
Barry Wyke, MD, BS Gale trusts (Fig. 1) (Stack, 1963) - Europe ...
26
THE NEUROLOGY OF JOINTS Arris and Gale Lecture delivered at the Royal College of Surgeons of England on 17th February 1966 by Barry Wyke, M.D., B.S. Neurological Laboratory, Department of Applied Physiology, Royal College of Surgeons of England 'But the expression ofa well-made man appears not only in his face, It is in his limbs andjoints also, it is curiously in the joints ofhis hips and wrists, It is in his walk, the carriage ofhis neck, theflex ofhis waist and knees....." -WALT WHITMAN, Leaves of Grass (1891-2) THE THEME I propose to pursue in this lecture-the neurology of joints- involves consideration of the innervation of the joints of the skeleton, and a description of the role of this innervation in the regulation of muscular *~~~~~~~~~~~~~~~~~~~~~..r;e.- _`kg'7- Reprodu.ced by permission of the Worshipful Company of Barbers Fig. 1. A portrait by Richard Greenbury (now in the Guildhall Art Museum, London) of Mr. Edward Arris (on the right) in attendance at one of the Muscular Lectures endowed by him. The lecture was delivered at Surgeon's Hall by Sir Charles Scarborough (on the left) in 1649-although the picture was not painted until 1651. activity. This would seem an appropriate modern counterpart to the three-hundred-year-old intentions of the co-founders of the Arris and Gale trusts (Fig. 1) (Stack, 1963)-which were intended at their inception to provide a series of lectures on muscular and osteological topics (Copeman, 1963; Dobson, 1964). 25
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Barry Wyke, MD, BS Gale trusts (Fig. 1) (Stack, 1963) - Europe ...

THE NEUROLOGY OF JOINTSArris and Gale Lecture delivered at the Royal College of Surgeons of England
on17th February 1966
byBarry Wyke, M.D., B.S.
Neurological Laboratory, Department of Applied Physiology,Royal College of Surgeons of England
'But the expression ofa well-made man appears not only in hisface,It is in his limbs andjoints also, it is curiously in thejoints ofhis hips and wrists,It is in his walk, the carriage ofhis neck, theflex ofhis waist and knees....."
-WALT WHITMAN, Leaves of Grass (1891-2)
THE THEME I propose to pursue in this lecture-the neurology ofjoints-involves consideration of the innervation of the joints of the skeleton, anda description of the role of this innervation in the regulation of muscular*~~~~~~~~~~~~~~~~~~~~~..r;e.-_`kg'7-
Reprodu.ced by permission of the Worshipful Company of BarbersFig. 1. A portrait by Richard Greenbury (now in the Guildhall Art Museum,London) of Mr. Edward Arris (on the right) in attendance at one of the MuscularLectures endowed by him. The lecture was delivered at Surgeon's Hall by SirCharles Scarborough (on the left) in 1649-although the picture was not painted
until 1651.
activity. This would seem an appropriate modern counterpart to thethree-hundred-year-old intentions of the co-founders of the Arris andGale trusts (Fig. 1) (Stack, 1963)-which were intended at their inceptionto provide a series of lectures on muscular and osteological topics(Copeman, 1963; Dobson, 1964).
25

BARRY WYKE
Our interest in the subject of articular neurology arises from realizationthat the neurology of joints, in spite of its obvious relevance to severalbranches of surgery, and to physical medicine generally, is a field ofknowledge that has never been adequately explored*; and for this reason,when the new Neurological Laboratory in this College was opened three
(a)
(c)
(b)Fig. 2. Dissections of the specific articular nerves supplying the knee joint (in thecat). (a) The left posterior articular nerve (indicated by arrow), arising from theposterior tibial nerve (n) at the lower end of the popliteal fossa, and branching toenter the posterior capsule of the joint (c) on the right. x 1. (b) The medialarticular nerve exposed on the antero-medial aspect of the right thigh after removalof the sartorius muscle. In this instance, the nerve arises from the obturator nervetrunk and passes anteriorly through the adductor musculature to the front of thethigh. It branches just above the medial condyle of the femur below, close to themedial superior genicular artery (a): (p) is the patella. x i. (c) The lateral articu-lar nerve (indicated by large arrow) supplying the left knee joint. It arises from thelateral popliteal nerve (n) on the surface of the lateral head of the gastrocnemiusmuscle (G) and ascends to branch into the inferior edge of th- lateral capsule of thejoint (c). A second articular branch (indicated by small arrow) arises distally fromthe lateral popliteal nerve, to supply the superior tibio-fibular joint (t). x .
years ago, we decided to devote its excellent resources to a co-ordinatedattack on the subject. To this end, we have pursued the general topicof articular neurology along three parallel lines, employing similar methodsof investigation in each case:
* Thus, in a recent comprehensive monograph on joints, Barnett et al. (1961) wereforced to write: " In the present state of knowledge, it is impossible to define with anyprecision the role of the receptors in the joint tissues in the control of posture, movement,tone of muscles and so forth."
26

THE NEUROLOGY OF JOINTS
1. Studies of the limb and vertebral joints, whose neurological func-tions are important to orthopaedic and neurological surgeons, and topractitioners of physical medicine.
2. Studies of the laryngeal joints, and of the ossicular joints of themiddle ear, in relation to the mechanisms of speech and hearing-a subjectthat is of obvious significance to otolaryngologists and speech therapists.
3. Studies of the temporo-mandibular joints, whose neurology is rele-vant to many problems of oral and facio-maxillary surgery.We are fortunate in having made a certain amount of progress in each
of these fields, the details of which are recounted in the papers and mono-graphs listed at the end of this lecture. In the course of this work,however, it has become clear that the neurological aspects of each of thesesystems of joints share certain basic features in common; and it is to thesemore general aspects of articular neurology that I here wish to drawattention.
THE INNERVATION OF JOINTSAs with every neurological investigation, the first step in this field is
anatomical, and involves the definition of the structural features of thenerve supply of each system of joints.
Extrinsic innervationThe extrinsic innervation of each joint is being investigated by macro-
and micro-dissection in animals and man. In this way, it has been foundthat each joint has a dual pattern of nerve supply-first, by specificarticular nerves that reach the joint capsule as independent branches ofadjacent peripheral nerves, often (but not exclusively) in company with thearticular blood vessels; and secondly, by non-specific articular branches ofrelated muscle nerves. These latter arise within some of the musclesthat are attached to each joint capsule as intramuscular branches of variousmuscle nerves, and reach the joint by running through the substance of themuscles, embedded in the interfascicular connective tissue. Two examplesfrom different joint systems will serve to illustrate these points.
1. The knee joint is supplied by three specific articular nerves-aposterior, a medial and a lateral articular nerve, derived respectively fromthe posterior tibial, femoral (or obturator) and lateral popliteal nerves.Dissections of each of these are displayed in Figure 2-the posteriorarticular nerve (in the popliteal fossa) on the upper left (Fig. 2a), the medialarticular nerve (on the antero-medial aspects of the thigh) on the right(Fig. 2b), and the lateral articular nerve (on the supero-lateral aspect of theleg) on the lower left (Fig. 2c). The knee joint is also supplied by numerousnon-specific intra-muscular branches derived from the nerves to thesartorius and quadriceps musculature, and by a few twigs derived from thecutaneous nerves innervating the skin over the joint.
2. Again, the laryngeal joints are each supplied by specific articularbranches of the recurrent and internal laryngeal nerves, and by intra-
27

