
Protection of Penaeus monodon against white spot syndrome virus by inactivated vaccine with herbal...
10
Protection of Penaeus monodon against white spot syndrome virus by inactivated vaccine with herbal immunostimulants Aiyakani Yogeeswaran, Subramanian Velmurugan, Stanislas Mary Josephine Punitha, Mariavincent Michael Babu, Thangaswamy Selvaraj, Thangamani Kumaran, Thavasimuthu Citarasu * Centre for Marine Science and Technology, Manonmaniam Sundaranar University, Rajakkamangalam, Kanyakumari District, Tamilnadu 629502, India article info Article history: Received 17 July 2011 Received in revised form 25 February 2012 Accepted 28 February 2012 Available online xxx Keywords: Immunostimulants Vaccine Penaeus monodon White spot syndrome virus abstract To improve the immune response in tiger shrimp Penaeus monodon against WSSV infection, juveniles (350 10 mg) were vaccinated with formalin-inactivated WSSV and fed with herbal immunostimulants. The methanolic extracts of herbal immunostimulants such as Acalypha indica, Cynodon dactylon, Picrorrhiza kurrooa, Withania somnifera and Zingiber officinalis were incorporated in formulated diets at different concentrations; 250 (ED 1 ), 500 (ED 2 ), 1000 (ED 3 ) and 2000 (ED 4 ) mg kg 1 of feed and fed for 60 days after vaccination. After 30 and 60 days intervals of feeding, the shrimps were challenged with WSSV, which were isolated and propagated from the infected crustaceans. The shrimps fed with control diets (C 1 ) succumbed to death within 5 days after WSSV challenge, when no vaccination and immunostimulations were given. The other control groups (C 2 and C 3 ) had slight improvements in all parameters including survival. The percentage survival was significantly (P < 0.05) increased to 30, 50 and 60% in the ED 2 , ED 3 and ED 4 diets respectively after 60 days challenging. The better haematological, biochemical and immunological parameters were also found in the herbal extracts supplemented diets fed vaccinated shrimps. The present study revealed that the combined effect of immunostimulation and vaccination helped to boost the immune system against WSSV infection and hence this application can be adopted for shrimp culture. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction White spot syndrome virus (WSSV) is an economically signifi- cant shrimp virus, which causes high mortalities and severe damages to shrimp culture. The clinical symptom of the affected shrimp bears white spots in the exoskeleton and epidermis [1]. The current disease treatment protocols are non-effective, cost effective and generate environmental hazards [2]. Considering this potential threat of disease on one hand and the environmental issues on the other hand, the disease management aspects should concentrate on eco-friendly with biotechnological methods like the development of rapid procedures for detection of diseases, prophylaxis (Vaccines, immunostimulants, bioremedia- tion of environment including the methods of administration) and disease treatment protocols [3]. To develop the alternative practices for disease management in aquaculture, attention should be diverted to find novel drugs, especially from plant sources. Citarasu et al. [4] studied antibacterial, antiviral, immunostimulant and anti- stress effect of herbal products that significantly influenced shrimp aquaculture. Prevention and control of diseases are now the priority for the revival of aquaculture industry. Within the past decade, intensification of the shrimp production, based on progress in zootechnology, has increased but with a little corresponding increase in scientific knowledge of shrimp physiology. Within this field, shrimp immunology is a key element in establishing strate- gies for the control of diseases in shrimp aquaculture [5]. The use of immunostimulants, in addition to chemotherapeutic agents and vaccines, has been widely accepted by the fish farmers. Further, immunostimulation and vaccination of shrimp with inac- tivated Vibrio spp. have been reported to provide some protection [6]. In vivo experiments with Marsupenaeus japonicus demonstrated the presence of a quasi-immune response when survivors of both natural and experimental WSSV infected shrimps were rechal- lenged with WSSV. Plasma from the surviving infected shrimp could neutralize WSSV from 20 days up to 2 months after infection [7]. Recently an interesting phenomenon, called quasi-immune response, has been reported against WSSV in M. japonicus [5]. The resistance against WSSV appeared 3 weeks after the virus exposure and was maintained for about 1 month, which was endorsed by the appearance of virus neutralizing factor(s) in the haemolymph of “immunized” kuruma shrimp [7]. * Corresponding author. Tel./fax: þ91 4652 253078. E-mail addresses: [email protected], [email protected] (T. Citarasu). Contents lists available at SciVerse ScienceDirect Fish & Shellfish Immunology journal homepage: www.elsevier.com/locate/fsi 1050-4648/$ e see front matter Ó 2012 Elsevier Ltd. All rights reserved. doi:10.1016/j.fsi.2012.02.029 Fish & Shellfish Immunology xxx (2012) 1e10 Please cite this article in press as: Yogeeswaran A, et al., Protection of Penaeus monodon against white spot syndrome virus by inactivated vaccine with herbal immunostimulants, Fish & Shellfish Immunology (2012), doi:10.1016/j.fsi.2012.02.029
Transcript of Protection of Penaeus monodon against white spot syndrome virus by inactivated vaccine with herbal...

