
Pulsed-field gel electrophoresis, plasmid profiles and phage types for the human isolates of...
12
Pulsed-field gel electrophoresis, plasmid profiles and phage types for the human isolates of Salmonella enterica serovar Enteritidis obtained over 13 years in Taiwan J.-C. Pang 1 , T.-H. Chiu 1 , C.-S. Chiou 2 , A. Schroeter 3 , B. Guerra 3 , R. Helmuth 3 and H.-Y. Tsen 4 1 Department of Food Science, National Chung-Hsing University, Taichung City, 2 The Central Branch Office, Center for Disease Control, Taichung City, Taiwan, 3 Federal Institute of Risk Assessment, National Veterinary Reference Laboratory for Salmonella, Berlin, Germany, and 4 Department of Food Science and Nutrition, Hung-Kuang University, Shalu, Taichung County, Taiwan 2005/0072: received 24 January 2005, revised 20 March 2005 and accepted 1 April 2005 ABSTRACT J.-C. PANG, T.-H. CHIU, C.-S. CHIOU, A. SCHROETER, B. GUERRA, R. HELMUTH AND H.-Y. TSEN. 2005. Aims: Plasmid profile, phage typing, and pulsed-field gel electrophoresis (PFGE) patterns of 124 Salmonella Enteritidis strains isolated in 1998–2002 in Taiwan were analysed and the results were compared with those of the 63 strains obtained in 1991–1997, so that molecular subtypes and epidemic strains for Salmonella Enteritidis over a 13-year period (1991–2002) could be elucidated. Methods and Results: A total of 124 strains of Salmonella Enteritidis isolated from human in Taiwan between 1998 and 2002 were analysed by PFGE, plasmid analysis and phage typing. The results obtained were compared with those of the 63 strains obtained in 1991–1997, so that the clonal relationships for a total of 187 strains obtained over 13 years could be elucidated. For PFGE, restriction enzymes XbaI, SpeI and NotI were used for chromosomal DNA digestion. Results showed 28 PFGE pattern combinations for the 187 Salmonella strains. Of them, pattern X3S3N3 was the major subtype as 130 strains isolated from different locations during 1991–2002 showed this PFGE pattern. For all these 187 strains, the genetic similarity was higher than 80%. Plasmid analysis showed 17 distinct types, which consist of one to four plasmids and the predominant phage type of those strains was PT4 (71Æ6%) and PT6a (13Æ4%). The three methods identified different degrees of polymorphism in the following order: plasmid profile (18 types, D ¼ 0Æ659) > PFGE (28 types, D ¼ 0Æ512) > phage typing (13 types, D ¼ 0Æ438). As PFGE patterns, phage type and plasmid profile were combined for subtyping, the 187 strains could be grouped into 46 subtypes and the discriminatory index was raised to 0Æ795. For these 46 subtypes, the predominant one was X3S3N3/P1/PT4, which contained 77 (41%) isolates. Conclusions: Most of the Salmonella Enteritidis strains from sporadic cases were with pattern X3S3N3. They were the prevalent and may be the epidemic strains found in Taiwan during 1991–2002. The present study suggested that the several variants were derived from a single clonal line and the genome for strains of Salmonella Enteritidis are highly conserved over a 13-year period (1991–2002). Significance and Impact of the Study: The results obtained here are useful for epidemiolgical study of salmonellosis caused by Salmonella Enteritidis in Taiwan. Comparing the data of the present study with those obtained for strains from other countries, the major subtypes for Salmonella Enteritidis infection in the world can be elucidated. Keywords: PFGE, plasmid phage typing, Salmonella Enteritidis, epidemic, subtypes. Correspondence to: Hau-Yang Tsen, Department of Food Science and Nutrition, Hung-Kuang University, No. 34, Chung-Chi Rd, Taichung County 433, Taiwan (e-mail: [email protected]). ª 2005 The Society for Applied Microbiology Journal of Applied Microbiology 2005, 99, 1472–1483 doi:10.1111/j.1365-2672.2005.02749.x
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Pulsed-field gel electrophoresis, plasmid profiles and phage types for the human isolates of...

Pulsed-field gel electrophoresis, plasmid profiles and phagetypes for the human isolates of Salmonella enterica serovarEnteritidis obtained over 13 years in Taiwan
J.-C. Pang1, T.-H. Chiu1, C.-S. Chiou2, A. Schroeter3, B. Guerra3, R. Helmuth3 and H.-Y. Tsen4
1Department of Food Science, National Chung-Hsing University, Taichung City, 2The Central Branch Office, Center for Disease Control,
Taichung City, Taiwan, 3Federal Institute of Risk Assessment, National Veterinary Reference Laboratory for Salmonella, Berlin,
Germany, and 4Department of Food Science and Nutrition, Hung-Kuang University, Shalu, Taichung County, Taiwan
2005/0072: received 24 January 2005, revised 20 March 2005 and accepted 1 April 2005
ABSTRACT
J . -C . PANG, T . -H . CHIU, C. -S . CHIOU, A . SCHROETER, B. GUERRA, R . HELMUTH AND H. -Y . TSEN. 2005.
Aims: Plasmid profile, phage typing, and pulsed-field gel electrophoresis (PFGE) patterns of 124 SalmonellaEnteritidis strains isolated in 1998–2002 in Taiwan were analysed and the results were compared with those of the
63 strains obtained in 1991–1997, so that molecular subtypes and epidemic strains for Salmonella Enteritidis over a
13-year period (1991–2002) could be elucidated.
Methods and Results: A total of 124 strains of Salmonella Enteritidis isolated from human in Taiwan between
1998 and 2002 were analysed by PFGE, plasmid analysis and phage typing. The results obtained were
compared with those of the 63 strains obtained in 1991–1997, so that the clonal relationships for a total of 187 strains
obtained over 13 years could be elucidated. For PFGE, restriction enzymes XbaI, SpeI and NotI were used for
chromosomal DNA digestion. Results showed 28 PFGE pattern combinations for the 187 Salmonella strains. Of
them, pattern X3S3N3 was the major subtype as 130 strains isolated from different locations during 1991–2002
showed this PFGE pattern. For all these 187 strains, the genetic similarity was higher than 80%. Plasmid analysis
showed 17 distinct types, which consist of one to four plasmids and the predominant phage type of those strains
was PT4 (71Æ6%) and PT6a (13Æ4%). The three methods identified different degrees of polymorphism in the
following order: plasmid profile (18 types, D ¼ 0Æ659) > PFGE (28 types, D ¼ 0Æ512) > phage typing (13 types,
D ¼ 0Æ438). As PFGE patterns, phage type and plasmid profile were combined for subtyping, the 187 strains could
be grouped into 46 subtypes and the discriminatory index was raised to 0Æ795. For these 46 subtypes, the
predominant one was X3S3N3/P1/PT4, which contained 77 (41%) isolates.
Conclusions: Most of the Salmonella Enteritidis strains from sporadic cases were with pattern X3S3N3. They
were the prevalent and may be the epidemic strains found in Taiwan during 1991–2002. The present study
suggested that the several variants were derived from a single clonal line and the genome for strains of SalmonellaEnteritidis are highly conserved over a 13-year period (1991–2002).
Significance and Impact of the Study: The results obtained here are useful for epidemiolgical study of
salmonellosis caused by Salmonella Enteritidis in Taiwan. Comparing the data of the present study with those
obtained for strains from other countries, the major subtypes for Salmonella Enteritidis infection in the world can be
elucidated.
Keywords: PFGE, plasmid phage typing, Salmonella Enteritidis, epidemic, subtypes.
Correspondence to: Hau-Yang Tsen, Department of Food Science and Nutrition, Hung-Kuang University, No. 34, Chung-Chi Rd, Taichung County 433, Taiwan
(e-mail: [email protected]).
ª 2005 The Society for Applied Microbiology
Journal of Applied Microbiology 2005, 99, 1472–1483 doi:10.1111/j.1365-2672.2005.02749.x

INTRODUCTION
Salmonella enterica serovar Enteritidis is one of the major
food pathogens. It may cause salmonellosis and gastroen-
teritis in humans and animals. Animals and their products,
especially meat and eggs from chickens, are considered as a
major source of infection with this pathogen for humans
(Rodrigue et al. 1990; Boonmar et al. 1998). In the past
10 years, Salm. Enteritidis is the most common Salmonellaserovars isolated from human, animal, food and feed in
many countries (http://www.who.int/salmsurv). In Tai-
wan, there has been a gradual significant increase in group D
Salmonella infection and the incidence of food-borne cases
caused by Salm. Enteritidis has increased dramatically after
1996 (Su et al. 2001). Between 1993 and 1997, Salm.Enteritidis was the major cause for food-borne gastroenter-
itis outbreaks (Wang et al. 1999). Because of the major role
of Salm. Enteritidis in Salmonella infection in Taiwan,
establishment of the epidemiological data and study of the
clonal relationship for Salm. Enteritidis strains is important.
