
TIME REPRODUCTION DISTURBANCES IN ADHD CHILDREN: AN ERP STUDY
18
Downloaded By: [Universidad de Sevilla] At: 04:47 30 November 2007 Intern. J. Neuroscience, 118:119–135, 2008 Copyright C 2008 Informa Healthcare USA, Inc. ISSN: 0020-7454 / 1543-5245 online DOI: 10.1080/00207450601042177 TIME REPRODUCTION DISTURBANCES IN ADHD CHILDREN: AN ERP STUDY A. A. GONZ ´ ALEZ-GARRIDO Instituto de Neurociencias Universidad de Guadalajara Guadalajara, M ´ exico and O.P.D.Hospital Civil de Guadalajara Guadalajara, M ´ exico F. R. G ´ OMEZ-VEL ´ AZQUEZ D. ZARABOZO Instituto de Neurociencias Universidad de Guadalajara Guadalajara, M ´ exico R. L ´ OPEZ-ELIZALDE A. ONTIVEROS O.P.D.Hospital Civil de Guadalajara Guadalajara, M ´ exico H. MADERA-CARRILLO O. L. VEGA Instituto de Neurociencias Universidad de Guadalajara Guadalajara, M ´ exico Received 4 July 2006. The authors thank Dr. Manuel Gonzalez for his helpful collaboration to perform this research. Address correspondence to Dr. Andr´ es Antonio Gonz´ alez-Garrido, Instituto de Neurociencias, Francisco de Quevedo 180. Col. Arcos Vallarta, Guadalajara Jalisco, 44130, M´ exico. E-mail: [email protected] 119
Transcript of TIME REPRODUCTION DISTURBANCES IN ADHD CHILDREN: AN ERP STUDY

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
Intern. J. Neuroscience, 118:119–135, 2008Copyright C© 2008 Informa Healthcare USA, Inc.ISSN: 0020-7454 / 1543-5245 onlineDOI: 10.1080/00207450601042177
TIME REPRODUCTION DISTURBANCES IN ADHDCHILDREN: AN ERP STUDY
A. A. GONZALEZ-GARRIDO
Instituto de NeurocienciasUniversidad de GuadalajaraGuadalajara, MexicoandO.P.D. Hospital Civil de GuadalajaraGuadalajara, Mexico
F. R. GOMEZ-VELAZQUEZD. ZARABOZO
Instituto de NeurocienciasUniversidad de GuadalajaraGuadalajara, Mexico
R. LOPEZ-ELIZALDEA. ONTIVEROS
O.P.D. Hospital Civil de GuadalajaraGuadalajara, Mexico
H. MADERA-CARRILLOO. L. VEGA
Instituto de NeurocienciasUniversidad de GuadalajaraGuadalajara, Mexico
Received 4 July 2006.The authors thank Dr. Manuel Gonzalez for his helpful collaboration to perform this research.Address correspondence to Dr. Andres Antonio Gonzalez-Garrido, Instituto de Neurociencias,
Francisco de Quevedo 180. Col. Arcos Vallarta, Guadalajara Jalisco, 44130, Mexico. E-mail:[email protected]
119

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
120 A. A. GONZALEZ-GARRIDO ET AL.
J. L. OROPEZA DE ALBA
O.P.D. Hospital Civil de GuadalajaraGuadalajara, Mexico
J. M. DE LA SERNA TUYA
Instituto de NeurocienciasUniversidad de GuadalajaraGuadalajara, MexicoandUniversidad de SevillaSeville, Spain
Sixteen ADHD children and a control group were asked to reproduce the varyingtime duration of successively presented visual stimuli. Time estimation was poorerin ADHD children, who showed more impulsive errors. ERPs exhibited similargrand-mean waveforms for both groups during the estimating period, but they weresignificantly different during the reproducing stage, when an early positive waveover frontal regions characterized the control group, interpreted as memory-guidedmotor output, followed by a slow negativity probably reflecting an inhibitory motorclosure process, both probably involving central executive networks that seem to beimproperly activated in ADHD children.
Keywords ADHD, central executive, children, ERP; time estimation, workingmemory
INTRODUCTION
Time perception can be considered as a complex cognitive and adaptive abilitythat assists the facility to predict, anticipate, and respond competently tofuture events. In this sense, accurate representation of transitory information isrequired to accomplish the ability to plan and organize adequate sequences ofbehavioral actions.
The neural processes underlying time perception and estimation remainunclear. Whether time perception relies on specific internal chronometricmechanisms or is a by-product of information processing is a matter ofcontroversy (Boltz, 1991; Church, 1984; Marmaras et al., 1995; Pastor &Artieda, 1996; Sawyer et al., 1994; Treisman et al., 1992; Zakay et al.,1994). However, it seems to be a general consensus that time perceptioncomprises several processes that engage multiple brain regions, including

