
Purification of functional T lymphocytes from splenocytes of the beluga whales ( Delphinapterus...
10
Purification of functional T lymphocytes from splenocytes of the beluga whales (Delphinapterus leucas ) Jacques Bernier a, *, Sylvain De Guise b , Daniel Martineau c , Pierre Be´land d , Martin Beaudet a , Michel Fournier a a INRS-Institut Armand Frappier, Centre Sante ´ humaine, Pointe-Claire, Quebec, Canada b Department of Pathobiology, University of Connecticut, Storrs, CT, USA c Faculte ´ de Me ´decine Ve ´te´rinaire, Universite ´ de Montre ´al, St-Hyacinthe, Quebec, Canada d St. Lawrence National Institute of Ecotoxicology, Montre ´al, Quebec, Canada Received 10 May 1999; received in revised form 17 November 1999; accepted 29 November 1999 Abstract In an eort to gain knowledge on immune functions in beluga whales, Delphinapterus leucas, we have used two physical methods for the purification of T lymphocytes of spleen cells. Isolation by sheep red blood cells (SRBC) rosetting and by adherence on nylon wool columns were tested. SRBC-rosetting gave unreliable results in obtaining purified T cells. Therefore, the purification of T cells was done using nylon wool columns. Less than 3% of the IgM B cells remained in euent populations. In the later population, 45% gave positive staining with mouse anti- human CD4 allowing us to verify functionality of the cells. The study of calcium mobilization and tyrosine kinase activation, mediated by CD4 cross-linking permitted verification of the functionality of cells. We also showed that upon activation with mitogens, beluga T cells upregulate the density of MHC class II molecules on their surfaces. CD4 cross-linking with a specific antibody inhibited the proliferation response. Overall, the activation of beluga whales lymphocytes did not dier markedly from what is known in other species. This study can help in the groundwork for functional investigation of the beluga whale’s immune system. 7 2000 Elsevier Science Ltd. All rights reserved. Keywords: Delphinapterus leucas; Beluga whales; T-cells; Activation Developmental and Comparative Immunology 24 (2000) 653–662 0145-305X/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved. PII: S0145-305X(99)00086-5 www.elsevier.com/locate/devcompimm * Corresponding author. Tel.: +1-514-630-8813; fax: +1-514-630-8850. E-mail address: [email protected] (J. Bernier). Abbreviations: AET, 2-aminoethylisothio-uronium bromide hydrobromide; FBS, fetal bovine serum; Ca 2+ , calcium; CON-A, concancavalin-A; DPM, disintegrations per minute (DPM); gp, glycoprotein; HIV, human immunodeficiency virus; Ig, immunoglo- bulin; INF, interferon; IL-2, interleukin 2; IL-2R, interleukin 2 receptors; kDa, kilo dalton; mAb, monoclonal antibody; MHC, major histocompatibility complex; PLC, phospholipase C; PHA, pPphytohemaglutinin; SRBC, sheep red blood cells; TBS-T, tris– buered solution with 0,01% Tween-20.
Transcript of Purification of functional T lymphocytes from splenocytes of the beluga whales ( Delphinapterus...

Puri®cation of functional T lymphocytes from splenocytesof the beluga whales (Delphinapterus leucas )
Jacques Berniera,*, Sylvain De Guiseb, Daniel Martineauc, Pierre Be landd,Martin Beaudeta, Michel Fourniera
aINRS-Institut Armand Frappier, Centre Sante humaine, Pointe-Claire, Quebec, CanadabDepartment of Pathobiology, University of Connecticut, Storrs, CT, USA
cFaculte de MeÂdecine VeÂteÂrinaire, Universite de MontreÂal, St-Hyacinthe, Quebec, CanadadSt. Lawrence National Institute of Ecotoxicology, MontreÂal, Quebec, Canada
Received 10 May 1999; received in revised form 17 November 1999; accepted 29 November 1999
Abstract
In an e�ort to gain knowledge on immune functions in beluga whales, Delphinapterus leucas, we have used twophysical methods for the puri®cation of T lymphocytes of spleen cells. Isolation by sheep red blood cells (SRBC)rosetting and by adherence on nylon wool columns were tested. SRBC-rosetting gave unreliable results in obtaining
puri®ed T cells. Therefore, the puri®cation of T cells was done using nylon wool columns. Less than 3% of theIgM� B cells remained in e�uent populations. In the later population, 45% gave positive staining with mouse anti-human CD4 allowing us to verify functionality of the cells. The study of calcium mobilization and tyrosine kinase
activation, mediated by CD4 cross-linking permitted veri®cation of the functionality of cells. We also showed thatupon activation with mitogens, beluga T cells upregulate the density of MHC class II molecules on their surfaces.CD4 cross-linking with a speci®c antibody inhibited the proliferation response. Overall, the activation of beluga
whales lymphocytes did not di�er markedly from what is known in other species. This study can help in thegroundwork for functional investigation of the beluga whale's immune system. 7 2000 Elsevier Science Ltd. Allrights reserved.
Keywords: Delphinapterus leucas; Beluga whales; T-cells; Activation
Developmental and Comparative Immunology 24 (2000) 653±662
0145-305X/00/$ - see front matter 7 2000 Elsevier Science Ltd. All rights reserved.
