Barry Wyke, MD, BS Gale trusts (Fig. 1) (Stack, 1963) - Europe ...
Upload
independentCategory
view
0download
0
BlOTROPlCA 36(1): 99-108 2004
Rain Forest Regeneration beneath the Canopy of Fig Trees Isolated in Pastures of Los Tuxtlas, Mexico'
Sergio Guevara2, Javier Laborde, and Graciela Sanchez-Rios
lnstituto de Ecologia, A.C., Krn 2.5 Carretera Antigua a Coatepec 351, Congregacion El Haya C.P. 91070 Xalapa, Veracruz, Mexico
ABSTRACT In the highly fragmented landscape of Los Tuxtlas, Mexico, we studied seed rain, seed bank, and vegetation established beneath the canopies of five fig trees (Ficw spp.) isolated in pastures. The area beneath the canopy of each fig tree was enclosed with a barbed wire fence to exclude cattle, and weeding with machete and herbicide spraying were stopped. Annual seed rain beneath the canopy of the five trees was 20,936 seeds (1066 seeds/m2) belonging to 149 species. Seed rain was dominated by woody species dispersed by frugivorous birds and bats. Most captured seeds (94%) were small (seed diam < 3 mm). A total of 6290 individuals from 78 species germinated from the seed bank (1258 seeds/m*). Ruderal herbs dominated the seed bank (52% of spp. and 77% of seeds). After three years of exclusion, a dense, 4-5 m tall understory comprised of woody plants developed beneath the canopy of the five fig trees. A total of 621 plants (4.6 plants/m2) from 95 species were recorded along 15 transects placed beneath the canopy of the fenced trees. Primary forest and pioneer tree and shrub species dominated species richness (55%) and abundance (37%) of established vegetation, but secondary and pioneer species had higher overall biomass. Most of the established plants (68%) had relatively large seeds (>3 mm). When cattle was excluded and management practices were stopped, remnant trees in Los Tuxtlas pastures effectively favored the early establishment of late successional species, including large-seeded canopy and subcanopy trees.
RESUMEN En el paisaje fragmentado en Los Tuxdas, Mkxico, se describe la lluvia de semillas, el contenido de semillas del suelo y la vegetacibn establecida bajo la copa de cinco irboles de EM spp. aislados en potreros. El irea bajo la copa de estos irboles fie cercada con el fin de excluir el @ado, el corte con machete y la aspersibn de herbicidas. Durante un afio se colectaron 20,936 semillas (1066 semillas/m2) pertenecientes a 149 especies. La lluvia de semillas estuvo dominada por especies leiiosas que son dispersadas por aves y murciklagos frugivoros. La mayoria de las semillas (94%) tuvieron un d ihe t ro menor a 3 mm. De las muestras de suelo germinaron 6290 semillas de 78 especies. El contenido de semillas del suelo estuvo dominado por hierbas ruderales (52% de las especies y 77% de las semillas). Despuks de tres afios de exclusibn, bajo la copa de 10s irboles estudiados, se desarrollb un denso sotobosque de 4-5 m de alrura, compuesto por plantas leiiosas. Se registrb un total de 621 plantas (4.6 plantas/m2) pertenecientes a 95 especies. Esta vegetacibn bajo la copa estuvo dominada tanto en nrimero de especies (55%) como en abundancia (37%) por plantas de la selva. Sin embargo, las especies secundarias y pioneras alcanzaron 10s valores de biomasa m k altos. La mayoria de las plantas establecidas bajo la copa de 10s irboles poseen semillas grandes (>3 mm de dihetro). Cuando el ganado y las pricticas de manejo son eliminados, se favorece bajo la copa de 10s irboles aislados en potreros de Los Tuxdas, el establecimiento de plantas de especies sucesionales tardias, que forman pane del dose1 y subdosel de la selva.
Ky word: Ficus sp.; isolated pasture trees; Los Turk; Mixico; regeneration; seed bank; seed rain; tropical rain forest.
THE MAIN ROUTES FOR THE REGENERATION OF TROP-
ICAL RAIN FOREST (TRF) plants after natural distur- bances are vegetative resprouting, germination of dormant seeds in soil, and germination of seeds recently dispersed from nearby seed sources (Mar- tinez-Ramos 1994). In abandoned pastures and crop fields, woody species regenerate by these same routes. The potential contribution of each route
' Received 16 September 2002; revision accepted 25 No- vember 2003.
Corresponding author; Tel. (228) 842 18 00; Fax (228) 8 18 78 09; e-mail: [email protected]
depends on clearing and management practices, in- cluding duration and intensity of agricultural use (Gbmez-Pompa et al. 1972, Purata 1986, Uhl et al. 1988), the extent, quality, and location of seed sources and the responses of seed dispersal agents to forest clearing and fragmentation (Janzen 1988, Guevara et al. 1998).
Frequent weeding with machete, spraying of se- lective herbicides, and prescribed burns on pas- tures, quickly eliminate the sprouting potential of woody plant remains (Uhl et al. 1988, Nepstad et al. 1990). After a few years, the soil seed bank of active pastures is exhausted of woody plants, while
99

100 Guevara, Laborde, and Sanchez-Rios
seeds of ruderal herbs and grasses build up (Kell- man 1980, Guevara 1986, Ganvood 1989). The only possible route for TRF regeneration in these pastures is seed dispersal from nearby forest rem- nants; however, pastures are commonly so extensive that the majority of their area is distant from po- tential seed sources (G6mez-Pompa et al. 1972, Holl 1999).