Such data may allow us to elucidate the major epidemic
subtypes and to trace the contamination source during
outbreaks.
Different phenotypic and genotypic methods have been
used for the subtyping of Salm. Enteritidis. These methods
include phage typing, serotyping, antibiogram, pulsed-field
gel electrophoresis (PFGE), plasmid profile, IS 200 typing,
ribotyping, random amplified polymorphic DNA, amplified
fragment length polymorphism and multisequence locus
typing, etc. (Helmuth and Schroeter 1994; Olsen et al.1994; Laconcha et al. 1998; Kotetishvili et al. 2002; Yan
et al. 2004). Because of the high genetic homogeneity
among the Salm. Enteritidis strains (Thong et al. 1995;
Bakeri et al. 2003), a single method cannot be relied on for
discriminating between strains. Combination of several
typing methods may provide a powerful discriminatory tool
for the epidemiological analysis of unrelated and related
strains of Salm. Enteritidis (Laconcha et al. 1998, 2000;
Liebana et al. 2001). However, phage typing has been a
common method for typing Salmonella strains since 1950.
The phage typing data, which suggest a clonal diffusion of
a limited number of strains in Europe (PT4) and USA
(PT8) (Laconcha et al. 2000; Patrick et al. 2004) was also
included in this study.
Previously, we used PFGE, phage typing and plasmid
profiling to analyse the 63 Salm. Enteritidis strains
isolated from patients suffering from food-borne poisoning
cases in Taiwan during 1991–1997. For PFGE analysis of
these 63 Taiwan isolates, XbaI, SpeI and NotI were used
for DNA digestion and the results showed that 10 PFGE
pattern combinations were found. Of them, pattern
X3S3N3 was the major subtype as 46 strains belong to
this PFGE type (Tsen and Lin 2001). There was a close
genetic similarity between epidemiologically unrelated and
outbreak strains of Salm. Enteritidis. This study was
carried out with an intention to extend the monitoring
work of genetic diversity to more recently collected
Taiwan strains of Salm. Enteritidis. The methods used
were the PFGE, phage typing and plasmid analysis. The
results obtained were then compared with those reported
previously (Tsen and Lin 2001) in order to establish the
subtyping data and to elucidate the epidemic strains of
Salm. Enteritidis over a 13-year period (1991–2002) in
Taiwan.
MATERIALS AND METHODS
Bacterial strains
A total of 100 clinical isolates of serogroup D1 Salmonellafrom stool samples of unrelated patients during 1998–2002
were used in this study. They were obtained from the
National Center for Disease Control (Taipei, Taiwan). Of
them, 84 clinical isolates were identified as Salm. Enteritidisby serotyping. Another 40 strains of Salm. Enteritidis wereisolated from unrelated patients of National Taiwan Uni-
versity Hospital, Taipei, Taiwan. For cultivation of the
bacteria, one loopful of the Salmonella strain was inoculated
into 3 ml Luria broth (LB) or tryptic soy broth (TSB) and
incubated at 37�C for 12–18 h. Stock cultures were kept at
)70�C with 25% glycerol.
Phage typing
Phage types of Salm. Enteritidis strains were determined by
the Federal Institute for Risk Assessment (BfR), Berlin-
Marienfelde, Germany, through the generous assistance
from Dr A. Schroeter and Dr R. Helmuth.
Polymerase chain reaction
Polymerase chain reaction (PCR) detection of the spv gene ofSalm. Enteritidis was performed as described by Mahon and
Lax (1993). The primers used were SPVR1 and SPVR2.
Their sequences are as follows: SPVR1: acaggttccttcagtatcgc
and SPVR2: ctgttgatatcaggttt. Molecular weight of the PCR
product amplified from these PCR primers is expected to be
491 bp. A thermal cycler (Perkin-Elmer 9600, Norwalk, CT,
USA) was used for automated temperature control of the
PCR. DNA was amplified through 35 cycles of denaturation
(94�C, 30 s), annealing (50�C, 30 s) and polymerization
(72�C, 30 s). A 5-ll aliquot of amplified DNA was examined
by electrophoresis through a 2% ultra-pure agarose gel in
0Æ5x TAE buffer (50x TAE: 242 g Tris base; 37Æ2 g, 100 ml
0.5 mol l)1 EDTA, pH 8Æ0; 57Æ1 glacial and adjusted to
1000 ml with H2O).
PFGE, PLASMID PROFILE, AND PHAGE TYPING OF SALM. ENTERITIDIS 1473
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x

Pulsed-field gel electrophoresis
Themethods described byTsen andLin (2001)weremodified
for use. Salmonella. Enteritidis strains were grown in 3 ml
TSB broth at 37�C for 3 h and the cells were collected by
centrifugation (6000 g, 10 min), washed and resuspended in
300 ll suspension solution (10 mmol l)1 Tris-HCl, 20 mmol
l)1NaCl, 100 mmol l)1 EDTA).Theywere heated at 55�C for
3 min and mixed with the same volume of 2% agarose gel
(pulsed-field grade; Bio-Rad, Hercules, CA, USA) prepared
with suspension solution. The mixture was poured into the
slots of a plastic mould (CHEF DRII; Bio-Rad). The agarose
plugs obtained were transferred into lysozyme solution
(2Æ5 mg ml)1 lysozyme, 10 mmol l)1 Tris-HCl, pH 7Æ2;50 mmol l)1 NaCl, 100 mmol l)1 EDTA; 0Æ2% sodium
deoxycholate and 0Æ5% N-laurylsarcosine) and cells were
lysed for 24 h at 37�C. The lysis buffer was then replaced by
proteolysis solution (0Æ1 mol l)1 EDTA, 1% N-laurylsarco-
sine, 0Æ2% sodium deoxycholate, 0Æ1 mg ml)1 proteinase K)
and proteolysis was carried out at 55�C for 24–48 h. Finally,
the agarose plugs were washed five times for 30 min with
washing buffer (20 mmol l)1 Tris-HCl; 50 mmol l)1 EDTA)
with gently shaking and stocked in storage solution (2 mmol
l)1 Tris-HCl; 5 mmol l)1 EDTA) at 4�C.The 1-mm slice of each agarose plug was equilibrated in
100 ll restriction buffer 3 or 2 (New England Biolabs,
Beverly, MA, USA) for 15 min and digested at 37�C for 3 h
with 20 unitsXbaI (New England Biolabs), 8 units SpeI (New
England Biolabs) or 10 units NotI (New England Biolabs) in
100 ll fresh buffer 3 or 2 respectively. After digestion, the
plugs were placed into the slots of a 1Æ2% agarose gel (pulsed-
field grade; Bio-Rad) in 0Æ5x TBE buffer and electrophoresis
was carried out by use of the CHEF DRII PFGE system
(Bio-Rad). The conditions used were 200 V for 24 h at 12�Cwith pulsed times ranging from 3 to 63 s for XbaI-digestedDNA, 200 V for 26 h at 12�Cwith pulsed times ranging from
2 to 45 s for SpeI-digested DNA and 200 V for 25 h at 12�Cwith pulsed times ranging from 2 to 10 s for NotI-digestedDNA. Lambda Ladder PFGMarker (New England Biolabs)
was used as molecular weight markers. The gels were stained
and photographed. PFGE patterns obtained were analysed by
the NTSYSpc software (Numerical taxonomy and multiva-
riate analysis system, version 2Æ01b, State University of New
York, Stony Brook, NY, USA). The relationship between
two given strains was scored by Dice coefficient of similarity,
and strains were clustered by the hierarchical clustering inter-
strain similarities based on the unweighted pair group
method with arithmetic averages (UPGMA)(UPGMA).
Plasmid DNA analysis
Plasmid DNA was extracted and analysed by a procedure
modified from the method of Kado and Liu (1981).
