
Re-evaluation of an animal model for ADHD using a free-operant choice task
6
Journal of Neuroscience Methods 176 (2009) 166–171 Contents lists available at ScienceDirect Journal of Neuroscience Methods journal homepage: www.elsevier.com/locate/jneumeth Re-evaluation of an animal model for ADHD using a free-operant choice task Margery C. Pardey, Judi Homewood, Alan Taylor, Jennifer L. Cornish ∗ Department of Psychology, Macquarie University, Sydney, New South Wales 2109, Australia article info Article history: Received 10 March 2008 Received in revised form 5 September 2008 Accepted 5 September 2008 Keywords: Attention Deficit Hyperactivity Disorder Animal model Spontaneously Hypertensive Rat Locomotor activity Delayed reinforcement abstract Previous research using free-operant procedures have reported that the Spontaneously Hypertensive Rat (SHR) is more impulsive and inattentive than the Wistar-Kyoto (WKY) rat. Recently these behavioural differences have been suggested to be a consequence of differences in the overall activity of these strains. This study compared SHRs to WKYs on locomotor activity and delay sensitivity using a delayed reinforce- ment (DR) and extinction (EXT) task. SHRs maintained higher locomotor activity than WKYs, however no significant group differences were found on the total lever presses in the DR or EXT tasks. During the DR task, SHRs shifted to selecting the immediate small reinforcer significantly faster than WKYs as the delay increased. WKYs predominantly selected the lever previously associated with the delayed large reinforcer throughout the EXT task, while the SHRs showed no such preference. The significant group differences found on lever selection during the DR and EXT tasks suggests that SHRs are more sensitive to delays, therefore providing further support for the face validity of the SHR as an animal model of ADHD. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Attention Deficit Hyperactivity Disorder (ADHD) is charac- terised by a pervasive pattern of inattention and/or hyperactivity and impulsivity, present before the age of seven years with the symptoms present in at least two settings (American Psychiatric Association, 2000). The symptoms are relatively stable, chronic, interfere with major life activities and are not better explained by another disorder. ADHD is subdivided into predominantly hyperactive-impulsive type (ADHD-HI), predominantly inattentive type (ADHD-I) and combined type (ADHD-C). The prevalence of ADHD is estimated to be 3–7% of school-aged children (American Psychiatric Association, 2000) and symptoms may persist into adulthood (van den Bergh et al., 2006). The disor- der has a higher prevalence in boys than girls (American Psychiatric Association, 2000) and data suggest that it has a genetic compo- nent (Biederman et al., 1990; Tannock, 1998), which may be linked to dysfunction of the central dopamine system (Cook et al., 1995; Volkow et al., 2001). This research was funded by the Post Graduate Research Fund provided by the Department of Psychology at Macquarie University (MCP) and was also supported by the Psychology Department Start-up grant (JLC). The findings of this research were presented in a poster format at the 28th Annual Australian Neuroscience Soci- ety’s meeting held in January 2008 in Hobart, Tasmania. All work was conducted in accordance with Macquarie University’s policies on animal welfare and husbandry. ∗ Corresponding author. Tel.: +61 2 9850 9467; fax: +61 2 9850 7759. E-mail address: [email protected] (J.L. Cornish). An appropriate animal model of ADHD is necessary to con- duct neurobiological investigations of this disorder, as current techniques limit similar investigations in human subjects. The Spontaneously Hypertensive Rat (SHR) was developed by inbreed- ing Wistar-Kyoto rats (WKY) selecting for hypertension, and unexpectedly resulted in increased motor activity (Davids et al., 2003). Davids et al. (2003) concluded from their literature review of animal models of ADHD that the SHR is the most thoroughly studied, and widely used animal model of ADHD. Animal models of human disorders must meet three validation criteria: face validity, construct validity, and predictive valid- ity (Sagvolden, 2000; Sagvolden et al., 2005; Alsop, 2007). The behavioural similarities between the SHR and human ADHD pro- vides support for the face validity of the SHR as a model of ADHD. Compared to its genetic control, the WKY, the SHR has been shown to be hyperactive, impulsive, and have a deficit of sustained atten- tion (Mook et al., 1993; Berger and Sagvolden, 1998; Boix et al., 1998; Sagvolden, 2000). These symptoms have also been shown to be ameliorated by administration of low doses of psychostimulants such as methylphenidate and d-amphetamine, common pharma- cotherapies for ADHD (Sagvolden et al., 1992, 2005; Boix et al., 1998; Sagvolden, 2000). While the characteristic higher prevalence of ADHD in boys over girls is not shared by the SHR, there is a differ- ence in the types of ADHD behavioural traits displayed by male and female animals (Berger and Sagvolden, 1998; Bucci et al., 2008). Altered dopamine function in the SHR conforms to the theoreti- cal rationale for ADHD, providing construct validity for this animal model (Sagvolden, 2000; Sagvolden et al., 2005). It is also important to note that prior to the availability of the SHR, the underlying pro- 0165-0270/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.jneumeth.2008.09.009
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Re-evaluation of an animal model for ADHD using a free-operant choice task

R
MD
a
ARRA
KAASLD
1
tasAibht
cmdAntV
Dbwea
0d
Journal of Neuroscience Methods 176 (2009) 166–171
Contents lists available at ScienceDirect
Journal of Neuroscience Methods
journa l homepage: www.e lsev ier .com/ locate / jneumeth
e-evaluation of an animal model for ADHD using a free-operant choice task�
argery C. Pardey, Judi Homewood, Alan Taylor, Jennifer L. Cornish ∗
epartment of Psychology, Macquarie University, Sydney, New South Wales 2109, Australia
r t i c l e i n f o
rticle history:eceived 10 March 2008eceived in revised form 5 September 2008ccepted 5 September 2008
a b s t r a c t
Previous research using free-operant procedures have reported that the Spontaneously Hypertensive Rat(SHR) is more impulsive and inattentive than the Wistar-Kyoto (WKY) rat. Recently these behaviouraldifferences have been suggested to be a consequence of differences in the overall activity of these strains.This study compared SHRs to WKYs on locomotor activity and delay sensitivity using a delayed reinforce-
eywords:ttention Deficit Hyperactivity Disordernimal modelpontaneously Hypertensive Ratocomotor activityelayed reinforcement
ment (DR) and extinction (EXT) task. SHRs maintained higher locomotor activity than WKYs, however nosignificant group differences were found on the total lever presses in the DR or EXT tasks. During the DRtask, SHRs shifted to selecting the immediate small reinforcer significantly faster than WKYs as the delayincreased. WKYs predominantly selected the lever previously associated with the delayed large reinforcerthroughout the EXT task, while the SHRs showed no such preference. The significant group differencesfound on lever selection during the DR and EXT tasks suggests that SHRs are more sensitive to delays,
r sup
dtSiu2os
cibvC
therefore providing furthe
. Introduction
Attention Deficit Hyperactivity Disorder (ADHD) is charac-erised by a pervasive pattern of inattention and/or hyperactivitynd impulsivity, present before the age of seven years with theymptoms present in at least two settings (American Psychiatricssociation, 2000). The symptoms are relatively stable, chronic,
nterfere with major life activities and are not better explainedy another disorder. ADHD is subdivided into predominantlyyperactive-impulsive type (ADHD-HI), predominantly inattentiveype (ADHD-I) and combined type (ADHD-C).