BARRY WYKE
muscular twigs running within some of the muscles attached to the laryn-geal cartilages. Thus, Figure 3 shows an articular branch of the internallaryngeal nerve that innervates the crico-arytenoid joint.As similar relationships appear to obtain with all other joints, it seems
that the basic implications of Hilton's Law (1863)* are applicable to all thejoints of the body, and not merely to those of the limbs-although modi-fications of detail, which I cannot describe here, have to be made in apply-ing this law to the individual joints.
Fig. 3. The innervation of the crico-arytenoid joint (cat). The articular nerve(indicated by arrow) arises from the internal laryngeal nerve (I). The larynx isviewed from the left, after removal of the thyroid cartilage (the cut edges of whichare seen at TC). (T) is the trachea, with the recurrent laryngeal nerve (R) beside it.
x2
Constitution of articular nervesNeurohistological studies have shown that each articular nerve contains
a mixture of myelinated and unmyelinated nerve fibres, whose diametersrange from less than lI up to 17ji. Considered in combination with theresults of oscillographic analyses of the impulse traffic in the articularnerves, of electrical stimulation procedures, and of other neurophysio-
* Hilton stated: "The same trunks of nerves whose branches supply the groups ofmuscles moving a joint, also furnish a distribution of nerves to the skin over the in-sertions of the same muscles and-what at the moment more especially merits ourattention-the interior of the joint receives its nerves from the same source."
28

THE NEUROLOGY OF JOINTS
logical investigations to be described later, these studies suggest that thefibres in articular nerves generally may be subdivided into three principalsize categories, as indicated in Table I, each of which has specific functionalcorrelates.
First, a large proportion (at least 45 per cent) of the total number offibres in each articular nerve has diameters of less than 51. Most of thesesmall myelinated and unmyelinated fibres, which are embraced in GroupIlI of the Table, are afferent in function and subserve articular painsensation; but a small proportion of the unmyelinated fibres in this groupconsists of visceral efferent fibres of sympathetic origin, that innervate thearticular blood vessels that is, these latter are articular vasomotor nervefibres. Thus far, we have no anatomical or physiological evidence of thepresence of secretomotor fibres in the articular nerves-or, indeed, of anydirect nervous influence on the production of synovial fluid.
TABLE IGENERAL COMPOSITION OF ARTICULAR NERVES
Group Diamleter Structure Functionnumber rai;ge
(p)I 13-i7 Large myelinated Mechanoreceptor afferent
(from o:nt ligaments)II 6- 12 Medium myelinated Mechanore eptor afferent
(from fibrous capsule and fatpat ))
III 2-5 Small myelinated Pai:t afferent<2 Unmyelinated Pain afferent
Vasomotor efferent
Secondly, another large proportion (some 45 to 55 per cent of the total)consists of medium-sized myelinated fibres between 6p and 12i in diameter,and forms Group II of our classificat on. All of these fibres are mechano-receptor afferents, innervating small corpuscular endorgans located in thefibrous capsules and fat pads of the joints that will be described in amoment.
Thirdly, a small proportion (some 10 per cent, or less) consists of largemyelinated fibres between 13 and 17p in diameter, and forms Group Iof our classification. These also are mechanoreceptor afferents, inner-vating large corpuscular endorgans that are confined to the joint ligaments.This last group of fibres is absent from the articular nerves of the larynxand middle ear, but is represented in most of the other articular nerves thatwe have examined.
In the case of the limb and vertebral joints, the nerve cell bodies fromwhich the articular afferent fibres derive are located in the segmentallyrelated dorsal root ganglia; and in the case of the laryngeal joints, they aresituated in the ganglion nodosum of the vagus nerve. We have notyet established the location of the nerve cell bodies of the afferentfibres innervating the temporo-mandibular and ossicular joints-but
29

BARRY WYKE
we have a priori reasons for expecting that they will be found in themesencephalic nucleus of the fifth nerve in the case of the mechanoreceptorafferents from the temporo-mandibular joints, and in the trigeminalganglion in the case of the pain afferent fibres from these same joints.
Articular receptor endingsUsing specially modified neurohistological staining techniques* in-
volving gold, silver and methylene blue procedures, it has been possible toshow that the articular nerve fibres just described terminate in fourstructurally distinct varieties of ending within the articular tissues (Fig. 4).Correlated neurophysiological investigations, by ourselves and by others,have further revealed that these morphologically distinct categories ofnerve ending possess individually distinct behavioural characteristics;and, on this basis, we have proposed a general structure-activity schemefor the classification of articular nerve endings, the basic criteria for whichare summarized in Table 1I.
Type I receptorsThe Type I receptors, a group of which is shown in Figure 4a, are
globular or ovoid corpuscles, similar to those described originally byRuffini (1905) in subcutaneous and fascial connective tissues. They arenumerous in relation to all the limb joints, the apophyseal joints of thevertebral column, and the temporo-mandibular joints; but in the larynx,they are restricted to the crico-arytenoid joint, being absent from thethyro-epiglottic, thyrohyoid and cricothyroid joints. In the middle ear,we have likewise failed to find them in the ossicular joints-but this nega-tive observation cannot yet be considered definitive, as our histologicalstudy of this system of joints is not yet complete. In the limbs, the Type Ireceptors appear to be more numerous in the proximal (for example, inthe shoulder and hip) joints than in the distal joints; and in the spine,they appear to be more numerous in the joints of the cervical region thanelsewhere-but again, this distribution requires further statistical analysisbefore it can be regarded as settled.
In each joint in which they are present, the Type I receptors are locatedmainly in the superficial (that is, in the external) layers of the fibrous cap-sule (as shown in Fig. 4a), within which they are distributed tridimension-ally in clusters of up to six corpuscles. Each member of such a clusteris supplied by terminal branches from a single Group II myelinated parentafferent axon some 6-9pi in diameter, that enters the joint capsule in oneof the articular nerves-as can be seen in Figure 4a. Within the capsuleof each individual joint, the clusters of Type I receptors show regionaldifferences in distribution density, in that they are more numerous on
* We are deeply grateful to Dr. M. J. F. Fitzgerald (Department of Anatomy,University College, Cork, Eire) and to Dr. C. Coers (Departement de Neurologie,Universit6 Libre de Bruxelles) for advice and assistance in the development of some ofthese procedures for use on articular tissues.
30

THE NEUROLOGY OF JOINTS
Fig. 4. The four varieties of articular receptor nerve ending (descriptions in text),as seen in the articular tissues of the cat. (a) A Type I receptor cluster (onecorpuscle in which is indicated by arrow), embedded in the superficial surface ofthe fibrous capsule of the joint. The myelinated afferent nerve fibre (N) can beseen joining an articular nerve trunk (A) on the right. Wyke gold chloride method.x 350. (b) A Type II corpuscle (indicated by arrow) in the joint capsule, at thejunction between the fibrous capsule (FC) and the sub-synovial fibro-adiposetissue (FA). The myelinated afferent nerve fibre (N) can be seen joining anarticular nerve branch at (A). Fitzgerald frozen silver method. x 150. (c) AType III receptor corpuscle (indicated by arrow) on the surface of a collateralligament (L), near its fusion with the fibrous capsule of the joint (FC). (N)indicatesthe myelinated afferent nerve fibre. Fitzgerald frozen silver method. x150.(d) Portion of the Type IV plexus system in the fibrous joint capsule, the super-ficial (external) surface of which is seen above and to the left. The fine nervefibres, weaving amongst the small blood vessels, arise from myelinated and unmye-linated parent nerve fibres within the fibrous capsule. Wyke gold chloride method.x 150. (e) Type IV free nerve endings ramifying amongst the fibres of a joint
ligament. Fitzgerald frozen silver method. x 150.
31