Generally, 50 ll of the stock culture (in 25% glycerol, at
)70�C) for each Salmonella strain were mixed with 3 ml of
LB broth and incubated at 37�C overnight. Cells in 1 ml of
overnight culture was harvested by centrifugation (10 000 g,5 min) at room temperature. The cell pellet was suspended
in 20 ll TE buffer (50 mmol l)1 Tris, 1 mmol l)1 EDTA,
pH 8Æ0), mixed gently with 100 ll of lysis solution
(50 mmol l)1 Tris, 3% SDS, pH 12Æ6) and incubated at
58�C for 30 min. Afterwards, 100 ll of phenol/chloroform(1 : 1) solution was mixed gently and the aqueous phase was
subjected to phenol/chloroform extraction followed by
centrifugation at 10 000 g for 30 min. The upper aqueous
phase was analysed by 0Æ8% agarose gel horizontal electro-
phoresis in 1x TBE buffer (10x buffer, 108 g Tris, 55 g
Boric acid, 9.3 g EDTA, pH 8Æ0) at 100 V for 3 h. Plasmid
sizes were determined by comparison of the DNA bands
with reference plasmid from strains of Escherichia coliR27, R1, RP4 and V517 kindly provided by Dr B. Guerra
of BfR.
Discriminatory power of the methods
The index of discrimination (D) for each typing method was
calculated according to Simpson’s index of diversity as
described by Liebana et al. (2001). It is based on the
probability that two unrelated strains randomly sampled
from the test population will be assigned to different typing
groups.
RESULTS
Pulsed-field gel electrophoresis
In order to obtain the genetic clonality and subtypes of the
Salm. Enteritidis isolates, chromosomal DNAs from 124
Salm. Enteritidis strains isolated between 1998 and 2002 in
Taiwan were digested with XbaI, SpeI and NotI, respect-ively, and analysed by PFGE. The data obtained were then
combined with those from 63 strains collected in 1991–1997.
With XbaI for digestion, for those 187 Salm. Enteritidisstrains, a total of 15 PFGE patterns (X1–X15) were
identified (Fig. 1). Each isolate generated 12 to 15 restriction
fragments with molecular sizes ranging from approx. 40 to
700 kb. Most of these strains, i.e. 140 (74Æ9%) of the total
187 strains analysed, belonged to a pattern termed as X3,
which is the major pattern. The next predominant pattern is
pattern X1, which was found for 19 of 187 isolates analysed.
Strains belonged to any of the other patterns counted <5%.
The discriminatory index (D) of XbaI was 0Æ428. The
patterns obtained with SpeI were characterized by 21–25
bands of restriction digested fragments with molecular
weights ranging from 40 to 700 kb. A total of 13 PFGE
patterns (S1–S13) were identified (Fig. 2). The predomin-
1474 J . -C .PANG ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x

ant type S3 was found for 145 (77Æ5%) of the total 187
isolates followed by pattern S2, which was found for 22
(11Æ8%) of the total strains. The discriminatory index (D) ofSpeI was 0Æ386. Finally, digestion with restriction enzyme
NotI generated 13 PFGE patterns (N1–N8, N10–N14).
Molecular weights of those digestion fragments were
between 10 and 300 kb (Fig. 3). Of the total 187 strains,
147 (78Æ6%) belong to a pattern N3, which is the major
pattern. The discriminatory index (D) of NotI was 0Æ377.For the 187 strains, XbaI, SpeI and NotI generated a total of
15, 13 and 13 PFGE patterns respectively. When those three
Pattern: X1 X2 X11 X3 X5 X12 X8 X7 X13 X14 X15 X9 X10 X1 X1 X4 X4 X6 X11 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
48·5
97·0
145·5
194·0
291·0
388·0
485·0
630·5
Size (kb)
Lane:
Fig. 1 Pulsed-field gel electrophoresis (PFGE) types for XbaI-digested genomic DNA of human isolates of Salmonella Enteritidis obtained in
Taiwan during 1991–2002. Lanes 1, 13, 14, and 23: Lambda ladder marker for PFGE; lane 2–12 and 15–22: the 15 XbaI-digested PFGE patterns for
the Salm. Enteritidis strains
Pattern:
48·5
97·0
145·5
194·0291·0
388·0
485·0
630·5
Size (kb)
Lane: 1 2S3 S2 S8 S4 S9 S10 S11 S12 S6 S7 S5 S1S13
3 4 5 6 7 8 9 10 11 12 13 14 15
Fig. 2 Pulsed-field gel electrophoresis (PFGE) types for SpeI-diges-
ted genomic DNA of human isolates of Salmonella Enteritidis obtained
in Taiwan during 1991–2002. Lanes 1 and 15: Lambda ladder marker
for PFGE; lane 2–14: the 13 SpeI-digested PFGE patterns for the
Salm. Enteritidis strains
Pattern: N3 N8 N2 N10 N11 N12 N13 N14 N1 N2 N4 N5 N6 N7
48·5
97·0
145·5194·0
Size (kb)
Lane: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Fig. 3 Pulsed-field gel electrophoresis (PFGE) types for NotI-diges-
ted genomic DNA of human isolates of Salmonella Enteritidis obtained
in Taiwan during 1991–2002. Lane 9: Lambda ladder marker for
PFGE; lane 2–8 and 10–15: the 13 SpeI-digested PFGE patterns for
the Salm. Enteritidis strains
PFGE, PLASMID PROFILE, AND PHAGE TYPING OF SALM. ENTERITIDIS 1475
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x

restriction enzyme digested patterns were combined, a total
of 28 PFGE patterns combinations were obtained. A
summary of these 28 PFGE pattern combinations is shown
in Table 1. It was found that 130 of 140 pattern X3 strains
belonged to a pattern combination of X3S3N3. For the rest
10 pattern X3 isolates, five additional patterns combinations,
i.e. X3S7N3, X3S3N8, X3S13N13, X3S7N14 and X3S5N3,
were obtained. When three enzyme patterns were combined
for subtyping, the discriminatory index (D) increased to
0Æ512. In addition to the above three restriction enzymes, we
also used restriction enzyme AvrII (BlnI) for the digestion of
43 randomly selected strains of subtype X3S3N3 and found
that only three AvrII patterns were obtained for these 43
strains. Forty-one of the 43 strains belonged to a pattern and
the rest two strains belonged to two separate patterns (data
not shown). Moreover, only one band shift was found
among those three AvrII digestion patterns.
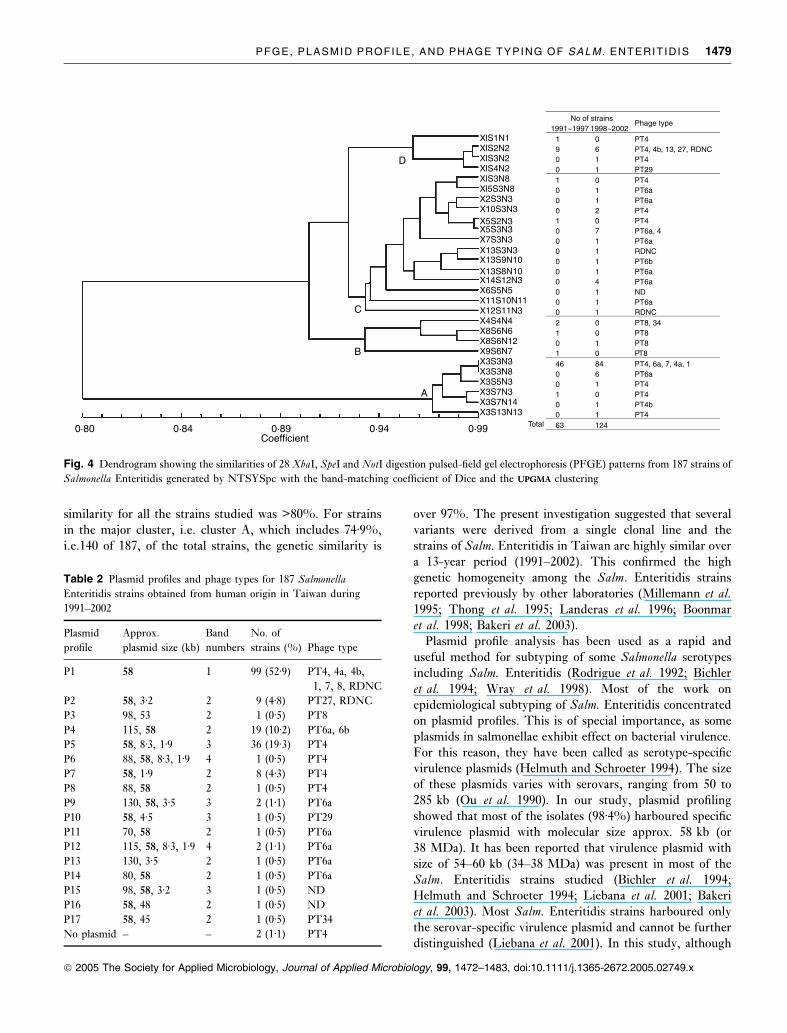
The PFGE patterns, obtained from XbaI, SpeI and NotIdigestion, were grouped into four main clusters by the
NTSYSpc, designated A to D. The similarity between
the different patterns was shown in Fig. 5. It was found that
the genetic similarity between clusters A and B, C, D is
80%. Cluster A was the major one, which encompassed 140
(74Æ9%) of the total 187 strains. All the strains with X3
subtype belonged to this cluster. Although combination of
the PFGE patterns obtained from XbaI, SpeI and NotI,respectively, could further discriminate the strains in pattern
X3 to six subtypes, the genetic similarity among these is
over 97%. For all these 187 strains, genetic similarity was
higher than 80%. Furthermore, strains isolated in 1991–
1997 and in 1998–2002 show a common major pattern, i.e.
subtype X3S3N3 (Table 1 and Fig. 4).
