
Protozoa of soil - Wilhelm FOISSNER
8
Protistological Actual ities (Proccedings of the Secorrd European Congress of Protistology Clernrortt-Ferrand, I 995) Ilds : G. Brugerolle and J.-P. Mignot Protozoaof Soil Symposium 5 WILHELM FOISSNER Universität Salzburg, Institut für Zoologie, Hellbrunnerstrasse 34, A-5020 Salzburg (Austria), MANFRED WANNER RWTH Aachcn, Institut für Biologic II, Kopcrnikusstrasse 16, D-52056 Aachcn (Gcrmany) With contributions from Erna Aescht, John F. Darbyshire, Julius G. Geltzer, and Ralf Meisterfeld Summary This paper reports on a symposium organizcd by W. Foissner (chair and report) and M. Wanner (co-chair) on thc occasion of the Sccond Europcan Congrcss of Protistology in Clermont-Ferrand,21-27 July 1995. The symposium emphasized aspects of applied soil proto- zoology and treated the following subjects in some detail: integrating soil protozoology with general soil and environmental sciences, soil protozoan diversity, bioindication and autecolo- gy, soil protozoology in Russia. A guidc to rcccnt rcvicws and books on soil protozoa is inclu- dcd. Introduction Soil protozoology is rejuvenating after a long period of stagnation between 1930 and 1970. This is reflected not only by several important recent reviews [1, 6-8, 13, '14, 16-18] but also by the Second European Congress of Protistology, which highlighted the importance of soil protozoa by dcdicating two symposia to thc subjcct (scc report by E. Piccini in this vo- lume). Rcscarch during the past 20 years providcd convincing cvidence that protozoa play an important role in the energy and nutrient flux of soil ecosystems [6-8, 18]. In spite of this, protozoa are still insufficiently recognized by general ecologists and soil scientists. Thus, John F. Darbyshire, the grand old man of soil protozoology, was invited to lecture on possible ways out of the isolation. A theme inhcrent also in thc contributions by Erna Acscht, Ralf Meister- feld, and Manfred Wanner, who prescnted aspects of applied soil protozoology, namely bioin- dication, an increasingly important field. However, bioindication requires, as the word implies, a profund knowledge of species and community structures. Unfortunately, soil protozoan di- versity is still very insufficiently known. This was highlighted by Wilhelm Foissner in his ti'o' 128
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Protozoa of soil - Wilhelm FOISSNER

Protistological Actual ities(Proccedings of the Secorrd European Congress of ProtistologyClernrortt-Ferrand, I 995)Ilds : G. Brugerolle and J.-P. Mignot
Protozoaof Soil
Symposium 5
WILHELM FOISSNERUniversität Salzburg, Institut für Zoologie, Hellbrunnerstrasse 34, A-5020 Salzburg
(Austria),
MANFRED WANNERRWTH Aachcn, Institut für Biologic II,
Kopcrnikusstrasse 16, D-52056 Aachcn (Gcrmany)
With contributions from Erna Aescht, John F. Darbyshire, Julius G. Geltzer,and Ralf Meisterfeld
Summary
This paper reports on a symposium organizcd by W. Foissner (chair and report) andM. Wanner (co-chair) on thc occasion of the Sccond Europcan Congrcss of Protistology inClermont-Ferrand,21-27 July 1995. The symposium emphasized aspects of applied soil proto-zoology and treated the following subjects in some detail: integrating soil protozoology withgeneral soil and environmental sciences, soil protozoan diversity, bioindication and autecolo-gy, soil protozoology in Russia. A guidc to rcccnt rcvicws and books on soil protozoa is inclu-dcd.
Introduction
Soil protozoology is rejuvenating after a long period of stagnation between 1930 and1970. This is reflected not only by several important recent reviews [1, 6-8, 13, '14, 16-18] butalso by the Second European Congress of Protistology, which highlighted the importance ofsoil protozoa by dcdicating two symposia to thc subjcct (scc report by E. Piccini in this vo-lume).
