
Genome-wide transcriptional changes induced by phagocytosis or growth on bacteria in Dictyostelium

ARTICLE IN PRESS
1438-4221/$ - se
doi:10.1016/j.ijm
�Correspondfax: +49531 39
E-mail addr
International Journal of Medical Microbiology 299 (2009) 489–508
www.elsevier.de/ijmm
Proteomic analysis of Legionella-containing phagosomes isolated from
Dictyostelium
Olga Shevchuka,b, Christoph Batzillab, Sonja Hagelec, Harald Kuschc,Susanne Engelmannd, Michael Heckerd, Albert Haase, Klaus Heunerb,Gernot Glocknerf, Michael Steinerta,�
aInstitut fur Mikrobiologie, Technische Universitat Braunschweig, Spielmannstr. 7, D-38106 Braunschweig, GermanybInstitut fur Molekulare Infektionsbiologie, Universitat Wurzburg, GermanycZentrum fur Physiologie und Pathophysiologie, Abteilung Herz-Kreislaufphysiologie, Gottingen, GermanydInstitut fur Mikrobiologie, Ernst-Moritz-Arndt-Universitat, Greifswald, GermanyeInstitut fur Zellbiologie, Universitat Bonn, GermanyfLeibniz Institute for Age Research – Fritz Lipmann Institute, Jena, Germany
Received 23 December 2008; received in revised form 2 March 2009; accepted 14 March 2009
Abstract
Legionella pneumophila, the agent of Legionnaires’ disease, replicates intracellularly within specialized phagosomesof human macrophages and amoebae. In this study, we have developed a protocol for the isolation of Legionella-containing phagosomes from Dictyostelium discoideum. Cell fractionation, two-dimensional gel electrophoresis andMALDI-TOF MS combined with genomic data identified 157 phagosome host proteins. In addition to proteins withan evident role in phagosome maturation, we identified proteins for which a function remains to be elucidated. Possibleinteractions of coronin with cytosolic NADPH oxidase components and protein kinase C inhibitors which togethermay lead to an inhibition of phagosomal superoxide generation are discussed. Comparative proteomics ofphagosomes containing highly virulent L. pneumophila Corby versus less virulent L. hackeliae revealed distinctiveprotein expression patterns, e.g., an abundance of RhoGDI in L. hackeliae degrading phagosomes versus littleRhoGDI in L. pneumophila Corby replicative phagosomes. We present a kinetic dissection of phagosome maturationincluding the complex alterations of the phagosome protein composition. A reference flow chart suggests so farunrecognized consequences of infection for host cell physiology, actin degradation on phagosomes, and a putativecysteine proteinase inhibitor interference with lysosomal enzyme sorting and activation processes.r 2009 Elsevier GmbH. All rights reserved.
Keywords: Legionella pneumophila; Dictyostelium discoideum; Phagosome; Proteome
e front matter r 2009 Elsevier GmbH. All rights reserved.
m.2009.03.006
ing author. Tel.: +49531 3915802;
15854.
ess: [email protected] (M. Steinert).
Introduction
Legionella pneumophila is an environmental Gram-negative bacterium that often causes a severe and life-threatening pneumonia in humans. The aetiologicalagent of the so-called Legionnaires’ disease is naturally

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508490
found in fresh water where the bacteria parasitizeintracellularly within various amoeba species. Transmis-sion of L. pneumophila to humans takes place byinhalation of aerosols from contaminated water sources(Steinert et al., 2007). After uptake by alveolar macro-phages the Legionella containing phagosomes bypassendolysosomal degradation, develop a rough endosomalreticulum-derived replicative niche, and undergo atransition from a tight to a spacious vacuole (Molmeretet al., 2005; Li et al., 2005). Experiments with severalcellular infection models including the soil amoebaDictyostelium discoideum have shown that the modula-tion of host cell functions is important for the biogenesisof the Legionella-specific replicative vacuole (Hilbi et al.,2007).
The haploid amoeba D. discoideum has proven to be arepresentative host model system to analyze cellularaspects of Legionella pathogenicity (Hagele et al., 2000;Solomon et al., 2000; Steinert and Heuner, 2005; Weberet al., 2006; Chen et al., 2007a, b). The complete genomesequence and the analysis of the Dictyostelium tran-scriptome upon infection with L. pneumophila provideda better understanding of the manifold host cellresponses (Farbrother et al., 2006). Functional annota-tion of differentially regulated genes revealed that byestablishing its replicative niche Legionella not onlyinterferes with bacterial degradation and intracellularvesicle trafficking but also profoundly influences andexploits the metabolism of its host. Functional studiesusing infection assays, pharmacological inhibitors,Dictyostelium mutants including GFP-tagged host cellproteins revealed that entry of L. pneumophila is anactin-mediated process which involves the phospholi-pase C pathway, that cytoplasmic calcium levels, thecytoskeleton proteins coronin, villidin, and a-actinin/filamin as well as the calcium-binding ER proteinscalreticulin and calnexin significantly influence thisprocess, and that the newly formed phagosome istransported along microtubules (Fajardo et al., 2004;Lu and Clarke, 2005). In addition, Dictyostelium
mutants with an altered ability to support Legionella
multiplication have been identified. The decreasedintracellular replication of L. pneumophila in aD. discoideum ratioA (rtoA–) mutant obviously resultsfrom a lowered efficiency of vesicle trafficking importantfor the integrity of the Legionella-containing compart-ment (Li et al., 2005). Of particular interest are thesubstrates of the Legionella Dot/Icm type IV secretionsystem. SidM/DrrA and LidA target the mammalianRab1, a small GTPase regulating ER-to-Golgi traffic.RalF recruits and activates ADP ribosylation factor 1(Arf1), a small GTPase involved in retrograde vesicletransport from the Golgi apparatus to the ER (Nagaiet al., 2002; Derre and Isberg, 2005; Machner andIsberg, 2006; Murata et al., 2006). Analysis of theD. discoideum transcriptome revealed that genes belong-
ing to the Dictyostelium ubiquitination machinery werestrongly upregulated after infection with L. pneumophila
wild type as compared to the dotA mutant (Farbrotheret al., 2006). Thus, DNA sequencing and transcriptomeanalysis identified new candidate genes which may beinvolved in host–pathogen interactions (Cazalet et al.,2004). The challenge now is to translate this body ofinformation into the protein equivalent.
In the present study, proteome analysis of isolatedDictyostelium phagosomes by two-dimensional gelelectrophoresis and mass spectrometry were used tocharacterize the reprogramming of this organelle afterinfection with L. pneumophila or L. hackeliae. Theidentified phagosome proteins belonged to a variety offunctional categories and exhibited characteristic tem-poral profiles. Moreover, this approach allowed us tomonitor specifically Legionella-induced pathogenic al-terations of phagosome maturation.
Materials and methods
Bacterial strains and cell culture conditions
L. pneumophila Corby (serogroup S1) (Jepras et al.,1985) and L. hackeliae (Ott et al., 1991) were grown onbuffered charcoal–yeast (BCYE) agar at 37 1C with 5%CO2 atmosphere for 3 days (Edelstein, 1981). TheD. discoideum wild-type strain AX2 was grown axenicallyat 24 1C in HL5 medium (Clarke et al., 1980) withagitation (150 rpm) for 3 days.
Antibodies
The monoclonal mouse anti-a-mitoporin 70-100-1and anti-calnexin 270-390-2 antibodies were a gift ofAnnette Muller-Taubenberger, Martinsried, Germany.Monoclonal mouse anti-actin-1 was a gift of MichaelSchleicher, Munchen, Germany. The polyclonal goatanti-mouse antibody HRP was from Dianova (Ham-burg, Germany).
Infection of D. discoideum cells
For proteome analysis of Legionella-containingphagosomes, D. discoideum AX2 was infected withL. pneumophila Corby or L. hackeliae. L. pneumophila
Corby is able to replicate to high numbers in theDictyostelium model while L. hackeliae is degraded.Previous studies have shown that knowledge acquiredthrough the study of infected Dictyostelium cells can beextended to mammalian phagosome biogenesis,although the course of infection is delayed (Hageleet al., 2000; Solomon et al., 2000). D. discoideum cellswere grown axenically in HL5 medium for 3 days,