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 121
the neocerebellum, basal ganglia, and prefrontal cortex (Casini & Ivry, 1999;Harrington et al., 1998; Xu et al., 2006).
It has been proposed that time perception relies on an internal clockemitting pulses at a particular rate, that are counted and employed subsequentlyby further components of the clock (Treisman et al., 1992). The temporalpacemaker seems to be sensitive to the arousal level, where, if attention isdedicated to the task of time estimation, the emitted clicks could pass uncheckedthus favoring underestimations (Treisman, 1984). In this regard, it is generallyaccepted that when another attentional demanding task is performed during theinterval that must be judged, attention is diverted from monitoring the passage oftime, so one could expect that monitoring the course of time requires attention.In fact, cortical systems had been proposed to be responsible for any attentionaleffect on time estimation (Chaston & Kingstone, 2004; Kingstone et al.,1995).
Attention deficit/hyperactivity disorder (ADHD) is a widespread develop-mental disorder that among other several clinical manifestations shows evidenceof time perception deficit. Barkley proposed the existence of a core deficitin ADHD: an impaired behavioral inhibition that is related to, and essentialfor, the executive functions that subserve self-regulation (Barkley 1997a,b,c).The Barkley’s model predicts deficits in working memory (WM) and in thedevelopment of sense of time and associated retrospective and prospectivefunctions as a consequence of poor behavioral inhibition, among other cognitivedeficits (Barkley et al., 1997).
There are multiple evidences of WM impairments in children with ADHD(Shallice et al., 2002; Martinussen et al., 2005) as well as time perception deficit(Barkley et al., 2001; Castellanos & Tannock, 2002; Kerns et al., 2001; Meaux& Chelonis, 2003; Smith et al., 2002; Toplak & Tannock, 2005). However,disturbance appears to depend on the type of task employed and the methodfor determining performance. Furthermore, Radonovich and Mostofsky (2004)postulated on page 162 that “there is a deficiency in the utilization of temporalinformation in ADHD (possibly secondary to deficits in working memoryand/or strategy utilization), rather than a problem involving a central timingmechanism.”
Earlier work suggests that ERP could measure possible physiologicalcorrelates of the time judgment neural basis, even suggesting that slow DCshifts in the ERP could be correlated with time-keeping functions (Brunia &Damen, 1988; Casini & Macar, 1993; Elbert et al., 1991; Johannes et al., 1997;Macar & Besson, 1985). On this basis the authors studied a time estimationtask in ADHD and normal children using ERP methods.

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
122 A. A. GONZALEZ-GARRIDO ET AL.
METHODS
Subjects
Sixteen male children aged from 7.5 to 10.5 years (X = 8.8; SD ± 1.5)were selected to integrate the experimental group. All children had a diagnosisof ADHD confirmed by a multidisciplinary clinical diagnostic assessment.The assessment comprised two semi-structured clinical diagnostic interviews(face-to-face interview with parents and teachers), standardized behavior ratingscales, and a comprehensive child assessment were completed by parentsand teacher. When applying the Diagnostic and Statistical Manual of MentalDisorders (DSM-IV-TR, APA, 2000) criteria, they were all diagnosed asADHD-predominantly inattentive type (314.00).
The ADHD group was matched to 16 healthy subjects based on age(X = 8.9; SD ± 1.4), handedness, gender, and educational level. Subjectsfrom both groups were right-handed, they all had a Wechsler IntelligenceScale for Children-Mexican Revision global IQ of 90 or higher, and they wereall regular students. They had no personal or familiar history of psychiatric,neurological, or degenerative illness, nor did they have received diagnosis ofbehavioral disorder. They all had a normal neurological examination and anormal baseline EEG. None of the participants were receiving medicationprior to the investigation. Experiment was conducted with the precedingfull understanding and written consent from the parents of the participants.The Neuroscience Institute ethical committee approved and supervised theexperiment.
Psychological Task Description
Subjects were tested seating in a comfortable chair at a sound-attenuated room.They were instructed to mentally calculate the duration of the stimulus in thescreen, and immediately reproduce its estimated time by pressing with theirright index finger a mouse-key. Subjects were comprehensively trained beforethe performance of the task. All stimuli were 25 millimeters diameter yellowcircles centered over a black background in a computer screen, located 60centimeters away from the subjects’ eyes. There were 80 stimuli presentedduring randomized periods of 2,500 ms (40 trials) or 5,500 ms (40 trials), withpost-stimulus interval of 5,000 and 8,000 ms, respectively.
Electrophysiological Methods
Recording. Electroencephalographic (EEG) activity was recorded from Fp1,Fp2, F3, F4, F7, F8, C3, C4, P3, P4, O1, O2, T3, T4, T5, T6, Fz, Cz, and Pz scalp