Rcscarch during the past 20 years providcd convincing cvidence that protozoa playan important role in the energy and nutrient flux of soil ecosystems [6-8, 18]. In spite of this,protozoa are still insufficiently recognized by general ecologists and soil scientists. Thus, JohnF. Darbyshire, the grand old man of soil protozoology, was invited to lecture on possible waysout of the isolation. A theme inhcrent also in thc contributions by Erna Acscht, Ralf Meister-feld, and Manfred Wanner, who prescnted aspects of applied soil protozoology, namely bioin-dication, an increasingly important field. However, bioindication requires, as the word implies,a profund knowledge of species and community structures. Unfortunately, soil protozoan di-versity is still very insufficiently known. This was highlighted by Wilhelm Foissner in his
ti'o'
128

lecture on soil ciliates from the Etosha National Park: 53 out of 153 specics found in 12 sam-plcs wcre ncw to sciencc.
Finally, Julius Celtzer was invitcd introduce us to the vast Eastern European litera-ture on soil protozoa, which has been widely neglected, probably because few colleagues cancope with the Russian language. Thus, our Russian colleagues should be encouraged to publishin English or at least to furnish their papers with extensive English summaries.
Integrating soil protozoology with general soil and environmental sciences
Dr John Darbyshire (Macauly Land Usc Rcscarch Institute, Scotland) prcscntcd a
storm of idcas as to how soil protozoology could bc more closcly relatcd to cnvironmcntalsciences in general and soil science in particular. He emphasized the need for soil protozoolo-gists to promote their discipline more rigorously and to improve the situation themselves byintegrating their studies more closely with the rest of soil ecology at the levels of ecotones,ecosystems and landscapes. John Darbyshire's papcr was read by H. G. Smith (Coventry Uni-vcrsity, England) bccause John was unable to attcnd the congress.
In order to examine ways in which soil protozoology might be integrated with wideraspects of soil ecology we need to be aware of the complex and heterogeneous nature of thesoil ecosystem and also that the species composition and the population fluctuations of theprotozoan fauna are still imperfectly known.
Estimatcs of soil protozoan populations may possibly bc improvcd by a two-phasepartitioning method devclopcd by Smith & Striblcy [19]. Air-dricd soil is added to an aqucousmixturc of dcxtran and polycthylcne glycol, microorganisms in the organic phase bcing rcmo-ved by pipette.
It may be rewarding to correlate changes in soil solution and atmosphere with proto-zoan populations as these may suggest hitherto unsuspected interactions which merit furtherinvcstigation. Suitablc mcthods for sampling soil solution and atmosphcrc alrcady exist: [-ow-spccd ccntrifugation on filtration can providc sufficient volumcs of solution for analysis byinductivcly-couplcd plasma atomic cmission spectrometry. Dcvclopments in gas chromatogra-phy enable the determination of the gas composition of soil atmosphere withdrawn by syringe
[11].There remains the problem of how to deal with soil heterogeneity. Geostatistics offer
a potcntial mcans of studying thc spatial rclationships bctwcen protozoan populations andcither soil micro.bial proccsscs or plant distribution. This has bcen uscd to link microbial bio-mass and carbon mincralization with thc dispcrsion of sage-bush plants in scmi-arid ecosys-tems [20], to relate collembolan and microbial populations to carbon content in cultivated soils
[10], and to study the distribution of soil invertebrates and litter accumulation under tropicalbull-oak plantations.
A third potentially fruitful field of work is protozoa-bactcria-plant pathogen relations.Lcvrat ct al. [15] havc shown that cell-frcc cxudatcs of Acanthamoeba castellani can
stimulate Pseudomonas putida to supprcss the growth and sporulation of the Fusarium wiltfungal pathogen. There is also evidence that fungal metabolites may stimulate the growth ofmycophagous giant amoebae. If the chemical identity and mode of action of microbial extra-cellular metabolites can be determined, then many new roles for soil protozoa may yet bediscovercd.
129

Soil protozoan diversity
Dr Wilhclm Foissner (Salzburg Univcrsity, Austria) hijackcd thc audicncc to onc ofthc niccst placcs of thc world, thc Etosha National Park in Namibia, Southwcst Africa, whcrehe started a project on taxonomy of tropical soil ciliates. His contribution, amended with manybeautiful slides from soil ciliates of the Etosha Pan, showed our profund ignorance of soilprotozoan diversity, especially in the tropics and subtropics, where a single sample may con-tain up to 80 ciliatc specics [9]. Dr Foissncr complained that too many pcoplc arc spcaking andpublishing about "biodivcrsity" and too fcw doing thc "hard work", i. c. dctcrmining andcventually dcscribing ncw spccics. Of'ten, biodivcrsity has bccn misuscd by ccologists toaquire money for pure ecological rcsearch, e. g. for studying cncrgy fluxes etc. at high taxalevel. This is by no means biodiversity! Biodiversity needs species and individuals which arethe centres of evolution; higher taxa, such as genera, orders, and functional groups are(hclpful) artificial constructs.