PII: S0145-305X(99 )00086-5
www.elsevier.com/locate/devcompimm
* Corresponding author. Tel.: +1-514-630-8813; fax: +1-514-630-8850.
E-mail address: [email protected] (J. Bernier).
Abbreviations: AET, 2-aminoethylisothio-uronium bromide hydrobromide; FBS, fetal bovine serum; Ca2+, calcium; CON-A,
concancavalin-A; DPM, disintegrations per minute (DPM); gp, glycoprotein; HIV, human immunode®ciency virus; Ig, immunoglo-
bulin; INF, interferon; IL-2, interleukin 2; IL-2R, interleukin 2 receptors; kDa, kilo dalton; mAb, monoclonal antibody; MHC,
major histocompatibility complex; PLC, phospholipase C; PHA, pPphytohemaglutinin; SRBC, sheep red blood cells; TBS-T, tris±
bu�ered solution with 0,01% Tween-20.

1. Introduction
Our knowledge in the ®eld of marine mam-mal immunology has increased in the last dec-ade, especially for cetaceans. The morphologyof lymphoid organs such as the thymus andthe spleen in cetaceans are not di�erent fromthat of other mammals [1±4]. Although di�er-ences reside in the composition of the belugawhale's serum when compared to that of othermammals [5,6], no notable di�erence is foundin the structure of immunoglobulins [2,7±11].We and others have shown that splenocytes,thymocytes and peripheral blood cells can beactivated with common mitogenic lectin, rein-forcing the similarities between the immunesystems of di�erent mammals [12±15]. More-over, our laboratory has demonstrated thatseveral immune parameters of beluga whalessuch as natural killer (NK) activity and pha-gocytosis can be evaluated using well charac-terized assays in rodents and humans withslight modi®cations [15,16]. Furthermore, wefound that NK activity in beluga whales wasenhanced in the presence of human IL-2, indi-cating that the general mechanism occurring inother mammalian species for control of theimmune response also exists in cetaceans [17].
Recently, while using a panel of monoclonalantibody (mAb) speci®c to di�erent CD anti-gens in mammalian species, we have identi®edseveral mAbs speci®c to CD antigens employedin the phenotyping of peripheral blood lym-phocytes of beluga whales [18]. Among them,we have demonstrated that an mAb speci®c tothe D1 domain of the CD4 molecules can beused for the identi®cation of CD4+ T cells.Interestingly, all lymphoid cells express CD2 mol-ecules as reported in rodents, but not in humans[19,20]. On the other hand, the majority of lym-phoid cells are positive for MHC class II mol-ecules as reported for di�erent species such asswine, cats and dogs but not in humans and mice[21±25]. These results indicate the importance ofcharacterizing the immune responses in cetaceansto better understand their immune systems.
In this paper we have used two widely knownmethods to obtain enriched T cells from the
spleen cells of beluga whales. We also addressbiochemical signal pathways in puri®ed T cellsand the functional role of CD4 molecules.
2. Material and methods
2.1. Animals, sampling and cell preparation
Two free ranging beluga whales were collectedby Inuit hunters for consumption purposes inArviat, NWT, Canada. Within 10 min followingthe death of the whales, the spleen was collectedand cut into 1 cm3 pieces that were stored in icecold RPMI 1640 supplemented with 10% fetalbovine serum (FBS), 100 U penicillin and 100 mgstreptomycin. Within 18 h after death, the spleenpieces were minced with forceps into completeRPMI, ®ltered through a gauze and through a®ne coat of nylon wool into a Pasteur pipette,before being suspended in fetal calf serum with7.5% DMSO (Sigma-Aldrich, Oakville, ON,Canada), and aliquoted in cryogenic vials (Nal-gene, Rochester, NY). The cell suspensions werefrozen and stored in liquid nitrogen. Cryopre-served splenocytes from the two free rangingbeluga whales were used for the di�erent assays.For subsequent cell culture, the cells werethawed, washed three times in complete RPMIand live mononuclear cells were isolated by den-sity gradient centrifugation on Ficoll (PharmaciaBiotech, Montre al, QC, Canada) for 35 min at900 � g. Splenocytes were washed three times,resuspended in complete RPMI, enumerated andtheir viability assessed with acridin orange-propi-dium iodide and a ¯uorescence microscope.
2.2. Cell puri®cation
2.2.1. Nylon wool columnsT cells were puri®ed from a spleen cell suspen-
sion using a nylon wool column [26,27]. Brie¯y,the nylon wool was washed with a detergent,then pre-treated with 0.1 N HCl, rinsed anddried at 378C. 0.6 g of dry nylon wool wasplaced in a volume of 5 ml in the barrel of a 10ml syringe (Becton Dickinson, Rutherford, NJ).Columns were washed with warm (378C ) com-
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662654

plete RPMI, ®lled and incubated for 60 min at378C. The columns were further washed with 50ml of warm complete RPMI. 2.8 � 108 belugasplenocytes were suspended in 1 ml of warmcomplete RPMI, placed in the column and incu-bated at 378C for 60 min. The column was elutedwith 30 ml of warm complete RPMI and the Tcells were recollected in a tube.
