC peripheral blood mononuclear cellsPG prostaglandinsPL phospholipasePMA phorbol 12-myristate 13-acetatePMNL polymorphonuclear leukocytesRANKL receptor activator of NF-kB ligandRIMEC rat intestinal microvascular endothelial cellsROS reactive oxygen speciesSAR structure–activity relationshipTAK-1 TGF-activated kinase-1TGF transforming growth factorTh 1 T-helper cells 1TNF-a tumor necrosis factor-aTPA 12-O-tetradecanoyl phorbol 13-acetateVLA very late activation antigen
ANTI-INFLAMMATORYNATURALPRODUCTS K 41
Medicinal Research Reviews DOI 10.1002/med

REFERENCES
1. Woolf AD, Pfleger B. Burden of major musculoskeletal conditions. Bull World Health Org
2003;81:646–656.
2. Smith RJ. Therapies for rheumatoid arthritis: Hope springs eternal. Drug Discov Today
2005;10:1598–1606.
3. Mantovani A, Pierotti MA. Cancer and inflammation: A complex relationship. Cancer Lett
2008;267:180–181.
4. Lawrence T. Inflammation and cancer: A failure of resolution? TRENDS Pharmacol Sci
2007;28:162–165.
5. Schimmer BP, Parker KL. Adrenocorticotropic hormone; adrenocortical steroids and their
synthetic analogs; Inhibitors of the synthesis and actions of adrenocortical hormones. In:
Hardman JG, Limbird LE, Gilman AG, editors. Goodman and Gilman’s The pharmacological
basis of therapeutics. 10th Edition. USA: McGraw; 2001. pp 1649–1677.
6. Jachak SM. Cyclooxygenase inhibitory natural products: Current status. Curr Med Chem
2006;13:659–678.
7. Strand V, Kimberly R, Isaacs JD. Biologic therapies in rheumatology: Lessons learned future
directions. Nature Rev Drug Discov 2007;6:75–92.
8. Tincani A, Andreoli L, Bazzani C, Bosiso D, Sozzani S. Inflammatory molecules: A target for the
treatment of systemic autoimmune diseases. Autoimmun Rev 2007;7:1–7.
9. Simmons, DL. What makes a good anti-inflammatory drug target? Drug Discov Today
2006;11:210–219.
10. McCulloch CA, Downey GP, El-Gabalawy H. Signaling platforms that modulate the
inflammatory response: New targets for drug development. Nature Rev Drug Discov
2006;5:864–876.
11. Schmitz ML, Bacher S. Novel molecular targets in the search for anti-inflammatory agents.
Phytochemistry Rev 2005;4:19–25.
12. Hume DA, Fairlie DP. Therapeutic targets in inflammatory disease. Curr Med Chem
2005;12:2925–2929.
13. Roman-Blass JA, Jimenez SA. NF-kB as a therapeutic target in osteoarthritis and rheumatoid
arthritis. Osteoarthritis Cartilage 2006;14:839–848.
14. Newman DJ, Cragg GM. Natural products as sources of new drugs over the last 25 years. J Nat
Prod 2007;70:461–477.
15. Jachak SM, Saklani A. Challenges and opportunities in drug discovery from plants. Curr Sci
2007;92:1251–1257.
16. Vane JR. Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs.
Nat New Biol 1971;231:232–235.
17. Schneider I, Bucar F. Lipoxygenase inhibitors from natural plant sources. Part 1: Medicinal plants
with inhibitory activity on arachidonate 5-lipoxygenase and 5-lipoxygenase/cyclooxygenase.
Phytother Res 2005;19:81–102.
18. Schneider I, Bucar F. Lipoxygenase inhibitors from natural plant sources. Part 2: Medicinal plants
with inhibitory activity on arachidonate 12-lipoxygenase, 15-lipoxygenase and leukotriene
antagonists. Phytother Res 2005;19:263–272.
19. Calixto JB, Otuki MF, Santos ARS. Anti-inflammatory compounds of plant origin. Part I. Action
on arachidonic acid pathway, nitric oxide and nuclear factor-kB (NF-kB). Planta Med
2003;69:973–983.
20. Bremner P, Heinrich M. Natural products and their role as inhibitors of the pro-inflammatory
transcription factor NF-kB. Phytochemistry Rev 2005;4:27–37.
21. Paul AT, Gohil VM, Bhutani KK. Modulating TNF-a signaling with natural products. Drug
Discov Today 2006;11:725–732.
42 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

22. Calixto JB, Campos MM, Otuki MF, Santos ARS. Anti-inflammatory compounds of plant origin.
Part II. Modulation of pro-inflammatory cytokines, chemokines and adhesion molecules. Planta
Med 2004;70:93–103.
23. Rahman I, Biswas SK, Kirkham PA. Regulation of inflammation and redox signaling by dietary
polyphenols. Biochem Pharmacol 2006;72:1439–1452.
24. Bucar F, Jachak SM, Noreen Y, Kartnig T, Perera P, Bohlin L, Schubert-Zsilavecz M.
Amentoflavone from Biophytum sensitivum and its effect on COX-1/COX-2 catalysed
prostaglandin biosynthesis. Planta Med 1998;64:373–374.
25. Selvam C, Jachak SM, Bhutani KK. Cyclooxygenase inhibitory flavonoids from the stem bark of
Semecarpus anacardium Linn. Phytother Res 2004;18:582–584.
26. Selvam C, Jachak SM. A cyclooxygenase (COX) inhibitory biflavonoid from the seeds of
Semecarpus anacardium. J Ethnopharmacol 2004;95:209–212.
27. Selvam C, Jachak SM, Oli RG, Thilagavathi R, Chakraborti AK, Bhutani KK. A new
cyclooxygenase (COX) inhibitory pterocarpan from Indigofera aspalathoides: Structure elucidation
and determination of binding orientations in the active sites of the enzyme by molecular docking.
Tetrahedron Lett 2004;45:4311–4314.
28. Selvam C, Jachak SM, Thilagavathi R, Chakraborti AK. Design, synthesis, biological evaluation
and molecular docking of curcumin analogues as antioxidant, cyclooxygenase inhibitory and anti-
inflammatory agents. Bioorg Med Chem Lett 2005;15:1793–1797.
29. Jachak SM, Selvam C, Srivastava A, Ahuja V. Evaluation of anti-inflammatory activity and
identification of bioactive compounds from Vitex negundo L., Cardiospermum helicacabum L. and
Tridax procumbens L. Planta Med 2007;73:802–803.
30. Danz H, Stoyanova S, Wippich P, Brattstrom A, Hamburger M. Identification and isolation of
the cyclooxygenase-2 inhibitory principle in Isatis tinctoria. Planta Med 2001;67:411–416.
31. Danz H, Baumann D, Hamburger M. Quantitative determination of the dual COX-2/5-LOX
inhibitor tryptanthrin in Isatis tinctoria by ESI-LC-MS. Planta Med 2002;68:152–157.
32. Danz H, Stoyanova S, Thomet OAR, Simon HU, Dannhardt G, Ulbrich H, Hamburger M.
Inhibitory activity of tryptanthrin on prostaglandin and leukotriene synthesis. Planta Med
2002;68:875–880.
33. Ishihara T, Kohno K, Ushio S, Iwaki K, Ikeda M, Kurimoto M. Tryptanthrin inhibits nitric oxide
and prostaglandin E2 synthesis by murine macrophages. Eur J Pharmacol 2000;407:197–204.
34. Micallef MJ, Iwaki K, Ishihara T, Ushio S, Aga M, Kunikata T, Koya-Miyata S, Kimoto T,
Ikeda M, Kurimoto M. The natural plant product tryptanthrin ameliorates dextran sodium
sulfate-induced colitis in mice. Int Immunopharmacol 2002;2:565–578.