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 491
collected by centrifugation for 5min at 1000 rpm (RT)and suspended in infection medium (1:1 solution of HL5medium and Soerensen phosphate buffer, pH 6.0) tofinal concentration of 1� 108 cells/ml. Three-day-oldLegionella colonies were suspended in 1ml sterile H2Oand cell density adjusted to 5� 109 cells/ml. In order toachieve a synchronization of infection, host cells wereco-incubated with Legionella at a MOI of 20. Infectionwas at 24 1C in 25 cm2 cell culture flasks. The overallprogression of infection was monitored by fluorescencemicroscopy (see below). Rhodamine labelling of bacte-ria was as described previously (Unal and Steinert, 2006).Colloidal iron particles (final concentration of 1mg/ml)were added 30min before harvesting D. discoideum cellsfor phagosome isolation.
Immunofluorescence
For immunofluorescence, 106 Dictyostelium cellsinfected with rhodamine labelled bacteria were allowedto settle onto glass cover slips for 30min. Cells werefixed with ice-cold methanol for 10min and post-fixedwith ice-cold acetone for 10min. Subsequently, cellswere washed in TBST buffer (50mM Tris, 0.2M NaCl,pH 7.4, 0.1% v/v Tween 20, pH 7.4) and stained withthe primary calnexin antibody. Secondary Cy2-conju-gated goat anti-mouse antibody was from Dianova.The cells were observed using an Zeiss Axioplan 2fluorescence microscope equipped with a 100� /1.3Plan-Neofluar objective. Number of infected and non-infected Dictyostelium cells as well as extra- andintracellular Legionella were calculated.
Preparation of iron particles
Dextran-coated colloidal iron particles with anaverage diameter of 8 nm were prepared as describedpreviously (Rodriguez-Paris et al., 1993). In brief, 10mlof 1.2M FeCl2 were mixed with 10ml of 1.8M FeCl3and agitated extensively while 10ml 25% NH3 wereadded. The suspension was divided into 6 aliquots andplaced on a magnet unit until the precipitate hadgathered on the bottom of the tube. After the super-natant was decanted, the precipitate was washed oncewith 5% NH3 and twice with deionized H2O. Thesediment was suspended in 80ml of 0.3M HCl andstirred for 30min. Then 4 g of dextran (64–76 kDa,Sigma-Aldrich, Schnelldorf, Germany) were added tothe suspension and stirred for another 30min. Thesample was dialyzed against cold water for 2 days withrepeated buffer changes and filtered through a 0.22 mmsterile filter (Millipore, Billerica, MA, USA). Thesolution was stored at 4 1C (maximum 3 months). Theiron concentration of this solution was 10mg/ml.
Purification of bacteria-containing phagosomes
Phagosomes were purified according to Luhrmannand Haas (2000) with modifications. All steps wereperformed on ice. Infected D. discoideum cells (108 cells)were harvested and washed twice in 30ml ice-cold 1�Soerensen buffer and once in ice-cold homogenizationbuffer HB (0.5mM Na2EGTA, 20mM HEPES,250mM Sucrose) containing EDTA-free protease in-hibitor cocktail (Roche Diagnostic, Penzberg, Ger-many). Washed cells were suspended in 2ml of HBcontaining 5mg/ml INT, broken by 12 strong strokes ina Dura Grind stainless-steel homogenizer and incubatedwith 50U Benzonase/ml for 7min at 37 1C. Nuclei andcell debris were removed from the homogenate bycentrifugation at 500g for 5min at 4 1C. Lysosomalcontamination in the obtained postnuclear supernatantwas eliminated by running the supernatant through aMiltenyi Biotec MiniMACS column in a magneticseparation unit (OctoMACS, Miltenyi Biotec, BergischGladbach, Germany). The resulting suspension wasadjusted to a volume of 3ml by HB, laid on top of an8-ml 5–30% OptiPrep gradient in HB and centrifuged at100,000g for 2 h at 4 1C (SW40 rotor, Beckmanultracentrifuge). The bottom of the Polyallomer cen-trifuge tube (Beckman, CA, USA) was pierced with aneedle (27G� 3/400; 0.4� 20mm) and 0.6–0.8ml frac-tions were collected. Division of the OpiPrep gradientinto numerous fractions with small volume improvedpurity of Legionella-containing phagosomes signifi-cantly. To determine the positioning of Legionella-containing phagosomes in the OptiPrep gradient, serialdilutions of the respective fractions were plated onBCYE agar plates. Purity of the phagosomal fractionswas assessed by transmission electron microscopy.
Preparation of fractions for transmission electron
microscopy
OptiPrep fractions were examined by transmissionelectron microscopy. Samples were fixed in 2.5%glutaraldehyde for 1 h and centrifuged at 5000 rpm for30min in a Heraeus biofuge 13R. The supernatant wasremoved, and the pellet was incubated in 2% osmiumte-troxid (in deionized H2O) overnight at 4 1C. Sampleswere dehydrated in an ethanol series (50%, 70%, 80%,90%, and twice with 100% ethanol) and finally withpropylenoxid. Subsequently, the pellet was embedded inEpon–propylene oxide (1:1). Following overnight in-cubation, this mixture was replaced by fresh Epon,incubated at room temperature for 2–3 h and thenpolymerised for 3 days at 60 1C. Examination of thinsections was performed with a Zeiss A 100 transmissionelectron microscope at 80 kV.

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508492
Preparation of the phagosome protein fraction
For separation of bacterial from host cell proteins,samples were treated with Triton X-100 (final concen-tration 0.5%), incubated on ice for 30min andcentrifuged for 5min at 25,000g and 4 1C. Proteinsfrom the resulting supernatant were precipitated over-night with 10% trichloroacetic acid solution (v/v),washed thoroughly with aceton, and resuspended inlysis buffer (30mM Tris, 7M urea, 2M thiourea, 4%(w/v) CHAPS(3-[(3-Cholamidopropyl)dimethylammo-nio]-1-propanesulfonate); pH adjusted to 8.0 with 1MHCl). The phagosome protein concentration was deter-mined using the Roti-Nanoquant (Roth, Karlsruhe,Germany). The protein solution was stored at �20 1C.
Protein labelling
Phagosome proteins (5 mg), solubilized in lysis buffer,were labelled with the dyes Cy3 or Cy5 (AmershamBiosciences, Freiburg, Germany) according to themanufacturer’s protocol. In brief, cysteine residues inthe extracted proteins were reduced by incubating with2mM TCEP (tris(2-caboxyethyl)phosphine), at 37 1Cfor 1 h. Proteins were labelled with 2mM CyDye DIGEFluor in dimethylformamide (DMF), at 37 1C for30min. To stop the labelling reaction, the doublevolume of 2� sample buffer (7M urea, 2M thiourea,4% (w/v) CHAPS, 2% (v/v) Pharmalyte, 13 mM DTT)was added. The samples were stored either on ice or forup to one month at –20 1C in the dark.
2D Gel electrophoresis
For analytical gels, labelled proteins (total 0.01mg)were separated by 2D gel electrophoresis. The firstdimension, isoelectric focusing (IEF), was performed ina 24-cm Immobiline DryStrip with a nonlinear pH 3–10gradient using Ettan IPGphor (Amersham Biosciences)at 20 1C. Immobiline strip rehydration was performedfor 18 h in a rehydration buffer (1% Pharmalyte, 7Murea, 2M thiourea, 4% (w/v) CHAPS, and 1mg/mlDTT (dithiothreitol) containing 0.01mg Cy3- and Cy5-labelled proteins. Isoelectric focusing was performed in3 steps: at 500V for 1 h, at 1000V for 1 h, and 8000Vuntil the bromphenol blue front migrated out of thegel. Before running the second dimension, immobilinestrips with separated proteins were equilibrated in asolution containing 6M Urea, 100mM Tris–HCl,pH 8.0, 2% SDS, 30% glycerol, and 0.5% (w/v) DTTfor 10min at room temperature. In the second dimen-sion, the proteins were separated on a 12.5% SDS-polyacrylamide gel in a 1� SDS running buffer [25mMTris, 192mM glycine, 0.2% (w/v) SDS] at 2W/gel
constant current until the dye front migrated out of thegel.
For preparative gels 500 mg of phagosome proteinswere solubilized in 450 ml rehydration buffer containing8M urea, 2M thiourea, 1% CHAPS, 20mM DTT,0.5% and bromphenol blue and rehydrated for 18 h atroom temperature. Isoelectric focusing was performedin a IPGphor system (Amersham Bioscience) at 20 1Cusing the parameters as described for analytical gels.After IEF, strips were incubated at room temperature inthe equilibration solution A (50mM Tris, 6M urea,30% glycerin, 4% SDS containing 3.5mg/ml DTT) for15min and in the equilibration solution B (50mg/mliodoacetamide instead of DTT) for additional 15min. Inthe second dimension, the proteins were separated on a12.5% SDS-polyacrylamide gel in a 1� SDS runningbuffer [25mM Tris, 192mM glycine, 0.1% (w/v) SDS].Protein spots were visualized by Coomassie staining forMALDI-TOF analysis.
Gel image analysis
The separated proteins labelled with Cy3 and Cy5fluorophores were detected in the gels with a Typhoon8600 imaging system (Amersham Biosciences). Data setswere analyzed, in order to screen for different proteinamounts. 2D gel image analysis was performed with theDelta2D Software (Decodon, Germany). A set of 18 gels(3 gels for every strain and every time point) wereselected for quantitative analysis. Quantitation wasbased on spot detection and filtering, whole imagewarping on a reference gel, background subtraction, andaverage gel creation. Only statistically significantchanges in spot representation of 2.0 or higher wereanalyzed.
Protein identification by mass spectrometry
For protein identification, spots from preparativeCoomassie-stained gels were excised using a Ettan SpotPicker (Amersham Biosciences). Proteins in the excisedgel pieces were digested using in-gel trypsin digest, andMALDI-TOF measurements were carried out on aProteome Analyzer 4700 (Applied Biosystems, FosterCity, CA, USA) as described previously (Eymannet al., 2004). The identification of spots was via thebatch mode using the Mascot protein identificationsystem (Matrix Science, London, UK) applying theD. discoideum database (http://dictybase.org/) (Chisholmet al., 2006). Search parameters were as described byEymann et al. (2004). Search results that yielded aMowse score of at least 54 (po0.05) for MS data and 25(po0.05) for MS/MS data, a sequence coverage of atleast 20% were regarded as positive identifications.Additionally, pI value and molecular weight of each

