
Rapid Increase of Interleukin-10 Plasma Levels After Combined Auxiliary Liver-Kidney Transplantation...
8
Rapid Increase of Interleukin-10 Plasma Levels After Combined Auxiliary Liver-Kidney Transplantation in Presensitized Patients Madeleine Ingelsten, 1 Karin Gustafsson, 2 Michael Olausson, 3 Bo ¨rje Haraldsson, 4 Alex Karlsson-Parra, 5 and Jenny Nystro ¨m 6,7 Background. After transplantation, donor dendritic cells (DCs) in the grafted organ are activated by an ischemia/ reperfusionYinduced inflammatory process that induces their migration to the recipient’s secondary lymphoid tissues. The subsequent interaction between migrated and mature donor DCs, recipient T cells, and natural killer (NK) cells is proposed to be crucial in directing host immune reactions toward allograft rejection. A liver transplant is less prone to induce rejection compared with most other solid organ transplants, and simultaneous transplantation of liver and kidney is known to improve the clinical outcome of kidney transplantation. Methods and Results. Here we show that liver as well as combined auxiliary liver-kidney transplantation in patients induces a rapid increase in plasma interleukin-10 (IL-10) to levels that are significantly higher than those seen after standard kidney transplantation. Addition of IL-10 during in vitro maturation of human monocyteYderived DCs with ischemia/reperfusion-associated factors was found to affect phenotypic DC maturation significantly. Addition of IL-10 inhibited DC production of the NK cell- and T cell-recruiting chemokines CXCL9, CXCL10 and CXCL11. Conclusion. Our findings indicate that liver transplantation induces a substantial systemic release of IL-10, which may inhibit T cell- and NK cell-mediated rejection processes toward the transplanted liver and concurrently transplanted kidney. Keywords: Dendritic cells, Allograft rejection, Tolerance, Chemokine, Interleukin-10. (Transplantation 2014;00: 00Y00) A fter transplantation, an inflammatory process caused by ischemia/reperfusion activate donor dendritic cells (DCs), inducing them to migrate to the recipient’s second- ary lymphoid tissues. The subsequent interaction between migrated donor DCs and recipient T cells that takes place in the lymphoid organs during the early posttransplantation period is proposed to be crucial in directing host immune reactions toward allograft rejection or acceptance (1). It is known that a transplanted liver is less prone to in- duce rejection than a renal graft (2). Several studies have shown that patients simultaneously transplanted with a liver and a kidney have a lower incidence of rejection than expected after regular kidney transplantation (3Y5). In a recent study by Leca et al., sensitized patients did not have a significantly re- duced survival after combined liver kidney transplantation compared with unsensitized patients (6). The tolerogenic and protective effect induced by the liver in combined liver kidney transplantation has made it possible to transplant highly sensitized kidney recipients using a combined auxiliary liver- kidney transplantation procedure (7). This procedure, in which a partial liver graft is transplanted shortly before a renal transplant from the same donor, has been highly successful in This study was supported by the Swedish Research Council 14764, the IngaBritt and Arne Lundberg Research Foundation, the Professor Lars- Erik Gelin Memorial Foundation, the John and Brit Wennerstro ¨m Re- search Foundation, the Royal Society of Arts and Sciences in Go ¨teborg, and the Sahlgrenska University Hospital Grant LUA-ALF. The authors declare no conflicts of interest. 1 Department of Infectious Medicine, Sahlgrenska Academy, Gothenburg University, Gothenburg, Sweden. 2 Department of Immunology, Sahlgrenska Academy, Gothenburg Uni- versity, Gothenburg, Sweden. 3 Department of Surgery, Sahlgrenska Academy, Gothenburg University, Gothenburg, Sweden. 4 Department of Clinical and Molecular Medicine (Nephrology), Sahlgrenska Academy, Gothenburg University, Gothenburg, Sweden. 5 Department of Immunology, Genetics, and Pathology, Uppsala Univer- sity, Uppsala, Sweden. CLINICAL AND TRANSLATIONAL RESEARCH Transplantation & Volume 00, Number 00, Month 00, 2014 www.transplantjournal.com 1 6 Department of Physiology, Sahlgrenska Academy, Gothenburg University, Gothenburg, Sweden. 7 Address correspondence to: Jenny Nystro ¨m, Ph.D., Gothenburg Univer- sity, Institute of Neuroscience and Physiology, Renal Physiology, P. O. Box 432, 40530 Gothenburg, Sweden. E-mail: [email protected] M.I. participated in research design, writing of the paper, performance of the research, and data analysis. K.G. participated in research design, writing of the paper, performance of the research, and data analysis. M.O., B.H., A.K.-P., and J.N. participated in research design, data analysis, and writing of the paper. Received 20 May 2013. Revision requested 12 June 2013. Accepted 23 December 2013. Copyright * 2014 by Lippincott Williams & Wilkins ISSN: 0041-1337/14/0000-00 DOI: 10.1097/TP.0000000000000038 Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
Transcript of Rapid Increase of Interleukin-10 Plasma Levels After Combined Auxiliary Liver-Kidney Transplantation...

Rapid Increase of Interleukin-10 Plasma Levels AfterCombined Auxiliary Liver-Kidney Transplantation in
Presensitized Patients
Madeleine Ingelsten,1 Karin Gustafsson,2 Michael Olausson,3 Borje Haraldsson,4 Alex Karlsson-Parra,5
and Jenny Nystrom6,7
Background. After transplantation, donor dendritic cells (DCs) in the grafted organ are activated by an ischemia/reperfusionYinduced inflammatory process that induces their migration to the recipient’s secondary lymphoid tissues.The subsequent interaction between migrated and mature donor DCs, recipient T cells, and natural killer (NK) cells isproposed to be crucial in directing host immune reactions toward allograft rejection. A liver transplant is less prone toinduce rejection compared with most other solid organ transplants, and simultaneous transplantation of liver andkidney is known to improve the clinical outcome of kidney transplantation.Methods and Results. Here we show that liver as well as combined auxiliary liver-kidney transplantation in patientsinduces a rapid increase in plasma interleukin-10 (IL-10) to levels that are significantly higher than those seen afterstandard kidney transplantation. Addition of IL-10 during in vitro maturation of human monocyteYderived DCs withischemia/reperfusion-associated factors was found to affect phenotypic DC maturation significantly. Addition of IL-10inhibited DC production of the NK cell- and T cell-recruiting chemokines CXCL9, CXCL10 and CXCL11.Conclusion. Our findings indicate that liver transplantation induces a substantial systemic release of IL-10, whichmay inhibit T cell- and NK cell-mediated rejection processes toward the transplanted liver and concurrentlytransplanted kidney.