The prevalence of ADHD is estimated to be 3–7% of school-agedhildren (American Psychiatric Association, 2000) and symptomsay persist into adulthood (van den Bergh et al., 2006). The disor-
er has a higher prevalence in boys than girls (American Psychiatric
ssociation, 2000) and data suggest that it has a genetic compo-ent (Biederman et al., 1990; Tannock, 1998), which may be linkedo dysfunction of the central dopamine system (Cook et al., 1995;olkow et al., 2001).� This research was funded by the Post Graduate Research Fund provided by theepartment of Psychology at Macquarie University (MCP) and was also supportedy the Psychology Department Start-up grant (JLC). The findings of this researchere presented in a poster format at the 28th Annual Australian Neuroscience Soci-
ty’s meeting held in January 2008 in Hobart, Tasmania. All work was conducted inccordance with Macquarie University’s policies on animal welfare and husbandry.∗ Corresponding author. Tel.: +61 2 9850 9467; fax: +61 2 9850 7759.
E-mail address: [email protected] (J.L. Cornish).
tt1bsc1oefAcmt
165-0270/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.jneumeth.2008.09.009
port for the face validity of the SHR as an animal model of ADHD.© 2008 Elsevier B.V. All rights reserved.
An appropriate animal model of ADHD is necessary to con-uct neurobiological investigations of this disorder, as currentechniques limit similar investigations in human subjects. Thepontaneously Hypertensive Rat (SHR) was developed by inbreed-ng Wistar-Kyoto rats (WKY) selecting for hypertension, andnexpectedly resulted in increased motor activity (Davids et al.,003). Davids et al. (2003) concluded from their literature reviewf animal models of ADHD that the SHR is the most thoroughlytudied, and widely used animal model of ADHD.
Animal models of human disorders must meet three validationriteria: face validity, construct validity, and predictive valid-ty (Sagvolden, 2000; Sagvolden et al., 2005; Alsop, 2007). Theehavioural similarities between the SHR and human ADHD pro-ides support for the face validity of the SHR as a model of ADHD.ompared to its genetic control, the WKY, the SHR has been showno be hyperactive, impulsive, and have a deficit of sustained atten-ion (Mook et al., 1993; Berger and Sagvolden, 1998; Boix et al.,998; Sagvolden, 2000). These symptoms have also been shown toe ameliorated by administration of low doses of psychostimulantsuch as methylphenidate and d-amphetamine, common pharma-otherapies for ADHD (Sagvolden et al., 1992, 2005; Boix et al.,998; Sagvolden, 2000). While the characteristic higher prevalencef ADHD in boys over girls is not shared by the SHR, there is a differ-nce in the types of ADHD behavioural traits displayed by male and
emale animals (Berger and Sagvolden, 1998; Bucci et al., 2008).ltered dopamine function in the SHR conforms to the theoreti-al rationale for ADHD, providing construct validity for this animalodel (Sagvolden, 2000; Sagvolden et al., 2005). It is also importanto note that prior to the availability of the SHR, the underlying pro-

roscien
cspi2
bhgtfwrtAhSdbS
amfo(Wlii
abracopd
da1sa
2
2
TVhlstdeadmw0
(mossfmC
2
2
b3ow1tow(iMo
2
oiNwwh6
2
2
iA(fTswtlAcias
2
ttion. Throughout this experiment the rats had ad libitum access
M.C. Pardey et al. / Journal of Neu
esses of the altered reinforcement mechanism integral to ADHDymptomatology was unknown. Additionally, the SHR assisted inredicting the developmental course of hyperactivity, establish-
ng the SHRs predictive validity (Sagvolden, 2000; Sagvolden et al.,005).
The face validity of the SHR as a model of ADHD has recentlyeen questioned. Previous researchers have not accounted for theigher level of activity in SHRs compared to WKYs, which may exag-erate the group differences reported. Alsop (2007) re-examinedhe data of selected papers that compared SHRs and WKYs onree-operant procedures. When the data from the selected papersas expressed as a proportion of baseline or maximum response
ates, Alsop (2007) found that the group differences reported byhe researchers were attenuated or in some instances removed.lsop also reported that while the SHRs total response rate wasigher, the overall pattern of responding was similar in both theHRs and WKYs. This finding is inconsistent with studies of chil-ren where different patterns of responding have been measuredetween ADHD and non-ADHD subjects (Sagvolden et al., 1998;agvolden, 2000; Alsop, 2007).
