
Direct binding of IL-12 to human and murine B lymphocytes
8
International Immunology, Vol. 8, No. 12, pp. 1955-1962 © 1996 Oxford University Press Direct binding of IL-12 to human and murine B lymphocytes Laura A. Vogel 2 , Louise C. Showe 1 , Terry L. Lester, Renee M. McNutt, Victor H. Van Cleave 3 and Dennis W. Metzger Department of Microbiology and Immunology, Medical College of Ohio, Toledo, OH 43699-0008, USA 1 The Wistar Institute, Philadelphia, PA 19104, USA 2 Pre-Clinical Biology Department, Genetics Institute, Cambridge, MA 01810, USA 3 Present address: Department of Microbiology, Dartmouth Medical School, Lebanon, NH 03756, USA Keywords: B lymphocytes, B-1 lymphocytes, cytokine receptors, IL-12 Abstract IL-12 has been shown to play a central role in cell-mediated inflammatory reactions through direct activation of T cells and NK cells. IL-12 also strongly influences humoral immunity but these effects have been thought to be indirect and caused by Intermediary cytokines. Using flow cytometry, we now show that IL-12 directly interacts with B cells. Freshly isolated murine peritoneal B-1 and conventional B lymphocytes bound IL-12, but splenic B cells failed to react unless first stimulated with lipopolysaccharide. All murine B cell sources were found to express IL-12R p1 subunit transcripts as detected by PCR and RNase protection assays. IL-12 binding was also detected on phytohemagglutinin-stimulated human T cell blasts and Staphylococcus aureus/ IL-2-stimulated B cell blasts but not on freshly isolated peripheral blood lymphocytes. Similarly, IL-12 directly bound to the human SKW6.4 Burkitt's B cell lymphoma line. In all cases positive staining was ablated by omitting IL-12 from the procedure, showing that it was not due to detection of endogenous IL-12. These findings indicate that B cells represent another major target for IL-12 in addition to T and NK cells, and that IL-12 can directly affect humoral immunity. Introduction IL-12 is being increasingly recognized as a pivotal cytokine that has dramatic and novel effects on the control of cellular immunity (reviewed in 1-3). Recent studies have shown that it also strongly regulates humoral immunity (4-7). Treatment with IL-12 has been found to cause long-term enhancement of antibody responses to protein and hapten antigens (6,8), while suppressing the in vivo and in vitro function of B-1 cells, the subset of B cells that is reported to be critically involved in the development of autoimmunity and chronic lymphocytic leukemia (8,9). It is generally believed that IL-12 mediates its effects on B cells through induction of intermediary cyto- kines, particularly IFN-y, rather than through direct binding (10). IFN-y is known to enhance production of lgG2a and to inhibit B-1 cell IgM secretion (11-14). However, we (6) and others (4,5,15) have found that the observed influences of IL-12 on antibody production are either not affected or incompletely inhibited by anti-IFN-y antibodies. Furthermore, a recent study (16) demonstrated that IL-12 acts as a co- stimulator for growth of purified human B cells. Because of this finding and the questionable role of IFN-y in mediating all of the effects of IL-12 on humoral immunity, we considered the possibility that IL-12 interacts directly with B cells. A previous study in humans identified an IL-12R on activated T cells and NK cells, but failed to detect any reactivity of IL-12 with B cells or B cell lines (10). However, the B cell-activating agents used in that study failed to induce significant B cell proliferation. Furthermore, not all B cell subsets were examined. Recently, Wu et al. (17) showed that a human Burkitt's lymphoma cell line and activated tonsillar B cells bear a subunit of the IL-12R but these investigators could not detect cellular binding of radiolabeled IL-12. We now demonstrate direct binding of IL-12 to human and murine B cells. This activity was detected with freshly isolated murine peritoneal B cells as well as appropriately activated murine splenic and human peripheral blood B cells. In addition, we show by RT-PCR and ribonuclease protection assay (RPA) that murine B cells express IL-12R |31 chain transcripts. The results suggest that B cells express an IL- Correspondence to: D. W. Metzger Transmitting editor. E. Sercarz Received 9 January 1996, accepted 6 September 1996 by guest on July 13, 2011 intimm.oxfordjournals.org Downloaded from
Transcript of Direct binding of IL-12 to human and murine B lymphocytes

International Immunology, Vol. 8, No. 12, pp. 1955-1962 © 1996 Oxford University Press
Direct binding of IL-12 to human and murineB lymphocytesLaura A. Vogel2, Louise C. Showe1, Terry L. Lester, Renee M. McNutt,Victor H. Van Cleave3 and Dennis W. Metzger
Department of Microbiology and Immunology, Medical College of Ohio, Toledo, OH 43699-0008, USA1The Wistar Institute, Philadelphia, PA 19104, USA
2Pre-Clinical Biology Department, Genetics Institute, Cambridge, MA 01810, USA
3Present address: Department of Microbiology, Dartmouth Medical School, Lebanon, NH 03756, USA
Keywords: B lymphocytes, B-1 lymphocytes, cytokine receptors, IL-12
Abstract
IL-12 has been shown to play a central role in cell-mediated inflammatory reactions through directactivation of T cells and NK cells. IL-12 also strongly influences humoral immunity but theseeffects have been thought to be indirect and caused by Intermediary cytokines. Using flowcytometry, we now show that IL-12 directly interacts with B cells. Freshly isolated murineperitoneal B-1 and conventional B lymphocytes bound IL-12, but splenic B cells failed to reactunless first stimulated with lipopolysaccharide. All murine B cell sources were found to expressIL-12R p1 subunit transcripts as detected by PCR and RNase protection assays. IL-12 binding wasalso detected on phytohemagglutinin-stimulated human T cell blasts and Staphylococcus aureus/IL-2-stimulated B cell blasts but not on freshly isolated peripheral blood lymphocytes. Similarly,IL-12 directly bound to the human SKW6.4 Burkitt's B cell lymphoma line. In all cases positivestaining was ablated by omitting IL-12 from the procedure, showing that it was not due to detectionof endogenous IL-12. These findings indicate that B cells represent another major target for IL-12in addition to T and NK cells, and that IL-12 can directly affect humoral immunity.