Keywords: Dendritic cells, Allograft rejection, Tolerance, Chemokine, Interleukin-10.
(Transplantation 2014;00: 00Y00)
A fter transplantation, an inflammatory process caused byischemia/reperfusion activate donor dendritic cells
(DCs), inducing them to migrate to the recipient’s second-ary lymphoid tissues. The subsequent interaction betweenmigrated donor DCs and recipient T cells that takes place inthe lymphoid organs during the early posttransplantationperiod is proposed to be crucial in directing host immunereactions toward allograft rejection or acceptance (1).
It is known that a transplanted liver is less prone to in-duce rejection than a renal graft (2). Several studies haveshown that patients simultaneously transplanted with a liver
and a kidney have a lower incidence of rejection than expectedafter regular kidney transplantation (3Y5). In a recent study byLeca et al., sensitized patients did not have a significantly re-duced survival after combined liver kidney transplantationcompared with unsensitized patients (6). The tolerogenic andprotective effect induced by the liver in combined liver kidneytransplantation has made it possible to transplant highlysensitized kidney recipients using a combined auxiliary liver-kidney transplantation procedure (7). This procedure, inwhich a partial liver graft is transplanted shortly before a renaltransplant from the same donor, has been highly successful in
This study was supported by the Swedish Research Council 14764, theIngaBritt and Arne Lundberg Research Foundation, the Professor Lars-Erik Gelin Memorial Foundation, the John and Brit Wennerstrom Re-search Foundation, the Royal Society of Arts and Sciences in Goteborg,and the Sahlgrenska University Hospital Grant LUA-ALF.
The authors declare no conflicts of interest.1 Department of Infectious Medicine, Sahlgrenska Academy, Gothenburg
University, Gothenburg, Sweden.2 Department of Immunology, Sahlgrenska Academy, Gothenburg Uni-
versity, Gothenburg, Sweden.3 Department of Surgery, Sahlgrenska Academy, Gothenburg University,
Gothenburg, Sweden.4 Department of Clinical and Molecular Medicine (Nephrology), Sahlgrenska
Academy, Gothenburg University, Gothenburg, Sweden.5 Department of Immunology, Genetics, and Pathology, Uppsala Univer-
sity, Uppsala, Sweden.
CLINICAL AND TRANSLATIONAL RESEARCH
Transplantation & Volume 00, Number 00, Month 00, 2014 www.transplantjournal.com 1
6 Department of Physiology, Sahlgrenska Academy, Gothenburg University,Gothenburg, Sweden.
7 Address correspondence to: Jenny Nystrom, Ph.D., Gothenburg Univer-sity, Institute of Neuroscience and Physiology, Renal Physiology, P. O.Box 432, 40530 Gothenburg, Sweden.
E-mail: [email protected]. participated in research design, writing of the paper, performance of the
research, and data analysis. K.G. participated in research design, writingof the paper, performance of the research, and data analysis. M.O., B.H.,A.K.-P., and J.N. participated in research design, data analysis, andwriting of the paper.
Received 20 May 2013. Revision requested 12 June 2013.Accepted 23 December 2013.Copyright * 2014 by Lippincott Williams & WilkinsISSN: 0041-1337/14/0000-00DOI: 10.1097/TP.0000000000000038
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
over 20 patients despite positive cross-match (7). The mech-anism behind the reduced risk for rejection is not knownbut tolerogenic processes in the liver are most likely involved
(8, 9). By comparing transcriptional changes in human liverand kidney grafts after combined auxiliary liver-kidneytransplantation, we recently demonstrated that transcrip-tional upregulation of potential DC-maturating factors, in-cluding interferon (IFN) F, tumor necrosis factor (TNF) >,and interleukin (IL) 1A, was similar in both organs (9). Here,we hypothesized that the immunosuppressive factor IL-10may explain the beneficial effect of simultaneous liver trans-plants on kidney transplantation. As previously shown by LeMoine et al. in orthopic liver transplantation, there is a rapidincrease in IL-10 release after allograft reperfusion, and theyshow that there is more IL-10 in right atrial vein blood than inportal blood, indicating that the release is originating from theliver (8). Here, we focused on the systemic release of IL-10 inpatients undergoing liver, kidney and auxiliary liver-kidneytransplantation. We also investigated the NK and T cell-recruiting ability and immune stimulatory function ofmonocyte-derived DCs activated with a combination of theischemia/reperfusion-associated inflammatory mediatorsIFN-F, TNF->, and IL-1A in the presence of IL-10.
RESULTS
