
inventory control and economic order quantity in - St Clements ...

ORIGINAL ARTICLE
Quality or quantity: is nutrient transfer driven moreby symbiont identity and productivity than bysymbiont abundance?
Christopher J Freeman1, Robert W Thacker1, David M Baker2,3 and Marilyn L Fogel2
1Department of Biology, University of Alabama at Birmingham, Birmingham AL, USA; 2GeophysicalLaboratory, Carnegie Institution of Washington, Washington, DC, USA and 3Smithsonian Institution,National Museum of Natural History, Washington, DC, USA
By forming symbiotic interactions with microbes, many animals and plants gain access to theproducts of novel metabolic pathways. We investigated the transfer of symbiont-derived carbon andnitrogen to the sponges Aplysina cauliformis, Aplysina fulva, Chondrilla caribensis, Neopetrosiasubtriangularis and Xestospongia bocatorensis, all of which host abundant microbial populations,and Niphates erecta, which hosts a sparse symbiont community. We incubated sponges in light anddark bottles containing seawater spiked with 13C- and 15N-enriched inorganic compounds and thenmeasured 13C and 15N enrichment in the microbial (nutrient assimilation) and sponge (nutrienttransfer) fractions. Surprisingly, although most sponges hosting abundant microbial communitieswere more enriched in 13C than N. erecta, only N. subtriangularis was more enriched in 15N thanN. erecta. Although photosymbiont abundance varied substantially across species, 13C and 15Nenrichment was not significantly correlated with photosymbiont abundance. Enrichment wassignificantly correlated with the ratio of gross productivity to respiration (P:R), which varied acrosshost species and symbiont phylotype. Because irradiance impacts P:R ratios, we also incubatedA. cauliformis in 13C-enriched seawater under different irradiances to determine whether symbiontcarbon fixation and transfer are dependent on irradiance. Carbon fixation and transfer to the spongehost occurred in all treatments, but was greatest at higher irradiances and was significantlycorrelated with P:R ratios. Taken together, these results demonstrate that nutrient transfer frommicrobial symbionts to host sponges is influenced more by host–symbiont identities and P:R ratiosthan by symbiont abundance.The ISME Journal (2013) 7, 1116–1125; doi:10.1038/ismej.2013.7; published online 14 February 2013Subject Category: microbe-microbe and microbe-host interactionsKeywords: cyanobacteria; microbial symbionts; nutrient exchange; porifera; stable isotopes;symbiosis
Introduction
Symbioses between prokaryotes and their eukaryo-tic hosts are found across disparate taxa and withindiverse environments ranging from deep-sea vents,coral reefs, terrestrial soils, marine and terrestrialinvertebrates, and the mammalian gut (Boucheret al., 1982; Zilber-Rosenberg and Rosenberg, 2008;Rosenberg et al., 2010). In many cases, interactionsbetween distantly related organisms are mutuallybeneficial, whereby the symbiont is provided aprotected, nutrient-rich habitat in exchange forproducts of its metabolism (Venn et al., 2008;Vrijenhoek, 2010). These interactions can be highly
coevolved, leading to adaptations such as verticaltransmission of symbionts and reduced symbiontgenomes (Nyholm and McFall-Ngai, 2004; Moyaet al., 2008; Zilber-Rosenberg and Rosenberg, 2008;Thacker and Freeman, 2012).
In the marine environment, nutritional symbiosesare exemplified by mutualisms between reef-build-ing corals and dinoflagellates. Indeed, reef-buildingcorals would be unlikely to grow and compete forsubstrates in oligotrophic waters without supple-mental nutrition provided by their symbionts(Muscatine and Cernichiari, 1969; Muscatine et al.,1989). Other diverse groups of marine organismsalso host microbial symbionts, but these interactionsare less well understood (Taylor et al., 2007a, b;Venn et al., 2008; Webster and Blackall, 2009;Webster and Taylor, 2012; Thacker and Freeman,2012). For instance, marine sponges can dominatebenthic communities throughout the world(Becerro, 2008) and often host abundant and diverse
Correspondence: C Freeman, Smithsonian Marine Station at FortPierce, 701 Seaway Drive, Fort Pierce, FL 34949, USA.E-mail: [email protected] 4 May 2012; revised 20 December 2012; accepted 3January 2013; published online 14 February 2013
The ISME Journal (2013) 7, 1116–1125& 2013 International Society for Microbial Ecology All rights reserved 1751-7362/13
www.nature.com/ismej

microbial communities that are capable of complexmetabolic pathways, including carbon fixation andnitrogen transformations (Taylor et al., 2007a, b;Webster et al., 2010; Siegl et al., 2011).
Two recent studies demonstrated that stableisotopes of carbon (13C) and nitrogen (15N) couldbe used to investigate the transfer of symbiontcarbon and nitrogen to the host sponge (Weiszet al., 2007; Freeman and Thacker, 2011). Isotopevalues of isolated sponge cell and microbial sym-biont cell fractions revealed that the transfer ofsymbiont carbon and nitrogen differs across hostspecies (Freeman and Thacker, 2011). Thus, whilesome sponge–microbe symbioses appear to befacultative mutualisms, other sponge hosts do notacquire the products of symbiont metabolism andform commensal interactions (Freeman andThacker, 2011).
Inorganic compounds enriched in 13C (for example,NaH13CO3) and 15N (for example, Na15NO3) can serveas metabolic ‘tracers’ to follow the transfer ofautotrophic symbiont nutrition to a host (Tanakaet al., 2006). Because these compounds are assimi-lated via photosynthesis, host sponge cells becomeisotopically enriched through the transfer of enrichedorganic biomolecules from photosynthetic symbionts.To date, studies using this approach have conducted‘pulse-chase’ experiments, whereby organisms are‘pulsed’ in water containing enriched tracer for ashort period of time and subsequently held in naturalseawater for a ‘chase’ period, during which thetransfer of enriched compounds is followed (Tanakaet al., 2006, Weisz et al., 2010). While informative,such experiments are most often conducted with asingle species and include relatively lengthy chaseperiods (1–2 weeks), which limit the contempora-neous determination of differences among closelyrelated species, and thus significant functional varia-tion within taxonomic groups.
