Impaired Working Memory for Location but not for Colour or Shape in Visual Neglect: a Comparison of...
12
INTRODUCTION A common outcome of acute right hemisphere damage in humans is spatial neglect, in which patients fail to respond to or acknowledge stimuli arising from the contralesional (left) side of space. Neglect may follow damage to a number of different brain areas, but is most common and severe following lesions of the right temporo- parietal region (Karnath et al., 2001; Vallar and Perani, 1986). It is now recognised that neglect is a heterogeneous disorder comprising several component deficits (Barbieri and De Renzi, 1989; Karnath et al., 2002; Vallar, 1998). Although the most striking of these involves a bias of selective attention that favours ipsilesional over more contralesional stimuli (Kinsbourne, 1993), many individuals with neglect also show reduced attentional capacity (Duncan et al., 1999; Karnath, 1988), impaired arousal (Robertson, 1993; Robertson et al., 1998), and a pathological tendency to focus on local details in a scene (Robertson et al., 1988; Halligan and Marshall, 1994; Marshall and Halligan, 1995). Two recent single-case studies have suggested that neglect may also involve an impairment of spatial working memory, which during exploration for visual targets (as in clinical cancellation tests) may cause patients to revisit previously detected targets and treat them as if they have not been seen before (Husain et al., 2001; Wojciulik et al., 2001). If confirmed, a deficit of spatial working memory in neglect would be consistent with findings from animal neurophysiology and human brain imaging, which suggest that regions of the parietal lobe are involved in maintaining stable representations of the locations of visual objects over time, and across changes in eye position (Duhamel et al., 1992; Colby et al., 1996; Gnadt and Andersen, 1988). In the present study, we investigated visual working memory for spatial location, colour and shape in right hemisphere patients with left neglect. The primary aim was to test whether spatial working memory is selectively impaired in neglect, relative to working memory for the visual attributes of colour and shape. A second aim was to compare the performances of patients with parietal versus non-parietal lesions, to determine whether any impairment of spatial working memory is specific to neglect arising from parietal damage. There are several reasons why an impairment of spatial working memory might be predicted in right hemisphere patients with neglect, and particularly in those with lesions of the parietal lobe. First, functional imaging studies in normals have shown that a right-hemisphere network of brain areas, including the posterior parietal and lateral prefrontal cortices, is selectively active when healthy participants are required to memorise the spatial locations of targets over time (Awh et al., 1999; Jonides et al., 1993; Owen et al., 1996). These same right hemisphere areas are also frequently damaged in patients with spatial neglect (Heilman and Valenstein, 1972; Mesulam, 1999; Vallar and Perani, Cortex, (2004) 40, 379-390 SPECIAL ISSUE IMPAIRED WORKING MEMORYFOR LOCATION BUT NOT FOR COLOUR OR SHAPE IN VISUAL NEGLECT: A COMPARISON OF PARIETALAND NON- PARIETAL LESIONS Laure Pisella 1 , Nadja Berberovic 2 and Jason B. Mattingley 2 ( 1 Espace et Action, INSERM U 534, Bron, France; 2 Cognitive Neuroscience Laboratory, Department of Psychology, University of Melbourne, Victoria, Australia) ABSTRACT Patients with spatial neglect due to right hemisphere pathology may show ‘revisiting’ behaviour during visual search and cancellation tasks, such that previously encountered targets are treated as if they are new discoveries. Revisiting behaviour is particularly evident when no visible trace is left to inform patients that a particular target has already been detected (Husain et al., 2001; Wojciulik et al., 2001), implying that spatial working memory may be impaired in neglect. To test whether working memory for location is selectively impaired relative to memory for colour and shape, we compared performances of right hemisphere neglect patients with parietal (n = 4) and non-parietal (n = 4) lesions on a change detection task. Patients were presented with a matrix containing four objects in different positions, and required to detect a change in the location, colour or shape of one of the objects following presentation of a brief visual mask. Parietal patients were selectively impaired in detecting location changes, regardless of the horizontal position of the object in the matrix, relative to colour and shape changes. This deficit of spatial working memory was not apparent for neglect patients with lesions that spared the parietal cortex. We conclude that the human parietal cortex is crucially involved in the updating and maintenance of spatial representations across saccades, and that neglect arising from parietal damage causes impairment in these re-mapping mechanisms. Key words: attention, change detection, parietal lobe, right hemisphere, spatial neglect, spatial working memory
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Impaired Working Memory for Location but not for Colour or Shape in Visual Neglect: a Comparison of...

INTRODUCTION
A common outcome of acute right hemispheredamage in humans is spatial neglect, in whichpatients fail to respond to or acknowledge stimuliarising from the contralesional (left) side of space.Neglect may follow damage to a number ofdifferent brain areas, but is most common andsevere following lesions of the right temporo-parietal region (Karnath et al., 2001; Vallar andPerani, 1986). It is now recognised that neglect is aheterogeneous disorder comprising severalcomponent deficits (Barbieri and De Renzi, 1989;Karnath et al., 2002; Vallar, 1998). Although themost striking of these involves a bias of selectiveattention that favours ipsilesional over morecontralesional stimuli (Kinsbourne, 1993), manyindividuals with neglect also show reducedattentional capacity (Duncan et al., 1999; Karnath,1988), impaired arousal (Robertson, 1993;Robertson et al., 1998), and a pathologicaltendency to focus on local details in a scene(Robertson et al., 1988; Halligan and Marshall,1994; Marshall and Halligan, 1995).
Two recent single-case studies have suggestedthat neglect may also involve an impairment ofspatial working memory, which during explorationfor visual targets (as in clinical cancellation tests)may cause patients to revisit previously detectedtargets and treat them as if they have not been seenbefore (Husain et al., 2001; Wojciulik et al., 2001).If confirmed, a deficit of spatial working memory
in neglect would be consistent with findings fromanimal neurophysiology and human brain imaging,which suggest that regions of the parietal lobe areinvolved in maintaining stable representations ofthe locations of visual objects over time, andacross changes in eye position (Duhamel et al.,1992; Colby et al., 1996; Gnadt and Andersen,1988). In the present study, we investigated visualworking memory for spatial location, colour andshape in right hemisphere patients with left neglect.The primary aim was to test whether spatialworking memory is selectively impaired in neglect,relative to working memory for the visual attributesof colour and shape. A second aim was to comparethe performances of patients with parietal versusnon-parietal lesions, to determine whether anyimpairment of spatial working memory is specificto neglect arising from parietal damage.