IL-10 Is Released After Liver TransplantationThe release of cytokines into serum was measured
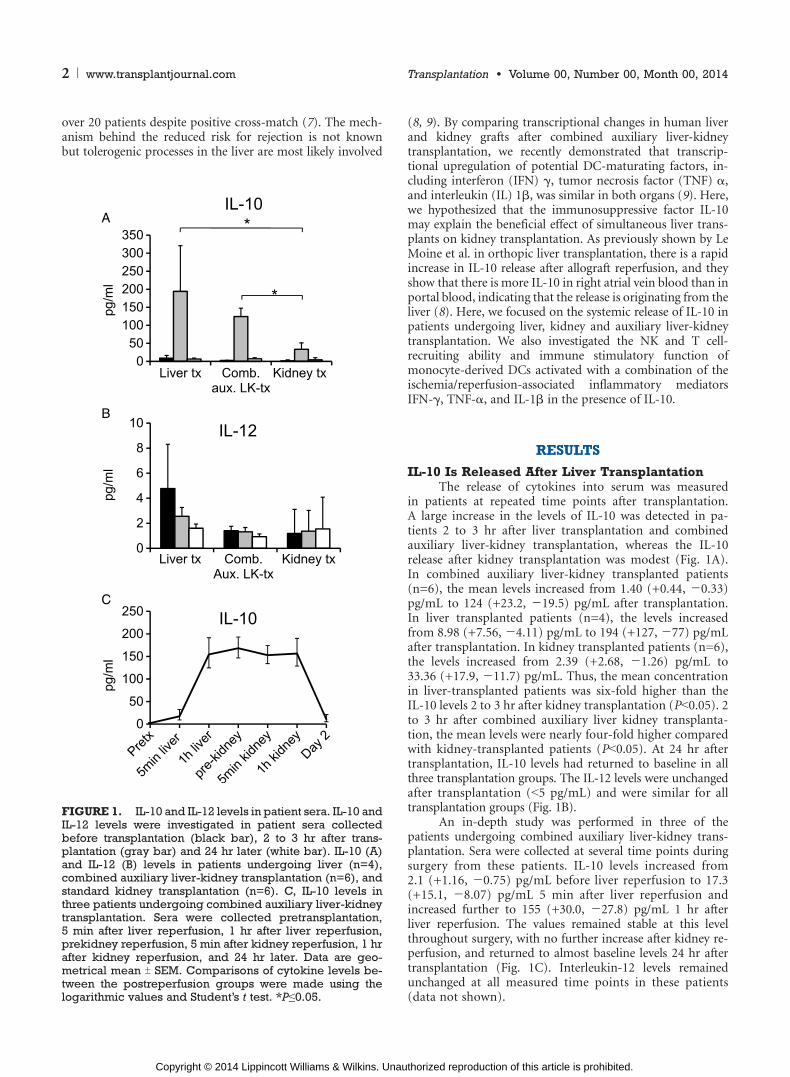
in patients at repeated time points after transplantation.A large increase in the levels of IL-10 was detected in pa-tients 2 to 3 hr after liver transplantation and combinedauxiliary liver-kidney transplantation, whereas the IL-10release after kidney transplantation was modest (Fig. 1A).In combined auxiliary liver-kidney transplanted patients(n=6), the mean levels increased from 1.40 (+0.44, j0.33)pg/mL to 124 (+23.2, j19.5) pg/mL after transplantation.In liver transplanted patients (n=4), the levels increasedfrom 8.98 (+7.56, j4.11) pg/mL to 194 (+127, j77) pg/mLafter transplantation. In kidney transplanted patients (n=6),the levels increased from 2.39 (+2.68, j1.26) pg/mL to33.36 (+17.9, j11.7) pg/mL. Thus, the mean concentrationin liver-transplanted patients was six-fold higher than theIL-10 levels 2 to 3 hr after kidney transplantation (PG0.05). 2to 3 hr after combined auxiliary liver kidney transplanta-tion, the mean levels were nearly four-fold higher comparedwith kidney-transplanted patients (PG0.05). At 24 hr aftertransplantation, IL-10 levels had returned to baseline in allthree transplantation groups. The IL-12 levels were unchangedafter transplantation (G5 pg/mL) and were similar for alltransplantation groups (Fig. 1B).
An in-depth study was performed in three of thepatients undergoing combined auxiliary liver-kidney trans-plantation. Sera were collected at several time points duringsurgery from these patients. IL-10 levels increased from2.1 (+1.16, j0.75) pg/mL before liver reperfusion to 17.3(+15.1, j8.07) pg/mL 5 min after liver reperfusion andincreased further to 155 (+30.0, j27.8) pg/mL 1 hr afterliver reperfusion. The values remained stable at this levelthroughout surgery, with no further increase after kidney re-perfusion, and returned to almost baseline levels 24 hr aftertransplantation (Fig. 1C). Interleukin-12 levels remainedunchanged at all measured time points in these patients(data not shown).
FIGURE 1. IL-10 and IL-12 levels in patient sera. IL-10 andIL-12 levels were investigated in patient sera collectedbefore transplantation (black bar), 2 to 3 hr after trans-plantation (gray bar) and 24 hr later (white bar). IL-10 (A)and IL-12 (B) levels in patients undergoing liver (n=4),combined auxiliary liver-kidney transplantation (n=6), andstandard kidney transplantation (n=6). C, IL-10 levels inthree patients undergoing combined auxiliary liver-kidneytransplantation. Sera were collected pretransplantation,5 min after liver reperfusion, 1 hr after liver reperfusion,prekidney reperfusion, 5 min after kidney reperfusion, 1 hrafter kidney reperfusion, and 24 hr later. Data are geo-metrical mean T SEM. Comparisons of cytokine levels be-tween the postreperfusion groups were made using thelogarithmic values and Student’s t test. *Pe0.05.
2 www.transplantjournal.com Transplantation & Volume 00, Number 00, Month 00, 2014
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

Release of Proinflammatory Cytokines AfterLiver Transplantation
The release of the proinflammatory cytokines IL-1A,IL-2, and IL-8 also differed after liver and combined auxil-iary liver-kidney transplantation compared with kidneytransplantation. IL-1A (PG0.05), IL-2 (PG0.001), and IL-8(PG0.001) increased 2 to 3 hr after liver reperfusion bothin liver and combined auxiliary liver-kidney patients but notin patients that only underwent a kidney transplantation.Moreover, 24 hr after liver or combined auxiliary liver-kidney transplantation, the levels of IL-1A had returned topretransplantation levels, whereas IL-2 (PG0.001) and IL-8(PG0.001) levels remained significantly higher in liver andcombined auxiliary liver-kidney transplanted patients com-pared with kidney transplanted patients (Fig. 2).
DC Phenotype Is Affected by IL-10To investigate how the maturation of passenger DCs
could be affected by the high levels of IL-10 in the graft,
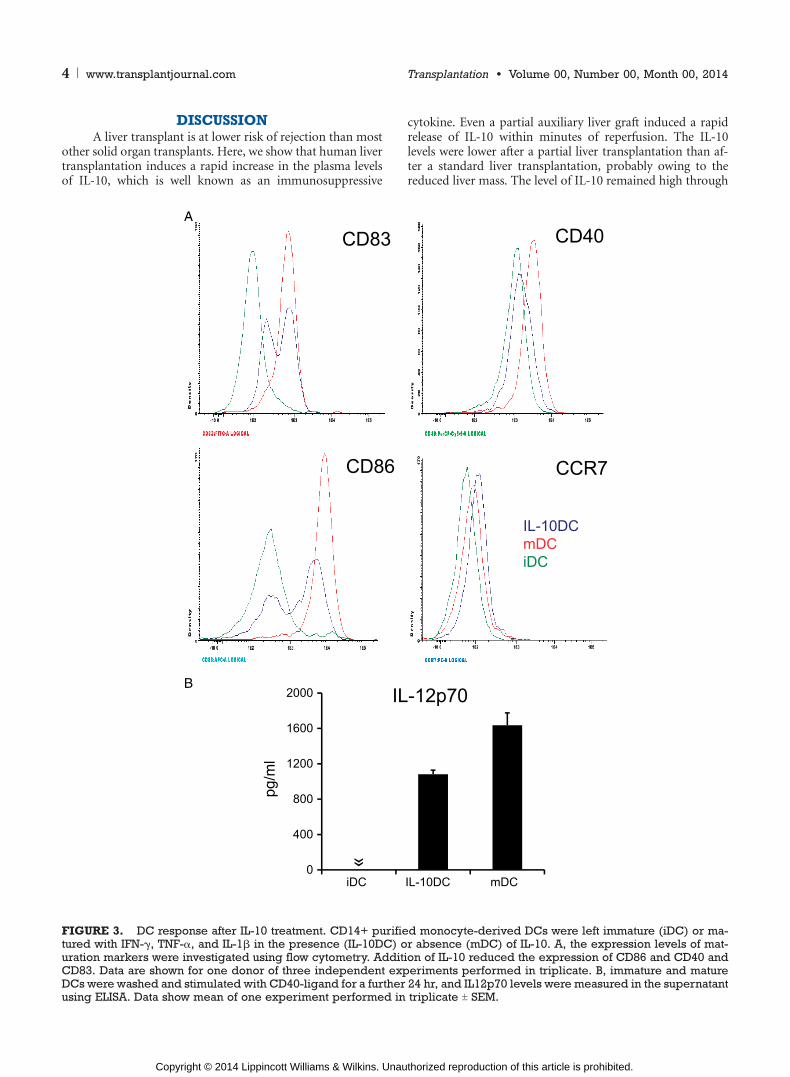
monocyte-derived DCs were studied in vitro. Immature DCswere incubated in the presence or absence of IL-10 andthereafter activated with a maturation cocktail consisting ofIFN-F, TNF->, and IL-1A. These cytokines were chosen as wepreviously showed that they are upregulated in liver and kid-ney grafts shortly after combined auxiliary liver-kidney trans-plantation (9). IL-10 treatment resulted in reduced expressionof the characteristic DC maturation marker CD83 and co-stimulatory molecules CD86 and CD40 (Fig. 3A). The ex-pression of CCR7, which indicates the ability of DCs to migratetowards lymphoid organs, was unaffected by IL-10 (Fig. 3A).
