
Proteomic analysis of the tetraspanin web using LC-ESI-MS/MS and MALDI-FTICR-MS
13
RESEARCH ARTICLE Proteomic analysis of the tetraspanin web using LC-ESI-MS/MS and MALDI-FTICR-MS Magali André 1 , Jean-Pierre Le Caer 2 , Céline Greco 1 , Sébastien Planchon 1 , Wassim El Nemer 3 , Claude Boucheix 1 , Eric Rubinstein 1 , Julia Chamot-Rooke 2 and Franc ¸ ois Le Naour 1 1 INSERM U602, Institut André Lwoff, Université Paris XI, Hôpital Paul Brousse, Villejuif Cedex, France 2 Laboratoire des Mécanismes Réactionnels, UMR CNRS 7651, Ecole Polytechnique, Palaiseau, France 3 INSERM U665, Institut National de la Transfusion Sanguine, Paris, France Tetraspanins are integral membrane proteins involved in a variety of physiological and patho- logical processes. In cancer, clinical and experimental studies have reported a link between tet- raspanin expression levels and metastasis. Tetraspanins play a role as organizers of a molecular network of interactions, the “tetraspanin web”. Here, we have performed a proteomic character- ization of the tetraspanin web using a model of human colon cancer consisting of two cell lines derived from primary tumor and metastasis from the same patient. The tetraspanin complexes were isolated after immunoaffinity purification and the proteins were identified by MS using LC- ESI-MS/MS and MALDI-FTICR. The high resolution and mass accuracy of FTICR MS allowed reliable identification using mass finger printing with only two peptides. Thus, it could be used to resolve the composition of complex peptide mixtures from membrane proteins. Different types of membrane proteins were identified, including adhesion molecules (integrins, Lu/B- CAM, GA733 proteins), receptors and signaling molecules (BAI2, PKC, G proteins), proteases (ADAM10, TADG15), and membrane fusion proteins (syntaxins) as well as poorly characterized proteins (CDCP1, HEM-1, CTL1, and CTL2). Some components were differentially detected in the tetraspanin web of the two cell lines. These differences may be relevant for tumor progres- sion and metastasis. Received: March 23, 2005 Revised: July 8, 2005 Accepted: July 25, 2005 Keywords: FTICR / LC-MS/MS / Membrane protein complexes / Metastasis / Tetraspanin Proteomics 2006, 6, 1437–1449 1437 1 Introduction Tetraspanins are integral membrane proteins characterized by the presence of four transmembrane domains. They ex- hibit significant sequence homology with conserved amino acids as well as specific structural features [1–4]. Tetra- spanins are highly expressed on many cell types and are involved in a large variety of physiological and pathological processes such as immune response, fertilization, infectious diseases (e.g., HCV, HIV, and malaria), and genetic diseases (e.g., X-linked mental retardation, retinal degeneration) [1, 3, Correspondence: Dr. Franc ¸ois Le Naour, INSERM U602, Hôpital Paul Brousse, 94807 Villejuif Cedex, France E-mail: [email protected] Fax: 133-1-45-59-53-29 Abbreviations: ADAM10, a disintegrin and metalloprotease 10; BAI2, brain angiogenesis inhibitor 2; CTL, choline transporter like; DHB, 2,5-dihydroxy benzoic acid; EGFR, epidermal growth factor receptor; EpCAM, epithelial cell adhesion molecule; Lu/B- CAM, Lutheran/B-cell adhesion molecule; TADG15, tumor asso- ciated differentially expressed gene 15; uPA, urokinase plasmino- gen activator DOI 10.1002/pmic.200500180 © 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Transcript of Proteomic analysis of the tetraspanin web using LC-ESI-MS/MS and MALDI-FTICR-MS

RESEARCH ARTICLE
Proteomic analysis of the tetraspanin web using
LC-ESI-MS/MS and MALDI-FTICR-MS
Magali André1, Jean-Pierre Le Caer2, Céline Greco1, Sébastien Planchon1,Wassim El Nemer3, Claude Boucheix1, Eric Rubinstein1, Julia Chamot-Rooke2
and Francois Le Naour1
1 INSERM U602, Institut André Lwoff, Université Paris XI, Hôpital Paul Brousse,Villejuif Cedex, France
2 Laboratoire des Mécanismes Réactionnels, UMR CNRS 7651, Ecole Polytechnique,Palaiseau, France
3 INSERM U665, Institut National de la Transfusion Sanguine, Paris, France
Tetraspanins are integral membrane proteins involved in a variety of physiological and patho-logical processes. In cancer, clinical and experimental studies have reported a link between tet-raspanin expression levels and metastasis. Tetraspanins play a role as organizers of a molecularnetwork of interactions, the “tetraspanin web”. Here, we have performed a proteomic character-ization of the tetraspanin web using a model of human colon cancer consisting of two cell linesderived from primary tumor and metastasis from the same patient. The tetraspanin complexeswere isolated after immunoaffinity purification and the proteins were identified by MS using LC-ESI-MS/MS and MALDI-FTICR. The high resolution and mass accuracy of FTICR MS allowedreliable identification using mass finger printing with only two peptides. Thus, it could be usedto resolve the composition of complex peptide mixtures from membrane proteins. Differenttypes of membrane proteins were identified, including adhesion molecules (integrins, Lu/B-CAM, GA733 proteins), receptors and signaling molecules (BAI2, PKC, G proteins), proteases(ADAM10, TADG15), and membrane fusion proteins (syntaxins) as well as poorly characterizedproteins (CDCP1, HEM-1, CTL1, and CTL2). Some components were differentially detected inthe tetraspanin web of the two cell lines. These differences may be relevant for tumor progres-sion and metastasis.
Received: March 23, 2005Revised: July 8, 2005
Accepted: July 25, 2005
Keywords:
FTICR / LC-MS/MS / Membrane protein complexes / Metastasis / Tetraspanin
Proteomics 2006, 6, 1437–1449 1437
1 Introduction
Tetraspanins are integral membrane proteins characterizedby the presence of four transmembrane domains. They ex-hibit significant sequence homology with conserved aminoacids as well as specific structural features [1–4]. Tetra-spanins are highly expressed on many cell types and areinvolved in a large variety of physiological and pathologicalprocesses such as immune response, fertilization, infectiousdiseases (e.g., HCV, HIV, and malaria), and genetic diseases(e.g., X-linked mental retardation, retinal degeneration) [1, 3,
Correspondence: Dr. Francois Le Naour, INSERM U602, HôpitalPaul Brousse, 94807 Villejuif Cedex, FranceE-mail: [email protected]: 133-1-45-59-53-29
Abbreviations: ADAM10, a disintegrin and metalloprotease 10;BAI2, brain angiogenesis inhibitor 2; CTL, choline transporterlike; DHB, 2,5-dihydroxy benzoic acid; EGFR, epidermal growthfactor receptor; EpCAM, epithelial cell adhesion molecule; Lu/B-
CAM, Lutheran/B-cell adhesion molecule; TADG15, tumor asso-ciated differentially expressed gene 15; uPA, urokinase plasmino-gen activator
DOI 10.1002/pmic.200500180
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

1438 M. André et al. Proteomics 2006, 6, 1437–1449
4]. In cancer, many clinical studies have reported that a highlevel of the tetraspanins CD9 and CD82/KAI-1 on tumorcells is associated with a favorable prognosis. Additionally, adecrease of expression level of these molecules is correlatedwith metastasis in many cancers. These observations havebeen corroborated using in vitro as well as in vivo experi-mental models [1, 5, 6]. By contrast, opposite effect wasobserved concerning the tetraspanins CD151 and Co-029 inclinical and/or experimental studies [7–12].