Table 1. Number of soil ciliatc specics in a transcct of thc Etosha National Park, Namibia
Biotope PI-I Number of ciliate speciesr
Pan (saline desert)
Pan margin, Suaeda zonc
Thorn bush savanna (1 kmpan margin)
distant from pan margin)
9.7 -9.7
8.6-8.4
distant from 7 .7
7.1
I -21
43-57
28
37t Obtaincd wilh the non-flooded pctri dish mcthod tts dcscribetl in [8].
Dr Foissncr invcstigatcd 12 soil samplcs for ciliatcs from thc ccntrc and pcriphcry ofthe Etosha Pan. The pan soil is a very special mixture of salt, clay, and lime having a pH rangeof about 8.0-9.7; the air-dried mixture is like a stone, but quickly doubles its volume and be-comes a fluffy pancake when it is rewetted. Most of the soil is covered with a more or lessdistinct laycr of filamcntous cyanobactcria. 153 ciliatc spccics wcrc found, 53 (!) of whichwcre ncw to scicnce. Most belonged to onc of thc following groups: hypotrichs (43 spccics),colpodids (35), gymnostomatids (33), nassulids (15).
The high number and frequency of nassulid ciliates, usually occurring sparsely insoil, was obviously related to the commonness of cyanobacteria, their preferred food. A tran-sect from the pan to the surrounding savanna showed that the salt shrub (Suaeda) region hadthe highcst spccics richncss and that thc numbcr of spccics sharply dccrcascd abovc pH 8.6(Tab. 1). Refined ccological rcscarch on these spccial ciliatc communitics is urgcntly nccdcdbut difTicult to rcalize because of the high number of ncw, as yct undcscribcd spccics.
Bioindication and autecology
Dr Erna Aescht (Biology Center of the Upper Austrian Natural History Museum)gave a general lecture on the potentialities and limits of soil protozoa as bioindicators in envi-ronmental field studies. Shc summarizcd pcrtincnt prcvious studics [1] and vcry reccnt data,mainly by Bcrthold & Palzcnbcrgcr [4].
Rcscarch in most groups of soil protozoa is hampcrcd by mcthodological problcmsand the lack of taxonomic guides for species. Dr Aescht emphasized that the widely used cul-
130

ture (dilution) tcchniqucs cannot rcliably discriminate active and cystic protozoa. Furthermore,thcy arc time-consuming and are oftcn statistically inadequate. Thus, direct counts in dilutcdsoil suspcnsions should be pret'erred. Dircct counting providcs data on activc individuals andspecies, which are prerequisites tbr using protozoa as bioindicators in the soil environment.Unfortunately, no reliable direct methods are known for counting naked amoebae and small,amoeboid flagellates which adhere to the soil particles. Thus, practical bioindication work is atprcsent almost cntircly restrictcd to ciliates and testate amoebae, for which standardized andpropcrly tcstcd direct counting methods havc bcen publishcd [2, 3]. Ciliatcs must be countedon thc day of sampling duc to thcir ability to cncyst and excyst rapidly, while testacean collec-tions can be conserved and stored for years. Ciliates and testate amoebae are equivalent indi-cator groups in raw soils, e. g. the litter layer. In evolved natural and cultivated soils testateamoebae are much more important than ciliates (and, probably, naked amoebae and flagellates,too), whosc activity is strongly supprcsscd by microbiostatic cffccts [8]. Thc data availableshow that total individual and spccics numbcrs and/or functional groups arc oftcn insufficientin rcvcaling trcatmcnt cfTccts.
These overall parameters fiequently obscure the fact that some indicator species de-crease or, respectively, increase. Bioindication thus needs to be done at species level. Howe-ver, this is time consuming and Dr Aescht thus suggested restricting identification to dominant(> 2%) spccics, at lcast in applicd environmcntal studics.