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 123
sites (Klem et al., 1999). The electrooculogram (EOG) was recorded from theouter canthus and infraocular orbital ridge of the right eye. Electrophysiologicalrecordings were done using 10 mm diameter gold disk electrodes (Grass TypeE5GH) and Grass electrode cream. All recording sites were referred to linkedmastoids. Interelectrode impedances were below 5 k� at 30 Hz. EEG and EOGsignals were amplified at a bandpass of 0.05–30 Hz (3-dB cuttoff points of6 dB/octave rolloff curves) with a sampling period of 4 ms on MEDICID-03E system. Data was collected as 1.100-s epochs for all recording channelsbeginning 100 ms prior to stimulus onset. Single trial data was analyzed off-linefor averaging and analysis.
Data Analysis
Behavioral Measures. The onset and conclusion of the stimuli and responseswere automatically marked on the EEG by software. The accuracy in timeestimation was measured through the following estimator:
Estimation Accuracy (EA) =Estimated time − Actual time
Actual time
Impulsive errors were defined as the trials in which the subjects starteda response before the conclusion of the stimulus presentation; they weremeasured and compared between the groups.
Signal Averaging. In this article two conditions were considered for furtheranalysis (estimating vs. reproducing time periods). Epochs of data on allchannels were excluded from averages when voltage in a given recordingepoch exceeded ±100 µV on any EEG or EOG channel. Epochs with artifactswere additionally rejected by visual inspection. In general, 6 to 10 epochs hadto be rejected in each condition per subject. Twenty free-artifact trials wereconsidered to obtain the individual ERP in each condition.
Statistical Analysis. Behavioral data comparisons were made using Student’st for paired groups. Electrophysiological data were analyzed using Split-plotAnalysis of Variance (Kirk, 1995). The Geisser–Greenhouse epsilon correctionwas utilized to modify the p values. Four time windows were used toexamine ERP-waveforms during reproducing-time condition: W1: 0–250; W2:250–450; W3: 450–650, and W4: 650–1000 ms respectively. Analyses ofvariance (ANOVAs: Group × Windows × Recording Site) were conductedon average voltage across each time window as dependent variable.

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
124 A. A. GONZALEZ-GARRIDO ET AL.
RESULTS
Behavioral Results
In general, time estimation accuracy was significantly poorer in ADHDgroup [t(15) = 3.314; p < 0.01)]. ADHD children made more impulsiveerrors than controls, but this tendency did not reach statistical significance.Time underestimations were predominant; however, their number was notsignificantly different between the groups. The estimation accuracy forunderestimated trials was significantly poorer in ADHD group [t(15) = 2.604;p < 0.05)].
On the other hand, the number of overestimations was significantly higherin the control group [t(15) = 3.156; p < 0.01)] but the inaccuracy in theoverestimated trials was slightly higher in ADHD children. See Table 1.
Event-Related Potentials Description
Group averaged event-related brain potentials showed remarkable similaritiesfor the examined groups during the estimating-time stage. Figure 1 representsERPs during the estimation period in both groups. There was a typical sequenceof peaks with magnitudes higher than 1 µv that was nominated as P1-N1-P2(with maximum at vertex over 220, at right frontal over 350, and 650 ms invertex, respectively) for further description.
In general, ERPs were almost similar for the two groups in P1 and morenegative in N1 only in the right prefrontal location. In addition, P2 was morepositive in frontocentral areas for the ADHD group.
The ERPs obtained during reproducing-time stage showed a distinguish-able different morphology than that described for the estimation period. In thissecond stage, ERPs in the control group exhibited an early P1 peaking around
Table 1. Behavioral results of ADHD and control groups in the time estimation task
Control (X ± SD) ADHD (X ± SD)
Underestimations 30.9 ± 7.8 34.4 ± 9.5E.A. of underestimations −.21 ± .14 −.31 ± .11∗Overestimations 14.0 ± 6.3 7.5 ± 5.7∗∗E.A. of overestimations .15 ± .06 .19 ± .09Impulsive errors 4.4 ± 4.1 7.5 ± 7.6
Total EA −0.07 ± 0.06 −0.19 ± 0.16∗∗
EA: estimation accuracy averaged; ∗p < 0.05; ∗∗p < 0.01.

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 125
Figure 1. Grand mean waveforms of the ERPs from ADHD children and Control Group duringthe time estimating period of the task.
140 ms in Fz and Cz, followed by a small negativity at vertex over 210 ms, anda subsequent very slow positive shift in parietal regions. See Figure 2.
The larger differences between the ERPs in the two groups, however, werelocated in the frontal areas, particularly in the prefrontal scalp sites where ERPsfrom the control group showed a long-lasting initial positivity, that extendedfrom the beginning of the trial until 450 ms approximately, with maximum over320 ms, followed by a slow and prolonged negative wave with a maximum at700 ms. Meanwhile, ERPs of the ADHD group exhibited a large slow negativeinitial shift with a maximum at 265 ms, followed by a lengthened low-voltagepositive wave with a maximum over 720 ms.
Electrophysiological Data Analysis
Visual inspection of the ERPs across the two conditions of the task showedthat main voltage changes occurred over frontal central regions. On this basis,statistical analysis was restricted to the difference-waveforms from locationsFp1, Fp2, F3, F4, Fz, Cz, and Pz at the four time windows previouslydefined.
Tests of within-subjects effects from split plot analysis of variance duringthe task estimation period showed that factor recording site was statistically