35. Heinemann C, Schliemann-Willers S, Oberthur C, Humburger M, Elsner P. Prevention of
experimentally induced irritant contact dermatitis by extracts of Isatis tinctoria compared to pure
tryptanthrin and its impact on UVB-induced erythema. Planta Med 2004;70:385–390.
36. Woo HG, Lee CH, Noh M-S, Lee JJ, Jung Y-S, Baik EJ, Moon C-H, Lee SH. Rutaecarpine, a
quinazolinocarboline alkaloid, inhibits prostaglandin production in RAW264.7 macrophages.
Planta Med 2001;67:505–509.
37. Choi YH, Shin EM, Kim YS, Cai XF, Lee JJ, KimHP. Anti-inflammatory principles from the fruits
of Evodia rutaecarpa and their cellular action mechanisms. Arch Pharm Res 2006;29:293–297.
38. Noh EJ, Ahn KS, Shin EM, Jung SH, Kim YS. Inhibition of lipopolysaccharide-induced iNOS
and COX-2 expression by dehydroevodiamine through suppression of NF-kB activation in RAW
264.7 macrophages. Life Sci 2006;79:695–701.
39. Ko H-C, Wang Y-H, Liou K-T, Chen C-M, Chen C-H, WangW-Y, Chang S, Hou Y-C, Chen K-T,
Chen C-F, Shen Y-C. Anti-inflammatory effects and mechanisms of the ethanol extract of Evodia
rutaecarpa and its bioactive components on neutrophils and microglial cells. Eur J Pharmacol
2007;555:211–217.
40. Moon TC, Murakami M, Kudo I, Son KH, Kim HP, Kang SS, Chang HW. A new class of COX-2
inhibitor, rutaecarpine from Evodia rutaecarpa. Inflamm Res 1999;48:621–625.
ANTI-INFLAMMATORYNATURALPRODUCTS K 43
Medicinal Research Reviews DOI 10.1002/med

41. Shin Y-K, Bae E-A, Cai XF, Lee JJ, Kim D-H. In vitro and in vivo antiallergic effect of the fructus
of Evodia rutaecarpa and its constituents. Biol Pharm Bull 2007;30:197–199.
42. Jeon TW, Jin CH, Lee SK, Jun IH, Kim GH, Lee DJ, Jeong HG, Lee K-B, Jahng Y, Jeong TC.
Immunosuppressive effects of rutaecarpine in female BALB/c mice. Toxicol Lett
2006;164:155–166.
43. Adams M, Kunert O, Haslinger E, Bauer R. Inhibition of leukotriene biosynthesis by quinolone
alkaloids from the fruits of Evodia rutaecarpa. Planta Med 2004;70:904–908.
44. Yarosh DB, Galvin JW, Nay SL, Pena AV, Canning MT, Brown DA. Anti-inflammatory activity
in skin by biomimetic of Evodia rutaecarpa extract from traditional Chinese medicine. J Dermatol
Sci 2006;42:13–21.
45. Wang Y, Fang Y, Huang W, Zhou X, Wang M, Zhong B, Peng D. Effect of sinomenine on
cytokine expression of macrophages and synoviocytes in adjuvant arthritis rats. J Ethnopharma-
col 2005;98:37–43.
46. Feng H, Yamaki K, Takano H, Inoue K, Yanagisawa R, Yoshino S. Effect of sinomenine on
collagen-induced arthritis in mice. Autoimmunity 2007;40:532–539.
47. Cheng H, Xia B, Guo Q, Zhang L, Wang F, Jiang L, Wang Z, Zhang Y, Li C. Sinomenine
attenuates 2,4,6-trinitrobenzene sulfonic acid-induced colitis in mice. Int Immunopharmacol
2007;7:604–611.
48. Chen Y, Yang C, Jin N, Xie Z, Fei L, Jia Z, Wu Y. Sinomenine promotes differentiation but
impedes maturation and co-stimulatory molecule expression of human monocyte-derived
dendritic cells. Int Immunopharmacol 2007;7:1102–1110.
49. Huang J, Lin Z, Luo M, Lu C, Kim MH, Yu B, Gu J. Sinomenine suppresses TNF-a-inducedVCAM-1 expression in human umbilical vein endothelial cells. J Ethnopharmacol 2007;114:
180–185.
50. Zeng Y, Gu B, Ji X, Ding X, Song C, Wu F. Sinomenine, an antirheumatic alkaloid, ameliorates
clinical signs of disease in the lewis rat model of acute experimental autoimmune encephalolmye-
litis. Biol Pharm Bull 2007;30:1438–1444.
51. Zhao Y, Li J, Yu K, Liu Y, Chen X. Sinomenine inhibits maturation of monocyte-derived
dendritic cells through blocking activation of NF-kB. Int Immunopharmacol 2007;7:
637–645.
52. Qian L, Xu Z, Zhang W, Wilson B, Hong J-S, Flood PM. Sinomenine, a natural dextrorotatory
morphinan analogue, is anti-inflammatory and neuroprotective through inhibition of microglial
NADPH oxidase. J Neuroinflamm 2007;4:23–37.
53. Tang Q, Luo J, Zhu Q, Li Y, Yin S. Synthesis and anti-inflammatory activities investigation of
sinomenine derivatives on ring C. Nat Prod Res 2006;20:1015–1023.
54. Choi HS, Kim HS, Min KR, Kim Y, Lim HK, Chang YK, Chung MW. Anti-inflammatory effects
of fangchinoline and tetrandrine. J Ethnopharmacol 2000;69:173–179.
55. Shen YC, Chou CJ, Chiou WF, Chen C. Anti-inflammatory effects of the partially purified extract
of radix Stephaniae tetrandrae: comparative studies of its active principles tetrandrine and
fangchinoline on human polymorphonuclear leukocyte functions. Mol Pharmacol 2001;60:
1083–1090.
56. Wu S-J, Ng L-T. Tetrandrine inhibits proinflammatory cytokines, iNOS and COX-2 expression in
human monocytic cells. Biol Pharm Bull 2007;30:59–62.
57. Ho L-H, Juan T-Y, Chao P, Wu W-L, Chang D-M, Chang S-Y, Lai J-H. Plant alkaloid
tetrandrine downregulates IkBa kinases-IkBa-NF-kB signaling pathway in human peripheral
blood T cell. Br J Pharmacol 2004;143:919–927.
58. Fukuda K, Hibiya Y, Mutoh M, Koshiji M, Akao S, Fujiwara H. Inhibition by berberine of
cyclooxygenase-2 transcriptional activity in human colon cancer cells. J Ethnopharmacol
1999;66:227–233.
59. Kuo C-L, Chi C-W, Liu T-Y. The anti-inflammatory potential of berberine in vitro and in vivo.
Cancer Lett 2003;203:127–137.
44 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

60. Choi BH, Ahn I-S, Kim Y-H, Park J-W, Lee S-Y, Hyun C-K, Do M-S. Berberine reduces the
expression of adipogenic enzymes and inflammatory molecules of 3T3-L1 adipocyte. Exp Mol
Med 2006;38:599–605.
61. Lee CH, Chen J-C, Hsiang C-Y, Wu S-L, Wu H-C, Ho T-Y. Berberine suppresses inflammatory
agents-induced interleukin-1b and tumor necrosis factor-a productions via the inhibition of IkBdegradation in human lung cells. Pharmacol Res 2007;56:193–201.
62. Kim KW, Ha K-T, Park C-S, Jin U-H, Chang HW, Lee I-S, Kim C-H. Polygonum cuspidatum,
compared with baicalin and berberine, inhibits inducible nitric oxide synthase and cycloox-
ygenase-2 gene expressions in RAW 264.7 macrophages. Vascul Pharmacol 2007;47:99–107.
63. Kang BY, Chung SW, Cho D, Kim TS. Involvement of p38 mitogen-activated protein kinase in
the induction of interleukin-12 p40 production in mouse macrophages by berberine, a
benzodioxoloquinolozine alkaloid. Biochem Pharmacol 2002;63:1901–1910.