Most woody plants from TRF species depend on frugivorous vertebrates for seed dispersal. These vertebrates commonly avoid open areas (Nepstad et af. 1990, Cardoso et al. 1996) and seed dispersal into these pastures is severely limited. Forest seeds reaching the pasture must also overcome seed pre- dation and seedling herbivory, periods of drought, and competition with grasses and heliophytic ru- deral plants (Janzen 1988, Uhl etal. 1988, Nepstad et af. 1990, Gerhardt 1993, Aide & Cavelier 1994, Guariguata et al. 1995, Gondez-Montagut 1996, Slocum 1997). These obstacles are particularly in- tense in pastures deprived of tall trees. In the hu- mid tropics of Mexico and Central America, how- ever, pastures commonly have numerous and di- verse trees that are remnants of the original TRF canopy (Guevara 1986, Guevara et al. 1998, Har- vey 2000a, Slocum & Horvin 2000). These iso- lated trees are frequently visited by several species of frugivorous birds and bats that deposit tree seeds beneath their canopies (Guevara & Laborde 1993, Galindo-Gonzdez et al. 2000, Slocum & Horvitz 2000).
Several studies have found that in comparison to the surrounding open pasture, the vegetation growing beneath the canopy of isolated trees has a much higher density and richness of zoochorous woody plants (Guevara et af. 1986, 1992; Janzen 1988; Uhl et af. 1988; Nepstad et al. 1990; Holl 1999; Otero-Amaiz et al. 1999; Harvey 2000a; Slocum 2001), and therefore these trees may func- tion as forest regeneration nuclei (Guevara et al. 1986, Janzen 1988) after cattle grazing and man- agement practices are stopped. The outcome of stopping these anthropogenic disturbances beneath the canopy of the isolated trees, however, is still unknown. The removal of the grazers may provoke the profuse growth of palatable grasses and other pasture weeds that will quickly outcompete all woody plants (Janzen 1988, Holl 1999), deflecting succession. On the other hand, woody plants may grow and prevail, but in this case the regenerating woody vegetation may be composed of a reduced number of pioneer or early secondary species or woody vegetation may be formed by a rich assem- blage of tree and shrub species (including late suc-
cessional species of the original TRF). If the latter case is true, then the role of isolated trees in pas- tures as regeneration nuclei of the TRF will be more effective, even more so if the establishment of canopy and subcanopy tree species is favored very early.
The goal of this study was to assess the efficacy of isolated trees as potential regeneration nuclei for the TRF. We enclosed the area beneath the canopy of five Ficus trees isolated within pastures of Los Tuxtlas, Mexico, for a period of three years in order to exclude cattle and stop pasture management practices. We first describe the seed rain, seed bank, and established vegetation beneath these these five isolated trees in terms of species composition and community structure (species number, plant den- sity, and type of species). We then describe in detail the structure of the vegetation established after three years of cattle exclusion, including small seed- lings, and determine the most likely source of the established species.
METHODS Los Tuxtlas is a volcanic mountain range 90 km long and 30-50 km wide, rising from the sea up to 1680 m elevation. It is located in the southeast of the state of Veracruz, Mexico, along the Gulf Coast and is surrounded by extensive lowland plains (<lo0 m asl). Mean annual precipitation exceeds 4000 mm and the mean annual tempera- ture is 25°C. Soils are volcanic in origin, relatively young, and classified as Vitric Andosols. They show poorly developed profiles and relatively high amounts of organic material and nutrients. The original vegetation in the area was tropical rain for- est with a high canopy of 25 to 35 m (Bongers et al. 1988). For a detailed description of Los Tuxtlas, see Gonzdez-Soriano et af. (1997).
This study used two pastures from the ejido Balzapote (18"35'37"N, 95"03'05''W), which is lo- cated on leeward slope of the Los Tuxtlas range at elevations ranging between 0 and 400 m and is limited to the South by the Los Tuxtlas Biological Research Station. Balzapote was founded in 1955 when the felling of the TRF for agricultural pur- poses was started. Pastures covered more than 80 percent of the ejido's area by 1991, with less than 100 ha of the original TRF divided into several small fragments. For a detailed description of study site, see Guevara et al. 1998. Pastures are weeded with machete twice a year, herbicides are applied occasionally, and pastures are not burned. As is common in Los Tuxtlas, these pastures contain nar-

Regeneration under Isolated Pasture Trees 101
row strips of trees that have been left along rivers and streams, as well as isolated trees with an average density of 3.3 treedha, most of them being TRF species (Guevara et al. 1992, 1998; Gonzdez-Mon- tagut 1996).
EXPERIMENTAL DEsrGN.-we selected five isolated remnant trees belonging to Ficw spp.: three E yo- ponensis Desv., one E petenensis Lundell, and one E tecolutensis (Liebm.) Miq. The first two species correspond to the subgenus Pbarmacosycea, whereas the last species belongs to the subgenus Urostigma. All selected trees were taller than 24 m and had trunk diameters at breast height greater than 80 cm. Canopy cover ranged between 246 and 1012 m2. The trees were isolated by forest felling be- tween 1960 and 1975. By the time this study was undertaken, both pastures had experienced more than ten years of uninterrupted cattle ranching with a density of 1 to 1.5 adult cows/ha. In Feb- ruary 1988, a barbed wire fence was built to ex- clude cattle in the area below the canopy of each isolated fig tree. Weeding with machete as well as herbicide spraying were also stopped.
SEED FwN.-Seed traps were placed randomly be- neath the canopy of the five trees located within the fenced area, using a combination of two ran- dom numbers in a coordinate system (X, Y) in which one of the corners of the fenced corral was designated as the origin. Traps consisted of a cloth cone fastened to a 50 cm diameter circular wire structure and placed 60 cm above the ground. The number of seed traps per tree was 22, 15, 35, 19, and 9 and was proportional to the canopy area, with a total trap area equivalent to 0.7 percent of the canopy area. Total sampling area was 19.6 m2 (4.3, 2.9, 6.9, 3.7, and 1.8 m2). From 15 March 1988 to 16 March 1989, seed traps were collected monthly. Captured seeds were counted first as morphospecies, and identified later by direct com- parison with a reference seed collection at the Tux- tlas Research Station. For more details on seed rain methodology, see Guevara and Laborde 1993.