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
126 A. A. GONZALEZ-GARRIDO ET AL.
Figure 2. Grand mean waveforms of the ERPs from ADHD children and Control Group duringthe time reproducing period of the task.
significant [F(2.9,81) = 5.715, p < 0.01] as well as the interaction betweenrecording site and time window [F(7.7,215) = 12.052, p < 0.01]. During thereproduction period, factor time window was statistically significant [F(2.4,68)= 6.34, p < 0.01], as well as the interaction between time window and recordingsite [F(6.8,189) = 4.808, p < 0.01], and the interaction between group and timewindow [F(2.4,68) = 7.801, p < 0.01].
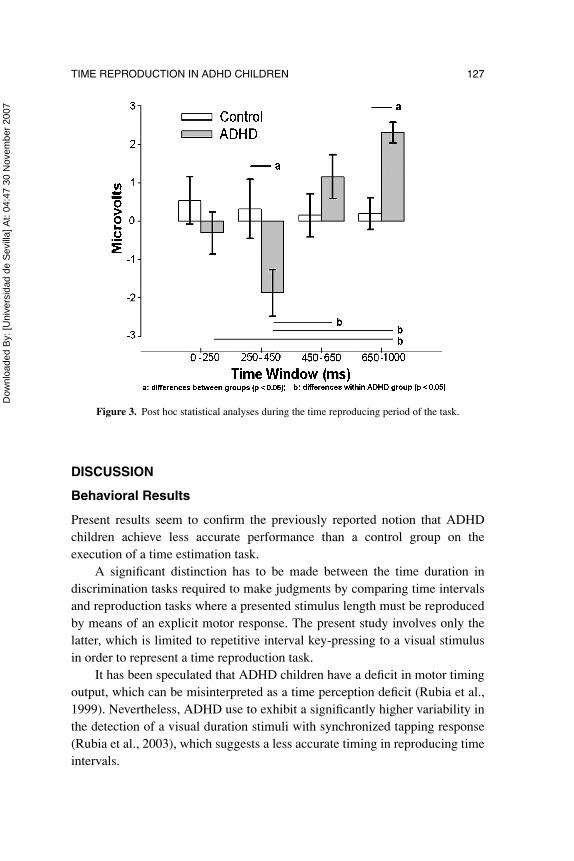
Differences between ADHD and the control group as well as withinADHD subjects were further analyzed for each time window and conditionwith randomized block factorial design ANOVAs considering two factors:recording site and group and with Tukey-Kramer post-hoc analyses. In thetime estimation period, voltage amplitude variations did not reach statisticalsignificance between groups in the studied recording sites, however, duringthe time reproduction period post-hoc analyses showed significant differencesbetween the groups in W2 and W4, whereas ADHD children exhibitedsignificant voltage magnitude variations in W4 with respect to W1 and W2,and between W2 and W3. (See Figure 3).
Finally, during the reproduction period of the task post-hoc analyses onlyshowed significant effects for recording site Fp1 in W2, and for Fp1, F3, F4,Fz, Cz, and Pz in W4.
Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 127
Figure 3. Post hoc statistical analyses during the time reproducing period of the task.
DISCUSSION
Behavioral Results
Present results seem to confirm the previously reported notion that ADHDchildren achieve less accurate performance than a control group on theexecution of a time estimation task.
A significant distinction has to be made between the time duration indiscrimination tasks required to make judgments by comparing time intervalsand reproduction tasks where a presented stimulus length must be reproducedby means of an explicit motor response. The present study involves only thelatter, which is limited to repetitive interval key-pressing to a visual stimulusin order to represent a time reproduction task.
It has been speculated that ADHD children have a deficit in motor timingoutput, which can be misinterpreted as a time perception deficit (Rubia et al.,1999). Nevertheless, ADHD use to exhibit a significantly higher variability inthe detection of a visual duration stimuli with synchronized tapping response(Rubia et al., 2003), which suggests a less accurate timing in reproducing timeintervals.