at SciVerse ScienceDirect
Fish & Shellfish Immunology xxx (2012) 1e10
Contents lists available
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate / fs i
Protection of Penaeus monodon against white spot syndrome virus by inactivatedvaccine with herbal immunostimulants
Aiyakani Yogeeswaran, Subramanian Velmurugan, Stanislas Mary Josephine Punitha,Mariavincent Michael Babu, Thangaswamy Selvaraj, Thangamani Kumaran, Thavasimuthu Citarasu*
Centre for Marine Science and Technology, Manonmaniam Sundaranar University, Rajakkamangalam, Kanyakumari District, Tamilnadu 629502, India
a r t i c l e i n f o
Article history:Received 17 July 2011Received in revised form25 February 2012Accepted 28 February 2012Available online xxx
Keywords:ImmunostimulantsVaccinePenaeus monodonWhite spot syndrome virus
* Corresponding author. Tel./fax: þ91 4652 253078E-mail addresses: [email protected], citarasu@m
1050-4648/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.fsi.2012.02.029
Please cite this article in press as: Yogeeswvaccine with herbal immunostimulants, Fish
a b s t r a c t
To improve the immune response in tiger shrimp Penaeus monodon against WSSV infection, juveniles(350 � 10 mg) were vaccinated with formalin-inactivated WSSV and fed with herbal immunostimulants.Themethanolic extracts of herbal immunostimulants such asAcalypha indica, Cynodon dactylon, Picrorrhizakurrooa, Withania somnifera and Zingiber officinalis were incorporated in formulated diets at differentconcentrations; 250 (ED1), 500 (ED2), 1000 (ED3) and 2000 (ED4) mg kg�1 of feed and fed for 60 days aftervaccination. After 30 and 60 days intervals of feeding, the shrimpswere challengedwithWSSV,whichwereisolated and propagated from the infected crustaceans. The shrimps fedwith control diets (C1) succumbedto death within 5 days after WSSV challenge, when no vaccination and immunostimulations were given.The other control groups (C2 and C3) had slight improvements in all parameters including survival. Thepercentage survival was significantly (P < 0.05) increased to 30, 50 and 60% in the ED2, ED3 and ED4 dietsrespectively after 60 days challenging. The better haematological, biochemical and immunologicalparameterswere also found in the herbal extracts supplemented diets fed vaccinated shrimps. The presentstudy revealed that the combined effect of immunostimulation and vaccination helped to boost theimmune system against WSSV infection and hence this application can be adopted for shrimp culture.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
White spot syndrome virus (WSSV) is an economically signifi-cant shrimp virus, which causes high mortalities and severedamages to shrimp culture. The clinical symptom of the affectedshrimp bears white spots in the exoskeleton and epidermis [1]. Thecurrent disease treatment protocols are non-effective, cost effectiveand generate environmental hazards [2].