The aim of the present study was to re-evaluate the SHR asmodel of ADHD, in light of Alsop’s findings. Comparisons wereade between SHR and WKY rats on three tasks. Firstly, to account
or the problems outlined by Alsop (2007) in the data analysisf previous research, locomotor activity and total lever pressesresponse rates) were measured in the same group of SHRs and
KYs. This was to determine whether overall activity levels areinked to the total response rates during the operant task employedn this study and allowed for statistical control during data analysis,f necessary.
Secondly, patterns of responding for SHRs and WKYs weressessed using a free-operant choice task. Children with ADHD haveeen reported to prefer small immediate reinforcers over largereinforcers delivered after a delay (Sonuga-Barke et al., 1992). SHRsnd WKYs were compared on an operant task which allowed freehoice between receiving a small reinforcer delivered immediately,r a larger reinforcer delivered after a delay. If the SHR is an appro-riate model of ADHD, the pattern of responses on the task shouldiffer between the groups.
Finally, as a measure of the deficit of sustained attention evi-ent in children with ADHD, SHRs and WKYs were compared onn extinction task (Berger and Sagvolden, 1998; Sagvolden et al.,998, 2005). If the SHR has a deficit of sustained attention, theyhould continue to respond at an increased level in the absence ofreinforcer when compared to the WKYs.
. Materials and methods
.1. Subjects
The present study used 12 male SHR and 12 male WKY rats.he rats were obtained from the Animal Resources Centre (Canningale, WA, Australia). Upon arrival in the laboratory, the rats wereoused in pairs in opaque, plastic cages (60 cm × 21.5 cm × 36 cm,
ength × height × width) containing sawdust, blocks of wood andhredded paper covered with a raised wire mesh roof (27 cmotal height). They were allowed free access to water and stan-ard laboratory rat chow, except during the operant conditioningxperiment. During operant conditioning the rats were individu-
lly housed and placed on food deprivation (given 8–17 g of foodaily to maintain their body weight at approximately 85%). The ani-al holding room was held at a constant temperature of 21 ◦C. Ratsere housed on a reverse light/dark cycle (lights on at 20:00 h until8:00 h) and experiments were conducted during the rats’ activetaofD
ce Methods 176 (2009) 166–171 167
dark) cycle. At the beginning of training, the rats were approxi-ately 56 days old, weighed 167–220 g, had been handled daily for
ne week by the experimenter and were experimentally naïve. Thetudy was conducted with the approval of the Macquarie Univer-ity Animal Ethic Committee (reference number AEC 2006/019) andollowed the Australian Code of Practice for the Care and Use of Ani-
als for Scientific Purposes (National Health and Medical Researchouncil, 2004).
.2. Locomotor activity
.2.1. ApparatusLocomotor activity was measured in eight identical purpose
uilt operant conditioning chambers. Each chamber measured0 cm × 47.5 cm × 25.5 cm (length × height × width) and consistedf aluminium sides and roof, clear polycarbonate front and backith a floor of 16 stainless steel bars (6 mm diameter) spaced
8 mm apart (centre to centre). Two passive infrared detec-ors were located 30 mm above the floor, opposite each other,n the side walls to record locomotor activity. Each chamberas located in a ventilated, sound attenuated outer housing
64 cm × 72 cm × 60 cm, length × height × width). Locomotor activ-ty was tracked via “Workbench Mac” software running on
acintosh Computers. Video cameras in each chamber allowedbservation of the rats’ behaviour.
.2.2. ProcedureThe rats’ locomotor activity was measured in these chambers
ver a period of 2 h. Locomotor activity was measured by passivenfrared detectors (PIR, Quantum passive infrared motion sensor,ess Security Products, Australia) located in the centre of each sideall approximately 30 mm above floor level. Locomotor activityas measured by detection of small movements of the subjects’ead and body. The locomotor data was recorded by computer in0 s bins.
.3. Delayed reinforcement (DR) and extinction (EXT) task
.3.1. ApparatusThe DR task and the EXT task were both conducted in six
dentical operant conditioning chambers (MED Associates, Inc., St.lbans, VT, USA). Each chamber measured 30 cm × 21 cm × 25.5 cm
length × height × width) and consisted of clear polycarbonateront, back and roof, and aluminium side walls (including test wall).he floor consisted of 19 stainless steel bars, 5 mm in diameter andpaced 16 mm apart (centre to centre). Each chamber was equippedith a house light (100 mA) on the wall opposite the test wall. The
est wall had a food magazine in the centre with a non-retractableever to either side, each requiring 25 g of weight to activate.bove both levers was a cue light (100 mA, 2.5 cm diameter). Eachhamber was located in a ventilated, sound attenuated outer hous-ng (64 cm × 60 cm × 40 cm, length × height × width). Lever pressctivity in these chambers was tracked using “MED-PC Version IV”oftware running on a PC computer.
.3.2. DR task procedureTwenty-four hours prior to the commencement of training,
he rats were individually housed and placed on food depriva-
o water, with their food restricted to maintain their body weightt approximately 85%. The procedure was based on the conceptf self-controlled choice (Evenden and Ryan, 1996). The food rein-orcer used was 45 mg Noyes Precision Pellets, Formula A (Researchiets, Inc., New Brunswick, NJ, USA).
168 M.C. Pardey et al. / Journal of Neuroscience Methods 176 (2009) 166–171
Table 1Duration of delay (s) experienced in the delayed reinforcement task before thereinforcer was delivered on each test day.