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
128 A. A. GONZALEZ-GARRIDO ET AL.
In the present study underestimated time reproductions were predominantin ADHD and control group. Considering that ADHD children who participatedin the experiment were previously classified as belonging to the inattentivesubtype, their particular inability to sustain attention could probably interferewith the “on-line” monitoring of the passage of time needed to fulfill thetask requirements. If the hypothesis of the temporal brain pacemaker isaccepted, one could expect that ADHD children errors could be mostly dueto underestimations of the time period. However, the results were similar withthe control group, so it seems more reasonable that the age of the subjects andthe nature of the task itself could better explain this sort of performance.
ADHD children had not only more errors on reproducing the presentedstimuli, but the inaccuracy of their responses was greater, regardless if timespan was under or overestimated. This result stress the temporal distortionproblems found in ADHD and it seems consistent with Barkley’s theory, whichemphasizes the presence of behavioral inhibition. In the other hand, the greaternumber of “impulsive errors” found in ADHD children appears to underpinthis idea.
ERPs Results
Regarding the behavioral task, one could expect that in order to establish thetime duration of the stimuli in the screen, children had, at least, visually identifythe stimulus occurrence and mark its onset and ending by probably upgradinga working memory store, which subsequently will serve to modulate a suitablemotor output in order to reproduce the estimated time interval.
Most studies on ERPs during non-spatial selection tasks had typicallyreported a prefrontal positivity also called as P2a; P3f, frontopolar component(FP), or frontal selection positivity (FSP) (Baas et al., 2002; Guillem et al.,2001; Jonkman et al., 2004; Kenemans et al., 1993; Makeig et al., 1999; Pottset al., 1996, 1998; Potts & Tucker, 2001), which are larger with task-relevantstimuli, and they have all been interpreted as indexing operations related to thetask-relevance of stimulus items and processes related to response production.In the other hand, a coexisting family of posterior negativities starting between150 and 200 ms, including N2b, N2pc, and “selection negativity” (SN) havebeen also described. These negative components are enhanced with the attendedstimuli and they are located over the superior parietal and inferior temporalcortical areas during visual tasks (Hopf et al., 2000; Potts et al., 1998; Tarkkaet al., 1995), probably reflecting the formation and categorization of perceptualrepresentations (Harter & Aine, 1984; Simson et al., 1977) even across sensory

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 129
modalities (Beer & Roder, 2005). Subsequently, both positive and negativeearly components are frequently followed by a negativity (N2b) between 250and 300 ms in adults (Potts, 2004).
It has been postulated that FSP, SN, and N2b could suggest the coexistenceof different modes of selection for the same ensemble of stimulus attributes(Smid et al., 1999). Thus it is not surprising that ERPs obtained from theestimation stage could have a similar morphology than those previouslydescribed. Nevertheless, the character of the present task let the subjectsjudge the time duration of each trial without performing a committed cognitiveprocessing of any of the physical characteristics of the stimuli.
Regarding the fact that a deficit to focus and sustained attention is themost characteristic clinical sign in ADHD, one could expect the appearance ofelectrophysiological differences between ADHD and control children whiledoing a task that required sustaining the attention to a visual stimulusrepetitively presented during a variable period of time. In addition, an absence ofthe FSP component in ADHD children has been described denoting that theirselective visual processing disturbances might be caused by a specific earlyfiltering deficit occurring around 200 ms (Jonkman et al., 2004). However, inthe present study, statistically significant differences between the ERPs fromthe two groups during the estimating time period were restricted to a recordingsite effect and its interaction with the analyzed time windows.
Behavioral results and the lack of additional differences between ERPs inthe first stage of the task seem to indicate that, at least in this case, differencesbetween the groups could depend more probably on the subsequent workingmemory storage, maintenance, and rehearsal of the estimated time period, oreven on the implementation and preservation of the mandatory motor output,than on an early attentional pure impairment. Recent findings revealed thatprospective duration reproduction efficiency could be related with coordinativeworking memory demands and working memory capacity (Dutke, 2005) mightsupport this hypothesis.
In the current control group of children, the most prominent ERPcomponents during time-reproducing were an early slow positivity peakingaround 320 ms, and a subsequent negative component with a bilateral maximumover 700 ms. They were both predominant at prefrontal locations and theyseemed to lateralize to the left side.
Because in the present study time reproduction was made through a motorresponse, one might expect that the initial prefrontal slow positivity (PSP) couldbe reflecting the motor response processing. Lateralization of the component tothe left seems to support this idea, particularly considering that the subjects were

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
130 A. A. GONZALEZ-GARRIDO ET AL.
instructed to respond with their right index finger. However, this component wasvirtually absent in ADHD children when they were emitting motor responseswhile its amplitude progressively decreased until it disappeared in the controlgroup, earlier than the end of the behavioral response. In addition, ERPscomponents related with motor responses usually have a more widespreaddistribution over the frontal areas, whereas the present slow positivity wasalmost restricted to the prefrontal sites. In this sense, time reproduction mayinvolve the activation and running of a memory-locked estimated time andit could be accomplished by consciously performing an alternative mentalprocess that could act as temporal marker (i.e., to count subvocally). Therefore,lateralization of the ERPs components to the left frontal brain region couldrepresent an activation of motor language area due to the selected cognitivestrategy. In this case, those who preferred this strategy could be more accurateand that might explain the ERPs differences between the groups, regardlessof the fact that children were instructed to avoid talking aloud while theyperformed the task.
It is generally accepted that selective attention affects visual processingeither at relatively early or late stages, depending on task demands. In this sense,functional neuroimaging studies have shown two main attentional systems, onebilateral dorsal attention system involved in top-down orienting of attentionand selective perception that seems to involve different posterior-ventral corticalnetworks, and a right-lateralized ventral attention system involved in reorientingattention in response to salient sensory stimuli (Fox et al., 2006; Posner &Petersen, 1990), which seems to regulate the selective (motoric) behavior onthe basis of combining external and internal stimuli (Posner, 1995). In thepresent context, the PSP component could also be interpreted as an indexof this regulation triggered by the internal activation of the memory content,indicating perhaps efficiency in the interaction between memory activation andmotor response. ERPs in the ADHD group failed to show this component, factthat seems to additionally support this idea.
Following this argument, subsequent prefrontal slow negativity in normalchildren might reflect the conclusion of the memory-guide motor activationprocess, or even the onset of an inhibitory mechanism devoted to discontinuethe programmed motor response. In this sense, ERPs from ADHD in this stageof the task seem to point out a probable timing distortion in neural substratesactivation processes, or less probably a discernible delay in their latencyonset.
In brief, the present results showed time estimation accuracy to bepoorer in ADHD children. In addition, differences were found on ERPs