Introduction
IL-12 is being increasingly recognized as a pivotal cytokinethat has dramatic and novel effects on the control of cellularimmunity (reviewed in 1-3). Recent studies have shown thatit also strongly regulates humoral immunity (4-7). Treatmentwith IL-12 has been found to cause long-term enhancementof antibody responses to protein and hapten antigens (6,8),while suppressing the in vivo and in vitro function of B-1 cells,the subset of B cells that is reported to be critically involvedin the development of autoimmunity and chronic lymphocyticleukemia (8,9). It is generally believed that IL-12 mediatesits effects on B cells through induction of intermediary cyto-kines, particularly IFN-y, rather than through direct binding(10). IFN-y is known to enhance production of lgG2a and toinhibit B-1 cell IgM secretion (11-14). However, we (6) andothers (4,5,15) have found that the observed influences ofIL-12 on antibody production are either not affected orincompletely inhibited by anti-IFN-y antibodies. Furthermore,a recent study (16) demonstrated that IL-12 acts as a co-stimulator for growth of purified human B cells. Because of
this finding and the questionable role of IFN-y in mediatingall of the effects of IL-12 on humoral immunity, we consideredthe possibility that IL-12 interacts directly with B cells. Aprevious study in humans identified an IL-12R on activated Tcells and NK cells, but failed to detect any reactivity of IL-12with B cells or B cell lines (10). However, the B cell-activatingagents used in that study failed to induce significant Bcell proliferation. Furthermore, not all B cell subsets wereexamined. Recently, Wu et al. (17) showed that a humanBurkitt's lymphoma cell line and activated tonsillar B cellsbear a subunit of the IL-12R but these investigators couldnot detect cellular binding of radiolabeled IL-12. We nowdemonstrate direct binding of IL-12 to human and murineB cells. This activity was detected with freshly isolatedmurine peritoneal B cells as well as appropriately activatedmurine splenic and human peripheral blood B cells. Inaddition, we show by RT-PCR and ribonuclease protectionassay (RPA) that murine B cells express IL-12R |31 chaintranscripts. The results suggest that B cells express an IL-
Correspondence to: D. W. Metzger
Transmitting editor. E. Sercarz Received 9 January 1996, accepted 6 September 1996
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from

1956 IL- 12R on B lymphocytes
12R and represent a major target for direct binding ofthis cytokine.
Methods
Mice
BALB/c mice (6-8 weeks old) were purchased from theNational Cancer Institute (Bethesda, MD) and housed inthe animal facility at the Medical College of Ohio. All miceused weighed at least 15 g.
Cell preparations and cultures
Peritoneal cells (PeC) were obtained from mice by i.p. injectionof 10 ml HBSS containing 0.1% gelatin. Single spleen cell(SpC) suspensions were prepared by passage through a wiremesh and red blood cells were removed by centrifugationon Lympholyte M (Cedarlane Laboratories, Hornby, Ontario,Canada). For activation with lipopolysaccharide (LPS), SpCwere cultured for 3 days at 37°C in 96-well microtiter plates(Costar Corning, Cambridge, MA), with each well containing2X105 cells in 0.2 ml RPMI 1640/10% FCS and 3 ng/mlSalmonella typhimurium LPS (Difco, Detroit, Ml). For enrich-ment of B cells, T and NK cells were depleted from SpCand PeC suspensions by complement-mediated lysis. T celldepletion was performed by incubating cells at 5X106 cells/ml in RPMI 1640-0.3% (w/v) BSA medium containing 2.5%(v/v) rabbit complement (Gibco/BRL, Grand Island, NY), 25%(v/v) anti-CD3 mAb supernatant (clone 145-2C11; ATCC,Rockville, MD) and 25% (v/v) anti-Thy-1.2 mAb supernatant(clone 30-H12; ATCC). NK cells were depleted in a similarmanner using rabbit anti-asialo-GM1 serum (Wako, Richmond,VA) at a 1:1000 dilution. The cells were subjected to two 30min rounds of depletion at 37°C and viable lymphocytes werethen recovered by density centrifugation over Lympholyte-M (Accurate Chemical, Westbury, NY). These procedurestypically reduced levels of T cells to <2% and led to undetect-able levels of IFN-y after culture of the cells with LPS plus IL-12.
Human peripheral blood mononuclear cells were isolatedfrom heparinized venous blood of a healthy adult volunteerby centrifugation on Ficoll-Paque (Pharmacia, Piscataway,NJ). The cells were activated with phytohemagglutinin (PHA)by incubation for 4 days in microtiter plates at a concentrationof 1.Ox 105 cells/well in RPMI 1640/10% FCS and 10 ng/mlPHA-M (Sigma, St Louis, MO). Activation with IL-2 and heat-killed Staphylococcus aureus was performed in a similarmanner by culturing 2.0x105 cells/well for 5 days with 100 U/ml recombinant human IL-2 (R&D Systems, Minneapolis, MN)and a 1.5x 10~5 dilution of heat killed Cowan I Staphylococcusaureus (kindly provided by Dr Michael D. P. Boyle, MedicalCollege of Ohio, Toledo, OH).
Cell lines and antibodies
The SKW6.4 human Burkitt's lymphoma cell line wasobtained from ATCC. The B3B4 anti-CD23 (18) and RA3-6B2.1anti-CD45R (B220) (19) hybridomas were kindly supplied byDrs Daniel H. Conrad (Medical College of Virginia, VirginiaCommonwealth University, Richmond, VA) and Robert L.Coffman (DNAX, Palo Alto, CA) respectively. The 2.4G2 anti-FcRyll (20), 53-7.313 anti-CD5 (21) and M1/70.15.11.5.HL
anti-MAC-1 (22) hybridoma cells were obtained from ATCC.The mAb were purified from culture supernatants by precipita-tion with ammonium sulfate and passage over Protein G-coupled columns (23).
Rabbit anti-mouse and anti-human IL-12 polyclonal anti-bodies were prepared as previously described (24). Briefly,rabbits were immunized s.c. with human or murine IL-12 incomplete Freund's adjuvant and boosted at bi-weekly inter-vals. Antisera were passed over Protein G (Pierce, Rockford,IL) using a FPLC system (Pharmacia, Piscataway, NJ) toisolate Ig. Antibodies were then coupled to biotin or FITC asdescribed (23). Anti-mouse IgM antibody conjugated to FITCwas purchased from Zymed (San Francisco, CA), anti-mouseCD69 mAb conjugated to FITC from PharMingen (San Diego,CA), and anti-human CD3 and CD19 mAb conjugated to FITCfrom Caltag (South San Francisco, CA).