SEED BANK.-Seven months after fencing, 25 soil samples (20 X 20 X 5 cm deep each) were taken randomly below each of the five trees. Sampling area per tree was 1 m2. Large plant debris (mainly grass roots) and stones were removed using a sieve with a 1 cm mesh. The sieved soil was placed in trays inside a greenhouse and kept moist. Seedlings were counted monthly and removed from the trays until no new seedlings germinated. Counted seed-
lings were transplanted in separate containers for later identification. Germination ceased six months later.
ESTABLISHED VEGETATION.-The vegetation that had established within the enclosed area was sampled three years after fencing (April 1991). Three 1 m wide transects were set out from the trunk to the fence limit beneath each tree. Sampling areas among trees were 28, 26, 37, 26, and 18.4 m2. All plants within transects that were taller than 5 cm were identified to species and mapped, and the to- tal height and crown diameter were recorded. In addition, each woody stem was carefully examined to verify if it had established from seed or had sprouted from a root or a stump. Plants with at- tached cotyledons or cotyledonary leaves were re- corded as seedlings.
Analysis of vegetation height was restricted to 541 individuals that established during the three years of exclusion (ie. , they were not present before fencing). Individuals previously established and re- sprouted were excluded ( N = 20) from this anal- ysis.
DATA ANALYSIS.-For each species recorded in the seed rain, seed bank, and established vegetation, the following characteristics were determined: life-form (tree, shrub, vine, liana, herb, or epiphyte), dis- persal mode (anemochory, endozoochory, or oth- er), successional status (primary, forest pioneer, sec- ondary, or ruderal), and seed size (based on seed diam: <3, 3-7, 7-11, and >11 rnm). Designation of species attributes was based on published liter- ature about Los Tuxtlas (Guevara 1986; Purata 1986; Bongers et al. 1988; Ibarra-Manriquez & Si- naca 1995, 1996a,b; Gonzalez-Soriano et al. 1997). Two categories, forest pioneer and secondary, in- cluded early successional species. The former was comprised of species common in natural canopy gaps inside the forest, whereas the latter included plant species that were common in abandoned crop fields and extremely rare in canopy gaps. Ruderal species are heliophytic plants that grow in sites dis- turbed by humans. The importance value (IV) for each species was calculated for the established veg- etation as a summation of its relative frequency, abundance, and cover. Finally, we used Smensen’s similarity index (ssi) to compare the species com- position among woody species in seed rain, seed bank, and established vegetation.
RESULTS SEED RAIN.-A total of 20,936 seeds belonging to 149 species (plus 660 seeds of 15 unidentified

102 Guevara, Laborde, and Sanchez-Rios
TABLE 1. Number of species and density (m’) f . r seed deposition, seed bank, and vegetation established ajer three years of cattle exclusion (beneath the canopy o f j v e Ficus trees isolated in pastures of Los Tmtku, Mexico). Recent seedlings @lants with cotylehns are grouped separately). Results are grouped by Ipecies attributes.
Established vegetation
Seed rain Seed bank All plants Seedlings
SPP. Dens. SPP. Dens. Spp. Dens. Spp. Dens.
Life-form Tree 42 429.7 9 13.8 55 2.36 31 0.72 Shrub 23 397.8 8 115.2 24 1.63 13 0.21 Liana 9 2.1 1 0.2 7 0.06 3 0.02 Herb 38 166.1 43 963.6 8 0.49 4 0.14 Vine 14 15.9 5 146.8 1 0.05 0
0 Epiphyte 8 21.1 0 0
- - - 0
unknown 15 33.6 12 18.4 0 -
- -
Dispersal mode Zoochory 74 779.5 28 157.8 71 3.30 39 0.80 Anemochory 39 111.7 16 25.2 12 0.22 5 0.05 Other 23 142.7 20 1050.8 10 0.98 6 0.23
unknown 13 32.4 14 24.2 2 0.07 1 0.01
Primary 52 181.7 6 4.4 52 1.74 32 0.55 Pioneer 24 5 19.4 8 20.4 19 1.37 10 0.29 Secondary 14 81.2 11 25 1.4 18 1.10 8 0.17 Ruderal 42 248.2 41 963.4 5 0.37 1 0.08
Successional status
- unknown 17 35.8 12 18.4 1 0.01 0
Seed diameter (mm) <3 95 1001.8 52 1209.8 27 1.46 9 0.14
3-7 44 51.3 12 21.6 39 2.25 25 0.65 7-1 1 9 12.4 0 0.0 17 0.65 13 0.23
>11 1 0.8 0 0.0 11 0.23 4 0.07 unknown 0 - 14 26.6 1 0.01 0
Total 149 1066.3 78 1258.0 95 4.59 51 1.09 -
morphospecies) were captured in seed traps over a year. Accumulated seed density during the entire year of capture was 1066.3 seeds/m2 with a daily deposition rate of 2.9 seeds/m2/d. The mean num- ber of species per tree was 87.6 (SD ? 4.3) and mean seed density per tree was 1282.8 seeds/m2 (SD ? 686.8).
Tree and herb species accounted for 28 and 25 percent of species, respectively. Density was highest for tree and shrub species (Table 1). Most captured seeds belonged to species that are dispersed by fru- givorous birds or bats (47% of spp. and 73% of seeds). From the total captured seeds, 94 percent were smaller than 3 mm in diameter. Relatively few forest pioneer species (16% of total spp.) repre- sented half of all seeds captured; however, primary rain forest species were the most common type of dispersed species.