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 131
that could be related more to the neural mechanisms responsible for internalmemory maintenance and rehearsal of the time duration period and its motoroutput, than to the sustained attention processes required to accomplishthe mental estimating task period. Distinctive behavioral results and frontalERPs waveforms during time reproduction in ADHD and controls appearsto be in line with Barkley’s model prediction of working memory deficitsin ADHD patients as a consequence of poor behavioral inhibition, probablyalso supporting Radonovich and Mostofsky’s explanation about the originof deficient utilization of temporal information in ADHD. Present resultsemphasize that ERPs represent an adequate tool to intensely explore thesecognitive processes in ADHD children to help to clarify the nature of theirdeficit.
REFERENCES
American Psychiatric Association. (2000). Diagnostic and statistical manual of mentaldisorders, 4th edition, Text revision. Washington, DC: American Psychiatric Press.
Baas, J. M., Kenemans, J. L., & Mangun, G. R. (2002). Selective attention to spatialfrequency: An ERP and source localization analysis. Clinical Neurophysiology,113(11), 1840–1854.
Barkley, R. (1997a). Attention-deficit/hyperactivity disorder, self regulation, and time:Toward a more comprehensive theory. Developmental and Behavioral Pediatrics,18, 271–279.
Barkley, R. (1997b). ADHD and the nature of self control. New York: Guilford.Barkley, R. (1997c). Behavioral inhibition, sustained attention, and executive functions:
Constructing a unifying theory of ADHD. Psychological Bulletin, 121, 65–94.Barkley, R., Koplowitz, S., Anderson, T., & McMurray, M. (1997). Sense of time in
children with ADHD: Effects of duration, distraction, and stimulant medication.Journal of the International Neuropsychological Society, 3, 359–369.
Barkley, R., Edwards, G., Laneri, M., Fletcher, K., & Metevia, L. (2001). Executivefunctioning, temporal discounting, and sense of time in adolescents with attentiondeficit hyperactivity disorder (ADHD) and oppositional defiant disorder (ODD).Journal of Abnormal Child Psychology, 29, 541–546.
Beer, A. L., & Roder, B. (2005). Attending to visual or auditory motion affects perceptionwithin and across modalities: An event-related potential study. European Journalof Neuroscience, 21(4), 1116–11130.
Boltz, M. (1991). Time estimation and attentional perspective. Perception & Psy-chophysics, 49, 422–433.
Brunia, C. H., & Damen, E. J. (1988). Distribution of slow brain potentials relatedto motor preparation and stimulus anticipation in a time estimation task.Electroencephalography and Clinical Neurophysiology, 69, 234–243.