2.2.2. SRBC separationBeluga whale lymphocytes were separated from
the spleen by rosetting with 2-aminoethylisothio-uronium bromide hydrobromide (AET)-treatedSRBC, as described by Pellegrino et al. [28].Brie¯y, 1 ml of washed SRBC suspension wasincubated with 0.14 M AET (Sigma-Aldrich) for20 min at 378C. SRBC were washed twice andbrought to a 10% SRBC solution in HBSS. Sple-nocytes (50 � 106 cells) were incubated in anequal volume of FBS previously absorbed onSRBC and 1% of AET-SRBC was added. Cellsuspensions were incubated for 10 min at 378C,spun at 500 � g for 10 min and incubated on icefor 1 h. Supernatants were aspirated and the pel-lets were overlaid with 5 ml of Percoll (sp.gr.1.078, Sigma-Aldrich), which were then overlaidwith 5% FBS-PBS and centrifuged at 550� g for20 min. Adherent cells to SRBC were isolatedfrom the pellet by osmotic lysis.
2.3. Immunophenotyping analysis
The T-cell enriched population and the unpuri-®ed splenocytes were then immuno-labeled with aphycoerythrin-labeled goat anti-mouse IgM(FisherBiotech, Orangeburg, NY), with an anti-human CD4 (SIM.4; Dr. Michel Tremblay,Centre Hospitalier de l'Universite Laval, QC,Canada) or with a mouse anti-bovine MHC II(TH21A, VMRD, Pullman, WA) and an FITC-labeled goat anti-mouse IgG (Gibco, Gaithers-berg, MD). We have demonstrated no cross-reac-tivities between FITC-labeled goat anti-mouseand beluga lymphocytes.
2.4. Measurement of intracellular calcium
The T-cell enriched population obtained after
nylon wool puri®cation was used for calcium mo-bilization assays. The cells were adjusted to aconcentration of 5 � 106/ml and were incubatedin presence of 3 mM Fluo-3 AM (MolecularProbes, Eugene, OR) in HBSS with 0.1% bovineserum albumin (BSA, Sigma-Aldrich) for 30 minat 378C. An equal volume of HBSS-BSA wasadded and the incubation was continued for 30min at 378C. The cells were washed and thenanalyzed for their ¯uorescence at 530 nm (FL1)after stimulation with either Concancavalin-A(Con-A, Sigma-Aldrich) or p-phytohemaglutinin(PHA, Sigma-Aldrich) for 17 min using a FACS-can ¯ow cytometer (Becton Dickinson, Ruther-ford, NJ). Results were analyzed with the LYSISII software.
2.5. Analysis of protein tyrosine phosphorylation
Tyrosine phosphorylation of cellular substrateswas analyzed in total cell lysates after CD4 mol-ecule cross-linked with speci®c mAb, SIM 4. Thereactions were terminated by rapid centrifugationand an addition of 100 ml of reducing loadingbu�er to the cell pellets. The proteins were thenresolved by 10% SDS±PAGE and subsequentlytransfered to nitrocellulose membranes (Bio-RadLaboratories, Mississauga, ON, Canada) by elec-troblotting following standard procedure. Mem-branes were blocked for 2 h in Tris±bu�eredsolution supplemented with 0.01% Tween-20(TBS-T) containing 2% gelatin. Phosphotyrosinewere determined with mAb PY-20 (Santa CruzBiotechnology, Santa Cruz, Ca) at a ®nal concen-tration of 1 mg/ml in blocking bu�er for 60 minat room temperature. After several washes inTBS-T, antibody binding was revealed with agoat anti-mouse IgG-horseradish peroxidase(Amersham Pharmacia Biotech, Baie d'Urfe , QC,Canada) for 45 min at room temperature. Anti-gen±antibody complexes were detected using anenhanced chemiluminescence kit (AmershamPharmacia Biotech, Baie d'Urfe , QC, Canada).
2.6. Proliferative response and CD4 cross-linkingexperiments
Beluga splenocytes were cultured with or with-
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662 655

out PHA (1 mg/ml) in the presence of di�erent di-lutions (1:10, 1:100, 1:1000, 1:10 000) of a rabbitpolyclonal antibody produced against humanCD4 (gift by Dr. Michel Tremblay). This anti-body used in ¯uorocytometry analysis gave thesame % of positive cells as obtained with theSIM.4 antibody used in this paper (data notshown) and has been heat inactivated and dia-lysed. No cytotoxic e�ects were demonstrated inhuman cell culture (data not shown). The cellswere incubated for 48 h at 378C with 5% CO2,and pulsed with 0.5 mCi of �3H]-thymidine (6.7Ci/mmol; ICN, Mississauga, ON, Canada) andplates were further incubated for 18 h at 378Cwith 5% CO2. The cells were then ®ltered using aTiterteck cell harvester, and radioactivity wasevaluated with a Beckman scintillation counter.The raw data was expressed in disintegrationsper minute (DPM). The triplicates were averagedand the data is presented as means2standard de-viation.
2.7. Statistical analysis
The signi®cances were con®rmed by Student'st-test. Di�erences were considered signi®cantwhen p was less than 0.05.