The most abundant species was Cecropia ob- tusifolia (25.4% of seeds), which was captured in all seed traps and had an accumulated total density
of 271.3 seeds/m2/yr. Piper auriturn was next most abundant species (10.5% of seeds), followed by Fi- cus spp. subgen. Urostigma (7.9% of seeds) and Piper hispidurn (5.9%).
SEED BANK.-A total of 6290 seedlings germinated from the soil samples, comprised of 78 species be- longing to 32 families. The best-represented family was Compositae with 12 species. Mean species richness per tree was 43.2 (SD t 4.7), and mean seed density was 1258 seeds/m2 (SD t 122.9). There were 92 seedlings from 15 unidentified morphospecies.
The seed bank was predominantly composed of herbaceous plants (61% of spp.). Most viable seeds in the soil (77% of germinated seedlings) were ruderal plants typical of Neotropical pastures (Table 1). Seeds from secondary species were the next most abundant type (20% of germinants). Particularly abundant taxa included several grasses (Paspalurn, Axonopus, and Panicurn), sedges

Regeneration under Isolated Pasture Trees 103
(Rhynchospora, Scleria, Killingia, and Fimbistrylis), and forbs (Desmodium, Mimosa, Borreria, and So- lanum).
Only 1 percent of the total viable seeds found in the soil were tree species and 9 percent were shrub species (Table 1). The most abundant woody species were the shrubs Conostegia xalapensis, Cor- dia spinescens, Neurolaena lobata and P+er hispi- dum. The last species is a pioneer species, while the others are secondary or ruderal.
ESTABLISHED VEGETATION.-A total of 62 1 individ- uals belonging to 95 species were established be- neath the canopy of the five fig trees after three years of cattle exclusion. The mean number of spe- cies established per fig tree was 36 (SD ? 7.5) species and mean plant density was 4.7 plants/m2 (SD 5 1.09).
Woody plants accounted for 90 percent of the total species (60% trees, 23% shrubs, and 7% lia- nas). Trees and shrubs accounted for 87 percent of all individuals (52% and 35%, respectively). Total vegetation cover for these two life-forms was 728 m2, which exceeded by more than five times the total sampled area.
Established vegetation was characterized by rainforest trees and shrubs with seeds larger than 3 mm in diameter (71% of spp.) and dispersed by frugivorous vertebrates (72% of spp. and 64% of individuals); however, most rain forest plants were small seedlings or saplings. Pioneer and secondary species attained higher biomass because they were taller and had larger canopies. Ruderal plants were poorly represented at the end of the study (only 49 plants representing 4 spp.; Table 1).
REsmovrrNc-Sixteen individuals (3%) of the es- tablished trees and shrubs resprouted from stumps. Ten of these were from the secondary forest tree Acacia cornigera. The other six represented the pri- mary forest tree species Lonchocarpus cruentus, Ro- llinia jimenezii, and Coccoloba m a d e . For each one of these tree species, recently established seed- lings were also present. Four additional trees (Cor- dia stenoclada, Diosmros digyna, and Pseudolmedia oxyphyllaria with an immature E tecolutensis stran- gler attached to it) were present before fencing.
Smxmcs.--.One hundred forty-eight plants from 51 species were in the seedling stage, resulting in a total density of 1.09 seedlings/m2 (Table 1). Mean seedling species richness beneath each fig tree was 15.8 (SD 2 8.9), and mean density was 1.2 seed- lings/m2 (SD ? 0.7). Most seedlings were TRF
primary tree species (with seeds larger than 3 mm) that were dispersed by frugivorous vertebrates. The most abundant species in this category were Cordia stenoclada, Cupania glabra, Bursera simaruba, and Cym bopetallum baillonii.
SPECIES coMrosiTioN.-P+er hispidum, Acalypha diversifolia, and Croton pyramidalis were recorded under all or most of the studied fig trees. These species were the only ones with an importance val- ue greater than 10 percent. Tabernaemontana ar- borea, Psychotria limonensis, Cupania glabra, Cym bopetalum bailhnii, and Parathesis lenticellata were the only primary forest species registered be- neath three or more of the study trees. The only ruderal herb with a high importance value was Pa- vonia schiedeana (Table 2).
VERTICAL PROFILE OF VEGETATIoN.-Height Of trees and shrubs ranged from 10 cm (seedlings) to 6.3 m. Vertical profiles showed that relatively tall in- dividuals of secondary forest and pioneer species formed the regrowth canopy characterized by a compact and dense foliage between 3 and 5 m above ground (Fig. 1). Beneath this regrowth can- opy, there were numerous seedlings and saplings of TRF primary species, whereas ruderal heliophytic plants were diminished by the dense shade. The outermost portion of each transect showed a higher density of plants, as well as a more crowded vertical profile, than the inner portions close to the trunk of the fig tree.
The height distribution of trees and shrubs showed a higher proportion of individuals shorter than 1.5 m (Fig. 2a). When seed size was taken into account, however, the latter tendency varied; most individuals of species with small seeds (<3 mm) were taller than 1.5 m (Fig. 2b). In contrast, the highest proportion of individuals for species with larger seeds (>3 mm) was less than 0.5 m tall (Fig. 2c, 2d).
COMPARISON OF SPECIES COMPOSITION AMONG SEED RAIN, SEED BANK AND ESTABLISHED VEGETATION.-A total of 117 woody species (trees, shrubs, and lia- nas) accumulated beneath the canopies of isolated trees when taking into account species in seed rain, seed bank, and established vegetation. Only 15 per- cent of these species appeared in the seed bank, whereas 62 percent were in seed rain and 72 per- cent in established vegetation.