Plasmid profile
Table 2 shows the distribution of plasmid types for the 187
isolates obtained in 1991–2002. Plasmid profiling of the 187
Salm. Enteritidis isolates produced 17 distinct types with
one to four plasmids for each type. A total of 15 different
plasmid sizes were found in those profiles. Pattern P1,
harboured a single plasmid with size approx. 58 kb, was
the major plasmid type which include 99 (52Æ9%) of 187
strains. Of these 187 strains, 36 (19Æ3%) and 19 (10Æ2%)
strains belonged to pattern P5 (plasmid sizes 58, 8Æ3,1Æ9 kb) and P4 (plasmid sizes 115, 58 kb) respectively. Two
isolates harbour no plasmid and 183 (98%) strains
harboured a 58-kb virulence plasmid. PCR amplification
of spv was performed on all isolates with bacterial lysate as
the template. All these 183 isolates, which harboured the
58-kb plasmid, possessed spv gene. No PCR product was
found for the rest of four Salm. Enteritidis strains, which
did not harbour the 58-kb plasmid (data not shown). The
D value for plasmid profiling was 0Æ659. For plasmid
profiling, it should be mentioned that for each strain, at
least duplicate studies were performed and the results
obtained were the same. In addition, to reduce the
possibility of picking a colony where plasmid loss has
occurred, we also picked 2–3 colonies for same strains or
cells after subculture and analysed their plasmid profiles,
the results were also the same.
Phage typing
Phage typing analysis of the total 187 Salm. Enteritidis
strains collected in 1991–2002 shows 12 phage types and one
RDNC type (Table 3). Of the total strains, 134 (71Æ6%)
belonged to phage type PT4, 25 (13Æ4%) belonged to type
PT6a followed by five (2Æ7%) to PT4b, four (2Æ1%) to PT8
respectively. Nine (4Æ8%) isolates were RDNC (for reacts
but does not confirm) and for the remaining phage types, i.e.
PT1, PT4a, PT6b, PT7, PT13, PT27, PT29, PT34, only a
single strain for each type was found. PT4 was the
predominant phage type for our strains, followed by PT6a,
PT4b and PT8. PT4 has been the major PT for strains
collected every year. The D value of phage typing was 0Æ438.
Combination of PFGE, plasmid profile and phagetypes
The results from combination of the PFGE patterns and
phage types were shown in Table 1 and Fig. 4. It was found
that the major PFGE patterns in cluster A were generated
by Salm. Enteritidis PT4, 4a, 4b, 6a, 7, 1 and PFGE
patterns in cluster B were generated by PT8, PT34 strains.
The PFGE patterns in cluster C were generated by strains
of PT6a, 4, 6b and RDNC. Phage type 4 and 6a were the
predominant PT for all the strains assayed. A total of 116
(86Æ6%) PT4 strains belong to cluster A and there were 15
(60%) PT6a isolates in the cluster C. All the four PT8
strains were found in cluster B.
When phage type and plasmid profiles were combined for
subtyping, it was found that except of one strain, all the
PT6a isolates harboured plasmid with molecular weight
>58 kb (Table 2). About 80% (20/25) of the PT6a isolates
harboured 115 kb plasmid (profile P4 and P12). Three PT6a
isolates harboured 130 kb plasmid (profile P13) and one
isolate harboured 70 kb or 80 kb plasmid (profile P11 or
P14). For PT4 strains, 62Æ7% (84/134) were with plasmid
profile P1 (58 kb), 26Æ9% (36/134) were with profile P5 (58,
8Æ3, 1Æ9 kb) and 6% (8/134) were with profile P7 (58,
1Æ9 kb). Only two PT4 isolates did not harbour any plasmid.
Finally, combination of the PFGE patterns, phage types
and plasmid profiles grouped the total 187 isolates into 46
subtypes (Table 1) and the discriminatory index was raised
to 0Æ795. The predominant subtype was X3S3N3/P1/PT4,
which encompassed 77 (41%) isolates.
1476 J . -C .PANG ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x

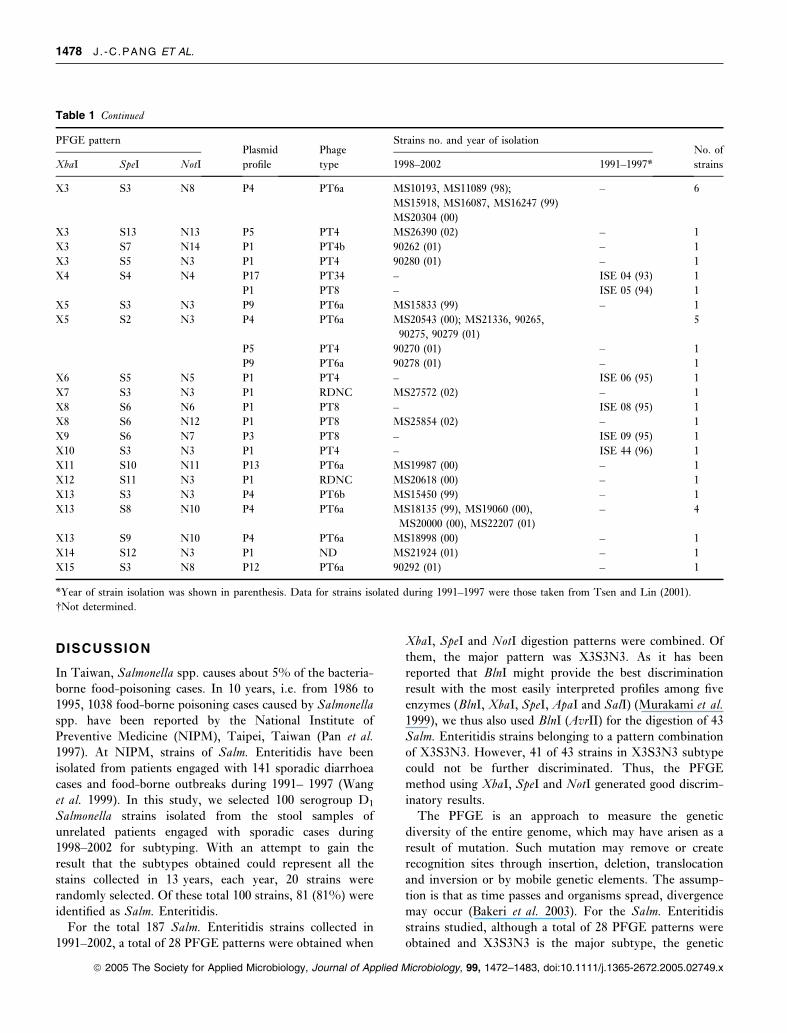
Table 1 Plasmid profiles, PFGE patterns and phage types for human isolates of Salmonella Enteritidis obtained from clinical samples in Taiwan
during 1991–2002
PFGE patternPlasmid
profile
Phage
type
Strains no. and year of isolationNo. of
strainsXbaI SpeI NotI 1998–2002 1991–1997*
X1 S1 N1 P1 PT4 – ISE01 (91) 1
X1 S2 N2 P1 PT4 – ISE02 (92) 1
P1 PT4b MS11088, MS11128, MS11213 (98);
MS20026 (00)
4
P15 PT13 – ISE07 (95) 1
P2 ND – ISE12 (96) 1
P2 PT27 – ISE29 (97) 1
P2 RDNC MS18654 (00); MS21871 (01) ISE 53–57 (97) 7
X1 S3 N8 P4 PT6a MS18529 (00) – 1
X1 S3 N9 P8 PT4 MS10856 (98) – 1
X1 S4 N9 P10 PT29 MS18218 (99) – 1
X2 S3 N3 – PT4 MS11104 (98); MS20243 (00) – 2
X3 S3 N3 P1 PT4 MS10198, MS10828 (98) ISE03 (92) 77
MS15425, MS15441, MS15470,
MS16464, MS17931, MS18174,
MS18246, MS18344 (99)
ISE34–37 (92)
MS18655, MS18678, MS18712, MS19094,
MS20232 (00)
ISE10 (95), ISE38–42 (95)
MS21465, MS21466, MS21515,
MS21608, MS21612, MS22100,
MS22379, MS22397, MS22404, MS22407,
90255, 90258, 90260, 90261, 90263, 90274,
90286, 90290 (01)
ISE14–19, ISE43,
45, 46, 47 (96)
MS25605, MS25709, MS26381 (02) ISE20–23, ISE25, 27, 28,
ISE 31–33, ISE48–52,
ISE58–62 (97)
P5 PT4 MS15380 (99); MS19409 (00) – 34
MS21132, MS22485, MS22569, 90251,
90252, 90253, 90256, 90257, 90259,
90264, 90267, 90268, 90269, 90272,
90273, 90277, 90283, 90288,
90293, 90294 (01)
MS25601, MS26322, MS26391, MS27653,
MS27669, MS27790,
MS27788, MS27791, MS27874, MS27875,
MS27912, MS27988 (02)
P7 PT4 MS20614 (00);
MS21513, MS21549, MS21997, 90266,
90276, 90284, 90285 (01)
– 8
P6 PT4 MS27891 (02) – 1
P4 PT6a 90271 (01) – 1
P1 PT7 – ISE13 (96) 1
P1 PT4a – ISE24 (97) 1
P1 ND� – ISE11 (96); ISE26 (97) 2
P1 PT1 MS20080 (00) – 1
P14 PT6a MS20093 (00) – 1
P11 PT6a 90281 (01) – 1