In addition, IL-12 production was reduced after CD40-stimulation of DCs matured in the presence of IL-10 com-pared with DCs incubated without IL-10 (Fig. 3B).
Production of CXCL9, CXCL10, and CXCL11 byDCs Is Repressed by IL-10
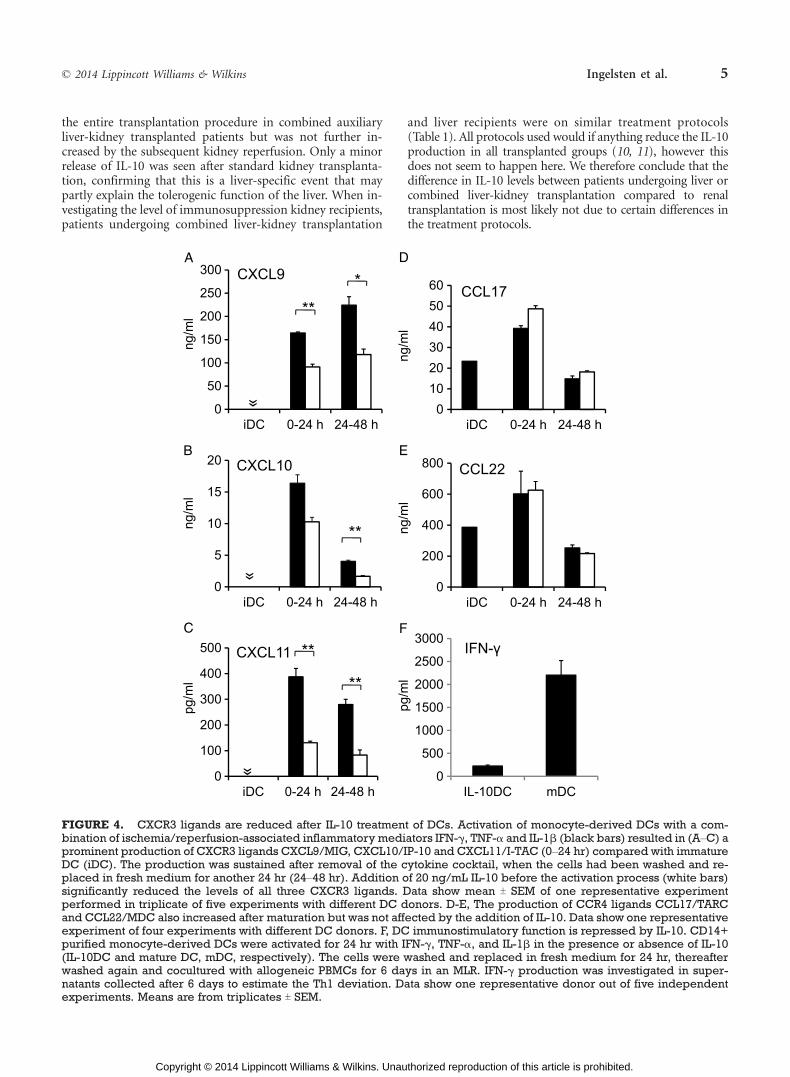
Dendritic cells activated with IFN-F, TNF->, and IL-1Afor 24 hr produced high levels of the NK and T cell-recruitingCXCR3 ligands CXCL9/MIG, CXCL10/IP-10 and CXCL11/I-TAC in the supernatant. Addition of IL-10 (20 ng/mL) beforeaddition of the cytokine cocktail reduced the production ofCXCL9/MIG by 33% (T4.3, PG0.01), CXLC10/IP-10 by 23%(T10.5, n.s.), and CXCL11/I-TAC by 62% (T9.2, PG0.01)(Fig. 4AYC).
The ability of DCs to produce chemokines was also in-vestigated after the removal of IL-10. In this setup, DCs werematured for 24 hr with IL-10, washed twice and replaced infresh medium. 24 hr later, the production of CXCR3-ligandswas still reduced in IL-10 treated cells compared with ma-tured DCs (CXCL9 reduced with 30% T 9, PG0.05, CXCL1044% T10, PG0.01, CXCL11 73% T7, PG0.01) (Fig. 4AYC). Theinhibitory effect of IL-10 was concentration dependent, andconcentrations higher than 20 ng/mL did not reduce theCXCR3-ligand production any further (data not shown).This was true after both 24 and 48 hr for all investigatedchemokines.
Finally, we showed that addition of IL-10 to DCs be-fore activation with the cytokine cocktail did not affect theproduction of the CCR4ligands CCL17/TARC and CCL22/MDC, chemokines associated with recruitment of Th2 andregulatory T cells in DCs (Fig. 4DYE).