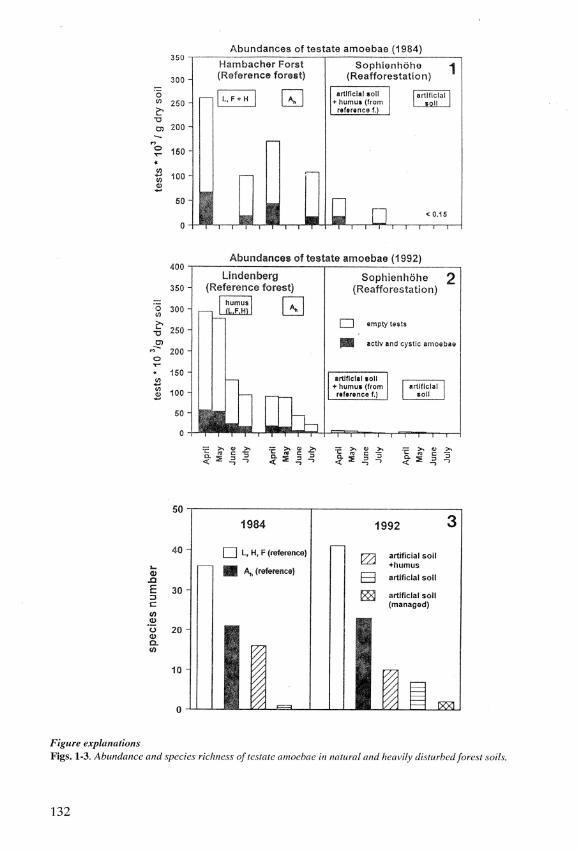
Dr Acscht's conclusions wcre impressivcly supportcd by an unpublishcd study of DrRalf Mcistcrfeld's group (Aachcn Univcrsity, Cermany) on thc rccolonization of heavily dis-turbed forest soils. His data showed convincingly that testate amoebae are more sensitive indi-cators and tär better suited to monitoring soil development than microbial activity andabundances of flagellates and naked amoebae (Figs. 1-3). Unfortunately, ciliates were notstudied.
As a rcsult of opcn cast mining of brown coal, hcaps of overburdcn (arca up to10 km2, height up to 200 m) werc deposited. To allow reafforcstation, thc almost stcrilc sandymound was covered with artificial soil made up of 20 7o loess and 80 Vo sand and more than 10million trees (mainly beech and oak) were planted afterwards. A small experimental area wasamended with humus from a primary tbrest to stimulate succession. To monitor the develo-pmcnt of soil biota, soil rcspiration, microbial biomass, flagellatcs as wcll as nakcd and tcstateamocbac wcrc studicd. During a sampling pcriod in .l984
almost all thcsc paramctcrs dcclinedsignificantly from primary forcst ovcr forcst humus on the mound to the artificial soil (Figs. 1,3). ln 7992 the study was repeated with somewhat different results (Figs. 2, 3). Only the orga-nic horizon of the primary forest had a significantly higher respiration than the soils on theheap. Microbial biomass followed a similar trend with only small differences between the Ahhorizon of thc primary forcst and thc top soil on thc heap. During summcr, microbial biomasscstimates for thc soils on thc hcap wcre higher. Flagcllatcs and nakcd amocbac of thc soils onthc hcap had similar or highcr abundanccs than those of thc Ah horizon of thc primary forcst.Testate amoebae showed a different trend. Abundances in the forest humus on the heap wereonly 7 Vo of lhat of the Ah horizon of the reference forest, and the artificial soils had even lessindividuals. Species numbers on the heap were only 25 Vo of those of the primary forest (Figs.2, 3). Furthcrmore, most forcst litter and humus-spccific spccics (e. g. Centropyxis spp., Cy-clopyxis spp., Plagiopyx rpp., Trigonopyxis spp., Nebela spp.) wcrc lost during thc first ycarof succcssion. Only a fcw ubiquists rcmaincd, viz. Trinen a spp., small Euglypha spp., andP hry ganel la acropodia.
As a result of this study, the amendment of artificial soils with forest humus does notseem to be the appropriate strategy. This technique is very expensive and its effects are quitetransicnt. It was not possible to cstablish a typical community of humus and litter tcstateamocbac. Although abundanccs and spccics numbcrs are still highcr on the trcatcd plots, dif-fbrcnccs bccomc smallcr and will probably disappcar within a fbw ycars. Community dcvclo
131
Abundances of testate amoebae (1984)
Hambacher Forst(Referenco forest)
S oph len höhe( Re affo restati o n )
=oo zsobE6l) 200
(.,
3 160Ioü 100o+,
=oUI
bEo)
(t,
oF
I
.h
.ao
350
300
0
400
350
300
250
200
150
100
50
0
LollE30=coo'6 zooo-o
Figure explanationsFigs. 1-3. Abundance and specics richncss of tcstate amoebae h natural and heavily disturbed forest soils.