There are several reasons why an impairment ofspatial working memory might be predicted in righthemisphere patients with neglect, and particularly inthose with lesions of the parietal lobe. First,functional imaging studies in normals have shownthat a right-hemisphere network of brain areas,including the posterior parietal and lateral prefrontalcortices, is selectively active when healthyparticipants are required to memorise the spatiallocations of targets over time (Awh et al., 1999;Jonides et al., 1993; Owen et al., 1996). These sameright hemisphere areas are also frequently damagedin patients with spatial neglect (Heilman andValenstein, 1972; Mesulam, 1999; Vallar and Perani,
Cortex, (2004) 40, 379-390
SPECIAL ISSUEIMPAIRED WORKING MEMORY FOR LOCATION BUT NOT FOR COLOUR OR
SHAPE IN VISUAL NEGLECT: A COMPARISON OF PARIETAL AND NON-PARIETAL LESIONS
Laure Pisella1, Nadja Berberovic2 and Jason B. Mattingley2
(1Espace et Action, INSERM U 534, Bron, France; 2Cognitive Neuroscience Laboratory, Department of Psychology,University of Melbourne, Victoria, Australia)
ABSTRACT
Patients with spatial neglect due to right hemisphere pathology may show ‘revisiting’ behaviour during visual searchand cancellation tasks, such that previously encountered targets are treated as if they are new discoveries. Revisitingbehaviour is particularly evident when no visible trace is left to inform patients that a particular target has already beendetected (Husain et al., 2001; Wojciulik et al., 2001), implying that spatial working memory may be impaired in neglect.To test whether working memory for location is selectively impaired relative to memory for colour and shape, we comparedperformances of right hemisphere neglect patients with parietal (n = 4) and non-parietal (n = 4) lesions on a changedetection task. Patients were presented with a matrix containing four objects in different positions, and required to detect achange in the location, colour or shape of one of the objects following presentation of a brief visual mask. Parietal patientswere selectively impaired in detecting location changes, regardless of the horizontal position of the object in the matrix,relative to colour and shape changes. This deficit of spatial working memory was not apparent for neglect patients withlesions that spared the parietal cortex. We conclude that the human parietal cortex is crucially involved in the updating andmaintenance of spatial representations across saccades, and that neglect arising from parietal damage causes impairment inthese re-mapping mechanisms.
Key words: attention, change detection, parietal lobe, right hemisphere, spatial neglect, spatial working memory

1986). Second, single-neuron recordings inmonkeys have revealed cells in the posterior parietalcortex (PPC) whose activity is correlated with theupdating and maintenance of spatial locations ofvisual targets across saccades (Colby et al., 1996;Duhamel, Colby et al., 1992). One of the primaryfunctions of these cell populations is to re-map thelocations of objects across saccades in order tomaintain a stable perceptual representation of thevisual world (Colby, 1998). The PPC in humansalso seems to be selectively involved in re-mappingspatial locations across saccades. Patients withlesions of the PPC are impaired in double-stepsaccade tasks in which the retinal location of thesecond of two sequential visual targets must beupdated with extraretinal information about eyedisplacement arising from the first saccade (Heideet al., 1995; Heide and Kompf, 1997). The role ofthe PPC in such re-mapping has recently beenconfirmed by a functional brain imaging study oftriple-step saccades, which showed selectiveactivation of the right PPC and inferior frontal gyrusunder conditions of retinospatial dissonance (Heideet al., 2001).
Previous investigations of spatial workingmemory in neglect have required patients to locatevisual targets in displays of scattered items, as inthe cancellation tasks that are commonly used inthe clinical assessment of neglect. In one suchstudy, Wojciulik et al. (2001) tested a neglectpatient on a task requiring cancellation of Osscattered on a page. In one condition, the patientused an inking pen that left a visible red mark oneach target cancelled. In a second condition, thesame pen was used with its cap on, so that novisible marks were left on the page. In both tasksthe patient was asked to cancel each target onceonly. Despite this instruction, the patient re-cancelled several targets in the invisible markingcondition, in which the locations of previouslyvisited items had to be remembered. Such re-cancellations did not occur in the visible markingcondition, in which a permanent visible trace wasleft for items already found. In a secondexperiment, the Os were replaced by small linedrawings of different common objects. Thisafforded the patient an opportunity to rememberwhich items had been cancelled based on theiridentity, rather than on their location, thuscircumventing the need to rely on spatial workingmemory to avoid re-cancellations. In this secondexperiment, the patient never re-cancelled targets inthe invisible marking condition. Based on thesefindings, Wojciulik et al. (2001) concluded thattheir patient’s working memory impairment wasspecific to the spatial attributes of visual arrays,rather than to other non-spatial features.
Another study by the same group (Husain et al.,2001) confirmed this conclusion in a neglectpatient with right parietal damage, by measuringeye movements during visual search for Ts
amongst Ls on a computer display. The patient wasrequired to fixate each target and to press aresponse button to indicate that it was a ‘new’ (i.e.,previously undetected) item. Crucially, no visibletrace of previously tagged targets was visible onthe display, so that the patient had to remember thelocations of all previously found items to avoidrevisiting them. In a series of experiments thepatient exhibited a striking pattern of revisitingbehaviour, fixating previously detected targets andtreating them as new discoveries. The frequency ofsuch revisiting was correlated with the severity ofhis neglect on standard clinical tests, suggestingthat an impairment of spatial working memory, incombination with an ipsilesional attentional bias,exacerbates the tendency to ignore stimuli on theleft contralesional side.
Taken together, the results of these previousstudies of spatial working memory in neglectsuggest that these right hemisphere patients may beimpaired in updating the locations of targets invisual space during active spatial exploration(manual or saccadic). It is important to note,however, that re-visiting behaviour duringcancellation often occurs even when visible marksare left on target items, raising the possibility thatsome instances of re-visiting may be due to motorperseveration rather than to a failure of spatialworking memory (Manly et al., 2002; Na et al.,1999; Rusconi et al., 2002). Moreover, in previousstudies of spatial working memory in neglect thepatients’ primary task was to locate and markindividual targets in a visual array, rather thanexplicitly to remember their locations. It is thus notclear whether any impairment of spatial workingmemory in neglect is restricted to situations inwhich remembering the locations of items isincidental to the primary task of visual search, orwhether it also arises when patients are explicitlyinstructed to remember the locations of objects in avisual array. In the present study, therefore, weused a change-detection task in which no directedmotor response was required, and in whichparticipants were instructed explicitly to remembereither the spatial locations, colours or shapes of asmall number of items in a visual array.
Change-detection tasks have been usedextensively in studies of visual short-term memoryin normals. In a series of pioneering experiments,Phillips presented normal participants withcheckerboard patterns consisting of differentnumbers of black and white squares (Phillips, 1974,1983; Phillips and Christie, 1977a, b). After aninitial period of encoding, the display disappearedfor a short time, and then re-appeared either in itsoriginal form, or with one of the squares changedfrom black to white, or vice versa. The task was toindicate whether a change had occurred in thesecond display, so that accurate performancedepended on maintaining a representation of theposition of each square in the matrix across the
380 Laure Pisella and Others

blank interval (Phillips, 1974). Two findings fromthis work are important in the present context. First,visual short-term memory was found to have alimited capacity; errors in change detection withinterstimulus intervals greater than 100-200 msincreased significantly with the number of squaresin the matrix (Phillips, 1974, 1983). Second, thesensory processing of visual information was foundto be virtually unlimited in capacity; accuracy inchange detection was near ceiling with interstimulusintervals of less than 100 ms, even for very largematrices, provided there was no intervening mask toremove the local transient associated with thechanged square (Phillips, 1974, 1983).