Ability of DCs to Stimulate an Immune ResponseIs Reduced by IL-10
The effect of IL-10 on the ability of passenger DCs tostimulate an allogeneic immune response was investigatedusing a mixed leukocyte reaction (MLR) model. CD14+monocyte-derived DCs were matured by IFN-F, TNF-> andIL-1A in the presence or absence of IL-10 for 24 hr. The cellswere washed and incubated for another 24 hr without cy-tokines to simulate a phase when the passenger DCs leavethe immune-activated graft and migrate to the lymphoidorgans. Thereafter, the cells were washed again and alloge-neic peripheral blood mononuclear cells (PBMCs) wereadded to the plate. IFN-F in supernatants collected after6 days was used to study the activation profile of alloreactiveT cells. In five independent experiments, IL-10-treated cellswere without exception less capable of activating IFN-Fproduction from T cells (10-59% of the response observedin the absence of IL-10) (one representative experiment isshown in Fig. 4F), PG0.01, n=5.
FIGURE 2. Cytokine levels in patient sera. IL-1A, IL-2,and IL-8 levels were investigated in patient sera collectedbefore transplantation (black bar), 2 to 3 hr after trans-plantation (gray bar) and 24 hr later (white bar). Data aregeometrical mean T SEM.
* 2014 Lippincott Williams & Wilkins Ingelsten et al. 3
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
DISCUSSIONA liver transplant is at lower risk of rejection than most
other solid organ transplants. Here, we show that human livertransplantation induces a rapid increase in the plasma levelsof IL-10, which is well known as an immunosuppressive
cytokine. Even a partial auxiliary liver graft induced a rapidrelease of IL-10 within minutes of reperfusion. The IL-10levels were lower after a partial liver transplantation than af-ter a standard liver transplantation, probably owing to thereduced liver mass. The level of IL-10 remained high through
FIGURE 3. DC response after IL-10 treatment. CD14+ purified monocyte-derived DCs were left immature (iDC) or ma-tured with IFN-F, TNF->, and IL-1A in the presence (IL-10DC) or absence (mDC) of IL-10. A, the expression levels of mat-uration markers were investigated using flow cytometry. Addition of IL-10 reduced the expression of CD86 and CD40 andCD83. Data are shown for one donor of three independent experiments performed in triplicate. B, immature and matureDCs were washed and stimulated with CD40-ligand for a further 24 hr, and IL12p70 levels were measured in the supernatantusing ELISA. Data show mean of one experiment performed in triplicate T SEM.
4 www.transplantjournal.com Transplantation & Volume 00, Number 00, Month 00, 2014
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
the entire transplantation procedure in combined auxiliaryliver-kidney transplanted patients but was not further in-creased by the subsequent kidney reperfusion. Only a minorrelease of IL-10 was seen after standard kidney transplanta-tion, confirming that this is a liver-specific event that maypartly explain the tolerogenic function of the liver. When in-vestigating the level of immunosuppression kidney recipients,patients undergoing combined liver-kidney transplantation
and liver recipients were on similar treatment protocols(Table 1). All protocols used would if anything reduce the IL-10production in all transplanted groups (10, 11), however thisdoes not seem to happen here. We therefore conclude that thedifference in IL-10 levels between patients undergoing liver orcombined liver-kidney transplantation compared to renaltransplantation is most likely not due to certain differences inthe treatment protocols.
FIGURE 4. CXCR3 ligands are reduced after IL-10 treatment of DCs. Activation of monocyte-derived DCs with a com-bination of ischemia/reperfusion-associated inflammatory mediators IFN-F, TNF-> and IL-1A (black bars) resulted in (AYC) aprominent production of CXCR3 ligands CXCL9/MIG, CXCL10/IP-10 and CXCL11/I-TAC (0Y24 hr) compared with immatureDC (iDC). The production was sustained after removal of the cytokine cocktail, when the cells had been washed and re-placed in fresh medium for another 24 hr (24Y48 hr). Addition of 20 ng/mL IL-10 before the activation process (white bars)significantly reduced the levels of all three CXCR3 ligands. Data show mean T SEM of one representative experimentperformed in triplicate of five experiments with different DC donors. D-E, The production of CCR4 ligands CCL17/TARCand CCL22/MDC also increased after maturation but was not affected by the addition of IL-10. Data show one representativeexperiment of four experiments with different DC donors. F, DC immunostimulatory function is repressed by IL-10. CD14+purified monocyte-derived DCs were activated for 24 hr with IFN-F, TNF->, and IL-1A in the presence or absence of IL-10(IL-10DC and mature DC, mDC, respectively). The cells were washed and replaced in fresh medium for 24 hr, thereafterwashed again and cocultured with allogeneic PBMCs for 6 days in an MLR. IFN-F production was investigated in super-natants collected after 6 days to estimate the Th1 deviation. Data show one representative donor out of five independentexperiments. Means are from triplicates T SEM.
* 2014 Lippincott Williams & Wilkins Ingelsten et al. 5
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
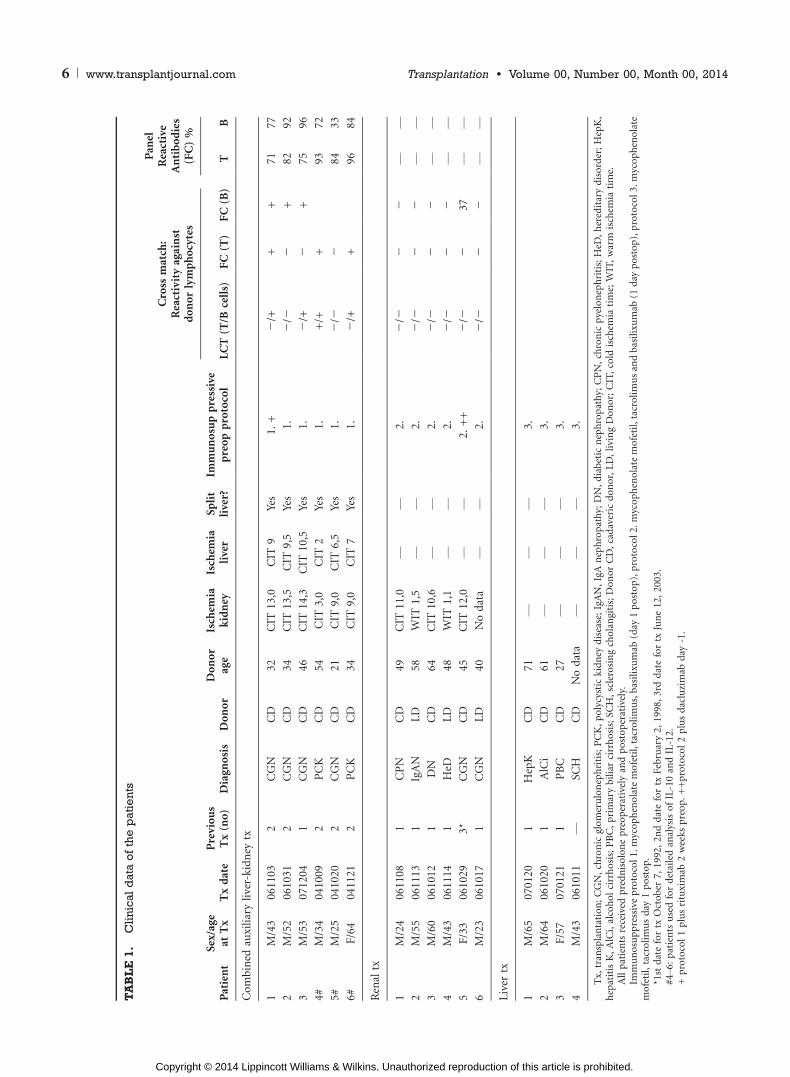
TABLE1.