Although the transfer of symbiont-derived carbonand nitrogen varies across sponges that host densemicrobial communities, and appears to be negligiblein species hosting sparse microbial communities(Weisz et al., 2007; Freeman and Thacker, 2011), thefactors shaping such variation remain unknown. Forexample, photosymbiont abundance, the gross pro-ductivity to respiration (P:R) ratio of the symbiosis(that is, whether photosymbionts can compensatefor holobiont respiration) and symbiont phylotypealso vary across host species (Wilkinson, 1983;Erwin and Thacker, 2007; 2008a; Freeman andThacker, 2011). To determine whether these factorsimpact nutrient transfer, we coupled short-termlight–dark bottle incubations with two tracer com-pounds (NaH13CO3 and Na15NO3), followed by theseparation of symbiont and host cell fractions(Freeman and Thacker, 2011) before stable isotopeanalysis. We compared carbon and nitrogen assim-ilation by microbial symbionts and nutrient transferto the host to photosymbiont abundance, P:R ratios(Wilkinson, 1983; Erwin and Thacker, 2008a) and
photosymbiont identity (Erwin and Thacker, 2007,2008b). Because P:R is strongly influenced byirradiance within single species (Erwin andThacker, 2008a), we also incubated Aplysina cauli-formis in seawater spiked with NaH13CO3 undervarying irradiances. This experimental designallowed us to determine whether symbiont carbonfixation and nutrient transfer increase with increas-ing P:R. We tested three hypotheses: (1) 13C and 15Nenrichment of the microbial cell fraction (assimila-tion of inorganic compounds) and the sponge cellfraction (transfer of symbiont nutrition to the host)will be highest in sponges hosting dense symbiontcommunities, but will be negligible in Niphateserecta, which hosts only a sparse symbiont commu-nity; (2) variability in 13C and 15N enrichment acrossspecies will be driven more by photosymbiontabundance than by the P:R ratio of the symbiosis;and (3) carbon assimilation and transfer inA. cauliformis will be highest at maximum irradi-ance and will be directly correlated to P:R ratios.
Materials and methods
Study organismsSponges were collected from shallow (2–4 m) reefsadjacent to the Smithsonian Tropical ResearchInstitute in Bocas del Toro, Panama. Six focalspecies differed in their growth forms, symbiontphylotypes and reliance on symbiont nutrition.A. cauliformis, Aplysina fulva and Neopetrosiasubtriangularis all host abundant microbial commu-nities that are dominated by unicellular cyano-bacteria (Erwin and Thacker, 2007); however, eachof these hosts forms a unique relationship with itssymbionts (Freeman and Thacker, 2011). Thesponge N. erecta hosts a sparse community of non-photosynthetic microbes (Erwin and Thacker, 2007;Weisz et al., 2007), and gains nutrients primarily byfilter feeding (Freeman and Thacker, 2011); takentogether, these characteristics indicate that thisspecies is a suitable negative control in our experi-ment. While these four sponges have rope-likegrowth forms, the sponges Xestospongia bocatorensisand Chondrilla caribensis grow as encrusting cush-ions and host abundant filamentous and unicellularcyanobacterial symbionts, respectively (Erwin andThacker, 2007; Thacker et al., 2007).
Five large individuals of each of these specieswere transported to aquaria, where each individualwas divided into three replicate pieces. An initialpiece was wrapped in aluminum foil and immedi-ately frozen for initial measurements of naturalabundance stable isotope ratios (d13C and d15N) aswell as chlorophyll a (chl a) concentrations, whiletwo pieces were used in the experimental incuba-tions. Experimental pieces were incubated in flow-through seawater tanks for at least 12 h overnight toallow sponges to acclimate and to ensure that onlyhealthy individuals were used in the experiment.
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1117
The ISME Journal

d13C and d15N tracer experimentBefore the experiment, 2.0 g of 98 at% 13C NaH13CO3
(1.18 mM) and 200 mg of 98 at% 15N Na15NO3
(0.117 mM) were added to 20 l of filtered (0.70 mmGF/F Whatman, GE Healthcare Life Sciences,Piscataway, NJ, USA) seawater collected from off-shore. These compounds were used to trace auto-trophic pathways from microbial symbionts to hostsponges. Concentrations were selected based onpreliminary data suggesting that these were thelowest concentrations of tracer that provided ade-quate signal during short-term incubations. This‘enriched seawater’ was partitioned equally among40, 500 ml Nalgene HDPE bottles (20 light (translu-cent, with B50% of ambient light-transmitted)bottles and 20 dark (amber, with 0% of ambientlight-transmitted) bottles). Initial dissolved oxygen(DO, mg l�1) concentrations were measured using aNexSens (NexSens Technology, Inc., Alpha, OH,USA) WQ-DO dissolved oxygen sensor. This experi-mental design separated replicates and species, butmeant that incubations could be conducted withonly four species at a time (five individuals of fourspecies split among light and dark bottle treat-ments). For this reason, two sets of sponges wereincubated on consecutive days (four species the firstday and two the following day). Light and darkbottles were incubated under natural sunlight in aflow-through seawater tank (functioning as a con-stant temperature bath) for 6.4 and 5.9 h on the firstand second days, respectively. Ambient irradianceand temperature was recorded every 2 min duringthese incubations using HOBO data loggers attachedto the incubation tank and within light and darkbottles. At the end of the incubation, all bottles wereremoved from the tank and final DO concentrationswere measured. Sponges were removed, blotted dry,weighed, wrapped in aluminum foil and frozen forfuture analyses. To ensure that only metabolicallyactive tissue (organic matter) was included in thesemeasurements, additional specimens of each spe-cies were collected for construction of standardregressions between blotted wet weights, dryweights and ash-free dry weights. Blotted wetweights and dry weights were normalized to ash-free dry weights using the species-specific standardcurves.
d13C assimilation and transfer under varyingirradiancesEight large (B50 cm) individuals of A. cauliformiswere collected at a 4 m deep reef near SmithsonianTropical Research Institute Point, and returned to thelaboratory. Each of these replicates was partitionedinto five pieces (one for initial measurements and fourexperimental pieces); the experimental pieces wereallowed to acclimate overnight in flow-through sea-water aquaria as above. Translucent and dark bottleswere filled with filtered (0.70mm GF/F Whatman)seawater that was enriched with NaH13CO3, as above.
Experimental pieces were allocated to each of fourtreatments (dark (amber), light (translucent), lightlyshaded (light bottle wrapped with two layers of shadecloth, approximately 50% of light bottle irradiance)and heavily shaded (light bottle wrapped with fivelayers of shade cloth, approximately 10% of lightbottle irradiance)). Ambient and shaded irradianceswere measured using HOBO data loggers (Onset) inlux and converted to mmol photons m�2 s� 1 usingpublished conversion factors (http: //www.licor.com/)that were validated by simultaneous measurementsfrom a HOBO data logger and a Licor light meter.Experimental irradiances were chosen because priorresearch demonstrated that sponges under the lightlyshaded treatment were near the compensation point ofphotosynthesis, while sponges under the highlyshaded treatment were below this compensation point(Erwin and Thacker, 2008a).
P:R ratios and chl aNet primary productivity was calculated for eachlight incubation bottle by subtracting the initial DOconcentration from the final DO concentration; thesame calculation was applied to dark bottles to yieldrespiration (R). Values of net primary productivityand R were corrected for ash-free dry weights andincubation time so that final values were expressedas mg O2 l� 1 g�1 h� 1. Gross primary productivitywas calculated by subtracting R from net primaryproductivity for each individual. Net primaryproductivity, R and gross primary productivityvalues for each species are provided asSupplementary Information S1. Gross photosynthe-sis:respiration ratios (P:R) were calculated bydividing gross primary productivity by respiration.Because the experiments were conducted over thecourse of 2 days, P:R values were corrected toaccount for differences in irradiance. Irradiance wasmeasured every 2 min during the incubation period,and then averaged for each day’s incubation period.P:R values were analyzed using an analysis ofcovariance, with mean light intensity as the covari-ate, to obtain an adjusted least-squares mean andstandard error for the P:R ratios of each species.