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 493
protein derived from the position on the gels wascompared with theoretically predicted data from theamino acid sequence.
Immunoblot analysis
For immunoblot analysis, equal concentrations ofphagosome protein were loaded and separated in a12.5% urea SDS-PAGE as described previously (Golds-brough et al., 1989). Proteins were electroblotted onto anitrocellulose membrane in Towbin buffer at 70mA for75min (Towbin et al., 1979), and the membrane wasblocked overnight in phosphate buffered saline (PBS),pH 7.4 (150mM NaCl, 5mM KCl, 0.8mM KH2PO4,3.2mM Na2HPO4) with 0.1% (w/v) Tween-20 and 1%(m/v) milk. Then the blot was incubated for 2 h with thefirst antibody, washed again, and incubated with thesecond antibody (polyclonal goat anti-mouse Immuno-globulins/HRP; 1:5000) for 2 h. Bound antibody wasdetected by ECL-advance. Stained gels and immuno-blots were scanned using a Chemi Lux-Imager(Intas Science Imaging Instruments GmbH, Gottingen,Germany).
Fig. 1. Distribution of Legionella-containing phagosomes at
2 h post-infection in a representative OptiPrep gradient
fractionation. (A) An aliquot of each OptiPrep fraction was
plated onto a BCYE-agar plate, and CFU/ml of Legionella
were counted. The highest bacterial concentration corre-
sponded to phagosome fractions which are indicated by
asterisks. (B) Immunoblot analysis of OptiPrep fractions.
Monoclonal antibodies against mitochondrial a-mitoporin
and the ER marker protein calnexin indicated the separation
of these organelles into different fractions; D. discoideum lysate
was used as a control.
Results
Isolation of Legionella-containing phagosomes
In order to analyze phagosome protein changesduring Legionella infection, we have used the well-characterized interaction of D. discoideum with highlyvirulent L. pneumophila Corby and less virulentL. hackeliae, respectively. Both species are taken up byphagocytosis, however, only L. pneumophila Corbyis capable of growing intracellularly within a phago-some that inhibits the fusion with lysosomes, whileL. hackeliae is degraded by D. discoideum (Hagele et al.,2000).
Infection of axenically grown Dictyostelium AX2 cellswas as previously described (Unal and Steinert, 2006).Isolation of Legionella-containing phagosomes was at 2,4, and 6 h post-infection since most fateful cellularchanges occur during this period of time. Afterelimination of extracellular bacteria by several washingsteps, infected D. discoideum were lysed mechanically,unbroken cells and nuclei were removed, and thepostnuclear supernatant was treated with Benzonasefor nucleic acid degradation. Lysosomes were eliminatedmagnetically after colloidal iron loading. Mitochondrialcontaminations were eliminated by iodophenylnitrophe-nyltetrazolium (INT) ‘heavy labelling’. Succinate dehy-drogenase activity of mitochondria reduces INT and theresulting product formazan is deposited on mitochon-drial membranes. This step increases the density of
mitochondria and allows their separation from bacterialphagosomes by a continous OpiPrep gradient.
The distribution of Legionella-containing phagosomesin the gradient was determined by plating 18 separateOptiPrep fractions onto BCYE agar plates and countingbacterial CFU/ml (Fig. 1A). Fractions 6 and 7 wereused for 2-DE and MALDI-TOF-MS. The purity ofthe phagosome preparation was analyzed by immuno-blotting and transmission electron microscopy. Westernblots with antibodies to the well-characterized ERmarker calnexin or to the mitochondrial markera-mitoporin confirmed the separation of these orga-nelles by ultracentrifugation (Fig. 1B). Transmissionelectron microscopy of Legionella containing fractions(fractions 6 and 7) usually revealed one single bacteriumper ‘spacious vacuole’ (Fig. 2A and B). The blebs on thephagosomes have previously been described as ER-derived vesicles (Tilney et al., 2001). Additionally, emptyvesicular structures are visible which resemble break-down vesicles of the ER. The respective heavily labelledOptiPrep fractions (fractions 3 and 4) revealed mito-chondria and different types of vesicles (Fig. 2C).Lysosomes were detected in a separate fraction afterthe magnetic treatment of the postnuclear supernatant(Fig. 2D) which confirmed the successful elimination ofthis organelle from phagosome preparations. Thus, wepresent a method for the isolation of Legionella-containing phagosomes with purity to permit proteomicanalysis.

ARTICLE IN PRESS
.
0.5µm0.5µm 0.5µm0.5µm
0.5µm0.5µm0.5µm0.5µm
Fig. 2. Representative transmission electron micrographs of different subcellular fractions after 2-h infection of D. discoideum with
Legionella pneumophila Corby. (A) and (B) Isolated Legionella-containing phagosomes (corresponds to fractions 6 and 7 in Fig. 1).
(C) Mitochondria from the OptiPrep fractions (corresponds to fractions 3 and 4 in Fig. 1). (D) Lysosomes eliminated from the
phagosome fraction by running the supernatant through a Miltenyi Biotec MiniMACS column in a magnetic separation unit.
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508494
2-D gel electrophoresis and identification of
phagosomal proteins
To obtain the phagosome proteome, we separated thebacteria from the phagosome membrane by detergenttreatment and subsequent sedimentation of the bacteria.We found that the use of harsher detergents to solubilizehydrophobic and membrane-associated proteins resultsin contamination with bacterial compounds. Such anartefact, however, would lead to the misinterpretationthat these contaminants are secreted or naturallyassociate with the phagosome. To map the proteome,representative fractions were separated by preparative2-DE. Subsequently, 439 protein spots were subjected toMALDI-TOF-MS analysis. Several proteins producedmore than one spot or even multiple spots which mightbe due to artificial (deamidations) or post-translationalmodifications. A total of 157 different proteins could beidentified by mass spectrometry. Using the Dictyoste-
lium genome database (http://dictybase.org/), a total of138 spots were identified as proteins or open readingframes with a known or predicted function. Thefunctions of the remaining 19 ‘hypothetical proteins’are unknown. Formally, it cannot be excluded that oneprotein, which has been identified as S-adenosyl-L-ho-mocysteine hydrolase, is a Legionella protein. However,since this protein is evolutionary highly conserved and
other putative Legionella contaminations could not bedetected, it is very likely that this is also a Dictyostelium
protein. All identified proteins including their accessionnumbers, their known or predicted functions, as well astheir molecular weights and pIs are listed in Table 1.
The identified phagosome proteins belong to a varietyof functional categories (Fig. 3). The largest groupcomprises 32 metabolic proteins. The 22 proteins of thesecond largest group are members of the proteinbiosynthesis category. The category cytoskeleton andsignal transduction comprises 28 proteins, and 10proteins were assigned to the proteasome and proteasecategory. Other proteins belong to the mitochondrialcategory (10 proteins), stress response category(11 proteins), oxidoreductase activity category (6 proteins),subunits of the H+ATPase (5 proteins), intracellularprotein transport category (3 proteins), ER category(5 proteins), iron transport category (2 proteins), proteinfolding category (2 proteins), and the transcriptionregulation and splicing category (2 proteins). Takentogether, the phagosome proteome mapping demon-strated that we have identified many proteins with anevident role in phagocytosis or phagosome maturation.In addition, we found proteins for which a link tophagosomal processes remains to be established. In thefollowing, we will describe temporal and pathologicalchanges of the phagosome proteome.

ARTICLE IN PRESS
Table 1. Phagosome proteins identified by MALDI MS.
Protein Acc. number Function MW pI
Metabolism (32)
Carbohydrate metabolism (8)
ALDHb,c DDB0231504 Aldehyde dehydrogenase 55.16 5.78
AlrAa,b,c DDB0215363 Aldehyde reductase 33.62 6.31
Fbab,c DDB0231387 Fructose-bisphosphate aldolase 38.88 7.01
GpdAc DDB0185087 Glyceraldehyde-3-phosphate dehydrogenase 36.53 6.46
IdhCc DDB0231401 Isocitrate dehydrogenase (NADP+) 46.34 6.36
Nag B1b DDB0186848 Glucosamine-6-phosphate isomerase 82.64 6.14
pckAc DDB0231108 Phosphoenolpyruvate carboxykinase 62.53 6.51
Zwfb,c DDB0231285 Glucose 6-phosphate-1-dehydrogenase 56.66 7.16
Fatty acid metabolism (3)
FcsAc DDB0191105 Fatty acyl-CoA synthetase 74.56 6.03
PccAb,c DDB0230063 Propionyl-CoA carboxylase 79.77 6.29
PccBb,c DDB0166993 Propionyl-CoA carboxylase beta chain 60.65 6.82
Vitamins, co-enzymes, and hormones (1)
JC3V2_0_00291 DDB0205386 Putative ATP citrate lyase 50.23 6.05
Purines and pyrimidines (8)
DDB0230174b,c DDB0230174 Adenosine kinase 56.27 5.57
DDB0230202b DDB0230202 5’-nucleotidase 34.44 6.09
DDB0231470b DDB0231470 Urate oxidase 33.01 8.95
GuaBb,c DDB0230098 IMP dehydrogenase 44.72 7.97
PrsAa DDB0186131 Phosphoribosyl pyrophosphate synthetase 37.10 7.68
Pyr56b,c DDB0214958 Bifunctional UMP-synthetase 35.37 6.21
ThyAb,c DDB0214905 Thymidylate synthase (FAD) 32.36 6.44
UdpBa,b DDB0230170 Uridine phosphorylase 33.01 5.69
Amino acid biosynthesis and catabolism (12)
ArgEc DDB0191165 Acetylornithine deacetylase 49.04 5.28
AsnA DDB0230140 Asparagine synthetase 63.45 5.90
DDB0230064 DDB0230064 Sulfate adenylyltransferase 65.58 6.18
DDB0230070c DDB0230070 S-adenosylmethionine synthetase 41.74 6.14
DDB0230072c DDB0230072 Serine hydroxymethyltransferase 50.66 6.54
DDB0231438b,c DDB0231438 NAD-dependent glutamate dehydrogenase 55.01 7.98
GlnBa DDB0231551 Glutamate-ammonia ligase 82.56 5.80
Hpdb DDB0203373 4-hydroxyphenylpyruvate dioxygenase 41.97 5.77
Lpdb,c DDB0216232 Dihydrolipoamide:NAD oxidoreductase 51.76 6.88
Pyd3b DDB0185221 Beta-alanine synthase 44.06 6.32
SahAc DDB0191108 S-adenosyl-L-homocysteine hydrolase 47.25 5.81
SerA DDB0230052 3-phosphoglycerate dehydrogenase 44.72 6.61
Protein biosynthesis (22)
Ribosome and elongation factors (15)
DDB0192007 DDB0192007 Poly(A)-mRNA binding protein 62.74 8.83
Efa1Bc DDB0191174 Elongation factor 1 beta 24.11 4.54
EfaAIa,c DDB0191135 Elongation factor 1 alpha 49.63 9.07
EfbAa,c DDB0191363 Elongation factor 2 92.60 6.22
eIF-3 betab DDB0186657 Eukaryotic translation initiation factor 3, subunit 2 37.01 6.56
EifEc DDB0191442 eifE 17.08 4.95
IfdAc DDB0191262 Eukaryotic translation initiation factor 4A 44.32 6.04
Moe1a,b DDB0219927 Eukaryotic translation initiation factor 3, subunit 7 60.26 5.67
Rpl11a,b,c DDB0214853 S60 ribosomal protein L11 22.38 10.12
RPLP0c DDB0219977 Ribosomal acidic phosphoprotein P0 32.83 9.01
Rps12a,c DDB0230044 40S ribosomal protein S12 14.90 6.73
Rps3b,c DDB0201667 40S ribosomal protein S3 24.20 9.83
Rps5b,c DDB0230022 40S ribosomal protein S5 21.20 9.93
RpsAa,c DDB0230016 40S ribosomal protein SA 27.10 5.29
TufMb,c DDB0219464 Elongation factor Tu domain-containing protein 46.60 7.2
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 495