64. Hu J-P, Nishishita K, Sakai E, Yoshida H, Kato Y, Tsukuba T, Okamoto K. Berberine inhibits
RANKL-induced osteoclast formation and survival through suppressing the NF-kB and Akt
pathways. Eur J Pharmacol 2008;580:70–79.
65. Kupeli E, Kosar M, Yesilada E, Baser KHC, Baser C. A comparative study on the anti-
inflammatory, antinociceptive and antipyretic effects of isoquinoline alkaloids from the roots of
Turkish Berberis species. Life Sci 2002;72:645–657.
66. Lee DU, Kang YJ, Park MK, Lee YS, Seo HG, Kim TS, Kim CH, Chang KC. Effects of 13-alkyl-
substituted berberine alkaloids on the expression of COX-II, TNF-a, iNOS, and IL-12 production
in LPS-stimulated macrophages. Life Sci 2003;73:1401–1412.
67. Chiou WF, Peng CH, Chen CF, Chou CJ. Anti-inflammatory properties of piperlactam S:
Modulation of complement 5a-induced chemotaxis and inflammatory cytokines production in
macrophages. Planta Med 2003;69:9–14.
68. Chiou W-F, Shum AYC, Peng C-H, Chen C-F, Chou C-J. Piperlactam S suppresses macrophage
migration by impeding F-actin polymerization and filopodia extension. Eur J Pharmacol
2003;458:217–225.
69. Yin W, Wang TS, Yin FZ, Cai BC. Analgesic and anti-inflammatory properties of brucine and
brucine N-oxide extracted from seeds of Strychnos nux-vomica. J Ethnopharmacol 2003;88:205–214.
70. Lee BG, Kim HS, Zee OP, Lee KR, Lee HY, Han JW, Lee HW. Suppression of inducible nitric
oxide synthase expression in RAW 264.7 macrophages by two b-carboline alkaloids extracted
from Melia azedarach. Eur J Pharmacol 2000;406:301–309.
71. Ringbom T, Huss U, Stenholm A, Flock S, Skattebol L, Perera P, Bohlin L. COX-2 inhibitory
effects of naturally occurring and modified fatty acids. J Nat Prod 2001;64:745–749.
72. Su B-N, Cuendet M, Farnsworth NR, Fong HHS, Pezzuto JM, Kinghorn AD. Activity-guided
fractionation of the seeds of Ziziphus jujuba using a cyclooxygenase-2 inhibitor assay. Planta Med
2002;68:1125–1128.
73. Jang DS, Cuendet M, Su B-N, Totura S, Riswan S, Fong HHS, Pezzuto JM, Kinghorn AD.
Constituents of the seeds of Hernandia ovigera with inhibitory activity against cyclooxygenase-2.
Planta Med 2004;70:893–896.
74. Shrestha R, Kim S-C, Dyer JM, Dixon RA, Chapman KD. Plant fatty acid (ethanol) amide
hydrolases. Biochim Biophys Acta Mol Cell Biol Lipids 2006;1761:324–334.
75. Marzo VD, Maccarrone M. FAAH and anandamide: is 2-AG really the odd one out? Trends
Pharmacol Sci 2008;29:229–233.
76. Chapman KD. Occurrence, metabolism, and prospective functions of N-acylethanolamines in
plants. Prog Lipid Res 2004;43:302–327.
77. Venables BJ, Waggoner CA, Chapman KD. N-acylethanolamines in seeds of selected legumes.
Phytochemistry 2005;66:1913–1918.
78. Verme JL, Fu J, Astarita G, La Rana G, Russo R, Calignano A, Piomelli D. The nuclear receptor
peroxisome proliferator-activated receptor-a mediates the anti-inflammatory actions of palmi-
toylethanolamide. Mol Pharmacol 2005;67:15–19.
ANTI-INFLAMMATORYNATURALPRODUCTS K 45
Medicinal Research Reviews DOI 10.1002/med

79. Shen CC, Wang Y-H, Chang T-T, Lin L-C, Don M-J, Hou Y-C, Lio K-T, Chang S, Wang W-Y,
Ko H-C, Shen Y-C. Anti-inflammatory ergostanes from the basidiomata of Antrodia salmonea.
Planta Med 2007;73:1208–1213.
80. Manjula N, Gayathri B, Vinaykumar KS, Shankernarayanan NP, Vishwakarma RA,
Balakrishnan A. Inhibition of MAP kinases by crude extract and pure compound isolated from
Commiphora mukul leads to down regulation of TNF-a, IL-1b and IL-2. Int Immunopharmacol
2006;6:122–132.
81. Singh BB, Mishra LC, Vinjamury SP, Aquilina M, Singh VJ, Shepard N. The effectiveness of
Commiphora mukul for osteoarthritis of the knee: An outcomes study. Altern Ther Health Med
2003;9:74–79.
82. Khanna D, Sethi G, Ahn KS, Pandey MK, Kunnumakara AB, Sung B, Aggarwal A, Aggarwal
BB. Natural products as a gold mine for arthritis treatment. Curr Opin Pharmacol
2007;7:344–351.
83. Bai L, Wang L, Zhao M, Toki A, Hasegawa T, Ogura H, Kataoka T, Hiroe K, Sakai J, Bai J,
Ando M. Bioactive pregnanes from Nerium oleander. J Nat Prod 2007;70:14–18.
84. Giang PM, Jin HZ, Son PT, Lee JH, Hong YS, Lee JJ. ent-Akurane diterpenoids from Croton
tonkinensis inhibit LPS-induced NF-kB activation and NO production. J Nat Prod
2003;66:1217–1220.
85. Hong SS, Lee SA, Han XH, Jin HZ, Lee JH, Lee D, Lee JJ, Hong JT, Kim Y, Ro JS, Hwang BY.
Akurane diterpenoids from Isodon excisus inhibit LPS-induced NF-kB activation and NO
production in macrophage RAW264.7 cells. J Nat Prod 2007;70:632–636.
86. Perez-Garcia F, Martin E, Parella T, Adzet T, Canigueral S. Activity of taraxasteryl acetate on
inflammation and heat shock protein synthesis. Phytomedicine 2005;12:278–284.
87. Suksamrarn A, Kumpun S, Kirtikara K, Yingyongnarongkul, Suksamrarn S. Iridoids with anti-
inflamatory activity from Vitex peduncularis. Planta Med 2002;68:72–73.
88. Yoshikawa K, Inoue M, Mastumoto Y, Sakkakibara C, Miyataka H, Mastumoto H, Arihara S.
Lanostane triterpenoids and triterpene glycosides from the fruit body of Fomitopsis pinicola and
their inhibitory activity against COX-1 and COX-2. J Nat Prod 2005;68:69–73.
89. Chao K-P, Hua K-F, Hsu H-Y, Su Y-C, Chang S-T. Anti-inflammatory activity of Sugiol, a
diterpene isolated from Calocedrus formosana bark. Planta Med 2005;71:300–305.
90. Ma J, Dey M, Yang H, Poulev A, Pouleva R, Dorn R, Lipsky PE, Kennelly EJ, Raskin I. Anti-
inflammatory and immunosuppressive compounds from Tripterygium wilfordii. Phytochemistry
2007;68:1172–1178.
91. Wu SX, Guo NR. Clinical observation on effect of triptolide tablet in treating patients with
psoriasis vulgaris. Chin J Integr Med 2005;11:147–148.
92. Corson TW, Crews CM. Molecular understanding and modern application of traditional
medicines: Triumphs and trials. Cell 2007;130:769–774.
93. Sethi G, Ahn KS, Pandey MK, Aggarwal BB. Celastrol, a novel triterpene, potentiates TNF-
induced apoptosis and suppresses invasion of tumor cells by inhibiting NF-kB-regulated gene
products and TAK1-mediated NF-kB activation. Blood 2007;109:2727–2735.
