
An anabaptist paradigm for conflict transformation - University ...
Upload
independentCategory
view
0download
0
Acetaldehyde Oral Self-Administration: Evidence from theOperant-Conflict Paradigm
Silvana Cacace, Fulvio Plescia, Ignazio Barberi, and Carla Cannizzaro
Background: Acetaldehyde (ACD), ethanol’s first metabolite, has been reported to interact withthe dopaminergic reward system, and with the neural circuits involved in stress response. Rats self-administer ACD directly into cerebral ventricles, and multiple intracerebroventricular infusions ofACD produce conditioned place preference. Self-administration has been largely employed to assessthe reinforcing and addictive properties of most drugs of abuse. In particular, operant conditioning isa valid model to investigate drug-seeking and drug-taking behavior in rats.
Methods: This study was aimed at the evaluation of (i) the motivational properties of oral ACD inthe induction and maintenance of an operant-drinking behavior; (ii) ACD effect in a conflict situationemploying the punishment-based Geller–Seifter procedure; and (iii) the onset of a relapse drinkingbehavior, following ACD deprivation. The lever-pressing procedure in a sound-attenuated operant-conditioning chamber was scheduled into 3 different periods: (i) training—rewarded responses with afixed ratio 1; (ii) conflict—rewarded responses periodically associated with a 0.2 mA foot-shock; and(iii) relapse—rewarded lever presses following 1-week ACD abstinence.
Results: Our results show that oral self-administrated ACD induced: a higher rate of punishedresponses in Geller–Seifter procedures; and the establishment of a relapse behavior following ACDdeprivation.
Conclusions: In conclusion, our results indicate that ACD is able to induce an operant-drinkingbehavior, which is also maintained besides the conflict procedure and enhanced by the deprivationeffect, supporting the hypothesis that ACD itself possesses motivational properties, such as alcohol andother substances of abuse.
Key Words: Acetaldehyde, Lever-Pressing, Punishment, Reinforcement, Relapse.
SEVERAL STUDIES IN the last 30 years have investi-gated the role and the effects of acetaldehyde (ACD) as
one of the mediators of the central and peripheral effects ofethanol (EtOH). ACD derives from the peripheral metabo-lism of EtOH by alcohol dehydrogenase and is convertedinto acetic acid by aldehyde dehydrogenase (ALDH). Theblock of ALDH by disulfiram leads to an accumulation ofACD following alcohol intake and this in turn causes a seri-ous adverse reaction which has represented the rationale fortreating alcoholics with disulfiram for decades (Eckard et al.,1998; Sippel, 1974; Westcott et al., 1980; Zimatkin et al.,1998). A slow but measurable accumulation of ACD in therat brain has also been demonstrated. Although otherenzymes from the microsomal EtOH-oxidizing system, suchas CYP2E1, and alcohol dehydrogenase I are also present in
the brain (Hansson et al., 1990; Martinez et al., 2001; Mon-toliu et al., 1995; Roberts et al., 1994; Tindberg and Ingel-man-Sundberg, 1996), even if a lesser extent than in theperiphery, the biochemical data suggest that ACD is formedin the brain mainly by catalase, when EtOH is administeredacutely (Aragon and Amit, 1992; Deng and Deitrich, 2008;Hamby-Mason, 1997; Zimatkin et al., 2006). Moreover,ACD itself, and as a consequence of themetabolism of EtOH,has been reported to possess reinforcing effects: Indeed, itactivates dopaminergic neuron firing in the ventral tegmentalarea (VTA; Foddai et al., 2004) and stimulates dopaminetransmission in the nucleus accumbens (Enrico et al., 2009;Melis et al., 2007). Rats self-administer 1 to 5% (v/v) ACDdirectly into cerebral ventricles (Brown et al., 1979; Smithet al., 1984), and intragastric administration of ACDproduceconditioned place preference (Peana et al., 2009).
Self-administration has been largely employed to assessthe reinforcing and addictive properties of most drugs ofabuse, such as cocaine (Koob et al., 2004), morphine, heroin(Cooper et al., 2010), ketamine (De Luca and Badiani,2011), nicotine, and EtOH (Burattini et al., 2006; Manzardoet al., 2002), providing essential information of the brain andbrain circuits involved in drug abuse and addiction. Indeed,drugs that are addictive at the human level are also voluntar-ily self-administered by laboratory animals (Gardner, 2002).
It is still under debate whether, under physiological condi-tions, ACD formed in the liver is able to penetrate into the
From the Department of Science for Health Promotion “G.D’Alessandro” (SC, FP, IB, CC), Section of Pharmacology, Universityof Palermo, Palermo, Italy.
Received for publication August 2, 2011; accepted November 21, 2011.Reprint requests: Carla Cannizzaro, MD, Department of Science for
Health Promotion “G. D’Alessandro”, Section of Pharmacology, Univer-sity of Palermo, Via del Vespro 129, 90127 Palermo, Italy; Tel.: +39-091-6553215; Fax: +39-091-591063; E-mails: [email protected], [email protected]
Copyright© 2012 by the Research Society on Alcoholism.
DOI: 10.1111/j.1530-0277.2011.01725.x
1278 Alcohol Clin Exp Res, Vol 36, No 7, 2012: pp 1278–1287
ALCOHOLISM: CLINICAL AND EXPERIMENTAL RESEARCH Vol. 36, No. 7July 2012

brain from the periphery in spite of the metabolic barrier pre-sented by ALDH in the blood–brain barrier (Hunt, 1996).Studies showing central effects of peripherally administeredACD seem to validate this possibility (Peana et al., 2009,2010).
Self-administration within operant conditioning is consid-ered a valid experimental model to investigate drug-seekingand drug-taking behavior in rats: Indeed, the frequency ofthe animal response is directly correlated with the rewardingand addictive properties of the test substance (Higgins et al.,1994). Although the dopamine neurocircuitry was originallyproposed to mediate “reward” and was referred to as the“dopamine reward pathway” (e.g., Ettenberg, 1989; Koob,1992; Wise and Bozarth, 1985), recent studies indicate thatdopamine is likely to play an important role in the processingof behaviorally relevant positive and negative stimuli and theselection of appropriate responding (Koch et al., 2000; Sala-mone, 1994; Salamone et al., 1994, 1997; Schultz, 1998;Spanagel and Weiss, 1999; Young et al., 1998). Previouslyneutral stimuli that have been conditioned to predict “moti-vationally salient events” have been shown to increase dopa-mine release in the nucleus accumbens (Young et al., 1998).
Despite the involvement of subjective factors that are diffi-cult to reproduce in the laboratory, animal studies help shed-ding light on incentive motivational factors in addiction andon the mechanisms underlying relapse to drug-taking behav-ior. Indeed, a topic and critical element of addiction is thatmore than80%of addicts relapse todrug seeking anduse, evenafter long periods of withdrawal and abstinence. The allevia-tion of the negative symptoms that occur during withdrawal,besides the drug motivational properties, triggers relapse todrug use (reward and relief craving;Heinz et al., 2003).Under-standing the neurobiological basis of drug seeking and relapse,following the deprivation effect, is a central issue of theresearch on addiction (Aston-Jones and Harris, 2004), and asfar as we know, no studies have been published on ACD-induced relapse using anoperant-drinkingbehavior paradigm.