ARTICLE IN PRESS
Table 1. (continued )
Protein Acc. number Function MW pI
TRNA synthetase (7)
ArgS1b DDB0231324 Arginyl-tRNA synthetase 67.00 6.43
AspS1 DDB0231308 Aspartyl-tRNA synthetase 64.89 6.10
CysSb DDB0231318 Cysteinyl-tRNA synthetase 74.62 6.63
GlyS DDB0231313 Glycyl-tRNA synthetase 76.17 6.44
LysS DDB0231263 Lysyl-tRNA synthetase 61.96 5.80
SerSc DDB0231305 Seryl-tRNA synthetase 51.55 6.13
ThrS1b DDB0231248 Threonyl-tRNA synthetase 81.14 6.66
Proteasome & proteases (10)
BC5V2_0_01884a,c DDB0189202 Peptidase M16 family protein 47.72 6.30
CpiAa,b,c DDB0220657 Cysteine protease inhibitor 10.41 6.40
JC1V2_0_01800b DDB0190923 Similar to methionine aminopeptidase 2 46.58 6.52
PrlAb DDB0204785 Proliferation associated protein 38.14 6.47
PsmA1b,c DDB0214956 20S proteasome subunit alpha-1 27.97 6.00
PsmC1 DDB0202018 26S protease regulatory subunit 4 49.16 5.90
PsmC2 DDB0168337 26S protease regulatory subunit 7 47.37 6.10
PsmC3b DDB0186002 26S protease regulatory subunit 6A 46.84 5.36
PsmC4b DDB0191435 26S protease regulatory subunit 6B 45.51 6.45
PsmD14 DDB0191298 26S proteasome nonATPase regulatory subunit 14 34.43 6.07
Protein folding (2)
Cct6 DDB0169209 Chaperonin containing TCP1 zeta subunit 59.35 6.20
Tcp1b DDB0191128 Chaperonin containing TCP1 alpha subunit 59.37 6.55
Oxidoreductase activity (6)
BC5V2_0_01017b DDB0188339 Putative amino oxidase 52.33 6.07
BEC6V2_0_00612a,b DDB0219748 Oxidoreductase activity 94.79 6.06
CatAc DDB0185123 Catalase 55.64 8.61
MfeBb DDB0214811 Hypothetical peroxisomal multifunctional enzyme 32.38 6.40
Prdx4b,c DDB0217516 Peroxiredoxin 4 22.95 6.09
SodAa,b,c DDB0191290 Superoxide dismutase 15.85 5.85
Vacuolar H+ATPase (5)
Atp5b DDB0190669 ATP synthase beta-subunit 70.80 5.62
Atp5C1 DDB0184316 ATP synthase F1 subunit alpha. 33.61 7.71
BC4V2_0_00187b,c DDB0185431 ATP synthesis coupled proton transport 33.11 6.86
VatAa,c DDB0201563 Vacuolar H+-ATPase A subunit 68.16 5.35
VatBa,c DDB0185207 Vacuolar H+ ATPase B subunit 54.84 5.53
Mitochondrial proteins (10)
Aassb DDB0218638 Lysine-ketoglutarate reductase activity 102.91 6.19
CxeAa,b,c DDB0191104 Cytochrome c oxidase subunit V 13.50 6.60
DDB0220638b,c DDB0220638 Citrate synthase 51.18 8.09
FumHa,b,c DDB0231400 Fumarate hydratase 50.69 6.81
IdhBb DDB0231294 Isocitrate dehydrogenase 38.66 8.48
MdhBc DDB0230188 Malate dehydrogenase 37.62 8.78
NdkCc DDB0185051 Nucleoside diphosphate kinase 16.79 7.97
NdkMa,b,c DDB0214817 Nucleoside diphosphate kinase 23.98 9.69
Oxct1b DDB0187790 3-oxoacid CoA-transferase 54.90 8.07
SdhAb,c DDB0214886 Succinate dehydrogenase 68.47 6.46
Cytoskeleton and signal transduction (28)
AbpBb,c DDB0214810 Actin bundling protein 33.33 6.19
AclAa,b,c DDB0219936 Component of the Arp2/3 complex 46.67 5.72
AcpAa,b,c DDB0191202 Actin capping protein 30.68 5.02
AcpBa,b,c DDB0191243 Actin capping protein 31.14 6.84
Acta,c Actin 41–43 –
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508496
ARTICLE IN PRESS
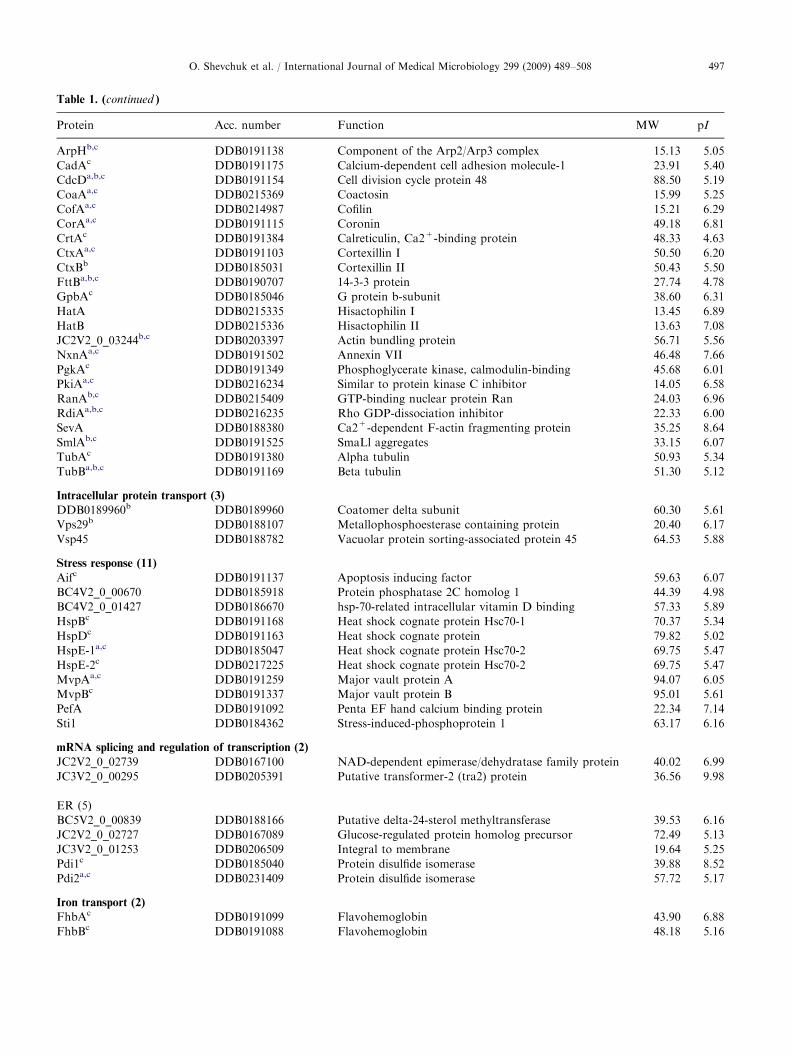
Table 1. (continued )
Protein Acc. number Function MW pI
ArpHb,c DDB0191138 Component of the Arp2/Arp3 complex 15.13 5.05
CadAc DDB0191175 Calcium-dependent cell adhesion molecule-1 23.91 5.40
CdcDa,b,c DDB0191154 Cell division cycle protein 48 88.50 5.19
CoaAa,c DDB0215369 Coactosin 15.99 5.25
CofAa,c DDB0214987 Cofilin 15.21 6.29
CorAa,c DDB0191115 Coronin 49.18 6.81
CrtAc DDB0191384 Calreticulin, Ca2+-binding protein 48.33 4.63
CtxAa,c DDB0191103 Cortexillin I 50.50 6.20
CtxBb DDB0185031 Cortexillin II 50.43 5.50
FttBa,b,c DDB0190707 14-3-3 protein 27.74 4.78
GpbAc DDB0185046 G protein b-subunit 38.60 6.31
HatA DDB0215335 Hisactophilin I 13.45 6.89
HatB DDB0215336 Hisactophilin II 13.63 7.08
JC2V2_0_03244b,c DDB0203397 Actin bundling protein 56.71 5.56
NxnAa,c DDB0191502 Annexin VII 46.48 7.66
PgkAc DDB0191349 Phosphoglycerate kinase, calmodulin-binding 45.68 6.01
PkiAa,c DDB0216234 Similar to protein kinase C inhibitor 14.05 6.58
RanAb,c DDB0215409 GTP-binding nuclear protein Ran 24.03 6.96
RdiAa,b,c DDB0216235 Rho GDP-dissociation inhibitor 22.33 6.00
SevA DDB0188380 Ca2+-dependent F-actin fragmenting protein 35.25 8.64
SmlAb,c DDB0191525 SmaLl aggregates 33.15 6.07
TubAc DDB0191380 Alpha tubulin 50.93 5.34
TubBa,b,c DDB0191169 Beta tubulin 51.30 5.12
Intracellular protein transport (3)
DDB0189960b DDB0189960 Coatomer delta subunit 60.30 5.61
Vps29b DDB0188107 Metallophosphoesterase containing protein 20.40 6.17
Vsp45 DDB0188782 Vacuolar protein sorting-associated protein 45 64.53 5.88
Stress response (11)
Aifc DDB0191137 Apoptosis inducing factor 59.63 6.07
BC4V2_0_00670 DDB0185918 Protein phosphatase 2C homolog 1 44.39 4.98
BC4V2_0_01427 DDB0186670 hsp-70-related intracellular vitamin D binding 57.33 5.89
HspBc DDB0191168 Heat shock cognate protein Hsc70-1 70.37 5.34
HspDc DDB0191163 Heat shock cognate protein 79.82 5.02
HspE-1a,c DDB0185047 Heat shock cognate protein Hsc70-2 69.75 5.47
HspE-2c DDB0217225 Heat shock cognate protein Hsc70-2 69.75 5.47
MvpAa,c DDB0191259 Major vault protein A 94.07 6.05
MvpBc DDB0191337 Major vault protein B 95.01 5.61
PefA DDB0191092 Penta EF hand calcium binding protein 22.34 7.14
Sti1 DDB0184362 Stress-induced-phosphoprotein 1 63.17 6.16
mRNA splicing and regulation of transcription (2)
JC2V2_0_02739 DDB0167100 NAD-dependent epimerase/dehydratase family protein 40.02 6.99
JC3V2_0_00295 DDB0205391 Putative transformer-2 (tra2) protein 36.56 9.98
ER (5)
BC5V2_0_00839 DDB0188166 Putative delta-24-sterol methyltransferase 39.53 6.16
JC2V2_0_02727 DDB0167089 Glucose-regulated protein homolog precursor 72.49 5.13
JC3V2_0_01253 DDB0206509 Integral to membrane 19.64 5.25
Pdi1c DDB0185040 Protein disulfide isomerase 39.88 8.52
Pdi2a,c DDB0231409 Protein disulfide isomerase 57.72 5.17
Iron transport (2)
FhbAc DDB0191099 Flavohemoglobin 43.90 6.88
FhbBc DDB0191088 Flavohemoglobin 48.18 5.16
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 497