Flow cytometry
Flow cytometry for detection of IL-12 binding was performedusing a sandwich procedure similar to that of Desai et al.(10). For staining of mouse lymphocytes, 5x105 PeC or SpCwere incubated for 45 min on ice in 50 \i\ of 500 nMmurine IL-12 that was diluted in 2.4G2 anti-FcR-yll mAbsupernatant to block FcR binding. Cells were washed withHBSS containing 0.1% (w/v) gelatin and then incubated with1 |ig biotinylated rabbit anti-murine IL-12 antibody or normalrabbit Ig for 15 min on ice. After washing, mAb conjugated toFITC was added, together with streptavidin-allophycocyanin(SA-APC) (Molecular Probes, Eugene, OR). In some experi-ments, staining was performed using murine IL-5 (R&DSystems, Minneapolis, MN) or IL-6 (Promega, Madison, Wl)instead of IL-12 in the procedure. The stained cells wereanalyzed on a Coulter EPICS Elite flow cytometer and thelymphocytes were gated based on 90° and forward lightscatter.
Human lymphocytes were stained in a similar manner withsome modifications. Cells (5x105) were incubated for 45 minon ice in 100 |xl of 500 nM human IL-12 in 10% normal rabbitserum. The cells were then washed with HBSS containing0.1% (w/v) gelatin and incubated with biotinylated rabbit anti-human IL-12 antibody or normal rabbit Ig for 15 min on ice.After washing, mAb conjugated to FITC was added togetherwith SA-APC for 15 min on ice and the cells were analyzedas described above.
The recombinant murine IL-12 used for these studies wasgenerously provided by Dr Brian Hubbard (Genetics Institute,Cambridge, MA) and was purified from CHO cells asdescribed (25). Murine rlL-12 was >96% pure by SDS-PAGEanalysis and contained <1.2 U/mg of endotoxin activity asdetermined by the Limulus amebocyte assay. Its specificactivity was 4.9x 106 U/mg as determined by a PHA prolifera-tion assay. Recombinant human IL-12 was purified from CHOcells and had a specific activity of 5.26x106 U/mg.
RT-PCR
RNA was extracted from frozen cells using Ultraspec(Biotecx, Houston, TX) according to the manufacturer'srecommendations. RNA was resuspended in diethyl pyro-carbonate-treated water and stored at -20° C. Approximately2 |ig of total cellular RNA was subjected to reverse
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from

IL- 12R on B lymphocytes 1957
PeC SPC
10* 10' f̂ iS1 it!1 ID*
IL-12 + Anti-IL-12
Z"53U
I D " T O * 1 0 * I D *
Anti-IL-12 only
IL-12 + Rabbit Ig
Fig. 1. In vivo expression of an IL-12R on murine B lymphocytes. Fresh PeC (left) and SpC (right) were stained for IL-12 binding by sequentialincubation with IL-12, biotinylated rabbit anti-IL-12 antibody and SA-APC (top row). The cells were then analyzed by flow cytometry. Controlsfor staining specificity included omitting IL-12 from the procedure (middle row) and using biotinylated normal rabbit Ig rather than anti-IL-12antibody (bottom row). Histograms represent CD45R(B220)+ lymphocytes.
transcription in a 20 nl total volume. PCR was performedusing 10% of the RT reaction volume with 0.3 ng of forwardand reverse primers and 1 U Taq polymerase (Gibco/BRL,Gaithersburg, MD) in 100 JLL) for 40 cycles. The p1 receptorchain forward primer sequence was GAACCACACA-CACTGTACCCTG and the reverse primer was TTT-AGTGGGTGGCACGAGCC. An aliquot of the amplifiedproduct was electrophoresed on a 1.5% agarose gel, theDNA was transferred to Immobilon N (Millipore, Bedford,MA), and the blot was hybridized with a 32P-labeled internalprimer CGTGTCGGTGAGGAACCAAACC to confirm thespecificity of the PCR product.
RPA
RPA was performed essentially as described (26). Briefly,antisense RNA probes were prepared using the T7 promoterin Bluescript (Stratagene, La Jolla, CA) or Sp6 promoter inPCRII (Invitrogen, San Diego, CA). The probes were labeledwith [32P]UTP and [32P]CTP, purified on a 6% (w/v) poly-acrylamide sequencing gel, and used the same day. TotalRNA (5-30 jig) was hybridized with 5X106 c.p.m. antisenseprobe in 80% (w/v) formamide buffer at 60°C. The IL-12R (31
probe was generated from a mouse cDNA clone (27) andincluded nucleotides 1301-1621; a mouse cyclophilin anti-sense probe (Ambion, Austin, TX) was used as an internalcontrol for standardization of expression levels betweensamples. Samples were processed as described (26) andfractionated on a 6% (w/v) polyacrylamide sequencing gel,the gel was dried and exposed to a phosphoimager screen,and the relative (31 signals were calculated using Image-Quant (Molecular Dynamics, Sunnyvale, CA).
Results
Binding of IL-12 to murine B cells
The ability of IL-12 to influence humoral immunity has beenthought to be indirect and mediated by IFN-y produced byTh1 cells. IFN-y is known to regulate B cell Ig secretion andto inhibit differentiation of B-1 cells (11-14). However, in manycases, the effects of IL-12 on B lymphocytes cannot be fullyeliminated by treatment with neutralizing anti-IFN-y antibody(4-6,15).