The function of tetraspanins is still not precisely knownat the molecular level. They may play a role in plasma mem-brane as organizers of multimolecular complexes that con-tain not only tetraspanins but also numerous transmem-brane cell surface proteins [1, 3, 4]. This network of interac-tions has been worded by the unifying concept of“tetraspanin web” [1, 13]. The down-regulation of some tet-raspanins during tumor progression may therefore pro-foundly modify the function of the tetraspanin web, thusfacilitating metastasis development. It is therefore of specialinterest to define the composition of the tetraspanin web incells originating from primary tumors and from metastasis.
In search for components of the tetraspanin web, we andothers have recently identified new proteins using MS.Among them, CD9P-1 [14, 15] and EWI-2 [16–20] belong to anovel family of proteins with Ig domains and of unknownfunction. Recently, other components of the tetraspanin webwere identified using proteomics such as surface immu-noglobulin in B cells [20] or GPCR and G proteins in thenervous system [21]. However, no extensive analysis of thetetraspanin web has been reported.
Proteomics is an emerging field allowing the globalcharacterization of subcellular compartments and proteincomplexes. However, the characterization of membrane pro-teins is still challenging. Indeed, most of them are notresolved using regular 2-D PAGE due to their low abundanceand high hydrophobicity [22–25]. Recently, strategies basedon LC-MS/MS were found to be well adapted for identifica-tion of membrane proteins. Indeed, LC-MS/MS allows theseparation of complex mixtures of peptides followed bysequence tag determination for each peptide analyzed (alsocalled shot gun), thus allowing identification of a proteinfrom only one peptide [25–29]. On the other hand, FTICRMS has emerged recently in proteomics [30]. The very highresolution and mass accuracy as well as the sensitivity of thispowerful device might open a new way for protein identifi-cation in complex peptide mixtures by mass finger printing.No study has yet addressed its potential for the identificationof transmembrane proteins.
In this report, we have implemented a proteomicapproach to address the composition of the tetraspanin webcombining immunoprecipitation experiments and SDS-PAGE with MS analysis using LC-MS/MS as well as MALDI-FTICR. Using human colon primary tumors and metastasisderived cells, we have identified novel proteins in the tetra-spanin web. Some of them were differentially expressed be-tween tumor and metastasis derived cells.
2 Materials and methods
2.1 Cell culture
The human colon carcinoma cell lines SW480, SW620, Caco-2, HT29, SW48, Colo205, and Lovo were obtained from theEuropean Collection of Cell Cultures (Salisbury, UK). Cellswere cultured in DMEM supplemented with 10% FCS, 2 mM
glutamine, and antibiotics (all from Invitrogen, Cergy-Pon-toise, France), and were maintained in a 377C humidifiedincubator in the presence of 5% CO2.
2.2 mAbs and flow-cytometric analysis
Anti-tetraspanin mAbs used in this study were ALB-6 (CD9)[31], TS9 (CD9), TS53 (CD53), TS81 (CD81), TS82 (CD82),and TS151 (CD151) [14]. Anti-integrins mAbs used were b1-vjf (integrin b1) [14], AK7 (integrin a2) (Diaclone, Besancon,France), M-KID-2 (integrin a3) [32], 4F10 (integrin a6) (Ser-otec, Oxford, UK), CD104 (integrin b4) (BD Biosciences, SanDiego, CA). Other mAbs were 1F11 (CD9P-1) [14], 8A12(EWI-2) [18], HEA125 (GA733-2/EpCAM) (Progen Bio-technik, Heidelberg, Germany), VIM15 (CDw92) (ResearchDiagnostics, Flanders, NJ), and 12A12 (CD55) [33]. MousemAb F241 (Lu/B-CAM) was produced by Institut National dela Transfusion Sanguine (INTS, Paris, France).
For flow-cytometric analysis, cells were detached using anon-enzymatic solution (Invitrogen), washed and stainedwith saturating concentrations of primary mAb. After wash-ing three times with media, cells were incubated with 10 mg/mL FITC-labeled goat antimouse antibody. After washing,cells were fixed with 1% formaldehyde in PBS. All incuba-tions were performed for 30 min at 47C. The analysis of cellsurface staining was performed using FACScalibur (BDBiosciences).
2.3 Cell surface labeling, immunoprecipitation and
Western-blot
Surface labeling of cells with EZ-link-Sulfo-NHS-LC-biotin(Pierce, Rockford, IL) was performed as previously described[14, 18]. Briefly, cells were washed three times in Hank’sbuffered saline and incubated 30 min in 10 mM HEPESpH 7.3, 150 mM NaCl, 0.2 mM CaCl2, 0.2 mM MgCl2 con-taining 0.5 mg/mL EZ-link-Sulfo-NHS-LC-biotin.
Cells were lysed directly in the flask in the lysis buffer(10 mM Tris pH 7.4, 150 mM NaCl, 1 mM CaCl2, 1 mM
MgCl2, 0.02% NaN3) containing 1% Brij97 (Sigma,St. Louis, MO) and proteases inhibitors. After 30 min at47C, insoluble material was removed by centrifugation at10 000g and cell lysate was precleared for 2 h by addition of1/1000 volume heat inactivated goat serum and 20 mL pro-tein G sepharose beads (Amersham-Pharmacia, Rainham,UK). Proteins were then immunoprecipitated by adding2 mg of specific antibody and 10 mL protein G-sepharose
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2006, 6, 1437–1449 Cell Biology 1439
beads to 200–400 mL lysate. After 2 h incubation at 47Cunder constant agitation, beads were washed five times inlysis buffer containing 1% Brij97. The immunoprecipitateswere separated by 5–15% SDS-PAGE under non-reducingconditions and transferred to a PVDF membrane (Amers-ham). Western-blotting was performed using specific bioti-nylated CD9 mAb. The membrane was incubated with astreptavidin-biotinylated horseradish peroxidase complex(SAv-PO) (Amersham), which was revealed by ECL (NEN,Boston, MA).
2.4 Immunoisolation of CD9-containing complexes
and in-gel tryptic digestion
For identification of CD9 associated molecules, 46108 cellswere lysed in 28 mL of lysis buffer containing 10 mM TrispH 7.4, 150 mM NaCl, 1 mM CaCl2, 1 mM MgCl2, 1% Brij97in the presence of protease inhibitors. Insoluble materialwas removed by centrifugation at 12 000g for 15 min, andthe lysates were precleared three times successively withsepharose 4B beads (Amersham, Saclay, France) coupled toBSA, to goat serum (Sigma), and then to an isotype-matched mAb. Isolation of CD9-containing complexes wasperformed using beads coupled to mAb ALB-6. The beadswere washed five times with lysis buffer and the proteinswere eluted using 1% Triton X-100 and then acetone pre-cipitated. The proteins were separated by 5–15% SDS-PAGE under nonreducing conditions. For profiling of thetetraspanin complexes, gels were silver stained as pre-viously described [18, 34]. For MS analysis, the gels werestained with colloidal CBB (BioRad, Hercules, CA). The gelpieces were excised and destained. They were incubated in100% ACN for 10 min, dried and incubated in 100 mM
ammonium bicarbonate containing 10 mM DTT for 30 minat 567C. After cooling to room temperature, the DTT solu-tion was replaced with 55 mM iodoacetamide in 100 mM
ammonium bicarbonate for 20 min at room temperaturein the dark. The gel pieces were washed in 100 mM
ammonium bicarbonate for 20 min, dehydrated in100% ACN, and dried. They were swollen in a digestionbuffer containing 25 mM ammonium bicarbonate and100 ng of trypsin (Roche, Meylan, France). Followingenzymatic digestion overnight at 377C, the resulting pep-tides were extracted with 50 mL of 5% formic acid for15 min at 377C followed by addition of 100 mL of100% ACN for another 15 min at 377C. The peptides werethen dried. They were rehydrated in 1% formic acid anddesalted using pipette tips C18 (Millipore, Bedford, MA)when analyzed using MALDI.