Considering this potential threat of disease on one hand and theenvironmental issues on the other hand, the disease managementaspects should concentrate on eco-friendly with biotechnologicalmethods like the development of rapid procedures for detection ofdiseases, prophylaxis (Vaccines, immunostimulants, bioremedia-tion of environment including the methods of administration) anddisease treatment protocols [3]. To develop the alternative practicesfor disease management in aquaculture, attention should bediverted to find novel drugs, especially from plant sources. Citarasuet al. [4] studied antibacterial, antiviral, immunostimulant and anti-stress effect of herbal products that significantly influenced shrimp
.suniv.ac.in (T. Citarasu).
All rights reserved.
aran A, et al., Protection of P& Shellfish Immunology (20
aquaculture. Prevention and control of diseases are now thepriority for the revival of aquaculture industry. Within the pastdecade, intensification of the shrimp production, based on progressin zootechnology, has increased but with a little correspondingincrease in scientific knowledge of shrimp physiology. Within thisfield, shrimp immunology is a key element in establishing strate-gies for the control of diseases in shrimp aquaculture [5].
The use of immunostimulants, in addition to chemotherapeuticagents and vaccines, has been widely accepted by the fish farmers.Further, immunostimulation and vaccination of shrimp with inac-tivated Vibrio spp. have been reported to provide some protection[6]. In vivo experimentswithMarsupenaeus japonicus demonstratedthe presence of a quasi-immune response when survivors of bothnatural and experimental WSSV infected shrimps were rechal-lenged with WSSV. Plasma from the surviving infected shrimpcould neutralize WSSV from 20 days up to 2 months after infection[7]. Recently an interesting phenomenon, called quasi-immuneresponse, has been reported against WSSV in M. japonicus [5]. Theresistance against WSSV appeared 3 weeks after the virus exposureand was maintained for about 1 month, which was endorsed by theappearance of virus neutralizing factor(s) in the haemolymph of“immunized” kuruma shrimp [7].
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e102
Plants are the storehouses and sources of safer and cheaperchemicals. The natural plant products have been reported to havevarious activities like anti-stress, growth promotor, appetizer, tonic,immunostimulant, aphrodisiac and antimicrobial in finfish and shell-fish larviculture due to the active principle natures such as alkaloids,flavonoids, pigments, phenolics, terpenoids, steroids and essential oils[2]. The present study focuses on screening of antiviral and immu-nostimulant active principles from the selected herbals against WSSVinfection and proves the combined effect of vaccination and immu-nostimulation on Penaeus monodon to enhance the disease resistant.
2. Materials and methods
2.1. Herbal extracts and in vitro antiviral activity
Five herbal extracts such as Acalypha indica, Cynodon dactylon,Withania somnifera, Zingiber officinalis and Picrorrhiza kurrooahaving antiviral and immunostimulant characteristics wereselected following Nadkarni [8] and Citarasu [2]. These wereextracted serially with hexane, ethyl acetate, methanol and distilledwater. Aqueous extracts were concentrated using lyophilizer andstored at 4 �C. The in vitro antiviral activity was performedfollowing the method of Balasubramanian et al. [9] using differentaqueous extracts of all herbals. Among the different solventextractions, methanolic extracts effectively suppressed the WSSVreplication and this was selected for further studies.
2.2. Experimental diets preparation
Selection, formulation and preparation of basal diets were doneaccording to the method described by Boonyaratpalin [10]. Fourexperimental diets were prepared by adding equal proportion ofthe five methanolic extracts and they were mixed with the basalingredients at the concentration of 250 (ED1), 500 (ED2) 1000 (ED3)and 2000 (ED4) mg kg�1 diets.
2.3. Experimental setup and vaccine preparation
Healthy P. monodon juveniles weighing about 350 � 10 mgcollected from a local shrimp farm at Thamaraikulam, Tamil Nadu,
Control-1 (C1)
Control-1 (C2)
Control-1 (C3)
Experimental Diet-1 (ED1)
Experimental Diet-2 (ED2)
Study Groups
Experimental Diet-3 (ED3)
Non
He
He(2
He(5
He(1
Experimental Diet-4 (ED4)
He(2
Fig. 1. Experimental setup for different treatment groups tow
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
India were acclimatized in the laboratory condition for five days.They were stocked in a FRP tank (5000 l capacity) in the laboratory.Uniform size of P. monodonwas selected from the stock culture andtransferred into individual experimental fibre tanks of 100 lcapacity. Four experimental groups, ED1, ED2, ED3, ED4, and threecontrol groups, C1 (no herbal extracts þ no vaccination), C2 (noherbal extracts þ with vaccination) and C3 (only herbal extractsenriched feed and no vaccination) were maintained (Fig. 1). Trip-licate culture (n ¼ 30 � 3 ¼ 90) was maintained in each group withcontinuous flow-through water and constant aeration system. Thewater quality parameters such as temperature (27� 1.0 �C), salinity(28 � 1.5&) and pH (8.2 � 0.1) were maintained daily.