Since the early work of Phillips and hiscolleagues, several subsequent studies haveconfirmed and extended these two major findings,employing the change-detection technique to studythe nature of visual perception and visual short-term memory (see O’Regan et al., 1999; Rensink,2002; Simons, 2000). One finding to emerge fromthese studies is that normal observers often fail todetect salient changes in a visual scene across briefmasked intervals, saccades, movie cuts and evenreal-life social interactions (O’Regan et al., 1999;Rensink, 2000; Simons and Levin, 1998). Theoccurrence of such ‘change blindness’ has led tothe hypothesis that only those objects in a visualscene that are selectively attended get stored inmemory; unattended objects are simply overwrittenupon presentation of a new scene, or following aneye movement or movie cut (Becker et al., 2000;Rensink, 2000; Simons, 2000). The change-detection technique thus provides a powerful toolfor exploring interactions between mechanisms ofselective attention and visual short-term memory.
In the present study we used a change-detectiontask to examine visual working memory in patientswith spatial neglect. Patients were shown an arrayof four different geometric shapes, each with aunique colour, and each occupying a specificlocation within a 4 × 4 matrix. Following a shortinitial exposure, the first display disappeared andwas replaced with a second such display, eitherimmediately or following a masked interstimulusinterval (ISI) of 1 s. The second display couldremain the same as the first (no-change trials), orelse one of the shapes could change its location,colour or shape (change trials). Patients wererequired to focus on just one of these attributes inseparate blocks of trials, and to indicate whether ornot a change had occurred. Using this approach wewere able to examine visual working memoryindependently for location, colour and shape, usingidentical displays. Since patients might be expectedto exhibit impairments in processing contralesionaltargets within the matrix due to perceptual orattentional deficits, the zero ISI condition wasincluded as a baseline measure of change detectionin the absence of a visual working memorycomponent.
We examined three principal hypotheses derivedfrom previous work. First, if spatial workingmemory in particular is impaired in neglect(Husain et al., 2001; Wojciulik et al., 2001),patients should have more difficulty detectingchanges in the spatial locations of targets over abrief delay, compared with changes to non-spatialfeatures such as colour and shape. Second, anyimpairment of spatial working memory shouldaffect targets across all locations within the matrix,not just those on the contralesional side. Thus,although a spatial gradient in change detectionmight be expected (with worse performance atmore contralesional locations), any differencebetween the zero- and delayed-ISI conditionsshould be equivalent across space for the location-change condition. Third, if parietal cortex is acrucial component of the network underlyingspatial working memory in humans (Awh et al.,1999; Husain et al., 2001), then neglect patientswith circumscribed parietal lesions should exhibit adeficit in spatial working memory, whereas thosein whom parietal cortex is spared should not.
METHOD
Patients
Eight patients (4 males, 4 females) who hadsuffered a first right-hemisphere stroke volunteeredtheir participation. Structural brain imaging (CT orMRI) was used to determine the locus and extentof damage, and lesioned areas were mapped onto astandard MRI template using MRIcro software(Rorden and Brett, 2000). Patients were dividedinto two subgroups based on lesion site: (1) thosewith damage involving the posterior parietal cortex(PPC); and (2) those with damage sparing the PPC(non-PPC group). The mean time interval betweenstroke onset and the experimental testing sessionswas 4.5 months for the PPC group (range: 2-9months) and 4 months for the non-PPC group(range: 2-8 months).
Each patient exhibited unilateral spatial neglecton admission to a neurological unit, as assessed bya standard battery of neuropsychological testsincluding line bisection, line cancellation, starcancellation, picture copying, and a perceptual biastest (the greyscales task; Mattingley et al., 1994).None of the patients had gaze paresis orhemianopia on confrontation testing. Patient detailsand clinical test performances are shown separatelyfor the PPC and non-PPC groups in Tables 1A and1B, respectively.
Stimuli
Visual displays were presented on the active-matrix screen of a laptop computer (Toshiba Tecra8200). A black outline grid consisting of a 4 × 4
Spatial working memory in neglect 381

382 Laure Pisella and Others
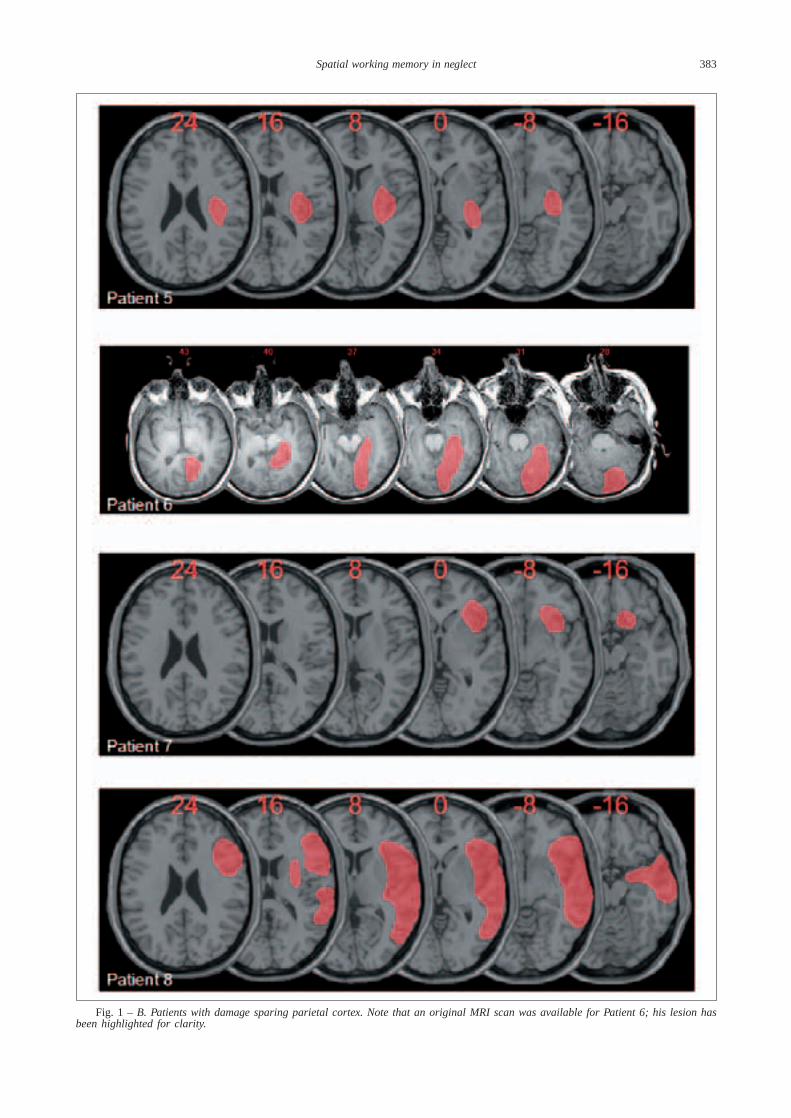
Fig. 1 – Lesion maps for the eight right hemisphere patients with spatial neglect, plotted onto a normal template brain using MRIcrosoftware (Rorden and Brett, 2000). Affected areas (translucent red) are plotted onto axial slices, with numbers in red above each sliceindicating Z-coordinates in Talairach space. A. Patients with damage affecting parietal cortex.