Test day Delay duration in seconds(each experienced 6 times)
1 0, 1, 2, 3, 42 0, 2, 4, 6, 83 0, 5, 10, 15, 204 0, 10, 20, 40, 605 0, 10, 20, 30, 406 0, 20, 40, 60, 8078
2pfcwtrtarlia13rr
lfrop
2ttiFtwhdidr
rlbedsrrpcwtt
FWs
2
tnlbtoeAf
2
MGrfpw
3
3
stwamaisipcd
0, 30, 60, 90, 12060
.3.2.1. Training phase. The rats were placed in the chambers onceer day. The house light was illuminated and the rat was given oneree food pellet. After 10 s, the house light was extinguished and aue light above one of the levers was turned on indicating the leveras now active. Once the rat pressed the active lever, they received
he food reinforcer associated with that lever. Pressing one leveresulted in a small (one pellet) immediate reinforcer, while pressinghe other lever resulted in a large (five pellets) reinforcer deliveredfter a delay of 2 s. During this delay, the cue light flashed. Once theeinforcer was delivered, the cue light was turned off and the houseight was illuminated signalling the end of that trial. There was annter-trial interval (ITI) of 21 s. The sequence of which lever wasctive was in a fixed random order, with each lever active a total of5 times. The daily session was complete when the rat had made0 successful responses, or 30 min had expired. The rats met crite-ia to move onto the testing phase when they made 30 successfulesponses within 30 min on three consecutive days.
The levers were counterbalanced such that for half the rats thearger, delayed reinforcer was associated with the left lever andor the remaining half it was associated with the right lever. Thisemained constant throughout the experiment. The total numberf reinforcers attained from each lever and the total number of leverresses on each lever were recorded.
.3.2.2. Test phase. On each test day, the rats received a reminderrial to begin. Following the procedure outlined above for theraining days, the rats were initially forced to choose the smallmmediate lever and then forced to choose the large delayed lever.ollowing these reminder trials, the testing trials began. In theserials both cue lights were turned on, signalling that both leversere active and allowing the rat to choose a lever. Once the ratad chosen a lever, the reinforcer associated with that lever waselivered, the cue lights were turned off and the house light was
lluminated, indicating the end of that trial. The ITI was 21 s. Theaily session was complete when the rat had made 32 successfulesponses, or 45 min had expired.
The reinforcer associated with the small immediate leveremained constant throughout the test phase. If the rats chose thearge delayed lever, they received five pellets, however the delayetween their response and reinforcer delivery increased duringach test session, with different delay durations on each of the testays. Table 1 shows the time in seconds that the rat must with-tand between selecting the large delayed lever and receiving theireinforcer on each test day. Each of these delay durations were expe-
ienced six times before proceeding to the longer delay on thatarticular test day. As with the training phase, during the delay theue light flashed until the reinforcer was delivered. The data thatas recorded included the longest delay accepted for each test day,he total number of reinforcers attained from each lever and theotal number of presses on each lever.
h
3
d
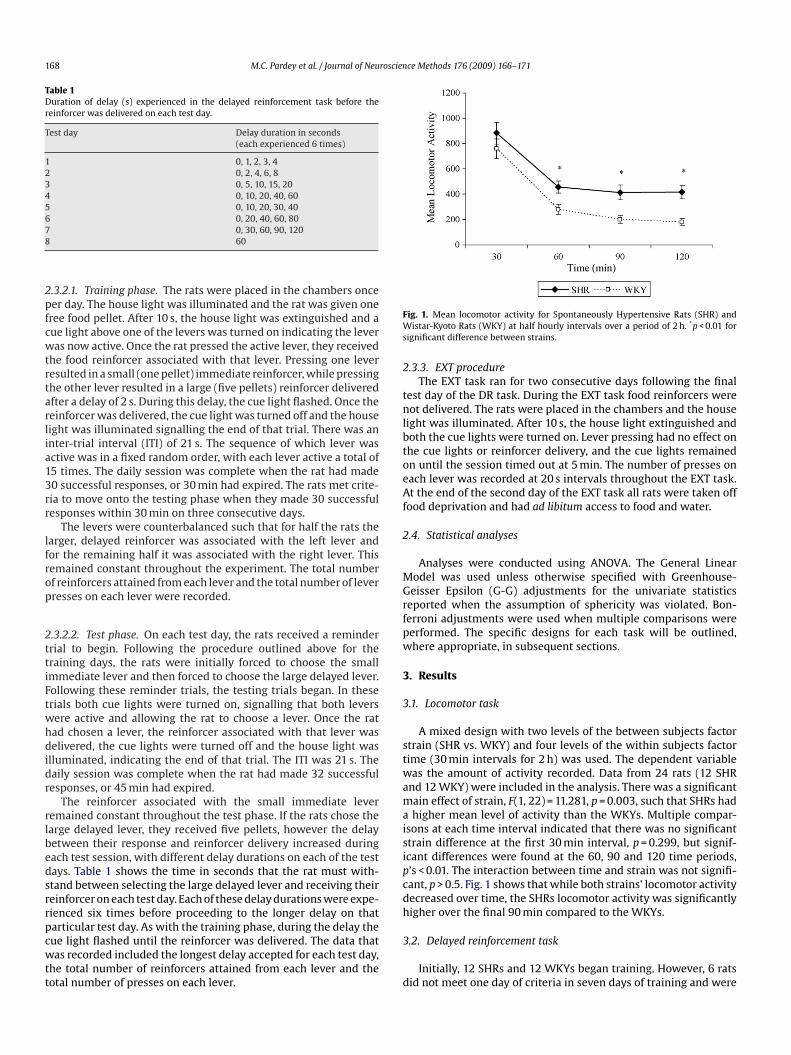
ig. 1. Mean locomotor activity for Spontaneously Hypertensive Rats (SHR) andistar-Kyoto Rats (WKY) at half hourly intervals over a period of 2 h. *p < 0.01 for
ignificant difference between strains.
.3.3. EXT procedureThe EXT task ran for two consecutive days following the final
est day of the DR task. During the EXT task food reinforcers wereot delivered. The rats were placed in the chambers and the house
ight was illuminated. After 10 s, the house light extinguished andoth the cue lights were turned on. Lever pressing had no effect onhe cue lights or reinforcer delivery, and the cue lights remainedn until the session timed out at 5 min. The number of presses onach lever was recorded at 20 s intervals throughout the EXT task.t the end of the second day of the EXT task all rats were taken off
ood deprivation and had ad libitum access to food and water.
.4. Statistical analyses
Analyses were conducted using ANOVA. The General Linearodel was used unless otherwise specified with Greenhouse-eisser Epsilon (G-G) adjustments for the univariate statistics
eported when the assumption of sphericity was violated. Bon-erroni adjustments were used when multiple comparisons wereerformed. The specific designs for each task will be outlined,here appropriate, in subsequent sections.