The infected haemolymph viral suspension was diluted with 10fold of 1� Phosphate Buffered Saline solution (PBS) and inactivatedby 0.5% V/v formalin for 10min at 25 �C. Formalinwas removed twotimes by centrifugation at 30,000� g for 1 h at 4 �C. The pellet wasre-suspended in PBS to furnish the viral concentration of one tenthof the original viral stock solution and used as a vaccine [11].
2.4. Vaccination and feeding
P. monodon juveniles of the experimental and control (C2) groupswere vaccinated through intra muscularly (IM) in the secondabdominal segment with 25 ml of formalin-inactivated WSSV(300 mg of total protein) per animal. The control C1 group of shrimpswas injected with diluted un-infected shrimp haemolymph simi-larly treated with formalin. The control C1 group was fed with thediet having no herbal antiviral and immunostimulant active prin-ciples. The other groups (C2 and ED1eED4) were fed with the diethaving herbal active principles. The shrimpswere fed thrice a day at8.00, 13.00 and 18.00 h on 10% of the body weight basis. Uneatenfood and waste materials were removed before feeding.
2.5. WSSV challenge
On the 30th day of post-vaccination (dpv), each group ofexperimental shrimps was divided into two subgroups. The firstsubgroup (45 shrimps in each diet) was challenged by IntraMuscular (IM) injection with 15 ml of WSSV infected haemolymphhaving a protein concentration of 300 mg. The second group was
Cum
ulat
ive
Mor
talit
y, h
aem
atol
ogic
al &
im
mun
olog
ical
impr
ovem
ent
stud
ies
afte
r 3
0 &
60
dpv
rmal diet (No vaccination & o herbal immunostimulant)
rbal immunostimulant diets(No vaccination)
rbal immunostimulant diets 50 mg kg-1) + Vaccination
rbal immunostimulant diets 00 mg kg-1) + Vaccination
rbal immunostimulant diets 000 mg kg-1)+ Vaccination
Normal diet (Vaccination only)
rbal immunostimulant diets 000 mg kg-1)+ Vaccination
ards WSSV inactivated vaccine with immunostimulants.
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029
A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e10 3
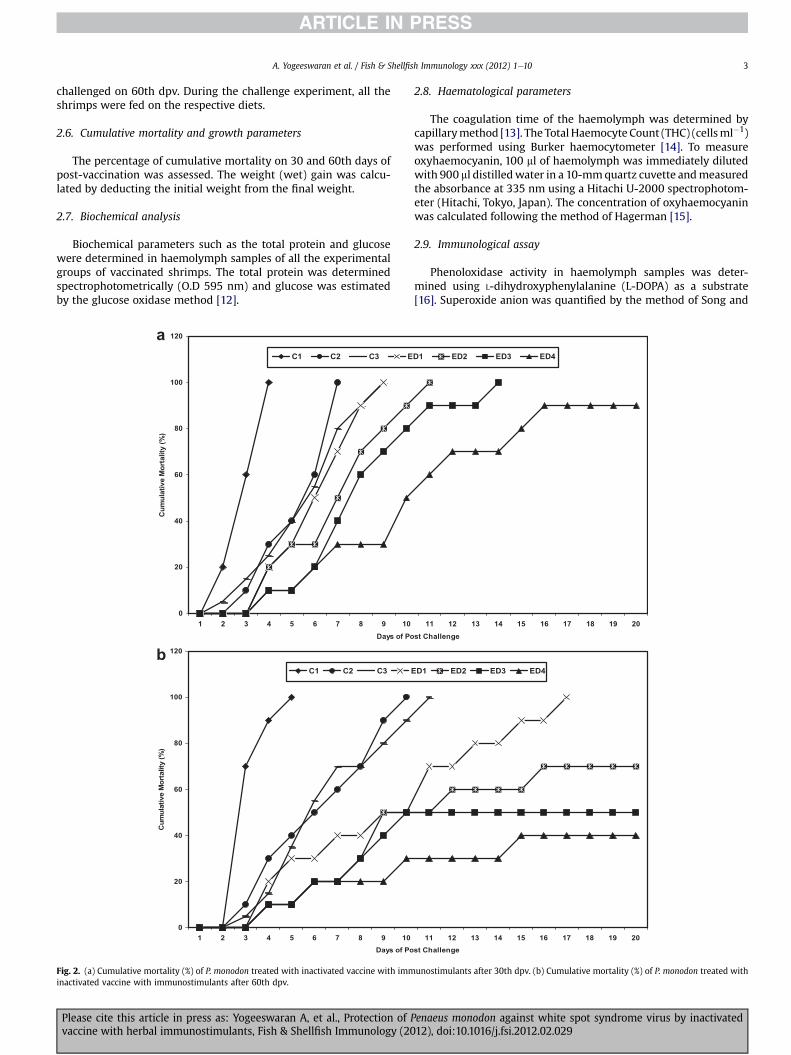