Based on a recent report that IL-12 can directly stimulate
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from
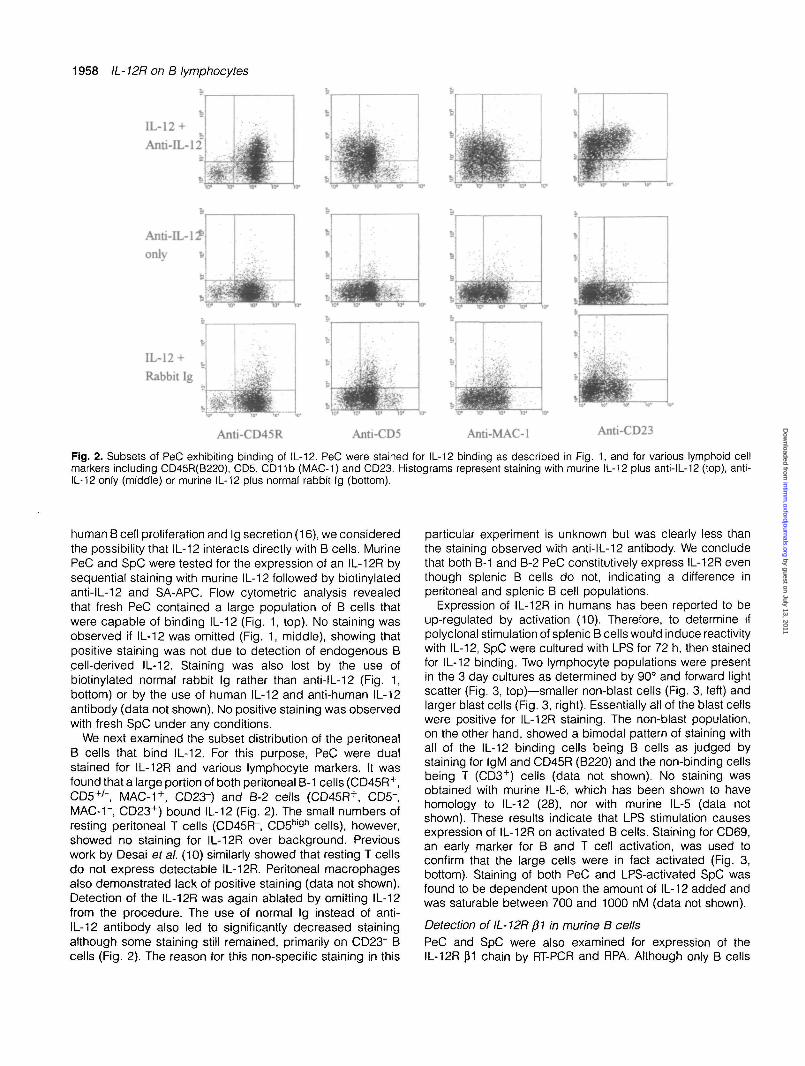
1958 IL- 12R on B lymphocytes
IL-12 +Anti-IL-12
10" 10* IG1
'0* 10*
IL-12 +Rabbit Ig
Anti-CD45R Anti-CD5 Anti-MAC-1 Anti-CD23
Fig. 2. Subsets of PeC exhibiting binding of IL-12. PeC were stained for IL-12 binding as described in Fig. 1, and for various lymphoid cellmarkers including CD45R(B220), CD5, CD11b (MAC-1) and CD23. Histograms represent staining with murine IL-12 plus anti-IL-12 (top), anti-IL-12 only (middle) or murine IL-12 plus normal rabbit Ig (bottom).
human B cell proliferation and Ig secretion (16), we consideredthe possibility that IL-12 interacts directly with B cells. MurinePeC and SpC were tested for the expression of an IL-12R bysequential staining with murine IL-12 followed by biotinylatedanti-IL-12 and SA-APC. Flow cytometric analysis revealedthat fresh PeC contained a large population of B cells thatwere capable of binding IL-12 (Fig. 1, top). No staining wasobserved if IL-12 was omitted (Fig. 1, middle), showing thatpositive staining was not due to detection of endogenous Bcell-derived IL-12. Staining was also lost by the use ofbiotinylated normal rabbit Ig rather than anti-IL-12 (Fig. 1,bottom) or by the use of human IL-12 and anti-human IL-12antibody (data not shown). No positive staining was observedwith fresh SpC under any conditions.
We next examined the subset distribution of the peritonealB cells that bind IL-12. For this purpose, PeC were dualstained for IL-12R and various lymphocyte markers. It wasfound that a large portion of both peritoneal B-1 cells (CD45R+,CD5+/", MAC-1+, CD23") and B-2 cells (CD45R+, CD5",MAC-r, CD23+) bound IL-12 (Fig. 2). The small numbers ofresting peritoneal T cells (CD45R", CD5h'9h cells), however,showed no staining for IL-12R over background. Previouswork by Desai ef al. (10) similarly showed that resting T cellsdo not express detectable IL-12R. Peritoneal macrophagesalso demonstrated lack of positive staining (data not shown).Detection of the IL-12R was again ablated by omitting IL-12from the procedure. The use of normal Ig instead of anti-IL-12 antibody also led to significantly decreased stainingalthough some staining still remained, primarily on CD23" Bcells (Fig. 2). The reason for this non-specific staining in this
particular experiment is unknown but was clearly less thanthe staining observed with anti-IL-12 antibody. We concludethat both B-1 and B-2 PeC constitutively express IL-12R eventhough splenic B cells do not, indicating a difference inperitoneal and splenic B cell populations.
Expression of IL-12R in humans has been reported to beup-regulated by activation (10). Therefore, to determine ifpolyclonal stimulation of splenic B cells would induce reactivitywith IL-12, SpC were cultured with LPS for 72 h, then stainedfor IL-12 binding. Two lymphocyte populations were presentin the 3 day cultures as determined by 90° and forward lightscatter (Fig. 3, top)—smaller non-blast cells (Fig. 3, left) andlarger blast cells (Fig. 3, right). Essentially all of the blast cellswere positive for IL-12R staining. The non-blast population,on the other hand, showed a bimodal pattern of staining withall of the IL-12 binding cells being B cells as judged bystaining for IgM and CD45R (B220) and the non-binding cellsbeing T (CD3+) cells (data not shown). No staining wasobtained with murine IL-6, which has been shown to havehomology to IL-12 (28), nor with murine IL-5 (data notshown). These results indicate that LPS stimulation causesexpression of IL-12R on activated B cells. Staining for CD69,an early marker for B and T cell activation, was used toconfirm that the large cells were in fact activated (Fig. 3,bottom). Staining of both PeC and LPS-activated SpC wasfound to be dependent upon the amount of IL-12 added andwas saturable between 700 and 1000 nM (data not shown).