2.5 LC-ESI-MS/MS on IT and data analysis
LC-MS/MS analyses were performed using an ESI IT massspectrometer (LCQ Deca XP, ThermoElectron, San Jose, CA)coupled online with a capillary nanoHPLC (LC Packings)(Dionex, Amsterdam, ND) for LC. The capillary column used
in this study was a PepMap C18 RP (75 mm ID, 15 cm) (LCPackings). A linear 20 min gradient (flow rate 170 nL/min)from 5 to 50% ACN in 0.1% v/v aqueous formic acid wasperformed. All data were collected in centroid mode usingdata dependent acquisition mode. After the acquisition of afull MS scan (m/z 400–2000 Da) in the first scan event, thethree most intense ions present in above a threshold of105 counts were subsequently isolated for fragmentation(MS/MS scan). The collision energy for the MS/MS scanevents was preset at a value of 35%. The sequences of theMS/MS spectra were identified by correlation with the pep-tide sequences from human proteins present in the non-redundant protein sequence database (nr from NCBI) usingthe SEQUEST algorithm incorporated into the FinniganBIOWORKS 3.1 software. The SEQUEST search results wereinitially assessed by examination of the Xcorr (cross correla-tion) and the DCn (delta normalized correlation) scores. As ageneral rule, an Xcorr value of greater than 1.5, 2.0, and 2.5 for11, 21, and 31 charged peptides, respectively, and a DCngreater than 0.1 were accepted as a positive identification [35,36].
2.6 MALDI-FTICR MS and data analysis
FTICR mass spectrometer used was an APEX III (BrukerDaltonics, Bremen, Germany) equipped with a 7 T activelyshielded superconducting magnet and an infinity cell [37].The SCOUT MALDI source (Bruker Daltonics) with pulsedin-source collision gas and in-source accumulation cap-ability was used for this study. This source uses collision gas(argon) for cooling the ions, introduced during ion forma-tion and accumulation [38]. Singly protonated peptides werethus externally produced using a 337 nm nitrogen laser(20 Hz). Ions were stored in the source region and pulsedinto the detection cell through a series of electrostaticlenses. Ions were finally trapped in the cell using SideKick™
and front and back trapping voltages of 0.9 and 0.95 V. Massspectra were acquired from m/z 500 to 4000 with 512 k datapoints and monoisotopic peaks were automatically labeledusing the software XMASS 6.1.4 (Bruker Daltonics). Foreach measurement, 20–30 scans (200–300 laser shots) werecoadded. 2,5-Dihydroxy benzoic acid (DHB) (Sigma-Aldrich)was used as the matrix. A 10 mg/mL solution of DHB inwater, ACN (70:30 v/v), with the addition of 0.2% TFA wasprepared. For each gel band, 0.5 mL of the desalted peptidesin (H2O, formic acid 1%) was vortexed with 0.5 mL of theDHB solution. 0.5 mL of the resultant mixture was loadedon the MALDI target for each spot. Anchorchips™ wereused for all experiments. External calibration was per-formed using the peptide calibration standard (Bruker)containing Angiotensin I and II, Substance P, Bombesin,ACTH clip 1–17, ACTH clip 18–39, and Somatostatin 28.Protein identification was performed by mass finger print-ing using the program Profound (http://prowl.rocke-feller.edu/profound_bin/WebProFound.exe) in the NCBIdatabase.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

1440 M. André et al. Proteomics 2006, 6, 1437–1449
3 Results and discussion
3.1 Proteomic strategy for analysis of the tetraspanin
web
To address the composition of the tetraspanin web, a prote-omic approach has been implemented (Fig. 1). The cellswere lysed using the mild detergent Brij97. We have pre-viously shown that in these conditions, tetraspanin–tetra-spanin interactions are preserved and tetraspanin complexescan be immunoprecipitated by mAbs directed against anytetraspanin [13]. After immunoprecipitation experiments,
the associated proteins were eluted using the more stringentdetergent Triton X-100, which dissociates tetraspanin–tetra-spanin associations [13, 39] (Fig. 1A). We have used theSW480 and SW620 cell lines as a biological model. Indeed,these cells have been established, respectively, from a pri-mary colon cancer and a derived lymph node metastasisfrom the same patient [40]. The target used in this study wasthe tetraspanin CD9, because it is highly expressed on thesecells. The CD9-associated proteins were separated usingSDS-PAGE and then the proteins were either transferred to aPVDF membrane and revealed by chemiluminescence whencell surface proteins were biotinylated before lysis, or stained
Figure 1. Proteomic strategy foranalysis of the tetraspanin com-plexes. (A) Tetraspanin com-plexes (schematically repre-sented in the dotted box) weresolubilized using the mild deter-gent Brij97 and isolated byimmunoprecipitation experi-ments using specific CD9 mAbswhich were covalently coupledto sepharose beads. Associatedproteins from the complexeswere eluted using the morestringent detergent Triton X-100. (B) Proteins were separatedby SDS-PAGE. Then, the pro-teins were transferred to a PVDFmembrane and revealed bystreptavidine peroxidase andchemiluminescence (when cellsurface proteins were biotiny-lated before lysis) (left panel), orstained with silver or colloidalCBB. Each lane was system-atically cut in 13 gel slices (rightpanel). Proteins were in-geltrypsin digested and the result-ing peptides were analyzedusing MS. (C) For LC-MS/MSanalysis, the peptides wereseparated by nanoHPLC allow-ing to measure total ion counton a chromatogram (upper pic-ture). For each retention time(e.g., 16.75 min, dotted straightline), a mass spectrum isrecorded (middle picture), inwhich a parent ion can be selec-ted (e.g., m/z = 641.4, blackarrow). Fragmentation of parentions led to MS/MS spectrumacquisition (lower picture). Thisspectrum contains sequenceinformation of the parent ionthat can be deduced. (D) MALDI-FTICR-MS analysis gave massspectrum exhibiting MH1 ionswith very high mass accuracy.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2006, 6, 1437–1449 Cell Biology 1441
with silver or CBB to visualize the pattern of tetraspanin-associated molecules. As shown in Fig. 1B, significant differ-ences of the tetraspanin complexes were observed betweenprimary tumor and metastasis derived cells. For MS analysis,each lane was systematically cut and the proteins were in-geldigested. The resulting peptides were analyzed using LC-MS/MS (Fig. 1C) and/or MALDI-FTICR-MS (Fig. 1D). To elim-inate background proteins from analysis, the IgG1-coatedbeads that were used in the last preclearing step were treatedidentically as CD9 beads. Any protein found in both sampleswas not considered as a tetraspanin-associated protein.
3.2 Proteomic analysis using LC-ESI-MS/MS and
MALDI-FTICR-MS
The use of MALDI-FTICR for identification of membraneproteins by mass finger printing depends on its ability toaccurately detect peptides from protein mixtures as found ina SDS-PAGE band.
The band 1, corresponding to CD9 antigen (Fig. 1B), wasused to determine the minimal number of peptides neces-sary for identification using MALDI-FTICR. Tetraspaninshave regularly been found difficult to identify by MS, possi-bly due to the fact that a major part of their sequence istransmembranous. Indeed, only two peptides correspondingto CD9 were characterized using LC-MS/MS analysis on IT(Fig. 2A and Table 1). The same peptides were observedusing MALDI-FTICR-MS, with a high intensity (Fig. 2B).The database search with those two masses allowed theidentification of CD9 using the program Profound (Table 1).This suggested that FTICR MS is accurate enough to allowprotein identification based on mass finger printing withonly two peptides.