ARTICLE IN PRESS
Table 1. (continued )
Protein Acc. number Function MW pI
Unknown function (19)
BC4V2_0_00250 DDB0218489 Unknown 26.24 5.27
BC4V2_0_01063 DDB0186311 Hemopexin domain signature 31.67 4.80
BC5V2_0_00316 DDB0219276 Unknown 37.50 5.17
BC5V2_0_00460c DDB0187793 Unknown 29.08 9.54
BC5V2_0_01650 DDB0188942 Unknown 53.33 6.06
BEC6V2_0_01236 DDB0219884 Hemopexin domain signature 80.34 5.19
CinBc DDB0220110 Esterase/lipase/thioesterase domain-containing protein 38.53 5.61
DD8-14 DDB0188434 AAA ATPase domain-containing protein 75.20 6.32
DDB0229909a,c DDB0229909 Putative O-methyltransferase 25.84 5.57
DscEa,c DDB0215382 Discoidin II 28.56 6.65
EppA DDB0218473 Erk2-dependent phosphoprotein 45.29 6.19
JC1V2_0_00135c DDB0189346 Unknown 39.08 5.40
JC1V2_0_00564 DDB0189754 Esterase/lipase/thioesterase domain-containing 24.53 6.71
JC1V2_0_00789 DDB0202301 Putative SAM dependent methyltransferase 33.78 8.28
JC2V2_0_00793 DDB0168923 Dihydropteridine reductase 24.65 6.90
JC2V2_0_01412b,c DDB0217373 Similar to discoidin I, A chain 28.24 6.20
JC2V2_0_01572 DDB0217484 Unknown 99.02 4.98
JC3V2_0_00478c DDB0206195 Unknown 25.97 7.60
JC3V2_0_01972 DDB0204655 Lupus La protein 40.32 8.55
aProteins which showed significant differences (2.0 fold or higher) between the L. pneumophila Corby and L. hackeliae-specific phagosome.bProteins which appear only in the Legionella phagosome but not in the latex bead-containing phagosome (Gotthardt et al., 2006).cProteins which were identified in the Legionella-containing phagosome purified by magnetic immunoseparation (Urwyler et al., 2009).
Fig. 3. Functional categories of identified phagosome proteins. The phagosome proteome was categorized based on gene ontology
annotations in the D. discoideum database (http://www.dictybase.org). The proteins (n ¼ 157) fell into 14 functional categories. The
number of proteins detected for each category is indicated.
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508498
Specific changes of the phagosomal proteome during
infection with L. pneumophila Corby or L. hackeliae
To describe protein differences between theL. pneumophila Corby and L. hackeliae containingphagosomes as well as temporal changes in protein
composition, phagosome proteins were systematicallyidentified at 2, 4, and 6 h post-infection. The overallprogression of infection was monitored by fluorescencemicroscopy using rhodamine labelled bacteria. Thisanalysis revealed an almost constant number of infectedhost cells (36.5% SD 4.2) and intracellular bacteria

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 499
(1.1 SD 0.3 bacteria per cell) between 2 and 6 h post-infection. These observations indicate that host cellinvasion did not occur later than 2 h post-infection.High accuracy in the identification of the respectiveprotein composition was achieved by combining thedata of 3 independent experiments. Equal amounts oftotal phagosome protein were used for each time point,and the resulting fluorescent 2D gels were analyzed withthe Delta2D software program (Decodon). The analysis
Fig. 4. Dual channel images of the D. discoideum phagosomal prote
infection. (C) Pattern 6 h post-infection. Phagosome proteins from
labelled with Cy3 (green), while phagosome proteins from L. hacke
expressed proteins are highlighted and listed in the Tables 1 and 2.
spot, excluding background. The signal intensity of all spots on th
expression (L. pneumophila Corby phagosome/L. hackeliae phagoso
legend, the reader is referred to the web version of this article.)
protocol included spot detection and filtering, wholeimage warping on a reference gel, background subtrac-tion, and average gel creation. Fig. 4A–C presents agallery of dual colour 2-DE images (green fluorescentCy3 for L. pneumophila Corby and red fluorescent Cy5for L. hackeliae) exhibiting the differences between theL. pneumophila Corby and L. hackeliae phagosomes.Only significant changes in spot representation of 2.0-fold or higher were further analyzed. By using this
in patterns. (A) Pattern 2 h post-infection. (B) Pattern 4 h post-
D. discoideum cells infected with L. pneumophila Corby were
liae infections were labelled with Cy5 (red). All differentially
The % volume in Table 2 indicates the relative quantity of the
e gel is 100%. The ratio represents a fold change of protein
me). (For interpretation of the references to color in this figure

ARTICLE IN PRESS
Fig. 4. (Continued)
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508500
criterion we found that infection with L. pneumophila
Corby versus L. hackeliae resulted in 22, 25, and 26differentially regulated protein spots after 2, 4, and 6 h,respectively (Table 2).
The comparison after infection with both species at2 h post-infection revealed that L. pneumophila Corbyinduced higher protein expression levels for the cysteineprotease inhibitor (CpiA), Cofilin (CofA), a peptidaseM16 family protein (DDB0189202), superoxide dismu-tase (SodA), and several actin spots (Act). Stronglyreduced protein expression levels were observed for the14-3-3 protein (FttB), uridine phosphorylase (UdpB),and one of the 2 spots which were identified as Rho-GDP-dissociation inhibitor (RdiA) (Table 2).
The comparison of the phagosome proteome afterinfection with L. pneumophila Corby and L. hackeliae at4 h post-infection revealed the enrichment of numerousproteins like the ribosomal protein small subunit(Rps12), several actin spots (Act), and the discoidin-like protein (DDB0217373) in the L. pneumophila Corbycontaining phagosome. Interestingly, the RdiA and theFttB were strongly reduced in the pathogen containingphagosome, while a prominent presence of both proteinswas observed in the L. hackeliae-containing phagosome.
The comparison of the phagosome proteome afterinfection with both species at 6 h post-infection revealedthe enrichment of proteins belonging to the proteinbiosynthesis category (Efa1b, Moe1, ribosomal proteins

ARTICLE IN PRESS
Fig. 4. (Continued)
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 501
and asparagenyl-tRNA synthetase) and the cyto-skeleton category (AcpB, CofA, CoaA, Act) in theL. pneumophila Corby-containing phagosome. Smallprotein amounts but a significant difference betweenL. pneumophila Corby and L. hackeliae containingphagosomes were observed for glutamate-ammonialigase. As already described for 4 h post-infection, weobserved a strong reduction of the Rho-GDP-dissocia-tion inhibitor in phagosomes containing L. pneumophila
Corby.Taken together, this analysis clearly indicated distinct
phases in the maturation programme. The changes inprotein composition suggest that L. pneumophila Corbycontaining phagosomes exhibit a higher upregulation of
proteins compared to phagosomes containing theapathogenic species. Moreover, it is obvious that therecruitment of factors involved in cytoskeleton organi-sation and protein biosynthesis predominate in phago-somes isolated from L. pneumophila Corby-infected hostcells.
Actin degradation caused by L. pneumophila Corby
infection
It has been shown that entry of L. pneumophila intoD. discoideum is an actin-mediated process and thatL. pneumophila-containing phagosomes are transported