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
132 A. A. GONZALEZ-GARRIDO ET AL.
Casini, L., & Ivry, R. B. (1999). Effects of divided attention on temporal processingin patients with lesions of the cerebellum or frontal lobe. Neuropsychology, 13,10–21.
Casini, L., & Macar, F. (1993). Behavioural and electrophysiological evidence for thespecific processing of temporal information. Psychologica Belgica, 33, 285–296.
Castellanos, F., & Tannock, R. (2002). Neuroscience of attention deficit/hyperactivitydisorder: The search for endophenotypes. National Review of Neuroscience, 3,617–628.
Chaston, A., & Kingstone, A. (2004). Time estimation: The effect of cortically mediatedattention. Brain and Cognition, 55, 286–289.
Church, R. M. (1984). Properties of the internal clock. In J. Gibbon & L. Allan (Eds.),Timing and time perception, Ann. N. Y. Acad. Sci., Vol. 423 (pp. 566–582). NewYork: New York, Academy of Sciences.
Dutke, S. (2005). Remembered duration: Working memory and the reproduction ofintervals. Perception & Psychophysics, 67(8), 1404–1413.
Elbert, T., Ulrich, R., Rockstroh, B., & Lutzenberger, W. (1991). The processing oftemporal intervals reflected by CNV-like brain potentials. Psychophysiology, 28,648–655.
Fox, M. D., Corbetta, M., Snyder, A. Z., Vincent, J. L., & Raichle, M. E. (2006).Spontaneous neuronal activity distinguishes human dorsal and ventral attentionsystems. Proceedings of the National Academy of Sciences of the United States ofAmerica, 103(26), 10046–10051.
Guillem, F., Bicu, M., & Debruille, J. B. (2001). Dissociating memory processesinvolved in direct and indirect tests with ERPs to unfamiliar faces. CognitiveBrain Research, 11(1), 113–125.
Harrington, D. L., Haaland, K. Y., & Knight, R. T. (1998). Cortical networksunderlying mechanisms of time perception. Journal of Neuroscience, 18, 1085–1095.
Harter, M. R., & Aine, C. J. (1984). Brain mechanisms of visual selective attention.In R. Parasuraman & D. R. Davies (Eds.), Varieties of attention. (pp. 293–321).London: Academic Press.
Hopf, J. M., Luck, S. J., Girelli, M., Hagner, T., Mangun, G. R., Scheich, H., & Heinze,H.-J. (2000). Neuronal sources of focused attention in visual search. CerebralCortex, 10(12), 1233–1241.
Johannes, S., Kube, C., Wieringa, B. M., Matzke, M., & Munte, T. F. (1997). Brainpotentials and time estimation in humans. Neuroscience Letters, 231, 63–66.
Jonkman, L. M., Kenemans, , J. L., Kemner, C., Verbaten, M. N., & van Engeland,H. (2004). Dipole source localization of event-related brain activity indicativeof an early visual selective attention deficit in ADHD children. ClinicalNeurophysiology, 115(7), 1537–1549.
Kenemans, J. L., Kok, A., & Smulders, F. T. (1993). Event-related potentials toconjunctions of spatial frequency and orientation as a function of stimulus

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 133
parameters and response requirements. Electroencephalography and ClinicalNeurophysiology, 88(1), 51–63.
Kerns, K., McInerney, R., & Wilde, N. (2001). Time reproduction, working memory,and behavioral inhibition in children with ADHD. Child Neuropsychology, 7, 21–31.
Kingstone, A., Enns, J., Mangun, G. R., & Gazzaniga, M. S. (1995). Guided visualsearch is a left hemisphere process in split-brain patients. Psychological Science,6, 118–121.
Kirk, R. E. (1995). Experimental design: Procedures for the behavioral sciences (3rd.ed.). Belmont: Brooks/Cole Publishing Company.
Klem, G. H., Luders, H. O., Jasper, H. H., & Elger, C. (1999). The ten-twenty electrodesystem of the International Federation. The International Federation of ClinicalNeurophysiology. Electroencephalography and Clinical Neurophysiology, 52,3–6.
Macar, F., & Besson, M. (1985). Contingent negative variation in processes ofexpectancy, motor preparation and time estimation. Biological Psychology, 21,293–307.
Makeig, S., Westerfield, M., Jung, T. P., Covington, J., Townsend, J., Sejnowski, T. J., &Courchesne, E. (1999). Functionally independent components of the late positiveevent-related potential during visual spatial attention. Journal of Neuroscience,19(7), 2665–2680.
Marmaras, N., Vassilakis, P., & Dounias, G. (1995). Factors affecting accuracy ofproducing time intervals. Perceptual and Motor Skills, 80, 1043–1056.
Martinussen, R., Hayden, J., Hogg-Johnson, S., & Tannock, R. (2005). A meta-analysisof working memory impairments in children with attention-deficit/hyperactivitydisorder. Journal of the American Academy of Child and Adolescent Psychiatry,44(4), 377–384.
Meaux, J. B., & Chelonis, J. J. (2003). Time perception differences in children with andwithout ADHD. Journal of Pediatric Health Care, 17(2), 64–71.
Pastor, M. A., & Artieda, J. (1996). Time, internal clocks and movement. Amsterdam,North-Holland: Elsevier.
Posner, M. I. (1995). Attention in cognitive neuroscience: An overview. In M. S.Gazzaniga (Ed.), The cognitive neurosciences. (pp. 615–624). Cambridge, MA:MIT Press.
Posner, M. I., & Petersen, S. E. (1990). The attention system of the human brain. AnnualReview of Neuroscience, 13, 25–42.
Potts, G. F. (2004). An ERP index of task relevance evaluation of visual stimuli. Brainand Cognition, 56(1), 5–13.
Potts, G. F., Hirayasu, Y., O’Donnell, B. F., Shenton, M. E., & McCarley, R. W. (1998).High-density recording and topographic analysis of the auditory oddball event-related potential in patients with schizophrenia. Biological Psychiatry, 44(10),982–989.