. Results
.1. Locomotor task
A mixed design with two levels of the between subjects factortrain (SHR vs. WKY) and four levels of the within subjects factorime (30 min intervals for 2 h) was used. The dependent variableas the amount of activity recorded. Data from 24 rats (12 SHR
nd 12 WKY) were included in the analysis. There was a significantain effect of strain, F(1, 22) = 11.281, p = 0.003, such that SHRs hadhigher mean level of activity than the WKYs. Multiple compar-
sons at each time interval indicated that there was no significanttrain difference at the first 30 min interval, p = 0.299, but signif-cant differences were found at the 60, 90 and 120 time periods,’s < 0.01. The interaction between time and strain was not signifi-ant, p > 0.5. Fig. 1 shows that while both strains’ locomotor activityecreased over time, the SHRs locomotor activity was significantlyigher over the final 90 min compared to the WKYs.
.2. Delayed reinforcement task
Initially, 12 SHRs and 12 WKYs began training. However, 6 ratsid not meet one day of criteria in seven days of training and were
M.C. Pardey et al. / Journal of Neuroscience Methods 176 (2009) 166–171 169
Table 2Total number of presses in the DR task for the immediate and delayed levers combined, mean (s.d.), for SHRs and WKYs on each test day.
Strain Day 1 Day 2 Day 3 Day 4 Day 5 Day 6 Day 7 Day 8
SHR 43.22 (6.28) 44.11 (5.97) 61.67 (9.18) 75.44 (28.14) 63.89 (34.07) 100.33 (17.64) 115.56 (40.22) 171.67 (34.99)W )
p
D oto Ra
daW
wvedvldtanpTa
dwpt6b
1Fdl
1iSW
Ffaf*
3
fdwvlti9tpdnsFtms
4
llDti
KY 50.67 (9.85) 55.22 (16.9) 69.11 (18.86) 80.33 (28.43
value 0.074 0.081 0.303 0.719
R, Delayed reinforcement; SHR, Spontaneously Hypertensive Rat; WKY, Wistar-Ky
iscontinued from the experiment, leaving N = 9 in each group. Onverage, to meet their first day of criteria, SHRs took 3 days and the
KYs took 3.33 days. This difference was not significant, p = 0.675.Separate analyses were run for each day of testing. The design
as mixed with two levels of the between subject factor strain (SHRs. WKY), five levels of the within subject factor delay (varied forach test day, see Table 1), with the total number of lever pressesuring that session initially included as a covariate. The dependentariable was the number of reinforcers attained from the delayedever. Analysis of the number of reinforcers attained from the imme-iate lever was not included as it would reflect identical significanceo the delayed lever, given that the maximum number of reinforcersrat could attain during a testing session was 30. There was no sig-ificant difference between the groups for total number of leverresses on any day, p > 0.07, so it was excluded from the analysis.able 2 shows the average total number of lever presses for SHRsnd WKYs on each test day.
Fig. 2 shows the mean number of reinforcers attained from theelayed lever on each test day for both strains. As expected, thereas a significant main effect of delay on every day of testing, all
’s ≤ 0.001, such that both strains pressed the delayed lever less ashe delay increased. With the exception of test day 1 and test day, which are discussed below, all main effects of strain, and strainy delay interactions were not significant, all p’s > 0.1.
On test day 1, there was a significant main effect of strain, F(1,6) = 6.084, p = 0.025, and a significant strain by delay interaction,(1, 16) = 6.084, p = 0.025. This occurred as the SHRs choose theelayed lever, on average, more often than the WKYs during the
ongest delay for this day (4 s, see Table 1).
On test day 6, there was a significant main effect of strain, F(1,6) = 4.717, p = 0.045, with the strain by delay interaction approach-ng significance, F(1.789, 28.625) = 2.901, p = 0.076. On average, theHRs selected the delayed lever significantly less compared to theKYs.
ig. 2. Mean number of reinforcers attained from the delayed lever on each test dayor Spontaneously Hypertensive Rats (SHR) and Wistar-Kyoto Rats (WKY), averagedcross delay periods in the delayed reinforcement task. The delay time increasedrom test day 1 to test day 8 (refer to Table 1 for delay durations on each test day).p < 0.05 for significant difference between strains.
fHlifT
FHt
83.11 (35.01) 123.33 (31.02) 142.67 (24.92) 181.89 (51.54)
0.255 0.071 0.105 0.629
t.