To estimate photosymbiont abundance withinthese sponges, we measured chl a concentrationsusing lyophilized samples of each piece of sponge,following methods described by Freeman andThacker (2011). Although chl a values can varyowing to the physiological condition of somesymbionts, this metric is widely cited as the optimalmethod for quantifying photosymbiont abundance(Becerro and Paul, 2004; Sacristan-Soriano et al.,2011; Hill and Scott, 2012). Values were correctedfor ash-free dry weights. Because mean chl a valuesdid not differ across treatment for each species (thatis, values from initial samples and both treatmentswere not significantly different, as determined by ageneral linear model (GLM; Systat, version 11;Systat Software, Inc., Chicago, IL, USA) testing
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1118
The ISME Journal

treatment nested within species (P¼ 0.84, F¼ 0.58)and species (Po0.001, F¼ 237), the chl a value foreach species represents the combined mean of allsubsamples from each species.
Stable isotope analysesSymbiont and host cell fractions were separatedfrom bulk sponge tissue in A. cauliformis, A. fulva,C. caribensis, N. erecta and N. subtriangularis usingmethods adapted from Freeman and Thacker (2011),provided as Supplementary Information S2. Newmethods were developed for X. bocatorensis, whichhosts filamentous cyanobacterial symbionts; thesenew methods are also provided in SupplementaryInformation S2. Lyophilized microbial and spongecell pellets were prepared for isotope analyses asdescribed in Freeman and Thacker (2011). Stableisotope compositions were measured at the Geo-physical Laboratory using a Thermo Delta V isotoperatio mass spectrometer (Thermo Scientific, WestPalm Beach, FL, USA) coupled to a Carlo-ErbaNC2500 elemental analyzer via a Conflo III open-split interface. Mean precision of d13C and d15N fromseven or eight sets of duplicate samples from eachtreatment was ±0.2% and ±0.1% in initials,±0.23% and ±2.25% in samples from the darktreatment. In the light treatment, samples fromspecies other than N. subtriangularis had meand13C and d15N precision values of ±15.8% and±3.8%. Given that the observed magnitude ofenrichment for N. subtriangularis was much higherthan the other samples, its precision was calculatedseparately (197% and 209% for d13C and d15N,respectively). Isotope enrichment (expressed asAPE (Atom Percent Excess)) was calculated usingformulas adapted from Fry (2006), Tanaka et al.(2006) and Weisz et al. (2010):
APE13C¼ atom % 13C in experimental sample-atom % 13C in initial sample
Atom % 13C¼ ½100�Rstandard�ðd13C sample�1000þ 1Þ��½1þRs tan dard�ðd13C sample�1000þ1Þ�
d13C ¼ ½ðRsample�RstandardÞ� 1��1000 with R¼ 13C=12C
In experiments using tracer compounds, enrichmentmeasured as APE assesses the increase in the atompercent of 13C in experimental samples comparedwith the initial (or natural abundance) samples;thus, ‘sample’ refers to either the microbial orsponge cell fraction from an individual and ‘stan-dard’ refers to values of international standards ofVienna Pee Dee Belemnite and atmospheric N2 ford13C and d15N, respectively (Fry, 2006). The formulasabove are adapted for nitrogen by replacing 15N and14N for 13C and 12C.
Data analysisAll raw data for this manuscript have been depos-ited at Dryad. Mean APE13C and APE15N values
across treatments (light and dark bottles) werecompared using GLMs, with treatment as the mainfactor and source individual as a blocking variable,for each fraction of each species. For these and allsubsequent analyses, residuals were tested fornormality and homogeneity of variances amonggroups. To determine if the microbial and host cellfractions of each species differed in APE13C orAPE15N values, GLMs, with fraction as the mainfactor and individual as a blocking variable, wereanalyzed for each species. Mean APE values ofsponge and microbial fractions across the six specieswere compared using four separate analyses ofvariance (ANOVA), followed by pairwise compar-isons across species using the Fisher’s least-signifi-cant difference (LSD) test.
For P:R measurements, the adjusted least-squaresmeans for each species from the analysis ofcovariance were compared using post hoc pairwisecomparisons. Chl a concentrations across specieswere compared using a GLM testing the effects ofspecies and treatment nested within species, fol-lowed by post hoc pairwise comparisons acrossspecies. Linear regressions determined whethertrends in mean APE13C and APE15N across specieswere related to photosymbiont abundance (mea-sured as chl a concentrations) or productivity(measured as P:R ratios). All pairwise comparisonswere conducted using the LSD test.
In the irradiance experiment with 13C tracer, P:Rratios across treatments were compared using a GLMtesting the effect of treatment blocked by individual.Mean APE13C values of the two fractions withineach treatment were compared with a GLM testingthe effect of fraction blocked by individual. Themean APE13C values of each fraction across treat-ments were compared using a GLM testing the effectof treatment blocked by individual. The relationshipbetween APE13C values and productivity was deter-mined using a GLM testing the effects of individualand P:R ratio on APE13C. Multiple pairwise compar-isons were conducted using an LSD test.
Results and discussion13C and 15N tracer experimentSix species of marine sponges spanning diversesymbiotic interactions were incubated in NaH13CO3
and Na15NO3 to trace the transfer of symbiont-derived carbon and nitrogen to the host. Ourobserved enrichment of 15N or 13C in sponge cellfractions is direct evidence for the translocation ofenriched biomolecules from the symbiont. Varia-tions in the extent of uptake of 15N or 13C could bedue to the fact that sponges hosting abundantmicrobial communities (termed high microbialabundance (HMA) sponges) are generally thoughtto obtain large amounts of symbiont-derivednutrients compared with sponges hosting sparsesymbiont communities (termed low microbial
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1119
The ISME Journal

abundance sponges) (Hentschel et al., 2006; Tayloret al., 2007a). Minimal experimental evidence,however, supports this hypothesis (Thacker andFreeman, 2012).
Therefore, we originally hypothesized that HMAsponges would have higher 13C and 15N enrichmentthan N. erecta, which lacks photosymbionts, containsbacteria that are most closely related to those foundin the water column (Weisz et al., 2007), grows fasterin the shade, and has natural abundance isotopevalues suggestive of a heavy reliance on filter feeding(Freeman and Thacker, 2011). Indeed, APE13C andAPE15N values in N. erecta were low regardless ofwhether they were incubated in a light or dark bottle(GLM: P¼ 0.064, F¼ 0.26 and P¼ 0.074, F¼ 11.99 forAPE13C of microbial and sponge fractions, respec-tively, and P¼ 0.706, F¼ 0.16 and P¼ 0.26, F¼ 2.38for APE15N of microbial and sponge fractions,respectively; Figures 1 and 2), suggesting thatinorganic C and N were largely inaccessible to thislow microbial abundance sponge.