3. Results
3.1. Cell puri®cation
Two common methods used in mice and inman to isolate the T-cell populations are theadherence procedure on nylon wool and AET-SRBC rosetting. We have applied these methods
to the splenocytes from beluga whales to obtainenriched populations of T-cells. The adherentand non-adherent cell fractions of the spleen cellsobtained after separation were stained with anti-CD4 and anti-IgM antibodies. The splenocytescontained 24% B lymphocytes, as demonstratedby coloration with the anti-IgM, and 33.12%CD4+ T lymphocytes (Table 1). After isolationby AET-SRBC assay, the proportion of B or T-cells in AET-SRBC non-binding or AET-SRBCbinding remained similar demonstrating the fail-ure of the technique to isolate an enriched popu-lation of T-cells. Separating splenocytes by nylonwool adherence has shown that the adherentpopulation contained 66.48% of B lymphocytesand only 6.30% of CD4+ T lymphocytes. Nylonnon-adherent populations were composed of45.39% of CD4+ T cells and only 2.90% B lym-phocytes. The cells recovered in the non-adherentpopulations represented about 35% of the totalpopulations applied on the columns.
3.2. Calcium mobilization
To determine whether T-cells function after thenylon wool puri®cation, calcium mobilizationwas studied with a FLUO-3 probe and FACSanalysis. The calcium ¯ux in the beluga whale'sT-lymphocytes enriched cell populations in re-sponse to stimulation by either Con-A or PHAwere monitored for 16 min. The Fluo-3 probewas used to determine the rise of intracellularcalcium. The ¯uorescence of the probe increaseswith a magnitude of 100 times after calciumbinding (Fig. 1). Both PHA and ConA stimu-lations caused a rapid increase of intracellularcalcium mobilization as shown by an increase of
Table 1
Isolation of T-cells by SRBC-rosetting and adherence on nylon wool
Cells % CD4 positive cells % IgMs positive cells
Total Spleen cells 33.12 24.00
Non-binding cells to SRBC 32.00 26.00
Binding cells to SRBC 32.80 21.00
Nylon binding cells 6.30 66.48
Nylon non-binding cells 45.39 2.90
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662656
¯uorescence. The majority of cells responded tothe stimuli, indicating that the non-adherent cellsobtained from the nylon wool column were T-cells. Although we had no speci®c pan-antibodiesto T-cells, this result indicates that the cell popu-lation recovered from the nylon wool columnrespond to mitogenic lectin speci®c to T-cells.
3.3. CD4 cross-linking studies
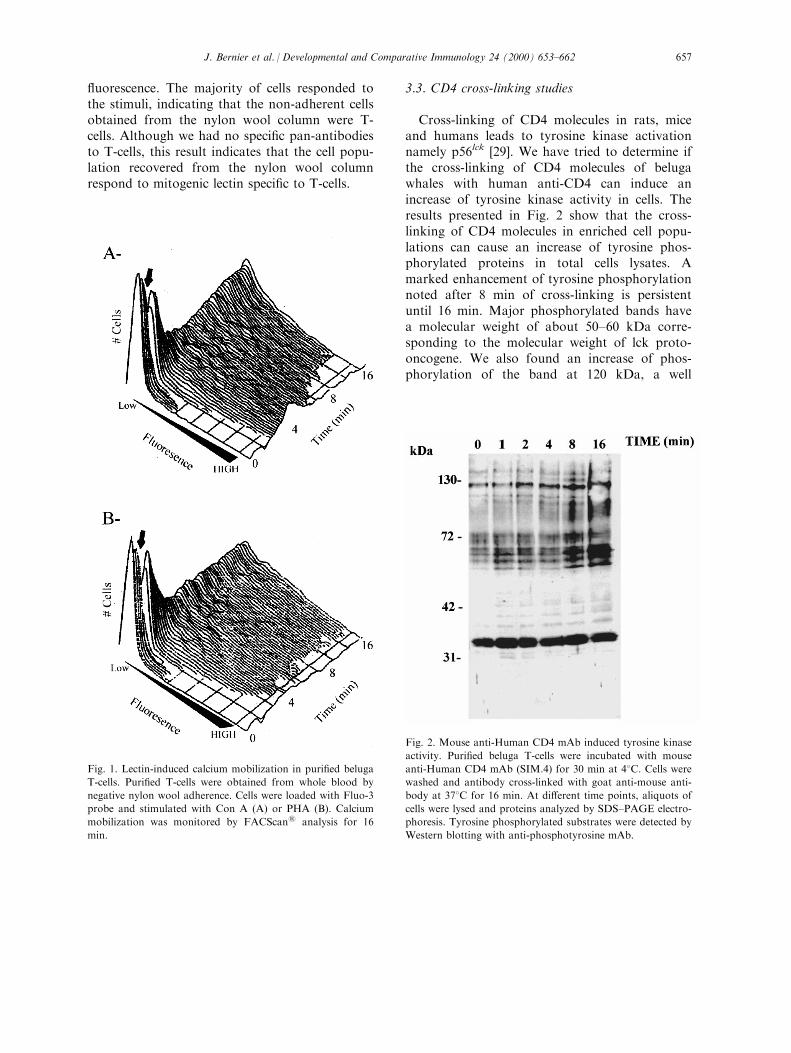
Cross-linking of CD4 molecules in rats, miceand humans leads to tyrosine kinase activationnamely p56lck [29]. We have tried to determine ifthe cross-linking of CD4 molecules of belugawhales with human anti-CD4 can induce anincrease of tyrosine kinase activity in cells. Theresults presented in Fig. 2 show that the cross-linking of CD4 molecules in enriched cell popu-lations can cause an increase of tyrosine phos-phorylated proteins in total cells lysates. Amarked enhancement of tyrosine phosphorylationnoted after 8 min of cross-linking is persistentuntil 16 min. Major phosphorylated bands havea molecular weight of about 50±60 kDa corre-sponding to the molecular weight of lck proto-oncogene. We also found an increase of phos-phorylation of the band at 120 kDa, a well
Fig. 2. Mouse anti-Human CD4 mAb induced tyrosine kinase
activity. Puri®ed beluga T-cells were incubated with mouse
anti-Human CD4 mAb (SIM.4) for 30 min at 48C. Cells werewashed and antibody cross-linked with goat anti-mouse anti-
body at 378C for 16 min. At di�erent time points, aliquots of
cells were lysed and proteins analyzed by SDS±PAGE electro-
phoresis. Tyrosine phosphorylated substrates were detected by
Western blotting with anti-phosphotyrosine mAb.