94. Tao X, Younger J, Fan FZ, Wang B, Lipsky PE. Benefit of an extract of Tripterygium wilfordii
Hook F in patients with rheumatoid arthritis: A double-blind, placebo-controlled study. Arthritis
Rheum 2002;46:1735–1743.
95. Heinrich M. Ethnopharmacology of Mexican asteraceae (compositae). Annu Rev Pharmacol
Toxicol 1998;38:539–565.
96. Kwok BHB, Koh B, Ndubuisi MI, Elofsson M, Crews CM. The anti-infammatory natural
product parthenolide from the medicinal herb Feverfew directly binds to and inhibits IkB kinase.
Chem Biol 2001;8:759–766.
97. Lindenmeyer MT, Kern C, Sparna T, Donauer J, Wilpert J, Schwager J, Porath D, Kreutz C,
Timmer J, Merfort I. Microarray analysis reveals influence of the sesquiterpene lactone
parthenolide on gene transcription profiles in human epithelial cells. Life Sci 2007;80:1608–1618.
46 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

98. Koo TH, Lee J-H, Park YJ, Hong Y-S, Kim HS, Kim K-W, Lee, JJ. A sesquiterpene lactone,
costunolide, from Magnolia grandiflora inhibits NF-kB by targeting IkB phosphorylation. Planta
Med 2001;67:103–107.
99. Castro V, Murillo R, Klaas CA, Meunier C, Mora G, Pahl HL, Merfort I. Inhibition of the
transcription factor NF-kB by sesquiterpene lactones from Podachaenium eminens. Planta Med
2000;66:591–595.
100. Bocca C, Gabriel L, Bozzo F, Miglietta A. A sesquiterpene lactone, costunolide, interacts with
microtubule protein and inhibits the growth of MCF-7 cells. Chem-Biol Interact 2004;147:79–86.
101. LyX G, Schmidt TJ, Merfort I, Pahl HL. Helenalin, an anti-inflammatory sesquiterpene lactone
from Arnica, selectively inhibits transcription factor NF-kB. Biol Chem 1997;378:951–961.
102. LyX G, Knorre A, Schmidt TJ, Merfort I, Pahl HL. The anti-inflammatory sesquiterpene lactone
helenalin inhibits the transcription factor NF-kB by directly targeting p65. J Biol Chem
1998;273:33508–33516.
103. Tornhamre S, Schmidt TJ, Nasman-Glaser B, Ericssona I, Lindgren JA. Inhibitory effects of
helenalin and related compounds on 5-lipoxygenase and leukotriene C4 synthase in human blood
cells. Biochem Pharmacol 2001;62:903–911.
104. De Marino S, Borbone N, Zollo F, Lanaro A, Di Meglio, Iorizzi M. New sesquiterpene lactones
from Laurus nobilis leaves as inhibitors of nitric oxide production. Planta Med 2005;71:
706–710.
105. Siedle B, Gustavsson L, Johansson S, Murillo R, Castro V, Bohlin L, Merfort I. The effect of
sesquiterpene lactones on the human neutrophil elastase. Biochem Pharmacol 2003;65:897–903.
106. Rungeler P, Castro V, Mora G, Goren N, Vichnewski W, Pahl HL, Merfort I, Schmidt TJ.
Inhibition of transcription factor NF-kB by sesquiterpene lactones: A proposed molecular
mechanism of action. Bioorg Med Chem 1999;7:2343–2352.
107. Liu J. Oleanolic acid and ursolic acid: Research perspectives. J Ethnopharmacol 2005;100:92–94.
108. Subbaramaiah K, Michaluart P, Sporn MB, Dannenberg AJ. Ursolic acid inhibits cycloox-
ygenase-2 transcription in human mammary epithelial cells. Cancer Res 2000;60:2399–2404.
109. Sosa S, Altinier G, Politi M, Braca A, Morelli I, Loggia RD. Extracts and constituents of
Lavandula multifida with topical anti-inflammatory activity. Phytomedicine 2005;12:271–277.
110. Baricevic D, Sosa S, Loggia RD, Tubaro A, Simonovska B, Krasna A, Zupancic A. Topical anti-
inflammatory activity of Salvia officinalis L. leaves: The relevance of ursolic acid.
J Ethnopharmacol 2001;75:125–132.
111. Alvarez ME, Rotelli AE, Pelzer LE, Saad JR, Giordano O. Phytochemical study and anti-
inflammatory properties of Lampaya hieronymi Schum. ex Moldenke. IL Farmaco 2000;55:
502–505.
112. Giner-Larza EM, Manez S, Recio MC, Giner RM, Prieto JM, Cerda-Nicolas M, Rios JL.
Oleanonic acid, a 3-oxotriterpene from Pistacia, inhibits leukotriene synthesis and has anti-
inflammatory activity. Eur J Pharmacol 2001;428:137–143.
113. Honda T, Finlay HJ, Gribble GW, Suh N, Sporn MB. New enone derivatives of oleanolic acid
and ursolic acid as inhibitors of nitric oxide production in mouse macrophages. Bioorg Med Chem
Lett 1997;7:1623–1628.
114. Honda T, Rounds BV, Gribble GW, Suh N, Wang Y, Sporn MB. Design and synthesis of 2-
cyano-3,12-dioxooolean-1,9-dien-28-oic acid, a novel and highly active inhibitors of nitric oxide
production in mouse macrophages. Bioorg Med Chem Lett 1998;8:2711–2724.
115. Honda T, Rounds BV, Bore L, Jr., Favaloro FG, Gribble GW, Suh N, Wang Y, Sporn MB.
Novel synthetic oleanane triterpenoids: A series of highly active inhibitors of nitric oxide
production in mouse macrophages. Bioorg Med Chem Lett 1999;9:3429–3434.
116. Honda T, Rounds BV, Bore L, Finlay HJ, Jr., Favaloro FG, Suh N, Wang Y, Sporn MB,
Gribble GW. Synthetic oleanane and ursane triterpenoids with modified ring A and C: A series of
highly active inhibitors of nitric oxide production in mouse macrophages. J Med Chem 2000;43:
4233–4246.
ANTI-INFLAMMATORYNATURALPRODUCTS K 47
Medicinal Research Reviews DOI 10.1002/med

117. Honda T, Gribble GW, Suh N, Finlay HJ, Rounds BV, Bore L, Jr., Favaloro FG, Wang Y, Sporn
MB. Novel synthetic oleanane and ursane triterpenoids with various enone functionalities in ring
A as inhibitors of nitric oxide production in mouse macrophages. J Med Chem
2000;43:1866–1877.
118. Honda T, Honda Y, Jr., Favaloro FG, Gribble GW, Suh N, Place AE, Rendi MH, Sporn MB. A
novel dicyanotriterpenoid, 2-Cyano-3,12-dioxooleana-1,9(11)-dien-28-onitrile, active at picomolar
concentrations for inhibition of nitric oxide production. Bioorg Med Chem Lett
2002;12:1027–1030.
119. http://www.clinicaltrials.gov/ct2/show/NCT00352040?term5CDDO&rank5 1 (accessed on 8-2-
2009, last updated on 6-2-2009).
120. Cichewicz RH, Kouzi SA. Chemistry, biological activity, and chemotherapeutic potential of
betulinic acid for the prevention and treatment of cancer and HIV infection. Med Res Rev
2004;24:90–114.
121. Sami A, Taru M, Salme K, Jari Y-K. Pharmacological properties of the ubiquitous natural
product betulin. Eur J Pharm Sci 2006;29:1–13.
122. Yogeeswari P, Sriram D. Betulinic acid and its derivatives: A review on their biological properties.
Curr Med Chem 2005;12:657–666.
123. Bernard P, Scior T, Didier B, Hibert M, Berthon J. Ethnopharmacology and bioinformatic
combination for leads discovery: Application to phospholipase A2 inhibitors. Phytochemistry
2001;58:865–874.