BARRY WYKE
those aspects of the joint capsule that undergo the greater changes instress during natural joint movement.
Physiologically, the Type I corpuscles behave as low-threshold, slowlyadapting mechanoreceptors responding to the mechanical stresses ob-taining in the part of the fibrous capsule in which they lie. For this reason,a proportion of the Type I receptors in each joint capsule is always activein every position of the joint, even when it is immobile. This restingdischarge usually has a frequency of some 10-20 impulses per sec., and isgenerated partly by the stresses created regionally within the jointcapsule by the varying degrees of tone in the muscles attached to it, andpartly by the overall capsular stress created by the fact that the intra-capsular (i.e. the intra-articular) pressure is normally some 5-10 mm. Hgless than the external atmospheric pressure. Alterations-either increases
TABLE IICLASSIFICATION OF ARTICULAR RECEPTOR SYSTEMS
Type Morphology Location Parent nervefibres Behavioural characteristicsI Thinly encapsulated globular Fibrous capsule Small mvelinated Static and dynamic
corpuscles (I00 x 40p), in ofjoint (mainly (6-9u) mechanoreceptors; low-clusters of 3-6 corpuscles superficial layers) threshold, slowly adapting
JI Thickly encapsulated conical Fibrous capsule Medium myelinated Dynamic mechanoreceptorscorpuscles (280u x 120Au), of joint (mainly (9-12A) low-threshold, rapidlyin clusters of 2-4 corpuscles deeper layers). adapting
Articular fat pads
III Thinly encapsulated fusiform Joint ligaments Large myelinated Dynamic mechanoreceptors-corpuscles (600g x 100l) (intrinsic and (13-17,) high-threshold, very slowly
extrinsic) adapting
IV Plexuses and free nerve Fibrous capsule. Very small myelinated Pain receptors; high-endings Articular fat pads. (2-5A) threshold, non-adapting
Ligaments. Walls Unmyelinatedof blood vessels (<2,u)
or decreases-in the rate of this resting discharge occur whenever the jointis moved actively or passively, whenever the tone in the related muscleschanges isotonically or isometrically, or whenever the pressure gradientbetween the interior of the joint and the atmosphere is altered sufficiently.When such changes in capsular stress are imposed, the Type I receptors inthe affected regions of the joint capsule respond with an abrupt rise or fallin their resting discharge rate, after which they adapt slowly (overseveral seconds) to a new discharge rate that is determined by the newdegree of stress now prevailing in the affected parts of the joint capsule.In the light of these data, then, the Type I articular receptors can be cate-gorized as static and dynamic mechanoreceptors, whose discharge patternsignals static joint position, intra-articular pressure changes, and thedirection, amplitude and velocity ofjoint movements.
Type II receptorsThe Type II receptors, one of which is illustrated in Fig. 4b, are elon-
gated conical corpuscles, with a thick, multi-laminated connective tissuecapsule enclosing a single (or sometimes multistranded) unmyelinatednerve terminal that ends in a bulb or Y-shaped bifurcation near the apex
32

THE NEUROLOGY OF JOINTS
of the corpuscle. * Within the capsule of the endorgan, the terminal axonis ensheathed by a single palisade-like layer of clear, columnar cells; andit is derived from an extra-corpuscular myelinated axon that is about 5jin diameter. These corpuscles have been observed in all the joints wehave examined, without exception. In the joints of the larynx and middleear they constitute the sole corpuscular endorgan present (except for thefew additional Type I corpuscles mentioned earlier as being present inthe capsule of the crico-arytenoid joint); and in the limbs, they are rela-tively more numerous in distal joints (for example, in the ankle joint) thanin more proximal joints. They are also particularly numerous in thetemporo-mandibular joints.
Like the Type I receptors, the Type II corpuscles are found in thefibrous capsules of all the joints, partly in the superficial layers-where,in the case of the limb, vertebral, temporo-mandibular and crico-arytenoidjoints, they are mixed with varying numbers of Type I corpuscles. Mostof the Type II corpuscles are located, however, in the deeper (that is, in theinternal) layers of the fibrous capsules of the joints, particularly (as can beseen in Fig. 4b) at the border between the fibrous capsule and the sub-synovial fibro-adipose tissue, where they often lie alongside, or coil around,the articular blood vessels. They are distributed in each joint capsule inclusters usually of 2-4 corpuscles, each member of such a cluster beinginnervated (as shown in Fig. 4b) by a branch of a Group II parent mye-linated articular nerve fibre whose diameter is between 9p and 12p. Similarclusters of Type IL endings are present also on the surfaces of all the fatpads related to the limb and temporo-mandibular joints, whether these beintra-articular or extra-articular.The Type II corpuscles behave as low-threshold, rapidly adapting
mechanoreceptors. They are entirely inactive in immobile joints,becoming active only at the onset or cessation ofjoint movement-that is,at the moment at which sudden changes of stress occur in the regions ofjoint capsule or fat pad in which they lie. When they are so stimulated,each cluster of Type II receptors emits a brief, high-frequency burst ofimpulses into the related afferent axon that lasts less than one second;and in rapidly moving joints, such as those in the larynx, the duration ofthis impulse burst may be less than 0.5 sec. Furthermore, as the diameterof the afferent nerve fibres innervating the clusters of Type II corpuscles isgreater than that of the fibres innervating the Type I clusters, the centri-petal conduction velocity of the Type II volley is faster (by some 20-40metres per sec.) than that of the impulses emanating from Type I corpuscles.
In summary, then, the Type II corpuscles can be regarded solely asdynamic mechanoreceptors, whose brief, high-velocity discharges signaljoint acceleration and deceleration.
* Some previous workers have regarded these corpuscles as modified forms of theVater-Pacinian corpuscle-but, for reasons discussed elsewhere (Wyke, 1967), wedo:not agree with this homology (see also Fitzgerald, 1962).
33

BARRY WYKE
Tjpe III receptorsWhereas the Type t and Type 11 corpuscles are joint capsule receptors,
the Type III corpuscles-an example of which is shown in Figure 4c-areconfined to the joint ligaments. They are the largest of the articularcorpuscles, and are identical structurally with the tendon organs of Golgi-of which they appear to be the articular homologue. As can be seen inFigure 4c, each Type III corpuscle is a fusiform endorgan applied longi-tudinally to the external surfaces of the joint ligaments, usually near theirbony attachments; and it consists of a filmy connective tissue capsule en-closing a mass of densely arborizing nerve filaments derived from a large,Group 1 myelinated parent axon that may be up to 17p in diameter.A few of these corpuscles are located on all the extrinsic (that is, the
collateral) ligaments of the limb and spinal apophyseal joints, and onintrinsic joint ligaments such as the cruciate ligaments in the knee joint.Some are also present in relation to the lateral ligament of the temporo-mandibular joint; but they are absent from the longitudinal ligaments ofthe vertebral column. The Type III corpuscles are also entirely absentfrom the ossicular joints of the middle ear, and from all the laryngealjoints.The stimulus-response characteristics of these endorgans have not yet
been studied as thoroughly as those of the Type I and Type II corpuscles;but the data available thus far suggest that the Type III endorgans behaveas high-threshold, slowly adapting mechanoreceptors-as do the Golgiendorgans in the tendons related to joints. Thus they are completelyinactive in immobile joints, and only become active towards the extremesofjoint movement-that is, when considerable stresses are generated in thejoint ligaments. In these circumstances, the Type III corpuscles emita stream of impulses that travel centripetally at high velocity in the largediameter afferent fibres in the articular nerves; and the frequency of thisdischarge adapts only very slowly (over many seconds), if the extremejoint displacement be maintained.
Type IV receptorsThe Type IV category of articular receptors embraces the non-corpus-
cular nerve endings in the joint tissues, being represented by lattice-likeplexuses (Fig. 4d) and free nerve endings (Fig. 4e). These terminationsare derived from the smallest (Group III) afferent fibres in the articularnerves-some of which (those between 2g and 5p in diameter) are thinlymyelinated, whilst the remainder (less than 2g in diameter) are unmye-linated.The plexus or network system of terminals, an example ofwhich is shown
in Figure 4d, is prominent in the limb, spinal apophyseal and temporo-mandibular joints, in each of which it is distributed throughout the fibrouscapsule and adjacent periosteum, the articular fat pads (both external andinternal), and the adventitial sheaths of the articular blood vessels. In the
34