.3. Extinction task
The design was mixed with two levels of the between subjectactor strain (SHR vs. WKY), two levels of the within subject factorsay (1 and 2) and lever (immediate and delay), and 15 levels of theithin subject factor time (20 s intervals for 5 min). The dependent
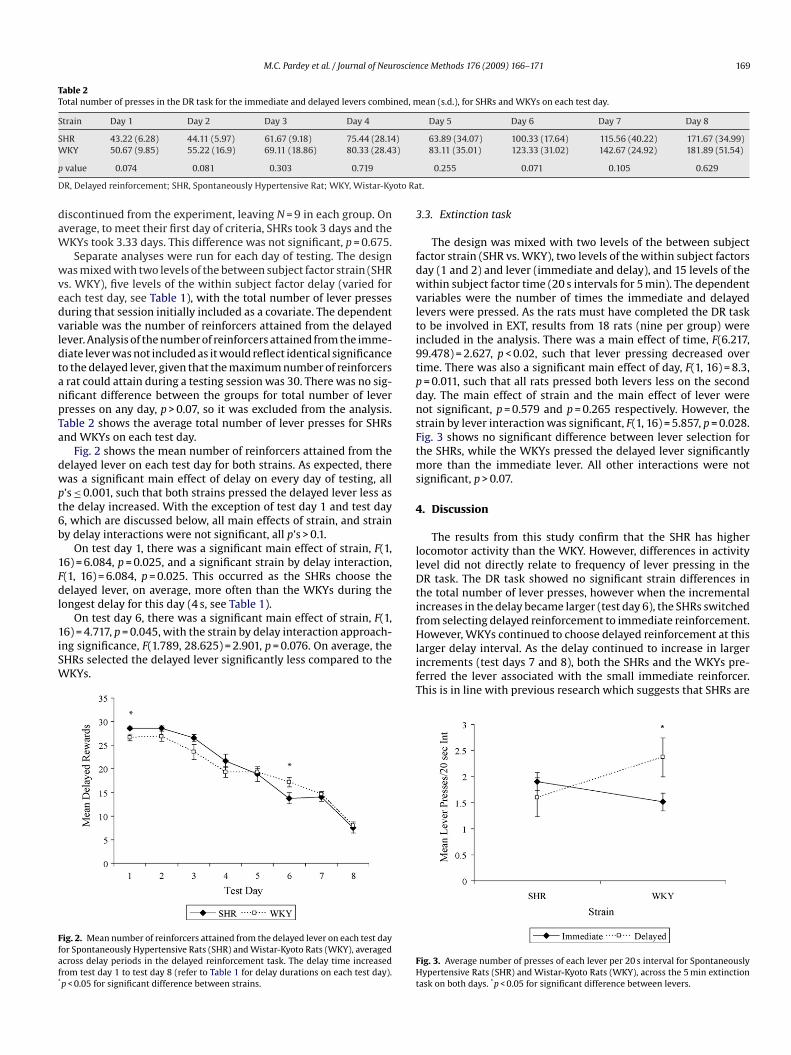
ariables were the number of times the immediate and delayedevers were pressed. As the rats must have completed the DR tasko be involved in EXT, results from 18 rats (nine per group) werencluded in the analysis. There was a main effect of time, F(6.217,9.478) = 2.627, p < 0.02, such that lever pressing decreased overime. There was also a significant main effect of day, F(1, 16) = 8.3,= 0.011, such that all rats pressed both levers less on the seconday. The main effect of strain and the main effect of lever wereot significant, p = 0.579 and p = 0.265 respectively. However, thetrain by lever interaction was significant, F(1, 16) = 5.857, p = 0.028.ig. 3 shows no significant difference between lever selection forhe SHRs, while the WKYs pressed the delayed lever significantly
ore than the immediate lever. All other interactions were notignificant, p > 0.07.
. Discussion
The results from this study confirm that the SHR has higherocomotor activity than the WKY. However, differences in activityevel did not directly relate to frequency of lever pressing in theR task. The DR task showed no significant strain differences in
he total number of lever presses, however when the incrementalncreases in the delay became larger (test day 6), the SHRs switchedrom selecting delayed reinforcement to immediate reinforcement.
owever, WKYs continued to choose delayed reinforcement at thisarger delay interval. As the delay continued to increase in largerncrements (test days 7 and 8), both the SHRs and the WKYs pre-erred the lever associated with the small immediate reinforcer.his is in line with previous research which suggests that SHRs are
ig. 3. Average number of presses of each lever per 20 s interval for Spontaneouslyypertensive Rats (SHR) and Wistar-Kyoto Rats (WKY), across the 5 min extinction
ask on both days. *p < 0.05 for significant difference between levers.

1 roscien
m1a
hSafdalsieli
iwtttgfiWe
fbStiaabStAptwTtoWStt1
saest2ifAbeo(sd
tflbteldratS
ia1ofrtsmwd1
baaepmtrsmtwAfftdsaSdj2mtdtTis
Sre
70 M.C. Pardey et al. / Journal of Neu
ore sensitive to increases in delays than WKYs (Sagvolden et al.,992; Johansen and Sagvolden, 2005; Johansen et al., 2005; Fox etl., 2008).
Using the present EXT model, the results do not support theypothesis that SHRs have a deficit of sustained attention as theHRs did not continue to respond in the absence of a reinforcert a significantly higher rate than the WKYs. However, the resultsrom the EXT task corroborate that SHRs are more sensitive to aelay than WKYs. While both groups pressed the levers with equiv-lent frequency, the WKYs were more likely to choose the delayedever over the immediate lever, while the SHRs did not demon-trate this preference. This finding suggests that the SHRs are morempulsive than the WKYs as they are less willing to wait for anxpected reinforcer. Significant strain differences were observed inever selection on both the DR and EXT task, despite no differencen lever pressing frequency.
Overall activity level was not directly linked to total lever press-ng during the operant task employed in the present study. In fact,
hile no significant difference was found between the strains inhe number of lever presses during the DR and EXT tasks, WKYsended to lever press more than SHRs during both DR and EXTasks. Additionally, there was no significant difference between theroups during the first 30 min of the locomotor task. Together, thesendings counter the argument that differences between SHRs andKYs occur due to the typical inactivity of the WKY (van den Bergh
t al., 2006; Alsop, 2007).The findings of the present study provide further support for the
ace validity of the SHR as a model of ADHD. Similarly to what haseen reported in children with ADHD (Sagvolden et al., 2005) theHRs did not show hyperactivity compared to controls when ini-ially placed in a novel environment, however as the time spentn this environment increased, they became increasingly hyper-ctive. The fact that not all behavioural characteristics of ADHDre present in novel situations (Sagvolden et al., 2005) may alsoe the explanation of the pattern of data in the DR task. For theHRs, the change in procedure to test day 1 may have been enougho make the task novel, therefore limiting the SHRs expression ofDHD behaviours. However, by test day 6, the novelty of the newrocedure would have been lost, altering the SHRs behaviour. Inhe future, re-introduction of the test day 1 procedure at this pointould be an appropriate way to demonstrate a true novelty effect.
he SHRs underlying pattern of responding differed from that ofhe WKY, similar to differences observed in children with and with-ut ADHD (Sagvolden et al., 1998; Sagvolden, 2000; Alsop, 2007).hen the DR task was novel and the delay period was short, the
HRs were more willing to maximise their reinforcer. However, ashe delay lengthened, the SHRs preferred to minimise the delayhey experienced, as do children with ADHD (Sonuga-Barke et al.,992).