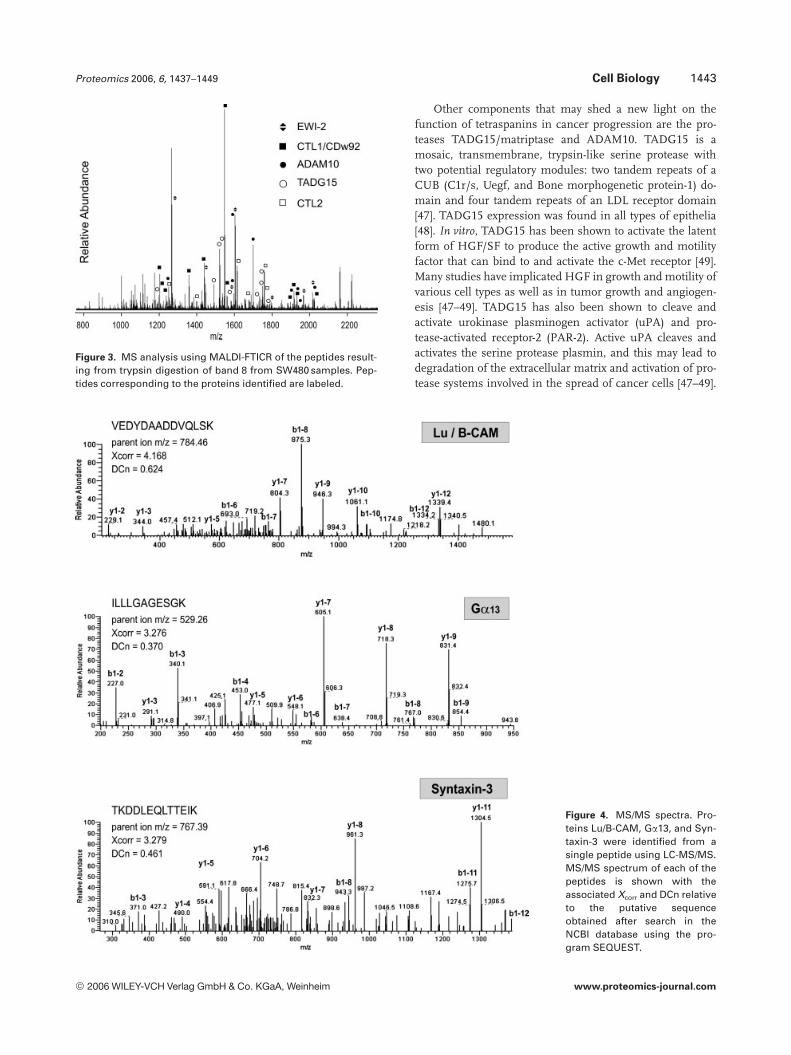
The band 8 from SW480 samples was used to demon-strate that MALDI-FTICR can resolve a mixture containingpeptides from different proteins by mass finger printing. LC-MS/MS analysis of this band yielded four different proteinsincluding EWI-2, CTL1/CDw92, ADAM10, and CTL2.MALDI-FTICR mass spectrum led to observe numerouspeptides and the database search with 5 ppm mass toleranceallowed to identify the previous proteins as well as TADG15.Each of them was identified with at least six matching pep-tides (Fig. 3 and Table 2).
A global comparison of the use of LC-MS/MS andMALDI-FTICR for the analysis of tetraspanin complexesisolated from SW480 cells (Table 3) indicated that among24 proteins, only four (Lu/B-CAM, CD82, Ga13, and Syn-taxin-3) were identified by LC-ESI-MS/MS. Their identifica-tion was based on a single peptide. The MS/MS spectra arereported in Fig. 4. This emphasizes the major advantage ofthe LC-MS/MS strategy, which is very powerful for proteinidentification from only one peptide. On the other hand,seven proteins (BAI2, HEM-1, TADG15, GA733-1, Gia2,Gia3, and Gb2) were identified only using MALDI-FTICR-MS. The MALDI-FTICR-MS analysis had several advantages.First, it does not require any step of separation of the pep-tides such as LC, thus avoiding time-consuming manipula-tion and loss of material (e.g., highly hydrophobic or coelutedpeptides). Second, the mass finger printing from complexmixture using MALDI-FTICR-MS does not require MS/MS,thus avoiding loss of information from poorly fragmentedpeptides resulting in MS/MS spectra that are not inter-pretable. This is all the more crucial since only few peptidesare obtained from some membrane proteins (such as CD9).Third, we observed better sensitivity of the MALDI-FTICRanalysis with the same samples thus leading to higher num-ber of detected peptides. Therefore, the proteomic analysis ofthe tetraspanin complexes from SW620 cells was performedusing only MALDI-FTICR-MS.
3.3 Novel proteins identified in the tetraspanin web
of human colon carcinoma cells
This proteomic analysis led to identify 27 different proteins(Table 3). Among them, three were tetraspanins (CD9,CD81, and CD82) and nine have previously been reported tobe associated with at least one tetraspanin, including themajor tetraspanin partners such as b1 integrins, CD9P-1,and EWI-2 [1, 14–20].
This study shows that tetraspanins associate with a so farunsuspected variety of membrane proteins including severalcategories of adhesion molecules, membrane proteases,receptor, and signaling molecules as well as proteinsinvolved in membrane fusion processes or with stillunknown function.
Table 1. Assignment of peptide masses from band 1 to CD9
LC-MS/MS MALDI-FTICR Residues Peptide sequence
Measuredmass
Chargestate
Xcorr DCn Measuredmass
Theoreticalmass
Delta,ppm
590.29 2 3.36 0.26 1179.6642 1179.6620 ,2 170–179 KDVLETFTVK782.48 2 3.31 0.45 1563.7352 1563.7326 ,2 120–131 EVQEFYKDTYNK
Analysis by LC-ESI-MS/MS on IT followed by search in the NCBI database using SEQUEST program and MALDI-FTICR-MS of the peptidemasses (MH1) followed by search in the NCBI database using the program Profound.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

1442 M. André et al. Proteomics 2006, 6, 1437–1449
Figure 2. MS analysis of thepeptides resulting from thetrypsin digestion of CD9 antigen(band 1). Peptides were ana-lyzed using LC-MS/MS (A) orMALDI-FTICR (B).
We observed the presence of the GA733 family mem-brane antigens GA733-1 and GA733-2. These proteins ex-hibit 50% identity with each other and some characteristicfeatures including an extracellular domain with 12 conservedcystein residues, a single transmembrane domain, a shortcytoplasm anchor region, and homology with the cystein-rich thyroglobulin [41]. The GA733-2, also called EpCAM, isa homophilic Ca21-independent cell–cell adhesion moleculethat is expressed in many human epithelial tissues [42].
EpCAM is abundantly expressed in colorectal carcinoma.This antigen can spontaneously induce a humoral and cel-lular antitumor immunity and constitutes therefore a com-mon target for immunotherapy [43]. We also observed thepresence of Lu/B-CAM. This protein belongs to the Ig superfamily and exhibits five Ig-like domains. It has been shownthat Lu/B-CAM is a laminin receptor with a restrictive speci-ficity to laminin a5 chain (a component of LN10 and LN11)[44–46].
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Proteomics 2006, 6, 1437–1449 Cell Biology 1443
Figure 3. MS analysis using MALDI-FTICR of the peptides result-ing from trypsin digestion of band 8 from SW480 samples. Pep-tides corresponding to the proteins identified are labeled.
Other components that may shed a new light on thefunction of tetraspanins in cancer progression are the pro-teases TADG15/matriptase and ADAM10. TADG15 is amosaic, transmembrane, trypsin-like serine protease withtwo potential regulatory modules: two tandem repeats of aCUB (C1r/s, Uegf, and Bone morphogenetic protein-1) do-main and four tandem repeats of an LDL receptor domain[47]. TADG15 expression was found in all types of epithelia[48]. In vitro, TADG15 has been shown to activate the latentform of HGF/SF to produce the active growth and motilityfactor that can bind to and activate the c-Met receptor [49].Many studies have implicated HGF in growth and motility ofvarious cell types as well as in tumor growth and angiogen-esis [47–49]. TADG15 has also been shown to cleave andactivate urokinase plasminogen activator (uPA) and pro-tease-activated receptor-2 (PAR-2). Active uPA cleaves andactivates the serine protease plasmin, and this may lead todegradation of the extracellular matrix and activation of pro-tease systems involved in the spread of cancer cells [47–49].