ARTICLE IN PRESS
Table 2. Phagosomal proteins significantly modulated during infection with L. pneumophila Corby and L. hackeliae.
Protein Function % volume L. pneumophila Corby % volume L. hackeliae Fold change
L. pneumophila Corby/L. hackeliae 2 h
CpiA Cysteine proteinase inhibitor 0.99 0.09 10.0
CofA Cofilin 0.11 0.01 10.0
DDB0189202 Peptidase M16 family protein 0.04 0.01 5.3
Act Actin 0.93 0.19 5.0
Act Actin 0.35 0.09 3.7
CorA Coronin 0.17 0.05 3.6
SodA Superoxide dismutase 0.68 0.19 3.6
Act Actin 0.64 0.19 3.4
Act Actin 0.16 0.05 3.2
Act Actin 0.97 0.32 3.0
Act Actin 2.01 0.71 2.8
NdkC Nucleoside diphosphate kinase 0.19 0.07 2.8
DDB0217373 Similar to discoidin I, A chain 0.08 0.03 2.7
FumH Fumarate hydratase 0.13 0.05 2.4
PkiA Similar to protein kinase C inhibitor 0.24 0.10 2.2
RdiA Rho GDP-dissociation inhibitor 0.54 0.25 2.1
EfbA Elongation factor 2 0.09 0.04 2.1
EfaA1 Elongation factor 1 alpha 1.29 3.06 0.4
FttB 14-3-3 protein 0.17 0.49 0.3
DDB0219748 Oxidoreductase activity 0.36 1.10 0.3
UdpB Uridine phosphorylase 0.01 0.04 0.1
RdiA Rho GDP-dissociation inhibitor 0.56 4.42 0.1
L. pneumophila Corby/L. hackeliae 4 h
CxeA Cytochrome c oxidase subunit V 1.00 0.45 2.2
PkiA Similar to protein kinase C inhibitor 0.23 0.11 2.2
HspE-1 Heat shock protein 0.004 0.0001 2.5
Rps12 Ribosomal Protein Small subunit 0.73 0.14 5.3
Act Actin 0.37 0.07 5.0
DDB0217373 Similar to discoidin I, A chain 0.07 0.01 5.0
CofB Cofilin 0.37 0.10 3.8
Act Actin 1.00 0.30 3.4
Act Actin 0.90 0.29 3.1
AxnA Annexin VII 0.32 0.13 2.4
Act Actin 0.14 0.06 2.3
NdkC Nucleoside diphosphate kinase 0.11 0.05 2.2
TubB Tubulin 0.04 0.02 2.2
AlrA Aldehyde reductase 0.42 0.19 2.2
NdkM Nucleoside diphosphate kinase 0.11 0.05 2.1
PrsA Phosphoribosyl pyrophosphate Synthetase 0.63 0.30 2.1
Act Actin 1.67 0.83 2.0
VatB Vacuolar H+ ATPase B subunit 0.08 0.16 0.5
MvpA Major vault protein A 0.05 0.10 0.5
Pdi2 Protein disulfide isomerase 0.10 0.21 0.5
AclA Component of Arp2/3 complex 0.12 0.26 0.4
FttB 14-3-3 protein 0.43 0.98 0.4
DDB0188166 Putative delta24sterol methyltransferase 0.02 0.04 0.4
RdiA Rho GDP-dissociation inhibitor 0.83 5.26 0.2
L. pneumophila Corby/L. hackeliae 6 h
GlnB Glutamate-ammonia ligase 0.03 0.001 25.0
AcpB Actin capping protein 0.06 0.006 10.0
CofA Cofilin 0.09 0.02 5.9
CdcD Cell division cycle protein 48 0.07 0.02 4.5
CoaA Coactosin 1.69 0.38 4.5
NdkM Nucleoside diphosphate kinase 0.12 0.03 4.5
Moe1 eIF-3, subunit 2 0.02 0.004 4.0
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508502

ARTICLE IN PRESS
Table 2. (continued )
Protein Function % volume L. pneumophila Corby % volume L. hackeliae Fold change
AspS1 Aspartyl-tRNA synthetase 0.02 0.005 3.3
Act Actin 0.19 0.06 3.3
CtxA Cortexillin I 0.05 0.02 2.9
VatA Vacuolar H+ ATPase B subunit 0.25 0.09 2.9
DDB0217373 Similar to discoidin I, A chain 0.18 0.07 2.6
Act Actin 0.74 0.28 2.6
AlrA Aldehyde reductase 0.68 0.26 2.6
Efa1b Elongation factor 1 beta 1.97 0.78 2.6
AcpA Actin capping protein 0.09 0.04 2.4
GlnB Glutamate-ammonia ligase 0.06 0.03 2.3
DscE Discoidin II 0.14 0.06 2.3
CorA Coronin 0.16 0.07 2.2
Rpl21 Ribosomal protein L11 0.27 0.12 2.2
RpsA 40S ribosomal protein SA 1.24 0.57 2.2
DDB0219748 Oxidoreductase activity 0.36 0.80 0.4
DDB0229909 Unknown function 0.002 0.01 0.2
RdiA Rho GDP-dissociation inhibitor 0.73 5.67 0.1
TubB Tubulin – 0.02 N
Pdi2 Protein disulfide isomerase – 0.24 N
The % volume indicates the relative quantity of the spot after subtraction of the background. The intensity of all spots on the gel sums up to 100%.
The ratio represents a fold change of protein expression (L. pneumophila Corby phagosome/L. hackeliae phagosome).
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 503
along microtubules (Lu and Clarke, 2005). Moreover, ithas been reported that Dictyostelium mutants lackingcertain actin-binding proteins are somewhat impaired inLegionella uptake and growth (Fajardo et al., 2004).Here, we examined the pathobiological changes of actinin the phagosome proteome after 2, 4, and 6 h ofinfection with L. pneumophila Corby and L. hackeliae,respectively. The comparison revealed additional actinspots in the L. pneumophila Corby phagosome pro-teome, which are not present in the L. hackeliae
phagosome proteome. The identified spots migratedfaster in the 2D SDS-PAGE than the 42-kDa actinspot, which suggests that they are degradation products(Fig. 5A). By using a specific D. discoideum anti-actinantibody, we demonstrated that the additional bandsindeed are actin fragments with a size of 36 kDa(Fig. 5B). We conclude that this degradation of actinis specific for the pathogen-containing phagosome.
Discussion
The success of L. pneumophila as pathogen dependslargely on its ability to avoid being killed in aphagolysosome (Skriwan et al., 2002; Steinert et al.,2007). D. discoideum, a well-investigated professionalphagocyte and a model host for Legionella, has beeninstrumental in providing new insights into phagosomeactivities and their regulation (Steinert and Heuner,2005; Chen et al., 2007a, b; de Felipe et al., 2008; Ragaz
et al., 2008). Exciting progress has been made by theanalysis of latex bead-containing Dictyostelium phago-somes which helped to define 3 maturation stages(Gotthardt et al., 2002). The first maturation stage ischaracterized by coronin and lysosomal glycoprotein(LmpB) acquisition. The second stage is characterizedby the transfer of lysosomal enzymes. The third stage ischaracterized by quantitative retrieval of hydrolasesfrom the phagolysosome and exocytosis of latexbeads. A subsequent proteomic study has been enligh-tening since almost 200 phagosomal proteins wereidentified and ordered by their temporal appearance(Gotthardt et al., 2006). Valuable insights intosubcellular vacuoles containing pathogenic microorgan-isms were also obtained by infection of mammalianphagocytes with Francisella tularensis, Mycobacterium
avium, Salmonella typhimurium, Listeria innocua,Rhodococcus equi, and Leishmania parasites (Sturgill-Koszycki et al., 1997; Mills and Finlay, 1998; Luhrmannand Haas, 2000; Fernandez-Mora et al., 2005; Kimaand Dunn, 2005). A recent proteome analysis ofLegionella-containing phagosomes purified by magneticimmuno-separation identified many known andnovel phagosome components including a number ofsmall GTPases or GTPase-associated proteins (Arf1,Rab1a, Rab1d, Rab7a, Rab8a, Rab14, RabC, RabGDI,RasG, RanA, Ran GAP, and Rho GDI). Moreover,the endosomal regulators Rab8 and Rab14 were foundto be present on phagosomes harbouring wild-type butnot Dot/Icm-deficient L. pneumophila (Urwyler et al.,2009).

ARTICLE IN PRESS
Fig. 5. Actin degradation caused by L. pneumophila Corby infection. (A) Actin spots on the 2D gels from L. pneumophila Corby
phagosome (left panel) and L. hackeliae phagosome (right panel). Arrows indicate additional spots, which migrated faster than
undegraded actin. (B) Western blot analysis of 1D SDS-PAGE demonstrated the appearance of degraded actin 2 h post-infection.
Lane 1: Isolated D. discoideum actin (control). Lane 2: L. pneumophila Corby phagosome fraction. Lane 3: L. hackeliae phagosome
fraction. Proteins were separated on SDS-polyacrylamide gel, blotted onto a nitrocellulose membrane and incubated with a specific
anti-actin-1 antibody.
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508504
To study the molecular processes that underly thedevelopment of an intracellular replicative niche, wehave developed an alternative preparative method forthe isolation of Legionella-containing phagosomeswhich may be applied to other pathogens as well. Ourmethod includes the loading of the endocytic compart-ments with colloidal iron, increasing the density ofmitochondria by a tetrazolium dye and the treatment ofthe postnuclear supernatant with Benzonase. Theseprocedures ensure the removal of nucleic acids andorganelles and improve the protein spot resolution after2D gel electrophoresis. The purity of the isolatedphagosomes after fractionation in an Opti-Prep gradientwas analysed by immunoblotting with specific organellemarkers and transmission electron microscopy. Theresults obtained with our method allowed to build atwo-dimensional phagosome proteome map that can beused to monitor L. pneumophila-induced changes inprotein composition. Using mass spectrometry andgenomic data analysis, we identified 157 phagosomeproteins, many of which have also been described forlatex bead-containing phagosomes and Legionella-con-taining phagosomes purified by magnetic immunosep-aration (Gotthardt et al., 2006; Urwyler et al., 2009). Acomparison of different and similar proteins after therespective phagosome preparations are highlighted inTable 1. Opposed to previous claims that the vacuolarH+-ATPase is not present in the phagosomal membranesurrounding L. pneumophila (Lu and Clarke, 2005), wehave detected vacuolar H+-ATPase A and B subunits.At the same time point, latex bead-containing phago-
somes similarly display numerous spots representingdifferent subunits of V-ATPase (Vat A, B, M and E)(Gotthardt et al., 2006). On the other hand, certain hostand bacterial factors were apparently below detectionlevel in the two-dimensional gel electrophoresis.
The phagosome proteins identified in this studybelong to a variety of functional categories. Proteinsinvolved in metabolism, protein biosynthesis as well ascytoskeleton and signal transduction make up the bulkof the Legionella phagosome proteome and agree wellwith the observed upregulation of their transcripts(Farbrother et al., 2006). The presence of elongationfactors, ribosomal proteins, t-RNA-synthetases as wellas proteins typically associated with ER is consistentwith the fact that the Legionella-containing phagosomeintercepts secretory vesicles from ER exit sites anddevelops into a compartment that shares many featureswith the rough ER (Machner and Isberg, 2006). More-over, our proteome data highlight the obvious relevanceof the ubiquitin–proteasome machinery. This view issupported by the upregulation of transcripts belongingto the ubiquitination machinery of D. discoideum afterinfection with Legionella wild type as compared to thedotA mutant (Farbrother et al., 2006) and also by theproteome analysis of Legionella-containing phagosomespurified by magnetic immunoseparation (Urwyler et al.,2009). Since the dotA mutant lacks a functional Dot/Icm type IV secretion system, respective Dot/Icm T4BSSsubstrates obviously play a role in this process. A recentpublication suggests that Legionella translocates an E3ubiquitin ligase that has 2 U-boxes. U-box 1 is critical to