The SHRs pattern of responses on the DR task conform to theteepened delay-of-reinforcement gradient postulated to drive theltered reinforcement pattern in children with ADHD (Sagvoldent al., 1998). The delay-of-reinforcement gradient is the relation-hip between the effect of the reinforcer and the interval betweenhe response and reinforcer (Johansen et al., 2002; Sagvolden et al.,005). The effect of the reinforcer is largest when the reinforcer
s delivered immediately and decreases as the delivery of the rein-orcer is delayed (Sagvolden et al., 2005). Therefore, if children withDHD have an altered reinforcement mechanism, as is illustratedy their steeper delay-of-reinforcement gradient, reinforcers deliv-
red immediately will be more effective, while the introductionf a delay will rapidly reduce the effectiveness of the reinforcerSagvolden et al., 1998). The results from the DR task in the presenttudy follow this pattern. When the delay was relatively short, testay 1, the SHRs chose the large delayed lever significantly more thant2Agn
ce Methods 176 (2009) 166–171
he WKYs, demonstrating a greater effectiveness of the larger rein-orcer. However, as the delay was increased, the effectiveness of thearger reinforcer decreased at a faster rate for the SHRs, illustratedy their significantly reduced selection of the large delayed lever onest day 6. As the delay increased further on test days 7 and 8, theffectiveness of the larger reinforcer had diminished to an equiva-ent point for both SHRs and WKYs, explaining the non-significantifferences reported for test days 7 and 8 (see Fig. 2). These resultseplicate the findings of previous research which have reported
steepened delay-of-reinforcement gradient in SHRs comparedo controls using related behavioural procedures (Johansen andagvolden, 2005; Johansen et al., 2005, 2007).
In contrast, results from the present study did not support find-ngs of previous research which have reported a deficit of sustainedttention in the SHR (Sagvolden et al., 1993; Berger and Sagvolden,998; Boix et al., 1998; Sagvolden, 2000). This null result may haveccurred for the following reasons: (i) the present study used aree-operant choice task, while previous research assessed rates ofesponding during fixed- and variable interval operant tasks; (ii)he duration of the EXT task (5 min) may have been too short tohow an effect using this different procedure; or (iii) our EXT taskay not have had sufficient sensitivity. In addition, that male ratsere used may also explain why the results of the present studyo not replicate those of previous research (Berger and Sagvolden,998).
Berger and Sagvolden (1998) assessed the effects of sex onehaviour of SHRs and WKYs. They reported that both malend female SHRs responded significantly more than both malend female WKYs during an operant task using a fixed interval-xtinction (FI-EXT) schedule of reinforcement. However, theattern of responses from the SHRs differed significantly betweenales and females. The authors’ interpretation of the results was
hat the responses of male SHRs may have occurred from an alteredeinforcement mechanism, reflecting the hyperactive and impul-ive type of ADHD (ADHD-HI), while the female SHRs responsesay have occurred from an attention deficit reflecting the inatten-
ive type (ADHD-I). Berger and Sagvolden (1998) concluded thathile male and female SHRs both display behavioural traits ofDHD, the underlying mechanism is quite different in males and
emales. While using a different procedure, the current findingsrom the EXT task support Berger and Sagvolden’s (1998) interpre-ation of their results. In the present study, while not showing aeficit in sustained attention, male SHRs were found to be sen-itive to a delay while male WKYs were not, demonstrating anltered reinforcement mechanisms as suggested by Berger andagvolden (1998). Therefore, the previous research reporting aeficit of sustained attention in SHRs, which all used males sub-
ects only (Sagvolden et al., 1993; Boix et al., 1998; Sagvolden,000), may indeed be misinterpreting an altered reinforcementechanism, as all of these studies used very similar procedures
o Berger and Sagvolden (1998). It is possible that the novel proce-ure used for the EXT task in the present study could tease aparthe different underlying mechanisms between males and females.his hypothesis may be tested in the future by using a designdentical to this study and incorporating both male and femaleubjects.
While many animal models of ADHD have been proposed, theHR is the most widely used and extensively studied. It has beenecognised that ADHD has a strong genetic component (Biedermant al., 1990; Tannock, 1998) with evidence of a dysfunction of
he central dopamine system (Cook et al., 1995; Volkow et al.,001). The benefit of the SHR over other animal models is that theDHD-like behaviours of the SHR strain are similarly set in theirenome rather than resulting from brain damage, previous surgery,eurotoxicity or social interactions (Boix et al., 1998). Addition-
roscien
aunrtcba
rgowligb
tTwiierA
A
ISB4
R
A
A
B
B
B
B
C
D
D
E
F
J
J
J
J
M
N
S
S
S
S
S
S
T
M.C. Pardey et al. / Journal of Neu
lly, the attenuation of these behaviours by the psychostimulantssed to treat humans implies similar neurobiological underpin-ings (Sagvolden et al., 1992). Experimental findings, including theesults of the present study, indicate that the SHRs behaviouralraits of ADHD may be caused by altered reinforcement pro-esses (Sagvolden et al., 1998) similar to the potential cause ofehavioural disturbances in children with ADHD (Sonuga-Barke etl., 1992).
Future research should consider including alternate strains ofats as an additional control group to the WKY. WKYs are distin-uishable by behavioural and neurochemical characteristics fromther ‘normal’ strains, such as Wistar rats (Drolet et al., 2002). Itould be prudent to include additional ‘normal’ strains to estab-
ish that reported group differences are not the result of deficitsn the WKY. Consideration must be given, however, to the largeenetic variation that would be introduced by including such out-red strains.
