
Quantitative and ultrastructural changes in glia and pericytes in the parietal cortex of the aging...
9
Quantitative and Ultrastructural Changes in Glia and Pericytes in the Parietal Cortex of the Aging Rat MARIA A. PEINADO, 1 ADORACION QUESADA, 1 JUAN A. PEDROSA, 1 MARIA I. TORRES, 1 MANUEL MARTINEZ, 1 FRANCISCO J. ESTEBAN, 1 MARIA L. DEL MORAL, 1 RAQUEL HERNANDEZ, 1 JOSE RODRIGO, 1,2 AND JOSE M. PEINADO 1,3 1 Department of Cell Biology, School of Experimental Sciences, University of Jae ´n, E-23071 Jae ´n, Spain 2 Instituto Cajal, Conseso Superior De Investigaciones Cientificas, E-28002 Madrid, Spain 3 Department of Biochemistry, F. Oloriz Institute of Neuroscience, University of Granada, E-18071 Granada, Spain KEY WORDS astrocytes; oligodendrocytes; microglia; pericytes; parietal cortex; aging; rat ABSTRACT The frequency of astrocytes, microglia plus oligodendrocytes, and pericytes display- ing nuclei was analyzed and quantified in 160-μm-wide strips of the parietal cortex (Par1 region) from young and aged Wistar rats. The study was performed on two groups of rats aged 3–4 and 32–36 months. Quantifications of the glial cell types and pericytes were made in 1-μm-thick sections stained with toluidine blue. Ultrathin sections were also made to analyze the ultrastructural features of these cells during aging. Astrocytes and pericytes increased in number by about 20% and 22%, respectively, with age. These increases were most significant in layers II–IV and V for both cellular types. Clusters of astrocytes were common in these layers of aging rats. The ultrastructural analysis also indicated changes in all cell types that stored inclusions and vacuoles with age, which were particularly abundant in microglial cells. End-feet astrocytes and pericytes surrounding the vascular wall also contained vacuoles and inclusions, and consequently the vascular wall increased in thickness. In conclusion, the aging process increased astrocyte and pericyte populations, but not microglia plus oligodendrocyte populations, in the rat parietal cortex. Although no significant change in nuclear size could be observed in any cell type, all glial cells as well as pericytes underwent morphological ultrastructural changes. These modifications may result from the need to correct possible homeostatic imbalances during aging. Microsc. Res. Tech. 43:34–42, 1998. r 1998 Wiley-Liss, Inc. INTRODUCTION Aging of the brain is characterized by a complex process that involves changes in several systems and structures (Amaducci and Tesco, 1994; Goldman et al., 1994). In addition to numerous neurochemical findings, aging is associated with a variety of morphological alterations in neuronal and glial populations, which reflect the neurodegeneration underlying this process. Neuronal cell loss (Amenta et al., 1994; Flood and Coleman, 1988), a decrease in the density of dendritic spines (Vaughan and Peters, 1974), glial hypertrophy and proliferation (Bjo ¨ rklund et al., 1985; Lolova, 1991), deposits of lipofuscin or amyloid proteins (Brizzee et al., 1976; Thal and Schlote, 1995), and vacuolar degenera- tion (Vincent et al., 1989) are among the many alter- ations described. The intensity of these changes varies with the area studied (Haug et al., 1983; Sturrock, 1980, 1990), animal used (Morgan et al., 1987), and methods of study (Haug et al., 1984). Studies of age-related changes in the glial cell population of the cerebral cortex have yielded conflicting results. In humans, either an in- crease in total glial cells in the cerebral cortex of aged subjects (Hansen et al., 1987; Terry et al., 1987) or no changes in this area (Haug et al., 1984) have been described. An increase in glial cells was found in monkeys (Brizzee, 1975; Peters et al., 1994) and mice (Heumann and Leuba, 1983). In the rat cerebral cortex, the astrocytic population seems to increase (Amenta et al., 1994), while other authors (Diamond et al., 1977) found no changes in astrocytes or oligodendrocytes. On the other hand, astrocytes in the rat cortex increase in cytoplasmic area and develop thick cell processes (Bjo ¨rklund et al., 1985). In our laboratory (Peinado et al., 1993, 1997), increases in the total glial population in the frontal and parietal cortices of the rat brain were described. However, the relative participa- tion of the different glial-cell subtypes in these age- related changes has not yet been elucidated. In the present study we analyzed, in semithin sec- tions, the differential contribution of astrocytes, microg- lia plus oligodendrocytes, and pericytes to the quantita- tive changes associated with aging. Glia were studied in 160-μm-wide cortical strips from the parietal cortex (Par1) of the rat. We also analyzed the ultrastructure of these glial-cell types in ultrathin sections taken from the same cortical levels of the cortex. MATERIALS AND METHODS Two groups of male albino Wistar rats aged 3–4 months (n 5 5; 269 6 27 g body weight) and 32–36 Contract grant sponsor: Direccion ´ General en Ciencia y Tecuologı ´a. Contract grant number: PM 90–0146; Contract grant sponsor: Junta de Andalucia. *Correspondence to: M.A. Peinado, Department of Cell Biology, University of Jae ´ n, Paraje Las Lagunillas s/n, E-23071 Jae ´ n, Spain. E-mail: [email protected] Received 5 June 1998; Accepted in revised form 21 July 1998 MICROSCOPY RESEARCH AND TECHNIQUE 43:34–42 (1998) r 1998 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Quantitative and ultrastructural changes in glia and pericytes in the parietal cortex of the aging...