THE NEUROLOGY OF JOINTS
tissues of these joints, free nerve endings are sparse, being confined largelyto the intrinsic and extrinsic joint ligaments, as shown in Figure 4e. In thecapsules of the laryngeal and ossicular joints, however, the networksystem is less well developed, and there are then many free nerve endingsdistributed throughout the fibrous joint capsule.
This Type IV category of ending constitutes the pain receptor system ofthe articular tissues. As such, the plexuses and free nerve endings areentirely inactive in normal circumstances; but they become active whenthe articular tissues containing this type of ending are subjected to markedmechanical deformation, or to direct mechanical or chemical irritation.In this connection, it should be emphasized that this variety of receptorending is entirely absent from the synovial lining of every joint that hasbeen examined, and is also lacking from the menisci present in the kneeand temporo-mandibular joints, and from the intervertebral discs. Thereis no mechanism, then, whereby articular pain can arise directly from thesynovial tissue or menisci in any joint; and surgical removal of synovialtissue or joint menisci likewise does not involve removal of pain-sensitivearticular tissues.
FUNCTIONS OF ARTICULAR RECEPTOR SYSTEMSHaving described the general pattern of innervation of the joints of the
body-which appears to be essentially similar in all mammalian jointsystems-I should like to devote the remainder of this lecture to a con-sideration of the functional significance of this articular innervation.
Articular kinaesthetic sensationAlthough Sherrington and his colleagues customarily included the
joints within the system of proprioceptive inputs that influence muscletone, his interests in this respect were largely in the receptors in the musclesthemselves. For this reason, it was for long believed that the muscular-and to a lesser extent the cutaneous-mechanoreceptors were the principalreflex regulators of striated muscle tone at a segmental level, and the majorcontributors to postural sensation.
In recent years, however, it has become apparent that mechanoreceptorsin the articular tissues make substantial contributions to the perceptionof joint posture and movement-a contribution that is not made by themuscle receptors. This suggestion-that receptors in the joints might besignificant contributors to kinaesthetic sensation-was originally madeat the end of the last century by Goldscheider (1898); but it is the con-siderable amount of recent work in experimental psychology and neurologyover the last decade that has provided ample confirmation of their im-portance in this respect-so much so, that the articular mechanoreceptorscan now be regarded as the major contributors to postural and kinaesthe-tic sensation, supplementary to the information provided by vision andfrom the skin.
35

BARRY WYKE
Articular mechanoreceptor (arthrokinetic) reflexesLess adequately delineated, however, is the possible r6le of the articular
receptors in the reflex regulation of muscle tone; and so it is to this aspectof their function that we have particularly directed our attention. Inthe remainder of this lecture, then, I should like to outline the results ofsome of the experimental and clinical studies that we have made in orderto throw light on the reflex significance of the articular receptor systemsthat I have just described.
Previous experiments have indicated that the discharges from the Type Itendorgans-which, you will recall, behave as low-threshold, rapidlyadapting mechanoreceptors-are transient, occurring only at the onset orcessation of movement. Our investigations have confirmed that this isindeed so; but they have further revealed that these transient dischargesfrom the Type II receptors are responsible for provoking brief reflexchanges in the tone of the muscles operating over the joints being moved-changes that may be facilitatory or inhibitory in nature, and which arereciprocally co-ordinated between individual muscle groups.One of the sites in which this effect can be conveniently demonstrated is
the larynx, where the joints (with the sole exception of the crico-arytenoidjoint) contain only Type II mechanoreceptors. If we take a joint such asthe cricothyroid joint, which we know to be provided solely with Type IIreceptors, it can be shown (Figs. 5, 6) that passive movements of this jointgive rise to very short-lived, reflexly co-ordinated changes in the tone ofthe intrinsic muscles of the larynx.
Figure 5a shows portion of an electromyogram recorded from thedetached left cricothyroid muscle, during the performance of passivemovements of the isolated left cricothyroid joint. It is apparent from thistracing that the onset and cessation of joint displacement is, in each in-stance, associated with a very brief burst of motor unit potentials in themuscle, the time course of which is compatible with their being provokedby the evanescent discharge of the Type II mechanoreceptors in the movingjoint capsule. Furthermore, as Figure Sb shows, movements of thecricothyroid joint in appropriate directions may produce equally short-lived changes in the tone of the other intrinsic muscles of the larynx thatare reciprocally co-ordinated. Thus, as can be seen in Figure Sb, antero-medial displacement of the cricothyroid joint produces a brief increase inmotor unit activity in the thyro-arytenoid muscle (upper tracing), that isaccompanied by an equally brief inhibition of motor unit activity in itsantagonist posterior crico-arytenoid muscle (lower tracing).That these changes in motor unit activity do in fact arise reflexly in
response to the discharge of mechanoreceptors located in the moving jointcapsule can be demonstrated in four types of control experiment.
First, we have shown that identical reflex responses can be provoked inthese same muscles by direct electrical stimulation (with appropriate
36

THE NEUROLOGY OF JOINTS
parameters) of the articular nerves supplying the cricothyroid joint.Secondly, we have shown that the reflex muscular responses to cricothyroidjoint movement (such as are depicted in Figure 5) can be totally and per-manently abolished by section of these same nerves. Thirdly, if thereceptor nerve endings in the joint capsule be destroyed by direct electro-coagulation, then all the reflex changes in laryngeal muscle tone thatotherwise occur when the joint is moved are abolished and this abolitionis permanent. On the other hand, reversible abolition of the rapidly
'
(a) Reproduced by couirtesy of Ann. Otol. Rhinol. Laryngol.
Fig. 5. Electromyograms of articular reflexes in the intrinsic muscles of thecat's larynx, recorded from pairs of needle electrodes inserted directly into themuscles. (a) Rapidly adapting motor unit discharges provoked in the detachedleft cricothyroid muscle at the onset and cessation of passive caudal displacementof the isolated ipsilateral cricothyroid joint. The interrupted line indicates aperiod of 60 sec. during which constant joint displacement was maintained. (FromKirchner and Wyke, 1965d.) (b) Reciprocally co-ordinated, rapidly adaptingarticular reflex responses in the thyro-arytenoid muscle (above) and posteriorcrico-arytenoid muscle (below), displayed in simultaneous recordings duringantero-medial displacement of the isolated ipsilateral crico-thyroid joint. Notethat, immediately following the joint movement, brief facilitation of motor unitactivity in the (adductor) thyro-arytenoid muscle is accompanied by transient in-hibition of activity in the (abductor) posterior crico-arytenoid muscle. Note alsothat slowly adapting changes in motor unit activity are absent in each of the
tracings in (a) and (b).
adapting reflex responses in the laryngeal muscles can be produced byinfiltration of the capsule of the cricothyroid joint with a very smallamount of a 1 per cent solution of Lignocaine hydrochloride-a potentlocal anaesthetic agent that has a short-lived action. Thus, in Figure 6you can see that the rapidly adapting motor unit response that is provokedreflexly in the thyro-arytenoid muscle by movement of the cricothyroidjoint (upper tracing) is abolished within six minutes of infiltrating the jointcapsule with this local anaesthetic agent (second tracing). Fifty minutes
37