P12 PT6a 90287 (01) – 1
P16 ND – ISE 30 (97) 1
X3 S7 N3 P1 PT4 – ISE 63 (97) 1
PFGE, PLASMID PROFILE, AND PHAGE TYPING OF SALM. ENTERITIDIS 1477
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x
DISCUSSION
In Taiwan, Salmonella spp. causes about 5% of the bacteria-
borne food-poisoning cases. In 10 years, i.e. from 1986 to
1995, 1038 food-borne poisoning cases caused by Salmonellaspp. have been reported by the National Institute of
Preventive Medicine (NIPM), Taipei, Taiwan (Pan et al.1997). At NIPM, strains of Salm. Enteritidis have been
isolated from patients engaged with 141 sporadic diarrhoea
cases and food-borne outbreaks during 1991– 1997 (Wang
et al. 1999). In this study, we selected 100 serogroup D1
Salmonella strains isolated from the stool samples of
unrelated patients engaged with sporadic cases during
1998–2002 for subtyping. With an attempt to gain the
result that the subtypes obtained could represent all the
stains collected in 13 years, each year, 20 strains were
randomly selected. Of these total 100 strains, 81 (81%) were
identified as Salm. Enteritidis.For the total 187 Salm. Enteritidis strains collected in
1991–2002, a total of 28 PFGE patterns were obtained when
XbaI, SpeI and NotI digestion patterns were combined. Of
them, the major pattern was X3S3N3. As it has been
reported that BlnI might provide the best discrimination
result with the most easily interpreted profiles among five
enzymes (BlnI, XbaI, SpeI, ApaI and SalI) (Murakami et al.1999), we thus also used BlnI (AvrII) for the digestion of 43
Salm. Enteritidis strains belonging to a pattern combination
of X3S3N3. However, 41 of 43 strains in X3S3N3 subtype
could not be further discriminated. Thus, the PFGE
method using XbaI, SpeI and NotI generated good discrim-
inatory results.
The PFGE is an approach to measure the genetic
diversity of the entire genome, which may have arisen as a
result of mutation. Such mutation may remove or create
recognition sites through insertion, deletion, translocation
and inversion or by mobile genetic elements. The assump-
tion is that as time passes and organisms spread, divergence
may occur (Bakeri et al. 2003). For the Salm. Enteritidisstrains studied, although a total of 28 PFGE patterns were
obtained and X3S3N3 is the major subtype, the genetic
Table 1 Continued
PFGE patternPlasmid
profile
Phage
type
Strains no. and year of isolationNo. of
strainsXbaI SpeI NotI 1998–2002 1991–1997*
X3 S3 N8 P4 PT6a MS10193, MS11089 (98);
MS15918, MS16087, MS16247 (99)
MS20304 (00)
– 6
X3 S13 N13 P5 PT4 MS26390 (02) – 1
X3 S7 N14 P1 PT4b 90262 (01) – 1
X3 S5 N3 P1 PT4 90280 (01) – 1
X4 S4 N4 P17 PT34 – ISE 04 (93) 1
P1 PT8 – ISE 05 (94) 1
X5 S3 N3 P9 PT6a MS15833 (99) – 1
X5 S2 N3 P4 PT6a MS20543 (00); MS21336, 90265,
90275, 90279 (01)
5
P5 PT4 90270 (01) – 1
P9 PT6a 90278 (01) – 1
X6 S5 N5 P1 PT4 – ISE 06 (95) 1
X7 S3 N3 P1 RDNC MS27572 (02) – 1
X8 S6 N6 P1 PT8 – ISE 08 (95) 1
X8 S6 N12 P1 PT8 MS25854 (02) – 1
X9 S6 N7 P3 PT8 – ISE 09 (95) 1
X10 S3 N3 P1 PT4 – ISE 44 (96) 1
X11 S10 N11 P13 PT6a MS19987 (00) – 1
X12 S11 N3 P1 RDNC MS20618 (00) – 1
X13 S3 N3 P4 PT6b MS15450 (99) – 1
X13 S8 N10 P4 PT6a MS18135 (99), MS19060 (00),
MS20000 (00), MS22207 (01)
– 4
X13 S9 N10 P4 PT6a MS18998 (00) – 1
X14 S12 N3 P1 ND MS21924 (01) – 1
X15 S3 N8 P12 PT6a 90292 (01) – 1
*Year of strain isolation was shown in parenthesis. Data for strains isolated during 1991–1997 were those taken from Tsen and Lin (2001).
�Not determined.
1478 J . -C .PANG ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x
similarity for all the strains studied was >80%. For strains
in the major cluster, i.e. cluster A, which includes 74Æ9%,
i.e.140 of 187, of the total strains, the genetic similarity is
over 97%. The present investigation suggested that several
variants were derived from a single clonal line and the
strains of Salm. Enteritidis in Taiwan are highly similar over
a 13-year period (1991–2002). This confirmed the high
genetic homogeneity among the Salm. Enteritidis strains
reported previously by other laboratories (Millemann et al.1995; Thong et al. 1995; Landeras et al. 1996; Boonmar
et al. 1998; Bakeri et al. 2003).Plasmid profile analysis has been used as a rapid and
useful method for subtyping of some Salmonella serotypes
including Salm. Enteritidis (Rodrigue et al. 1992; Bichleret al. 1994; Wray et al. 1998). Most of the work on
epidemiological subtyping of Salm. Enteritidis concentratedon plasmid profiles. This is of special importance, as some
plasmids in salmonellae exhibit effect on bacterial virulence.
For this reason, they have been called as serotype-specific
virulence plasmids (Helmuth and Schroeter 1994). The size
of these plasmids varies with serovars, ranging from 50 to
285 kb (Ou et al. 1990). In our study, plasmid profiling
showed that most of the isolates (98Æ4%) harboured specific
virulence plasmid with molecular size approx. 58 kb (or
38 MDa). It has been reported that virulence plasmid with
size of 54–60 kb (34–38 MDa) was present in most of the
Salm. Enteritidis strains studied (Bichler et al. 1994;
Helmuth and Schroeter 1994; Liebana et al. 2001; Bakeriet al. 2003). Most Salm. Enteritidis strains harboured only
the serovar-specific virulence plasmid and cannot be further
distinguished (Liebana et al. 2001). In this study, although
Table 2 Plasmid profiles and phage types for 187 Salmonella
Enteritidis strains obtained from human origin in Taiwan during
1991–2002
Plasmid
profile
Approx.