Quantitative and Ultrastructural Changes in Glia and Pericytesin the Parietal Cortex of the Aging RatMARIA A. PEINADO,1 ADORACION QUESADA,1 JUAN A. PEDROSA,1 MARIA I. TORRES,1 MANUEL MARTINEZ,1FRANCISCO J. ESTEBAN,1 MARIA L. DEL MORAL,1 RAQUEL HERNANDEZ,1 JOSE RODRIGO,1,2
AND JOSE M. PEINADO1,3
1Department of Cell Biology, School of Experimental Sciences, University of Jaen, E-23071 Jaen, Spain2Instituto Cajal, Conseso Superior De Investigaciones Cientificas, E-28002 Madrid, Spain3Department of Biochemistry, F. Oloriz Institute of Neuroscience, University of Granada, E-18071 Granada, Spain
KEY WORDS astrocytes; oligodendrocytes; microglia; pericytes; parietal cortex; aging; rat
ABSTRACT The frequency of astrocytes, microglia plus oligodendrocytes, and pericytes display-ing nuclei was analyzed and quantified in 160-µm-wide strips of the parietal cortex (Par1 region)from young and aged Wistar rats. The study was performed on two groups of rats aged 3–4 and32–36 months. Quantifications of the glial cell types and pericytes were made in 1-µm-thick sectionsstained with toluidine blue. Ultrathin sections were also made to analyze the ultrastructuralfeatures of these cells during aging. Astrocytes and pericytes increased in number by about 20% and22%, respectively, with age. These increases were most significant in layers II–IV and V for bothcellular types. Clusters of astrocytes were common in these layers of aging rats. The ultrastructuralanalysis also indicated changes in all cell types that stored inclusions and vacuoles with age, whichwere particularly abundant in microglial cells. End-feet astrocytes and pericytes surrounding thevascular wall also contained vacuoles and inclusions, and consequently the vascular wall increasedin thickness. In conclusion, the aging process increased astrocyte and pericyte populations, but notmicroglia plus oligodendrocyte populations, in the rat parietal cortex. Although no significantchange in nuclear size could be observed in any cell type, all glial cells as well as pericytesunderwent morphological ultrastructural changes. These modifications may result from theneed to correct possible homeostatic imbalances during aging. Microsc. Res. Tech. 43:34–42,1998. r 1998 Wiley-Liss, Inc.
INTRODUCTIONAging of the brain is characterized by a complex
process that involves changes in several systems andstructures (Amaducci and Tesco, 1994; Goldman et al.,1994). In addition to numerous neurochemical findings,aging is associated with a variety of morphologicalalterations in neuronal and glial populations, whichreflect the neurodegeneration underlying this process.Neuronal cell loss (Amenta et al., 1994; Flood andColeman, 1988), a decrease in the density of dendriticspines (Vaughan and Peters, 1974), glial hypertrophyand proliferation (Bjorklund et al., 1985; Lolova, 1991),deposits of lipofuscin or amyloid proteins (Brizzee et al.,1976; Thal and Schlote, 1995), and vacuolar degenera-tion (Vincent et al., 1989) are among the many alter-ations described.
The intensity of these changes varies with the areastudied (Haug et al., 1983; Sturrock, 1980, 1990),animal used (Morgan et al., 1987), and methods ofstudy (Haug et al., 1984). Studies of age-related changesin the glial cell population of the cerebral cortex haveyielded conflicting results. In humans, either an in-crease in total glial cells in the cerebral cortex of agedsubjects (Hansen et al., 1987; Terry et al., 1987) or nochanges in this area (Haug et al., 1984) have beendescribed. An increase in glial cells was found inmonkeys (Brizzee, 1975; Peters et al., 1994) and mice(Heumann and Leuba, 1983). In the rat cerebral cortex,
the astrocytic population seems to increase (Amenta etal., 1994), while other authors (Diamond et al., 1977)found no changes in astrocytes or oligodendrocytes.
On the other hand, astrocytes in the rat cortexincrease in cytoplasmic area and develop thick cellprocesses (Bjorklund et al., 1985). In our laboratory(Peinado et al., 1993, 1997), increases in the total glialpopulation in the frontal and parietal cortices of the ratbrain were described. However, the relative participa-tion of the different glial-cell subtypes in these age-related changes has not yet been elucidated.
In the present study we analyzed, in semithin sec-tions, the differential contribution of astrocytes, microg-lia plus oligodendrocytes, and pericytes to the quantita-tive changes associated with aging. Glia were studiedin 160-µm-wide cortical strips from the parietal cortex(Par1) of the rat. We also analyzed the ultrastructure ofthese glial-cell types in ultrathin sections taken fromthe same cortical levels of the cortex.