As expected, microbial and sponge cell fractionsfrom almost all (except the sponge fraction ofC. caribensis) of the HMA species were significantlymore enriched in 13C than the same fractions in N.erecta (Figure 1). In addition, there was significantvariation in APE13C values of the microbial andsponge cell fractions across the HMA species held inlight bottles (ANOVA: Po0.001, F¼ 45.6 andPo0.001, F¼ 15.43 for microbial and sponge cellfractions, respectively), as A. cauliformis, A. fulva,C. caribensis and X. bocatorensis had similar values,
but were all significantly less enriched compared toN. subtriangularis (Figure 1). Furthermore, unlike N.erecta, APE13C values of HMA species were sig-nificantly higher in light than in dark bottles,suggesting that carbon assimilation and transfer islight-dependent in these species and may be drivenby photosymbiont metabolism. For all species, weobserved minimal enrichment in 13C in dark bottles,suggesting that heterotrophic bacteria contributelittle to carbon assimilation.
Counter to our original hypothesis, however,although we observed substantial variation innitrate assimilation and transfer across these sixspecies (ANOVA: Po0.001, F¼ 56.8 and Po0.001,F¼ 17.5 for microbial and sponge fractions, respec-tively), and moderate 15N enrichment of HMAspecies implies that these sponges host microbescapable of nitrate assimilation, N. subtriangulariswas the only HMA sponge that was significantlymore enriched in 15N than N. erecta (Figure 2). 15Nenrichment was significantly lower in the darkbottles of most species (GLM: Po0.05; except thebacterial fraction of A. fulva), suggesting that nitrateassimilation may be light dependent, but that someheterotrophic assimilation in the dark may beoccurring. Although our data indicate that diversemicrobes, including both photosymbionts and pos-sibly heterotrophic microbes within some spongehosts are capable of assimilating nitrate as a sourceof inorganic nitrogen, regardless of irradiance, ourunderstanding of nitrogen assimilation withinmarine sponges remains relatively limited (Thackerand Freeman, 2012).
Because specific symbiont groups, like cyanobac-teria, are dominant members of the microbial
Figure 1 Mean (±s.e.) APE13C values for microbial and spongecell fractions from the sponges A. cauliformis (Acau), A. fulva(Aful), C. caribensis (Cnuc), N. erecta (Nere), N. subtriangularis(Nsub) and X. bocatorensis (Xboc) incubated in light (clear) anddark (amber) bottles. Values from the dark treatment are includedfor reference. Different letters within columns indicate statisti-cally significant differences in the APE13C of the sponge (lowercase) or microbial fraction (upper case) values across species, asdetermined by an ANOVA on each fraction followed by an LSDmultiple pairwise comparison.An asterisk indicates significantlydifferent APE13C values between the fractions of one species.N¼ for 5 for the microbial and sponge fractions of Acau, Aful,Cnuc, Nsub and Xboc and the microbial fraction of Nere. N¼ 3 forthe sponge fraction of Nere.
Figure 2 Mean (±s.e.) APE15N values for microbial and spongecell fractions from six species of sponges held in light (clear) anddark (amber) bottles. Values from the dark treatment are includedfor reference. Different letters within columns indicate statisti-cally significant differences in the APE15N of the sponge (lowercase) or microbial fraction (upper case) as determined by ANOVAon each fraction followed by an LSD multiple pairwise compar-ison. An asterisk indicates significantly different APE13C valuesbetween the fractions of one species. Abbreviations of speciesnames and sample sizes are identical to Figure 1.
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1120
The ISME Journal
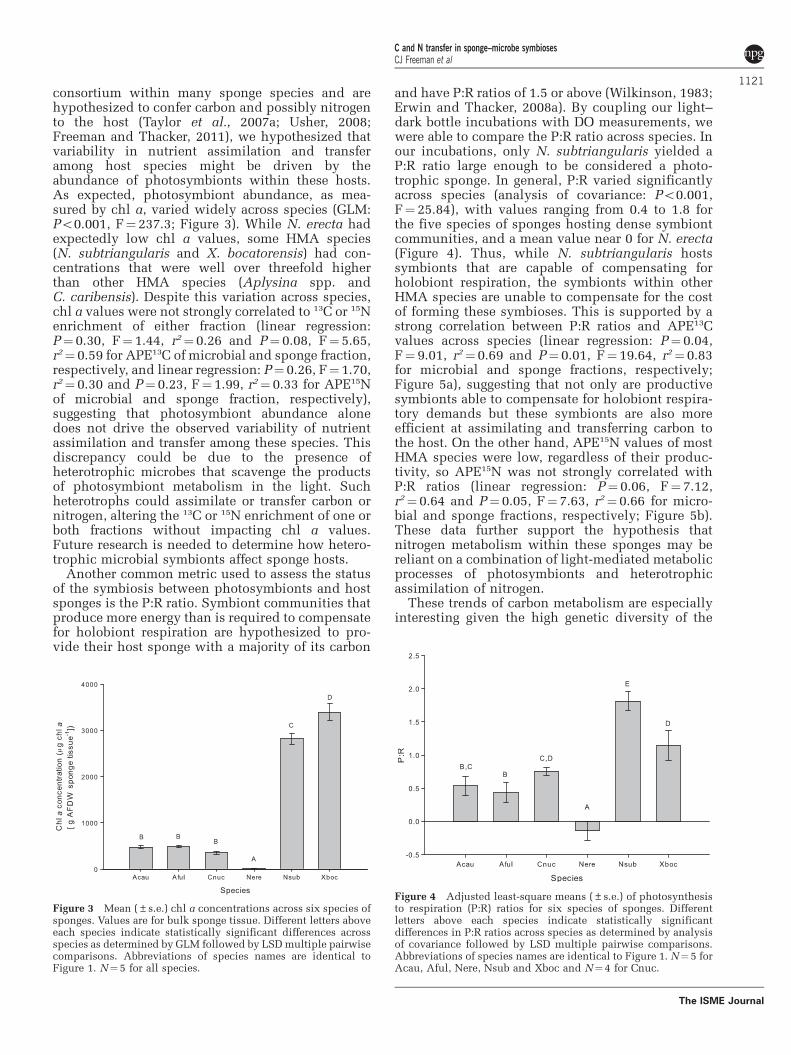
consortium within many sponge species and arehypothesized to confer carbon and possibly nitrogento the host (Taylor et al., 2007a; Usher, 2008;Freeman and Thacker, 2011), we hypothesized thatvariability in nutrient assimilation and transferamong host species might be driven by theabundance of photosymbionts within these hosts.As expected, photosymbiont abundance, as mea-sured by chl a, varied widely across species (GLM:Po0.001, F¼ 237.3; Figure 3). While N. erecta hadexpectedly low chl a values, some HMA species(N. subtriangularis and X. bocatorensis) had con-centrations that were well over threefold higherthan other HMA species (Aplysina spp. andC. caribensis). Despite this variation across species,chl a values were not strongly correlated to 13C or 15Nenrichment of either fraction (linear regression:P¼ 0.30, F¼ 1.44, r2¼ 0.26 and P¼ 0.08, F¼ 5.65,r2¼ 0.59 for APE13C of microbial and sponge fraction,respectively, and linear regression: P¼ 0.26, F¼ 1.70,r2¼ 0.30 and P¼ 0.23, F¼ 1.99, r2¼ 0.33 for APE15Nof microbial and sponge fraction, respectively),suggesting that photosymbiont abundance alonedoes not drive the observed variability of nutrientassimilation and transfer among these species. Thisdiscrepancy could be due to the presence ofheterotrophic microbes that scavenge the productsof photosymbiont metabolism in the light. Suchheterotrophs could assimilate or transfer carbon ornitrogen, altering the 13C or 15N enrichment of one orboth fractions without impacting chl a values.Future research is needed to determine how hetero-trophic microbial symbionts affect sponge hosts.