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
134 A. A. GONZALEZ-GARRIDO ET AL.
Potts, G. F., Liotti, M., Tucker, D. M., & Posner, M. I. (1996). Frontal and inferiortemporal cortical activity in visual target detection: Evidence from high spatiallysampled event-related potentials. Brain Topography, 9(1), 3–14.
Potts, G. F., & Tucker, D. M. (2001). Frontal evaluation and posterior representation intarget detection. Cognitive Brain Research, 11(1), 147–156.
Radonovich, K. J., & Mostofsky, S. H. (2004). Duration judgments in children withADHD suggest deficient utilization of temporal information rather than generalimpairment in timing. Neuropsychology, Development, and Cognition. Section C,Child Neuropsychology, 10(3), 162–172.
Rubia, K., Noorloos, J., Smith, A., Gunning, B., & Sergeant, J. (2003). Motor timingdeficits in community and clinical boys with hyperactive behavior: The effect ofmethylphenidate on motor timing. Journal of Abnormal Child Psychology, 31(3),301–313.
Rubia, K., Taylor, A., Taylor, E., & Sergeant, J. A. (1999). Synchronization, anticipation,and consistency in motor timing of children with dimensionally defined attentiondeficit hyperactivity behaviour. Perceptual and Motor Skills, 89(3), 1237–1258.
Sawyer, T. F., Meyers, P. J., & Huser, S. J. (1994). Contrasting task demands alterthe perceived duration of brief time intervals. Perception & Psychophysics, 56,649–657.
Shallice, T., Marzocchi, G. M., Coser, S., Del Savio, M., Meuter, R. F., & Rumiati, R. I.(2002). Executive function profile of children with attention deficit hyperactivitydisorder. Developmental Neuropsychology, 21(1), 43–71.
Simson, R., Vaughan, H. G., & Ritter, W. (1977). The scalp topography of potentialsin auditory and visual discrimination tasks. Electroencephalography and ClinicalNeurophysiology, 42, 528–535.
Smid, H. G., Jakob, A., & Heinze, H. J. (1999). An event-related brain potential studyof visual selective attention to conjunctions of color and shape. Psychophysiology,36(2), 264–279.
Smith, A., Taylor, E., Rogers, J., Newman, S., & Rubia, K. (2002). Evidence for a puretime perception deficit in children with ADHD. Journal of Child Psychology andPsychiatry, 43, 529–542.
Tarkka, I. M., Stokic, D. S., Basile, L. F., & Papanicolaou, A. C. (1995). Electricsource localization of the auditory P300 agrees with magnetic source localization.Electroencephalography and Clinical Neurophysiology, 96(6), 538–545.
Toplak, M. E., & Tannock, R. (2005). Time perception: Modality and duration effectsin attention-deficit/hyperactivity disorder (ADHD). Journal of Abnormal ChildPsychology, 33(5), 639–654.
Treisman, M. (1984). Temporal rhythms and cerebral rhythms. In J. Gibbon & I. Allan(Eds.), Timing and time perception, Ann. N. Y. Acad. Sci., Vol. 423 (pp. 542–565).New York: New York Academy of Sciences.
Treisman, M., Faulkner, A., & Naish, P. L. (1992). On the relation between timeperception and the timing of motor action: Evidence for a temporal oscillator

Dow
nloa
ded
By:
[Uni
vers
idad
de
Sev
illa]
At:
04:4
7 30
Nov
embe
r 200
7
TIME REPRODUCTION IN ADHD CHILDREN 135
controlling the timing of movement. The Quarterly Journal of ExperimentalPsychology, 45A, 235–263.
Xu, D., Liu, , T., Ashe, J., & Bushara, K. O. (2006). Role of the olivo-cerebellar systemin timing. The Journal of Neuroscience, 26(22), 5990–5995.
Zakay, D., Tsal, Y., Moses, M., & Shahar, I. (1994). The role of segmentation inprospective and retrospective time estimation processes. Memory & Cognition,22, 344–351.

This article was downloaded by:[Universidad de Sevilla]On: 30 November 2007Access Details: [subscription number 773444423]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
International Journal of NeurosciencePublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713644851
TIME REPRODUCTION DISTURBANCES IN ADHDCHILDREN: AN ERP STUDYA. A. González-Garrido ab; F. R. Gómez-Velázquez a; D. Zarabozo a; R.López-Elizalde b; A. Ontiveros b; H. Madera-Carrillo a; O. L. Vega a; J.L. OropezaDe Alba b; J.M. De La Serna Tuya aca Instituto de Neurociencias, Universidad de Guadalajara, Guadalajara, Méxicob O.P.D. Hospital Civil de Guadalajara, Guadalajara, Méxicoc Universidad de Sevilla, Seville, Spain
Online Publication Date: 01 January 2008To cite this Article: González-Garrido, A. A., Gómez-Velázquez, F. R., Zarabozo,D., López-Elizalde, R., Ontiveros, A., Madera-Carrillo, H., Vega, O. L., De Alba,J.L. Oropeza and Tuya, J.M. De La Serna (2008) 'TIME REPRODUCTION
DISTURBANCES IN ADHD CHILDREN: AN ERP STUDY', International Journal of Neuroscience, 118:1, 119 - 135To link to this article: DOI: 10.1080/00207450601042177URL: http://dx.doi.org/10.1080/00207450601042177
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.




