Given these premises, the aims of this study are as follows:(i) to investigate the ability of ACD to induce and maintainoral self-administration in an operant-conditioning para-digm; (ii) to explore ACD effect in a conflict situation,employing a modification of the punishment-based Geller–Seifter procedure; and (iii) to evaluate the onset of relapse inACD operant-drinking behavior, following repeated depri-vation periods.
These investigations can contribute to the definition ofACD as a substance of abuse, to of its overlapping with alco-hol activity in the brain, and therefore the role of ACD in theinduction and maintenance of alcohol addiction.
MATERIALS ANDMETHODS
Adult male Wistar rats (Harlan, Udine, Italy) weighing 250 to300 g, were housed 2 per cage and maintained on a reverse 12-hourlight/dark cycle under controlled environmental conditions (temper-ature 22 ± 2°C; humidity 60 to 65%) with food and water freely
available. All subjects were experimentally naive and were randomlyassigned to 1 of the 2 following groups (n = 16): ACD-drinkingrats (ACD) which self-administered a solution of ACD and water-drinking rats (CTR) which self-administered water.
On the test days, the animals were brought into the laboratoryand allowed to acclimatize for at least 60 minutes prior to theexperimental session. The experiments were performed in a sound-isolated chamber between 9:00 and 14:00. Animal performance wasrecorded on a counter connected to the chamber. The devices werethoroughly cleaned before the introduction of each animal to ensurethat the particular rat’s behavior was not affected by the detectionof another rat’s scent. All experiments were in accordance with theregulations of the Committee for Use of Experimental Animals ofthe University of Palermo.
Open Field Test
Locomotor activity was measured in an open field (OF) with acontrast-sensitive, video tracking system, the Opto-Varimex(Columbus Instruments, Columbus, OH). This apparatus is asquare box, 44 cm wide, 44 cm long, and 20 cm high, whose per-pendicular sides have 15 infrared emitters. Each beam is separatedfrom the next one by a distance of 2.54 cm. The apparatus producesa qualitative mapping of the motor patterns, measuring differentparameters simultaneously: total distance traveled (TDT), numberof transitions (NTC) from peripheral to central squares of the arena,and amount of time spent in the central areas (ATC). Each experi-mental session lasted 5 minutes; animal motor patterns wererecorded 1 minute after the rats were placed in the box and dis-played on a PC.
Apparatus
The experimental sessions were conducted in a custom-builtsound-attenuated operant-conditioning chamber (30 9 28 937 cm), placed in a sound-attenuating cubicle so as to isolate ratsfrom outside noises. The chamber was also fitted with an exhaustfan, 1 operant response lever, and diffuse house-light. The cham-ber was equipped with a liquid solenoid-actuated delivery system,which assured the delivery of 0.05 ml of solution for each leverpress; moreover, the chamber was fitted to deliver a constant-current, intermittent, inescapable, foot-shock (0.2 mA) through ascrambler to the grid floor. Besides, a stimulus light above thelever lights up during the punished period, allowing the animals toassociate the aversive stimulus to the light. A cork built for thereservoir enabled to maintain ACD concentration, preserving itfrom evaporation.
Preparation and Composition of ACD Solution
ACD (Sigma-Aldrich SRL, Milan Italy) was stocked at �20°Cand the solution was prepared with distilled water, maintained atthe same temperature. ACD concentration (0.90%; 0.450 ml in50 ml of distilled water) was controlled and measured with UVspectrum analysis, either at the beginning or at the end of the oper-ant session. The procedure confirmed that ACD concentration waspreserved along the 20-minute experimental procedure (Fig. 1).
Experimental Procedures
Before starting the paradigm, rats were water-deprived for24 hours. Then, they underwent 3 cycles of testing, and each cyclewas composed by 3 sections, as reported in the timeline of theexperimental procedure (Fig. 2). At the end of each experimentalsession, rats were allowed to drink for 1 hour, and the intake ofwater was recorded.
ACETALDEHYDE-INDUCED OPERANTCONDITIONING 1279

Operant Self-Administration
Shaping and Training. Animals were shaped to lever press toobtain water on a continuous reinforcement schedule (fixed ratio 1,FR-1), until they reached the same level of performance. Afterwardtraining started and rats orally self-administered ACD solution orwater in the operant chamber (FR-1) along 21 days. For each tap-ping, the system delivered 0.05 ml of water or 0.90%ACD solution.The number of lever presses was automatically recorded; water andACD intake were measured by multiplying the number of leverpresses per 0.05 ml, and this value corresponded to the amount ofliquid missing from the reservoir at the end of each session. Animalswere tested each day at the same time (9:00 to 14:00).
Conflict
This protocol represented a modification of the Geller–Seifterparadigm, a conflict procedure where a positive reinforcer (water orACD solution) is earned by an operant response; however, during acomponent of the test session, delivery of the positive reinforcer is
coupled with delivery of an aversive stimulus, commonly foot-shock. In our study, the animals underwent to an alternation ofunpunished and punished responses according to “3 minutes–1 minute” schedule as reported in (Fig. 2). The session started andended with an unpunished interval.
The punished responses were signaled by a cue-light triggered bythe response. During the “punished” response interval, lever presseswere rewarded with the solution and delivered with a mild foot-shock of 0.2 mA. The number of unpunished and punishedresponses was automatically recorded. Data have also been pre-sented as a ratio between ACD and CTR’ unpunished and punishedresponses.
Deprivation
During the 7-day deprivation period, rats were left undisturbedin their home cages and received water and food ad libitum. ACDself-administration was suspended to achieve a forced abstinence.
Relapse
After each deprivation period, rats were exposed again to lever-pressing in the operant chamber with a FR-1 response schedule, for7 days. Responses were then recorded along the 3 relapse experi-mental sessions and compared among them and to the lever pressesin the last week of the training period. Water and ACD solutionintake was also recorded.
Statistical Analysis
Operant-Drinking behavior. A 2-way analysis of variance(ANOVA) was conducted on the number of lever presses during thetraining, conflict, and relapse, as dependent variables, with “ACDself-administration” (treatment) as between-subjects factor, and“days” as repeated measurement factor. When necessary, simplemain effects and post hoc comparisons were calculated with Bonfer-roni post-test (a = 0.05). Differences were considered statisticallysignificant if p < 0.05. Furthermore, to compare ACD intake during
Fig. 1. UV spectrum analysis of acetaldehyde concentration in the drinksolution at the beginning (t = 0) and at the end (t = 1) of the 20 minuteself-administration sessions.
Fig. 2. Timeline showing the schematic sequence of behavioral testing (A) and the modification of the Geller–Seifter conflict paradigm (B).
1280 CACACE ET AL.
the relapse session with basal intake a 2-tailed Student’s t-test forpaired measures was employed.
OF test. Locomotor activity evaluation was conducted perform-ing a 2-tailed Student’s t-test for unpaired measures on TDT, NTC,and ACT of the arena.
RESULTS
Open Field
Rats were tested in the OF arena at the end of the trainingsession of the operant task, to assess locomotor activity andbehavioral reactivity in response to novelty. Statistical analy-sis by a 2-tailed Student’s t-test, performed on the differentparameters, indicated that there were not significant differ-ences in TDT (t = 0.3428, df = 30; p < 0.7341), NTC(t = 1.424, df = 30; p < 0.1657), and ATC (t = 0.3669,df = 30; p < 0.7163), between the 2 groups of animals.