challenged on 60th dpv. During the challenge experiment, all theshrimps were fed on the respective diets.
2.6. Cumulative mortality and growth parameters
The percentage of cumulative mortality on 30 and 60th days ofpost-vaccination was assessed. The weight (wet) gain was calcu-lated by deducting the initial weight from the final weight.
2.7. Biochemical analysis
Biochemical parameters such as the total protein and glucosewere determined in haemolymph samples of all the experimentalgroups of vaccinated shrimps. The total protein was determinedspectrophotometrically (O.D 595 nm) and glucose was estimatedby the glucose oxidase method [12].
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9 10
Days of Po
Cu
mu
lativ
e M
ortality (
%)
C1 C2 C3 E
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9 10
Days of Po
Cu
mu
lativ
e M
ortality (
%)
C1 C2 C3 E
a
b
Fig. 2. (a) Cumulative mortality (%) of P. monodon treated with inactivated vaccine with imminactivated vaccine with immunostimulants after 60th dpv.
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
2.8. Haematological parameters
The coagulation time of the haemolymph was determined bycapillarymethod [13]. The TotalHaemocyte Count (THC) (cellsml�1)was performed using Burker haemocytometer [14]. To measureoxyhaemocyanin, 100 ml of haemolymph was immediately dilutedwith 900 ml distilledwater in a 10-mmquartz cuvette andmeasuredthe absorbance at 335 nm using a Hitachi U-2000 spectrophotom-eter (Hitachi, Tokyo, Japan). The concentration of oxyhaemocyaninwas calculated following the method of Hagerman [15].
2.9. Immunological assay
Phenoloxidase activity in haemolymph samples was deter-mined using L-dihydroxyphenylalanine (L-DOPA) as a substrate[16]. Superoxide anion was quantified by the method of Song and
11 12 13 14 15 16 17 18 19 20
st Challenge
D1 ED2 ED3 ED4
11 12 13 14 15 16 17 18 19 20
st Challenge
D1 ED2 ED3 ED4
unostimulants after 30th dpv. (b) Cumulative mortality (%) of P. monodon treated with
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

Table 1aGrowth characteristics of P. monodon after 30th day of post-vaccination (dpv).