Figure 4. MS/MS spectra. Pro-teins Lu/B-CAM, Ga13, and Syn-taxin-3 were identified from asingle peptide using LC-MS/MS.MS/MS spectrum of each of thepeptides is shown with theassociated Xcorr and DCn relativeto the putative sequenceobtained after search in theNCBI database using the pro-gram SEQUEST.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

1444 M. André et al. Proteomics 2006, 6, 1437–1449
Table 2. Assignment of peptide masses from band 8
Measuredmass
TheoreticalMass
Delta, ppm Residues Peptide sequence Identifiedprotein
1271.69905 1271.6994 20.27 29–39 EVLVPEGPLYR EWI-21448.70126 1448.7016 20.23 224–237 SDLAVEAGAPYAER1615.9166 1615.9166 0 148–163 VLPDVLQVSAAPPGPR1800.8731 1800.8731 0 174–189 M*TVHEGQELALGCLAR1981.92853 1981.9337 22.61 338–357 HAAYSVGWEMAPAGAPGPGR1997.93182 1997.9287 1.56 338–357 HAAYSVGWEM*APAGAPGPGR2018.0189 2018.0189 0 358–377 LVAQLDTEGVGSLGPGYEGR
1203.5957 1203.59729 21.32 109–119 SVALCVAACPR CTL1/CDw921223.60676 1223.60889 21.74 621–630 VLMEFVENSR1239.60402 1239.6038 1.77 621–630 VLMEFVENSR1359.76414 1359.76306 0.79 517–528 DAFVILVENALR1441.81976 1441.81616 2.50 422–434 NLPFTPILASVNR1548.87101 1548.86853 1.60 162–175 LPVPASAPIPFFHR1568.74605 1568.74854 21.59 76–89 LEAIPNSGMDHTQR1584.74168 1584.7435 1.15 76–89 LEAIPNSGM*DHTQR1898.94417 1898.9469 21.44 91–105 YVFFLDPCNLDLINR1918.9988 1919.0022 21.77 187–203 FAEALITFVSDNSVLHR1927.93269 1927.9291 1.86 73–89 NTKLEAIPNSGM*DHTQR2027.03723 2027.04187 22.29 90–105 KYVFFLDPCNLDLINR2027.03723 2027.04187 22.29 91–106 YVFFLDPCNLDLINRK
1257.79404 1257.79297 0.85 710–721 LPPPKPLPGTLK ADAM101597.60072 1597.60059 0.08 481–493 DECCFDANQPEGR1605.77214 1605.76892 2.01 623–636 TITLQPGSPCNDFR1700.84686 1700.849 21.26 252–266 AIDTIYQTTDFSGIR1913.84255 1913.8480 22.85 544–560 KM*DPSTCASTGSVQWSR1927.93269 1927.93713 22.30 220–234 NTCQLYIQTDHLFFK1961.85899 1961.86267 21.88 558–574 HTQVCINGQCAGSICEK2025.07542 2025.07642 20.49 357–375 SLNTGIITVQNYGSHVPPK
1199.49739 1199.49976 21.98 262–271 SFDLASCDER TADG151492.65963 1492.65979 20.11 209–221 TQDNSCSFGLHAR1546.78265 1546.78015 1.62 324–336 HPGFEATFFQLPR1583.83186 1583.83154 0.20 190–204 SFVVTSVVAFPTDSK1731.76254 1731.76428 21.00 400–413 DYVEINGEKYCGER1750.82894 1750.83301 22.32 818–833 IFQAGVVSWGDGCAQR1760.83664 1760.83911 21.40 228–243 FTTPGFPDSPYPAHAR1795.90481 1795.90869 22.16 384–399 FFYLLEPGVPAGTCPK
1262.64368 1262.6463 22.07 171–180 RCFPAIHAYK CTL21397.64116 1397.6406 0.40 395–406 IFDDSPCPFTAK1625.68265 1625.686 22.06 407–420 TCNPETFPSSNESR1678.70165 1678.7067 23.00 427–440 CQFAFYGGESGYHR1777.92368 1777.9272 1.98 481–496 KPDDLPAFPLFSAFGR1788.93068 1788.9319 20.68 81–94 NENKPYLFYFNIVK
Analysis by MALDI-FTICR-MS of the peptide masses followed by search in the NCBI database using the programProfound with 5 ppm mass tolerance. Matching of peptides masses labeled with (*) was compatible with 1 Met-oxmodification. All masses are given for MH1 ions.
Therefore, the protease TADG15 may play a role in growthand invasion of cancer cells during tumor progression.Interestingly, a recent report has suggested that the expres-sion of tetraspanin CD82 is correlated with inhibition ofplasminogen activation by redistribution of uPA receptor(uPAR) with a resultant loss of uPA binding [50].
ADAM proteins are membrane-anchored metallopro-teases that process and shed ectodomains of membrane-anchored growth factors, cytokines, and receptors. ADAMsalso have essential roles in cell–cell and cell–matrix interac-tions and therefore in a variety of physiological and patho-logical processes including angiogenesis and cancer [51–53].
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2006, 6, 1437–1449 Cell Biology 1445
Table 3. Identified proteins in the tetraspanin complexes from SW480 and SW620 cells
Protein name gi no. SW480LC-MS/MS
SW480MALDI-FTICR
SW620MALDI-FTICR
nb ofpeptides
aa(% max)
MW(obs.)
MW(theo.)
BAI2 4502357 2 1 1 2, 5, 12 10 160 175Integrin a6 33942 2 2 1 2, 2, 5 6 130 120Integrin a3 220141 1 1 1 3, 3, 4 6 130 115Integrin b4 7459693 2 2 1 2, 2, 9 11 120 110Integrin b1 19743819 1 1 1 1, 7, 11 17 120 92CD9P-1 22760207 1 1 1 3, 6, 21 26 120 97CDCP1 14328879 1 1 1 1, 5, 8 10 90 94HEM-1 2506502 2 1 1 2, 3, 4 4 80 130PKC-b 20141488 2 2 1 2, 2, 2 4 80 78Lu/B-CAM 1708887 1 2 2 1, 2, 2 2 80 67CTL2 31377727 1 1 2 2, 6, 2 11 70 82CTL1/CDw92 18034692 1 1 1 2, 11, 14 22 70 75ADAM10 1616601 1 1 1 1, 8, 14 23 70 80TADG15 10257390 2 1 1 2, 8, 3 13 70 97EWI-2 16445029 1 1 1 2, 6, 5 14 70 66CD82 54696390 1 2 2 1, 2, 2 4.5 60 25GA733-1 4505057 2 1 2 2, 2, 2 10 50 37GA733-2 4505059 1 1 2 1, 2, 2 7 40 35Ga13 2494886 1 2 2 1, 2, 2 3 40 44Gaq 2286213 1 1 2 1, 3, 2 12 40 42Gia3 5729850 2 1 2 2, 6, 2 20 35 41Gia2 4218034 2 1 2 2, 5, 2 22 35 38Gb2 20357529 2 1 2 2, 4, 2 16 35 38Syntaxin-4A 7447076 1 1 2 1, 4, 2 14 35 34Syntaxin-3 6175045 1 2 2 1, 2, 2 4.5 35 33CD9 4502693 1 1 1 2, 2, 2 10 27 25CD81 4557944 1 1 1 1, 2, 2 17 26 26
Tetraspanin complexes were analyzed by LC-ESI-MS/MS on IT and/or MALDI-FTICR-MS. Resulting spectra wereused for searching in the NCBI database using, respectively, SEQUEST program (MS/MS spectra) or the programProfound for mass finger printing with 5 ppm mass tolerance. 1: identified; 2: not detected. Newly identifiedproteins are in bold.