Training Period
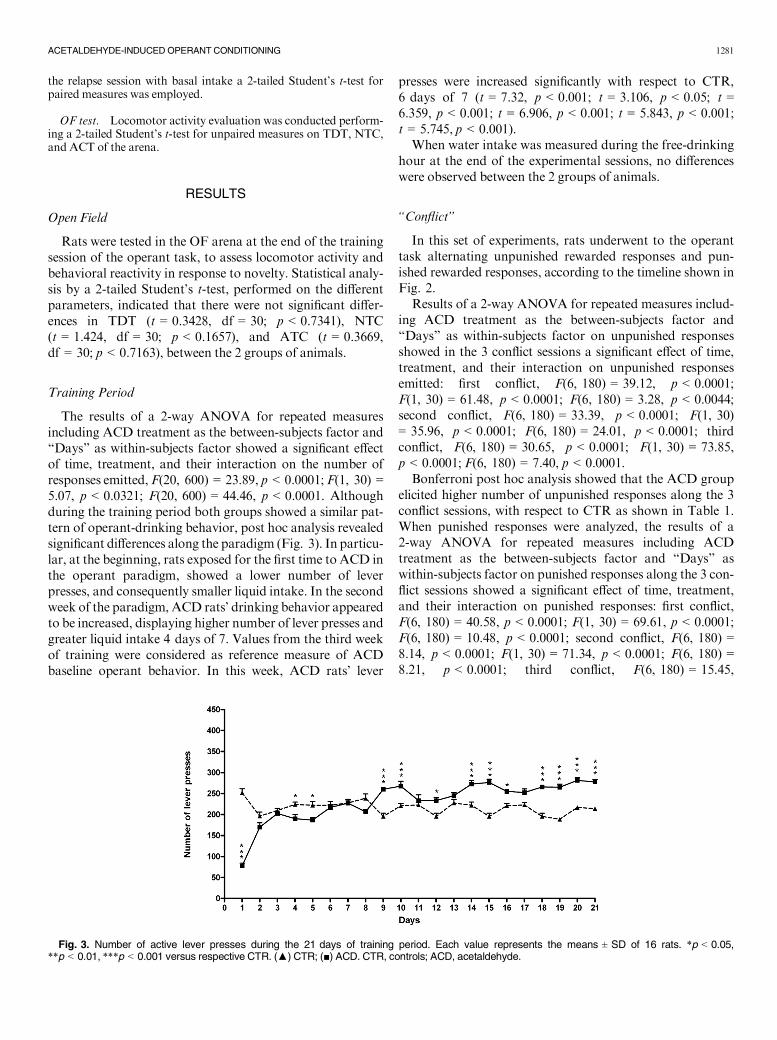
The results of a 2-way ANOVA for repeated measuresincluding ACD treatment as the between-subjects factor and“Days” as within-subjects factor showed a significant effectof time, treatment, and their interaction on the number ofresponses emitted, F(20, 600) = 23.89, p < 0.0001; F(1, 30) =5.07, p < 0.0321; F(20, 600) = 44.46, p < 0.0001. Althoughduring the training period both groups showed a similar pat-tern of operant-drinking behavior, post hoc analysis revealedsignificant differences along the paradigm (Fig. 3). In particu-lar, at the beginning, rats exposed for the first time to ACD inthe operant paradigm, showed a lower number of leverpresses, and consequently smaller liquid intake. In the secondweek of the paradigm, ACD rats’ drinking behavior appearedto be increased, displaying higher number of lever presses andgreater liquid intake 4 days of 7. Values from the third weekof training were considered as reference measure of ACDbaseline operant behavior. In this week, ACD rats’ lever
presses were increased significantly with respect to CTR,6 days of 7 (t = 7.32, p < 0.001; t = 3.106, p < 0.05; t =6.359, p < 0.001; t = 6.906, p < 0.001; t = 5.843, p < 0.001;t = 5.745, p < 0.001).
When water intake was measured during the free-drinkinghour at the end of the experimental sessions, no differenceswere observed between the 2 groups of animals.
“Conflict”
In this set of experiments, rats underwent to the operanttask alternating unpunished rewarded responses and pun-ished rewarded responses, according to the timeline shown inFig. 2.
Results of a 2-way ANOVA for repeated measures includ-ing ACD treatment as the between-subjects factor and“Days” as within-subjects factor on unpunished responsesshowed in the 3 conflict sessions a significant effect of time,treatment, and their interaction on unpunished responsesemitted: first conflict, F(6, 180) = 39.12, p < 0.0001;F(1, 30) = 61.48, p < 0.0001; F(6, 180) = 3.28, p < 0.0044;second conflict, F(6, 180) = 33.39, p < 0.0001; F(1, 30)= 35.96, p < 0.0001; F(6, 180) = 24.01, p < 0.0001; thirdconflict, F(6, 180) = 30.65, p < 0.0001; F(1, 30) = 73.85,p < 0.0001; F(6, 180) = 7.40, p < 0.0001.
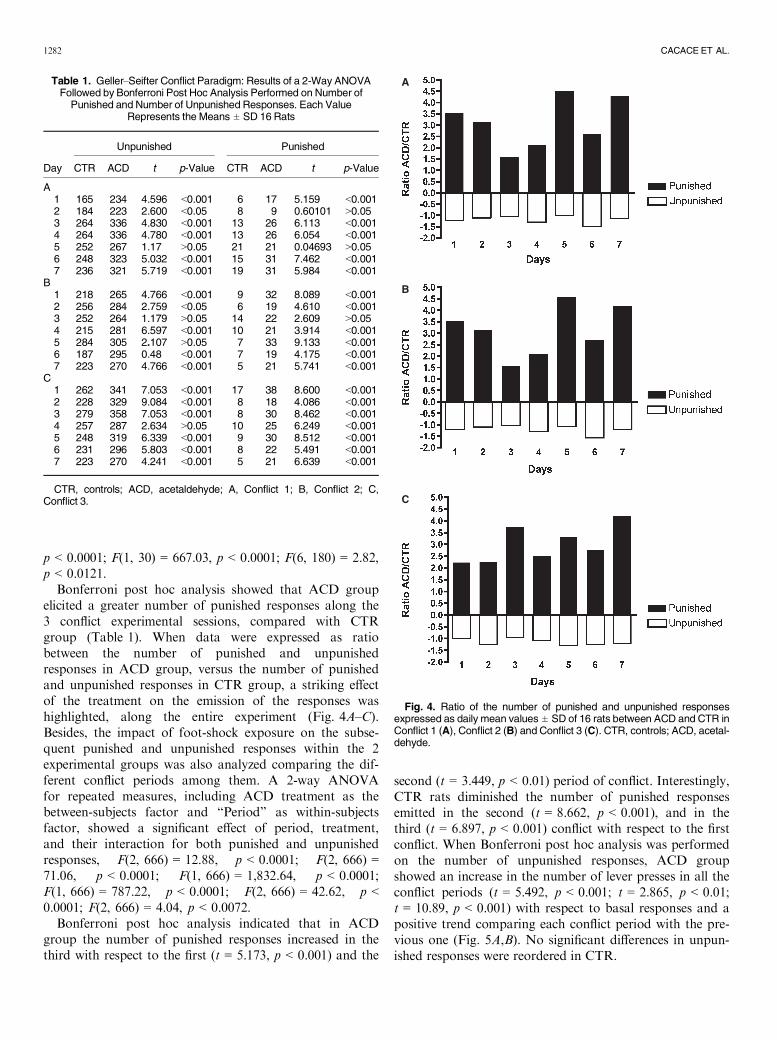
Bonferroni post hoc analysis showed that the ACD groupelicited higher number of unpunished responses along the 3conflict sessions, with respect to CTR as shown in Table 1.When punished responses were analyzed, the results of a2-way ANOVA for repeated measures including ACDtreatment as the between-subjects factor and “Days” aswithin-subjects factor on punished responses along the 3 con-flict sessions showed a significant effect of time, treatment,and their interaction on punished responses: first conflict,F(6, 180) = 40.58, p < 0.0001; F(1, 30) = 69.61, p < 0.0001;F(6, 180) = 10.48, p < 0.0001; second conflict, F(6, 180) =8.14, p < 0.0001; F(1, 30) = 71.34, p < 0.0001; F(6, 180) =8.21, p < 0.0001; third conflict, F(6, 180) = 15.45,
Fig. 3. Number of active lever presses during the 21 days of training period. Each value represents the means ± SD of 16 rats. *p < 0.05,**p < 0.01, ***p < 0.001 versus respective CTR. (▲) CTR; (■) ACD. CTR, controls; ACD, acetaldehyde.