Treatment Length (mm) Wet weight (mg) Weight gain (mg day�1)
Initial Final Initial Final
C1 38.6 � 1.02 49.00 � 0.81 376.0 � 20.46 1966.0 � 124.72 53.00 � 1.69a
C2 35.00 � 0.81 50.00 � 0.81 350.00 � 20.41 1950.00 � 40.52 53.33 � 1.69a
C3 36.08 � 1.3 52.45 � 0.50 359.03 � 12.85 2340.0 � 50.34 66.03 � 0.86b
ED1 37.6 � 2.05 53.00 � 0.81 364.00 � 11.44 2476.00 � 92.24 70.36 � 1.69c
ED2 38.00 � 1.63 54.00 � 0.81 353.6 � 5.56 2585.00 � 172.72 74.40 � 1.69c
ED3 38.0 � 1.62 54.30 � 0.47 362.00 � 9.93 2811.00 � 50.88 81.63 � 1.69d
ED4 38.6 � 1.02 56.3 � 0.47 368.00 � 15.57 3050.00 � 40.82 89.4 � 6.05e
Means with the same superscripts (aee) do not differ from each other (P < 0.05).
A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e104
Hsieh [17]. The optical density of the dissolved formazan was readat 630 nm and the effects of different treatments on the generationof O2
� in all the tested groups were compared.
2.10. Data analysis
All data obtained from the experiments were analyzed usingone way ANOVA (P < 0.05; P < 0.01 and P < 0.001 as significantlevel) in Statistica 6.0 computer package (Statsoft, UK). The Meanswere also compared using SNK test.
3. Results
3.1. Cumulative mortality after the 30th and the 60th days of post-vaccination
All the P. monodon juveniles succumbed to cent percent deathwithin 5 days by WSSV challenge, due to devoid of pre vaccinationand immunostimulant treatment after 30 and 60 dpv. The C2 and C3groups helped toprolong the survival times of 7, 9 days in 30 dpvand10, 11 days in 60 dpv. The Control groups (C2 and C3) had almost thesame value of cumulative mortality. The experimental diets(ED1eED4) fed P. monodon juveniles increased the survival ratesgradually from the lower concentration to higher concentration. Thecombined effect of vaccination and immunostimulation improvedthe survival rate at highly significant (P < 0.05) level. Whencompared to the 30 dpv challenge, the juvenile of P. monodon afterthe60dpvhadmuch improvement in survival. Thegroups suchasC1,C2 andED1 survived for5,10 and17days, respectively. Themaximumsurvivals were recorded in ED2 (30%), ED3 (50%) and ED4 (60%)groups of shrimps. The animals experimented with herbals havingthe antiviral and immunostimulant characteristics and the vacci-nations were highly influenced to resist the WSSV (Fig. 2a and b).
3.2. Growth parameters after the 30th and the 60th days of post-vaccination
The weight gain (mg day�1) was analyzed after the 30 and 60thdpv with WSSV challenging and given in the Tables 1a and 1b. The
Table 1bGrowth characteristics of P. monodon after 60th day of post-vaccination (dpv).
Treatment Length (mm) Wet w
Initial Final Initia
C1 49.00 � 0.816 60.00 � 0.816 196C2 50.00 � 0.816 59.66 � 1.247 1950C3 52.50 � 0.33 61.45 � 1.06 223ED1 53.00 � 0.816 64.00 � 0.816 2479ED2 54.00 � 0.816 66.6 � 0.942 258ED3 54.3 � 0.471 71.3 � 1.247 281ED4 56.3 � 0.471 74.6 � 1.247 3050
Means with the same superscripts (aed) do not differ from each other (P < 0.05).
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
weight gain recorded in the control group of C1 and C2 was53.0mg day�1 whereas the C3 had significantly (P< 0.05) increased(66.03 mg day�1) due to immunostimulative active principles inthe diets after 30 dpv. The experimental groups, ED1, ED2 ED3 andED4 attained significant weight gain (P < 0.05) of 70, 74, 81 and89 mg day�1, respectively. The juveniles of P. monodon after 60 dpvWSSV challenging achieved better growth performance as in the30 dpv. Theweight gain of 45mg day�1 was observed in the controlgroups, C1 and C2, but the weight gain in the experimental groupsincreased significantly (P < 0.05) as 57, 70 and 90 mg day�1 in thegroups such as ED1, ED2 and ED4, respectively.