132
AbundaRcoo of tostate arnoebae (1992)
Lindenberg(Reference forest)
Sophienhöhe 2(Reaffo restatio n)
f] omps tests
M rctlv and cystlc amoebae
ftrttfirl.l r"ll I1.1;;;;iä, I t;trl.TI rcfrrence f.) I I roll I
I 984
f] t- H, F (rederence)
E d (reference)
1ee2 3
f7V artificial soil(rr +humus
t arttficlal soll
ffi artificlal soll(managed)
FfiI E I artlflclal roll I Fdm"t"ill+ humur (from I I sorr I
I rsfore nce f.) I
mtr

pment in this stagc of succössion docs not dcpcnd on immigration but on availability of suita-ble microhabitats. Typical forcst spccics can obviously only survivc and cstablish larger po-pulations in evolved, humus rich zonal soils having a distinct litter layer. In young standsannual litter input is too low and the rate of decay too high to allow the accumulation of a
sufficiently thick litter and fermentation layer. As a consequence, adverse abiotic factors likedcsiccation have an immediate efTect on the tcstacean community. The mincral soil bclow 5cm was still almost sterile, compact and had a low porc volume and could thus not scrve as a
rcfugc for mcdium and largc-sizcd protozoan specics.Finally, Dr Wanner (Aachen University, Germany) enlarged on the practical aspects
discussed by Dr Aescht and Dr Meisterfeld. He presented a very fine piece of work on theecology and taxonomy of a common soil testate amoebae,Cyclopyxis kahli. The results pre-sented wcrc bascd on 500 000 (!) morphomctrically analyzed shclls and includcd both publis-hcd [21] and unpublishcd data. Dr Wanncr cmphasized that for ccological (e. g. bioindication)and taxonomical purposcs thc proportion of cnvironmentally dcpcndcnt variability to the totalmust be known.
In a first experiment with clonal batch cultures of Cyclopyxis kahli, the influence oflime and mineral fertilizer was tested by measuring shell size of the amoebae using an image-analyzing systcm [21]. Thc control showcd thc smallcst mcan shcll diamctcr (82 ptm), whilc an
avcragc of 93 pm was mcasurcd in the fertilizcd culturcs, a significant diffcrencc of more than10 mikrons. Thc shcll opcning was invcrsely affccted, with thc largcst opcning in the controlgroup. Because the food yeast was more abundant in the treated cultures than in the controlgroup, amoebae growth may have been influenced indirectly by food supply. Therefore a se-cond experiment with tbod supply changing across dif'ferent temperatures (15"C,20'C) wascstablishcd. Amocbae fcd by bactcria formcd significantly smaller shells than those fcd byycast across thc two tcmpcraturcs. Bactcria fcd amocbac kcpt at 20"C had significantly smallershells than thosc kcpt at 15"C, whercas no tcmpcraturc cffccts occurred when ycast was fed.Food and temperature interacted highly significantly: temperature effects were compensatedby the influence of yeast well utilized by the amoebae with the consequence that the smallestshells occurred when bacterial food was provided at the higher temperature. Inverse relationswcrc obscrvcd conccrning shcll opcning.
A complcx set of consccutive cxpcrimcnts [21] allowcd a dctailcd statistical analysisof intcraction betwcen and adaptations to scvcral cnvironmcntal factors. Significant changcs inculture growth (lag time, generation time, final culture density) and shell size parameters de-pended primarily on food and temperature, whereas insecticides had a minor but significantinfluence. Lime and fertilizer had no direct effects. Interaction between all tested factors oc-currcd frcqucntly, but no consistcnt adaptation phcnomcna could be observed. Thc shell sizecfTccts wcrc revcrsible within a fcw days. Shcll sizc of Cyclopyxis kahli was significantlyaffcctcd by cnvironmcntal conditions, in particular food and tempcrature. This was also provcnon other clones and taxa of testate amoebae. Therefbre new possibilities in bioindication maybe conceivable on the one hand, but on the other taxonomical problems may arise becauseseparation of closely related taxa depends largely on shell size. Therefore a genetic approach,bascd on RAPD-PCR 122),was dcveloped to complcte convcntional ccological and taxonomi-cal results.