ACETALDEHYDE-INDUCED OPERANTCONDITIONING 1281
p < 0.0001; F(1, 30) = 667.03, p < 0.0001; F(6, 180) = 2.82,p < 0.0121.
Bonferroni post hoc analysis showed that ACD groupelicited a greater number of punished responses along the3 conflict experimental sessions, compared with CTRgroup (Table 1). When data were expressed as ratiobetween the number of punished and unpunishedresponses in ACD group, versus the number of punishedand unpunished responses in CTR group, a striking effectof the treatment on the emission of the responses washighlighted, along the entire experiment (Fig. 4A–C).Besides, the impact of foot-shock exposure on the subse-quent punished and unpunished responses within the 2experimental groups was also analyzed comparing the dif-ferent conflict periods among them. A 2-way ANOVAfor repeated measures, including ACD treatment as thebetween-subjects factor and “Period” as within-subjectsfactor, showed a significant effect of period, treatment,and their interaction for both punished and unpunishedresponses, F(2, 666) = 12.88, p < 0.0001; F(2, 666) =71.06, p < 0.0001; F(1, 666) = 1,832.64, p < 0.0001;F(1, 666) = 787.22, p < 0.0001; F(2, 666) = 42.62, p <0.0001; F(2, 666) = 4.04, p < 0.0072.
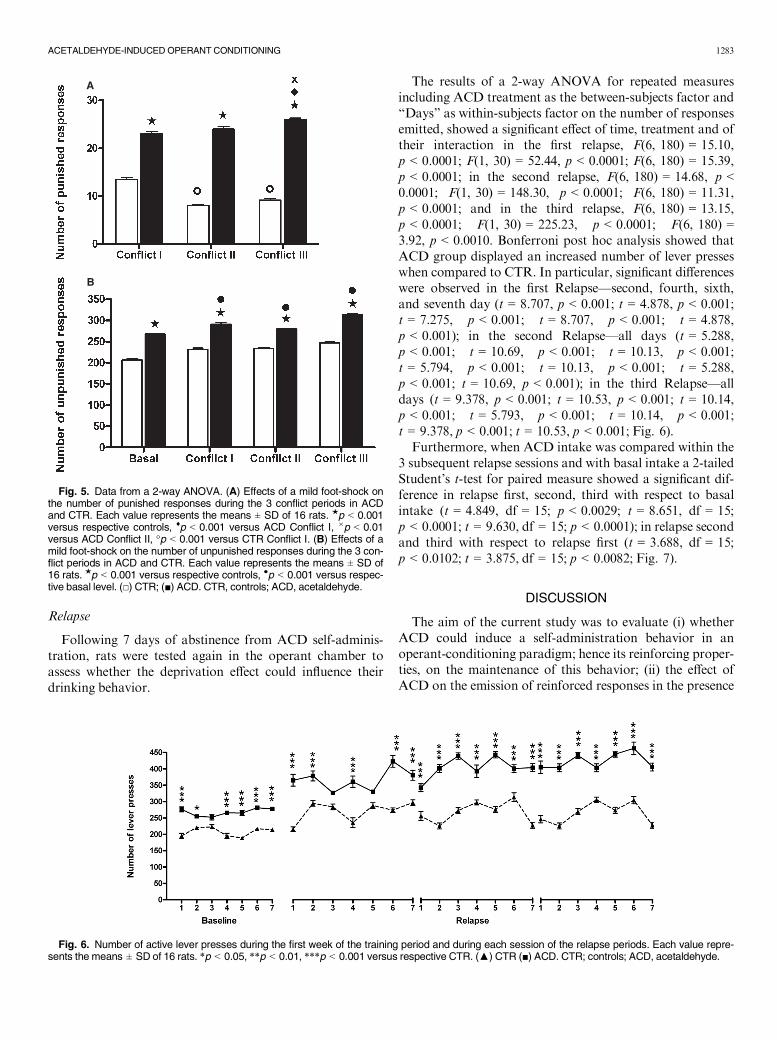
Bonferroni post hoc analysis indicated that in ACDgroup the number of punished responses increased in thethird with respect to the first (t = 5.173, p < 0.001) and the
second (t = 3.449, p < 0.01) period of conflict. Interestingly,CTR rats diminished the number of punished responsesemitted in the second (t = 8.662, p < 0.001), and in thethird (t = 6.897, p < 0.001) conflict with respect to the firstconflict. When Bonferroni post hoc analysis was performedon the number of unpunished responses, ACD groupshowed an increase in the number of lever presses in all theconflict periods (t = 5.492, p < 0.001; t = 2.865, p < 0.01;t = 10.89, p < 0.001) with respect to basal responses and apositive trend comparing each conflict period with the pre-vious one (Fig. 5A,B). No significant differences in unpun-ished responses were reordered in CTR.
Table 1. Geller–Seifter Conflict Paradigm: Results of a 2-Way ANOVAFollowed by Bonferroni Post Hoc Analysis Performed on Number of
Punished and Number of Unpunished Responses. Each ValueRepresents the Means ± SD 16 Rats
Day
Unpunished Punished
CTR ACD t p-Value CTR ACD t p-Value
A1 165 234 4.596 <0.001 6 17 5.159 <0.0012 184 223 2.600 <0.05 8 9 0.60101 >0.053 264 336 4.830 <0.001 13 26 6.113 <0.0014 264 336 4.780 <0.001 13 26 6.054 <0.0015 252 267 1.17 >0.05 21 21 0.04693 >0.056 248 323 5.032 <0.001 15 31 7.462 <0.0017 236 321 5.719 <0.001 19 31 5.984 <0.001
B1 218 265 4.766 <0.001 9 32 8.089 <0.0012 256 284 2.759 <0.05 6 19 4.610 <0.0013 252 264 1.179 >0.05 14 22 2.609 >0.054 215 281 6.597 <0.001 10 21 3.914 <0.0015 284 305 2.107 >0.05 7 33 9.133 <0.0016 187 295 0.48 <0.001 7 19 4.175 <0.0017 223 270 4.766 <0.001 5 21 5.741 <0.001
C1 262 341 7.053 <0.001 17 38 8.600 <0.0012 228 329 9.084 <0.001 8 18 4.086 <0.0013 279 358 7.053 <0.001 8 30 8.462 <0.0014 257 287 2.634 >0.05 10 25 6.249 <0.0015 248 319 6.339 <0.001 9 30 8.512 <0.0016 231 296 5.803 <0.001 8 22 5.491 <0.0017 223 270 4.241 <0.001 5 21 6.639 <0.001
CTR, controls; ACD, acetaldehyde; A, Conflict 1; B, Conflict 2; C,Conflict 3.