A small proportion of all established woody species was present in the seed bank, and all of these also occurred in the seed rain. Seeds from half
104 Guevara, Laborde, and Sanchez-Rios
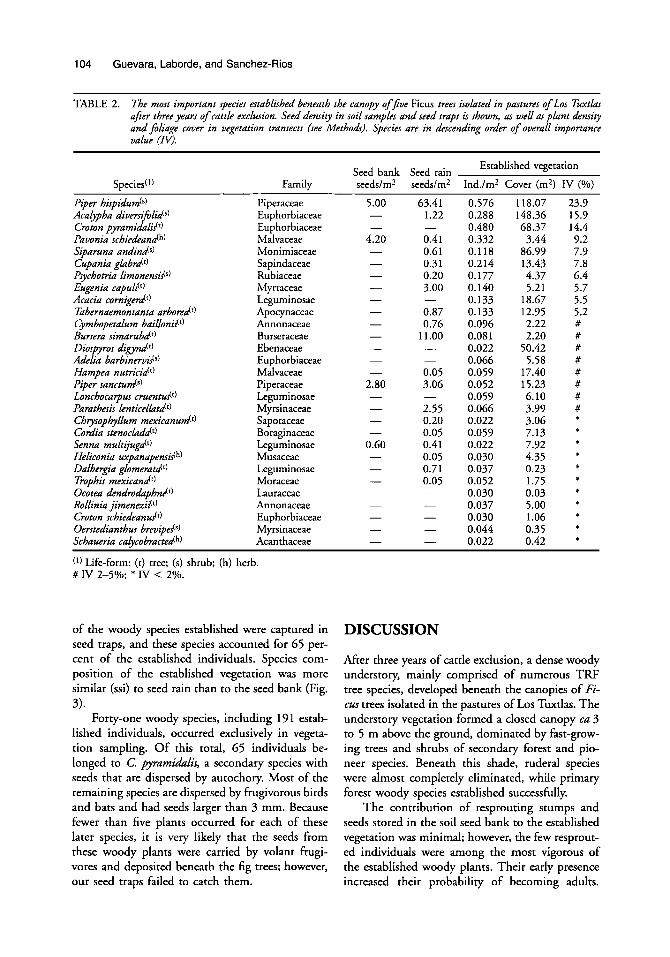
TABLE 2. The most important species established beneath the canopy offive Ficus trees isolated in pastures of Los Twtka afier three years of cattle exclusion. Seed density in soil samples and seed traps is shown, as well as plant density and filiage cover in vegetation transects (we Methodr). Species are in descending order of overall importance value (N).
~ ~~~~~
Established vegetation Seed bank Seed rain Family seeddm2 seeddm2 Ind./m2 Cover (m2) IV (%)
Piper hispidum(l) Acalypha diversi$lids) Croton pyramidalis!') Pavonia schiedeandh) Siparuna andin&) Cupania glabrd') Pychotria limonensik) Eugenia capulbl) Acacia cornigerd') Tabernaemontanta arbored') Cymbopetalum baillonib') Bursera simarubd') Diospyros d i a d ) Adelia barbinerv&) Hampea nutricid') Piper sanctum(s) Lonchocarpus cruend') Parathesis lenticellatd') Chrysopbyllum mexicanurn(') Cordia stenocla&') Senna multijugd') Heliconia wpanapensikh) Dalbergia glomeratd') Fophis mexirand') Ocotea dendrodaphnP) Rollinia jirnenezib') Croton schiedeand') Oerstedianthw brevipek) Schaueria calycobractedh)
Piperaceae Euphorbiaceae Euphorbiaceae Malvaceae Monimiaceae Sapindaceae Rubiaceae Myrtaceae Leguminosae Apocynaceae Annonaceae Burseraceae Ebenaceae Euphorbiaceae Malvaceae Piperaceae Leguminosae Myrsinaceae Sapotaceae Boraginaceae Leguminosae Musaceae Leguminosae Moraceae Lauraceae Annonaceae Euphorbiaceae Myrsinaceae Acanthaceae
63.41 1.22
0.41 0.61 0.3 1 0.20 3.00
0.87 0.76
1 1 .oo
-
-
- -
0.05 3.06
2.55 0.20 0.05 0.41 0.05 0.71 0.05
-
-
- - - -
0.576 0.288 0.480 0.332 0.118 0.214 0.177 0.140 0.133 0.133 0.096 0.081 0.022 0.066 0.059 0.052 0.059 0.066 0.022 0.059 0.022 0.030 0.037 0.052 0.030 0.037 0.030 0.044 0.022
118.07 148.36 68.37
3.44 86.99 13.43 4.37 5.21
18.67 12.95 2.22 2.20
50.42 5.58
17.40 15.23 6.10 3.99 3.06 7.13 7.92 4.35 0.23 1.75 0.03 5.00 1.06 0.35 0.42
23.9 15.9 14.4 9.2 7.9 7.8 6.4 5.7 5.5 5.2 # # # # # # # # t
* t
* * * * f
f
* t
( l ) Life-form: (r) tree; (s) shrub; (h) herb. # IV 2-5%; * W < 2%.
of the woody species established were captured in seed traps, and these species accounted for 65 per- cent of the established individuals. Species com- position of the established vegetation was more similar (ssi) to seed rain than to the seed bank (Fig. 3).
Forty-one woody species, including 19 1 estab- lished individuals, occurred exclusively in vegeta- tion sampling. Of this total, 65 individuals be- longed to C. pyramidalis, a secondary species with seeds that are dispersed by autochory. Most of the remaining species are dispersed by frugivorous birds and bats and had seeds larger than 3 mm. Because fewer than five plants occurred for each of these later species, it is very likely that the seeds from these woody plants were carried by volant frugi- vores and deposited beneath the fig trees; however, our seed traps failed to catch them.
DISCUSSION
After three years of cattle exclusion, a dense woody understory, mainly comprised of numerous TRF tree species, developed beneath the canopies of Fi- cus trees isolated in the pastures of Los Tuxtlas. The understory vegetation formed a closed canopy ca 3 to 5 m above the ground, dominated by fast-grow- ing trees and shrubs of secondary forest and pio- neer species. Beneath this shade, ruderal species were almost completely eliminated, while primary forest woody species established successfidly.