124. Takada Y, Aggarwal BB. Betulinic acid suppresses carcinogen-induced NF-kB activation through
inhibition of IkBa kinase and p65 phosphorylation: Abrogation of cyclooxygenase-2 and matrix
metalloprotease-9. J Immunol 2003;171:3278–3286.
125. Ryu SY, Oak M-H, Yoon S-K, Cho D-I, Yoo G-S, Kim T-S, Kim K-M. Anti-allergic and anti-
inflammatory triterpenes from the herb of Prunella vulgaris. Planta Med 2000;66:358–360.
126. Huguet A-I, Recio MC, Manez S, Giner RM, Rios J-L. Effect of triterpenoids on the
inflammation induced by protein kinase C activators, neuronally acting irritants and other agents.
Eur J Pharmacol 2000;410:69–81.
127. Honda T, Liby KT, Su X, Sundararajan C, Honda Y, Suh N, Risingsong R, Williams CR,
Royce DB, Sporn MB, Gribble GW. Design, synthesis, and anti-inflammatory activity both
in vitro and in vivo of new betulinic acid analogues having an enone functionality in ring A. Bioorg
Med Chem Lett 2006;16:6306–6309.
128. Poeckel D, Werz O. Boswellic Acids: Biological actions and molecular targets. Curr Med Chem
2006;13:3359–3369.
129. Ammon HPT. Boswellic acid in chronic inflammatory diseases. Planta Med 2006;72:
1100–1116.
130. Safayhi H, Mack T, Sabieraj J, Anazodo MI, Subramanian LR, Ammon HPT. Boswellic acids:
Novel, specific, nonredox inhibitors of 5-lipoxygenase. J Pharmacol Exp Ther 1992;261:
1143–1146.
131. Sailer ER, Subramanian LR, Rall B, Hoernlein RF, Ammon HPT, Safayhi H. Acetyl-11-keto-
beta-boswellic acid (AKBA): Structure requirements for binding and 5-lipoxygenase inhibitory
activity. Br J Pharmacol 1996;117:615–618.
132. Poeckel D, Tausch L, Kather N, Jauch J, Werz O. Boswellic acids stimulate arachidonic acid
release and 12-lipoxygenase activity in human platelets independent of Ca21 and differentially
interact with platelet-type 12-lipoxygenase. Mol Pharmacol 2006;70:1071–1078.
133. Siemoneit U, Hofmann B, Kather N, Lamkemeyer T, Madlung J, Franke L, Schneider G, Jauch J,
Poeckel D, Werz O. Identification and functional analysis of cyclooxygenase-1 as a molecular
target of boswellic acids. Biochem Pharmacol 2008;75:503–513.
134. Singh S, Khajuria A, Taneja SC, Khajuria RK, Singh J, Qazi GN. Boswellic acids and
glucosamine show synergistic effect in preclinical anti-inflammatory study in rats. Bioorg Med
Chem Lett 2007;17:3706–3711.
48 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

135. Kimmatkar N, Thawani V, Hingorani L, Khiyani R. Efficacy and tolerability of Boswellia serrata
extract in treatment of osteoarthritis of knee—a randomized double blind placebo controlled trial.
Phytomedicine 2003;10:3–7.
136. Gupta I, Parihar A, Malhotra P, Gupta S, Ludtke R, Safayhi H, Ammon HPT. Effects of gum
resin of Boswellia serrata in patients with chronic colitis. Planta Med 2001;67:391–405.
137. Ende C, Gebhardt R. Inhibition of matrix metalloproteinase-2 and -9 activities by selected
flavonoids. Planta Med 2004;70:1006–1008.
138. Ziyan L, Yongmei Z, Nan Z, Ning T, Bohlin L. Evaluation of the anti-inflammatory activity of
luteolin in experimental animal models. Planta Med 2007;73:221–226.
139. Jang J, Kim HP, Park H. Structure and anti-inflammatory activity relationships of wogonin
derivatives. Arch Pharm Res 2005;28:877–884.
140. Yamaki K, Kim DH, Ryu N, Kim YP, Shin KH, Ohuchi K. Effects of naturally occurring
isoflavones on prostaglandin E2 production. Planta Med 2002;68:97–100.
141. Laupattarakasem P, Houghton PJ, Hoult JR. Anti-inflammatory isoflavonoids from the stems of
Derris scandens. Planta Med 2004;70:496–501.
142. Min B, Oh SR, Lee HK, Takatsu K, Chang IM, Min KR, Kim Y. Sophoricoside analogues as the
IL-5 inhibitors from Sophora japonica. Planta Med 1999;65:408–412.
143. Tuchinda P, Reutrakul V, Claeson P, Pongprayoon U, Sematong T, Santisuk T, Taylor WC.
Anti-inflammatory cyclohexenyl chalcone derivatives in Boesenbergia pandurata. Phytochemistry
2002;59:169–173.
144. Yun J-M, Kwon H, Hwang J-K. In vitro anti-inflammatory activity of panduratin A isolated from
Kaemferia pandurata in RAW264.7 cells. Planta Med 2003;69:1102–1108.
145. Chung M-I, Weng J-R, Wang J-P, Teng C-M, Lin C-N. Antiplatelet and anti-inflammatory
constituents and new oxygenated xanthones from Hypericum geminiflorum. Planta Med
2002;68:25–29.
146. Ha SK, Lee P, Park JA, Oh HR, Lee SY, Park J-H, Lee EH, Ryu JH, Lee KR, Kim SY. Apigenin
inhibits the production of NO and PGE2 in microglia and inhibits neuronal cell death in a
middle cerebral artery occlusion-induced focal ischemia mice model. Neurochem Int 2008;52:
878–886.
147. Lee J-H, Zhou HY, Cho SY, Kim YS, Lee YS, Jeong CS. Anti-inflammatory mechanisms of
apigenin: Inhibition of cyclooxygenase-2 expression, adhesion of monocytes to human umbilical
vein endothelial cells, and expression of cellular adhesion molecules. Arch Pharm Res
2007;30:1318–1327.
148. Nicholas C, Batra S, Vargo MA, Voss OH, Gavrilin MA, Wewers MD, Guttridge DC, Grotewold E,
Doseff AI. Apigenin blocks lipopolysaccharide-induced lethality in vivo and pro-inflammatory
cytokines expression by inactivating NF-kB through the suppression of p65 phosphorylation.
J Immunol 2007;179:7121–7127.
149. Kim HP, Son KH, Chang HW, Kang SS. Anti-inflammatory plant flavonoids and cellular action
mechanisms. J Pharmacol Sci 2004;96:229–245.
150. Kim HK, Cheon BS, Kim YH, Kim SY, Kim HP. Effects of naturally occurring flavonoids on
nitric oxide production in the macrophage cell line RAW 264.7 and their structure-activity
relationships. Biochem Pharmacol 1999;58:759–765.
151. Matsuda H, Morikawa T, Ando S, Toguchida I, Yoshikawa M. Structural requirements of
flavonoids for nitric oxide production inhibitory activity and mechanism of action. Bioorg Med
Chem 2003;11:1995–2000.
152. Matsuda H, Morikawa T, Ueda K, Managi H, Yoshikawa M. Structural requirements of
flavonoids for inhibition of antigen-induced degranulation, TNF-a and IL-4 production from
RBL-2H3 cells. Bioorg Med Chem 2002;10:3123–3128.
153. Taleb-Contini SH, Kanashiro A, Kabeya LM, Polizello ACM, Lucisano-Valim YM, Oliveira
DCR. Immunomodulatory effects of methoxylated flavonoids from two Chromolaena species:
Structure-activity relationships. Phytother Res 2006;20:573–575.
ANTI-INFLAMMATORYNATURALPRODUCTS K 49
Medicinal Research Reviews DOI 10.1002/med

154. Chen C-C, Chow M-P, Huang W-C, Lin Y-C, Chang Y-J. Flavonoids inhibit tumor necrosis
factor-a-induced upregulation of intercellular adhesion molecule-1 (ICAM-1) in respiratory
epithelial cells through activator protein-1 and nuclear factor-kB: Structure-activity relationships.