Clinical
dataofthepatients
Cro
ssm
atch
:R
eact
ivit
yag
ain
std
on
or
lym
ph
ocy
tes
Pan
elR
eact
ive
An
tib
od
ies
(FC
)%
Pat
ien
tS
ex/a
ge
atT
xT
xd
ate
Pre
vio
us
Tx
(no
)D
iag
no
sis
Do
no
rD
on
or
age
Isch
emia
kid
ney
Isch
emia
live
rS
pli
tli
ver?
Imm
un
osu
pp
ress
ive
pre
op
pro
toco
lL
CT
(T/B
cell
s)F
C(T
)F
C(B
)T
B
Co
mb
ined
auxi
liar
yli
ver-
kid
ney
tx
1M
/43
06
11
03
2C
GN
CD
32
CIT
13
,0C
IT9
Yes
1.
+j
/++
+7
17
7
2M
/52
06
10
31
2C
GN
CD
34
CIT
13
,5C
IT9
,5Y
es1
.j
/jj
+8
29
2
3M
/53
07
12
04
1C
GN
CD
46
CIT
14
,3C
IT1
0,5
Yes
1.
j/+
j+
75
96
4#
M/3
40
41
00
92
PC
KC
D5
4C
IT3
,0C
IT2
Yes
1.
+/+
+9
37
2
5#
M/2
50
41
02
02
CG
NC
D2
1C
IT9
,0C
IT6
,5Y
es1
.j
/jj
84
33
6#
F/6
40
41
12
12
PC
KC
D3
4C
IT9
,0C
IT7
Yes
1.
j/+
+9
68
4
Ren
altx
1M
/24
06
11
08
1C
PN
CD
49
CIT
11
,0V
V2
.j
/jj
jV
V
2M
/55
06
11
13
1Ig
AN
LD
58
WIT
1,5
VV
2.
j/j
jj
VV
3M
/60
06
10
12
1D
NC
D6
4C
IT1
0,6
VV
2.
j/j
jj
VV
4M
/43
06
11
14
1H
eDL
D4
8W
IT1
,1V
V2
.j
/jj
jV
V
5F
/33
06
10
29
3*
CG
NC
D4
5C
IT1
2,0
VV
2.
++
j/j
j3
7V
V
6M
/23
06
10
17
1C
GN
LD
40
No
dat
aV
V2
.j
/jj
jV
V
Liv
ertx
1M
/65
07
01
20
1H
epK
CD
71
VV
V3
.
2M
/64
06
10
20
1A
lCi
CD
61
VV
V3
.
3F
/57
07
01
21
1P
BC
CD
27
VV
V3
.
4M
/43
06
10
11
VS
CH
CD
No
dat
aV
VV
3.
Tx,
tran
spla
nta
tio
n;
CG
N,
chro
nic
glo
mer
ulo
nep
hri
tis;
PC
K,
po
lycy
stic
kid
ney
dis
ease
;Ig
AN
,Ig
An
eph
rop
ath
y;D
N,
dia
bet
icn
eph
rop
ath
y;C
PN
,ch
ron
icp
yelo
nep
hri
tis;
HeD
,h
ered
itar
yd
iso
rder
;H
epK
,h
epat
itis
K,
AlC
i,al
coh
ol
cirr
ho
sis;
PB
C,
pri
mar
yb
ilia
rci
rrh
osi
s;S
CH
,sc
lero
sin
gch
ola
ngi
tis;
Do
no
rC
D,
cad
aver
icd
on
or,
LD
,li
vin
gD
on
or;
CIT
,co
ldis
chem
iati
me;
WIT
,w
arm
isch
emia
tim
e.A
llp
atie
nts
rece
ived
pre
dn
iso
lon
ep
reo
per
ativ
ely
and
po
sto
per
ativ
ely.
Imm
un
osu
pp
ress
ive
pro
toco
l1
.m
yco
ph
eno
late
mo
feti
l,ta
cro
lim
us,
bas
ilix
um
ab(d
ay1
po
sto
p),
pro
toco
l2
.m
yco
ph
eno
late
mo
feti
l,ta
cro
lim
us
and
bas
ilix
um
ab(1
day
po
sto
p),
pro
toco
l3
.m
yco
ph
eno
late
mo
feti
l,ta
cro
lim
us
day
1p
ost
op
.*1
std
ate
for
txO
cto
ber
7,
19
92,
2n
dd
ate
for
txF
ebru
ary
2,
19
98,
3rd
dat
efo
rtx
Jun
e1
2,
20
03
.#
4Y6
:p
atie
nts
use
dfo
rd
etai
led
anal
ysis
of
IL-1
0an
dIL
-12
.+
pro
toco
l1
plu
sri
tuxi
mab
2w
eek
sp
reo
p.
++
pro
toco
l2
plu
sd
aclu
zim
abd
ay-1
.
6 www.transplantjournal.com Transplantation & Volume 00, Number 00, Month 00, 2014
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

Interleukin-10 has previously been reported to down-regulate the immune stimulatory function of DCs in anumber of ways (12Y14). In this study, we investigated theeffect of IL-10 on DCs that are activated with IFN-F, TNF->,and IL-1A, a cytokine cocktail relevant when studying DCsactivated after graft reperfusion. These cytokines are well-known DC-maturating factors that show similar increasesin mRNA expression in human liver and kidney grafts incombined auxiliary liver-kidney transplanted patients (9).To confirm the proper activation of DCs in our in vitromodel, we studied phenotypic maturation markers on thecells by flow cytometry. Stimulation with the cytokinecocktail led to an increase in the characteristic maturationmarker CD83. Costimulatory molecules CD86 and CD40were also increased, consistent with proper maturation.Pretreatment with IL-10 resulted in a decreased level of co-stimulatory molecules, indicating that IL-10 pretreatmentlikely reduces the ability of DCs to stimulate an immuneresponse. It should be noted that the concentration of IL-10used for in vitro experiments is higher than what was seensystemically, however there is a possibility that the levelsmay be higher within the liver graft from which the IL-10production is supposed to emanate (8).