In conclusion, the results of the present study provide addi-ional support for the face validity of the SHR as a model of ADHD.he SHRs were found to have increased locomotor activity andere more sensitive to delays, making more impulsive choices
n comparison to the WKYs, independent of lever pressing activ-ty. Furthermore, while further research is required, the proceduremployed in the EXT task of this study together with previousesearch, could potentially distinguish the different subtypes ofDHD.
cknowledgements
We would like to thank Wayne McTegg for animal care, Professorain McGregor and the School of Psychology at the University ofydney for the use of their locomotor activity chambers and Richardamber for his contribution to programming the MED-PC Versionsoftware.
eferences
lsop B. Problems with spontaneously hypertensive rats (SHR) as a modelof Attention-Deficit/Hyperactivity Disorder (AD/HD). J Neurosci Meth2007;162:42–8.
merican Psychiatric Association. Diagnostic and statistical manual of mentaldisorders. 4th ed., text revision ed. Washington, DC: American Psychiatric Asso-ciation; 2000.
erger D, Sagvolden T. Sex differences in operant discrimination behaviour inan animal model of Attention-Deficit Hyperactivity Disorder. Behav Brain Res1998;94:73–82.
iederman J, Faranone SV, Keenan K, Knee D, Tsuang MT. Family-genetic and psy-chosocial risk factors in DSM-III Attention-Deficit Disorder. J Am Acad ChildAdolesc Psychiatry 1990;29:526–33.
v
V
ce Methods 176 (2009) 166–171 171
oix F, Qiao S, Kolpus T, Sagvolden T. Chronic l-deprenyl treatment alters brainmonoamine levels and reduces impulsiveness in an animal model of Attention-Deficit/Hyperactivity Disorder. Behav Brain Res 1998;94:153–62.
ucci DJ, Hopkins ME, Keene CS, Sharma M, Orr LE. Sex differences in learningand inhibition in spontaneously hypertensive rats. Behav Brain Res 2008;187:27–32.
ook EHJ, Stein MA, Krasowski MD, Cox NJ, Olkon DM, Kieffer JE, et al. Associations ofAttention-Deficit Disorder and the dopamine transporter gene. Am J Hum Genet1995;56:993–8.
avids E, Zhang K, Tarazi FI, Baldessarini RJ. Animal models of Attention-DeficitHyperactivity Disorder. Brain Res Rev 2003;42:1–21.
rolet G, Proulx K, Pearson D, Rochford J, Deschepper CF. Comparisons of behavioraland neurochemical characteristics between WKY, WKHA, and Wistar rat strains.Neuropsychopharmacology 2002;27:400–9.
venden JL, Ryan CN. The pharmacology of impulsive behaviour in rats: the effectsof drugs on response choice with varying delays of reinforcement. Psychophar-macology 1996;128:161–70.
ox AT, Hand DJ, Reilly MP. Impulsive choice in a rodent model of Attention-Deficit/Hyperactivity Disorder. Behav Brain Res 2008;187:146–52.
ohansen EB, Sagvolden T. Behavioral effects of intra-cranial self-stimulation in ananimal model of Attention-Deficit/Hyperactivity Disorder (ADHD). Behav BrainRes 2005;162:32–46.
ohansen EB, Aase H, Meyer A, Sagvolden T, Attention-Deficit/Hyperactivity Disor-der (ADHD) behavior explained by dysfunctioning reinforcement and extinctionprocesses. Behav Brain Res 2002;130:37–45.
ohansen EB, Sagvolden T, Kvande G. Effects of delayed reinforcers on the behaviorof an animal model of Attention-Deficit/Hyperactivity Disorder (ADHD). BehavBrain Res 2005;162:47–61.
ohansen EB, Killeen PR, Sagvolden T. Behavioral variability, elimination of responses,and delay-of-reinforcement gradients in SHR and WKY rats. Behav Brain Funct2007;3:60–90.
ook DM, Jeffrey J, Neuringer A. Spontaneously hypertensive rats (SHR) read-ily learn to vary but not repeat instrumental responses. Behav Neural Biol1993;59:126–35.
ational Health and Medical Research Council. Australian code of practice for thecare and use of animals for scientific purposes, 7th ed. 2004.
agvolden T. Behavioral validation of the spontaneously hypertensive rat (SHR) asan animal model of Attention-Deficit/Hyperactivity Disorder (AD/HD). NeurosciBiobehav Rev 2000;24:31–9.
agvolden T, Metzger MA, Schiorbeck HK, Rugland A-L, Spinnangr I, Sagvolden G. Thespontaneously hypertensive rat (SHR) as an animal model of childhood hyperac-tivity (ADHD): changed reactivity to reinforcers and to psychomotor stimulants.Behav Neural Biol 1992;58:103–12.
agvolden T, Petterson MB, Larsen MC. Spontaneously hypertensive rats (SHR) as aputative animal model of childhood hyperkinesis: SHR behavior compared tofour other rat strains. Physiol Behav 1993;54:1047–55.
agvolden T, Aase H, Zeiner P, Berger D. Altered reinforcement mechanisms inAttention-Deficit/Hyperactivity Disorder. Behav Brain Res 1998;94:61–71.
agvolden T, Russell VA, Aase H, Johansen EB, Farshbaf M. Rodent models ofAttention-Deficit/Hyperactivity Disorder. Biol Psychiatry 2005;57:1239–47.
onuga-Barke EJS, Taylor E, Sembi S, Smith J. Hyperactivity and delay aversion. I. Theeffect of delay on choice. J Child Psychol Psychiatry 1992;33:387–98.
annock R. Attention Deficit Hyperactivity Disorder: advances in cognitive, neuro-biological and genetic research. J Child Psychol Psychiatry 1998;39:65–99.
an den Bergh FS, Bloemarts E, Chan JSW, Groenink L, Olivier B, Oosting RS.Spontaneously hypertensive rats do not predict symptoms of Attention-DeficitHyperactivity Disorder. Pharmacol Biochem Behav 2006;83:380–90.
olkow ND, Wang G, Fowler JS, Logan J, Gerasimov M, Maynard L, et al. Therapeuticdoses of oral methylphenidate significantly increase extracellular dopamine inthe human brain. J Neurosci 2001;21:RC121.




