3.3. Biochemical analysis after the 30th and the 60th days of post-vaccination
The biochemical parameters such as total protein and glucosecontents were studied in the haemolymph samples after the 30 andthe 60 dpv challenging. The total protein recorded in the haemo-lymph of control groups (C1eC3) was 121.6, 113.6 and 110 mg ml�1
respectively on the 4th day of 30 dpv challenged P. monodon. Theprotein levels were significantly decreased (P < 0.01) to 99, 96 and96 mg ml�1 in the ED2, ED3 and ED4 groups, respectively. Theglucose level also varied (P < 0.01) significantly among the groups.After the 60th dpv challenge, the protein content of the controlgroups were 122 mgml�1 in the C1 and C2 groups and 119 mgml�1 inthe C3 group on the 4th day after challenging. The protein contentwas significantly (P< 0.01) decreased to 120,101, 94 and 93 mgml�1
in ED1, ED2, ED3 and ED4 groups respectively on 10th day afterchallenging. The glucose values were recorded 114 and 129 mg ml�1
in the haemolymph of C1 and C2 groups, respectively. The valuesignificantly decreased (P < 0.01) to 117, 100, 96 and 88 mg ml�1 ofhaemolymph in the ED1eED4 groups respectively on the 10th dayafter the 60th dpv (Tables 2a and 2b).
3.4. Haematological parameters after the 30th and the 60th days ofpost-vaccination
The haemolymph coagulated within 171 s when no herbalextracts and immunostimulants were given in the diet (C1). The
eight (mg) Weight gain (mg day�1)
l Final
6.6 � 124.7 3316.6 � 102.7 45.00 � 1.08a
.00 � 40.82 3300.00 � 70.71 45.00 � 4.08a
4.0 � 67.22 3862.3 � 123.5 54.26 � 0.77b
.00 � 92.2 4193.3 � 140.5 57.14 � 1.95b
5.3 � 172.59 4616.6 � 102.7 70.31 � 2.28c
1.6 � 50.8 5050.00 � 147.1 67.82 � 1.94c
.00 � 40.82 5750.00 � 204.1 90.00 � 2.44d
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

Table 2aBiochemical changes in the haemolymph of P. monodon juveniles fed with different herbal extracts diets after 30th dpv.
Days Treatments
C1 C2 C3 ED1 ED2 ED3 ED4
P G P G P G P G P G P G P G
2 116.6a � 1.24 91.00a � 1.62 111.6b � 0.47 91.00a � 1.41 105.0c � 0.33 93.00a � 0.55 112.3b � 1.699 92.3a � 1.24 111.0b � 0.816 93.00a � 0.81 112.0b 0.816 90.00a � 0.81 112.6b � 1.247 92.00a � 0.814 121.6a � 0.471 104.6a � 2.05 113.6b � 1.247 100.6b � 0.94 110.5c � 0.50 102.6b � 1.30 116.0c � 0.816 101.6b � 0.47 115.3c � 0.471 105.3c � 1.24 112.3b 1.247 101.6b � 1.69 107.3d � 1.247 102.0b � 0.818 116.5a � 1.35 108.9a � 0.88 117.5a � 0.408 111.0b � 0.81 102.6b � 1.247 100.3c � 1.24 101.3b 1.247 92.00d � 0.81 96.6c � 1.247 91.6d � 0.4710 99.0a � 0.816 90.6a � 0.47 96.0b 0.816 88.00b � 0.81 96.0b � 1.414 86.00c � 0.81
P e protein; G e glucose. Units are expressed as mg ml�1.Means with the same superscripts (aed) do not differ from each other (P < 0.05).
Table 2bBiochemical changes in the haemolymph of P. monodon juveniles fed with different herbal extracts diets after 60th dpv.