Detection of IL- 12R p1 in murine B cellsPeC and SpC were also examined for expression of theIL-12R pi chain by RT-PCR and RPA. Although only B cells
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from
IL- 12R on B lymphocytes 1959
FALS
!£• - • • -
101 ID* W ID* ° IQ* ~^ xa* ^ ~ \o* io* ib
IL-12R Fluorescence Intensity
CD69 Fluorescence Intensity
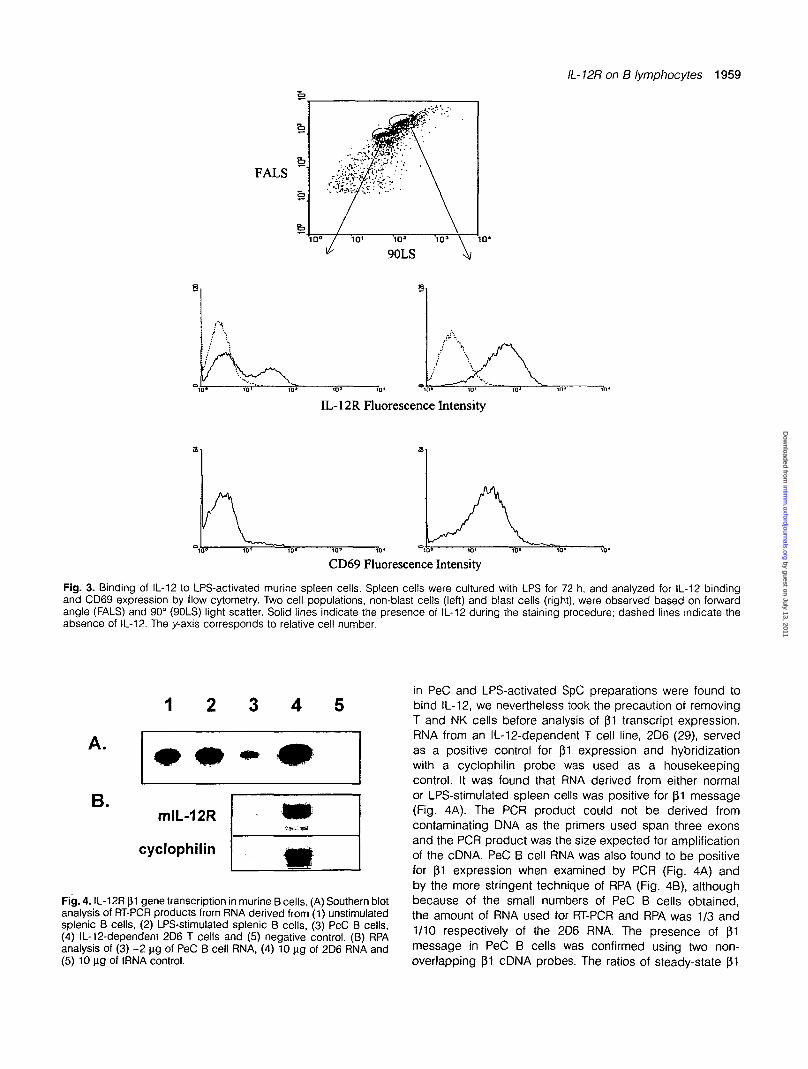
Fig. 3. Binding of IL-12 to LPS-activated murine spleen cells. Spleen cells were cultured with LPS for 72 h, and analyzed for IL-12 bindingand CD69 expression by flow cytometry. Two cell populations, non-blast cells (left) and blast cells (right), were observed based on forwardangle (FALS) and 90° (90LS) light scatter. Solid lines indicate the presence of IL-12 during the staining procedure; dashed lines indicate theabsence of IL-12. The y-axis corresponds to relative cell number.
2 3 4 5
A.
B.
m m
mlL-12R
cyclophilin
- m
•Fig. 4. IL-12R f$1 gene transcription in murine B cells. (A) Southern blotanalysis of RT-PCR products from RNA derived from (1) unstimulatedsplenic B cells, (2) LPS-stimulated splenic B cells, (3) PeC B cells,(4) IL-12-dependent 2D6 T cells and (5) negative control. (B) RPAanalysis of (3) ~2 ng of PeC B cell RNA, (4) 10 ng of 2D6 RNA and(5) 10 ng of tRNA control.
in PeC and LPS-activated SpC preparations were found tobind IL-12, we nevertheless took the precaution of removingT and NK cells before analysis of p1 transcript expression.RNA from an IL-12-dependent T cell line, 2D6 (29), servedas a positive control for pi expression and hybridizationwith a cyclophilin probe was used as a housekeepingcontrol. It was found that RNA derived from either normalor LPS-stimulated spleen cells was positive for pi message(Fig. 4A). The PCR product could not be derived fromcontaminating DNA as the primers used span three exonsand the PCR product was the size expected for amplificationof the cDNA. PeC B cell RNA was also found to be positivefor pi expression when examined by PCR (Fig. 4A) andby the more stringent technique of RPA (Fig. 4B), althoughbecause of the small numbers of PeC B cells obtained,the amount of RNA used for RT-PCR and RPA was 1/3 and1/10 respectively of the 2D6 RNA. The presence of p1message in PeC B cells was confirmed using two non-overlapping p1 cDNA probes. The ratios of steady-state pi
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from
1960 IL- 12R on B lymphocytes
CD3+ CD19+
FreshPHA Activated
S. aureus* IL-2
FreshS. aureus* IL-2
IL-12 Fluorescence Intensity
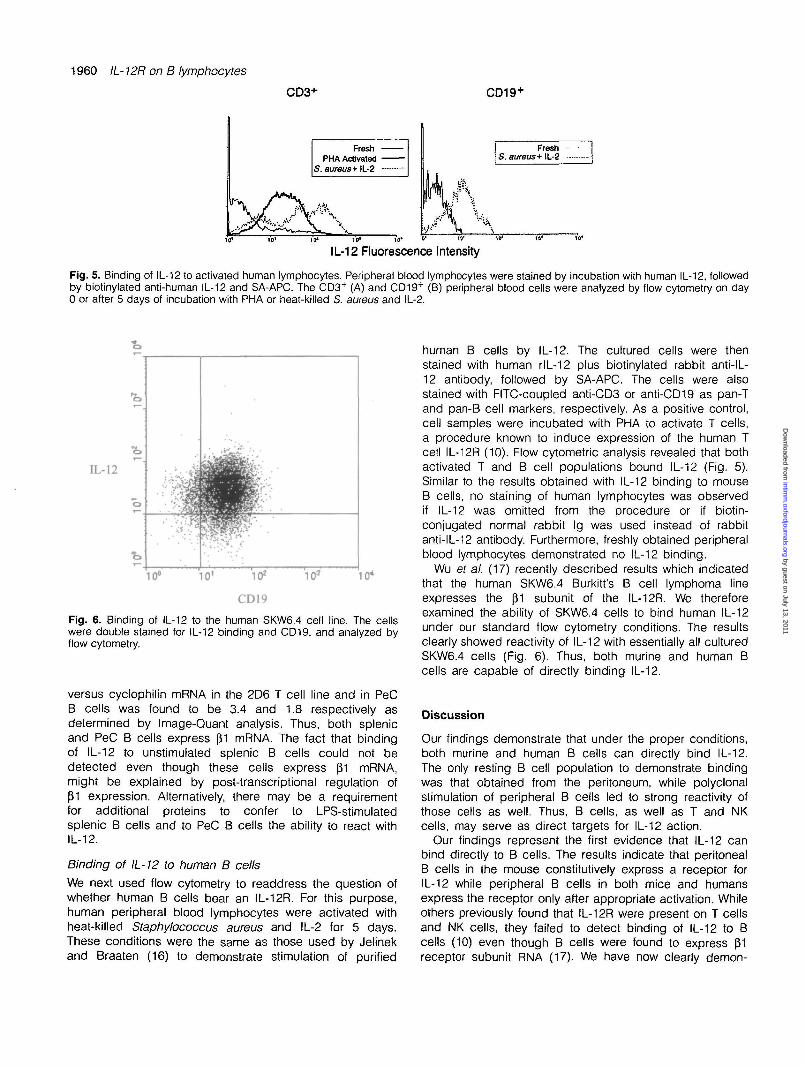
Fig. 5. Binding of IL-12 to activated human lymphocytes. Peripheral blood lymphocytes were stained by incubation with human IL-12, followedby biotinylated anti-human IL-12 and SA-APC. The CD3+ (A) and CD19+ (B) peripheral blood cells were analyzed by flow cytometry on day0 or after 5 days of incubation with PHA or heat-killed S. aureus and IL-2.
IL-12
Fig. 6. Binding of IL-12 to the human SKW6.4 cell line. The cellswere double stained for IL-12 binding and CD19, and analyzed byflow cytometry.
versus cyclophilin mRNA in the 2D6 T cell line and in PeCB cells was found to be 3.4 and 1.8 respectively asdetermined by Image-Quant analysis. Thus, both splenicand PeC B cells express (31 mRNA. The fact that bindingof IL-12 to unstimulated splenic B cells could not bedetected even though these cells express pi mRNA,might be explained by post-transcriptional regulation ofpi expression. Alternatively, there may be a requirementfor additional proteins to confer to LPS-stimulatedsplenic B cells and to PeC B cells the ability to react withIL-12.