Fig. 1. Lectin-induced calcium mobilization in puri®ed beluga
T-cells. Puri®ed T-cells were obtained from whole blood by
negative nylon wool adherence. Cells were loaded with Fluo-3
probe and stimulated with Con A (A) or PHA (B). Calcium
mobilization was monitored by FACScan1 analysis for 16
min.
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662 657

recognized consequence of CD4-lck activation[30].
3.4. Upregulation of MHC II in stimulated cells
T-cell activation leads to an increase in ex-pression of several proteins such IL-2 receptorsand transferin receptors. In some species, cell ac-tivation with T-cell mitogens was associated withthe upregulation of MHC class II molecules[23,31]. Since phenotyping analysis has shownthat the majority of lymphoid cells in belugawhale express MHC class II molecules [18], wehave addressed the level of expression of thismolecule after mitogenic stimulation. The stimu-lation of splenocytes with PHA for 48 h resulted
of a marked increase of MHC class II molecule(Fig. 3).
3.5. In¯uence of cross-linked CD4 on cellproliferation
The role of CD4 molecules in the regulation ofT-cell activities are well documented [32]. Wehave determined the e�ects of CD4 cross-linkingin beluga whale T-cells activation. Stimulation ofsplenocytes with an increasing amount of humanCD4 speci®c polyclonal antibody in the presenceof PHA caused an inhibition of the proliferativeresponse as compared to pre-immune serum(Fig. 4). This inhibition was dose-dependant andcan be observed at the lowest dilution of 1/10,000 of immune serum. Anti-CD4 dilutions at1/100 caused an inhibition of 75% of normal re-sponses and dilutions below 1/100 abrogated thecell proliferation. No di�erences regarding theproliferative responses were observed with thepre-immune serum.
4. Discussion
The use of puri®ed cell populations is a com-mon method to understand immune responsesand to characterize the changes in the immuneparameters at the cellular level. Since no T cell-speci®c antibody was available to obtain a puri-®ed population, we have applied two convention-al techniques for the isolation of this population.Isolation by SRBC rosetting failed to provide anenriched T-cell population (Table 1). This resultis not surprising since all spleen cells expressCD2, the key molecule in adherence to AET-SRBC [33]. Separation of T and B lymphocyteson nylon wool columns gave the best results.Although no T pan-antibody existed to con®rmthe purity of the cell populations, the staining ofe�uent populations showed that IgM positivecells represented only 3% as compared to CD4+
which constituted 45% of cells obtained. Adher-ent population was positive at 67% for IgM ex-pression. The proportion of B cells obtained ine�uent population is in the range of results
Fig. 3. PHA stimulation of beluga whale's splenocytes caused
an increase of MHC class II expression. Spleen cells were
stimulated with PHA for three days and the expression of
MHC class II molecule determined by immunostaining and
FACS analysis. Unstimulated T-cells (A) or PHA-stimulated
T-cells (B).
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662658

reported for mice and humans (5% B cells inpuri®ed T cells; [27]).
The functionality of lymphocytes after thepuri®cation procedure was established by studiesof the biochemical signaling pathways. We ®rststudied calcium ¯ux after lectin stimulation. Ourresults showed a rapid and sustained increase ofcytoplasmic calcium after stimulation. The re-sponse obtained con®rmed the purity of the e�u-ent population since a majority of the cellsresponded to these T-cells mitogens.