MATERIALS AND METHODSTwo groups of male albino Wistar rats aged 3–4
months (n 5 5; 269 6 27 g body weight) and 32–36
Contract grant sponsor: Direccion General en Ciencia y Tecuologıa. Contractgrant number: PM 90–0146; Contract grant sponsor: Junta de Andalucia.
*Correspondence to: M.A. Peinado, Department of Cell Biology, University ofJaen, Paraje Las Lagunillas s/n, E-23071 Jaen, Spain. E-mail: [email protected]
Received 5 June 1998; Accepted in revised form 21 July 1998
MICROSCOPY RESEARCH AND TECHNIQUE 43:34–42 (1998)
r 1998 WILEY-LISS, INC.

months (n 5 5; 534 6 23 g body weight) were used inthis study. The animals were caged under a 12-hour/12-hour light/dark cycle, with room temperature at 20–22°C, and with water and standard food pellets freelyavailable. The animal were housed, supervised, andhandled according to the approved national guidelinesfor animal care.
Rats were anesthetized intraperitoneally with Equith-esin (2 ml/kg body weight) and perfused through theascending aorta with phosphate-buffered saline (PBS),pH 7.4, followed by a fixative containing 4% paraformal-dehyde and 0.5% glutaraldehyde in PBS. The brainswere removed and postfixed by immersion in 4% para-formaldehyde in the same buffer for 4 hours at roomtemperature. After that the brains were sectionedtransversely with a vibratome (Vibracut FTB, Leica,
Barcelona, Spain). A 1-mm-thick slice taken at the levelof the optic chiasma (corresponding to interaural coordi-nates 9.7–7.7 from Paxinos and Watson, 1986) was selectedfor quantification. From this slice (Fig. 1), a 1.5-mm-wideblock was carefully obtained, which included all corticallayers from the Par1 area of the right hemisphere cortex(Zilles and Wree, 1985). These blocks of cortex were im-mersed in osmium, dehydrated in ascending ethanol se-ries, immersed in propylene oxide, and embedded inepoxy resin. They were then sectioned with an ultrami-crotome (Reichert-Jung Leica, Barcelona, Spain) to obtainserial 1-µm-thick sections positioned in a plane perpen-dicular to the pial surface and in a plane parallel to thelength of the apical dendrites of the pyramidal neurons.
The first sections from this series were discarded,and six equidistant and uniformly sampled sections (1
Fig. 1. Schematic illustration of the procedure used to obtain the semithin sections and to performcounts in the Par1 cortex.
35AGING GLIAL CHANGES IN PARIETAL CORTEX

µm thick, 50 µm apart) were taken for quantitativestudies. The remainder of the blocks were used toobtain ultrathin sections, which were contrasted withuranyl acetate and lead citrate; these sections, used inultrastructural studies, were examined with a PhilipsEM300 electron microscope (Philips, Madrid, Spain).
The semithin sections (Fig. 1) were stained withtoluidine blue and observed with a 1003 objective and a103 eyepiece fitted with a micrometer grid of 160 3 80µm (12,800 µm2); the dimensions of the grid were usedas the count unit (CU). The upper edge of the grid wasplaced at the pial surface and lowered successivelyalong 20 CU, which involved the entire cortical thick-ness at the level of the Par1 area. Only cells interceptedby the right vertical and top grid bars were included inthe counts; those intercepted by the left vertical andbottom bar were not. This procedure was repeatedalong the 160-µm-wide cortical strips from the pialsurface to the white matter in the six semithin sectionsfrom each animal. Each time, all glial nuclear profilesin the different CU were drawn with a camera lucida(O’Kusky and Colonnier, 1982). This procedure allowedus to quantify not only the mean number of thedifferent glial types in the cortical strip, but also thedensity distribution of glial cells (mean number of cellsin each CU) across the different cortical layers. Inaddition, the area of a significant number of nuclearprofiles from each cellular type was systematicallymeasured with the aid of a Videoplan Kontron analyzer(Zeiss, Madrid, Spain).
The cell nucleus was considered the test object (Chowand Dewson, 1966; O’Kusky and Colonnier, 1982) forcounting. Glial cells were identified according to thestandard criteria of Ling and Leblond (1973), as modi-fied by Sturrock (1983) and Peters et al. (1991), basedon nuclear chromatin patterns and the texture anddarkness of the cytoplasm. To avoid possible mistakesin identifying microglial cells and oligodendrocytes,these cell types were quantified as a single type.
Statistical analyses were performed by a computer-ized Kolmogorov-Smirnov test to determine whetherthe data followed a normal distribution. Because thistest refuted the hypothesis of a normal distribution, theresults in both age groups were compared by theMann-Whitney U-test.