A
B
C
Fig. 4. Ratio of the number of punished and unpunished responsesexpressed as daily mean values ± SD of 16 rats between ACD and CTR inConflict 1 (A), Conflict 2 (B) and Conflict 3 (C). CTR, controls; ACD, acetal-dehyde.
1282 CACACE ET AL.
Relapse
Following 7 days of abstinence from ACD self-adminis-tration, rats were tested again in the operant chamber toassess whether the deprivation effect could influence theirdrinking behavior.
The results of a 2-way ANOVA for repeated measuresincluding ACD treatment as the between-subjects factor and“Days” as within-subjects factor on the number of responsesemitted, showed a significant effect of time, treatment and oftheir interaction in the first relapse, F(6, 180) = 15.10,p < 0.0001; F(1, 30) = 52.44, p < 0.0001; F(6, 180) = 15.39,p < 0.0001; in the second relapse, F(6, 180) = 14.68, p <0.0001; F(1, 30) = 148.30, p < 0.0001; F(6, 180) = 11.31,p < 0.0001; and in the third relapse, F(6, 180) = 13.15,p < 0.0001; F(1, 30) = 225.23, p < 0.0001; F(6, 180) =3.92, p < 0.0010. Bonferroni post hoc analysis showed thatACD group displayed an increased number of lever presseswhen compared to CTR. In particular, significant differenceswere observed in the first Relapse—second, fourth, sixth,and seventh day (t = 8.707, p < 0.001; t = 4.878, p < 0.001;t = 7.275, p < 0.001; t = 8.707, p < 0.001; t = 4.878,p < 0.001); in the second Relapse—all days (t = 5.288,p < 0.001; t = 10.69, p < 0.001; t = 10.13, p < 0.001;t = 5.794, p < 0.001; t = 10.13, p < 0.001; t = 5.288,p < 0.001; t = 10.69, p < 0.001); in the third Relapse—alldays (t = 9.378, p < 0.001; t = 10.53, p < 0.001; t = 10.14,p < 0.001; t = 5.793, p < 0.001; t = 10.14, p < 0.001;t = 9.378, p < 0.001; t = 10.53, p < 0.001; Fig. 6).
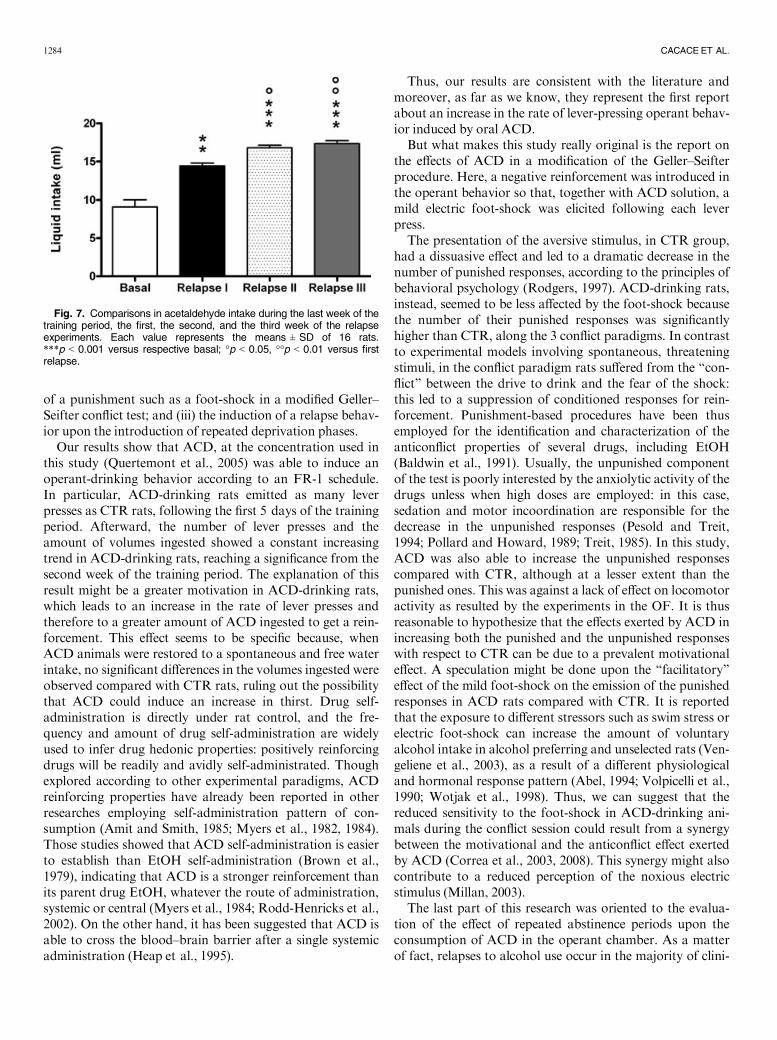
Furthermore, when ACD intake was compared within the3 subsequent relapse sessions and with basal intake a 2-tailedStudent’s t-test for paired measure showed a significant dif-ference in relapse first, second, third with respect to basalintake (t = 4.849, df = 15; p < 0.0029; t = 8.651, df = 15;p < 0.0001; t = 9.630, df = 15; p < 0.0001); in relapse secondand third with respect to relapse first (t = 3.688, df = 15;p < 0.0102; t = 3.875, df = 15; p < 0.0082; Fig. 7).
DISCUSSION
The aim of the current study was to evaluate (i) whetherACD could induce a self-administration behavior in anoperant-conditioning paradigm; hence its reinforcing proper-ties, on the maintenance of this behavior; (ii) the effect ofACD on the emission of reinforced responses in the presence
A
B
Fig. 5. Data from a 2-way ANOVA. (A) Effects of a mild foot-shock onthe number of punished responses during the 3 conflict periods in ACDand CTR. Each value represents the means ± SD of 16 rats. ★p < 0.001versus respective controls, ♦p < 0.001 versus ACD Conflict I, 9p < 0.01versus ACD Conflict II, °p < 0.001 versus CTR Conflict I. (B) Effects of amild foot-shock on the number of unpunished responses during the 3 con-flict periods in ACD and CTR. Each value represents the means ± SD of16 rats. ★p < 0.001 versus respective controls, ●p < 0.001 versus respec-tive basal level. (□) CTR; (■) ACD. CTR, controls; ACD, acetaldehyde.
Fig. 6. Number of active lever presses during the first week of the training period and during each session of the relapse periods. Each value repre-sents the means ± SD of 16 rats. *p < 0.05, **p < 0.01, ***p < 0.001 versus respective CTR. (▲) CTR (■) ACD. CTR; controls; ACD, acetaldehyde.
ACETALDEHYDE-INDUCED OPERANTCONDITIONING 1283
of a punishment such as a foot-shock in a modified Geller–Seifter conflict test; and (iii) the induction of a relapse behav-ior upon the introduction of repeated deprivation phases.