Mol Pharmacol 2004;66:683–693.
155. Lotito SB, Frei B. Dietary flavonoids attenuate tumor necrosis factor a-induced adhesion
molecule expression in human aortic endothelial cells. Structure–function relationship and activity
after first pass metabolism. J Biol Chem 2006;281:37102–37110.
156. Jung S-H, Cho S-H, Dang TH, Lee J-H, Ju J-H, Kim M-K, Lee S-H, Ryu J-C, Kim Y. Structural
requirement of isoflavonones for the inhibitory activity of interleukin-5. Eur J Med Chem
2003;38:537–545.
157. Sadik CD, Sies H, Schewe T. Inhibition of 15-lipoxygenases by flavonoids: Structure-activity
relations and mode of action. Biochem Pharmacol 2003;65:773–781.
158. Martinez YV, Ohri RV, Kenyon V, Holman TR, Boza SS. Structure-activity relationship studies
of flavonoids as potent inhibitors of human platelet 12-hLO, reticulocyte 15-hLO-1, and prostate
epithelial 15-hLO-2. Bioorg Med Chem 2007;15:7408–7425.
159. Ishikawa YT, Goto M, Yamaki K. Structure–activity relations of inhibitory effects of various
flavonoids on lipopolysaccharide-induced prostaglandin E2 production in rat peritoneal
macrophages: Comparison between subclasses of flavonoids. Phytomedicine 2006;13:310–317.
160. Chen J, Chang HW, Kim HP, Park H. Synthesis of phospholipase A2 inhibitory biflavonoids.
Bioorg Med Chem Lett 2006;16:2373–2375.
161. Agli MD, Bellosta S, Rizzi L, Galli GV, Canavesi M, Rota F, Parente R, Bosisio E, Romeo S.
A structure-activity study for the inhibition of metalloproteinase-9 activity and gene expression by
analogues of gallocatechin-3-gallate. Cell Mol Life Sci 2005;62:2896–2903.
162. Sartor L, Pezzato E, Aica ID, Caniato R, Biggin S, Garbisa S. Inhibition of matrix-proteases by
polyphenols: Chemical insights for anti-inflammatory and anti-invasion drug design. Biochem
Pharmacol 2002;64:229–237.
163. Kang BY, Chung SW, Kim SH, Cho D, Kim TS. Involvement of nuclear factor-kappaB in the
inhibition of interleukin-12 production from mouse macrophages by baicalein, a flavonoid in
Scutellaria baicalensis. Planta Med 2003;69:687–691.
164. Deschamps JD, Kenyon VA, Holman TR. Baicalein is a potent in vitro inhibitor against both
reticulocyte 15-human and platelet 12-human lipoxygenases. Bioorg Med Chem
2006;14:4295–4301.
165. Huang W-H, Lee A-R, Chien P-Y, Chou T-Z. Synthesis of baicalein derivatives as potential anti-
aggregatory and anti-inflammatory agents. J Pharm Pharmacol 2005;57:219–225.
166. Cho H, Yun C-W, Park W-K, Kong J-Y, Kim KS, Park Y, Lee S, Kim B-K. Modulation of the
activity of pro-inflammatory enzymes, COX-2 and iNOS, by chrysin derivatives. Pharmacol Res
2004;49:37–43.
167. Cho JY, Baik KU, Yoo ES, Yoshikawa K, Park MH. In vitro anti-inflammatory effects
of neolignan woorenosides from the rhizomes of Coptis japonica. J Nat Prod 2000;63:
1205–1209.
168. Cho JY, Kim AR, Park MH. Lignans from the rhizomes of Coptis japonica differentially act as
anti-inflammatory principles. Planta Med 2001;67:312–316.
169. Saleem M, Kim HJ, Ali MS, Lee YS. An update on bioactive plant lignans. Nat Prod Rep
2005;22:696–716.
170. Prieto JM, Giner RM, Recio MC, Schinella G, Manez S, Rios JL. Diphyllin acetylapioside, a
5-lipoxygenase inhibitor from Halophyllum hispanicum. Planta Med 2002;68:359–360.
171. Hwang BY, Lee J-H, Nam JB, Hong Y-S, Lee JJ. Lignans from Saururus chinensis inhibiting the
transcription factor NF-kB. Phytochemistry 2003;64:765–771.
172. Zschocke S, van Staden J, Paulus K, Bauer R, Horn MM, Munro OQ, Brown NJ, Drewes SE.
Stereostructure and anti-inflammatory activity of three diastereomers of ocobullenone from
Ocotea bullata. Phytochemistry 2000;54:591–595.
50 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

173. Weng J-R, Tsao L-T, Wang J-P, Wu R-R, Lin C-N. Anti-inflammatory phloroglucinols and
terpenoids from Garcinia subelliptica. J Nat Prod 2004;67:1796–1799.
174. Appendino G, Ottino M, Marquez N, Bianchi F, Giana A, Ballero M, Sterner O, Fiebich BL,
Munoz E. Arzanol, an anti-inflammatory and anti-HIV phloroglucinol a-pyrone from
Helichrysum italicum ssp. microphyllum. J Nat Prod 2007;70:608–612.
175. Kang BK, Chung SW, Kim TS. Inhibition of interleukin-12 production in lipopolysaccharide-
activated mouse macrophages by hypericin, an active component of Hypericum perforatum. Planta
Med 2001;67:364–366.
176. Wube AA, Bucar F, Asres K, Gibbons S, Adams M, Streit B, Bodensieck A, Bauer R.
Knipholone, a selective inhibitor of leukotriene metabolism. Phytomedicine 2006;13:452–456.
177. Abourashed EA, Muhammad I, Resch M, Bauer R, El-Feraly FS, Hufford CD. Inhibitory effects
of maesanin and analogues on arachidonic acid metabolizing enzymes. Planta Med
2001;67:360–361.
178. Marsik P, Kokoska L, Landa P, Nepovim A, Soudek P, Vanek T. In vitro effects of thymol and
quinones of Nigella sativa seeds on cyclooxygenase-1 and -2 catalysed prostaglandin E2
biosynthesis. Planta Med 2005;71:739–742.
179. Aggarwal BB, Shishodia S. Molecular targets of dietary agents for prevention and therapy of
cancer. Biochem Pharmacol 2006;71:1397–1421.
180. Folmer F, Jaspars M, Dicato M, Diederich M. Marine natural products as targeted modulators of
the transcription factor NF-kB. Biochem Pharmacol 2008;75:603–617.
181. Huang Q, Shen H-M, Ong C-N. Inhibitory effect of emodin on tumor invasion through
suppression of activator protein-1 and nuclear factor-kB. Biochem Pharmacol 2004;68:361–371.
182. Matsui T, Ito C, Itoigawa M, Okada T, Furukawa H. Anti-inflammatory activity of
phenylpropanoids and plastoquinoids from Illicium species in RBL-2H3 cells. Planta Med
2007;73:662–665.
183. Lee JY, Woo E-R, Kang KW. Inhibition of lipopolysaccharide-inducible nitric oxide synthase
expression by acteoside through blocking of AP-1 activation. J Ethnopharmacol 2005;97:
561–566.
184. Baur JA, Sinclair DA. Therapeutic potential of resveratrol: The in vivo evidence. Nat Rev Drug
Discov 2006;5:493–506.
185. Bi XL, Yang JY, Dong YX, Wang JM, Cui YH, Ikeshima T, Zhao YQ, Wu CF. Resveratrol
inhibits nitric oxide and TNF-a production by lipopolysaccharide-activated microglia. Int
Immunopharmacol 2005;5:185–193.
186. Csaki C, Keshishzadeh N, Fischer K, Shakibaei M. Regulation of inflammation signaling by
resveratrol in human chondrocytes in vitro. Biochem Pharmacol 2008;75:677–687.