RESULTSThe results obtained (mean 6 SEM) for glial cell
number and nuclear size are summarized in Figures 2and 3. Figure 2 shows the number (mean 6 SEM) ofastrocytes, microglial cells plus oligondredocytes, andpericytes counted in the cortical 160-µm-wide strips ofthe Par1 region in young and aged rats. Aging affectedonly astrocyte and pericyte populations, which showed20% (P , 0.01) and 22% (P , 0.01) increases respec-tively. Figure 3 shows the nuclear area (mean 6 SEM)of the different cell types; this cytomorphometric param-eter was not significantly affected by aging. This fact, aswell as the large number of nuclear profiles analyzed,supported the choice of the nucleus as test object(Weibel, 1979).
Distributions of the different glial-cell types andpericytes through the thickness of the cortex are shownin Figure 4. Only astrocytes and pericytes showedsignificant differences in the two age groups. Aging
increased astrocyte numbers, mainly in CU levels 4, 9,and 10, located in layers II–IV and V. The greatestincrease in pericyte numbers with aging was observedat CU levels 7 and 14, also included in layers II–IVand V.
Ultrastructural analyses revealed differences be-tween glial cells from young and aged rats. All glial-celltypes in aging rats showed cytoplasmic inclusions,although the number of these inclusions was significantonly in microglial cells. Astrocytes in aging rats hadoval or round nuclei with a typical chromatin pattern,consisting of an irregular ring beneath the nuclearenvelope and evenly dispersed accumulations in theinterior of the nucleus (Fig. 5A). The perinuclear cyto-plasmic region contained normal organelles, and there
Fig. 2. Number of nuclear profiles of astrocytes, microglial cellsplus oligodendrocytes, and pericytes (mean 6 SEM) in cortical 160-µm-wide strips of Par1 region in young (n 5 30 sections) and aged (n 5 30sections) rats. **P , 0.01.
Fig. 3. Nuclear area (mean 6 SEM) of astrocytes, microglial cellsplus oligodendrocytes, and pericytes of Par1 region in young (astro-cytes, n 5 200; microglia plus oligodendrocytes, n 5 100; pericytes, n 550) and aged (astrocytes, n 5 200; microglia plus oligodendrocytes, n 5100; pericytes, n 5 50) rats. Units, µm2.
36 M.A. PEINADO ET AL.

were no noteworthy differences from young animals,except for a greater dispersion and dilation of the Golgicomplex vesicles (Fig. 5B) as well as some dispersedareas of vacuolation. These vacuolated areas containeda few (2–5) pale inclusions that appeared to be empty(Fig. 5A).
Oligodendrocytes showed their characteristic struc-ture with an irregular, dense pattern of chromatin anddark cytoplasm. In aging rats some of these cells haddense, irregular inclusions (Fig. 6A).
In ultrastructural terms, microglial cells were themost strongly affected by aging: all of them accumu-lated many different types of inclusions in the cyto-plasm. These cells contained a few lysosomes andphagolysosomes even in young control animals; withaging, because of their phagocytic funtion, their cyto-plasm became filled with debris that in some casesdisplaced the nucleus (Fig. 6B,C).
Pericytes were not considered to be glial cells. Theywere located within the perivascular space surroundedby the basal lamina and the end-feet of astrocytes.Their characteristics coincided with those described inthe brains of other animals (Peters et al., 1991, 1994),although in the rat cortex their nucleus containeddispersed clumps of chromatin, mainly distributed atthe periphery. In aged rats, some pericytes bore pro-cesses containing inclusions of different textures (Fig.7A).
The vascular wall was hardly affected by aging,although vacuolar degenerated zones surrounding thewall were frequently seen (Fig. 7A). These zones wereabsent in the vascular wall of the young rat (Fig. 7B).The pericytes accumulated inclusions with age, andendothelial cells also acquired inclusions, althoughthese were fewer in number. In addition, we detectedpale vacuolated zones outside the basal lamina. Thesezones were probably located inside end-feet of theastrocytes, because they were very similar to those seenat perinuclear levels, although present in greater num-bers. This accumulation caused hypertrophy of thepericytes and astrocyte end-feet; consequently, the vas-cular wall appeared thicker in aging rats.
DISCUSSIONWe analyzed the possible influence of aging on the
number and structural features of different glial cells inthe parietal cortex (Par1) of the rat (Zilles and Wree,1985). The results show that aging produces an in-crease in the number of astrocytes and pericytes. Inaddition, all glial-cell types displayed a large number ofcytoplasmic inclusions, although no cell type showed asignificant change in nuclear size.
Previous studies in our laboratory demonstrated thatthe total glial population increases during aging in thefrontal (Peinado et al., 1993) and parietal (Peinado etal., 1997) cortex. Although the method used in those
Fig. 4. Number of astrocytes, microglial cells plus oligodendro-cytes, and pericytes (mean 6 SEM) obtained in each of the 20 countunits (CU) in the cortex. Each graph shows the distributions of nuclearprofiles of astrocytes, microglia plus oligodendrocytes, and pericytesacross the cortical thickness in young (n 5 30 sections) and aged (n 530 sections) rats. *P , 0.05; **P , 0.01. I, II–IV, V, and VI signifycortical layers.