Binding of IL-12 to human B cells
We next used flow cytometry to readdress the question ofwhether human B cells bear an IL-12R. For this purpose,human peripheral blood lymphocytes were activated withheat-killed Staphylococcus aureus and IL-2 for 5 days.These conditions were the same as those used by Jelinekand Braaten (16) to demonstrate stimulation of purified
human B cells by IL-12. The cultured cells were thenstained with human rlL-12 plus biotinylated rabbit anti-IL-12 antibody, followed by SA-APC. The cells were alsostained with FITC-coupled anti-CD3 or anti-CD19 as pan-Tand pan-B cell markers, respectively. As a positive control,cell samples were incubated with PHA to activate T cells,a procedure known to induce expression of the human Tcell IL-12R (10). Flow cytometric analysis revealed that bothactivated T and B cell populations bound IL-12 (Fig. 5).Similar to the results obtained with IL-12 binding to mouseB cells, no staining of human lymphocytes was observedif IL-12 was omitted from the procedure or if biotin-conjugated normal rabbit Ig was used instead of rabbitanti-IL-12 antibody. Furthermore, freshly obtained peripheralblood lymphocytes demonstrated no IL-12 binding.
Wu ef al. (17) recently described results which indicatedthat the human SKW6.4 Burkitt's B cell lymphoma lineexpresses the (31 subunit of the IL-12R. We thereforeexamined the ability of SKW6.4 cells to bind human IL-12under our standard flow cytometry conditions. The resultsclearly showed reactivity of IL-12 with essentially all culturedSKW6.4 cells (Fig. 6). Thus, both murine and human Bcells are capable of directly binding IL-12.
Discussion
Our findings demonstrate that under the proper conditions,both murine and human B cells can directly bind IL-12.The only resting B cell population to demonstrate bindingwas that obtained from the peritoneum, while polyclonalstimulation of peripheral B cells led to strong reactivity ofthose cells as well. Thus, B cells, as well as T and NKcells, may serve as direct targets for IL-12 action.
Our findings represent the first evidence that IL-12 canbind directly to B cells. The results indicate that peritonealB cells in the mouse constitutively express a receptor forIL-12 while peripheral B cells in both mice and humansexpress the receptor only after appropriate activation. Whileothers previously found that IL-12R were present on T cellsand NK cells, they failed to detect binding of IL-12 to Bcells (10) even though B cells were found to express pireceptor subunit RNA (17). We have now clearly demon-
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from

IL-12Ron B lymphocytes 1961
strated by flow cytometry that IL-12 can indeed bind to Bcells and B cell lines. Our results with the human SKW6.4cell line have recently been independently confirmed (J. P.Leonard, pers. commun.). We believe that our success indemonstrating IL-12 binding to B cells was due to: (i)appropriate activation of peripheral B cells to inducesignificant proliferation, (ii) examination of peritoneal B cellpopulations and (iii) use of a highly sensitive flow cytometrytechnique employing the biotin-avidin system. We havealso detected IL-12 binding to lymph node B cells fromimmunized mice (unpublished observations) and othershave now found IL-12R on mouse (M. K. Gately, pers.commun.) and human (31) B cells using radiolabeled IL-12 binding assays.