Nearly all analyzcd cultures of tcstate amocbae (Cyclopyxis kahli, C. eurystoma,Eu-glypha slrigosa,Trürcma lineare) provided specific fingerprints. It was, however, surprisingthat some cultures of Cyclopyxis kahli showed similar, clone-specific patterns of amplifiedDNA-fragments with and without the amoebae, although no eukaryotic contamination wascvidcnt. Additional cxpcrimcnts corroboratcd the assumption that some cxternal but clonespecific DNA was located within thc amocba casing, and was therefore inevitably transferredwith the amoebac to cach new subculture. This problcm was circumventcd by using isolated
133

nuclei of testatc amoebae. At present, diffcrcnt DNA-extraction protocols have been testedusing isolatcd nuclci a§ a rcfcrence.
Soil protozoology in Russia
Dr Julius Geltzer (Moscow State University) was invited for a lecture on "Soil Proto-zoology in Russia". Unfortunately, he could not attend, but he provided a written versionwhich is reported here, albeit much abbreviated. The development of soil protozoology inRussia is historically connectcd with agriculturc and the nccd for its intensification, the role ofprotozoa bcing considcrcd to bc regulators ofbacteria.
Some local faunistic and ecological invcstigations wcrc carried out in thc twenticsand thirties by, e. 9., Nowikoff, Losina-Losinsky, Martynov, Strelkov, and Belajeva. Thesestudies were critically reviewed and extended by Brodsky [5] in his monograph "Soil protozoaand their relative importance on soil activity". He postulated some general principles, such astrophic activity of soil protozoa in conditions of normal humidity, dcpcndcncc of protozoanabundancc on physical and chemical soil propcrtics, and influcncc of protozoa on nitrification.
ln thc sixtics, Nikolyuk and his studcnts dcmonstratcd thc stimulating cffcct of mcta-bolites from soil protozoa on bacteria and higher plants and the influence ofsuch processes onsoil biodynamics. Local faunistic studies proceeded during this time, e. 9., by Reinhard inUkrainia, Lepinis in the Baltic countries [14], Ibadov in Azerbaidjan, and Mordkovich in Sibe-ria. Thcse and thc formcr studics scrvcd as basis for important rcvicws on thc ccology andtaxonomy of soil protozoa [14, 76, 17].
Sincc thc sixtics, Gcltzcr and Korganova and their students invcntcd mcthods ofcombined cultivation of protozoa, bacteria and plants to observe the behaviour of protozoanpopulations in the rhizosphere and to reveal the protistocidic activity of soil fungi and actino-mycetes. Under their guidance several studies were pcrlormed, or arc in progress, relating soilprotozoology to gencral soil scicnce and cnvironmental problcms [12, 131.
Etna Acscht, Obcröstcrrcichischcs73, A - 4040 Linz (Austria).
John F. Darbyshire, The MacaulayJulius G. Gelt.zer, Moscow State
1 19899 Moscow (Russia)"Ralf Mcistcrfcld, RWTH Aachcn, Institut
Aachcn (Gcrrnany).
References
für Biologic II Kopcrnikusstrassc 1b, D-52056
Acscht E., and Foissner W. (1991): Bioindikation mit mikroskopisch kleincn Bodcnticrcn.VDI Ber. 901,985-1002.Aescht E., and Foissner W. (1992a): Enumerating soil testate amoebae by direct counting.In: Lee J. J., and Soldo A. T. (eds.): Protocols in protozoology, pp. 8-6.1 - 8-6.4. Societyof Protozoologists, AIlcn Prcss, Lawrence, Kansas.
3. Acscht 8., and Foissner W. (1992b): Enumcrating activc soil ciliatcs by dircct counting. ln:Lcc J. J., and Soldo A. T. (cds.): Protocols in protozoology, pp.B-7.1 - B-7.4. Socicty ofProtozoologists, Allen Press, Lawrence, Kansas.
4. Berthold 4., and Palzenberger M. (1 995): Comparison between direct counts of active soilciliates (Protozoa) and most probable number estimates obtained by Singh's dilution culturemcthod. Biol. Fertil. Soils /9, 348-356.
Land clcsmuscuffi, Biologiczcntrun!, J.-W. - Klcin - Strasss
Land Use Research [nstitute, AtreruJeen, Scotland AB9 zQJ.lJniversity, Department of Soil Science, Vorobjouy Hills,
1.
2.
134

5.
(1.