37AGING GLIAL CHANGES IN PARIETAL CORTEX

Fig. 5. Electron micrographs of the Par1 cortex from a 32-month-old rat. A: Two astrocytes (AS) and a neuron (N). Note the typicalpattern of chromatin in the astrocyte nucleus and some pale vacuoles(V) in the perinuclear cytoplasm. Also note the presence of lipofuscin
granules (L) in the cytoplasm of the neuron (35,400). B: A zonebelonging to the cytoplasm of two astrocytes. Note the distribution andspread of the Golgi vesicles (G) and the characteristics of the remain-ing cytoplasmic components (340,500).
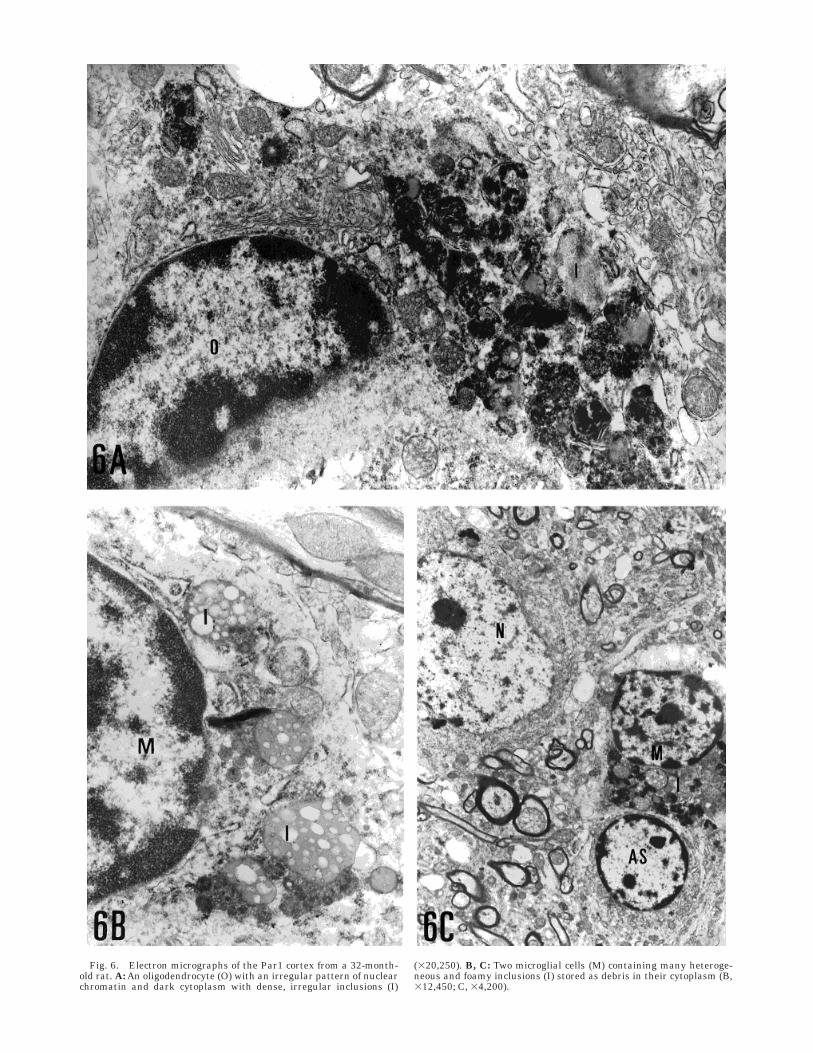
Fig. 6. Electron micrographs of the Par1 cortex from a 32-month-old rat. A: An oligodendrocyte (O) with an irregular pattern of nuclearchromatin and dark cytoplasm with dense, irregular inclusions (I)
(320,250). B, C: Two microglial cells (M) containing many heteroge-neous and foamy inclusions (I) stored as debris in their cytoplasm (B,312,450; C, 34,200).

Fig. 7. Electron micrographs of the Par1 cortex from a 32-month-old rat (A) and a 3-month-old rat (B) showing two blood vessels (BV).The blood vessel in A shows some zones of vacuolar degeneration (VD)that are absent from the vessel in B. In A, note a portion of a pericyte(P) filled with different inclusions and vacuoles within the perivascu-
lar space and surrounded by the basal lamina (arrowheads). B showsanother pericyte (P), with a clearly visible nucleus and a thin processextending from the soma between the endothelial and basal lamina. E,endothelial cell; AS, astrocyte; N, neuron (A, 36,500; B, 36,300).

studies did not allow us to distinguish between differ-ent glial-cell types, the current results are in agreementwith the earlier findings.
Our data are also consistent with the findings ofother authors, who described glial increases with agingof the human cortex (Terry et al., 1987), monkey cortex(Peters et al., 1994), and rat cortex (Brizzee, 1975;Brizzee et al., 1968; Landfield et al., 1977; Peters et al.,1991). However, exactly which glial-cell types are in-volved in these changes remains controversial.