We have shown that murine B cells express mRNA forthe p1 subunit of the IL-12R. The fact that unstimulatedsplenic B cells do not bind IL-12 yet display large amountsof (31 chain transcript suggests that (31 chain post-transcriptional regulation and/or expression of other receptorcomponents are required for IL-12 binding, and it is thesechanges that are induced by LPS activation. In contrast tothe findings of Chua et al. (30) who reported that COS-7cells transfected with the murine (31 subunit could reactwith human IL-12, we found that human IL-12 was unableto bind to murine B cells. Thus, expression of additionalreceptor components may be required to confer high-affinity IL-12 binding and it likely that B cells can expressthese subunits since high-affinity IL-12R have now beenfound on human B cell lines (31) and on activated mousesplenic B cells (M. Gately, pers. commun.). It is as yetunknown whether such binding tranduces activating signalsin the target B cells.
The discovery of an IL-12R on B cells indicates thathumoral immunity may be directly influenced by IL-12 andthat the Th1/Th2 model of immunity does not explain all ofthe influences of IL-12 on B cells. We have previouslyshown that peritoneal B-1 cells are inhibited by in vivo orin vitro exposure to IL-12 (8,9), but lgG2a antibodyresponses by conventional B cells to protein and haptenantigens are stimulated (6-8). Expression of the Th2-associated isotypes, lgG1 and IgE, is temporarily sup-pressed by IL-12 but production of these antibodies isultimately enhanced by the cytokine (6,32). The reason forthe distinct effects of IL-12 on B-1 and conventional B cellsubsets is currently unknown but may reflect differences inreceptor subunit expression and/or signaling pathways. Theability of IL-12 to act directly on B cells may also accountfor the observed increase in levels of lymph node B cellsearly after treatment of mice with IL-12, an effect thatappears to be independent of IFN-y (33). Since IL-12 maybe produced by B cells (34,35) and B cells also bear anIL-12R, this cytokine may act as an autocrine/paracrinegrowth factor, analogous to the action of IL-2 on T cells.It is also possible that B cells serve to dampen the effectsof IL-12 on T cells since they are a major source of IL-10which suppresses Th1-like responses (36-38) and numbersof splenic IL-10-secreting cells are increased after IL-12administration (5). Further knowledge about the IL-12R(s) onB cells may provide important insight into the physiological
differences between B-1 and conventional B cells, and theinfluence of IL-12 on humoral immunity.
Acknowledgements
We thank Donna Williams and Bei Wang for outstanding technicalassistance.
Abbreviations
LPSPeCPHARPASA-APCSpC
lipopolysaccharideperitoneal cellsphytohemagglutininribonuclease protection assaystreptavidin-allophycocyaninspleen cells
References
1 Trinchieri, G. 1995. lnterleukin-12: a proinflammatory cytokinewith immunoregulatory functions that bridge innate resistance andantigen-specific adaptive immunity. Annu. Rev. Immunol. 13:251.
2 Buchanan, J. M., Humphrey, G. B. and Metzger, D. W. 1996.Biological activity and potential therapeutic applications ofinterleukin-12. Int. J. Pediat. Hematol. Oncol. 3:123.
3 Hendrzak, J. A. and Brunda, M. J. 1995. Interleukin-12: biologicactivity, therapeutic utility, and role in disease. Lab. Invest. 72:619.
4 McKnight, A. J., Zimmer, G. J., Fogelman, I., Wolf, S. F. andAbbas, A. K. 1994. Effects of IL-12 on helper T cell-dependentimmune responses in vivo. J. Immunol. 152:2172.
5 Morris, S. C, Madden, K. B., Adamovicz, J. J., Gause, W. C,Hubbard, B. R., Gately, M. K. and Finkelman, F. D. 1994. Effectsof IL-12 on in vivo cytokine gene expression and Ig isotypeselection. J. Immunol. 152:1047.
6 Buchanan, J. M., Vbgel, L. A., Van Cleave, V. H. and Metzger,D. W. 1995. Interleukin-12 alters the isotype-restricted antibodyresponse of mice to hen eggwhite lysozyme. Int. Immunol. 7:1519.
7 Germann, T, Bongartz, M., Dlugonska, H., Hess, H., Schmitt, E.,Kolbe, L, Kolsch, E., Podlaski, F. J., Gately, M. K. and Rude, E.1995. Interleukin-12 profoundly up-regulates the synthesis ofantigen-specific complement-fixing lgG2a, lgG2b, and lgG3antibody subclasses in vivo. Eur. J. Immunol. 25:823.
8 Metzger, D. W., Buchanan, J. M., Collins, J. L, Lester, T. L,Murray, K. S., Van Cleave, V. H., Vbgel, L. A. and Dunnick, W. A.1995. Enhancement of humoral immunity by IL-12. Ann. NYAcad.Sci., in press.
9 Vbgel, L. A., Lester, T. L, Van Cleave, V. H. and Metzger, D. W.1996. Inhibition of murine B-1 lymphocytes by interleukin-12. Eur.J. Immunol. 26:219.
10 Desai, B. B., Quinn, P. M., Wolitzky, A. G., Mongini, P. K. A.,Chizzonite, R. and Gately, M. K. 1992. IL-12R. II. Distribution andregulation of receptor expression. J. Immunol. 148:3125.
11 Finkelman, F. D., Katona, I. M., Mosmann, T. R. and Coffman,R. L. 1988. IFN-y regulates the isotypes of Ig secreted duringin vivo humoral immune responses. J. Immunol. 140:1022.
12 Snapper, C. M. and Paul, W. E. 1987. Interferon-y and B cellstimulatory factor-1 reciprocally regulate Ig isotype production.Science 236:944.
13 Snapper, C. M., Peschel, C. and Paul, W. E. 1988. IFN-y stimulateslgG2a secretion by murine B cells stimulated with bacteriallipopolysaccharide. J. Immunol. 140:2121.
14 Chace, J. H.. Abed, N. S., Adel, G. L. and Cowdery, J. S. 1993.Regulation of differentiation in CD5+ and conventional B cells.Sensitivity to LPS-induced differentiation and interferon-v-mediated inhibition of differentiation. Clin. Immunol.Immunopathol. 68:327.
15 Vezzio, N., Gately, M., Kiniwa, M., Wu, C. Y. and Delespesse, G.1993. IFN-y-independent suppression of IgE synthesis by IL-12.J. Immunol. 150:61A (Abstr.).
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from

1962 IL- 12R on B lymphocytes
16 Jelinek, D. F. and Braaten, J. K. 1995. Role of IL-12 in human Blymphocyte proliferation and differentiation. J. Immunol. 154:1606.