Here, we show that ADAM10 is a component of the tetra-spanin web in colon carcinoma cells. It has been reportedthat ADAM10 plays a role in a crosstalk between GPCR andepidermal growth factor receptor (EGFR) [54]. Indeed, GPCRactivates ADAM10, which in turn, releases the EGFR ligandheparin-binding EGF (HB-EGF) to activate EGFR. GPCRsplay a role as receptor of bioactive peptides or growth factorsand they are involved in many cellular mechanisms aftersignal transduction via coupled G proteins. It is interestingto note that GPCRs were also identified in the tetraspaninweb. We have identified the GPCR BAI2 and several G pro-teins such as Gaq, Ga13, Gia2, Gia3, Gb2 in colon carci-noma cells, and the presence of the GPCR GPR56, and ofGaq and Gb in the nervous system has been recently report-ed [21].
Interestingly, the proteins syntaxin-3 and syntaxin-4Awere identified. Syntaxins are transmembrane proteinsexhibiting a carboxy-terminal tail-anchor and multiplecoiled-coil domains. They mediate vesicle fusion in diverse
vesicular transport processes along the exocytic and theendocytic pathways [55]. It should be noted that syntaxin-3and -4A are known to be expressed at the plasma membraneof colon carcinoma cells. This is the first evidence of theirpresence in the tetraspanin web. This observation is inter-esting in relation to the involvement of tetraspanins – inparticular CD9 – in membrane fusion events [1, 3, 56, 57].
Finally, some very poorly characterized proteins wereidentified such as CDCP1, HEM-1, CTL1/CDw92, and CTL2.The protein CDCP1 (CUB-domain-containing protein) is anovel transmembrane molecule containing three CUBdomains in the extracellular part most likely involved in celladhesion or interacting with the extracellular matrix [58, 59].This molecule has been reported to be highly expressed inlung, breast, and colon tumors [58, 59]. However, little isknown about the biological function of CDCP1. The HEM-1molecule belongs to the HEM family membrane proteins,which is conserved throughout evolution. These proteinsharbor several conserved membrane spanning domains [60].
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

1446 M. André et al. Proteomics 2006, 6, 1437–1449
Figure 5. Expression of components of the tetraspanin web onSW480 and SW620. (A) Expression levels at the cell surface werechecked by flow cytometry using specific mAbs directed againsttetraspanins CD9, CD81, CD82, and CD151, integrins b1, a3, anda6, the Ig proteins CD9P-1 and EWI-2, as well as EpCAM, CDw92,and integrin b4. (B) SW480 or SW620 cells were lysed using themild detergent Brij97 and immunoprecipitation experiments (IP)were performed using specific mAbs directed against CD9, CD9P-1, EpCAM, CDw92, integrin b4, and integrin a2 (negative control).Proteins were separated by SDS-PAGE and transferred to a PVDFmembrane. Association of these proteins with CD9 antigen wasrevealed by Western-blotting using biotin labeled CD9 mAbs.
The CTL1 and CTL2 molecules exhibit 43% sequencehomology and belong to the CTL family containing fourmembers. These are membrane proteins with ten trans-membrane domains. High levels of CTL1 expression wereobserved in the rat colon [61].
3.4 Comparative analysis of colon tumor and
metastasis derived cells
The proteomic analysis of the CD9 associated proteins inSW480 and SW620 cells suggested significant variation interms of the composition of the complexes (Table 3). Indeed,some components were differentially detected such asintegrins a6 and b4, PKC-b, B-CAM, CTL2, CD82,GA733 proteins, G proteins, and syntaxins. Other compo-nents were identified in both cells. However, MS is not
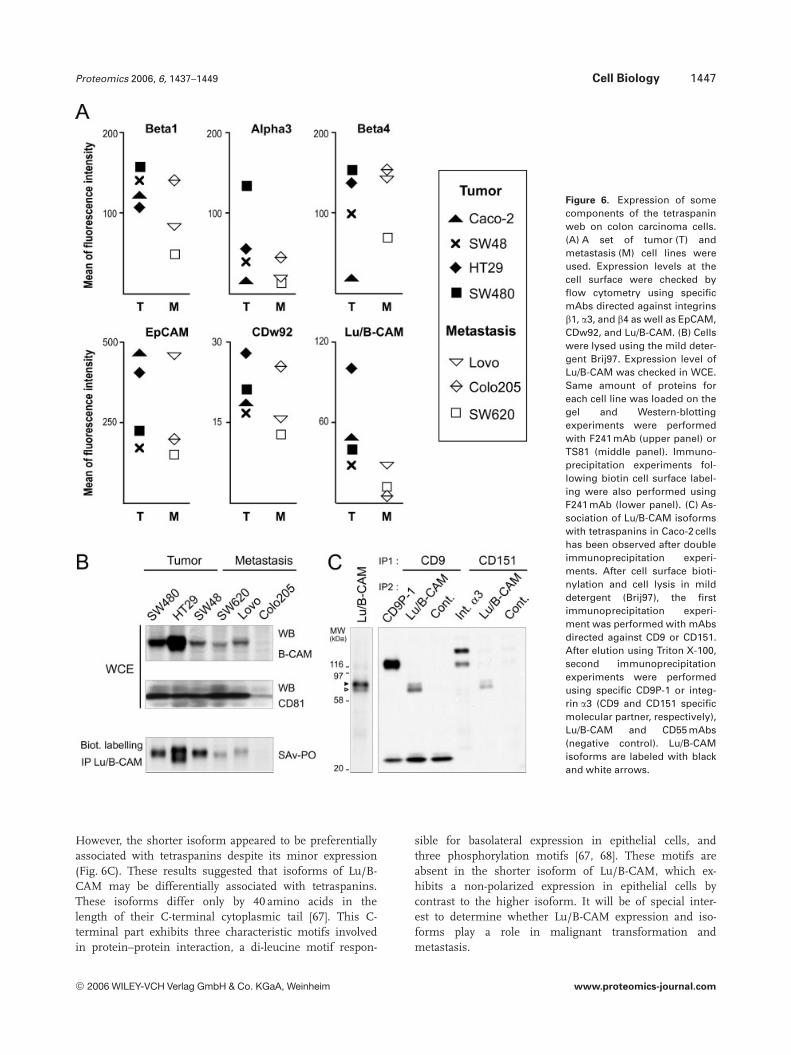
intrinsically a quantitative method. Thus, we sought toinvestigate the expression levels of some components of thetetraspanin web at the cell surface of SW480 and SW620 cells(when antibodies were available). We checked the expressionof the tetraspanins CD9, CD81, CD82, and CD151, theintegrins a3, a6, b1, and b4, the IgSF proteins CD9P-1 andEWI-2, as well as the newly identified components EpCAMand CTL1/CDw92 (Fig. 5A). We observed significant lowerexpression levels of all tested components of the tetraspanincomplexes on SW620 cells as compared to SW480 cells. Thepresence in the tetraspanin web of EpCAM, CTL1/CDw92,and integrin b4 was confirmed by immunoprecipitationexperiments using mAbs directed against these proteins fol-lowed by Western-blotting experiments using a CD9 mAb(Fig. 5B). Furthermore, the association of integrin b4 withCD9 was induced in SW620 cells, despite a decrease of theexpression at the cell surface. These results suggested a spe-cific recruitment of the integrin b4 in the tetraspanin com-plexes of SW620 cells. Integrin a6b4 is a laminin receptorthat participates in hemidesmosomes formation [62]. It hasbeen reported that integrin a6b4 is involved in proinvasivesignals with receptor tyrosine kinase (such as c-Met) andpromotes cell migration and invasion [63]. Integrin a6b4expression increases proliferation of epithelial cells and con-fers resistance to apoptosis on carcinoma cells [63]. Thepresence of the integrin a6b4 has previously been reportedin the tetraspanin complexes [64, 65] and a functional rele-vance has been suggested in terms of adhesion and migra-tion [66].