Our results show that ACD, at the concentration used inthis study (Quertemont et al., 2005) was able to induce anoperant-drinking behavior according to an FR-1 schedule.In particular, ACD-drinking rats emitted as many leverpresses as CTR rats, following the first 5 days of the trainingperiod. Afterward, the number of lever presses and theamount of volumes ingested showed a constant increasingtrend in ACD-drinking rats, reaching a significance from thesecond week of the training period. The explanation of thisresult might be a greater motivation in ACD-drinking rats,which leads to an increase in the rate of lever presses andtherefore to a greater amount of ACD ingested to get a rein-forcement. This effect seems to be specific because, whenACD animals were restored to a spontaneous and free waterintake, no significant differences in the volumes ingested wereobserved compared with CTR rats, ruling out the possibilitythat ACD could induce an increase in thirst. Drug self-administration is directly under rat control, and the fre-quency and amount of drug self-administration are widelyused to infer drug hedonic properties: positively reinforcingdrugs will be readily and avidly self-administrated. Thoughexplored according to other experimental paradigms, ACDreinforcing properties have already been reported in otherresearches employing self-administration pattern of con-sumption (Amit and Smith, 1985; Myers et al., 1982, 1984).Those studies showed that ACD self-administration is easierto establish than EtOH self-administration (Brown et al.,1979), indicating that ACD is a stronger reinforcement thanits parent drug EtOH, whatever the route of administration,systemic or central (Myers et al., 1984; Rodd-Henricks et al.,2002). On the other hand, it has been suggested that ACD isable to cross the blood–brain barrier after a single systemicadministration (Heap et al., 1995).
Thus, our results are consistent with the literature andmoreover, as far as we know, they represent the first reportabout an increase in the rate of lever-pressing operant behav-ior induced by oral ACD.
But what makes this study really original is the report onthe effects of ACD in a modification of the Geller–Seifterprocedure. Here, a negative reinforcement was introduced inthe operant behavior so that, together with ACD solution, amild electric foot-shock was elicited following each leverpress.
The presentation of the aversive stimulus, in CTR group,had a dissuasive effect and led to a dramatic decrease in thenumber of punished responses, according to the principles ofbehavioral psychology (Rodgers, 1997). ACD-drinking rats,instead, seemed to be less affected by the foot-shock becausethe number of their punished responses was significantlyhigher than CTR, along the 3 conflict paradigms. In contrastto experimental models involving spontaneous, threateningstimuli, in the conflict paradigm rats suffered from the “con-flict” between the drive to drink and the fear of the shock:this led to a suppression of conditioned responses for rein-forcement. Punishment-based procedures have been thusemployed for the identification and characterization of theanticonflict properties of several drugs, including EtOH(Baldwin et al., 1991). Usually, the unpunished componentof the test is poorly interested by the anxiolytic activity of thedrugs unless when high doses are employed: in this case,sedation and motor incoordination are responsible for thedecrease in the unpunished responses (Pesold and Treit,1994; Pollard and Howard, 1989; Treit, 1985). In this study,ACD was also able to increase the unpunished responsescompared with CTR, although at a lesser extent than thepunished ones. This was against a lack of effect on locomotoractivity as resulted by the experiments in the OF. It is thusreasonable to hypothesize that the effects exerted by ACD inincreasing both the punished and the unpunished responseswith respect to CTR can be due to a prevalent motivationaleffect. A speculation might be done upon the “facilitatory”effect of the mild foot-shock on the emission of the punishedresponses in ACD rats compared with CTR. It is reportedthat the exposure to different stressors such as swim stress orelectric foot-shock can increase the amount of voluntaryalcohol intake in alcohol preferring and unselected rats (Ven-geliene et al., 2003), as a result of a different physiologicaland hormonal response pattern (Abel, 1994; Volpicelli et al.,1990; Wotjak et al., 1998). Thus, we can suggest that thereduced sensitivity to the foot-shock in ACD-drinking ani-mals during the conflict session could result from a synergybetween the motivational and the anticonflict effect exertedby ACD (Correa et al., 2003, 2008). This synergy might alsocontribute to a reduced perception of the noxious electricstimulus (Millan, 2003).
The last part of this research was oriented to the evalua-tion of the effect of repeated abstinence periods upon theconsumption of ACD in the operant chamber. As a matterof fact, relapses to alcohol use occur in the majority of clini-
Fig. 7. Comparisons in acetaldehyde intake during the last week of thetraining period, the first, the second, and the third week of the relapseexperiments. Each value represents the means ± SD of 16 rats.***p < 0.001 versus respective basal; °p < 0.05, °°p < 0.01 versus firstrelapse.
1284 CACACE ET AL.

cal cases if no appropriate treatment is provided, indicatingthat relapse following periods of abstinence should beconsidered as one of the central features of the addictivebehavior. In our model, ACD-deprived rats underwent to arepresentation of ACD, and their ACD intake wasmeasured. According to evidence of increased alcohol con-sumption following periods of abstinence, ACD-drinkingrats displayed higher number of lever presses and a signifi-cant increase in liquid intake compared to CTR. Moreover,when ACD intake during the different relapse sessions wascompared with basal ACD intake, a significant effect ofdeprivation was recorded. Interestingly, ACD intake in thesecond and third week of relapse was significantly higherthan in the first week. The increase in ACD intake followingeach deprivation time might be caused by an increase in thereinforcing value of ACD; alternatively ACD itself may beacting as a cue or as a priming stimulus, as reported withalcohol. Indeed, alcohol deprivation effect in long-term vol-untary alcohol drinking rats is used as a measure of craving;that is, high motivation to drink alcohol as a measure forrelapse-like behavior (Holter et al., 1997, 1998; Spanagel andHolter, 1999). In summary, these findings suggest a moti-vated operant-drinking behavior which reflects an incentivedemand for ACD, apart from any other possible reason thanmotivation (no thirst-quenching effect). As far as we know,this is the first report about relapse in ACD-drinking behav-ior using an operant paradigm, and we believe that this evi-dence represent a further prove of the relevance of ACD inthe mediation of alcohol effects in the brain. Accordingly,recent reports show that EtOH acts as a prodrug and mustbe metabolized into ACD in the brain to generate rewardand reinforcement (Karahanian et al., 2011).
Indeed, the role of ACD in the central effects of EtOH isstill a matter of debate and this topic has been investigated onbehavioral and biochemical fields. In particular, studies basedon the use of alcohol or ACD dehydrogenases inhibitors (i.e.,4-methyl pyrazole or cyanamide), of catalase inhibitors (i.e.,3-amino triazole or sodium azide), or of ACD-sequesteringagents (D-penicillamine) have consistently shown that theeffects of EtOH on locomotor activity (Arizzi-LaFranceet al., 2006), anxiety (Arizzi et al., 2003; Correa et al., 2003,2008), HPA axis activation (Cannizzaro et al., 2010), andmotivation (Peana et al., 2008, 2009) appear to be mediatedby ACD (Aragon and Amit, 1992). In addition, although themechanism(s) by which ACD elicits its central effects remainslargely obscure, this compoundmay be responsible of its ownand also EtOH’s motivational properties, by activating dopa-minergic firing in the VTA (Foddai et al., 2004; Melis et al.,2007), like all drugs of abuse.
In conclusion, our results indicate that ACD is able toinduce, as well as alcohol, an operant-drinking behavior,which is also maintained besides the conflict procedure, andenhanced by the deprivation effect, supporting the hypothe-sis that ACD itself possesses motivational properties such usalcohol and other substances of abuse.