The contribution of resprouting stumps and seeds stored in the soil seed bank to the established vegetation was minimal; however, the few resprout- ed individuals were among the most vigorous of the established woody plants. Their early presence increased their probability of becoming adults.

Regeneration under Isolated Pasture Trees 105
h
E Y
E 0) a, I .-
FIGURE 1 . Vegetation profile of plants established beneath the canopy of one Firw yoponensis tree (3) isolated in a Los Tuxtlas pasture after three years of cattle exclusion. The figure shows one 1 m wide transect from the fig tree trunk to its canopy periphery. The height, canopy diameter, and successional status of each established plant are shown (see text).
Most established individuals within our transects belonged to tree and shrub species that usually do not form part of the soil seed bank; their relatively large seeds lack dormancy and germinate promptly after being deposited on the ground (Guevara 1986, Garwood 1989, Kitajima 1996). Woody plants established beneath the canopies of the iso- lated fig trees came mostly from recently dispersed seeds, which were deposited there by frugivorous birds or bats (Guevara & Laborde 1993, Galindo- Gonzdez et al. 2000).
As in other studies of seed rain in pastures, our seed rain data showed an overwhelming majority of small seeds (<3 mm), most of which belonged to early successional species (Nepstad et al. 1990, Guevara & Laborde 1993, Gonzdez-Montagut 1996, Harvey 2000b, Slocum & Horvitz 2000). Larger seeds from TRF trees were captured in such low numbers that their relative contribution to seed rain could be regarded as negligible. This trend, however, has also been documented beneath ma- ture TRF canopy within undisturbed areas (Smythe 1970, Jackson 1981, Soto 1992), since dispersal potential is inversely correlated with seed size (Ki- tajima 1996). The probability that a particular seed trap will capture a small seed can be orders of mag- nitude higher than that of capturing a large one. Therefore, before concluding that TRF tree species with large seeds are not dispersed effectively
throughout pastures (based solely on the analysis of seed rain data), we must consider the low dis- persal potential of these species.
Data on seed availability should be used care- fully when predictions about future vegetation of a site are attempted. Seed quantities, or their relative abundance, tell us nothing without knowing the seed biology and characteristics of the species in- volved. One particularly important characteristic is seed size. Our data demonstrate that once seeds were deposited beneath the canopy of an isolated tree, the rate of success in seedling establishment for large seeds was higher than those with small seeds, despite the low availability of the former and the high densities of the latter.
Small-seeded pioneer species should be abun- dant in the soil beneath the canopies of the five study trees because of their abundant seed rain and dormancy capabilities, but our data showed the op- posite. The case of Cecropia obtusifoolia is particu- larly illustrative. Although we captured more than 250 seeds/m2 from this species in the seed rain, less than 3 seeds/m2 occurred in the seed bank. These seeds are extremely abundant in soil samples from the forest floor as well as in crop fields of Los Tux- tlas (Guevara 1986). A possible explanation is that heavy seed predation of small seeds occurred within the pastures. In pastures from Balzapote, experi- ments with C. obtusfoolia and Heliocaps appendi-

106 Guevara, Laborde, and Sanchez-Rios
HEIGHT (rn)
2.5-3.5
1.5-2.5
0.5-1.5
SO.5
2.5-1.5
1.5-2.5
0.5-1.5
CO.5 >3'5E 2.5-3.5
1.5-2.5
0.5-1.5
0.0% 10.0% 20.0% 30.0% 40.0% 50.0% %Total
~~
FIGURE 2. Height distributions of all trees and shrubs established beneath the canopy of five Ficus trees isolated in pastures of Los Tuxtlas. (a) All species (521 individu- als), and species grouped by seed size (mm): (b) 3 (196); (c) >3 but <7 (N = 228); (d) >7 (97).
culatw confirmed fast seed removal (Gonzalez- Montagut 1996) by Solpnopsis geminata, the most abundant ant in Los Tuxtlas pastures, which is al- most absent in the forest (Quiroz & Valenzuela 1995). Preferential seed predation by ants may ex- plain the scarcity of seeds of pioneer species in our soil samples (20 seeds/m2), even though they were abundantly deposited (>500 seeds/m2) in our seed traps.
The area beneath the canopies of isolated trees also has a high cover of grasses and tuderal plants in actively managed pastures of Los Tuxtlas (Gue- vara et al. 1992). Frequent weeding and cattle graz- ing maintain this situation. When these anthro- pogenic disturbances were eliminated, however, the competitive outcome within our exclosures was switched in favor of small-seeded woody plants that were able to shade the grasses and ruderal herbs.
Established (4.0y vegetation
FIGURE 3. Total species richness and density (m2) of all woody plants (trees, shrubs, and lianas; N = 117 spp.) in seed rain, seed bank, and established vegetation be- neath the canopies of five isolated Ficus trees. Within cir- cles, are the number of species shared among sampled components (overlapping areas) and those exclusive to each component. * = density in established vegetation; ssi = Ssrensen similarity index.
Microenvironmental conditions beneath the rem- nant trees were substantially different than in open pasture. Canopy opennness is lower beneath trees compared to open pasture (closer to the trunk, PAR is 1.2-9.3% of sunlight in the pasture, where- as under the crown edge PAR is 10.9-33.5%). Rel- ative humidity decreases and air temperature in- creases with the distance from the trunk toward the pasture, and soil temperature is higher at 0 to 1 m from the tree trunk; (Williams-Linera et a/. 1998). The exclusion of cattle and the shade provided by the canopy of the fig tree, together with heavy seed rain, were the key factors that allowed the estab- lishment of fast-growing trees and shrubs of pio- neer and secondary species. After three years of cat- tle exclusion, the dense shade of fast-growing woody species also limited their own establishment, creating better conditions for the establishment of large-seeded plants with slower growth rates, such as those of TRF woody species.