187. Kundu JK, Shin YK, Surh Y-J. Resveratrol modulates phorbol ester-induced pro-inflammatory
signal transduction pathways in mouse skin in vivo: NF-kB and AP-1 as prime targets. Biochem
Pharmacol 2006;72:1506–1515.
188. Wu CC, Wu CI, Wang WY, Wu YC. Low concentrations of resveratrol potentiate the antiplatelet
effect of prostaglandins. Planta Med 2007;73:439–443.
189. Elmali N, Baysai O, Harma A, Esenkaya I, Mizrak B. Effects of resveratrol in inflammatory
arthritis. Inflammation 2007;1–2:1–6.
190. Elmali N, Esenkaya I, Harma A, Ertem K, Turkoz Y, Mizrak B. Effect of resveratrol in
experimental osteoarthritis in rabbits. Inflamm Res 2005;54:158–162.
191. Chung K-O, Kim B-Y, Lee M-H, Kim Y-R, Chung H-Y, Park J-H, Moon J-O. In-vitro and
in vivo anti-inflammatory effect of oyresveratrol from Morus alba L. J Pharm Pharmacol
2003;55:1695–1700.
192. Patel B, Patel S, Hoffman R. Inhibition of cyclooxygenase-2 expression in mouse macrophages
by 4-(3-methyl-but-1-enyl)-3,5,30,40-tetrahydroxystilbene, a resveratrol derivative from peanuts.
Phytother Res 2005;19:552–555.
ANTI-INFLAMMATORYNATURALPRODUCTS K 51
Medicinal Research Reviews DOI 10.1002/med

193. Chung EY, Kim BH, Lee MK, Yun Y-P, Lee SH, Min KR, Kim Y. Anti-inflammatory effect of
the oligomeric stilbene a-Viniferin and its mode of action through inhibition of cyclooxygenase-2
and inducible nitric oxide synthase. Planta Med 2003;69:710–714.
194. Lee JY, Kim J-H, Kang SS, Bae CS, Choi SH. The effects of a-Viniferin on adjuvant-induced
arthritis in rats. Am J Chin Med 2004;32:521–530.
195. Murias M, Handler N, Erker T, Pleban K, Ecker G, Saiko P, Szekeresb T, Jager W. Resveratrol
analogues as selective cyclooxygenase-2 inhibitors: Synthesis and structure–activity relationship.
Bioorg Med Chem 2004;12:5571–5578.
196. Handler N, Brunhofer G, Studenik C, Leisser K, Jager W, Parth S, Erker T. ‘‘Bridged’’ stilbene
derivatives as selective cyclooxygenase-1 inhibitors. Bioorg Med Chem 2007:15:6109–6118.
197. Cho D-I, Koo N-Y, Chung WJ, Kim T-S, Ryu SY, Im SY, Kim K-M. Effects of resveratrol-
related hydroxystilbenes on the nitric oxide production in macrophage cells: Structural
requirements and mechanism of action. Life Sci 2002;71:2071–2082.
198. Goel A, Kunnumakkara AB, Aggarwal BB. Curcumin as ‘‘Curecumin’’: From kitchen to clinic.
Biochem Pharmacol 2008;75:787–809.
199. Singh S. From exotic spice to modern drug? Cell 2007;130:765–768.
200. Maheshwari RK, Singh AK, Gaddipati J, Srimal RC. Multiple biological activities of curcumin: A
short review. Life Sci 2006;78:2081–2087.
201. Chattopadhyay I, Biswas K, Bandyopadhyay U, Banerjee RK. Turmeric and curcumin: Biological
actions and medicinal applications. Curr Sci 2004;87:44–53.
202. Natrajan C, Bright JJ. Curcumin inhibits experimental allergic encephalomyelitis by blocking IL-12
signaling through janus kinase-ATAT pathway in T lympbocytess. J Immunol 2002;169:6503–6513.
203. Liacini A, Sylvester J, Qing LW, Huang W, Dehnade F, Ahmad M, Zafarullah M. Induction of
matrix metalloproteinase-13 gene expression by TNF-alpha is mediated by MAP kinases, AP-1,
and NF-B transcription factors in articular chondrocytes. Exp Cell Res 2003;25:208–217.
204. Banerjee M, Tripathi LM, Srivastava VM, Puri A, Shukla R. Modulation of inflammatory
mediators by ibuprofen and curcumin treatment during chronic inflmammation in rat.
Immunopharmacol Immunotoxicol 2003;25:213–224.
205. Durgaprasad S, Pai CG, Vasanthkumar, Alvres JF, Namitha S. A pilot study of the antioxidant
effect of curcumin in tropical pancreatitis. Indian J Med Res 2005;122:315–318.
206. Lee S-L, Huang W-J, Lin WW, Lee S-S, Chen C-H. Preparation and anti-inflammatory activities
of diarylheptanoid and diarylheptylamine analogues. Bioorg Med Chem 2005;13:6175–6181.
207. Lee M-W, Kim N-Y, Park M-S, Ahn K-H, Toh S-H, Hahn D-R, Kim Y-C, Chung H-T.
Diarylheptanoids with in vitro inducible nitric oxide synthesis inhibitory activity from Alnus
hirsuta. Planta Med 2000;66:551–553.
208. Gafner S, Lee S-K, Cuendet M, Barthelemy S, Vergnes L, Labidalle S, Mehta RG, Boone CW,
Pezzuto JM. Biologic evaluation of curcumin and structural derivatives in cancer chemopreven-
tion model systems. Phytochemistry 2004;65:2849–2859.
209. Handler N, Jaeger W, Puschacher H, Leisser K, Erker T. Synthesis of novel curcumin analogues
and their evaluation as selective cyclooxygenase-1 (COX-1) inhibitors. Chem Pharm Bull
2007;55:64–71.
210. Liang G, Li X, Chen L, Yang S, Wu X, Studer E, Gurley E, Hylemon PB, Ye F, Li Y, Zhou H.
Synthesis and anti-inflammatory activities of mono-carbonyl analogues of curcumin. Bioorg Med
Chem Lett 2008;18:1525–1529.
211. Pearce An, Chia EW, Berridge MW, Clark GR, Harper JL, Larsen L, Maas EW, PageMJ, Perry NB,
Webb VL, Copp BR. Anti-inflammatory thiazine alkaloids isolated from the New Zealand ascidian
Alpidium sp.: Inhibitors of the neutrophil respiratory burst in a model of gouty arthritis. J Nat Prod
2007;70:936–940.
212. Harinantenaina L, Asakawa Y. Chemical constituents of Malagasy liverworts. 6. A myltaylane
caffeate with nitric oxide inhibitory activity from Bazzania nitida. J Nat Prod 2007;70:856–858.
52 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med

213. Nakao Y, Fusetani N. Enzyme inhibitors from marine invertebrates. J Nat Prod 2007;70:689–710.
214. Shen Y-C, Chen S-Y, Kuo Y-H, Huang T-L, Chiang MY, Khalil AT. Chemical transformation
and biological activities of ambrein, a major product of ambergris from Physeter macrocephalus
(Sperm Whale). J Nat Prod 2007;70:147–153.
215. Fujita M, Nakao Y, Matsunaga S, Nishikawa T, Fusetani N. Sodium 1-(12-hydroxy)octadecanyl
sulfate, an MMP-2 inhibitor, isolated from tunicate of the family polyclinidae. J Nat Prod
2002;65:1936–1938.
216. Ortiz AR, Pisabarro MT, Gago F. Molecular model of the interaction of bee venom
phospholipase A2 with manoalide. J Med Chem 1993;36:1866–1879.
217. Newman DJ, Cragg GM. Advanced preclinical and clinical trials of natural products and related
compounds from marine sources. Curr Med Chem 2004;11:1693–1713.