We showed that DCs activated in the presence of IL-10produced significantly lower levels of the NK and T cell-recruiting chemokines MIG/CXCL9, IP-10/CXCL10 and I-TAC/CXCL11. These CXCR3 ligands are required for DC-mediated NK cell recruitment to lymph nodes. CXCR3 is alsohighly expressed on both resting and activated CD8+ T cellsand is strongly upregulated on CD4+ T cells during activation(15). The influence of NK cells on allograft outcome has beenstudied in several publications and NK cells have repeatedlybeen shown to enhance the adaptive immune responsesleading to rejection (reviewed in (16)). Allogeneic DCs inducea CXCR3-dependent recruitment of NK cells to draininglymph nodes that is necessary for subsequent T helper 1(Th1)-deviated alloresponse (17). In line with these findings,the generation of human alloreactive cytotoxic T cells in vitrorequires the participation of NK cells (18). Thus, the reducedability of DCs activated in the presence of IL-10 to produceNK-recruiting chemokines, as shown in the present study,may have an impact on graft outcome. We could not observeany significant difference in the levels of the Th2 and Tregassociated cytokines CCL17 and CCL22 when treating theDCs with IL-10. This may indicate that Th2 cells or Tregs arenot involved or that other nonstudied, immunoregulatoryfactors are needed for the induction of tolerogenic dendriticcells that are able to recruit regulatory T cells. We also ob-served that the ability of DCs to produce the Th1-deviatingcytokine IL-12 after CD40-stimulation was reduced in IL-10-treated DCs. Th1 deviation of allogeneic T cells was alsoweakened in an MLR with IL-10-treated DCs as stimulatorycells, as indicated by the reduced ability of the responder cellsto produce IFN-F, a characteristic Th1 cytokine. Thus, ourfindings demonstrates that DCs that are matured with a cy-tokine cocktail relevant to liver transplantation are less able toboth recruit and activate allospecific T cells compared withDCs matured with a cocktail relevant to renal transplantation.
The proinflammatory cytokines IL-1A, IL-2, and IL-8were also released at significantly higher levels after liver andcombined auxiliary liver-kidney transplantation compared
with renal transplantation. IL-2 is secreted by activated T cellsand stimulates T cell proliferation and survival in an autocrinefashion. In transplantation, IL-2 has been found to have im-mune modulatory effects, as IL-2 is necessary for long-termgraft survival in certain transplant settings (19). IL-2 also fa-cilitates activation-induced T cell apoptosis and stimulatesregulatory T cells, two pathways for tolerance induction(19Y21). The differential secretion of cytokines after livercompared with renal reperfusion may thus modulate the ac-tivation of both antigen-presenting cells and effector cellsinfluencing graft survival.
To conclude, a rapid release of IL-10 is induced afterhuman liver transplantation, even with a small partial aux-iliary liver graft. This will affect the activation of passengerDCs, which are considered the major instigators of earlyallograft rejections. Our data suggest that passenger DCsthat migrate from the liver graft shortly after reperfusion areless able to recruit NK and T cells. This mechanism mayprevent harmful cross talk between DCs and effector cells,promoting graft acceptance.
MATERIALS AND METHODS
Cytokine Analysis of Patient SerumThe study subjects were patients undergoing combined auxiliary liver-
kidney transplantation, liver transplantation, and kidney transplantation.
For clinical characteristics of patients included in the study, see Table 1. Sera
were collected before surgery, 1 to 2 hr after surgery and 24 hr after surgery
from three combined auxiliary liver-kidney transplanted patients. Sera from
six renal transplantation recipients and four liver transplantation recipients
were collected at similar time points and used as controls. The levels of IL-
10, IL-12 p70, IFN-F, IL-13, IL-1A, IL-2, IL-4, IL-5, IL-8, and TNF-> were
measured in patient sera using the human TH1/TH2 multiplex panel (Meso
Scale Discovery, Gaithersburg, MD).
An in-depth study was performed in three additional patients undergoing
combined auxiliary liver-kidney transplantation. From these patients, sera were
collected at seven time points during and after surgery: before liver reperfusion,
5 min and 1 hr after liver reperfusion, before kidney reperfusion, 5 min and 1 hr
after kidney reperfusion and 24 hr after surgery. In these samples, IL-10 and IL-
12 levels were measured using Luminex-technology (Bio-Rad, Hercules, CA).
Generation of DCsPBMCs from healthy blood donors were isolated on density gradients
with Ficoll-Paque (GE Healthcare, Uppsala, Sweden). Cells were seeded in
24-well plastic culture plates at 2.5 x 106 cells per well and allowed to adhere
for 1.5 hr. After removal of non-adherent cells, the remaining cells were
cultured in AIM-V medium (Invitrogen/Life Technologies, Carlsbad, CA)
supplemented with recombinant human granulocyte macrophage colony-
stimulating factor and IL-4 both at 1,000 U/mL (R&D Systems, Minneapolis,
MN) for 5 days, resulting in immature DCs.
When indicated, DCs were generated from monocytes purified with
positive selection for CD14+ cells using magnetic-activated cell sorting
(Miltenyi Biotech, Germany). The purified cells were plated at 3�105 cells
per well and differentiated as described above.
Immature DCs were treated with 20 ng/mL IL-10 for 90 min before
addition of a maturation-cocktail consisting of IFN-F (1,000 U/mL), TNF->
(50 ng/mL) and IL-1A (25 ng/mL) (all from R&D Systems). After 24 hr the
supernatants were collected, the cells were washed and cultured for another
24 hr in AIM-V medium.
DC PhenotypeFor flow cytometry, CD14+ monocyte-derived cells were grown in
Cellgro DC medium (Cellgenix Technologie Transfer GmbH, Germany).
The cells were differentiated and activated as described above. After 24 hr in-
cubation in the presence or absence of IL-10, the cells were harvested and stained
with fluorescein isothiocyanate-conjugated anti-CD83, phycoerytrin-conjugated
* 2014 Lippincott Williams & Wilkins Ingelsten et al. 7
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

anti-CCR7, allophycocyanin-conjugated anti-CD86 and phycoerythrin-Cy5-
conjugated anti-CD40 (BD Biosciences, San Jose, CA). Cells were incubated at
4-C for 20 min and washed. Flow cytometry data were acquired on a
FACSAria cytometer (BD Biosciences) with Diva software (BD Biosciences).