In our study, the major impediments to TRF regeneration in pastures were surmounted by the presence of large trees within pastures. Our results confirmed that large and tall trees isolated in pas- tures effectively facilitate forest recovery in aban- doned pastures by enhancing seed dispersal of TRF tree species into the pasture and by shading out grasses and ruderal forbs beneath their canopies (see also Holl et al. 2000). The presence per se of a tree in a pasture does not, however, determine the successful establishment of TRF plants beneath

Regeneration under Isolated Pasture Trees 107
its canopy. Other necessary conditions are the pres- ence of seed sources within dispersal distance limits and the action of seed dispersal agents. But in order for trees to be effective regeneration nuclei, cattle browsing and weeding with machete must be stopped beneath the canopies of the isolated trees; otherwise, the established TRF trees and shrubs will not survive past the seedling stage.
It was impossible to include control plots in areas of open pasture in this study because farmers granted permission for us to use areas only around the five selected trees. Los Tuxtlas pastures are com- monly overgrazed, and therefore, open sites are very valuable to the farmers. Many other studies have demonstrated that seed deposition of woody plants is orders of magnitude higher beneath the canopy of large pasture trees than in the open grassland, by studying the seed rain directly (Uhl et al. 1988, Nepstad et al. 1990, Gonzda-Montagut 1996, Harvey 2OOOb, Holl et al. 2000, Slocum & Horvitz 2000), or indirectly by sampling the established seedlings or saplings in active pastures (Guevara et al. 1986, 1992; Janzen 1988; Otero-Amaiz et al. 1999). Several authors agree that large isolated trees in pastures may help overcome the main barriers to rain forest regeneration in pastures (Guevara et al. 1992, 1998; Aide & Cavelier 1994; Cardoso et al. 1996; Holl 1999). Before this study, however, there was no clear indication of what would actu- ally happen beneath the canopy of isolated trees if the pasture were to be abandoned.
The potential function of isolated trees as re- generation nuclei for the TRF is present from the moment at which the tree is isolated in the middle of a cleared area and may continue until the tree
dies, provided that seed input is maintained. Since the studied pastures are going to be maintained as such for a long time, we should ensure replacement of the remnant tree within the pastures. Ideally, the original tree should be replaced by another tall tree of a TRF canopy species; beneath the canopy of the five studied fig trees, we found seedlings and saplings of 32 tree species that reach the forest can- opy or subcanopy as adults, which represent feasi- ble candidates for the replacement of the original tree. Simple practices such as fencing the area be- neath the canopy of the isolated tree for a few years (.g., 5 yr), as well as selective thinning, can facil- itate the replacement of the original tree by another tall tree of a TRF species. These easy and extremely inexpensive maintenance practices could be imple- mented immediately in many areas of tropical America, particularly in fragmented landscapes in which TRF fragments are surrounded by extensive pastures. The goal will be to preserve a relatively high density of isolated trees within today's pastures in order to secure a faster forest recovery in exten- sive areas that can be reclaimed for restoration in the future.
ACKNOWLEDGMENTS We are grateful to Robin Chazdon for her valuable revi- sion of the manuscript. We also thank Patricia Moreno- Casasola, Bianca Delfosse, and Mariano Guevara for their assistance in the field. For species identification we are indebted to Santiago Sinaca and especially to Guillermo Ibarra. Many thanks to the students for their invaluable help in sorting the seeds. Financial support was provided by the National Council of Science and Technology- CONACYT (4127P-N9607) and by the Instituto de Ecologia, A.C. (I. E. 902-16).
LITERATURE CITED AIDE, M., AND J. CAVELIER. 1994. Barriers to lowland tropical forest restoration in the Sierra Nevada de Santa Marta,
BONGERS, F., J. POPMA, J. MEAVE DEL CASTILLO, AND J. CARABIAS. 1988. Structure and floristic Composition of the
CARDOSO, J. M., C. UHL, AND G. MURRAY. 1996. Plant succession, landscape management and the ecology of frugiv-
GALINDO-GON~LEZ, J., S. GUEVARA, AND V. SOSA. 2000. Bat and bird generated seed rains at isolated trees in pastures
GARWOOD, N. C. 1989. Tropical soil seed banks: a review. In L.A.R. Leck, R.L. Simpson and V.T. Parker (Eds.).
GERHARDT, K. 1993. Tree seedling development in tropical dry abandoned pasture and secondary forest in Costa Ria.
G~MEZ-POMPA, A., c. VAZQUEZ-YANES, AND s. GUEVARA. 1972. The tropic4 rain forest: A nonrenewable resource.
GONZALEZ-MONTAGUT, R. 1996. Establishment of three rain forest species along the riparian corridor-pasture gradient
GONZ~EZ-SORLANO, E., R. DIRZO, AND R. VOCT. 1997. Historia Natural de Los Tuxtlas. Instituto de Biologia, Uni-
Colombia. Rest. Ecol. 2: 219-229.
lowland rain forest of Los Tuxtlas, Mtxico. Vegetatio 74: 55-80.
orous birds in abandoned Amazonian pastures. Conserv. Biol. 10: 491-503.
in a tropical rain forest. Conserv. Biol. 14: 1693-1703.
Ecology of seed banks, pp. 149-190. Academic Press, San Diego, California.
J. Veg. Sci. 4: 95-102.
Science 177: 762-765.
in LQS Tuxtlas, Mtxico. Ph.D. dissertation. Harvard University, Cambridge, Massachusetts.
versidad Nacional Autonoma de Mtxico, Mtxico, D. F., Mexico.