The modulation of alcohol-derived ACD, either by reduc-ing its production and/or by using sequestrating agents, mayexert a profound influence on the reinforcing and discrimina-tive effects of alcohol and might also accelerate the develop-ment of more effective therapeutic interventions to reducethe incidence of alcohol abuse and alcoholism.
ACKNOWLEDGMENTS
Supported by a grant from PRIN (MIUR, 2006). Wewould like to thank Dr. Giuseppe Avellone for his precioustechnical help.
REFERENCES
Abel EL (1994) A further analysis of physiological changes in rats in the
forced swim test. Physiol Behav 56:795–800.Amit Z, Smith BR (1985) A multi-dimensional examination of the positive
reinforcing properties of acetaldehyde. Alcohol 2:367–370.Aragon CMG, Amit Z (1992) The effect of 3-amino-1,2,4-triazole on volun-
tary ethanol consumption: evidence for brain catalase involvement in the
mechanism of action. Neuropharmacology 31:709–712.Aragon CMG, Rogan F, Amit Z (1992) Ethanol metabolism in rat brain
homogenates by a catalase-H2O2 system. Biochem Pharmacol 44:93–98.Arizzi MN, Correa M, Betz AJ, Wisniecki A, Salamone JD (2003)
Behavioral effects of intraventricular injections of low doses of ethanol,
acetaldehyde, and acetate in rats: studies with low and high rate operant
schedules. Behav Brain Res 147:203–210.Arizzi-LaFrance MN, CorreaM, Aragon CMG, Salamone JD (2006) Motor
stimulant effects of ethanol injected into the substantia nigra pars
reticulata: importance of catalase-mediated metabolism and the role of
acetaldehyde. Neuropsychopharmacology 31:997–1008.Aston-Jones G, Harris GC (2004) Brain substrates for increased drug seeking
during protracted withdrawal. Neuropharmacology 47:167–179.Baldwin HA,Wall TL, Schuckit MA, Koob GF (1991) Differential effects of
ethanol on punished responding in the P and NP rats. Alcohol Clin Exp
Res 15:700–704.
Brown ZW, Amit Z, Rockman GE (1979) Intraventricular self-administra-
tion of acetaldehyde, but not ethanol, in naive laboratory rats. Psycho-
pharmacology 64:271–276.Burattini C, Gill TM, Aicardi G, Janak PH (2006) The ethanol self-adminis-
tration context as a reinstatement cue: acute effects of naltrexone. Neuro-
science 139:877–887.Cannizzaro C, La BarberaM, Plescia F, Cacace S, Tringali G (2010) Ethanol
modulates corticotropin releasing hormone release from the rat hypothala-
mus: does acetaldehyde play a role? Alcohol Clin Exp Res 34:1–6.Cooper ZD, Shi YG, Woods JH (2010) Reinforcer-dependent enhancement
of operant responding in opioid-withdrawn rats. Psychopharmacology
212:369–378.Correa M, Arizzi MN, Betz A, Mingote S, Salamone JD (2003) Open field
locomotor effects in rats after intraventricular injections of ethanol and the
ethanol metabolites acetaldehyde and acetate. Brain Res Bull 62:197–202.
CorreaM,Manrique HM, Font L, Escrig MA, Aragon CMG (2008) Reduc-
tion in the anxiolytic effects of ethanol by centrally formed acetaldehyde:
the role of catalase inhibitors and acetaldehyde-sequestering agents.
Psychopharmacology 200:455–464.De Luca MT, Badiani A (2011) Ketamine self-administration in the rat:
evidence for a critical role of setting. Psychopharmacology 214:549–556.
Deng XS, Deitrich RA (2008) Putative role of brain acetaldehyde in ethanol
addiction. Curr Drug Abuse Rev 1:3–8.Eckard MJ, File SE, Gessa GL, Grant KA, Guerri C, Hoffman PL, Kalant
H, Kado GF, Li TK, Tobakoff B (1998) Effects of moderate alcohol
ACETALDEHYDE-INDUCED OPERANTCONDITIONING 1285

consumption on the central nervous system. Alcohol Clin Exp Res 22:998–1040.
Enrico P, Sirca D, Mereu M, Peana AT, Lintas A, Golosio A, Diana M
(2009) Acetaldehyde sequestering prevents ethanol-induced stimulation of
mesolimbic dopamine transmission. Drug Alcohol Depend 100:265–271.Ettenberg A (1989) Dopamine, neuroleptics and reinforced behavior. Neuro-
sci Biobehav Rev 13:105–111.Foddai M, Dosia G, Spiga S, Diana M (2004) Acetaldehyde increases dopa-
minergic neuronal activity in the VTA. Neuropsychopharmacology 29:
530–536.Gardner EL (2002) Addictive potential of cannabinoids: the underlying
neurobiology. Chem Phys Lipids 121:267–290.Hamby-Mason R, Chen JJ, Schenker S, Perez A, Henderson GI (1997) Cata-
lase mediates acetaldehyde formation from ethanol in fetal and neonatal
rat brain 1997. Alcohol Clin Exp Res 21:1063–1072.Hansson T, Tindberg N, Ingelman-Sundberg M, Kohler C (1990) Regional
distribution of ethanol-inducible cytochrome P450 IIE1 in the rat central
nervous system. Neuroscience 34:451–463.Heap L, Ward RJ, Abiaka C, Dexter D, Lawlor M, Pratt O, Thomson A,
ShawK, Peters TJ (1995) The influence of brain acetaldehyde on oxidative
status, dopamine metabolism and visual discrimination task. Biochem
Pharmacol 50:263–270.Heinz A, Lober S, Georgi A, Wrase J, Hermann D, Rey E-R, Wellek S,
Mann K (2003) Reward craving and withdrawal relief craving: assessment
of different motivational pathways to alcohol intake. Alcohol Alcohol
38:35–39.Higgins GA, Joharchi N, Wang Y, Corrigall WA, Sellers EM (1994) The
CCK(A) receptor antagonist devazepide does not modify opioid self-
administration or drug discrimination: comparison with the dopamine
antagonist haloperidol. Brain Res 640:246–254.Holter SM, Engelmann M, Kirschke C, Liebsch G, Landgraf R, Spanagel R
(1998) Long-term ethanol self-administration with repeated ethanol depri-
vation episodes changes ethanol drinking pattern and increases anxiety-
related behaviour during ethanol deprivation in rats. Behav Pharmacol
9:41–48.Hölter SM, Landgraf R, Zieglgänsberger W, Spanagel R (1997) Time course
of acamprosate action on operant ethanol self- administration after etha-
nol deprivation. Alcohol Clin Exp Res 21:862–868.Hunt WA (1996) Role of acetaldehyde in the actions of ethanol on the brain
—a review. Alcohol 13:147–151.
Karahanian E, Quintanilla ME, Tampier L, Rivera-Meza M, Bustamante
D, Lira VG, Morales P, Herrera-Marschitz M, Israel Y (2011) Ethanol as
a prodrug: brain metabolism of ethanol mediates its reinforcing effects.
Alcohol Clin Exp Res 35:606–612.KochM, Schmid A, Schnitzler HU (2000) Role of nucleus accumbens dopa-
mine D1 and D2 receptors in instrumental and pavlovian paradigms of
conditioned reward. Psychopharmacology (Berl) 152:67–73.Koob GF (1992) Dopamine, addiction and reward. Semin Neurosci 4:139–148.