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 505
the ubiquitin ligation and U-box 2 mediates interactionwith Clk1 (Cdc2-like kinase 1) (Kubori et al., 2008;Ivanov and Roy, 2009; Al-Khodor et al., 2008).
To determine how far protein expression patternsreally are a consequence of pathogen-induced events, wehave compared L. pneumophila Corby-containing pha-gosomes with phagosomes of L. hackeliae, a Legionella
species which is degraded by Dictyostelium. Kineticanalysis clearly showed that the L. pneumophila Corbyphagosome accumulated ER markers and ribosomalconstituents including several elongation factors. Thiscorrelates with the progression of infection as visualizedby confocal microscopy of living D. discoideum cells(Lu and Clarke, 2005). At the same time points, thedegradative L. hackeliae phagosome eliminated compo-nents of the protein biosynthesis machinery.
Lysosomal enzymes are initially synthesized as pre-cursors on the rough endoplasmic reticulum, a cellularcompartment which during Legionella infection associ-ates with the L. pneumophila-specific phagosome.L. pneumophila Corby may inhibit proteolytic proces-sing events which are required for proper sorting andmaturing of lysosomal enzymes by inducing theupregulation of the Dictyostelium cysteine proteaseinhibitor CpiA expression (Richardson et al., 1988;North et al., 1996). The observation that Legionella
phagosomes avoid the phagolysosome formation only atan early stage of infection supports our hypothesis sincethe increase of the cysteine protease inhibitor was onlyobserved until the 2-h time point post-infection.Additionally, it is worth mentioning that L. pneumophila
Corby induces a 3.6-fold increase of superoxide dis-mutase SodA when compared to L. hackeliae containingphagosomes. This may contribute to the evasion ofL. pneumophila Corby from the bactericidal system byreactive oxygen species (ROS) as proposed earlier(Harada et al., 2007).
One of the most obvious differences in our proteomicanalysis is the prominent abundance of the Rho-GTPaseinhibitor RhoGDI on L. hackeliae-containing phago-somes versus its minimal presence on phagosomescontaining L. pneumophila Corby at all time pointstested. Rho-GTPases are small GTPases of the Rassuperfamily, which act as molecular switches in theregulation of many cellular processes by cycling betweenthe inactive GDP- and the active GTP-bound form(D’Souza-Schorey et al., 1997; Duhon and Cardelli,2002). RhoGDIs are able to inhibit their cognateGTPases by several modes of action like preventingGTP-for-GDP exchange and by extracting their GTPasepartners from the membranes (Dovas and Couchman,2005). These inhibitors are ubiquitously expressed ineukaryotes including humans, and RhoGDI ofD. discoideum was shown to interact with 6 of 19 knownDictyostelium GTPases (Vlahou and Rivero, 2006).RhoGDI deficient Dictyostelium cells showed a striking
defect in cytokinesis with the phenotype of multi-nucleated cells, prominent lamellar actin-rich structures,and giant vacuoles. Our proteomic data suggest acentral importance for this protein in the regulation ofthe host response to infections by phagosomal patho-gens that is abrogated by the virulent L. pneumohila
Corby strain.During bacteria or latex beads uptake by Dictyoste-
lium, actin is recruited to the nascent phagosome. TheArp2/3 complex, responsible for nucleation of actin andvarious actin-interacting proteins including coronin, istransiently enriched on the nascent phagosome. Afterengulfment of the particle, actin and associated proteinsdissemble, however, become recruited again at a laterstages of the endocytic transit (Lu and Clarke, 2005).These findings are consistent with our proteome datawhich revealed that the Arp2/3 complex, actin, actin-binding proteins, coronin, and others are present inLegionella-containing phagosomes. Since actin as-sembled by phagosomes can provide tracks for lyso-somes to move vectorially toward them, it is aninteresting observation that the pathogenic L. pneumo-
phila Corby infection causes degradation of actin in thephagosomal fraction. The fact that phagosomes con-taining non-replicative L. hackeliae do not displaydegraded actin, highlights the importance of this finding.Whether the cAMP-signaling system of D. discoideum
plays a role in phagosome actin assembly and onphagosome maturation, as described for macrophagesinfected with mycobacteria, remains to be established(Kalamidas et al., 2006). However, since elevation ofintracellular cAMP can reverse the ability of macro-phages to inhibit Legionella growth, we hypothesize thatthe same mechanism also occurs in Dictyostelium
(Egawa et al., 1992).The appearance of the actin-binding protein coronin
on the phagosome gives rise to another hypothesis.Coronin interacts with cytosolic NADPH oxidasecomponents p40phox and p67phox and is known to beinvolved in their translocation to the phagosome (Kaul,2008). Previous observations that Legionella replicateswell in Dictyostelium mutants lacking coronin may thusbe due to improper superoxide generation at thephagosome (Solomon et al., 2000; Fajardo et al.,2004). Moreover, in neutrophils phosphorylatedp47phox protein has been shown to allow appropriatelocalization of protein kinase C to coronin, leading tocoronin phosphorylation. The phosphorylated coroninsolubilizes, and the subsequent removal of the coronincoat facilitates phagolysosome fusion (Grogan et al.,1997; Kaul, 2008). Since we observed coronin as well asprotein kinase C inhibitors on Legionella phagosomes,we may have a first indication how the Legionella-specific inhibition of the phagolysosome fusion may beregulated. Further support comes from a recentpublication (Harada et al., 2007) that showed that the

ARTICLE IN PRESS
Fig. 6. Graphic representation of the Legionella-containing phagosome including some of the proteins identified in this study. After
the actin-mediated uptake, L. pneumophila avoids endosomal fusion and intersepts ER-derived vesicles that normally traffic to the
Golgi. The newly internalized phagosome is rapidly transported about the cell on microtubules. The proteasome and ubiquitinated
proteins are recruited to the Legionella-containing phagosome and are required for efficient bacterial replication.
O. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508506
production of reactive oxygen species (ROS) by macro-phages infected with a L. pneumophila wild-type strainwas suppressed. The results suggest that the interferenceis due to a failure of NADPH oxidase activationthrough inhibition of p47phox recruitment to phago-somes containing the bacteria (Harada et al., 2007).Further research is needed to reveal the details of thisand other proposed mechanisms (Fig. 6). Nevertheless,this proteomic analysis of a pathogen-containingphagosome will surely open new avenues for futureprojects.
Acknowledgements
The work was supported by grants from the DeutscheForschungsgemeinschaft (STE 838/3-4) to M.S., to A.H(SFB 670), and from the BMBF (PTJ-Bio/0313807A).We cordially thank Dirk Albrecht for mass spectro-metry analyses, Ralf Schnabel and Thomas Zobel forsupport in fluorescence microscopy, and MichaelSchleicher for helpful discussions.
References
Al-Khodor, S., Price, C.T., Habyarimana, F., Kalia, A., Abu
Kwaik, Y., 2008. A Dot/Icm-translocated ankyrin protein
of Legionella pneumophila is required for intracellular
proliferation within human macrophages and protozoa.
Mol. Microbiol. 70, 908–923.
Cazalet, C., Rusniok, C., Bruggemann, H., Zidane, N.,
Magnier, A., Ma, L., Tichit, M., Jarraud, S., Bouchier,
C., Vandenesch, F., Kunst, F., Etienne, J., Glaser, P.,
Buchrieser, C., 2004. Evidence in the Legionella
pneumophila genome for exploitation of host cell
functions and high genome plasticity. Nat. Genet. 36,
1165–1173.
Chen, J., Reyes, M., Clarke, M., Shuman, H.A., 2007a. Host
cell-dependent secretion and translocation of the LepA and
LepB effectors of Legionella pneumophila. Cell Microbiol.
9, 1660–1671.
Chen, G., Zhuchenko, O., Kuspa, A., 2007b. Immune-like
phagocyte activity in the social amoeba. Science 317,
678–681.
Chisholm, R.L., Gaudet, P., Just, E.M., Pilcher, K.E., Fey, P.,
Merchant, S.N., Kibbe, W.A., 2006. dictyBase, the model
organism database for Dictyostelium discoideum. Nucleic
Acids Res. 34, D423–427.