108 Guevara, Laborde, and Sanchez-Rios
GUARICUATA, M., R. RmiNC;ANs, AND F. MoNTAcNiNi. 1995. Early woody invasion under tree plantations in Costa Rica: Implications for forest restoration. Rest. Ecol. 3: 252-260.
GUEVARA, S. 1986. Plant species availability and regeneration in Mexican tropical rain forest. Ph.D. dissertation. Uppsala University, Uppsala, Sweden. , AND J. LABORDE. 1993. Monitoring seed dispersal at isolated standing trees in tropical pastures: Consequences for local species availability. Vegetatio 107/108: 319-338.
-- , AND G. SANCHEZ. 1998. Are isolated remnant trees in pastures a fragmented canopy? Selbyana 19: 34-43. , J. MEAVE, I? MORENO-CASASOLA, A N D J. LABORDE. 1992. Floristic composition and structure of vegetation under isolated trees in Neotropical pastures. J. Veg. Sci. 3: 655-664. , E. PURATA, AND E. VANDER MAAREL. 1986. The role of remnant forest trees in tropical secondary succession. Vegetatio 66: 77-84.
HARVEY, C. 2000a. Colonization of agricultural windbreaks by forest trees. Effects of connectivity and remnant trees. Ecological Applications lO(6): 1762-1793. . 2000b. Windbreaks enhance seed dispersal into agricultural landscapes in Monteverde, Costa Rica. Ecol.
HOLL, K. D. 1999. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate and soil. Biotropica 31: 229-242. , M. E. LOIK, E. H. V. LIN, AND I. A. SAMUELS. 2000 Tropical montane forest restoration in Costa Rica: Overcoming barriers to dispersal and establishment. Rest. Ecol. 8: 339-349.
IBARRA-MANR~QUEZ, G., AND S. S i N A a . 1995. Lista floristica de la estaci6n de Biologia Tropical Los Tuxtlas, Veracruz. Mtxico. Rev. Biol. Trop. 43: 75-1 15. , AND - . 1996a. Lista floristica de la estaci6n de Biologia Tropical Los Tuxtlas, Veracruz. Mtxico. Mimosaceae-Verbenaceae. Rev. Biol. Trop. 44: 41-60. , AND - . 1996b. Lista floristica de la estaci6n de Biologia Tropical Los Tuxtlas, Veracruz. Mtxico. Violaceae-Zingiberaceae. Rev. Biol. Trop. 44: 427-447.
JACKSON, J. F. 1981. Seed size as a correlate of temporal and spatial patterns of seed fall in a tropical forest. Biotropica
JANZEN, D. 1988. Management of tropical fragments in a tropical dry forest: Growth. Ann. Mo. Bot. Gard. 75: 105-
KELLMAN, M. 1980. Geographic patterning in tropical weed communities and early secondary successions. Biotropica
KITAJIMA, K. 1996. Ecophysiology of tropical tree seedlings. In S. Mulkey, R. Chazdon, and A. Smith (Eds.). Tropical rain forest plant ecophysiology, pp. 559-596. Chapman and Hall, New York, New York.
~ R T ~ N E Z - R A M O S , M. 1994. Regeneraci6n natural y diversidad de especies arb6reas en selvas hbmedas. Bol. SOC. Bot. Mtx. 54: 179-224.
NEPSTAD, D., C. UHL, AND E. A. S. SERRAO. 1990. Surmounting barriers to forest regeneration in abandoned, highly degraded pastures: A case study from Paragominas, Pad, Brazil. In A. B. Anderson (Ed). Alternatives to deforestation: Steps toward sustainable use of the Amazon rain forest, pp. 21 5-229. Columbia University Press. New York, New York.
OTERO-ARNAIZ, A., S. CASTILLO, J. MEAVE, AND G. IBAm-MANRiQuEz. 1999. Isolated pasture trees and the vegetation under their canopies in the Chiapas Coastal Plain, MCxico. Biotropica 31: 243-254.
PURATA, S. 1986. Floristic and structural changes during old-field succession in the Mexican tropics in relation to site history and species availability. J. Trop. Ecol. 2: 257-276.
QuiRoz, L., AND J. VALENZUEIA. 1995. A comparison of ground ant communities in a tropical rainforest and adjacent grassland in Los Tuxtlas, Veracruz, Mtxico. Southwest. Entomol. 20: 203-213.
SLOCUM, M. G. 1997. Reforestation in a Neotropical pasture: Differences in the ability of four tree taxa to hnction as recruitment foci. Ph.D. dissertation. Miami University, Oxford, Ohio. . 2001. How trees species differ as recruitment foci in a tropical pasture. Ecol. 82: 2547-2559. , AND C. HORVITZ. 2000. Seed arrival under different genera of trees in a Neotropical pasture. Plant Ecol. 149: 51-62.
SMYTHE, N. 1970. Relationships between fruiting seasons and seed dispersal methods in a Neotropical forest. Am. Nat. 104: 25-35.
SOTO, C. 1992. Patrones espaciales y temporales de la lluvia de semillas de grupos de irboles en una selva hbmeda de Mtxico. Tesis profesional. Universidad Autbnoma de Mtxico, Facultad de Ciencias, Mtxico, D.F., Mexico.
UHL, C., R. BUSCHBACHER, AND E. A. S. SERRAO. 1988. Abandoned pastures in Eastern Amazonia I: Patterns of plant succession. J. Ecol. 76: 663481.
WiLLws-LiNEra, G., v. DOMINGUEZ-GASTEL~, AND M. E. GARC~A-ZURITA. 1998. Microenvironment and floristics of different edges in a fragmented tropical rain forest. Conserv. Biol. 12: 1091-1102.
Appl. 10: 155-173.
13: 121-130.
116.
12(Suppl.): 34-39.
Copyright © 2022 FDOKUMEN