Koob GF, Ahmed SH, Boutrel B, Chen SA, Kenny PJ, Markou A, O’Dell
LE, Parsons LH, Sanna PP (2004) Neurobiological mechanisms in the
transition from drug use to drug dependence. Neurosci Biobehav Rev
27:739–749.Manzardo AM, Stein L, Belluzzi JD (2002) Rats prefer cocaine over nicotine
in a two-lever self-administration choice test. Brain Res 924:10–19.Martınez SE, Vaglenova J, Sabria J, Martınez MC, Farres J, Pares X (2001)
Disribution of alcohol dehydrogenase mRNA in the rat central nervous
system. Consequences for brain ethanol and retinoid metabolism. Eur J
Biochem 268:5045–5056.
Melis M, Enrico P, Peana AT, Diana M (2007) Acetaldehyde mediates
alcohol activation of the mesolimbic dopamine system. Eur J Neurosci
26:2824–2833.Millan MJ (2003) The neurobiology and control of anxious states. Prog
Neurobiol 70:83–244.Montoliu C, Sancho-Tello M, Azorin I, Burgal M, Valles S, Renau-Piqueras
J, Guerri C (1995) Ethanol increases cytochrome P4502E1 and induces
oxidative stress in astrocytes. J Neurochem 65:2561–2570.
Myers WD, Ng KT, Singer G (1982) Intravenous self-administration of
acetaldehyde in the rat as a function of schedule, food deprivation and
photoperiod. Pharmacol Biochem Behav 17:807–811.Myers WD, Ng KT, Singer G (1984) Effects of naloxone and buprenorphine
on intravenous acetaldehyde self-injection in rats. Physiol Behav 33:449–455.
Peana AT, Assaretti AR, Muggironi G, Enrico P, Diana M (2009) Reduc-
tion of ethanol-derived acetaldehyde induced motivational properties by
L-cysteine. Alcohol Clin Exp Res 33:43–48.Peana AT, Enrico P, Assaretti AR, Pulighe E, Muggironi G, Nieddu M,
Piga A, Lintas A, Diana M (2008) Key role of ethanol-derived acetalde-
hyde in the motivational properties induced by intragastric ethanol: a
conditioned place preference study in the rat. Alcohol Clin Exp Res
32:249–258.
Peana AT, Muggironi G, Diana M (2010) Acetaldehyde-reinforcing effects:
a study on oral self-administration behavior. Front Psychiatry 16:1–23.Pesold C, Treit D (1994) The septum and amygdala differentially mediate the
anxiolytic effects of benzodiazepines. Brain Res 638:295–301.
Pollard GT, Howard JL (1990) Effects of drugs on punished behavior: pre-
clinical test for anxiolytics. Pharmacol Ther 45:403–424.
Quertemont E, Grant KA, CorreaM, ArizziMN, Salamone JD, Tambour S,
Aragon CMG,McBrideWJ, Rodd ZA, Goldstein A, Zaffaroni A, Li T-K,
PisanoM, DianaM (2005) The role of acetaldehyde in the central effects of
ethanol. Alcohol Clin ExpRes 29:221–234.Roberts BJ, Shoaf SE, Jeong KS, Song BJ (1994) Induction of CYP2E1 in
liver, kidney, brain and intestine during chronic ethanol administration
and withdrawal: evidence that CYP2E1 possesses a rapid phase half-life of
6 hours or less. Biochem Biophys Res Commun 205:1064–1071.Rodd-Henricks ZA, Melendez RI, Zaffaroni A, Goldstein A, McBride WJ,
Li T-K (2002) The reinforcing effects of acetaldehyde in the posterior ven-
tral tegmental area of alcohol-preferring rats. Pharmacol Biochem Behav
72:55–64.Rodgers RJ (1997) Animal models of “anxiety”: where next? Behav Pharma-
col 8:477–496.Salamone JD (1994) The involvement of nucleus accumbens dopamine in
appetitive and aversive motivation. Behav Brain Res 61:117–133.Salamone JD, Cousins MS, McCullough LD, Carriero DL, Berkowitz RJ
(1994) Nucleus accumbens dopamine release increases during instrumental
lever pressing for food but not free food consumption. Pharmacol
Biochem Behav 49:25–31.Salamone JD, Cousins MS, Snyder BJ (1997) Behavioral functions of
nucleus accumbens dopamine: empirical and conceptual problems with
the anhedonia hypothesis. Neurosci Biobehav Rev 21:341–359.Schultz W (1998) Predictive reward signal of dopamine neurons. J Neuro-
physiol 80:1–27.Sippel HW (1974) The acetaldehyde content in rat brain during ethanol
metabolism. J Neurochem 23:451–452.Smith BR, Amit Z, Splawinsky J (1984) Conditioned place prefer-
ence induced by intraventricular infusions of acetaldehyde. Alcohol 1:193–195.
Spanagel R, Holter SM (1999) Long-term alcohol self-administration with
repeated alcohol deprivation phases: an animal model of alcoholism?
Alcohol Alcohol 34:231–243.Spanagel R, Weiss F (1999) The dopamine hypothesis of reward: past and
current status. Trends Neurosci 22:521–527.Tindberg N, Ingelman-SundbergM (1996) Expression, catalytic activity, and
inducibility of cytochrome P450 2E1 (CYP2E1) in the rat central nervous
system. J Neurophysiol 67:2066–2073.Treit D (1985) The inhibitory effect of diazepam on defensive burying: anxio-
lytic vs. analgesic effects. Pharmacol Biochem Behav 22:47–52.
Vengeliene V, Siegmund S, Singer MV, Sinclair JD, Li T-K, Spanagel RA
(2003) A comparative study on alcohol-preferring rat lines: effects of depri-
vation and stress phases on voluntary alcohol intake. Alcohol Clin Exp
Res 27:1048–1054.Volpicelli JR, Ulm RR, Hopson N (1990) The bidirectional effects of shock
on alcohol preference in rats. Alcohol Clin Exp Res 14:913–916.
1286 CACACE ET AL.

Westcott JY, Weiner H, Shultz J, Myers RD (1980) In vivo acetaldehyde in
the brain of the rat treated with ethanol. Biochem Pharmacol 29:411–417.Wise RA, BozarthMA (1985) Brain mechanisms of drug reward and eupho-
ria. Psychiatr Med 3:445–460.Wotjak CT, Ganster J, Kohl G, Holsboer F, Landgraf R, Engelmann M
(1998) Dissociated central and peripheral release of vasopressin, but not
oxytocin, in response to repeated swim stress: new insights into the secre-
tory capacities of peptidergic neurons. Neuroscience 85:1209–1222.
Young AMJ, Ahier RG, Upton RL, Joseph MH, Gray JA (1998) Increased
extracellular dopamine in the nucleus accumbens of the rat during associa-
tive learning of neutral stimuli. Neuroscience 83:1175–1183.Zimatkin SM, Liopo AV, Deitrich RA (1998) Distribution and kinetics of
ethanol metabolism in rat brain. Alcohol Clin Exp Res 22:1623–1627.Zimatkin SM, Pronko SP, Vasiliou V, Gonzalez FJ, Deitrich RA (2006)
Enzymatic mechanisms of ethanol oxidation in the brain. Alcohol Clin
Exp Res 30:1500–1505.
ACETALDEHYDE-INDUCED OPERANTCONDITIONING 1287
Copyright © 2022 FDOKUMEN




