BARRY WYKE
later, however, as the effects of the local anaesthetic agent begin to wear off,the motor unit response to joint movement can be seen to be re-emerging(third tracing); and this process continues, so that after the lapse of aboutone hour the motor unit potentials are well on the way back to theiroriginal pattern (fourth tracing).
Reproduced by courtesy of Ann. Otol. Rhinol. Laryngol.
Fig. 6. Reversible suppression of articular reflex responses in the thyro-arytenoidmuscle to passive movement of the isolated ipsilateral cricothyroid joint by localanaesthesia of the joint capsule with 0.1 ml. of 1 per cent Lignocaine hydrochloridesolution. Note that the rapidly adapting motor unit response to caudal jointdisplacement seen in the upper electromyogram is absent in the second tracing: thethird and fourth tracings show the gradual return of the motor unit responses as theeffect of the anaesthetic agent on the nerve endings in the joint capsule wanes.
(From Kirchner and Wyke, 1965d.)
In our opinion, this type of experimental sequence-which we havealso applied to the limb and temporo-mandibular joints (vide infra)-lendspowerful support to the view that the Type 11 receptors are in fact rapidlyadapting mechanoreceptors that are provoked into transient dischargesat the onset and cessation of joint movement. Further, such experimentsreveal that these brief discharges reflexly provoke short-lived bursts of
38

THE NEUROLOGY OF JOINTS
motor unit activity in some of the muscles related to the joint, with coin-cident short-lived inhibition in other muscles functionally related to thesame joint.Thus the Type II receptors may be regarded as behaving (in most sites,
at least) as " booster" mechanoreceptors, evoking supplementary motorunit activity transiently in the prime movers of a joint at the onset ofmovement (with coincident brief inhibition of their antagonists) in order toovercome the inertia of the immobile parts; but once the joint is in motiontheir activity ceases, until the parts are brought to rest once more. In thelight of this information it will be apparent that the Type IL articularmechanoreceptors can make no contribution to the perception of staticjoint position; nor is it likely, in view of the very brief duration of theirafferent discharge, that they make any significant contribution to per-ception of joint movement. In short, the Type II mechanoreceptorsappear to be purely reflexogenic.
In the joints of the body other than the larynx and the middle ear, how-ever, the reflex operations of the rapidly adapting Type 1I mechanorecep-tors are supplemented, in normal circumstances, by those of the slowlyadapting Type I mechanoreceptors; and I should like to illustrate thesecombined effects in two different joint systems-beginning with the tem-poro-mandibular joints.As Figure 7 shows, stimulation of the Type It receptors in one temporo-
mandibular joint by movement of the ipsilateral half of the mandibleprovokes transient reflex changes in motor unit activity in the relatedmasticatory muscles at the onset ofjaw movement, similar to those I havedemonstrated in the laryngeal muscles. For example, in the upper partof Figure 7 you can see that slow centric opening of the mouth provokes,in the isolated ipsilateral temporalis muscle, a transient discharge of motorunits at the beginning of the jaw movement. The bottom half of Figure 7shows, however, that centric closure of the mouth provokes a slowlyadapting (and fluctuating) increase in motor unit activity in the samemuscle, that persists throughout the time that the jaw is kept closed butstops when the mouth is re-opened. In this case, then, passive movementsof the temporo-mandibular joint in appropriate directions give rise bothto rapidly adapting and to slowly adapting reflex changes in the tone ofthe masticatory musculature-and this correlates with our histologicalobservations that the temporo-mandibular joint capsule contains a mixtureof Type II and Type I mechanoreceptors. That these reflex effects arisefrom mechanoreceptors located in the capsule of the temporo-mandibularjoint has been confirmed by an experimental procedural sequence (in-volving articular nerve stimulation and section, and electrocoagulation andlocal anaesthesia of the joint capsule) similar to that already described inrespect of the laryngeal joints.
This group of experiments suggests, then, that in those joints equippedboth with Type I and Type II mechanoreceptors, the Type I receptors
39

BARRY WYKE
exercise a continuous (or tonic) influence over the reflex regulation of thetone of the muscles operating over the joint in question-in contrast (andsupplementary) to the discontinuous (or phasic) influence exerted by theType II receptors. This combination of influences is particularly im-portant in respect of the mechanoreceptors located in the capsules of thelimb joints; and one further set of experiments will serve to demonstratethe co-ordinated reflex effects of the mixture of rapidly adapting Type 11and slowly adapting Type I mechanoreceptors that is present in all the limbjoints.
These effects can be illustrated in respect of any limb joint; but theone I have chosen for present purposes is the ankle joint. For example(Fig. 8a), an electromyogram of the tenotomized tibialis anterior muscleduring passive plantarflexion of the related foot shows that, as the move-
TZM~ALE.
~'lsdOpeuil1-
Centric~~~~~~~~Cnti~~~~~~~~~~~~~~~~~~~~~~centric 1 9vOpen I sec.
Fig. 7. Electromyograms of temporo-mandibular articular reflexes in thedetached temporalis muscle of the cat, provoked by passive movements of the iso-lated ipsilateral ramus of the mandible. Both tracings are from the same prepara-tion, with bipolar needle electrodes inserted directly into the muscle: all move-ments were made without occlusal contact of the teeth. The upper tracing, andthe right-hand end of the lower tracing, shows rapidly adapting motor unit responsesprovoked by passive opening of the mouth: the principal part of the lower tracingshows the fluctuating, slowly adapting responses to passive closure of the mouth.
ment begins, there is a brief, high-amplitude discharge of motor unitsthat dies away within about two seconds, melting into a more prolongeddischarge of motor units that persists-but with a slow diminution infrequency-as long as the foot is held in the plantarflexed position. Con-versely, as Figure 8b shows, passive dorsiflexion of the same foot provokesa similar dual pattern of discharge in the tenotomized gastrocnemiusmuscle. As our histological investigations show that the capsule of theankle joint contains both Type I and Type II receptors, this dual patternof muscle reflex response to ankle joint movement is to be expected; andagain, in the light of the data already presented, it seems reasonable toattribute the initial rapidly adapting motor unit discharge to the reflexeffects of Type II receptor activity; whilst the slowly adapting, lower-frequency discharge that persists during maintenance of the joint in its
40