Another common metric used to assess the statusof the symbiosis between photosymbionts and hostsponges is the P:R ratio. Symbiont communities thatproduce more energy than is required to compensatefor holobiont respiration are hypothesized to pro-vide their host sponge with a majority of its carbon
and have P:R ratios of 1.5 or above (Wilkinson, 1983;Erwin and Thacker, 2008a). By coupling our light–dark bottle incubations with DO measurements, wewere able to compare the P:R ratio across species. Inour incubations, only N. subtriangularis yielded aP:R ratio large enough to be considered a photo-trophic sponge. In general, P:R varied significantlyacross species (analysis of covariance: Po0.001,F¼ 25.84), with values ranging from 0.4 to 1.8 forthe five species of sponges hosting dense symbiontcommunities, and a mean value near 0 for N. erecta(Figure 4). Thus, while N. subtriangularis hostssymbionts that are capable of compensating forholobiont respiration, the symbionts within otherHMA species are unable to compensate for the costof forming these symbioses. This is supported by astrong correlation between P:R ratios and APE13Cvalues across species (linear regression: P¼ 0.04,F¼ 9.01, r2¼ 0.69 and P¼ 0.01, F¼ 19.64, r2¼ 0.83for microbial and sponge fractions, respectively;Figure 5a), suggesting that not only are productivesymbionts able to compensate for holobiont respira-tory demands but these symbionts are also moreefficient at assimilating and transferring carbon tothe host. On the other hand, APE15N values of mostHMA species were low, regardless of their produc-tivity, so APE15N was not strongly correlated withP:R ratios (linear regression: P¼ 0.06, F¼ 7.12,r2¼ 0.64 and P¼ 0.05, F¼ 7.63, r2¼ 0.66 for micro-bial and sponge fractions, respectively; Figure 5b).These data further support the hypothesis thatnitrogen metabolism within these sponges may bereliant on a combination of light-mediated metabolicprocesses of photosymbionts and heterotrophicassimilation of nitrogen.
These trends of carbon metabolism are especiallyinteresting given the high genetic diversity of the
Figure 3 Mean (±s.e.) chl a concentrations across six species ofsponges. Values are for bulk sponge tissue. Different letters aboveeach species indicate statistically significant differences acrossspecies as determined by GLM followed by LSD multiple pairwisecomparisons. Abbreviations of species names are identical toFigure 1. N¼5 for all species.
Figure 4 Adjusted least-square means (±s.e.) of photosynthesisto respiration (P:R) ratios for six species of sponges. Differentletters above each species indicate statistically significantdifferences in P:R ratios across species as determined by analysisof covariance followed by LSD multiple pairwise comparisons.Abbreviations of species names are identical to Figure 1. N¼ 5 forAcau, Aful, Nere, Nsub and Xboc and N¼4 for Cnuc.
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1121
The ISME Journal
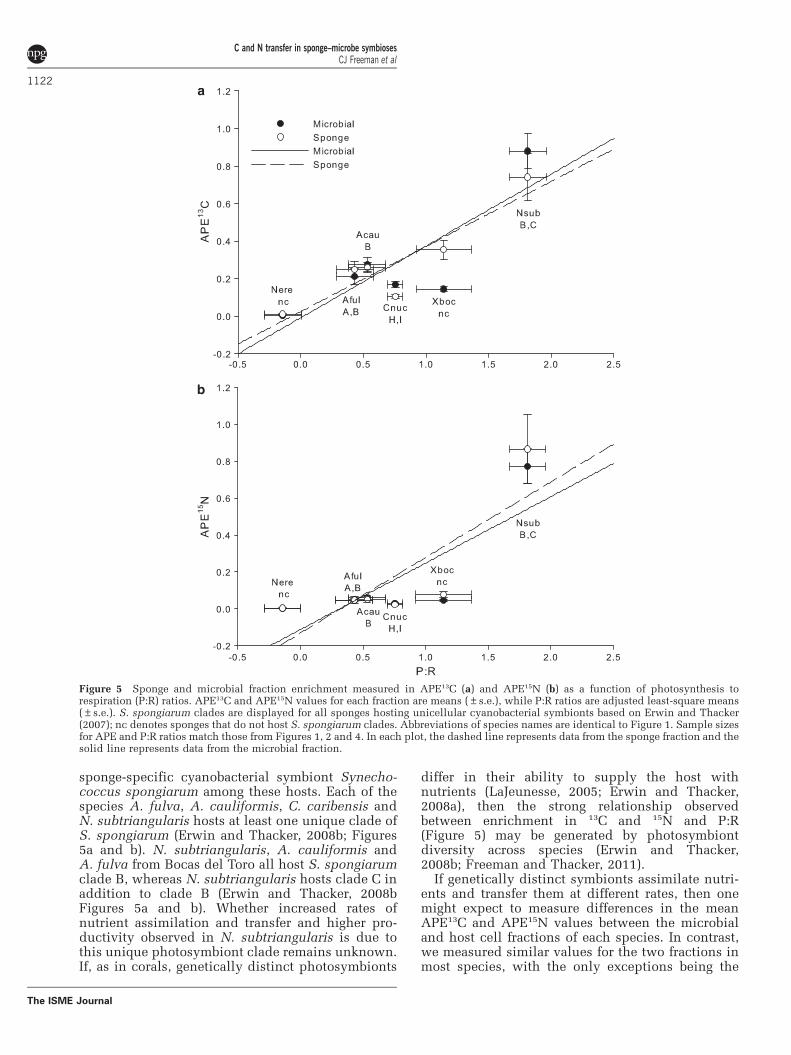
sponge-specific cyanobacterial symbiont Synecho-coccus spongiarum among these hosts. Each of thespecies A. fulva, A. cauliformis, C. caribensis andN. subtriangularis hosts at least one unique clade ofS. spongiarum (Erwin and Thacker, 2008b; Figures5a and b). N. subtriangularis, A. cauliformis andA. fulva from Bocas del Toro all host S. spongiarumclade B, whereas N. subtriangularis hosts clade C inaddition to clade B (Erwin and Thacker, 2008bFigures 5a and b). Whether increased rates ofnutrient assimilation and transfer and higher pro-ductivity observed in N. subtriangularis is due tothis unique photosymbiont clade remains unknown.If, as in corals, genetically distinct photosymbionts
differ in their ability to supply the host withnutrients (LaJeunesse, 2005; Erwin and Thacker,2008a), then the strong relationship observedbetween enrichment in 13C and 15N and P:R(Figure 5) may be generated by photosymbiontdiversity across species (Erwin and Thacker,2008b; Freeman and Thacker, 2011).