In several mammalian species, CD4 moleculesare associated in the intracellular portion withtyrosine kinase proto-oncogen p56lck, a memberof the src family [34,35]. The cross-linking ofCD4 molecules of an enriched population with apolyclonal antibody leads to an increase of theintracytoplasmic protein phosphorylated on tyro-sine residues. Major phosphorylations wereobserved around 50±65 kDa, this molecularweight corresponding to several tyrosines kinasespresent in T-cells, such as lck and fyn. Interest-ingly, a major phosphorylated band at 120 kDawas present after 2 min of stimulation. This mol-
ecular weight corresponds to PLC-g, a well-characterized substrate of p56lck [30]. Moreover,Ca2+ mobilization is triggered mainly by a sec-ond messenger, inositol 1,4,5-triphosphate, uponcell stimulation and activation of PLC-g, [36].Activation of tyrosine kinase has been demon-strated to take place before the calcium ¯ux [37].Cross-linking of CD4 molecules does not inducecalcium ¯ux, but leads to p56lck activity.Although we failed to demonstrate the presenceof p56lck with an antibody speci®c to the humanisoform (data not shown), our results illustratedthat the cross-linking of CD4 molecules is associ-ated with an increase of tyrosine kinase activity.
To better understand the activation of T lym-phocytes from beluga whales we have monitoredthe modulation of the cell's surface expression ofMHC class II and the consequences of CD4cross-linking on the proliferative response. Thestimulation of splenocytes or peripheral bloodcells with mitogenic lectins caused an increase ofseveral molecules on the cells surface such as IL-2R and transferin receptor [38]. No antibodyspeci®c to IL-2R of mouse, rat or human origin
Fig. 4. CD4 molecules cross-linking inhibit the mitogenic response of beluga whales splenocytes. Spleen cells were stimulated with
PHA in the presence of an increasing amount of cross-reactive rabbit polyclonal antibody speci®c to human CD4 for 48 h. Cells
were pulsed with �3H]- thymidine for an additional 18 h. Results are presented as mean DPM from triplicates. Signi®cant (�: p<0.05, �� p<0.001).
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662 659

reacted with beluga IL-2R (data not shown)although our results on NK activity showed thathuman IL-2 increased the response [17]. Acti-vation of human lymphocyte was also associatedwith the upregulation of the expression of othermolecules such as MHC class II [31]. WhileMHC II is present on virtually 100% of the per-ipheral blood lymphocytes [18] and the spleno-cytes (Fig. 3) of beluga whales, the stimulationwith PHA increased the intensity of the ex-pression of the molecule at the cell's surface. Anincreased proportion of cells expressing thedi�erent forms of MHC class II has also beenobserved in human T lymphocytes stimulated byPHA and IL-2 compared to unstimulated cells[31]. A similar upregulation of the proportion ofcells expressing MHC class II was noted in nor-mal cat peripheral blood lymphocytes stimulatedin vitro with PHA and IL-2 compared to unsti-mulated cells [23]. The increase in expression ofMHC class II upon stimulation in beluga whalesappears to be a mechanism shared with othermammalian species. Since our phenotypingincluded all splenocytes, our results suggested thepossibility that the production of cytokines suchas INF-g might be responsible for the upregula-tion of these molecules. The e�ects of INF-g onMHC class II expression is well documented[39,40].
Lymphocyte proliferation upon stimulation byPHA was partly inhibited by cross-linking of theCD4 receptor in beluga whales. Such an inhi-bition of T-cell proliferation was also observed inhumans by di�erent anti-CD4 monoclonal anti-bodies [41,42]. CD4 molecules play the role ofco-receptors in T cells activation [43]. In somecircumstances, CD4 molecules are associatedwith a negative regulatory role [43]. Severalauthors have suggested that the cross-linking ofCD4 dissociated the signaling pathway mediatedby TCR resulting in a negative e�ect [43]. Ourresults on the activation of tyrosine kinase afterCD4 oligomerization and the negative role in theproliferative response has shown that the re-sponse of T-cells in beluga whales was similar tothe response obtained in other mammalianspecies like humans and mice.
The immune system of cetaceans seems to be
similar to that of other mammals. Ultrastructuralstudy of the blood cells revealed that all majorperipheral cell types in circulation are commonto all mammalian species [44]. Proteins expressedon the lymphocyte's cell surface such as CD45have similar isoform distribution as observed inthat of humans and mice [45]. With regard to T-cells, the CD4 molecule of beluga whales shareda common epitope with humans as suggested bythe positive reaction obtained with SIM.4 mono-clonal antibody. Interestingly, this antibody mapsat the binding site of HIV-gp120 molecules [46].However, no binding of HIV-gp120 in belugawhales was observed (data not shown). Anti-CD4 polyclonal antibody produced in rabbit wasobtained after immunization with recombinantdomains D1 and D2 of human CD4 (Dr Trem-blay, personal communication) demonstratingthat these two domains were conserved duringevolution. Amino acid comparative analysis in®ve di�erent species (human, Rhesus, Chimpan-zee, Mouse and Rat) demonstrates a homologybetween 50% and 89% with the extracellulardomain of human CD4. Although the cetaceansdiverged evolutionarily from other mammalsover 55 million years ago [47], their immune sys-tems seem to be similar.
In this paper we have shown that the belugawhale's T-lymphocytes could be puri®ed usingnylon wool columns, and that the di�erent stepsof activation of T-lymphocytes can be studied.No major di�erences were noted with the mech-anisms known in other mammalian species. Thisstudy can help in laying the groundwork forfunctional investigations of the beluga whale'simmune system, particularly as it relates to thedi�erences between healthy and stranded ani-mals.