THE NEUROLOGY OF JOINTS
new position can be attributed to the reflex effects of the Type I receptordischarge.
Proof that these reflexes are in fact derived from mechanoreceptors inthe capsule of the ankle joint is provided by the same experimental pro-cedures as before, of which Figure 9 provides a typical illustration. Thus,as Figure 9 shows, electrocoagulation of the ankle joint capsule per-manently abolishes the rapidly and slowly adapting motor unit responsesin the tenotomized tibialis anterior muscle during plantarflexion of thefoot. Again, as in the experiments on the larynx (Fig. 6) and with thetemporo-mandibular joints, the motor unit responses in the leg muscles
(a) , 00jIOOpV2 Sec.
(b) , O#2'Sec.
Fig. 8. Electromyograms of articular reflex responses in the leg muscles of thecat to passive movements of the ipsilateral ankle joint. Prior to the recordings, acuff of skin was removed from around the joint region, and all tendons operatingover the joint were divided and freed. (a) Rapidly and slowly adapting motor unitresponses in the tenotomized tibialis anterior muscle to plantarflexion of the foot.(b) Rapidly and slowly adapting motor unit responses in the tenotomized gastroc-
nemius muscle to dorsiflexion of the foot.
to passive movement of the ankle are temporarily abolished by infiltrationof the capsule of the joint with a small quantity of local anaestheticsolution, and permanently abolished by articular neurectomy.
Finally, it should perhaps be pointed out that direct mechanical stimu-lation of the capsule of any of the joints is capable of provoking similarreflex responses in the related muscles. Thus, as Figure 10 shows, directgentle compression of the capsule of the ankle joint provokes both rapidlyand slowly adapting increases in the motor unit activity of the relatedgastrocnemius muscle (a)-and these responses can no longer be obtainedafter electrocoagulation of the nerve endings in the joint capsule (b).As a result of these experimental studies, I hope you will allow that it is
reasonable to propose that the mechanoreceptors in the capsules of all the
41

BARRY WYKE
joints make significant contributions to the reflex regulation of the toneof the muscles related to those joints-provoking either facilitation orinhibition during joint displacement, depending upon the direction andamplitude of the movement of the joint in question. At the onset ofmovement, the Type II receptors in the joint capsule and fat pads areprovoked into a brief episode of high-velocity impulse discharge that givesrise reflexly to a short-lived, reciprocally co-ordinated alteration in motor
Before Electrocoogulation Of Toint Capsule
(a) L L.JlOO)1V2 Sec.
After Electrocoagulation Of 3oinl Capsule
(b) . , 10OO)JV2 Sec.
Fig. 9. Abolition of articular reflex responses in the leg muscles to passive move-ment of the ankle joint by electrocoagulation of the joint capsule. Both electro-myograms were recorded from the same muscle (tibialis anterior). (a) Motor unitresponses evoked in the tenotomized tibialis anterior muscle by passive plantar-flexion of the ipsilateral foot. (b) Following capsular electrocoagulation, the motorunit responses are absent. The myotatic reflex (not illustrated) was still elicitable
in the muscle.
unit activity in the related muscles. At the same time, and throughout thewhole course of joint movement, the Type I receptors continuously con-tribute a fluctuating discharge whose precise frequency varies with thedegree and velocity of joint displacement; and this gives rise reflexly toan equally continuous but fluctuating alteration-involving either facilita-tion or inhibition-in the activity of the motor units in the muscles oper-ating over the joint. The Type III receptors, which (as I have mentioned)are present only in the joint ligaments, are not provoked into activity inthe circumstances of experiments such as those I have described-unlessthe joint manipulations produce extreme degrees of joint displacement:
42

THE NEUROLOGY OF JOINTS
then, and only then, do these high threshold receptors discharge, to pro-duce reflex inhibition of motor unit activity in the muscles operating overthe joint. The Type IV pain receptors in the articular tissues are likewisenot activated in these experiments; but other experiments that we haveperformed show that they can be-by sufficiently intense irritation of thejoint capsule, fat pads or ligaments-and their discharge then provokesintense non-adapting motor unit responses simultaneously in all themuscles related to the joint, as well as in more remote muscles elsewherein the body.
Compression Of Joint Capsule Before Electrocoagulation
(a) on Off,,fff
2 Sec. I
Compression Of Joint Capsule After Electrocoagulotion
on off(b) . 10gO,,#
2 Sec.Fig. 10. Evocation of articular reflex responses in the leg muscles by directmechanical stress applied to the capsule of the ipsilateral ankle joint. Bothelectromyograms were recorded from the same tenotomized muscle (gastrocnemius).(a) Rapidly and slowly adapting motor unit discharges provoked by direct gentlecompression of the capsule of the ipsilateral ankle joint (maximal posteriorly).Note that the pattern of discharge is similar to that evoked in the gastrocnemiusmuscle when the posterior capsule of the ankle joint is stretched by passive dorsi-flexion of the foot (see Fig. 8b). (b) Following electrocoagulation of the jointcapsule, similar compression no longer evokes any motor unit responses. (Theincreased amplitude of the activity throughout record (b) is due to the fact thatgeneral anaesthesia was allowed to decrease between (a) and (b), so that even slight
reflex effects could be detected, if present.)
The precise nature of the reflex pathway through which these effects areaccomplished is not yet determined; but a variety of incidental observa-tions that we have made in the course of our experiments-observationsthat I cannot detail here-suggests to us that the articular mechanoreceptor(i.e. the arthrokinetic) reflex pathway is polysynaptic, and that it projectsto gamma (fusimotor) rather than to alpha motoneurones. If this proposalcan be substantiated in our later experiments, then the articular mechano-receptors can be added to the cutaneous mechanoreceptors in the segmentalreflex systems driving the gamma motoneurone loops.
43

BARRY WYKE
In the light of the observations that we have made to date, however-and of the information now available from other laboratories-it seemslikely, then, that the Type I mechanoreceptors contribute reflexly to themaintenance of tone in the muscles of the limbs, spine and jaws at rest,and to the co-ordinated reflex regulation of isotonic and isometric changesin the tone of those muscles when movements are made, or attempted;and in addition, these same receptors contribute to conscious perceptionof static joint position and of dynamic joint movement-that is to say, tokinaesthesis. In contrast, the Type II and Type III mechanoreceptors arereflexogenic only, the former provoking transient " booster" changes inthe tone of the muscles operating over a joint during the actual period ofacceleration or deceleration of the articulated bones, whilst the latter actas " brakes " to limit excessive joint displacement.
THE CLINICAL SIGNIFICANCE OF ARTHROKINETIC REFLEXESIt may rightly be argued, however, that whilst these observations may be
pertinent to the experimental situations on which they are based, they donot provide definitive evidence that arthrokinetic reflexes are of practicalsignificance in the circumstances of everyday life. In order to investigatetheir possible significance in this respect, then, we have undertaken afurther set of experimental and clinical investigations, of which I shouldlike to show two examples as a conclusion to this lecture.
If the functional proposals I have made thus far are to be regarded asapplicable to the normal living subject, it should follow that interruptionof the flow of impulses from the mechanoreceptors in a joint capsule intothe central nervous system should result in clinically evident disturbancesof perception of joint position and movement, and of re1texes of postureand gait. That this is indeed so can be shown in two ways.
First, the centripetal flow of such mechanoreceptor afferent impulses canbe interrupted by surgical denervation of a particular joint in animals, andthe subsequent behaviour of the animals can be studied for prolongedperiods, in recovery experiments. Secondly, in animals and in man, it ispossible to demonstrate, by appropriate methods of testing, that destruc-tion of receptor endings in the capsule of a joint gives rise to persistingabnormalities of kinaesthesis and of postural reflexes.
The effects of articular neurectomyThe first of these situations has been examined by carrying out aseptic
partial neurectomy of one knee joint in a series of cats, followed by studiesof the postural reflexes and behaviour of the experimental animals there-after, for periods of up to two years.
In some of these animals, the posterior articular nerve to the left kneejoint was divided in the popliteal fossa; and in a second series of animals,the medial articular nerve to the left knee joint was divided in the front o,the thigh.
44