We further checked the expression levels of integrins b1,a3, and b4 as well as EpCAM, CTL1/CDw92, and Lu/B-CAMat the cell surface of a set of colon carcinoma cells includingprimary tumor (Caco-2, HT29, and SW48) and metastasis(Lovo and Colo205) derived cells (Fig. 6A). We observed sig-nificant lower expression levels of Lu/B-CAM on metastasiscell lines as compared with primary tumor derived cells. Wefurther checked the expression level of Lu/B-CAM usingWestern-blotting experiments in whole cell extracts (WCEs)from each cell line as well as after cell surface biotin labellingfollowed by Lu/B-CAM immunoprecipitation experiments(Fig. 6B). Again the level of Lu/B-CAM was lower on the celllines originated from metastasis as compared with tumorcells. In addition to a variation of Lu/B-CAM expressionlevel, we observed the presence of isoforms. Indeed, twobands were revealed in HT29 and Caco-2 cells (Fig. 6B, C). Ithas been previously reported that Lu/B-CAM is expressed astwo membrane glycoproteins (85 and 78 kDa) resulting fromalternative splicing of a primary transcript [67]. Thus, wesought to investigate the association of both isoforms withtetraspanin. Additional immunoprecipitation experimentswere performed in Caco-2 cells using mild detergent Brij97and mAbs directed against tetraspanins CD9 or CD151. Afterelution of the complexes with Triton X-100, a second immu-noprecipitation experiment was performed with F241 mAb.We observed the presence of the two Lu/B-CAM isoformssuggesting that both are components of the tetraspanin web.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Proteomics 2006, 6, 1437–1449 Cell Biology 1447
Figure 6. Expression of somecomponents of the tetraspaninweb on colon carcinoma cells.(A) A set of tumor (T) andmetastasis (M) cell lines wereused. Expression levels at thecell surface were checked byflow cytometry using specificmAbs directed against integrinsb1, a3, and b4 as well as EpCAM,CDw92, and Lu/B-CAM. (B) Cellswere lysed using the mild deter-gent Brij97. Expression level ofLu/B-CAM was checked in WCE.Same amount of proteins foreach cell line was loaded on thegel and Western-blottingexperiments were performedwith F241 mAb (upper panel) orTS81 (middle panel). Immuno-precipitation experiments fol-lowing biotin cell surface label-ing were also performed usingF241 mAb (lower panel). (C) As-sociation of Lu/B-CAM isoformswith tetraspanins in Caco-2 cellshas been observed after doubleimmunoprecipitation experi-ments. After cell surface bioti-nylation and cell lysis in milddetergent (Brij97), the firstimmunoprecipitation experi-ment was performed with mAbsdirected against CD9 or CD151.After elution using Triton X-100,second immunoprecipitationexperiments were performedusing specific CD9P-1 or integ-rin a3 (CD9 and CD151 specificmolecular partner, respectively),Lu/B-CAM and CD55 mAbs(negative control). Lu/B-CAMisoforms are labeled with blackand white arrows.
However, the shorter isoform appeared to be preferentiallyassociated with tetraspanins despite its minor expression(Fig. 6C). These results suggested that isoforms of Lu/B-CAM may be differentially associated with tetraspanins.These isoforms differ only by 40 amino acids in thelength of their C-terminal cytoplasmic tail [67]. This C-terminal part exhibits three characteristic motifs involvedin protein–protein interaction, a di-leucine motif respon-
sible for basolateral expression in epithelial cells, andthree phosphorylation motifs [67, 68]. These motifs areabsent in the shorter isoform of Lu/B-CAM, which ex-hibits a non-polarized expression in epithelial cells bycontrast to the higher isoform. It will be of special inter-est to determine whether Lu/B-CAM expression and iso-forms play a role in malignant transformation andmetastasis.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

1448 M. André et al. Proteomics 2006, 6, 1437–1449
4 Concluding remarks
We have investigated the composition of the tetraspanin webin human colon carcinoma cells using proteomics. In thisstudy, “shot gun” and mass finger printing strategies wereemployed for protein identification, using two different typesof mass spectrometry, LC-ESI-MS/MS and MALDI-FTICR.This is the first study using MALDI-FTICR for analysis ofcomplex mixtures of membrane proteins from mammaliancells. We have demonstrated that MALDI-FTICR-MS led toreliable protein identification by mass finger printing with afew peptides. Although LC-MS/MS allows protein identifica-tion from a single peptide, MALDI-FTICR exhibits numer-ous advantages such as versatility, rapidity, sensitivity, highresolution, and mass accuracy. It is likely that a hybrid devicecombining advantages of FTICR MS with MS/MS informa-tion obtained from IT would constitute a significant progressin proteomics.
This study is the first extensive analysis of the composi-tion of the tetraspanin web. The proteomic analysis of thetetraspanin web showed that tetraspanins associate with avariety of membrane proteins including adhesion molecules,membrane proteases, receptor, and signaling molecules aswell as proteins involved in fusion processes or with stillunknown function. Furthermore, this study highlights var-iations of the tetraspanin web between primary colon tumorand metastasis cell lines. The functional relevance of the as-sociation in the plasma membrane of the various compo-nents of the tetraspanin web remains to be addressed in fur-ther studies. This may give new insights on the role of tetra-spanins in the process of cancer progression and metastasis.
We are grateful to Zohar Mishal for assistance in MS and toPierre Eid for helpful discussions. This work was supported by anAction Concertée Incitative (ACI, 035109) of Ministère de laRecherche, Gefluc, Institut du Cancer et d’Immunogénétique(ICIG), Association pour la Recherche contre le Cancer (ARC,4735), and Nouvelles Recherches Biomédicales-Vaincre le Cancer(NRB). M.A. was recipient of a grant from NRB.
5 References
[1] Boucheix, C., Rubinstein, E., Cell. Mol. Life Sci. 2001, 58,1189–1205.
[2] Stipp, C. S., Kolesnikova, T. V., Hemler, M. E., Trends Bio-chem. Sci. 2003, 28, 106–112.
[3] Hemler, M. E., Annu. Rev. Cell. Dev. Biol. 2003, 19, 397–422.
[4] Levy, S., Shoham, T., Nat. Rev. Immunol. 2005, 5, 136–148.
[5] Dong, J. T., Lamb, P. W., Rinker-Schaeffer, C. W., Vukanovic, J.et al., Science 1995, 268, 884–886.
[6] Boucheix, C., Duc, G. H., Jasmin, C., Rubinstein, E., ExpertRev. Mol. Med. 2001, 1–17.
[7] Testa, J. E., Brooks, P. C., Lin, J. M., Quigley, J. P., Cancer Res.1999, 59, 3812–3820.
[8] Kohno, M., Hasegawa, H., Miyake, M., Yamamoto, T., Fujita,S., Int. J. Cancer 2002, 97, 336–343.
[9] Tokuhara, T., Hasegawa, H., Hattori, N., Ishida, H. et al., Clin.Cancer Res. 2001, 7, 4109–4114.
[10] Hashida, H., Takabayashi, A., Tokuhara, T., Hattori, N. et al.,Br. J. Cancer 2003, 89, 158–167.
[11] Ang, J., Lijovic, M., Ashman, L. K., Kan, K., Frauman, A. G.,Cancer Epidemiol. Biomarkers 2004, 13, 1717–1721.
[12] Kanetaka, K., Sakamoto, M., Yamamoto, Y., Takamura, M. etal., J. Gastroenterol. Hepatol. 2003, 18, 1309–1314.
[13] Rubinstein, E., Le Naour, F., Lagaudrière-Gesbert, C., Billard,M. et al., Eur. J. Immunol. 1996, 26, 2657–2665.