Spatial working memory in neglect 383
Fig. 1 – B. Patients with damage sparing parietal cortex. Note that an original MRI scan was available for Patient 6; his lesion hasbeen highlighted for clarity.
matrix of 16 cells (10º × 10º) was presented in thecentre of a uniform grey background. Fourdifferent objects were presented within the matrix,with the constraint that only one object couldappear in each of the four vertical columns. For thepurposes of the change detection task, each objectwas defined by three visual attributes: (1) location(one of the four cells within a vertical column); (2)colour (red, blue, green or yellow); and shape(cross, circle, square or triangle). There were thusfour different features for each of the three visualattributes. A masking display was used in theconditions involving a 1 s ISI, to prevent anyvisual persistence. This consisted of a 10º × 10ºgrid filled with randomly positioned fragments ofeach of the four colours and shapes used as targetobjects.
Procedure
Patients were seated comfortably in front of thecomputer screen, at a viewing distance ofapproximately 57 cm. After instructing the patientto fixate the centre of the blank computer screen,the experimenter initiated the trial. Each trialcommenced with the presentation of the initialencoding matrix containing four objects, theduration of which (1-3 s) was determinedindividually for each patient in a series of pilottests designed to avoid floor and ceiling effects inthe zero-ISI condition. This was followed by an ISIof either 0 s or 1 s. In the 1 s condition, themasking display replaced the matrix until the onsetof the second target display, which remained on thescreen until the patient responded (see Figure 2).Patients were not required to maintain fixation afterthe encoding display appeared; in fact they wereencouraged to explore the display thoroughly andto try and memorise its contents. They were
instructed that their task was to indicate whether achange had occurred to any of the target objects inthe matrix, according to a two-alternative forced-choice procedure (“change” or “no change”).Patients’ responses were keyed into the computerby the experimenter prior to the next trial beinginitiated. Patients received a number of practicetrials with feedback at the beginning of the sessionto familiarise themselves with the nature of thechanges for each attribute. In the experimentaltrials, no time constraint was imposed onresponses, and no feedback concerning accuracywas provided.
In separate blocks of trials patients wereinstructed to focus their attention on a singlestimulus attribute (location, colour or shape), andto report any change in the relevant attribute only.If a change occurred, it only ever involved achange in the cued attribute. (A pilot study inwhich changes occurred unpredictably in any of thethree attributes had to be discontinued because thetask proved too difficult for patients.) Within eachblock 80% of trials consisted of a change in theattended attribute, and 20% of trials consisted ofno change. On change trials, a single feature of therelevant attribute was changed in one of the fourobjects. The change was equally likely to occur forany of the four objects (i.e., within any of the fourvertical columns in the matrix). The no-changetrials were included as catch trials to provide anindex of false alarms.
Each patient completed six blocks of trials, oneat each ISI (0 and 1 s) for each stimulus attribute(location, colour, shape), according to an ABCCBAdesign. The order of blocks was randomised acrosspatients in each of the two lesion subgroups, withthe constraint that for each change attribute the 0ISI condition was always conducted first to providea baseline measure of performance. Each block
384 Laure Pisella and Others
TABLE IA
Age, sex and clinical details for neglect patients with right parietal lesions
Patient Sex Age Post-CVA Visual Clinical neglect tests(years) (months) fields
LB AL SC GS1 M 56 4 NAD + 27 12 0 + 1.002 F 72 9 NAD + 13 0 11 + 0.943 F 71 2 NAD + 48 62 79 + 1.004 M 72 3 NAD + 1 0 0 + 1.00
TABLE IB
Age, sex and clinical details for neglect patients with right non-parietal lesions
Patient Sex Age Post-CVA Visual Clinical neglect tests(years) (months) fields
LB AL SC GS5 M 59 2 NAD + 68 45 62 + 1.006 M 67 2 NAD + 58 40 2 + 1.007 F 61 4 NAD + 40 65 75 + 1.008 F 34 8 NAD – 5 0 14 + 0.06
Abbreviations: CVA – cerebrovascular accident; NAD – no abnormalities detected; LB – line bisection (% deviation to right (+) or left (–) of midpoint); AL –percentage of targets omitted on Albert’s line cancellation test; SC – percentage of targets omitted on the star cancellation test; GS – bias score on the greyscalestask (scores vary between – 1.00 (maximum left bias) and + 1.00 (maximum right bias), with 0 indicating no bias).

consisted of 50 trials (10 change trials for each ofthe four columns in the matrix, plus a further 10trials with no change), making 300 trials in total.For all patients testing lasted between two and fourhours, and was conducted in separate sessions ondifferent days.
RESULTS
The percentage of change trials in which achange was correctly reported (i.e, the percentageof hits) provided an estimate of the accuracy ofchange detection for each column in the matrix;this percentage was derived separately for eachchange attribute (location, colour, shape). The no-change trials were used to evaluate the global falsealarm rate, defined as the percentage of trials inwhich the patient indicated that a change hadoccurred when there was no change. The overallpercentage of false alarms was calculatedseparately for each patient, as a function of ISI andchange attribute. For purposes of analysis, the falsealarm rate thus obtained was assumed to behomogeneous across the four columns of thematrix, and so was divided by four to give anestimate of the false alarm rate for each columnalone. The resulting false alarm rates were thensubtracted from the hit rate obtained for each
column to yield a criterion-free estimate of changedetection performance. In the event, false-alarmrates were extremely low for all patients, and thecorrection made no difference to the overall patternof results.
Separate repeated-measures ANOVAs wereconducted on change detection scores from thePPC and non-PPC groups, with the factors of ISI(zero, delayed), Change Attribute (location, colour,shape), and Matrix Position (outer left, inner left,inner right, outer right). Change detectionperformance for the PPC and non-PPC groups isshown in Figures 3 and 4, respectively. Each graphshows the percentage of correct detections forlocation, colour and shape change according to theposition of the changed object within the matrix,with separate lines showing performance in thezero- and delayed-ISI conditions.