DC production of IL-12 was also investigated in CD14+ monocyte-
derived DCs activated as indicated above. After 24 hr, DCs were washed
and stimulated with histidine-tagged CD40 ligand (200 ng/mL, R&D Sys-
tems) for 20 min. Thereafter, anti-polyhistidine monoclonal antibody
(4Kg/mL, R&D Systems) was added to the wells. After another 24 hr, IL-12p70
levels were measured in the supernatants by ELISA (R&D Systems) according
to manufacturer’s instructions.
Chemokine DeterminationThe production of chemokines was determined in cell culture supernatants
collected 24 hr after addition of cytokines. To determine the continuous
production of chemokines, the cells were washed and cultured for another
24 hr in AIM-V medium without addition of cytokines. Chemokine production
was analyzed by commercially available ELISAs specific for MIG/CXCL9, IP-10/
CXCL10, I-TAC/CXCL11, TARC/CCL17, and MDC/CCL22 (all from R&D
Systems) according to the manufacturer’s instructions.
Mixed Leukocyte ReactionTo investigate the ability of DCs matured in the presence or absence of
IL-10 to activate allogeneic T cells, an MLR was performed. CD14+
monocyte-derived DCs were differentiated for 5 days, matured by cytokine
stimulation for 24 hr, washed and incubated for another 24 hr and
rewashed. 1�106 freshly isolated PBMCs were added to the 24 well-plate for
a 1:5 ratio of DC:PBMC. Supernatants were collected from the MLRs after
6 days for IFN-F analysis using ELISA (R&D Systems).
StatisticsSerum cytokine levels are presented as geometrical means T SEM. Stu-
dent’s t-test was used to compare cytokine levels in the different groups.
Chemokine levels and IFN-F production in cell culture supernatants are
presented as mean TSEM, and the statistical significance of differences were
determined using Student’s paired t-test. Statistical analysis was performed
with SPSS v 15.0. P valuesG0.05 were considered statistically significant.
ACKNOWLEDGMENTSThe authors thank Lena Heijdenberg, Katarina Junevik,
and Kerstin Ebefors for technical assistance.
REFERENCES1. Larsen CP, Morris PJ, Austyn JM. Migration of dendritic leukocytes
from cardiac allografts into host spleens. A novel pathway for initia-tion of rejection. J Exp Med 1990; 171: 307.
2. Gordon RD, Fung JJ, Markus B, et al. The antibody crossmatch in livertransplantation. Surgery 1986; 100: 705.
3. Gonwa TA, Nery JR, Husberg BS, et al. Simultaneous liver and renaltransplantation in man. Transplantation 1988; 46: 690.
4. Vogel W, Steiner E, Kornberger R, et al. Preliminary results withcombined hepatorenal allografting. Transplantation 1988; 45: 491.
5. Rasmussen A, Davies HF, Jamieson NV, et al. Combined transplan-tation of liver and kidney from the same donor protects the kidneyfrom rejection and improves kidney graft survival. Transplantation1995; 59: 919.
6. Leca N, Warner P, Bakthavatsalam R, et al. Outcomes of simultaneousliver and kidney transplantation in relation to a high level ofpreformed donor-specific antibodies. Transplantation 2013; 96: 914.
7. Olausson M, Mjornstedt L, Norden G, et al. Successful combinedpartial auxiliary liver and kidney transplantation in highly sensitizedcross-match positive recipients. Am J Transplant 2007; 7: 130.
8. Le Moine O, Marchant A, Durand F, et al. Systemic release ofinterleukin-10 during orthotopic liver transplantation. Hepatology1994; 20: 889.
9. Ingelsten M, Gustafsson K, Oltean M, et al. Is indoleamine 2,3-dioxygenase important for graft acceptance in highly sensitized patientsafter combined auxiliary liver-kidney transplantation? Transplantation2009; 88: 911.
10. Weimer R, Mytilineos J, Feustel A, et al. Mycophenolate mofetil-basedimmunosuppression and cytokine genotypes: effects on monokinesecretion and antigen presentation in long-term renal transplant re-cipients. Transplantation 2003; 75: 2090.
11. Chhabra D, Grafals M, Cabral B, et al. Late conversion of tacrolimus tosirolimus in a prednisone-free immunosuppression regimen in renaltransplant patients. Clin Transplant 2010; 24: 199.
12. Faulkner L, Buchan G, Baird M. Interleukin-10 does not affectphagocytosis of particulate antigen by bone marrow-derived dendriticcells but does impair antigen presentation. Immunology 2000; 99: 523.
13. Haase C, Jorgensen TN, Michelsen BK. Both exogenous and endog-enous interleukin-10 affects the maturation of bone-marrow-deriveddendritic cells in vitro and strongly influences T-cell priming invivo. Immunology 2002; 107: 489.
14. Morel AS, Quaratino S, Douek DC, et al. Split activity of interleukin-10 on antigen capture and antigen presentation by human dendriticcells: definition of a maturative step. Eur J Immunol 1997; 27: 26.
15. Rabin RL, Park MK, Liao F, et al. Chemokine receptor responses on Tcells are achieved through regulation of both receptor expression andsignaling. J Immunol 1999; 162: 3840.
16. Kitchens WH, Uehara S, Chase CM, et al. The changing role of naturalkiller cells in solid organ rejection and tolerance. Transplantation2006; 81: 811.
17. Martin-Fontecha A, Thomsen LL, Brett S, et al. Induced recruitmentof NK cells to lymph nodes provides IFN-gamma for T(H)1 priming.Nat Immunol 2004; 5: 1260.
18. Kos FJ, Engleman EG. Requirement for natural killer cells in the in-duction of cytotoxic T cells. J Immunol 1995; 155: 578.
19. Dai Z, Konieczny BT, Baddoura FK, et al. Impaired alloantigen-mediated T cell apoptosis and failure to induce long-term allograftsurvival in IL-2-deficient mice. J Immunol 1998; 161: 1659.
20. Kang HG, Zhang D, Degauque N, et al. Effects of cyclosporine ontransplant tolerance: the role of IL-2. Am J Transplant 2007; 7: 1907.
21. Furtado GC, de Lafaille MAC, Kutchukhidze N, et al. Interleukin 2signaling is required for CD4+ regulatory T cell function. J Exp Med2002; 196: 851.
8 www.transplantjournal.com Transplantation & Volume 00, Number 00, Month 00, 2014
Copyright © 2014 Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.




