Acknowledgements
The authors wish to acknowledge the ArviatHunters and Trappers Association for their helpon sampling Arctic belugas in Western HudsonBay. This project was funded by the U.S. En-vironmental Protection Agency, Canadian Wild-life Toxicology Fund, Environment Canada,
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662660

World Wildlife Fund-Washington. Biodoà me deMontre al and Ordre des Me decins ve te rinairesdu Que bec.
References
[1] Simpson JG, Gardner MB. Comparative anatomy of
selected marine mammals. In: Ridgway SH, editor.
Mammals of the sea: biology and medicine. CC Thomas:
Spring®eld, 1972. p. 298±418.
[2] Cavagnolo RZ. The immunology of marine mammals.
Dev Comp Immunol 1979;3:245±57.
[3] Schumacher U, Welsch U. Histological, histochemical,
and ®ne structural observations on the spleen of seals.
Am J Anat 1987;179:356±68.
[4] Romano T, Felten SY, Olschowka JA, Felten DL. A
microscopic investigation of the lymphoid organs of the
beluga, Delphinapterus leucas. J Morphol 1993;215:261±
87.
[5] Cornell LH, Du�eld DS, Joseph BE, Stark B, Cornell et
al. Hematology and serum chemistry values in the beluga
(Delphinapterus leucas ). J. Wildl. Dis. 1988;24:220±4.
[6] Cook RA, Stoskopf Mk, Dierenfeld ES. Circulating
levels of vitamin E, cholesterol, and selected minerals in
captive and wild beluga whales (Delphinapterus leucas ). J
Zoo Wild Med 1990;21:65±9.
[7] Nash DR, Mack J-P. Immunoglobulin classes in aquatic
mammals characterized by serologic cross-reactivity, mol-
ecular size and binding of human free secretory com-
ponent. J Immunol 1971;107:1424±30.
[8] Suer LD, Vedros NA, Schroeder JP, Dunn JL.
Erysipelothrix rhusiopathiae. Part II: Enzyme immunoas-
say of sera from wild and captive marine mammals. Dis
Aquat Organ 1988;5:7±13.
[9] Travis JC, Sanders BG. Whale immunoglobulins. Part II:
Heavy chain structure. Comp Biochem Physiol
1972;43B:637.
[10] Andresdottir V, Magnadottir B, Andresson O, Petursson
G. Subclasses of IgG from whales. Dev Comp Immunol
1986;11:801±6.
[11] Boyden A, Gemeroy D. The relative position of the ceta-
cea among the other of mammalia as indicated by preci-
pitin tests. Zoologica 1950;35:145.
[12] Mumford DM, Stockman GD, Barsales PB, Whitman T,
Wilbur JR. Lymphocyte transformation studies of sea
mammal blood. Experientia 1975;31:498±500.
[13] Colgrove GS. Stimulation of lymphocytes from a dolphin
(Tursiops truncatus ) by phytomitogens. Am J Vet Res
1978;39:141±4.
[14] Lahvis GP, Wells RS, Casper D, Via CS. In-vitro lym-
phocyte response of bottlenose dolphins (Tursiops trunca-
tus ): mitogen-induced proliferation. Mar Env Res
1993;35:115±9.
[15] De Guise S, Flipo D, Boehm J, Martineau D, Be land P,
Fournier M. Immune functions in beluga whales
(Delphinapterus leucas ): evaluation of phagocytosis and
respiratory burst with peripheral blood using ¯ow cyto-
metry. Vet Immunol Immunopathol 1995;47:351±62.
[16] De Guise S, Bernier J, Dufresne MM, Martineau D,
Be land P, Fournier M. Immune functions in beluga
whales (Delphinapterus leucas ): evaluation of mitogen-
induced blastic transformation of lymphocytes from per-
ipheral blood, spleen and thymus. Vet Immunol
Immunopathol 1996;50:117±26.
[17] De Guise S, Ross PS, Osterhaus ADME, Martineau D,
Be land P, Fournier M. Immune functions in beluga
whales (Delphinapterus leucas ): evaluation of natural
killer (NK) cell activity. Vet Immunol Immunopathol
1997;58:345±54.
[18] De Guise S, Bernier J, Martineau D, Beland P, Fournier
M. Phenotyping of beluga whale blood lymphocytes
using monoclonal antibodies. Dev Comp Immunol
1997;21:425±33.
[19] Yagita H, Nakamura T, Karasuyama H, Okumura K.
Monoclonal antibodies speci®c for murine CD2 reveal its
presence on B as well as T cells. Proc Natl Acad Sci
USA 1989;86:645±9.
[20] Sen J, Rosenberg N, Burako� SJ. Expression and onto-
geny of CD2 on murine B cells. J Immunol
1990;144:2925±30.
[21] Tizard I. In: Veterinary immunology: an introduction.
4th ed. W. B. Saunders: Montreal, 1992.
[22] Barbis DP, Bainbridge D, Crump AL, Zhang CH,
Antczak DF. Variation in expression of MHC class II
antigens on horse lymphocytesdetermined by MHC hap-
lotype. Vet Immunol Immunopathol 1994;42:103±14.
[23] Rideout BA, Moore PF, Pedersen NC. Persistent upregu-
lation of MHC class II antigen expression on T-lympho-
cytes from cats experimentally infected with feline
immunode®ciency virus. Vet Immunol Immunopathol
1992;35:71±81.