If genetically distinct symbionts assimilate nutri-ents and transfer them at different rates, then onemight expect to measure differences in the meanAPE13C and APE15N values between the microbialand host cell fractions of each species. In contrast,we measured similar values for the two fractions inmost species, with the only exceptions being the
Figure 5 Sponge and microbial fraction enrichment measured in APE13C (a) and APE15N (b) as a function of photosynthesis torespiration (P:R) ratios. APE13C and APE15N values for each fraction are means (±s.e.), while P:R ratios are adjusted least-square means(±s.e.). S. spongiarum clades are displayed for all sponges hosting unicellular cyanobacterial symbionts based on Erwin and Thacker(2007); nc denotes sponges that do not host S. spongiarum clades. Abbreviations of species names are identical to Figure 1. Sample sizesfor APE and P:R ratios match those from Figures 1, 2 and 4. In each plot, the dashed line represents data from the sponge fraction and thesolid line represents data from the microbial fraction.
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1122
The ISME Journal

microbial fraction of C. caribensis, which was sig-nificantly enriched in 13C and 15N relative to thesponge fraction (GLM: P¼ 0.003, F¼ 38.51 andP¼ 0.009, F¼ 22.9 for 13C and 15N, respectively;Figure 1) and the sponge fraction of X. bocatorensis,which was enriched in 13C relative to the microbialfraction (GLM: P¼ 0.007, F¼ 25.4). While theseobservations might suggest that the symbiosis inC. caribensis is less efficient than that in X. bocator-ensis or that X. bocatorensis might be able to stimulatenutrient transfer from its symbionts, future work isneeded to explicitly test these hypotheses. Suchstudies should quantify productivity, nutrient assim-ilation and transfer to the host at a wider variety oftime scales than reported here to assess the lag timebetween symbiont and host enrichment.
It is not surprising that the results from this studydo not exactly mirror the trends for the Aplysina spp.,N. subtriangularis and N. erecta seen in Freeman andThacker (2011). For instance, using natural abun-dance isotope values, we have shown that, innature, A. cauliformis obtains both C and N fromits symbionts, while A. fulva obtains only N andN. subtriangularis obtains only C from its symbiontcommunity and N. erecta obtains both C and N fromparticulate organic matter (Freeman and Thacker,2011). These d13C and d15N ratios represent natural,integrated records detailing whether these spongesare assimilating their C and/or N from particulateorganic matter or from symbiont sources over time.These values thus account for the fact that nutrientexchange may be due to factors such as ambientnutrient levels, particulate organic matter feeding,nutrient recycling within the symbiosis or othercurrently unknown factors. These light–dark bottleincubations, on the other hand, are assessments ofhow efficient the symbionts within these sponges areat assimilating and transferring nutrients whensupplied with low, but non-limiting, levels of C-and N-based compounds enriched in 13C and 15N.Comparisons of data from across these two studies,however, may help to explain some of these trends.
Nutrient assimilation and transfer to the host maybe driven more by symbiont productivity andphylotype than by symbiont abundance, implyingthat that these relationships are complex and tightlycoevolved symbioses (Knowlton and Rohwer, 2003;Thacker and Freeman, 2012). However, it remainsunclear why some sponges host less productiveclades and what processes might prevent a widerdistribution of the potentially more productiveS. spongiarum clade C. In a tightly coevolvedrelationship, selection might favor sponge–microbesymbioses with minimal cost:benefit ratios from thehost’s perspective. Because both heterotrophic filterfeeding and abundant communities of autotrophicsymbionts present metabolic costs to the hostsponge, we hypothesize that the host will balancethese two feeding modes to receive sufficientnutrients at minimal metabolic cost. For instance,N. subtriangularis hosts the highly productive
combination of clades B and C, which provides thehost with more carbon than is required to compen-sate for holobiont respiration. The cost of hostingthese symbionts might be reflected in lower hetero-trophic feeding, as this sponge assimilates o25% ofits carbon from filter-feeding (Freeman and Thacker2011). However, A. fulva hosts the less productiveclades A and B, and cannot compensate forholobiont respiration using symbiont-derived nutri-tion alone; reduced autotrophy might driveincreased heterotrophy, as this host assimilates over50% of its carbon from filter-feeding (Freeman andThacker, 2011). Indeed, these trade-offs betweenautotrophic and heterotrophic feeding modesappear to be well established; even under shadedconditions, N. subtriangularis maintains significantassimilation of symbiont-derived carbon, whileA. fulva becomes more reliant on heterotrophicfeeding (Freeman and Thacker, 2011).
Although abundant populations of microbialsymbionts are present within many sponge species,the relative benefits of these symbioses are drivenmore by the presence or absence of particular groupsof symbionts than overall symbiont abundance.These findings imply that the host may exert a levelof control over symbiont community composition inan attempt to maximize the benefits and minimizethe costs associated with different symbiont phylo-types (Bronstein, 2001; Cowden and Peterson, 2009;Thacker and Freeman, 2012).