THE NEUROLOGY OF JOINTS
The resulting abnormalities of postural reflex behaviour that persistafter these two operations, and which are not demonstrable in controlanimals subjected to comparable dummy operations, are summarized inTable III. From this Table, in which plus symbols indicate the severity ofthe post-operative disturbances, you can see that both posterior and medialarticular neurectomy result in significant and persistent changes in certainaspects of postural reflex behaviour. I do not propose to go into thedetailed analysis of these findings here (see Freeman and Wyke, 1966); in-stead, I should simply like to show you two examples of the abnormalreflexes that develop following this type of articular neurectomy.
TABLE IIIREFLEX EFFECTS OF ARTICULAR NEURECTOMY
Behaviour Posterior Medialneurectomy neurectomy
Stance .. + 0Reaching .. o oGait .. .. + +Jumping .. + +Bar walking.. ++ +
Posterior neurectomy Medial neurectomyReflex Ipsi- Contra- Ipsi- Contra-
lateral lateral lateral lateralPreening .. .. o o o oKnee-jerks .. .. o o o oWithdrawal .. o o o oSuspension .. ++ o + oStutz.. .. .. +++ o +Placing .. ..+++ o +++ oCrossed extensor . . o - + +
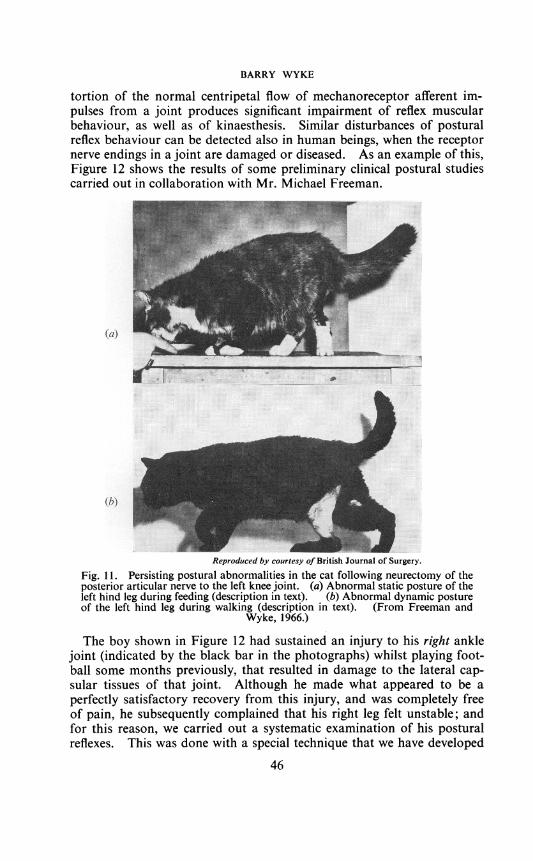
Figure 11 depicts two cats, in each of which the posterior articular nervesupplying the left knee joint has been divided. You will notice that theanimal in the upper illustration keeps its left hind leg in an abnormalposture while eating-the limb being externally rotated, and the kneeflexed, to an extent greater than in the opposite normal leg. Likewise, inthe animal in the lower illustration, you can see that the posture of theleft leg when the animal is walking is abnormal-the whole of the footbeing placed on the walking surface, instead ofjust the toe pads. Further-more, as the data in Table III indicate, these animals are generally un-steady in their gait, and in jumping; and furthermore, they clearly behaveas if they had a persisting kinaesthetic sensory deficit related to the lefthind leg-in that when walking, for instance, they frequently stop andshake the limb, and turn round to look at it.*
The effects of articular receptor damageIn our view, this series of experiments provides clear evidence that dis-* In relation to the above observations, it should be noted that none of these animals
developed pathological changes in the structure of the articular surfaces, or of theconnective tissues of the joint capsule-although these were sought when the animalswere destroyed at various intervals after operation.
45
BARRY WYKE
tortion of the normal centripetal flow of mechanoreceptor afferent im-pulses from a joint produces significant impairment of reflex muscularbehaviour, as well as of kinaesthesis. Similar disturbances of posturalreflex behaviour can be detected also in human beings, when the receptornerve endings in a joint are damaged or diseased. As an example of this,Figure 12 shows the results of some preliminary clinical postural studiescarried out in collaboration with Mr. Michael Freeman.
ii: ~~.
(a)
ia...................................................
Reproduced by courtesy of British Journal of Surgery.
Fig. 1 1. Persisting postural abnormalities in the cat following neurectomy of theposterior articular nerve to the left knee joint. (a) Abnormal static posture of theleft hind leg during feeding (description in text). (b) Abnormal dynamic postureof the left hind leg during walking (description in text). (From Freeman and
Wyke, 1966.)
The boy shown in Figure 12 had sustained an injury to his right anklejoint (indicated by the black bar in the photographs) whilst playing foot-ball some months previously, that resulted in damage to the lateral cap-sular tissues of that joint. Although he made what appeared to be aperfectly satisfactory recovery from this injury, and was completely freeof pain, he subsequently complained that his right leg felt unstable; andfor this reason, we carried out a systematic examination of his post-uralreflexes. This was done with a special technique that we have developed
46

THE NEUROLOGY OF JOINTS
(a)
V~~~~~~~~~~~~~~~~~* :~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ ..... .... .... .::.......:
(b)i :
Fig. 12. Persisting postural abnormalities in man resulting from disturbance ofarticular reflexes operated from the ankle joint, following injury to the lateralcapsule of the right joint. The two illustrations on the left show the normal posturewhen standing (with the eyes closed) on the unaffected left foot, with the arms atthe sides (a) and elevated (b). The two illustrations on the right show the differ-ences when standing on the affected right foot. Partial description in text: inaddition, note (i) Depression of shoulders when standing on right foot, indicatedby a decrease of 100 in the obtuse angle between the shoulder tangents drawn in onthe left and right of Figure 12a. (ii) Adduction and supination of ipsilateral arm,and extension of ipsilateral fingers, with flexion of contralateral elbow and fingers,when standing on right foot (compare Bl and B2). (iii) External rotation andabduction of contralateral leg, when standing on right foot (compare Cl and C2).(iv) External rotation of ipsilateral leg, and flexion of ipsilateral toes, when standingon right foot (compare Dl and D2). (v) Partial extension of contralateral knee, andabduction of contralateral thigh, when standing on right foot (compare El and E2).(vi) Abduction of ipsilateral outstretched arm, and adduction and elevation ofcontralateral arm, when standing on right foot (compare Xl and Y2; Yl and X2).
47

BARRY WYKE
for this purpose in our laboratory, in which postural electromyography iscombined with electronic flash photography of the patient during assump-tion of a variety of standardized postures.As you can see on the left of Figures 12a and 12b, when the boy stands on
his normal leg with his eyes closed, and his arms at his sides (Fig. 12a) orelevated (Fig. 12b), the vertical axis of his body (represented by the dottedline A1) is very close to the true vertical axis. The other interrupted linerepresents the axis of his supporting leg; and I would like to draw yourattention particularly to the angle (of 6°) that this supporting axis normallymakes with the body axis. On the other hand, when the boy stands onthe right foot-the side that had been injured-you will notice (on theright of Figures 12a and 12b) that the body axis (A2) tilts away from thetrue vertical axis (by some 70 in fact) and moves towards the axis of thesupporting leg, so that the normal 6° angle between the body axis and theaxis of the supporting leg is reduced (for instance, by 50 per cent inFig. 12a). At the same time, the axis of the supporting leg deviates awayfrom its normal angle with the true vertical axis, by about 3°.
In addition to these particular disturbances, there are a number of otherpostural reflex changes to be seen in this boy when he is standing on hisaffected leg, some of which are indicated by the numbered arrows in theillustrations and noted in the legend to the figure; but I cannot analysethem further here. I simply wish to show this case, in order to lend weightto the hypothesis advanced earlier-namely, that when injury to or diseaseof a joint is such as to cause degeneration of the nerve endings in itstissues, then this may result in postural reflex abnormalities (as well askinaesthetic disturbances) that are of clinical significance.
CONCLUSIONTaken in conjunction with the animal experiments that I have described
to you, and in the light of work that has been done in other laboratories todate, these observations can leave little doubt that the mechanoreceptorslocated in the fibrous capsules, fat pads and ligaments ofjoints are of con-siderable importance in the circumstances of everyday life-not only inrespect of their contribution to perceptual awareness of static joint positionand of joint movement, but also in the reciprocal reflex regulation ofmuscle tone in posture and movement. In the face of this evidence, Iwould suggest that serious attention must now be given by clinicians-particularly by orthopaedic and neurological surgeons, and by practitionersof physical medicine-to the role of articular reflexes in the regulation ofnormal and abnormal muscle tone, in addition to the role of the articularreceptors in kinaesthetic perception. I hope you will regard it as reason-able to assert that articular reflexes must now assume a position of greaterimportance in the physiology and pathology of movement at every joint inthe body than has been the case in the past; and that in many branches ofsurgery-ranging from otology and laryngology, through orthodontics, to
48