Patients in the PPC group exhibited asignificant spatial gradient as a function of matrixposition, with performance becoming graduallyworse toward the contralesional (left) side of thedisplay, F (3, 9) = 7.632, p < 0.05. The PPC groupalso showed a trend toward worse change detectionin the delayed-ISI condition than in the zero-ISIcondition, F (1, 3) = 7.632, p < 0.07. Mostimportantly, although there was no significant maineffect of Change Attribute, F (2, 6) < 1, n.s., therewas a significant two-way interaction between
Spatial working memory in neglect 385
Fig. 2 – Sequence of events in a typical trial of the working memory task. A. Location change. B. Colour change. C. Shape change.After ensuring that patients were fixating the centre of the display, the initial encoding matrix was presented briefly (1-3 s for differentindividuals). In the critical working memory trials, a masking display was then presented for 1 s. This was followed by the comparisonmatrix, in which a single object changed its location, colour or shape with a probability of 0.8. In the baseline (0 ISI) condition, themasking display was not presented. In the example sequences illustrated here, the change object is on the outer left of the matrix. Patientswere encouraged to explore and memorise the encoding display, and were permitted an unlimited time in which to respond.

Change Attribute and ISI, F (2, 6) = 13.472, p < 0.01. Inspection of Figure 3 suggests that thecost in change detection in the delayed- versuszero-ISI condition was greater for location changesthan for colour or shape changes. Figure 3 alsosuggests that although performance in the zero-ISIcondition was somewhat better for location changesthan for colour or shape changes (probably due tothe salient visual transient associated with locationchanges in the absence of a mask), performance inthe delayed-ISI condition was actually worse forlocation changes than colour or shape changes.Separate pairwise comparisons conducted for eachchange attribute confirmed these observations: PPCpatients showed a significant cost in the delayed-ISI condition for location changes, F (1) = 14.2, p < 0.05, but not for colour, F (1) = 2.59, p > 0.1,or shape changes, F (1) = 3.13, p > 0.1. Therewere no significant interactions involving the factorof Matrix Position [ISI × Matrix Position, F (3, 9)< 1, n.s.; ISI × Matrix Position × StimulusAttribute, F (6, 18) < 1, n.s.], indicating that PPCpatients’ poor performance in detecting locationchanges in the delayed-ISI condition wasequivalent across the four horizontal positionswithin the matrix.
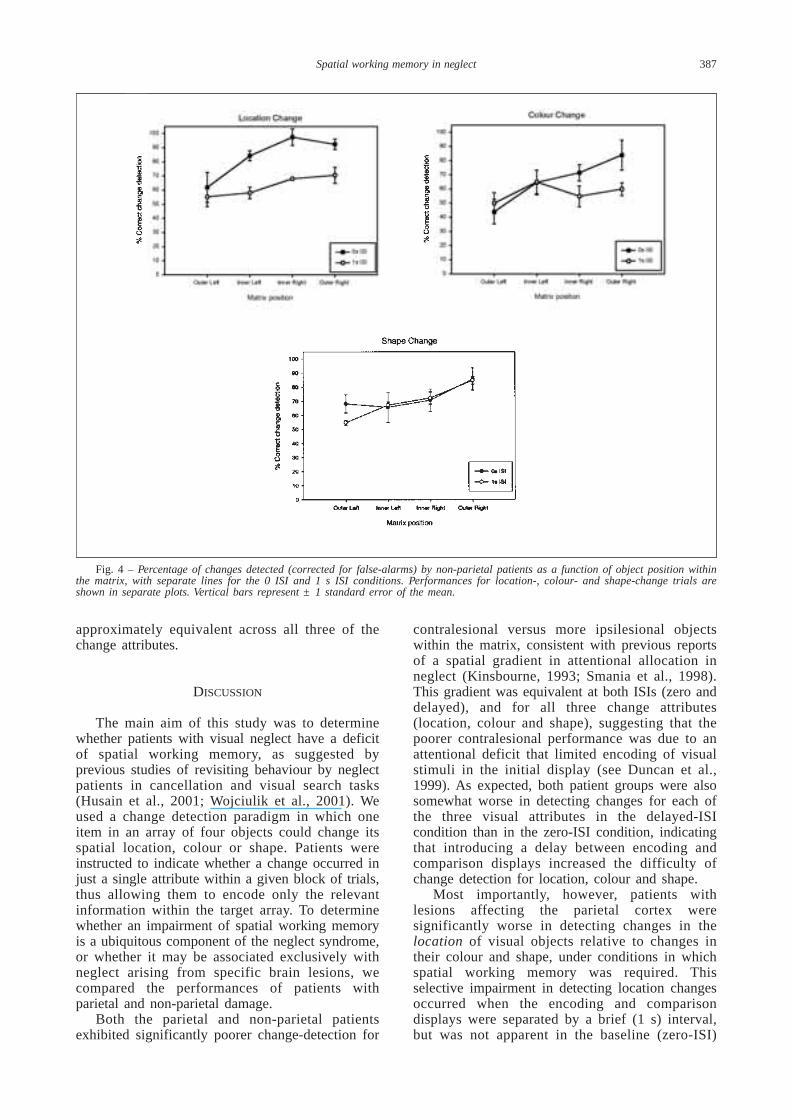
Analysis of data from the non-PPC grouprevealed a significant main effect of MatrixPosition, F (3, 9) = 5.065, p < 0.05, againindicating a spatial gradient of performance withpoorer change detection at more contralesionallocations within the matrix. There was nosignificant main effect of ISI, however, F (1, 3) =2.418, p < 0.10, or of Change Attribute, F (2, 6) =4.166, p > 0.05. Thus, in contrast with the PPCgroup, there was no trend for a difference betweenthe delayed- and zero-ISI conditions in the non-PPC group, suggesting no significant impairmentof visual working memory in these individuals (seeFigure 4). Most importantly, although detection oflocation changes by the non-PPC group appearssomewhat poorer in the delayed-ISI condition thanin the zero-ISI condition, particularly for the innerleft, inner right and outer right matrix positions,this difference was not statistically significant, asindicated by the absence of significant interactionsinvolving the factor of ISI (both Fs < 1). Moreover,inspection of Figure 4 suggests that in the zero-0ISI condition the non-PPC patients tended to bebetter in detecting location changes than colour orshape changes (as for the PPC group), whereas inthe delayed-ISI condition their performance was
386 Laure Pisella and Others
Fig. 3 – Percentage of changes detected (corrected for false-alarms) by parietal patients as a function of object position within thematrix, with separate lines for the 0 ISI and 1 s ISI conditions. Performances for location-, colour- and shape-change trials are shownin separate plots. Vertical bars represent ± 1 standard error of the mean.
approximately equivalent across all three of thechange attributes.