218. De Rosa M, Giordano S, Scettri A, Sodano G, Soriente A, Pastor PG, Alcaraz MJ, Paya M.
Synthesis and comparison of the antiinflammatory activity of manoalide and cacospongionolide B
analogues. J Med Chem 1998;41:3232–3238.
219. Giannini C, Debitus C, Lucas R, Ubeda A, Paya M, Hooper JNA, D’Auria MV. New
sesquiterpene derivatives from the sponge Dysidea species with a selective inhibitor profile against
human phospholipase A2 and other leukocyte functions. J Nat Prod 2001;64:612–615.
220. Monti MC, Casapullo A, Cavasotto CN, Napolitano A, Riccio R. Scalaradial, a dialdehyde-
containing marine metabolite that causes an unexpected noncovalent PLA2 inactivation.
Chembiochem 2007;8:1585–1591.
221. Xie Y, Liu L, Huang X, Guo YW, Lou L. Scalaradial inhibition of epidermal growth factor
receptor-mediated Akt phosphorylation is independent of secretory phospholipase A2.
J Pharmacol Exp Ther 2005;314:1210–1217.
222. Xie Y, Yu Z, Liu L, Guo Y, Lou L. Biphasic regulation of extracellular signal-regulated kinases
by scalaradial, a secretory phospholipase A2 inhibitor. Cancer Biol Ther 2006;5:988–992.
223. Hussein G, Sankawa U, Gato H, Matsumoto K, Watanabe H. Astaxanthin, a carotenoid with
potential in human health and nutrition. J Nat Prod 2006;69:443–449.
224. Terracciano S, Aquino M, Rodriquez M, Monti MC, Casapullo A, Riccio R, Gomez-Paloma L.
Chemistry and biology of anti-inflammatory marine natural products: Molecules interfering with
cyclooxygenase, NF-kB and other unidentified targets. Cur Med Chem 2006;13:1947–1969.
225. Mayer AMS, Rodriguez AD, Berlinck RGS, Hamann MT. Marine pharmacology in 2003-4:
Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory,
antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the
cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action.
Comp Biochem Physiol C 2007;145:553–581.
226. Lucas R, Giannini C, D’auria MV, Paya M. Modulatory effect of bolinaquinone, a marine
sesquiterpenoid, on acute and chronic inflammatory processes. J Pharmacol Exp Ther
2003;304:1172–1180.
227. Mayer AMS, Hamann MT. Marine pharmacology in 2001–2002: Marine compounds with
anthelmintic, antibacterial, anticoagulant, antidiabetic, antifungal, anti-inflammatory, antimalar-
ial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovas-
cular, immune and nervous systems, and other miscellaneous mechanisms of action. Comp
Biochem Physiol C 2005;140:265–286.
228. Potterat O, Hamburger M. Drug discovery and development with plant-derived compounds. In:
Petersen F, Amstutz R, editors. Progress in drug research. Vol. 65. Switzerland: Birkhauser
Verlag; 2008. pp 46–118.
229. Boonphong S, Puangsombat P, Baramee A, Mahidol C, Ruchirawat S, Kittakoop P. Bioactive
compounds from Bauhinia purpurea possessing antimalarial, antimycobacterial, antifungal, anti-
inflammatory, and cytotoxic activities. J Nat Prod 2007;70:795–801.
230. Chen J-J, Lin W-J, Liao C-H, Shieh P-C. Anti-inflammatory benzenoids from Antrodia
camphorata. J Nat Prod 2007;70:989–992.
ANTI-INFLAMMATORYNATURALPRODUCTS K 53
Medicinal Research Reviews DOI 10.1002/med

231. Zhang X, Xu J-K, Wang J, Wang N-L, Kurihara H, Kitanaka S, Yao X-S. Bioactive bibenzyl
derivatives and fluorenones from Dendrobium nobile. J Nat Prod 2007;70:24–28.
232. Jeenapongsa R, Yoovathaworn K, Sriwatanakul KM, Pongprayoon U, Kampon SK. Anti-
inflammatory activity of (E)-1-(3,4-dimethoxyphenyl) butadiene from Zingiber cassumunar Roxb.
J Ethnopharmacol 2003;87:143–148.
233. Xu M-Z, Lee WS, Han J-M, Oh H-W, Park D-S, Tian G-R, Jeong T-S, Park H-Y. Antioxidant
and anti-inflammatory activities of N-acetyldopamine dimers from Periostracum cicadae. Bioorg
Med Chem 2006;14:7826–7834.
234. Rios MY, Aguilar-Guadarrama B. Nitrogen-containg phorbol esters from Croton ciliatoglandu-
lifer and their effects on cyclooxygenases-1 and -2. J Nat Prod 2006;69:887–890.
235. Han M, Wen J-K, Zheng B, Zhang D-Q. Acetylbritannilatone suppresses NO and PGE2 synthesis
in RAW264.7 macrophages through the inhibition of iNOS and COX-2 gene expression. Life Sci
2004;75:675–684.
236. Duan H, Zhang Y, Xu J, Qiao J, Suo Z, Hu G, Mu X. Effect of anemonin on NO, ET-1 and
ICAM-1 production in rat intestinal microvascular endothelial cells. J Ethnopharmacol
2006;104:362–366.
237. Son HJ, Lee HJ, Yun-Choi HS, Ryu J-H. Inhibitors of nitric oxide synthesis and TNF-aexpression from Magnolia obovata in activated macrophages. Planta Med 2000;66:469–471.
238. Lee J, Jung E, Park J, Jung K, Lee S, Hong S, Park J, Park E, Kim J, Park S, Park D. Anti-
inflammatory effects of magnolol and honokiol are mediated through inhibition of the
downstream pathway of MEKK-1 in NF-kB activation signaling. Planta Med 2005;71:338–343.
239. Tse AK-W, Wan C-K, Shen X-L, Yang M, Fong W-F. Honokiol inhibits TNF-a-stimulated NF-
kB activation and NF-kB-regulated gene expression through suppression of IKK activation.
Biochem Pharmacol 2005;70:1443–1457.
240. Costa B, Trovato AE, Comelli F, Giagnoni G, Colleoni M. The non-psychoactive cannabis
constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and
neuropathic pain. European J Pharmacol 2007;556:75–83.
241. Darmani NA, Izzo AA, Degenhardt B, Valenti M, Scaglione G, Capasso R, Sorrentini I, Marzo VD.
Involvement of the cannabimimetic compound, N-palmitoyl-ethanolamine, in inflammatory and
neuropathic conditions: Review of the available pre-clinical data, and first human studies.
Neuropharmacol 2005;48:1154–1163.
Raju Gautam was born in 1980 in Bulandshahar district, Uttar Pradesh, India. He received hisB. Pharm. from Delhi Institute of Pharmaceutical Sciences And Research (DIPSAR),University of Delhi in June 2003, and his M. S. (Pharm.) from National Institute ofPharmaceutical Education and Research (NIPER), Mohali, India in June 2005. Aftercompletion of Master degree, he joined Ph.D. program under the supervision of AssociateProfessor Dr. Sanjay M. Jachak at NIPER in July 2005. Currently he is a fourth year Ph.D.student working on isolation, characterization, semi-synthesis, and biological evaluation ofcompounds from plants for anti-inflammatory activity.
Sanjay M. Jachak is Associate Professor of Natural Products at the National Institute ofPharmaceutical Education and Research (NIPER), SAS Nagar, Punjab, India. He receivedPh.D. in Pharmacy (specialization: Phytochemistry) from the Karl Franzens University, Graz,Austria in 1997. He was lecturer at the College of Pharmacy, Nashik, India from 1998– 1999.He joined NIPER, India as Assistant Professor of Natural Products in 1999 and becameAssociate Professor in 2006. His research interests include characterization of anti-inflammatory and antimycobacterial natural products, design and synthesis of bioactive naturalproducts.
54 K GAUTAMAND JACHAK
Medicinal Research Reviews DOI 10.1002/med




