THE NEUROLOGY OF JOINTS
orthopaedic and neurological surgery-the arthrokinetic reflexes must beconsidered as matters of moment in clinical practice. Certainly, a greatdeal still remains to be done in this field, especially in defining more pre-cisely the contribution of disordered articular reflexes to particular clinicalabnormalities of posture and movement in the various joint systems of thebody; but it is my firm belief that sufficient evidence is now available atleast to suggest that such an approach should be made. We hope, by ourcontinued efforts in the Neurological Laboratory of this College, to makesome small contribution to the further development of the subject ofarticular neurology-and I should like to thank the President and Councilof the College for the opportunity of presenting this epitome of our pro-gress to date.
ACKNOWLEDGEMENTSMay I conclude by thanking the Council of the College for the privilege
of delivering this Arris and Gale Lecture. The honour thus conferredshould not, however, be regarded as personal, but rather as a corporatetribute to the work of the Research Fellows whose names are listed inTable IV, the fruits of whose labours in the Neurological Laboratory in
TABLE IVRESEARCH FELLOWS IN THE NEUROLOGICAL LABORATORYDepartment of Applied Physiology, Royal College of Surgeons of EnglandMichael A. R. Freeman, M.A., M.D., F.R.C.S.(Department of Orthopaedic Surgery, Westminster Hospital, London).
John A. Kirchner, M.D., M.S., F.A.C.S.(Department of Otolaryngology, Yale University School of Medicine).
Bernard Greenfield, B.D.S., F.D.S.R.C.S.(University College Hospital School of Dentistry, London).
Brian Reeves, M.B., B.S., F.R.C.S.(Leverhulme Research Fellow in Orthopaedics).
M. A. Abo-El-Enein, M.B., Ch.B., Dip. Anat.(Department of Anatomy, Al-Azhar University, Cairo).
the Department of Applied Physiology I have attempted to summarizein this Lecture. I should also like to take this opportunity of acknowledg-ing publicly the generosity of the College in constructing the speciallydesigned laboratory suite in which our researches are conducted; of thePostgraduate Medical Federation of the University of London for pro-viding a capital grant for our equipment; and of the Camilla SamuelFund for a recent grant to provide supplementary laboratory staff. Last,but by no means least, I should like to pay tribute to the facilities affordedus by Professor David Slome, and to the devoted technical skill of Mr.S. P. Steward* and Mr. Johanna Gitau, without which the work I havedescribed would have been impossible.
* Mr. Steward recently retired from the staff in November 1965, with the remarkablerecord of 53 years of continuous service in the College.
49

BARRY WYKE
REFERENCESMost of these publications are from the Neurological Laboratory, Royal College of
Surgeons ofEngland.ABO-EL-ENEIN, M. A., and WYKE, B. D. (1966) Nature, 209, 683.BARNETT, C. H., DAVIES, D. V., and MACCONAILL, M. A. (1961) Synovial Joints. Their
Structure and Mechanics. London, Longmans Green.COPEMAN, W. S. C. (1963) Ann. Roy. Coll. Surg. Engl. 32, 1.DOBSON, J. (1964) Ann. Roy. Coll. Surg. Engl. 34, 1.FITZGERALD, M. J. T. (1962) J. Anat. Lond. 96, 189.FREEMAN, M. A. R. (1964) Ligamentous Injury. A Study of Injuries to the Lateral
Ligament of the Ankle. M.D. Thesis, University of Cambridge.(1965)Physiotherapy, 51, 393.(1967) Proc. Roy. Soc. Med. (in the press).DEAN, M. R. E., and HANHAM, I. W. F. (1965). J. Bone Jt. Surg
47B, 678.and WYKE, B. D. (1963) J. Anat., Lond. 98, 299 (1964).
(1964a) J. Physiol., Lond. 170, 20P.(1964b) J. Physiol., Lond., 171, 56P.(1965) Nature, 207, 196.(1966) Brit. J. Surg. 53, 61.(1967a) J. Bone. Jt. Surg. 49B, 185.(1967b) J. Anat., Lond. 101, Pt. 3 (in the press).(1967c) Acta anat. Basel. (in the press).(1968) Brit. J. Surg. (in the press).
GOLDSCHEIDER, A. (1898) Physiologie des Muskelainnes. Leipzig, Barth.GREENFIELD, B. E. (1961) J. Bone Jt. Surg. 43B, 396.
and WYKE, B. D. (1963) J. Anat., Lond. 98, 300 (1964).(1966) Nature (in the press).
KIRCHNER, J. A., and WYKE, B. D. (1964a) Nature, 201, 506.(1964b) J. Anat., Lond. 98, 684.(1964c) Nature, 202, 600.(1964d) Nature, 203, 1243.(1965a) Nature, 205, 86.(1965b) Proc. 8th Int. Congr. Otorhinolaryngol.,
Tokyo (Oct.).Tokyo (Oct.).
(1965c) Trans. Ann. Meet. Amer. Laryngol. Assoc.86, 16.
(1965d) Ann. Otol. Rhinol. Laryngol. 74, 749.REEVES, B., and WYKE, B. D. (1967) Nature (to be published).RUFFINI, A. (1905) Rev. gen. d'Hist. 1,420.STACK, H. G. (1963) Ann. Roy. Coll. Surg. Engl. 33, 307.WYKE, B. D. (1961) J. Bone Jt. Surg. 43B, 396.
(1962) Ann. Roy. Coll. Surg. Engl. 30, 135.(1965) In General Anaesthesia. 2nd ed. Vol. 1, p. 157. Ed. by F. T.
Evans and T. C. Gray. London, Butterworth.(1967a) J. Roy. Coll. Surg. Irel. (in the press).(1967b) Principles of General Neurology. Amsterdam, Elsevier (in the
press).(1967c) Brit. J. Dis. Communic. 2, 1.(1967 d) Studies in the Neurology ofJoints. Vol. 1. The Linib and Spinal
Joints. Amsterdam, Elsevier (to be published).and GREENFIELD, B. E. (1967) Studies in the Neurology ofJoints. Vol. III.
The Temporo-mandibular Joints. Amsterdam, Elsevier (to be published).and KIRCHNER, J. A. (1967) Studies in the Neurology of Joints. Vol. II.
The Laryngeal and Ossicular Joints. Amsterdam, Elsevier (to be published).
A comprehensive bibliography of the subject of articular neurology is provided in thereferences appended to these publications.
50




