DISCUSSION
The main aim of this study was to determinewhether patients with visual neglect have a deficitof spatial working memory, as suggested byprevious studies of revisiting behaviour by neglectpatients in cancellation and visual search tasks(Husain et al., 2001; Wojciulik et al., 2001). Weused a change detection paradigm in which oneitem in an array of four objects could change itsspatial location, colour or shape. Patients wereinstructed to indicate whether a change occurred injust a single attribute within a given block of trials,thus allowing them to encode only the relevantinformation within the target array. To determinewhether an impairment of spatial working memoryis a ubiquitous component of the neglect syndrome,or whether it may be associated exclusively withneglect arising from specific brain lesions, wecompared the performances of patients withparietal and non-parietal damage.
Both the parietal and non-parietal patientsexhibited significantly poorer change-detection for
contralesional versus more ipsilesional objectswithin the matrix, consistent with previous reportsof a spatial gradient in attentional allocation inneglect (Kinsbourne, 1993; Smania et al., 1998).This gradient was equivalent at both ISIs (zero anddelayed), and for all three change attributes(location, colour and shape), suggesting that thepoorer contralesional performance was due to anattentional deficit that limited encoding of visualstimuli in the initial display (see Duncan et al.,1999). As expected, both patient groups were alsosomewhat worse in detecting changes for each ofthe three visual attributes in the delayed-ISIcondition than in the zero-ISI condition, indicatingthat introducing a delay between encoding andcomparison displays increased the difficulty ofchange detection for location, colour and shape.
Most importantly, however, patients withlesions affecting the parietal cortex weresignificantly worse in detecting changes in thelocation of visual objects relative to changes intheir colour and shape, under conditions in whichspatial working memory was required. Thisselective impairment in detecting location changesoccurred when the encoding and comparisondisplays were separated by a brief (1 s) interval,but was not apparent in the baseline (zero-ISI)
Spatial working memory in neglect 387
Fig. 4 – Percentage of changes detected (corrected for false-alarms) by non-parietal patients as a function of object position withinthe matrix, with separate lines for the 0 ISI and 1 s ISI conditions. Performances for location-, colour- and shape-change trials areshown in separate plots. Vertical bars represent ± 1 standard error of the mean.

condition in which a visual transient between thedisplays provided a cue to indicate the changelocation. The dissociation between delayed andimmediate change detection suggests that parietalpatients are impaired in maintaining representationsof object locations over time, and/or in comparingthese representations with those of subsequentdisplays. Crucially, this impairment in detectinglocation changes over a brief delay was equivalentacross all four horizontal positions of the matrix,consistent with the view that spatial workingmemory impairment in neglect is not restricted tocontralesional locations (De Renzi and Nichelli,1975; Husain et al., 2001).
In contrast to the parietal patients, there was noselective impairment of spatial working memoryfor individuals with lesions that spared the parietalcortex, even though the two groups showed clinicalneglect of equivalent severity (compare Tables 1Aand 1B). This suggests that the spatial workingmemory deficit is not a ubiquitous feature of theneglect syndrome, but rather a specific concomitantof parietal damage.
This finding is consistent with previous studieson the role of the parietal cortex in spatial workingmemory. Functional brain imaging studies haveshown that the posterior parietal cortex is part of afronto-parietal network associated with visualspatial short-term memory (Awh et al., 1999;Jonides et al., 1993; Owen et al., 1996), and withthe updating and maintenance of the locations ofvisual targets across saccades (Heide et al., 2001).In addition, neuropsychological studies suggest thatdeficits of visuospatial working memory tend to beassociated with right posterior hemispheric lesions,whereas impairments of auditory-verbal workingmemory are typically associated with left inferiorparietal damage (Basso et al., 1982; De Renzi andNichelli, 1975; Hanley et al., 1991; see also Vallarand Papagno, 1995 for a review). The visuospatialmemory deficits observed in these studies of righthemisphere patients have not been restricted to thecontralesional side of space (De Renzi and Nichelli,1975; Hanley et al., 1991), again suggesting thatspatial working memory impairments can bedissociated from the bias of attention thatcharacterises the clinical syndrome of spatialneglect.
We must be cautious in ruling out all possibleinvolvement of non-parietal structures in spatialworking memory, however, given that previousstudies have included parts of the prefrontal cortex,and in particular the inferior and middle frontalgyri, in the network subserving visual workingmemory (Heide et al., 2001; Leung et al., 2002;Wojciulik et al., 2001). Our own data from non-PPC patients indicated a non-significant trend forpoorer performance in the delayed- versus zero-ISIcondition for location changes that was notapparent for colour and shape changes. It isimportant to note, however, that this trend can be
largely attributed to the non-PPC group’s betterperformance for location changes than for colour orshape changes in the zero-ISI condition (see Figure4), in which salient visual transients could haveacted as attentional cues, rather than to a selectivedeficit in detecting location changes in the delayed-ISI condition (as shown by the PPC group; seeFigure 3). Future research will need to test largergroups of patients with small, well-circumscribedlesions of different brain regions to determine theanatomical specificity of spatial working memoryimpairments in neglect. It will also be important todetermine whether left parietal patients with right-sided neglect have impaired spatial workingmemory, rather than (or perhaps in addition to)problems with auditory-verbal material assuggested by previous investigations (De Renzi andNichelli, 1975; Basso et al., 1982).
A recent study of visual working memory inneglect revealed effects that are broadly similar tothose reported here, though with several importantdifferences. Denis et al. (2002) had righthemisphere patients recall the names and locationsof four common objects positioned in 2x2 squarematrix. In a ‘perceptual’ condition, patientsinspected each array for 90 seconds and named theobjects while they remained visible. Patients withneglect were significantly worse for objects locatedon the left (contralesional) side than on the right,as expected given their attentional impairment.This left-sided impairment persisted in two furtherconditions: first, when the patients had to reportthe names and locations of objects from memory,following an initial 90 second visual encodingphase; and second, when patients had to report thenames and locations of objects from memory afterreceiving a verbal description of the objects in thearray (e.g., “The cake is in front of the biscuit. Thebiscuit is on the left of the ice-cream.”). Thefindings from the latter two conditions suggest animpairment in visual working memory in neglectthat is significantly worse for objects locatedtoward the contralesional side of a display. This isclearly inconsistent with the findings of our study,which suggest that spatial working memory inparietal cases applies equally across horizontalspace. It should be noted, however, that Denis etal. (2002) also found that although their patientstypically failed to remember the locations ofobjects within the matrix, they often correctlyrecalled their names. This supports our generalconclusion that parietal damage affects spatialworking memory while leaving working memoryfor other visual attributes, such as colour andshape, intact. It is also noteworthy that 10 of the 11neglect patients tested by Denis et al. (2002) hadparietal lesions.
Our findings are also broadly consistent withfunctional imaging studies of change detection inneurologically healthy participants. For instance,Huettel et al. (2001) found that a network of brain
388 Laure Pisella and Others

regions including the cortex surrounding theinferior parietal sulcus was active during visualsearch for change across flickering displays of acomplex, naturalistic scene. A similar region ofparietal activity has been shown to be associatedwith the conscious detection of change in visualdisplays containing face or house stimuli (Beck etal., 2001). Our data extend these findings bysuggesting that the parietal cortex may bespecifically involved in maintaining the spatiallocations of visual objects rather than other visualattributes such as colour and shape.