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508 507
Clarke, M., Bazari, W.L., Kayman, S.C., 1980. Isolation and
properties of calmodulin from Dictyostelium discoideum.
J. Bacteriol. 141, 397–400.
de Felipe, K.S., Glover, R.T., Charpentier, X., Anderson,
O.R., Reyes, M., Pericone, C.D., Shuman, H.A., 2008.
Legionella eukaryotic-like type IV substrates interfere with
organelle trafficking. PLoS Pathog. 4, e1000117.
Derre, I., Isberg, R.R., 2005. LidA, a translocated substrate of
the Legionella pneumophila type IV secretion system,
interferes with the early secretory pathway. Infect. Immun.
73, 4370–4380.
Dovas, A., Couchman, J.R., 2005. RhoGDI: multiple func-
tions in the regulation of Rho family GTPase activities.
Biochem. J. 390, 1–9.
D’Souza-Schorey, C., Boshans, R.L., McDonough, M., Stahl,
P.D., Van Aelst, L., 1997. A role for POR1, a Rac1-
interacting protein, in ARF6-mediated cytoskeletal rear-
rangements. Embo J. 16, 5445–5454.
Duhon, D., Cardelli, J., 2002. The regulation of phagosome
maturation in Dictyostelium. J. Muscle Res. Cell Motil. 23,
803–808.
Edelstein, P.H., 1981. Improved semiselective medium for
isolation of Legionella pneumophila from contaminated
clinical and environmental specimens. J. Clin. Microbiol.
14, 298–303.
Egawa, K., Klein, T.W., Yamamoto, Y., Newton, C.A.,
Friedman, H., 1992. Cyclic AMP inhibition of lipopoly-
saccharide-induced restriction of Legionella pneumophila
growth in macrophage cultures. Infect. Immun. 60,
1936–1940.
Eymann, C., Dreisbach, A., Albrecht, D., Bernhardt, J.,
Becher, D., Gentner, S., Tam le, T., Buttner, K., Buurman,
G., Scharf, C., Venz, S., Volker, U., Hecker, M., 2004. A
comprehensive proteome map of growing Bacillus subtilis
cells. Proteomics 4, 2849–2876.
Fajardo, M., Schleicher, M., Noegel, A., Bozzaro, S., Kill-
inger, S., Heuner, K., Hacker, J., Steinert, M., 2004.
Calnexin, calreticulin and cytoskeleton-associated proteins
modulate uptake and growth of Legionella pneumophila in
Dictyostelium discoideum. Microbiology 150, 2825–2835.
Farbrother, P., Wagner, C., Na, J., Tunggal, B., Morio, T.,
Urushihara, H., Tanaka, Y., Schleicher, M., Steinert, M.,
Eichinger, L., 2006. Dictyostelium transcriptional host cell
response upon infection with Legionella. Cell Microbiol. 8,
438–456.
Fernandez-Mora, E., Polidori, M., Luhrmann, A., Schaible,
U.E., Haas, A., 2005. Maturation of Rhodococcus equi-
containing vacuoles is arrested after completion of the early
endosome stage. Traffic 6, 635–653.
Goldsbrough, A.P., Bulleid, N.J., Freedman, R.B., Flavell,
R.B., 1989. Conformational differences between two wheat
(Triticum aestivum) ‘high-molecular-weight’ glutenin sub-
units are due to a short region containing six amino acid
differences. Biochem. J. 263, 837–842.
Gotthardt, D., Warnatz, H.J., Henschel, O., Bruckert, F.,
Schleicher, M., Soldati, T., 2002. High-resolution dissection
of phagosome maturation reveals distinct membrane
trafficking phases. Mol. Biol. Cell 13, 3508–3520.
Gotthardt, D., Blancheteau, V., Bosserhoff, A., Ruppert, T.,
Delorenzi, M., Soldati, T., 2006. Proteomics fingerprinting
of phagosome maturation and evidence for the role
of a Galpha during uptake. Mol. Cell Proteom. 5,
2228–2243.
Grogan, A., Reeves, E., Keep, N., Wientjes, F., Totty, N.F.,
Burlingame, A.L., Hsuan, J.J., Segal, A.W., 1997. Cytosolic
phox proteins interact with and regulate the assembly of
coronin in neutrophils. J. Cell Sci. 110, 3071–3081.
Hagele, S., Kohler, R., Merkert, H., Schleicher, M., Hacker, J.,
Steinert, M., 2000. Dictyostelium discoideum: a new host
model system for intracellular pathogens of the genus
Legionella. Cell Microbiol. 2, 165–171.
Harada, T., Miyake, M., Imai, Y., 2007. Evasion of Legionella
pneumophila from the bactericidal system by reactive
oxygen species (ROS) in macrophages. Microbiol. Immu-
nol. 51, 1161–1170.
Hilbi, H., Weber, S.S., Ragaz, C., Nyfeler, Y., Urwyler, S.,
2007. Environmental predators as models for bacterial
pathogenesis. Environ. Microbiol. 9, 563–575.
Jepras, R.I., Fitzgeorge, R.B., Baskerville, A., 1985. A
comparison of virulence of two strains of Legionella
pneumophila based on experimental aerosol infection of
guinea-pigs. J. Hyg. (Lond.) 95, 29–38.
Ivanov, S.S., Roy, C.R., 2009. Modulation of ubiquitin
dynamics and suppression of DALIS formation by the
Legionella pneumophila Dot/Icm system. Cell Microbiol.
11, 261–278.
Kalamidas, S.A., Kuehnel, M.P., Peyron, P., Rybin, V.,
Rauch, S., Kotoulas, O.B., Houslay, M., Hemmings,
B.A., Gutierrez, M.G., Anes, E., Griffiths, G., 2006. cAMP
synthesis and degradation by phagosomes regulate actin
assembly and fusion events: consequences for mycobacter-
ia. J. Cell Sci. 119, 3686–3694.
Kaul, D., 2008. Coronin-1A epigenomics governs mycobacte-
rial persistence in tuberculosis. FEMS Microbiol. Lett. 278,
10–14.
Kima, P.E., Dunn, W., 2005. Exploiting calnexin expression
on phagosomes to isolate Leishmania parasitophorous
vacuoles. Microb. Pathog. 38, 139–145.
Kubori, T., Hyakutake, A., Nagai, H., 2008. Legionella
translocates an E3 ubiquitin ligase that has multiple
U-boxes with distinct functions. Mol. Microbiol. 67,
1307–1319.
Li, Z., Solomon, J.M., Isberg, R.R., 2005. Dictyostelium
discoideum strains lacking the RtoA protein are defective
for maturation of the Legionella pneumophila replication
vacuole. Cell Microbiol. 7, 431–442.
Lu, H., Clarke, M., 2005. Dynamic properties of Legionella-
containing phagosomes in Dictyostelium amoebae. Cell
Microbiol. 7, 995–1007.
Luhrmann, A., Haas, A., 2000. A method to purify bacteria-
containing phagosomes from infected macrophages. Meth-
ods Cell Sci. 22, 329–341.
Machner, M.P., Isberg, R.R., 2006. Targeting of host Rab
GTPase function by the intravacuolar pathogen Legionella
pneumophila. Dev. Cell. 11, 47–56.
Mills, S.D., Finlay, B.B., 1998. Isolation and characterization
of Salmonella typhimurium and Yersinia pseudotuberculosis-
containing phagosomes from infected mouse macrophages:
Y. pseudotuberculosis traffics to terminal lysosomes where
they are degraded. Eur. J. Cell. Biol. 77, 35–47.

ARTICLE IN PRESSO. Shevchuk et al. / International Journal of Medical Microbiology 299 (2009) 489–508508
Molmeret, M., Horn, M., Wagner, M., Santic, M., Abu
Kwaik, Y., 2005. Amoebae as training grounds for
intracellular bacterial pathogens. Appl. Environ. Micro-
biol. 71, 20–28.
Murata, T., Delprato, A., Ingmundson, A., Toomre, D.K.,
Lambright, D.G., Roy, C.R., 2006. The Legionella pneu-
mophila effector protein DrrA is a Rab1 guanine nucleo-
tide-exchange factor. Nat. Cell Biol. 8, 971–977.
Nagai, H., Kagan, J.C., Zhu, X., Kahn, R.A., Roy, C.R.,
2002. A bacterial guanine nucleotide exchange factor
activates ARF on Legionella phagosomes. Science 295,
679–682.
North, M.J., Nicol, K., Sands, T.W., Cotter, D.A., 1996.
Acid-activatable cysteine proteinases in the cellular slime
mold Dictyostelium discoideum. J. Biol. Chem. 271,
14462–14467.
Ott, M., Messner, P., Heesemann, J., Marre, R., Hacker, J.,
1991. Temperature-dependent expression of flagella in
Legionella. J. Gen. Microbiol. 137, 1955–1961.
Ragaz, C., Pietsch, H., Urwyler, S., Tiaden, A., Weber, S.S.,
Hilbi, H., 2008. The Legionella pneumophila phosphatidyl-
inositol-4 phosphate-binding type IV substrate SidC
recruits endoplasmic reticulum vesicles to a replication-
permissive vacuole. Cell Microbiol. 10, 2416–2433.
Richardson, J.M., Woychik, N.A., Ebert, D.L., Dimond,
R.L., Cardelli, J.A., 1988. Inhibition of early but not
late proteolytic processing events leads to the missorting
and oversecretion of precursor forms of lysosomal
enzymes in Dictyostelium discoideum. J. Cell Biol. 107,
2097–2107.
Rodriguez-Paris, J.M., Nolta, K.V., Steck, T.L., 1993.
Characterization of lysosomes isolated from Dictyostelium
discoideum by magnetic fractionation. J. Biol. Chem. 268,
9110–9116.
Skriwan, C., Fajardo, M., Hagele, S., Horn, M., Wagner, M.,
Michel, R., Krohne, G., Schleicher, M., Hacker, J.,
Steinert, M., 2002. Various bacterial pathogens and
symbionts infect the amoeba Dictyostelium discoideum.
Int. J. Med. Microbiol. 291, 615–624.
Solomon, J.M., Rupper, A., Cardelli, J.A., Isberg, R.R., 2000.
Intracellular growth of Legionella pneumophila in Dictyo-
stelium discoideum, a system for genetic analysis of host-
pathogen interactions. Infect. Immun. 68, 2939–2947.
Steinert, M., Heuner, K., 2005. Dictyostelium as host model
for pathogenesis. Cell. Microbiol. 7, 307–314.
Steinert, M., Heuner, K., Buchrieser, C., Albert-Weissenbe-
rger, C., Glockner, G., 2007. Legionella pathogenicity:
genome structure, regulatory networks and the host cell
response. Int. J. Med. Microbiol. 297, 577–587.
Sturgill-Koszycki, S., Haddix, P.L., Russell, D.G., 1997. The
interaction between Mycobacterium and the macrophage
analyed by two-dimensional poly-acrylamide gel electro-
phoresis. Electrophoresis 18, 2558–2565.
Tilney, L.G., Harb, O.S., Connelly, P.S., Robinson, C.G.,
Roy, C.R., 2001. How the parasitic bacterium Legionella
pneumophila modifies its phagosome and transforms it into
rough ER: implications for conversion of plasma mem-
brane to the ER membrane. J. Cell Sci. 114, 4637–4650.
Towbin, H., Staehelin, T., Gordon, J., 1979. Electrophoretic
transfer of proteins from polyacrylamide gels to nitrocellu-
lose sheets: procedure and some applications. Proc. Natl.
Acad. Sci. USA 76, 4350–4354.
Unal, C., Steinert, M., 2006. Dictyostelium discoideum as a
model to study host–pathogen interactions. Methods Mol.
Biol. 346, 507–515.
Urwyler, S., Nyfeler, Y., Ragaz, C., Lee, H., Mueller, L.N.,
Aebersold, R., Hilbi, H., 2009. Proteome analysis of
Legionella vacuoles purified by magnetic immuno-separa-
tion reveals secretory and endosomal GTPases. Traffic 10,
76–87.
Weber, S.S., Ragaz, C., Reus, K., Nyfeler, Y., Hilbi, H., 2006.
Legionella pneumophila exploits PI(4)P to anchor secreted
effector proteins to the replicative vacuole. PLoS Pathog. 2,
e46.
Copyright © 2022 FDOKUMEN




