[24] Hewitt CRA, Fedlman M. Human T cell clones present
antigen. J Immunol 1989;142:1429±36.
[25] Klein J, Figueroa F. Evolution of the major histocom-
patibility complex. Crit Rev Immunol 1986;6:295±386.
[26] Julius MH, Simpson E, Herzenberg LA. A rapid method
for the isolation of functional thymus-derived murine
lymphocytes. Eur J Immunol 1973;3:645±9.
[27] Trizio D, Cudkowicz G. Separation of T and B lympho-
cytes by nylon wool columns: evaluation of e�cacy by
functional assays in vivo. J Immunol 1974;113:1093±7.
[28] Pellegrino MA, Ferrone S, Theo®lopoulos AN. Isolation
of human T and B lymphocytes by rosette formation
with 2-aminoethylisothiquronium bromide (AET) -trea-
ted sheep red blood cells with monkey red blood cells. J
Immunol Methods 1976;11:273±9.
[29] Ravichandran KS, Collins TL, Burako� SJ. CD4 and
signal transduction. Curr Top Microbiol Immunol
1996;205:47±62.
[30] Weber JR, Bell GM, Han MY, Pawson T, Imboden JB.
Association of the tyrosine kinase LCK with phospho-
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662 661

lipase C-gamma 1 after stimulation of the T cell antigen
receptor. J Exp Med 1992;176:373±9.
[31] Gansbacher B, Zier KS. Regulation of HLA-DR, DP,
and DQ expression in activated T cells. Cell Immunol
1988;117:22±34.
[32] Killeen N, Littman DR. The regulation and function of
the CD4 coreceptor during T lymphocyte. Curr Top
Microbiol Immunol 1996;205:89±106.
[33] Plunkett ML, Sanders ME, Selvaraj P, Dustin ML,
Springer TA. Rosetting of activated human T lympho-
cytes with autologous erythrocytes. De®nition of the
receptor and ligand molecules as CD2 and lymphocyte-
function-associated antigen 3 (LFA-3). J Exp Med
1987;165:664±76.
[34] Veillette A, Abraham N, Caron L, Davidson D. The
lymphocyte-speci®c tyrosine protein kinase p56lck. Semin
Immunol 1991;3:143±52.
[35] Bolen JB, Veillette A. A function for the lck proto-onco-
gene. Trends Biochem Sci 1989;14:404±7.
[36] Weiss A, Littman DR. Signal transduction by lympho-
cyte antigen receptors. Cell 1994;76:263±74.
[37] June CH, Fletcher MC, Ledbetter JA, Schieven GL,
Siegel JN, Phillips AF, Samelson LE. Inhibition of tyro-
sine phosphorylation prevents T-cell receptor-mediated
signal transduction. Proc Natl Acad Sci USA
1990;87:7722±6.
[38] Alberola-Ila J, Places L, de la Calle O, Romero M,
Yague J, Gallart T, Vives J, Lozano F. Stimulation
through the TCR/CD3 complex up-regulates the CD2
surface expression on human T lymphocytes. J Immunol
1991;146:1085±92.
[39] Guo TL, Mudzinski SP, Lawrence DA. Regulation of
HLA-DR and invariant chain expression by human per-
ipheral blood mononuclear cells with lead, interferon-
gamma, or interleukin-4. Cell Immunol 1996;171:1±9.
[40] Basham TY, Merigan TC. Recombinant interferon-
gamma increases HLA-DR synthesis and expression. J
Immunol 1983;130:1492±4.
[41] Bank I, Chess L. Perturbation of the T4 molecule trans-
mits a negative signal to T cells. J Exp Med
1985;162:1294±303.
[42] Neudorf SML, Jones MM, McCarthy BM, Harmony
JA, Choi EM. The CD4 molecule transmits biochemical
information important in the regulation of T lymphocyte
activity. Cell Immunol 1990;125:301±14.
[43] Julius M, Maroun CR, Haughn L. Distinct roles for
CD4 and CD8 as co-receptors in antigen receptor signal-
ling. Immunol Today 1993;14:177±83.
[44] Williams CR, Chapman GB, Blake AS. Ultrastructural
study of the blood cells of the beluga whale,
Delphinapterus leucas. J Morphol 1991;209:97±110.
[45] De Guise S, Erickson K, Blanchard M, Dimolfetto L,
Lepper H, Wang J, Stott JL, Ferrick DA.
Characterization of a monoclonal antibody that recog-
nizes a lymphocyte surface antigen for the cetacean hom-
ologue to CD45R. Immunology 1998;94:207±12.
[46] Barbeau B, Fortin JF, Genois N, Tremblay MJ.
Modulation of human immunode®ciency virus type 1-
induced syncytium formation by the conformational
state of LFA-1 determined by a new luciferase-based syn-
cytium quantitative assay. J Virol 1998;72:7125±36.
[47] Romano TA, Felten SY, Olschowka JA, Felten DL. A
microscopic investigation of the lymphoid organs of the
beluga, Delphinapterus leucas. J Morphol 1993;215:261±
87.
J. Bernier et al. / Developmental and Comparative Immunology 24 (2000) 653±662662




