An important feature of our change-detectionparadigm was that patients were simply required toindicate whether a change had occurred, using atwo-alternative forced-choice decision. They werenot required to indicate where any perceivedchange had occurred within the matrix; nor werethey required to indicate what had changed.Previous research has shown that normal observersare sometimes able to detect changes in visualdisplays without being aware of what has changed(Fernandez-Duque and Thornton, 2000). It istherefore possible that our performance measuresreflected both explicit perception of location,colour or shape change, as well as any implicitknowledge of change in the absence of awareness.Future experiments could examine the relativecontributions of explicit and implicit processes inneglect by having patients indicate the object ofany change and its location, and comparingperformance with simple detection for the samedisplays.
Another important finding to emerge from ourstudy is that spatial working memory in parietalneglect can be impaired over a delay of just 1 sbetween encoding and comparison displays. Infuture studies it will be important to determinewhether spatial working memory in these patientsdeclines further over longer delays, and inparticular whether the rate of decline is more rapidfor location information compared with other visualattributes (see also Halligan et al., 1992). The factthat we found a spatial working memory deficitover a very short delay is consistent with the workof Phillips and colleagues (Phillips, 1974; Phillipsand Christie, 1977a, b; Phillips, 1983), whoshowed in normals that a masked delay of just100-200ms between encoding and comparisondisplays is sufficient to reveal capacity limits inspatial working memory. Taken together, the spatialworking memory results from parietal neglectpatients and normals fit with neurophysiologicalfindings on the role of parietal areas in maintainingstable visual representations across saccades (Colbyet al., 1996; Duhamel et al., 1992; Heide et al.,1995; Heide and Kompf, 1997; Heide et al., 2001).If parietal lesions impair the ability to maintain andupdate visual representations across eyemovements, this would account for a number ofthe classical features of clinical neglect, including
patients’ failure to maintain awareness of stimulithey previously fixated (Halligan et al., 1992;Halligan and Marshall, 1993; Husain et al., 2001).
In conclusion, we have shown that neglect inright parietal patients is accompanied by a selectivedeficit of spatial working memory that is notrestricted to the contralesional side of space. Inagreement with previous studies (e.g., Husain etal., 2001; Wojciulik et al., 2001), we suggest thatproblems in remembering the locations of targetobjects may exacerbate the spatial bias thatcharacterises neglect behaviour, effectively makingalready detected items seem like new discoveriesduring cancellation and visual search tasks.Because the right parietal cortex is crucial for bothselective attention and spatial working memory,damage to this region will tend to produce both aspatial bias in selectivity and a deficit of spatialworking memory. However, we have also shownthat the two impairments can be dissociated, andthat neglect arising from non-parietal lesions is lesslikely to involve a spatial working memorycomponent.
Acknowledgements. The authors wish to thank all theparticipants for devoting several hours of their time to thisstudy. LP was supported by an NHMRC (Australia) –INSERM (France) post-doctoral fellowship. JBM wassupported by a grant from the Australian ResearchCouncil.
REFERENCES
AWH E, JONIDES J, SMITH EE, BUXTON RB, FRANK LR, LOVE T,WONG EC and GMEINDL L. Rehearsal in spatial workingmemory: Evidence from neuroimaging. PsychologicalScience, 10: 433-437, 1999.
BARBIERI C and DE RENZI E. Patterns of neglect dissociation.Behavioral Neurology, 2: 13-24, 1989.
BASSO A, SPINNLER H, VALLAR G and ZANOBIO ME. Lefthemisphere damage and selective impairment of auditory-verbal short-term memory. Neuropsychologia, 20: 263-274,1982.
BECK DM, REES G, FRITH CD and LAVIE N. Neural correlates ofchange detection and change blindness. Nature Neuroscience,4: 645-650, 2001.
BECKER MW, PASHLER H and ANSTIS SM. The role of iconicmemory in change-detection tasks. Perception, 29: 273-286,2000.
COLBY CL. Action-oriented spatial reference frames in cortex.Neuron, 20: 15-24, 1998.
COLBY CL, DUHAMEL J-R and GOLDBERG ME. Visual, presaccadic,and cognitive activation of single neurons in monkey lateralintraparietal area. Journal of Neurophysiology, 76: 2841-2852,1996.
DENIS M, BESCHIN N, LOGIE RH and DELLA SALA S. Visualperception and verbal descriptions as sources for generatingmental representations: Evidence from representationalneglect. Cognitive Neuropsychology, 19: 97-112, 2002.
DE RENZI E and NICHELLI P. Verbal and non-verbal short-termmemory impairment following hemispheric damage. Cortex,11: 341-354, 1975.
DUHAMEL J-R, COLBY CL and GOLDBERG ME. The updating of therepresentation of visual space in parietal cortex by intendedeye movements. Science, 255: 90-92, 1992.
DUNCAN J, BUNDESEN C, OLSON A, HUMPHREYS G, CHAVDA S andSHIBUYA H. Systematic analysis of deficits in visual attention.Journal of Experimental Psychology: General, 128: 450-478,1999.
FERNANDEZ-DUQUE D and THORNTON IM. Change detectionwithout awareness: Do explicit reports underestimate therepresentation of change in the visual system? VisualCognition, 7: 323-344, 2000.
Spatial working memory in neglect 389

GNADT JW and ANDERSEN RA. Memory related motor planningactivity in posterior parietal cortex of macaque. ExperimentalBrain Research, 70: 216-220, 1988.
HALLIGAN PW, DONEGAN CA and MARSHALL JC. When is a cuenot a cue? On the intractability of visuospatial neglect.Neuropsychological Rehabilitation, 2: 283-293, 1992.
HALLIGAN PW and MARSHALL JC. Homing in on neglect: A casestudy of visual search. Cortex, 29: 167-174, 1993.
HALLIGAN PW and MARSHALL JC. Focal and global attentionmodulate the expression of visuo-spatial neglect: A case study.Neuropsychologia, 32: 13-21, 1994.
HANLEY JR, YOUNG AW and PEARSON NA. Impairment of thevisuo-spatial sketch pad. Quarterly Journal of ExperimentalPsychology, 43A: 101-125, 1991.
HEIDE W, BINKOFSKI F, SEITZ RJ, POSSE S, NITSCHKE MF, FREUNDHJ and KOMPF D. Activation of frontoparietal cortices duringmemorized triple-step sequences of saccadic eye movements:An fMRI study. European Journal of Neuroscience, 13: 1177-1189, 2001.
HEIDE W, BLANKENBURG M, ZIMMERMANN E and KOMPF D.Cortical control of double-step saccades: Implications forvisuo-spatial orientation. Annals of Neurology, 38: 739-748,1995.