Our results support the idea that only astrocytessignificantly increase in number with age in the Par1cortex. Peters et al. (1994) found that in the frontalcortex of old rhesus monkeys, the astrocyte populationdid not change in number with age, although theseauthors noted a tendency toward a slight increase inthe number of neuroglial cells with dark cytoplasm(oligodendrocytes and microglial cells). In previousstudies (Peters et al., 1991), these authors also indi-cated that only microglia increased in number with age.In relation to ultrastructural features, they found, asalso described here, that all glial-cell types accumu-lated inclusions in their cytoplasm, especially micro-glial cells. In addition, they found an increase inastrocyte filaments, particularly in the outer glial-limiting membrane, where astrocyte processes prolifer-ate with age. Another ultrastructural study that quan-tified astrocytes, oligodendrocytes, and microgliaseparately in the cerebral auditory cortex (Vaughanand Peters, 1974) found that only microglia increasedin number with age.
Increases with age in the size of the soma and theprocesses of glial cells have been reported in studieswith antibodies against glial fibrillary acidic protein(GFAP) (Berciano et al., 1995; Bjorklund et al. 1985); inaddition, the population of astrocytes was found toincrease with age in the rat cortex (Amenta et al., 1994)and in the human cortex (Hansen et al., 1987). Thesechanges may be correlated with increases in glucoseuptake (Shimada et al., 1992), as well as in the activityof enzymes such as glutamine synthetase (Cao et al.,1985; Kanai et al., 1994; Peinado and Myers, 1988) andmonoamine oxidase B (Amenta et al., 1994), bothmarkers of glial function.
In relation to the cell distribution throughout thethickness of the cortex, the astrocytes in aged rats weremore numerous in all cortical layers, although thesedifferences were significant only in layers II–IV and V.These data also confirm the frequent images of pairedor clustered astrocytes found in these layers in semi-thin and ultrathin sections. It is noteworthy thatpericytes also showed significant increases in layersII–IV and V.
Astrocytes have been proposed to act as protectiveagents of neurons (Norenberg, 1994). In fact, these glialcells act as buffers of ions released by neurons duringelectrical activity (Tardy, 1991), and inactivate amino-acid neurotransmitters (Kanai et al., 1994; Worral andWilliams, 1994), store glycogen (Magistretti, 1988), andpossibly release neurotrophic factors that support neu-ronal function and survival (Hatten, 1990; Schwartzand Mishler, 1990; Shao and McCarthy, 1994). Thus,astrocyte proliferation can be interpreted as a conse-quence of increasing neuronal protection of the agingbrain. On the other hand, it has recently been demon-
strated that hypoxia produces pericyte proliferation(Nomura et al., 1995). In this context, the increasednumber of pericytes reported here could be interpretedas a possible consequence of hypoperfusion.
In relation to oligodendrocytes and microglial cells,our data show stability with age in the different corticallayers. In contrast with our resuls, studies in monkeys(Vaughan and Peters, 1974; Peters et al., 1991, 1994)showed an increase with age in oligodendrocytes andmicroglia, but not in astrocytes or pericytes. Otherstudies, however, reported no change in the microglialpopulation with age in the mouse brain (Lawson et al.,1992). In addition, Ogura et al. (1994), using OX-42 andOX-6 antibodies, showed that these cells do not changein number, although aging induces their transforma-tion into reactive cells. This was also found by Perry etal. (1993), who detected MCH-II antigen in some micro-glial cells in the brains of aged rodents.
In summary, we have shown that in the parietalcortex of the aging rat, there is a significant increase inthe number of astrocytes and pericytes. This increasemay be a consequence of neuronal damage produced bydifferent insults that underlie the aging process, suchas hypoxia, hypoglycemia, or increases in amino-acidrelease (Finch, 1993; Knight, 1995; Kowald and Kirk-wood, 1994).
ACKNOWLEDGMENTSThe authors are indebted to Dr. Jose Rodriguez Avi
for his invaluable help in the statistical study, and toMs. M. Angeles Cruz for revising the English usage ofthe manuscript.
REFERENCESAmaducci, L., and Tesco, G. (1994) Aging as a major risk for degenera-
tive diseases of the CNS. Curr. Opin. Neurol., 7:283–286.Amenta, F., Bograni, S., Cadel, S., Ferrante, F., Valsecchi, B., and
Vega, J.A. (1994) Microanatomical changes in the frontal cortex ofaged rats: Effect of L-deprenyl treatment. Brain Res. Bull., 34:125–131.
Berciano, M.T., Andres, M.A., Calle, E., and Lafarga, M. (1995)Age-induced hypertrophy of astrocytes in rat supraoptic nucleus: Acytological, morphometric, and immunocytochemical study. Anat.Rec., 243:129–144.
Bjorklund, H., Eriksdotter-Nilsson, M., Dahl, D., Rose, G., Hoffer, B.,and Olson, L. (1985) Image analysis of GFAP-positive astrocytesfrom adolescence to senescence. Exp. Brain Res., 58:163–170.