13C assimilation and transfer under varying irradianceOur coupling of short-term incubations withenriched isotope tracers can be readily adapted toexamine how nutrient exchange differs underchanging environmental conditions. Given thatA. cauliformis maintains high levels of symbiont-derived carbon inputs, even under shaded conditions(Freeman and Thacker, 2011), we chose this speciesto test the hypothesis that symbiont carbon fixationand nutrient transfer to the host sponge would varyacross discrete irradiance levels and would bedirectly related to productivity. As expected, twoand five layers of shade cloth provided enough lightto be near (0.8) and well below (0.17) the compensa-tion point (Erwin and Thacker, 2008a). Takentogether with the light bottles, these treatmentsprovided three distinct light environments, and weobserved three distinct P:R ratios (GLM for treatment:Po0.001, F¼ 49.8; Figure 6a). As hypothesized,carbon fixation and transfer differed across lighttreatments (GLM for treatment: P¼ 0.001, F¼ 11.52and P¼ 0.001, F¼ 11.6 for microbial and spongefractions, respectively), was highest at the maximumirradiance (Figure 6b), and was correlated with P:Rratio (GLM testing the effects of individual and P:Rratio on APE13C: Po0.01, F¼ 16.6, r2¼ 0.62 andPo0.001, F¼ 21.7, r2¼ 0.69 for microbial and spongefractions, respectively). In fact, while the spongefraction was approximately 50% as enriched as the
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1123
The ISME Journal
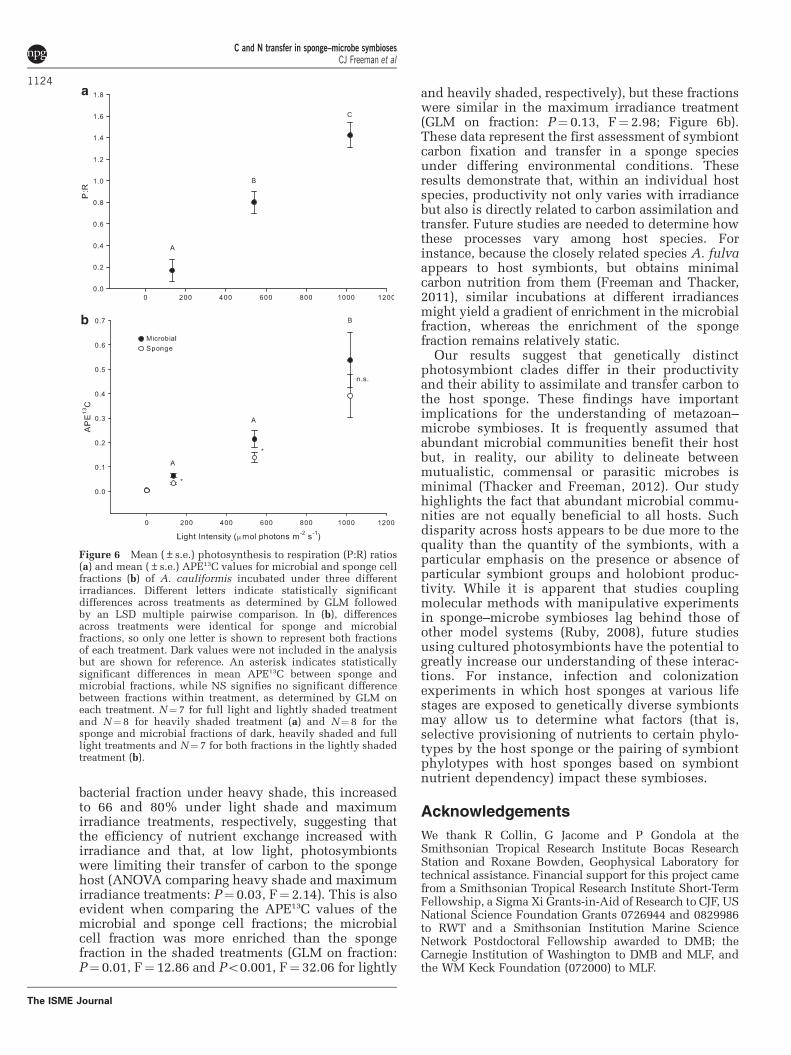
bacterial fraction under heavy shade, this increasedto 66 and 80% under light shade and maximumirradiance treatments, respectively, suggesting thatthe efficiency of nutrient exchange increased withirradiance and that, at low light, photosymbiontswere limiting their transfer of carbon to the spongehost (ANOVA comparing heavy shade and maximumirradiance treatments: P¼ 0.03, F¼ 2.14). This is alsoevident when comparing the APE13C values of themicrobial and sponge cell fractions; the microbialcell fraction was more enriched than the spongefraction in the shaded treatments (GLM on fraction:P¼ 0.01, F¼ 12.86 and Po0.001, F¼ 32.06 for lightly
and heavily shaded, respectively), but these fractionswere similar in the maximum irradiance treatment(GLM on fraction: P¼ 0.13, F¼ 2.98; Figure 6b).These data represent the first assessment of symbiontcarbon fixation and transfer in a sponge speciesunder differing environmental conditions. Theseresults demonstrate that, within an individual hostspecies, productivity not only varies with irradiancebut also is directly related to carbon assimilation andtransfer. Future studies are needed to determine howthese processes vary among host species. Forinstance, because the closely related species A. fulvaappears to host symbionts, but obtains minimalcarbon nutrition from them (Freeman and Thacker,2011), similar incubations at different irradiancesmight yield a gradient of enrichment in the microbialfraction, whereas the enrichment of the spongefraction remains relatively static.
Our results suggest that genetically distinctphotosymbiont clades differ in their productivityand their ability to assimilate and transfer carbon tothe host sponge. These findings have importantimplications for the understanding of metazoan–microbe symbioses. It is frequently assumed thatabundant microbial communities benefit their hostbut, in reality, our ability to delineate betweenmutualistic, commensal or parasitic microbes isminimal (Thacker and Freeman, 2012). Our studyhighlights the fact that abundant microbial commu-nities are not equally beneficial to all hosts. Suchdisparity across hosts appears to be due more to thequality than the quantity of the symbionts, with aparticular emphasis on the presence or absence ofparticular symbiont groups and holobiont produc-tivity. While it is apparent that studies couplingmolecular methods with manipulative experimentsin sponge–microbe symbioses lag behind those ofother model systems (Ruby, 2008), future studiesusing cultured photosymbionts have the potential togreatly increase our understanding of these interac-tions. For instance, infection and colonizationexperiments in which host sponges at various lifestages are exposed to genetically diverse symbiontsmay allow us to determine what factors (that is,selective provisioning of nutrients to certain phylo-types by the host sponge or the pairing of symbiontphylotypes with host sponges based on symbiontnutrient dependency) impact these symbioses.
Acknowledgements
We thank R Collin, G Jacome and P Gondola at theSmithsonian Tropical Research Institute Bocas ResearchStation and Roxane Bowden, Geophysical Laboratory fortechnical assistance. Financial support for this project camefrom a Smithsonian Tropical Research Institute Short-TermFellowship, a Sigma Xi Grants-in-Aid of Research to CJF, USNational Science Foundation Grants 0726944 and 0829986to RWT and a Smithsonian Institution Marine ScienceNetwork Postdoctoral Fellowship awarded to DMB; theCarnegie Institution of Washington to DMB and MLF, andthe WM Keck Foundation (072000) to MLF.
Figure 6 Mean (±s.e.) photosynthesis to respiration (P:R) ratios(a) and mean (±s.e.) APE13C values for microbial and sponge cellfractions (b) of A. cauliformis incubated under three differentirradiances. Different letters indicate statistically significantdifferences across treatments as determined by GLM followedby an LSD multiple pairwise comparison. In (b), differencesacross treatments were identical for sponge and microbialfractions, so only one letter is shown to represent both fractionsof each treatment. Dark values were not included in the analysisbut are shown for reference. An asterisk indicates statisticallysignificant differences in mean APE13C between sponge andmicrobial fractions, while NS signifies no significant differencebetween fractions within treatment, as determined by GLM oneach treatment. N¼ 7 for full light and lightly shaded treatmentand N¼8 for heavily shaded treatment (a) and N¼8 for thesponge and microbial fractions of dark, heavily shaded and fulllight treatments and N¼7 for both fractions in the lightly shadedtreatment (b).
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1124
The ISME Journal

References
Becerro MA, Paul VJ. (2004). Effects of depth and light onsecondary metabolites and cyanobacterial symbiontsof the sponge Dysidea granulosa. Mar Ecol Prog Ser280: 115–128.
Becerro MA. (2008). Quantitative trends in sponge ecologyresearch. Mar Ecol 29: 67–177.