17 Wu, C. Y, Warrier, R. R., Carvajal, D. M., Chua, A. O., Minetti,L J., Chizzonite, R., Mongini, P. K. A., Stern, A. S., Gubler, U.,Presky, D. H. and Gately, M. K. 1996. Biological function anddistribution of human interleukin-12 receptor p chain. Eur. J.Immunol. 26:345.
18 Rao, M., Lee, W. T. and Conrad, D. H. 1987. Characterization ofa monoclonal antibody directed against the murine B lymphocytereceptor for IgE. J. Immunol. 138:1845.
19 Coffman, R. L. and Weissman, I. L. 1981. B220: a B cell-specificmember of the T200 glycoprotein family. Nature 289:681.
20 Unkeless, J. C. 1979. Characterization of a monoclonal antibodydirected against mouse macrophage and lymphocyte Fcreceptors. J. Exp. Med. 150:580.
21 Ledbetter, J. A. and Herzenberg, L. A. 1979. Xenogeneicmonoclonal antibodies to mouse lymphoid differentiation antigens.Immunol. Rev. 47:63.
22 Springer, T., Galfre, G., Secher, D. S. and Milstein, C. 1978.Monoclonal xenogeneic antibodies to murine cell surfaceantigens: identification of novel leukocyte differentiation antigens.Eur. J. Immunol. 8:539.
23 Vogel, L. A., Sercarz, E. E. and Metzger, D. W. 1995. Antibodyresponse of murine B-1 cells to hen eggwhite lysozyme. Cell.Immunol. 161:88.
24 Hunter, C. A., Subauste, C. S., Van Cleave, V. H. and Remington,J. S. 1994. Production of gamma interferon by natural killercells from Toxoplasma gondii-lniected SCID mice: regulation byinterleukin-10, interleukin-12, and tumor necrosis factor alpha.Infect. Immun. 62:2818.
25 Gately, M. K., Warrier, R. R., Honasoge, S., Carvajal, D. M.,Faherty, D. A., Connaughton, S. E., Anderson, T. D., Sarmiento,U., Hubbard, B. R. and Murphy, M. 1994. Administration ofrecombinant IL-12 to normal mice enhances cytolytic lymphocyteactivity and induces production of IFN-y in vivo. Int. Immunol.6:157.
26 Gilman, M. 1996. Ribonuclease protection assay. In Ausubel, F.M., Biert, R., Kingston, R. E., Moore, D. D., Siedman, J. G., Smith,J. A. and Struhl, K., eds, Current Protocols in Molecular Biology,John Wiley & Sons, New York, p. 4.7.1.
27 Showe, L. C, Wysocka, M., Wang, B., Lineman-Williams, D., Peritt,D., Showe, M. K. and Trinchieri, G. 1996. Structure of the mouseIL-12RP1 chain and regulation of its expression in BCG/LPStreated mice. Ann. NY Acad. Sci., in press.
28 Merberg, D. M., Wolf, S. F. and Clark, S. C. 1992. Sequencesimilarity between NKSF and the IL-6/G-CSF family. Immunol.Today 13:77.
29 Maruo, S., Toyo-Oka, K., Oh-Hora, M., Tai, X. G., Iwata, H.,Takenaka, H., Yamada, S., Ono, S., Hamaoka, T., Kobayashi, M.,Wysocka, M., Trinchieri, G. and Fujiwara, H. 1996. IL-12 producedby antigen-presenting cells induces IL-2-independent proliferationof T helper cell clones. J. Immunol. 156:1748.
30 Chua, A. O., Wilkinson, V. L. Presky, D. H. and Gubler, U.1995. Cloning and characterization of a mouse IL-12 receptor-pcomponent. J. Immunol. 155:4286.
31 Benjamin, D., Sharma, V., Kubin, M., Klein, J. L., Sartori, A.,Holliday, J. and Trinchieri, G. 1996. IL-12 expression in AIDS-related lymphoma B cell lines. J. Immunol. 156:1626.
32 Germann, T., Guckes, S., Bongartz, M., Dlugonska, H., Schmitt,E., Kolbe, L, Kolsch, E., Podlaski, F. J., Gately, M. K. andRude, E. 1995. Administration of IL-12 during ongoing immuneresponses fails to permanently suppress and can even enhancethe synthesis of antigen-specific IgE. Int. Immunol. 7:1649.
33 Chung, C. L, Schopf, L. R., Goldman, S. J., Leonard, J. P. andSypek, J. P. 1995. Interleukin-12 expands B220+ cell populationsin mice infected with Leishmania major. 9th Int. Congress Immunol.894 (Abstr.).
34 D'Andrea, A., Rengaraju, M., Valiante, N. M., Chehimi, J., Kubin,M., Aste, M., Chan, S. H., Kobayashi, M., Young, D., Nickbarg,E. B., Chizzonite, R., Wolf, S. F. and Trinchieri, G. 1992. Productionof natural killer cell stimulatory factor (interleukin 12) by peripheralblood mononuclear cells. J. Exp. Med. 176:1387.
35 Mengel, J., Dare, L., Dare, G. M., Delgado, M., Nomizo, A., Silva,J. S. and Campos-Neto, A. 1992. An activated murine B celllymphoma line (A-20) produces a factor-like activity which isfunctionally related to human natural killer cell stimulatory factor.Eur. J. Immunol. 22:3173.
36 Mosmann, T. R. and Coffman, R. L. 1989. Heterogeneity ofcytokine secretion patterns and functions of helper T cells. Adv.Immunol. 46:111.
37 O'Garra, A., Chang, R., Go, N., Hastings, R., Haughton, G. andHoward, M. 1992. Ly-1 B (B-1) cells are the main source of Bcell-derived interleukin 10. Eur. J. Immunol. 22:711.
38 D'Andrea, A., Aste-Amezaga, M., Valiante, N. M., Ma, X., Kubin,M. and Trinchieri, G. 1993. Interleukin-10 inhibits humanlymphocyte IFN-y production by suppressing natural killer cellstimulatory factor/interleukin-12 synthesis in accessory cells. J.Exp. Med. 178:1041.
by guest on July 13, 2011intim
m.oxfordjournals.org
Dow
nloaded from




