Brizzee, K.R. (1975) Gross-morphometric analyses and quantitativehistology of the aging brain. In: Neurobiology of Aging. J.M. Ordyand K.R. Brizzee, eds. Plenum Press, New York, pp. 401–423.
Brizzee, K.R., Sherwood, N., and Timiras, P.S. (1968) Comparison ofcell populations at various depth levels in cerebral cortex of youngadult and aged Long-Evans rats. J. Gerontol., 23:289–297.
Brizzee, K.R., Ordy, J.M., Hansche, J., and Kaack, B. (1976) Quantita-tive assessment of changes in neurons and glial cell packing densityand lipofuscin accumulation with age in the cerebral cortex of anon-human primate (Macaca mulatta). In: Neurobiology of Aging.R.D. Terry and S. Gershon, eds. Raven Press, New York, pp.229–244.
Cao, D.H., Strolin, M., and Dostert, P. (1985) Age-related changes inglutamine synthetase activity of rat brain, liver, and heart. Gerontol-ogy, 31:95–100.
Chow, K.L., and Dewson, J.M. (1966) Numerical estimation of neuronsand glia in lateral geniculate body during retrograde degeneration.J. Comp. Neurol., 128:63–74.
Diamond, M.C., Johnson, R.E., and Gold, M.W. (1977) Changes inneuron number and size and glial number in the young, adult andageing rat medial occipital cortex. Behav. Biol., 20:409–418.
Finch, C.E. (1993) Neurons atrophy during aging: programmed orsporadic? Trends Neurosci., 16:104–110.
41AGING GLIAL CHANGES IN PARIETAL CORTEX

Flood, D.G., and Coleman, P.D. (1998) Neuron numbers and sizes inaging brain: Comparisons of human, monkey and rodent data.Neurobiol. Aging, 9:453–463.
Goldman, J.E., Calingasan, N.Y., and Gibson, G.E. (1994) Aging andthe brain. Curr. Opin. Neurol., 7:287–293.
Hansen, L.A., Armstrong, D.M., and Terry, R.D. (1987) An immunohis-tochemical quantification of fibrous astrocytes in the aging humancerebral cortex. Neurobiol. Aging, 8:1–6.
Hatten, M.E. (1990) Riding the glial monorail: A common mechanismfor glial-guided neuronal migration in different regions of thedeveloping mammaliam brain. Trends Neurosci., 13:179–184.
Haug, H., Barmwater, U., Eggerse, R., Fischer, D., Kuhl, S., and Sass,N.L. (1983) Anatomical changes in aging brain: Morphometricanalysis of the human prosencephalon. In: BrainAging, Neuropathol-ogy and Neuropharmacology. J. Cervos-Navarro and H.I. Sar-kander, eds. Raven Press, New York, pp. 1–12.
Haug, H., Kuhl, S., Mecke, E., Sass, N.L., and Wasner, K. (1984) Thesignificance of morphometric procedures in the investigation of agechanges in cytoarchitectonic structures of human brain. Z. Hirn-forsch., 25:353–374.
Heumann, D., and Leuba, G. (1983) Neuronal death in the develop-ment and aging of the cerebral cortex of the mouse. Neuropathol.Appl. Neurobiol., 5:433–456.
Kanai, Y., Smith, C.P., and Hediger, M.A. (1994) A new family ofneurotransmitter transporters: The high-affinity glutamate trans-porters. FASEB J., 8:1450–1459.
Knight, J.A. (1995) The process and theories of aging. Ann. Clin. Lab.Sci., 25:1–11.
Kowald, A., and Kirkwood, T.B.L. (1994) Towards a network of aging: Amodel combining the free radical theory and the protein errortheory. J. Theor. Biol., 168:75–94.
Landfield, P.W., Rose, G., Sandles, L., Wohstadter, T.C., and Lynch, G.(1977) Patterns of astroglial hypertrophy and neuronal degenera-tion in the hippocampus of aged memory deficient rats. J. Gerontol.,32:3–12.
Lawson, L.J., Perry, V.H., and Gordon, S. (1992) Turnover of residentmicroglia in the normal adult mouse brain. Neuroscience, 48:505–415.
Ling, E.A., and Leblond, C.P. (1973) Investigation of glial cells insemithin sections. II. Variation with age in the numbers of thevarious glial cells types in rat cortex and corpus callosum. J. Comp.Neurol., 149:73–82.
Lolova, I. (1991) Qualitative and quantitative glial changes in thehippocampus of aged rats. Anat. Anz., 172:263–271.
Magistretti, P.J. (1988) Regulation of glycogenolysis by neurotransmit-ters in the central nervous system. Diabete Metab., 14:237–246.
Morgan, D.G., May, P.C., and Finch, C.E. (1987) Dopamine andserotonine systems in human and rodent brain: Effects of aging andneurodegenerative diseases. J. Am. Gerontol. Soc., 35:334–345.