Boucher DH, James S, Keeler KH. (1982). The ecology ofmutualism. Annu Rev Ecol Syst 13: 315–347.
Bronstein JL. (2001). The costs of mutualism. Amer Zool41: 825–839.
Cowden CC, Peterson CJ. (2009). A multi-mutualist simula-tion: applying biological market models to diversemycorrhizal communities. Ecol Model 220: 1522–1533.
Erwin PM, Thacker RW. (2007). Incidence and identity ofphotosynthetic symbionts in Caribbean coral reefsponge assemblages. J Mar Biol Ass UK 87: 1683–1692.
Erwin PM, Thacker RW. (2008a). Phototrophic nutrition andsymbiont diversity of two Caribbean sponge–cyano-bacteria symbioses. Mar Ecol Prog Ser 362: 139–147.
Erwin PM, Thacker RW. (2008b). Cryptic diversity of thesymbiotic cyanobacterium Synechococcus spon-giarum among sponge hosts. Mol Ecol 17: 2937–2947.
Freeman CJ, Thacker RW. (2011). Complex interactionsbetween marine sponges and their symbiotic microbialcommunities. Limnol Oceanogr 56: 1577–1586.
Fry B. (2006). Stable Isotope Ecology. Springer: New York.Hentschel U, Usher KM, Taylor MW. (2006). Marine
sponges as microbial fermenters. FEMS Microbiol Ecol55: 167–177.
Hill R, Scott A. (2012). The influence of irradiance on theseverity of thermal bleaching in sea anemones thathost anemonefish. Coral Reef 31: 273–284.
Knowlton N, Rohwer F. (2003). Multispecies microbialmutualisms on coral reefs: the host as a habitat. AmNat 162: S51–562.
LaJeunesse TC. (2005). ‘‘Species’’ radiations of symbioticdinoflagellates in the Atlantic and Indo-Pacific since theMiocene–Pliocene transition. Mol Biol Evol 22: 570–581.
Moya A, Pereto J, Gil R, Latorre A. (2008). Learning how tolive together: genomic insights into prokaryote–animalsymbioses. Nat Rev 9: 218–228.
Muscatine L, Cernichiari E. (1969). Assimilation ofphotosynthetic products of zooxanthellae by a reefcoral. Biol Bull 137: 506–523.
Muscatine L, Porter JW, Kaplan IR. (1989). Resourcepartitioning by reef corals as determined from stableisotope composition I. d13C of zooxanthellae andanimal tissue vs depth. Mar Biol 100: 185–193.
Nyholm SV, McFall-Ngai MJ. (2004). The winnowing:establishing the squid–Vibrio symbiosis. Nat RevMicrobiol 2: 623–642.
Rosenberg E, Sharon G, Atad I, Zilber-Rosenberg I. (2010).The evolution of animals and plants via symbiosis withmicroorganisms. Environ Microbiol Rep 2: 500–506.
Ruby EG. (2008). Symbiotic conversations are revealed undergenetic interrogation. Nat Rev Microbiol 6: 752–762.
Sacristan-Soriano O, Banaigs B, Casamayor EO, Becerro MA.(2011). Exploring the links between natural products andbacterial assemblages in the sponge Aplysina aerophoba.Appl Environ Microbiol 77: 862–870.
Siegl A, Kamke J, Hochmuth T, Piel J, Richter M, Liang Cet al. (2011). Single-cell genomics reveals the lifestyle
of Poribacteria, a candidate phylum symbioticallyassociated with marine sponges. ISME J 5: 61–70.
Tanaka Y, Miyajima T, Koike I, Hayashibara Y, Ogawa H.(2006). Translocation and conservation of organicnitrogen within the coral–zooxanthella symbioticsystem of Acropora pulchra, as demonstrated by dualisotope-labeling techniques. J Exp Mar Biol Ecol 336:110–119.
Taylor MW, Hill RT, Piel J, Thacker RW, Hentschel U.(2007b). Soaking it up: the complex lives ofmarine sponges and their microbial associates. ISME J1: 187–190.
Taylor MW, Radax R, Steger D, Wagner M. (2007a).Sponge-associated microorganisms: evolution, ecol-ogy, and biotechnological potential. Microbiol Mol BiolRev 71: 295–347.
Thacker RW, Diaz MC, Rutzler K, Erwin PM, Kimble SJA,Pierce MJ et al. (2007). Phylogenetic relation-ships among the filamentous cyanobacterial sym-bionts of Caribbean sponges and a comparison ofphotosynthetic production between sponges hostingfilamentous and unicellular cyanobacteria. In: Hajdu E,Muricy G (eds) Porifera Research: Biodiversity, Inno-vation, and Sustainability. Museu Nacional: Rio deJaneiro, pp 621–626.
Thacker RW, Freeman CJ. (2012). Sponge–microbe sym-bioses: recent advances and new directions. In:Becerro MA, Uriz MJ, Maldonado M, Turon X (eds),Advances in Sponge Science: Phylogeny, Systematics,Ecology. Advances in Marine Biology Vol. 62. Elsevier:Amsterdam, pp 57–111.
Usher KM. (2008). The ecology and phylogeny of cyanobac-terial symbionts in sponges. Mar Ecol 29: 178–192.
Venn AA, Loram JE, Douglas AE. (2008). Photosyntheticsymbioses in animals. J Exp Bot 59: 1069–1080.
Vrijenhoek RC. (2010). Genetics and evolution of deep-seachemosynthetic bacteria and their invertebrate hosts.In: Kiel S (ed), The Vent and Seep Biota, Topics inGeobiology. Springer: Berlin, pp 15–49.
Webster NS, Blackall LL. (2009). What do we really knowabout sponge–microbial symbioses? ISME J 3: 1–3.
Webster NS, Taylor MW, Behnam F, Lucker S, Rattei T,Whalan S et al. (2010). Deep sequencing revealsexceptional diversity and modes of transmission forbacterial sponge symbionts. Environ Microbiol 12:2070–2082.
Webster NS, Taylor MW. (2012). Marine sponges and theirmicrobial symbionts: love and other relationships.Environ Microbiol 14: 335–346.
Weisz JB, Hentschel U, Lindquist N, Martens CS. (2007).Linking abundance and diversity of sponge-associatedmicrobial communities to metabolic differences inhost sponges. Mar Biol 152: 475–483.
Weisz JB, Massaro AJ, Ramsby BD, Hill MS. (2010).Zooxanthellar symbionts shape host sponge trophicstatus through translocation of carbon. Biol Bull 219:189–197.
Wilkinson CR. (1983). Net primary productivity in coralreef sponges. Science 219: 410–412.
Zilber-Rosenberg I, Rosenberg E. (2008). Role of micro-organisms in the evolution of animals and plants: thehologenome theory of evolution. FEMS Microbiol Rev32: 723–735.
Supplementary Information accompanies the paper on The ISME Journal website (http://www.nature.com/ismej)
C and N transfer in sponge–microbe symbiosesCJ Freeman et al
1125
The ISME Journal
Copyright © 2022 FDOKUMEN




