plasmid size (kb)
Band
numbers
No. of
strains (%) Phage type
P1 58 1 99 (52Æ9) PT4, 4a, 4b,
1, 7, 8, RDNC
P2 58, 3Æ2 2 9 (4Æ8) PT27, RDNC
P3 98, 53 2 1 (0Æ5) PT8
P4 115, 58 2 19 (10Æ2) PT6a, 6b
P5 58, 8Æ3, 1Æ9 3 36 (19Æ3) PT4
P6 88, 58, 8Æ3, 1Æ9 4 1 (0Æ5) PT4
P7 58, 1Æ9 2 8 (4Æ3) PT4
P8 88, 58 2 1 (0Æ5) PT4
P9 130, 58, 3Æ5 3 2 (1Æ1) PT6a
P10 58, 4Æ5 3 1 (0Æ5) PT29
P11 70, 58 2 1 (0Æ5) PT6a
P12 115, 58, 8Æ3, 1Æ9 4 2 (1Æ1) PT6a
P13 130, 3Æ5 2 1 (0Æ5) PT6a
P14 80, 58 2 1 (0Æ5) PT6a
P15 98, 58, 3Æ2 3 1 (0Æ5) ND
P16 58, 48 2 1 (0Æ5) ND
P17 58, 45 2 1 (0Æ5) PT34
No plasmid – – 2 (1Æ1) PT4
A
B
C
D
No of strains1991~1997 1998~2002
Phage type
1 0 PT49 6 PT4, 4b, 13, 27, RDNC0 1 PT40 1 PT291 0 PT40 1 PT6a0 1 PT6a0 2 PT41 0 PT40 7 PT6a, 40 1 PT6a0 1 RDNC0 1 PT6b0 1 PT6a0 4 PT6a0 1 ND0 1 PT6a0 1 RDNC2 0 PT8, 341 0 PT80 1 PT81 0 PT846 84 PT4, 6a, 7, 4a, 10 6 PT6a0 1 PT41 0 PT40 1 PT4b0 1 PT463 124Total
Coefficient0·80 0·84 0·89 0·94 0·99
XlS1N1 XlS2N2 XlS3N2 XlS4N2 XlS3N8 Xl5S3N8 X2S3N3X10S3N3X5S2N3X5S3N3X7S3N3X13S3N3X13S9N10X13S8N10X14S12N3X6S5N5X11S10N11X12S11N3X4S4N4X8S6N6X8S6N12X9S6N7X3S3N3X3S3N8X3S5N3X3S7N3X3S7N14X3S13N13
Fig. 4 Dendrogram showing the similarities of 28 XbaI, SpeI and NotI digestion pulsed-field gel electrophoresis (PFGE) patterns from 187 strains of
Salmonella Enteritidis generated by NTSYSpc with the band-matching coefficient of Dice and the UPGMAUPGMA clustering
PFGE, PLASMID PROFILE, AND PHAGE TYPING OF SALM. ENTERITIDIS 1479
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x
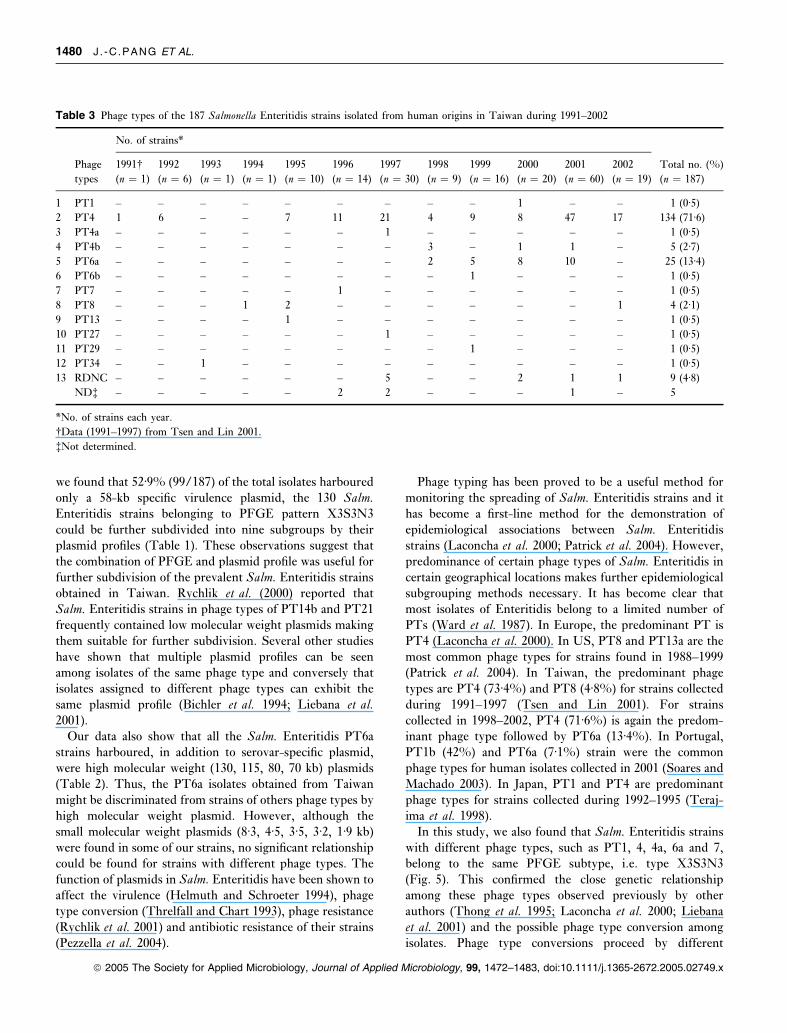
we found that 52Æ9% (99/187) of the total isolates harboured
only a 58-kb specific virulence plasmid, the 130 Salm.Enteritidis strains belonging to PFGE pattern X3S3N3
could be further subdivided into nine subgroups by their
plasmid profiles (Table 1). These observations suggest that
the combination of PFGE and plasmid profile was useful for
further subdivision of the prevalent Salm. Enteritidis strainsobtained in Taiwan. Rychlik et al. (2000) reported that
Salm. Enteritidis strains in phage types of PT14b and PT21
frequently contained low molecular weight plasmids making
them suitable for further subdivision. Several other studies
have shown that multiple plasmid profiles can be seen
among isolates of the same phage type and conversely that
isolates assigned to different phage types can exhibit the
same plasmid profile (Bichler et al. 1994; Liebana et al.2001).
Our data also show that all the Salm. Enteritidis PT6a
strains harboured, in addition to serovar-specific plasmid,
were high molecular weight (130, 115, 80, 70 kb) plasmids
(Table 2). Thus, the PT6a isolates obtained from Taiwan
might be discriminated from strains of others phage types by
high molecular weight plasmid. However, although the
small molecular weight plasmids (8Æ3, 4Æ5, 3Æ5, 3Æ2, 1Æ9 kb)
were found in some of our strains, no significant relationship
could be found for strains with different phage types. The
function of plasmids in Salm. Enteritidis have been shown to
affect the virulence (Helmuth and Schroeter 1994), phage
type conversion (Threlfall and Chart 1993), phage resistance
(Rychlik et al. 2001) and antibiotic resistance of their strains
(Pezzella et al. 2004).
Phage typing has been proved to be a useful method for
monitoring the spreading of Salm. Enteritidis strains and it
has become a first-line method for the demonstration of
epidemiological associations between Salm. Enteritidis
strains (Laconcha et al. 2000; Patrick et al. 2004). However,
predominance of certain phage types of Salm. Enteritidis incertain geographical locations makes further epidemiological
subgrouping methods necessary. It has become clear that
most isolates of Enteritidis belong to a limited number of
PTs (Ward et al. 1987). In Europe, the predominant PT is
PT4 (Laconcha et al. 2000). In US, PT8 and PT13a are the
most common phage types for strains found in 1988–1999
(Patrick et al. 2004). In Taiwan, the predominant phage
types are PT4 (73Æ4%) and PT8 (4Æ8%) for strains collected
during 1991–1997 (Tsen and Lin 2001). For strains
collected in 1998–2002, PT4 (71Æ6%) is again the predom-
inant phage type followed by PT6a (13Æ4%). In Portugal,
PT1b (42%) and PT6a (7Æ1%) strain were the common
phage types for human isolates collected in 2001 (Soares and
Machado 2003). In Japan, PT1 and PT4 are predominant
phage types for strains collected during 1992–1995 (Teraj-
ima et al. 1998).In this study, we also found that Salm. Enteritidis strains
with different phage types, such as PT1, 4, 4a, 6a and 7,
belong to the same PFGE subtype, i.e. type X3S3N3
(Fig. 5). This confirmed the close genetic relationship
among these phage types observed previously by other
authors (Thong et al. 1995; Laconcha et al. 2000; Liebanaet al. 2001) and the possible phage type conversion among
isolates. Phage type conversions proceed by different
Table 3 Phage types of the 187 Salmonella Enteritidis strains isolated from human origins in Taiwan during 1991–2002
No. of strains*
Total no. (%)
(n ¼ 187)
Phage
types
1991�(n ¼ 1)
1992
(n ¼ 6)
1993
(n ¼ 1)
1994
(n ¼ 1)
1995
(n ¼ 10)
1996
(n ¼ 14)
1997
(n ¼ 30)
1998
(n ¼ 9)
1999
(n ¼ 16)
2000
(n ¼ 20)
2001
(n ¼ 60)
2002
(n ¼ 19)
1 PT1 – – – – – – – – – 1 – – 1 (0Æ5)2 PT4 1 6 – – 7 11 21 4 9 8 47 17 134 (71Æ6)3 PT4a – – – – – – 1 – – – – – 1 (0Æ5)4 PT4b – – – – – – – 3 – 1 1 – 5 (2Æ7)5 PT6a – – – – – – – 2 5 8 10 – 25 (13Æ4)6 PT6b – – – – – – – – 1 – – – 1 (0Æ5)7 PT7 – – – – – 1 – – – – – – 1 (0Æ5)8 PT8 – – – 1 2 – – – – – – 1 4 (2Æ1)9 PT13 – – – – 1 – – – – – – – 1 (0Æ5)10 PT27 – – – – – – 1 – – – – – 1 (0Æ5)11 PT29 – – – – – – – – 1 – – – 1 (0Æ5)12 PT34 – – 1 – – – – – – – – – 1 (0Æ5)13 RDNC – – – – – – 5 – – 2 1 1 9 (4Æ8)
ND� – – – – – 2 2 – – – 1 – 5
*No. of strains each year.