HEIDE W and KOMPF D. Specific parietal lobe contribution tospatial constancy across saccades. In P Thier and H-O Karnath(Eds), Parietal Lobe Contributions to Orientation in 3d Space.Heidelberg: Springer-Verlag, 1997, pp. 149-172.
HEILMAN KM and VALENSTEIN E. Frontal lobe neglect in man.Neurology, 22: 660-664, 1972.
HUETTEL SA, GUZELDERE G and MCCARTHY G. Dissociating theneural mechanisms of visual attention in change detectionusing functional MRI. Journal of Cognitive Neuroscience, 13:1006-1018, 2001.
HUSAIN M, MANNAN S, HODGSON T, WOJCIULIK E, DRIVER J andKENNARD C. Impaired spatial working memory acrosssaccades contributes to abnormal search in parietal neglect.Brain, 124: 941-952, 2001.
JONIDES J, SMITH EE, KOEPPE RA, AWH E, MINOSHIMA SE andMINTUN MA. Spatial working memory in humans as revealedby PET. Nature, 363: 623-625, 1993.
KARNATH H-O. Deficits of attention in acute and recovered hemi-neglect. Neuropsychologia, 20: 27-45, 1988.
KARNATH H-O, FERBER S and HIMMELBACH M. Spatial awarenessis a function of the temporal not the posterior parietal lobe.Nature, 411: 950-953, 2001.
KARNATH H-O, MILNER AD and VALLAR G (Eds). The Cognitiveand Neural Bases of Spatial Neglect. Oxford: OxfordUniversity Press, 2002.
KINSBOURNE M. Orientational bias model of unilateral neglect:Evidence from attentional gradients within hemispace. In IHRobertson and JC Marshall (Eds), Unilateral Neglect: Clinicaland Experimental Studies. Hove: Lawrence ErlbaumAssociates, 1993, pp. 63-86.
LEUNG HC, GORE JC and GOLDMAN-RAKIC PS. Sustainedmnemonic response in the human middle frontal gyrus duringon-line storage of spatial memoranda. Journal of CognitiveNeuroscience, 14: 659-671, 2002.
MANLY T, WOLDT K, WATSON P and WARBURTON EF. Is motorperseveration in unilateral neglect ‘driven’ by the presence ofneglected left-sided stimuli? Neuropsychologia, 40: 1794-1803, 2002.
MARSHALL JC and HALLIGAN PW. Seeing the forest but only halfthe trees. Nature, 373: 521-523, 1995.
MATTINGLEY JB, BRADSHAW JL, NETTLETON NC and BRADSHAWJA. Can task specific perceptual bias be distinguished from unilateral neglect? Neuropsychologia, 32: 805-817,1994.
MESULAM MM. Spatial attention and neglect: Parietal, frontal andcingulate contributions to the mental representation andattentional targeting of salient extrapersonal events.
Philosophical Transactions of the Royal Society of London -Series B: Biological Sciences, 354: 1325-1346, 1999.
NA DL, ADAIR JC, KANG Y, CHUNG CS, LEE KH and HEILMANKM. Motor perseverative behaviour on a cancellation task.Neurology, 52: 1569-1576, 1999.
O’REGAN JK, RENSINK RA and CLARK JJ. Change-blindness as aresult of ‘mudsplashes’. Nature, 398: 34, 1999.
OWEN AM, EVANS AC and PETRIDES M. Evidence for a two-stagemodel of spatial working memory processing within thelateral frontal cortex. A positron emission tomography study.Cerebral Cortex, 6: 31-38, 1996.
PHILLIPS WA. On the distinction between sensory storage andshort-term visual memory. Perception & Psychophysics, 16:283-290, 1974.
PHILLIPS WA. Short-term visual memory. PhilosophicalTransactions of the Royal Society of London – Series B:Biological Sciences, 302: 295-309, 1983.
PHILLIPS WA and CHRISTIE DFM. Components of visual memory.Quarterly Journal of Experimental Psychology, 29: 117-133,1977a.
PHILLIPS WA and CHRISTIE DFM. Interference with visualisation.Quarterly Journal of Experimental Psychology, 29: 637-650,1977b.
RENSINK RA. Seeing, sensing, and scrutinizing. Vision Research,40: 1469-1487, 2000.
RENSINK RA. Change detection. Annual Review of Psychology, 53:245-277, 2002.
ROBERTSON IH. The relationship between lateralised and non-lateralised attentional deficits in unilateral neglect. In IHRobertson and JC Marshall (Eds), Unilateral Neglect: Clinicaland Experimental Studies. Hove: Lawrence ErlbaumAssociates, 1993, pp. 257-275.
ROBERTSON IH, MATTINGLEY JB, RORDEN C and DRIVER J. Phasicalerting of neglect patients overcomes their spatial deficit invisual awareness. Nature, 395: 169-172, 1998.
ROBERTSON LC, LAMB MR and KNIGHT RT. Effects of lesions oftemporo-parietal junction on perceptual and attentionalprocessing in humans. Journal of Neuroscience, 8: 3757-3769,1988.
RORDEN C and BRETT M. Stereotaxic display of brain lesions.Behavioral Neurology, 12: 191-200, 2000.
RUSCONI ML, MARAVITA A, BOTTINI G and VALLAR G. Is the intact side really intact? Perseverative responses in patients with unilateral neglect: A productive manifestation.Neuropsychologia, 40: 594-604, 2002.
SIMONS DJ. Current approaches to change blindness. VisualCognition, 7: 1-15, 2000.
SIMONS DJ and LEVIN DT. Failure to detect changes to people in areal-world interaction. Psychonomic Bulletin & Review, 5:644-649, 1998.
SMANIA N, MARTINI MC, GAMBINA G, TOMELLERI G, PALAMARA A,NATALE E and MARZI CA. The spatial distribution of visualattention in hemineglect and extinction patients. Brain, 121:1759-1770, 1998.
VALLAR G. Spatial hemineglect in humans. Trends in CognitiveScience, 2: 87-97, 1998.
VALLAR G and PAPAGNO C. Neuropsychological impairments ofshort-term memory. In A Baddeley, BA Wilson and FN Watts(Eds), Handbook of Memory Disorders. Wiley: Chichester, pp.135-165.
VALLAR G and PERANI D. The anatomy of unilateral neglectafter right-hemisphere stroke lesions. A clinical/CT-scancorrelation study in man. Neuropsychologia, 24: 609-622, 1986.
WOJCIULIK E, HUSAIN M, CLARKE K and DRIVER J. Spatial workingmemory deficit in unilateral neglect. Neuropsychologia, 39:390-396, 2001.
Jason B. Mattingley, Cognitive Neuroscience Laboratory, Department of Psychology,University of Melbourne, Victoria 3010, Australia. e-mail: [email protected]
390 Laure Pisella and Others