Nomura, M., Yamagishi, S., Harada, S., Hayashi, Y., Yamashima, T.,Yamashita, J., and Yamamoto, H. (1995) Possible participation ofautocrine and paracrine vascular endothelial growth factors inhypoxia-induced proliferation of endothelial cells and pericytes. J.Biol. Chem., 270:28316–28324.
Norenberg, M.D. (1994) Astrocyte responses to CNS injury. J. Neuro-pathol. Exp. Neurol., 53:213–220.
Ogura, K., Ogawa, M., and Yoshida, M. (1994) Effects of aging onmicroglia in the normal rat brain: Immunohistochemical observa-tions. Neuroreport, 5:1224–1226.
O’Kusky, J., and Colonnier, M.A. (1982) A laminar analysis of thenumber of neurons, glia and synapses in the visual cortex (area 17)of adult macaque monkeys. J. Comp. Neurol., 210:278–290.
Paxinos, G., and Watson, C. (1986) The Rat Brain in StereotaxicCoordinates. Academic Press, New York, pp: 25–27.
Peinado, J.M., and Myers, R.D. (1988) Cortical amino acid neurotrans-mitter release is altered by CCK perfused in frontal region ofunrestrained aged rat. Peptides, 9:631–636.
Peinado, M.A., Martınez, M., Pedrosa, J.A., Quesada, A., and Peinado,J.M.(1993) Quantitative morphological changes in neurons and gliain the frontal lobe of the aging rat. Anat. Rec., 237:104–108.
Peinado, M.A., Quesada, A. Pedrosa, J.A., Martınez, M., Esteban, F.J.,Del Moral, M.L., and Peinado, J.M. (1997) Light microscopic quanti-fication of morphological changes during aging in neurons and gliaof the rat parietal cortex. Anat. Rec., 247:420–425.
Perry, U.H., Matiszak, M.K., and Feurn, S. (1993) Altered antigenexpression of microglia in the aged rodent CNS. Glia, 7:60–67.
Peters, A., Josephson, K., and Vincent, S.L. (1991) Effects of aging onthe neuroglial cells and pericytes within area 17 of the rhesusmonkeys cerebral cortex. Anat. Rec., 9:384–398.
Peters, A., Leahu, D., Moss, M.B., and McNally, K.J. (1994) The effectsof aging on area-46 of the frontal-cortex of the rhesus monkey.Cereb. Cortex, 6:621–635.
Schwartz, J.P., and Mishler, K. (1990) Beta-adrenergic receptor regula-tion, through cyclic AMP, of nerve growth factor expression in ratcortical and cerebellar astrocytes. Cell. Mol. Neurobiol., 10:447–457.
Shao, Y., and McCarthy, K.D. (1994) Plasticity of astrocytes. Glia,11:147–155.
Shimada, M., Akagi, N., Goto, H., Watanabe, H., Nakanishi, M.,Hirose, Y., and Watanabe, M. (1992) Microvessel and astroglial celldensities in the mouse hippocampus. J. Anat., 180:89–95.
Sturrock, R.R. (1980) Comparative quantitative and morphologicalstudy of aging in the mouse neostriatum, indusium griseum andanterior commissure. Neuropathol. Appl. Neurobiol., 6:415–420.
Sturrock, R.R. (1983) Problems of glial identification and quantifica-tion. In: Brain Aging: Neuropathology and Neuropharmacology. J.Cervos-Navarro and H.I. Sarkander, eds. Raven Press, New York,pp. 179–209.
Sturrock, R.R. (1990) A comparison of age-related changes in neuronnumber in the dorsal motor nucleus of the vagus and the nucleusambiguus of the mouse. J. Anat., 173:169–176.
Tardy, M. (1991) Astrocyte et homeostasie. Med. Sci., 8:799–804.Terry, R.D., DeTeresa, R., Laurence, A., and Hansen, M.D. (1987)
Neocortical cell count in normal human adult aging. Ann. Neurol.,21:530–539.
Thal, D.R., and Schlote, W. (1995) Ultrastructural differences ofhippocampal lipofuscin in human develoment. Mech. Ageing Dev.,79:59–70.
Vaughan, D.W., and Peters, A. (1974) Neuroglial cells in the cerebralcortex of rats from young adulthood age: An electron microscopestudy. J. Neurocytol., 3:405–429.
Vincent, S.L., Peters, A., and Tigges, J. (1989) Effects of aging on theneurons within area 17 of rhesus monkey cerebral cortex. Anat.Rec., 223:329–341.
Weibel, E.R. (1979) Stereological Methods. Volume II. Practical Meth-ods for Biological Morphometry. Academic Press, London, New York,Sydney, and San Francisco, pp. 140–172.
Worral, M., and Williams, C. (1994) Sodium ion-dependent transport-ers for neurotransmitters: A review of recent developments. Bio-chem. J., 297:425–436.
Zilles, K., and Wree, A. (1985) Cortex: areal and laminar structure. In:The Rat Nervous System, Volume 1. G. Paxinos, ed. Academic Press,Sydney, pp. 375–415.
42 M.A. PEINADO ET AL.




