�Data (1991–1997) from Tsen and Lin 2001.
�Not determined.
1480 J . -C .PANG ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x
mechanisms, for example, the interrelationships between
PT4, 7, 7a, 8, 13, 13a, 23, 24 and 30 caused by the loss or
acquisition of an incN plasmid (Frost et al. 1989; Threlfalland Chart 1993), the conversion of Salm. enterica serovar
Enteritidis PT1, 4, 6 to PT 7 involved the loss of
lipopolysaccharide (Baggesen et al. 1997) and phage con-
version by the temperate phages. Phage types PT4, 6a, 6a,
13 and 15 were converted to PT 8, 4, 7, 13a and 11
respectively (Rankin and Platt 1995). In addition, none of
the four PT8 strains showed PFGE pattern of X3S3N3,
which was the most common subtype for strains of other
phage types. Similar results have been reported by Laconcha
et al. (2000).Many reports mentioned that a single method cannot be
relied on for discriminating between Salm. Enteritidis
strains. Combination of several typing methods could
provide a powerful discriminatory tool for the epidemiolog-
ical analysis of unrelated and related strains of Salm.Enteritidis (Laconcha et al. 1998, 2000; Liebana et al. 2001).In this study, we combined the data of PFGE, plasmid
profile and phage typing for the epidemiological typing of
Salm. Enteritidis isolates. The three methods identified
different degrees of polymorphism in the following order:
plasmid profile (18 types, D ¼ 0Æ659) > PFGE (28 types,
D ¼ 0Æ512) > phage typing (13 types, D ¼ 0Æ438). Combi-
nation of the three methods generated 47 subtypes (Table 1)
and the discriminatory power was raised (D ¼ 0Æ795). Thepredominant subtype was X3S3N3/P1/PT4, which consists
of 77 (41%) isolates. Finally, it should be mentioned that the
discriminatory power of PFGE for our isolates is lower than
those reported by other laboratories (Laconcha et al. 1998,2000). This might be because of the fact that our Salm.Enteritidis strains were obtained from human origin only
and recirculating of certain major subtype of strains in
Taiwan is possible.
The subtyping data obtained from this study may be
useful when compared with the epidemiological data
obtained from other countries. Through such comparison,
the major subtypes for human infection by Salm. Enteritidismay be elucidated. The data established here may also be
useful for tracing the transmission route for Salm. Enterit-idis infection.
In conclusion, we found that certain subtypes of Salm.Enteritidis prevailed in Taiwan. This might be due to the
environmental factors or the pathogenic properties linked
to Salm. Enteritidis isolates. Olsen et al. (1999) have
reported that levels of virulence are not determined by
genomic lineage of Salm. Enteritidis strains. Thus, the
presence of major subtypes and limited clonal diversity for
Salm. Enteritidis strains may be simply due to the limited
genetic rearrangement or recombination during their
evolution.
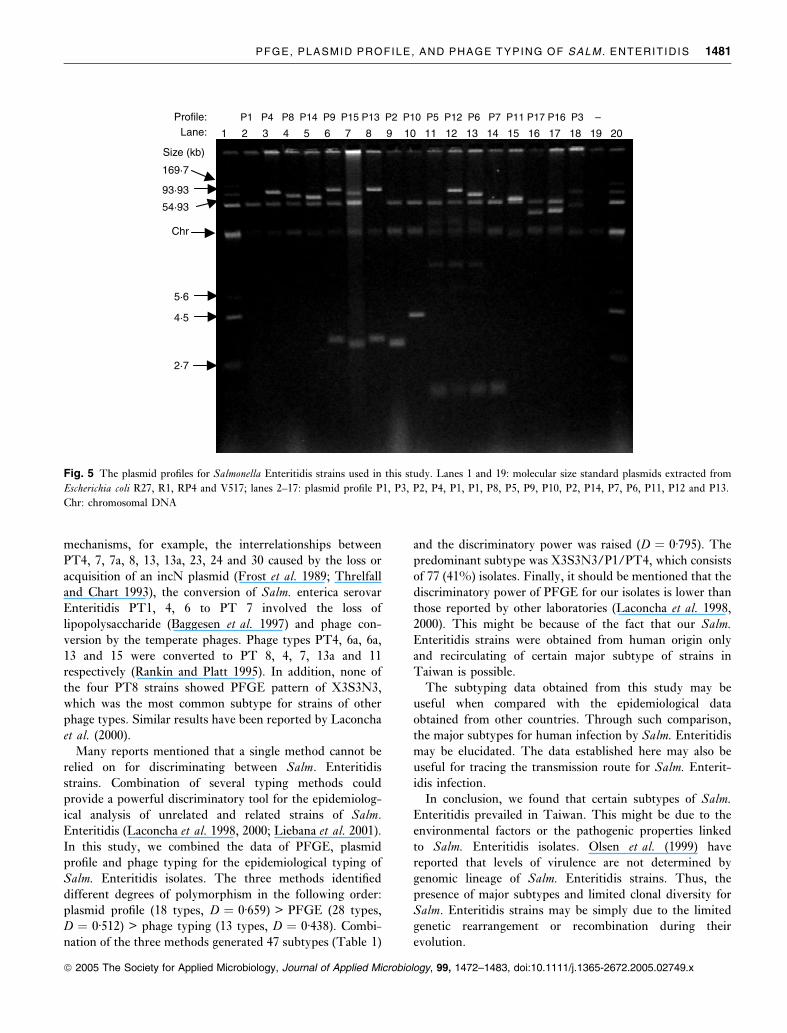
Profile:Lane:
Size (kb)
4·5
2·7
5·6
93·93
169·7
54·93
Chr
1
P1 P4 P8 P14 P9 P15 P13 P2 P5 P6 P7 P3 –P10 P12 P11 P17 P16
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Fig. 5 The plasmid profiles for Salmonella Enteritidis strains used in this study. Lanes 1 and 19: molecular size standard plasmids extracted from
Escherichia coli R27, R1, RP4 and V517; lanes 2–17: plasmid profile P1, P3, P2, P4, P1, P1, P8, P5, P9, P10, P2, P14, P7, P6, P11, P12 and P13.
Chr: chromosomal DNA
PFGE, PLASMID PROFILE, AND PHAGE TYPING OF SALM. ENTERITIDIS 1481
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x

ACKNOWLEDGEMENTS
This research was supported by a grant from the National
Science Council, Taipei, Taiwan, ROC. The project
numbers were NSC 92-2313-B-241-007 and 93-2313-B-
241-001.
REFERENCES
Baggesen, D.L., Wegener, H.C. and Madsen, M. (1997) Correlation of
conversion of Salmonella enterica serovar Enteritidis phage type 1, 4,
or 6 to phage type 7 with loss of lipopolysaccharide. J Clin Microbiol
35, 330–333.
Bakeri, S.A., Yasin, R.M., Koh, Y.T., Puthucheary, S.D. and Thong,
K.L. (2003) Genetic diversity of human isolates of Salmonella
enterica serovar Enteritidis in Malaysia. J Appl Microbiol 95, 773–
780.
Bichler, L.A., Nafaraja, K.V. and Pomeroy, B.S. (1994) Plasmid
diversity in Salmonella Enteritidis of animal, poultry and human
origin. J Food Prot 57, 4–11.
Boonmar, S., Bangtrakulnonth, A., Pornrunangwong, S., Terajima, J.,
Watanabe, H., Kaneko, K.I. and Ogawa, M. (1998) Epidemiological
analysis of Salmonella enteritidis isolates from humans and broiler
chickens in Thailand by phage typing and pulsed-field gel electro-
phoresis. J Clin Microbiol 36, 971–974.