[14] Charrin, S., Le Naour, F., Oualid, M., Billard, M. et al., J. Biol.Chem. 2001, 276, 14329–14337.
[15] Stipp, C. S., Orlicky, D., Hemler, M. E., J. Biol. Chem. 2001,276(7) 4853–4862.
[16] Stipp, C. S., Kolesnikova, T. V., Hemler, M. E., J. Biol. Chem.2001, 276, 40545–40554.
[17] Clark, K. L., Zeng, Z., Langford, A. L., Bowen, S. M., Todd, S.C., J. Immunol. 2001, 167, 5115–5121.
[18] Charrin, S., Le Naour, F., Billard, M., Labas, V. et al., Biochem.J. 2003, 373, 409–421.
[19] Zhang, X. A., Lane, W. S., Charrin, S., Rubinstein, E., Liu, L.,Cancer Res. 2003, 63, 2665–2674.
[20] Le Naour, F., Charrin, S., Labas, V., Le Caer, J. P. et al., CancerImmunol. Immunother. 2004, 53, 148–152.
[21] Little, K. D., Hemler, M. E., Stipp, C. S., Mol. Biol. Cell. 2004,15, 2375–2387.
[22] Santoni, V., Molloy, M., Rabilloud, T., Electrophoresis 2000,6, 1054–1070.
[23] Hanash, S., Proteomics 2001, 5, 635–637.
[24] Whitelegge, J. P., Gomez, S. M., Faull, K. F., Adv. ProteinChem. 2003, 65, 271–307.
[25] Rabilloud, T., Nat. Biotechnol. 2003, 21, 508–510.
[26] Simpson, R. J., Connolly, L. M., Eddes, J. S., Pereira, J. J. etal., Electrophoresis 2000, 21, 1707–1732.
[27] Adam, P. J., Boyd, R., Tyson, K. L., Fletcher, G. C. et al., J.Biol. Chem. 2003, 278, 6482–6489.
[28] Aebersold, R., Mann, M., Nature 2003, 422, 198–207.
[29] Wu, C. C., Yates III, J. R., Nat. Biotechnol. 2003, 21, 262–267.
[30] Bergquist, J., Curr. Opin. Mol. Ther. 2003, 5, 310–314.
[31] Boucheix, C., Perrot, J. Y., Mirshahi, M., Giannoni, F. et al.,Leuk. Res. 1985, 9, 597–604.
[32] Bartolazzi, A., Fraioli, R., Tarone, G., Natali, P. G., Hybridoma1991, 6, 707–720.
[33] Lozahic, S., Christiansen, D., Manie, S., Gerlier, D. et al., Eur.J. Immunol. 2000, 30, 900–907.
[34] Shevchenko, A., Wilm, M., Vorm, O., Mann, M., Anal. Chem.1996, 68, 850–858.
[35] Eng, J. K., McCornack, A. L., Yates III, J. R., J. Am. Soc. MassSpectrom. 1994, 5, 976–989.
[36] Ducret, A., Van Oostveen, I., Eng, J. K., Yates III, J. R.,Aebersold, R., Prot. Sci. 1998, 7, 706–719.
[37] Caravatti, P., Allemann, M., Org. Mass Spectrom. 1991, 26,514–518.
[38] Baykut, G., Jertz, R., Witt, M., Rapid Commun. Mass Spec-trom. 2000, 14, 1238–1247.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

Proteomics 2006, 6, 1437–1449 Cell Biology 1449
[39] Serru, V., Le Naour, F., Billard, M., Azorsa, D. O. et al., Bio-chem. J. 1999, 340, 103–111.
[40] Leibovitz, A., Stinson, J. C., McCombs, W. B. 3rd, McCoy, C.E. et al., Cancer Res. 1976, 36, 4562–4569.
[41] Szala, S., Froehlich, M., Scollon, M., Kasai, Y. et al., Proc.Natl. Acad. Sci. USA 1990, 87, 3542–3546.
[42] De Boer, C. J., van Krieken, J. H., Janssen-van Rhijn, C. M.,Litvinov, S. V., J. Pathol. 1999, 188, 201–206.
[43] Mellstedt, H., Fagerberg, J., Frodin, J. E., Hjelm-Skog, A. L. etal., Ann. NY Acad. Sci. 2000, 910, 254–261.
[44] Moulson, C. L., Li, C., Miner, J. H., Dev. Dyn. 2001, 222, 101–114.
[45] El Nemer, W., Gane, P., Colin, Y., Bony, V. et al., J. Biol. Chem.1998, 273, 16686–16693.
[46] El Nemer, W., Gane, P., Colin, Y., D’Ambrosio, A. M. et al., J.Biol. Chem. 2001, 276, 23757–23762.
[47] Oberst, M., Anders, J., Xie, B., Singh, B. et al., Am. J. Pathol.2001, 158, 1301–1311.
[48] Oberst, M. D., Singh, B., Ozdemirli, M., Dickson, R. B. et al., J.Histochem. Cytochem. 2003, 51, 1017–1025.
[49] Lee, S. L., Dickson, R. B., Lin, C. Y., J. Biol. Chem. 2000, 275,36720–36725.
[50] Bass, R., Werner, F., Odintsova, E., Sugiura, T. et al., J. Biol.Chem. 2005, 280, 14811–14818.
[51] Primakoff, P., Myles, D. G., Science 2002, 296, 2183–2185.
[52] White, J. M., Curr. Opin. Cell Biol. 2003, 15, 598–606.
[53] Blobel, C. P., Nature Rev. 2005, 6, 32–43.
[54] Yan, Y., Shirakabe, K., Werb, Z., J. Cell Biol. 2002, 158, 221–226.
[55] Teng, F. Y., Wang, Y., Tang, B. L., Genome Biol. 2001, 2,3012.1–3012.7.
[56] Le Naour, F., Rubinstein, E., Jasmin, C., Prenant, M., Bou-cheix, C., Science 2000, 287, 319–321.
[57] Miyado, K., Yamada, G., Yamada, S., Hasuwa, H. et al., Sci-ence 2000, 287, 321–324.
[58] Conze, T., Lammers, R., Kuci, S., Scherl-Mostageer, M. et al.,Ann. NY Acad. Sci. 2003, 996, 222–226.
[59] Buhring, H. J., Kuci, S., Conze, T., Rathke, G. et al., Stem Cells2004, 22, 334–343.
[60] Baumgartner, S., Martin, D., Chiquet-Ehrismann, R., Sutton,J. et al., J. Mol. Biol. 1995, 251, 41–49.
[61] O’Regan, S., Traiffort, E., Ruat, M., Cha, N. et al., Proc. Natl.Acad. Sci. USA 2000, 97, 1835–1840.
[62] Sterk, L. M., Geuijen, C. A., van den Berg, J. G., Claessen, N.et al., J. Cell Sci. 2002, 115, 1161–1173.
[63] Guo, W., Giancotti, F. G., Nat. Rev. Mol. Cell Biol. 2004, 10,816–826.
[64] Sincock, P. M., Fitter, S., Parton, R. G., Berndt, M. C., Gamble,J. R., Ashman, L. K., J. Cell. Sci. 1999, 112, 833–844.
[65] Baudoux, B., Castanares-Zapatero, D., Leclercq-Smekens,M., Berna, N., Poumay, Y., Eur. J. Cell. Biol. 2000, 79, 41–51.
[66] Yang, X., Kovalenko, O. V., Tang, W., Claas, C. et al., J. CellBiol. 2004, 167, 1231–1240.
[67] El Nemer, W., Colin, Y., Bauvy, C., Codogno, P. et al., J. Biol.Chem. 1999, 274, 31903–31908.
[68] Gauthier, E., Rahuel, C., Wautier, M. P., El Nemer, W. et al., J.Biol. Chem. 2005, 280, 30055–30062.
© 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com




