Brodsky A. L. (1935): Soil protozoa and thcir rclative importance on soil activity (Soilprotozoa of thc Ccntral Asia). Uzbckistan Comrnittcc Sci., Tashkcnt 20,99-182 (in Russianwith English summary).DarbyshireJ. F., ed., (1 994): Soil protozoa. CAB Int., Oxon, England. T Ekelund F., andRonn R. (1994): Notes on protozoa in agricultural soil with emphasis on heterotrophic fla-gellates and naked amoebae and their ecology. FEMS Microbiol. Rev. 15,331-353.Ekclund and ROnn R. (1994): Notcs on protozoa in agricultural soil with cmphasis on het-crotrophic flagcllatcs and nakcd amocbac and thcir ccology. FEMS Microbiol Rcv. /5,331-353.Foissner W. (1987): Soil protozoa: fundamental problems, ecological significance, adapta-tions in ciliates and testaceans, bioindicators, and guide to the literature. Progr. Protistol.2,69-212.Foissncr W. (.1995): Tropical protozoan divcrsity:80 ciliatc spccics (Protozoa, Ciliophora)in a soil samplc from a tropical dry fbrcst of Costa Rica, with dcscriptions of four ncw ge-ncra and scvcn ncw spccics. Arch. Protistcnk. 145,37-79.
10.Fromm H., Winter K., Filser J., Hantschcl R., and Beese F. (1 993): The influence of soiltype and cultivation system on the spatial distributions of the soil fauna and microorga-nisms and their interactions. Geoderma 60, 109-l 18.
ll.Gardini F., Antisari L.Y.,Cucrzoni M. 8., and Scqui P. (1991): A simple gas chromato-graphic approach to cvaluatc CO2 rclcasc, N2O cvolution, and 02 uptakc from soil. Biol.Fcrtil. Soils 12,1-4.
12.Geltzer J. G. (1991): Indices of biological activity in soil investigations. Soviet Soil Sci. 23,6-1 9 (translated by Scripta Technica, Inc. from Pochvovedeniye 23,47-60,1990).
l3.Geltzer J. G. (1 992): Free-living protozoa as a component of soil biota. Eurasian Soil Sci.24,'l-16 (translatcd by Scripta Technica, Inc. from Pochvovedcniyc 24,66-79,'1991).
14.Lcpinis A. K., GcltzcrJ. G., Chibisova O. 1., and Gcptncr V. A. (1973): Kcy to soilproto-zoaof thc European part of thc USSR. Mintis, Vilnius (in Russian).
15. Levrat P., Pussard M., and Alabouvctte C. (1992): Enhanced bacterial metabolism of aPseudomonas strain in response to the addition of culture filtrate of a bacteriophagousamoeba. Europ. J. Protistol. 28,79-84.
16. Nikolyuk V . F. (1972): Soil protozoa of thc USSR. Fan, Tashkent (in Russian).17. Nikolyuk V. F. (1980): Soil protozoa. Nauka, Moscow (in Russian).18.Schönborn W. (1992): Comparativc studics on thc production biology of protozoan com-
munities in freshwater and soil ecosystems. Arch. Protistenk. l4l ,187-214.19.Smith D. C., and Stribley D. P. (1994): A new approach to direct extraction of microorga-
nisms fiom soil. In: Ritz K., Dighton J., and Ciller K. E. (eds.) : Beyond the biomass, pp.49-55. British SoilScicncc Socicty, Wilcy Saycc, Chichcstcr.
20.Smith J. L., Halvorson J. J., and Bolton H. (199a): Spatial rclationships of soil microbialbiomass and C and N mincralization in a scmi-arid shrub-stcppc ccosystcm. Soil Biol. Bio-chem. 26, 1 15.1-1
.159.
21 .Wanner M., Eßer S., and Meisterf'eld R. (1994): Effects of light, temperature, fertilizersand pesticides on growth of the common freshwater and soil species Cyclopyxis kahli(Rhizopoda, Testaccalobosia), intcractions and adaptations. Limnologica 24,239-250.
22. Williams J. G. K., Kubclik A. R., Livak K. J., Rafalski J. A., and Tingcy S.V. (1990): DNApolymorphisms amplificd by arbitrary primers arc uscful as genctic markcrs. Nuclcic AcidsRes. 18, 6531-6535.
Key word
Bioindication - Cyclopyxis kahli- Etosha National Park -
Open cast mining - Soil proto-
zoan diversity
7.
8.
9.
135
]5
I
ü
{




