Frost, J.A., Ward, L.R. and Rowe, B. (1989) Acquisition of a drug
resistance plasmid converts Salmonella Enteritidis phage type 4 to
phage type 24. Epidemiol Infect 103, 243–248.
Helmuth, R. and Schroeter, A. (1994) Molecular typing methods for
Salmonella Enteritidis. Int J Food Microbiol 21, 69–77.
Kado, C.I. and Liu, S.T. (1981) Rapid procedure of detection and
isolation of large and small plasmids. J Bacteriol 145, 1365–1373.
Kotetishvili, M., Stine, O.C., Kreger, A., Morris, J.G. Jr and
Sulakvelidze, A. (2002) Multilocus sequence typing for character-
ization of clinical and environmental Salmonella strains. J Clin
Microbiol 40, 1626–1635.
Laconcha, I., Lopez-Molina, N., Rementeria, A., Audicana, A.,
Perales, I. and Garaizar, J. (1998) Phage typing combined with
pulsed-field gel electrophoresis and random amplified polymorphic
DNA increases discrimination in the epidemiological analysis of
Salmonella enteritidis strains. Int J Food Microbiol 40, 27–34.
Laconcha, I., Baggesen, D.L., Rementeria, A. and Garaizar, J. (2000)
Genotypic characterisation by PFGE of Salmonella enterica serotype
Enteritidis phage types 1, 4, 6 and 8 isolated from animal and human
sources in three European countries. Vet Microbiol 75, 155–165.
Landeras, E., Gonzalez-Hevia, M.A., Alzugaray, R. and Mendoza,
M.C. (1996) Epidemiological differentiation of pathogenic strains
of Salmonella Enteritidis by ribotyping. J Clin Microbiol 34, 2294–
2296.
Liebana, E., Garcia-Migura, L., Breslin, M.F., Davies, R.H. and
Woodward, M.J. (2001) Diversity of Salmonella enterica serotype
Enteritidis from English poultry farms assessed by multiple genetic
fingerprinting. J Clin Microbiol 39, 154–161.
Mahon, J. and Lax, A.J. (1993) A quantitative polymerase chain
reaction method for the detection in avian faeces of salmonellas
carrying the spvR gene. Epidemiol Infect 111, 455–464.
Millemann, Y., Lesage, M.C., Chaslus-Dancla, E. and Lafont, J.P.
(1995) Value of plasmid profiling, ribotyping and detection of IS200
for tracing avian isolates of Salmonella Typhimurium and
S. Enteritidis. J Clin Microbiol 33, 173–179.
Murakami, K., Horikawa, K. and Otsuki, K. (1999) Epidemiological
analysis of Salmonella Enteritidis from human outbreaks by pulsed-
field gel electrophoresis. J Vet Med Sci 4, 439–442.
Olsen, J.E., Skov, M.N., Threlfall, E.J. and Brown, D.J. (1994) Clonal
lines of Salmonella enterica serotype Enteritidis documented by
IS200-, ribo-, pulsed-field gel electrophoresis and RFLP typing.
J Med Microbiol 40, 15–22.
Olsen, J.E., Tiainen, T. and Brown, D.J. (1999) Levels of virulence are
not determined by genomic lineage of Salmonella enterica serotype
Enteritidis strains. Epidemiol Infect 123, 423–430.
Ou, J.T., Baron, L.S., Dai, X. and Life, C.A. (1990) The virulence
plasmids of Salmonella serovars Typhimurium, Choleraesuis, Dub-
lin, and Enteritidis, and the cryptic plasmids of Salmonella serovars
Copenhagen and Sendai belong to the same incompatibility group,
but not those of Salmonella serovars Durban, Gallinarum, give,
Infantis and Pullorum. Microb Pathol 8, 101–107.
Pan, T.M., Wang, T.K., Lee, C.L., Chien, S.W. and Horng, C.B.
(1997) Food-borne disease outbreaks due to bacteria in Taiwan, 1986
to 1995. J Clin Microbiol 35, 1260–1262.
Patrick, M.E., Adcock, P.M., Gomez, T.M., Altekruse, S.F.,
Holland, B.H., Tauxe, R.V. and Swerdlow, D.L. (2004) Salmon-
ella Enteritidis infections, United States, 1985–1999. Emerg Infect
Dis 10, 1–7.
Pezzella, C., Ricci, A., DiGiannatale, E., Luzzi, I. and Carattoli, A.
(2004) Tetracycline and streptomycin resistance genes, transposons,
and plasmids in Salmonella enterica isolates from animals in Italy.
Antimicrob Agents Chemother 48, 903–908.
Rankin, S. and Platt, D.J. (1995) Phage conversion in Salmonella
enterica serotype Enteritidis: implications for epidemiology. Epidem-
iol Infect 114, 227–236.
Rodrigue, D.C., Tauxe, R.V. and Rowe, B. (1990) International
increase in Salmonella Enteritidis: a new pandemic? Epidemiol Infect
105, 21–27.
Rodrigue, D.C., Cameron, D.N., Puhr, N.D., Brenner, F.W., St.
Louis, M.E., Wachsmuth, I.K. and Tauxe, R.V. (1992) Comparison
of plasmid profiles, phage types, and antimicrobial resistance
patterns of Salmonella Enteritidis isolates in the United States.
J Clin Microbiol 30, 854–857.
Rychlik, I., Svestkova, A. and Karpiskova, R. (2000) Subdivision of
Salmonella enterica serovar Enteritidis phage types PT14b and PT21
by plasmid profiling. Vet Microbiol 74, 217–225.
Rychlik, I., Sebkova, A., Gregorova, D. and Karpiskova, R. (2001)
Low-molecular-weight plasmid of Salmonella entericaserovar
Enteritidis codes for retron reverse transcriptase and influences
phage resistance. J Bacteriol 183, 2852–2858.
Soares, A.R. and Machado, J. (2003) Molecular epidemiology of
Salmonella Enteritidis phage type 1b and 6a isolates in Portugal.
Epidemiol Infect 131, 607–612.
Su, L.H., Chiu, C.H., Kuo, A.J., Chia, J.H., Sun, C.F., Leu, H.S. and
Wu, T.L. (2001) Secular trends in incidence and antimimicrobial
resistance among clinical isolates of Salmonella at a university hospital
in Taiwan, 1983–1999. Epidemiol Infect 127, 207–213.
1482 J . -C .PANG ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x

Terajima, J., Nakamura, A. and Watanabe, H. (1998) Epidemiological
analysis of Salmonella enterica Enteritidis isolates in Japan by phage-
typing and pulsed-field gel electrophoresis. Epidemiol Infect 120,
223–229.
Thong, K.L., Ngeow, Y.F., Altwegg, M., Navaratnam, P. and Pang,
T. (1995) Molecular analysis of Salmonella Enteritidis by pulsed-
field gel electrophoresis and ribotyping. J Clin Microbiol 33, 1070–
1074.
Threlfall, E.J. and Chart, H. (1993) Interrelationships between strains
of Salmonella Enteritidis. Epidemiol Infect 111, 1–8.
Tsen, H.Y. and Lin, J.S. (2001) Analysis of Salmonella Enteritidis
strains isolated from food-poisoning cases in Taiwan by pulsed field
gel electrophoresis, plasmid profile and phage typing. J Appl
Microbiol 91, 72–79.
Wang, T.K., Tsai, C.L., Lin, C.S., Ho, S.I., Chiu, C.S., Hsu, S.Y.,
Huang, H.C. and Pan, T.M. (1999) Recent trend of Salmonella food
poisoning and serovars. Epidemiol Bull 15, 1–6.
Ward, L.R., de Sa, J.D. and Rowe, B. (1987) A phage-typing scheme
for Salmonella Enteritidis. Epidemiol Infect 99, 291–294.
Wray, C., McLaren, I.M. and Jones, Y.E. (1998) The epidemiology of
Salmonella Typhimurium in cattle: plasmid profile analysis of
definitive phage type (DT) 204c. J med microbiol 47, 483–487.
Yan, S.S., Pendrak, M.L., Abela-Ridder, B., Punderson, J.W.,
Fedorko, D.P. and Foley, S.L. (2004) An overview of Salmonella
typing: public health perspectives. Clin Appl Immunol Rev 4, 189–
204.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 99, 1472–1483, doi:10.1111/j.1365-2672.2005.02749.x
PFGE, PLASMID PROFILE, AND PHAGE TYPING OF SALM. ENTERITIDIS 1483




