Days Treatments
C1 C2 C3 ED1 ED2 ED3 ED4
P G P G P G P G P G P G P G
2 121.3a � 1.247 100.3a � 0.47 117.6b � 1.247 102a � 1.63 113.5c � 0.54 98.00a � 0.54 117.0b � 0.816 101.0a � 1.62 113.6c � 1.247 102.3a � 2.0 114.6c 1.247 103.3a � 1.69 113.6c � 1.247 102.3a � 1.244 122.6a � 1.247 114.3a � 0.94 122.6a � 1.247 116.3a � 0.47 119.8a � 0.34 107.8b � 0.85 123.0a � 0.816 111.6c � 0.47 122.3a � 2.05 116.0d � 0.81 117.0b 0.816 115.0d � 3.26 115.6b � 1.247 110.6c � 0.948 e 125.0a � 0.816 122.6a � 1.69 116.5b � 1.5 119.3a � 1.56 121.6b � 1.247 130.6b � 0.94 111.6c � 1.247 110.3c � 0.47 106.0d 0.816 104.0d � 1.63 101.3e � 2.05 95.3e � 1.2410 e 124.6a � 0.47 129.3a � 2.05 111.0a � 0.85 112.6a � 0.98 120.3b � 1.24 117.0b � 0.81 101.6c � 2.49 100.6c � 1.24 94.6d 0.471 96.3d � 1.88 93.0d � 2.44 88.6e � 1.24
P e protein; G e glucose. Units are expressed as mg ml�1.Means with the same superscripts (aee) do not differ from each other (P < 0.05).
A.Yogeesw
aranet
al./Fish
&Shellfish
Immunology
xxx(2012)
1e10
5
Pleasecite
thisarticle
inpress
as:Yogeesw
aranA,et
al.,Protectionof
Penaeusmonodon
againstwhite
spotsyndrom
evirus
byinactivated
vaccinewith
herbalimmunostim
ulants,Fish&Shellfi
shIm
munology
(2012),doi:10.1016/j.fsi.2012.02.029
����
����

Table 3aHaematological changes in the haemolymph of P. monodon juveniles fed with different herbal extracts diets after 30th dpv.
Days Treatments
C1 C2 C3 ED1
C THC OXY C THC OXY C THC OXY C
2 164.0a � 2.44 25.0a � 0.81 1.1a � 0.016 162.0a � 0.816 27.0a � 0.81 1.06a � 0.094 160.5b � 1.05 24.0ab � 0.81 0.96a � 0.01 161.6ab � 0.474 171.6a � 1.69 22.0a � 0.81 0.89a � 0.008 169.0a � 0.81 24.4b � 0.94 0.96b � 0.001 150.0c � 1.40 20.5ab � 0.51 0.91b � 0.05 143.6d � 0.948 e e e e e e 142.7a � 3.44 1.05a � 0.06 132.0b � 0.81610 e e e e e e 111.4a � 2.40 131.6b � 1.24
C e Coagulase activity (seconds); THC e Total haemocyte count (X106 cells ml�1); OH e oxyhaemocyanin (mmol 1�1).Means with the same superscripts (aee) do not differ from each other (P < 0.05).
A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e106
time for coagulation decreased to 169 s in C2 group. Further it wassignificantly (P< 0.01) decreased to 150 s in the C3 group due to theinclusion of immunostimulants in the diets on 4th day after 30 dpv.The time of coagulation was 131, 109, 109 and 90 s in the ED1eED4groups respectively after WSSV challenge. The decreasing time ofcoagulation is responsible for the decreased viral load in the hae-molymph. The total Haemocyte Count (THC) of 22, 24 and26 � 106 cells ml�1 was observed in the control groups (C1eC3) onthe 4th day after 30 dpv challenge, and the THC was significantly(P < 0.01) increased to 36, 42 and 43 � 106 cells ml�1 in ED1eED4groups, respectively, in the 10th day of challenging. The averagevalue of oxyhaemocyanin is 1.0 mmol l�1 in the second day afterchallenging. The values were decreased to a minimum of0.86 mmol l�1 in other control and experimental groups at earlierdays after challenge. The original level was maintained in theexperimental groups from 8 to 10 days after challenge due to thecombined effects. The same manner was reflected in P. monodonchallenged after 60 dpv also (Tables 3a and 3b).
3.5. Immunological parameters after the 30th and the 60th days ofpost-vaccination
The prophenol oxidase activity (proPO) value observed washigher in the herbal extracts incorporated diets fed groups than thecontrol groups during different days of challenging. At the 4th dayafter 30 dpv challenging, the less proPO value of 0.32 was observedin the C1 group, 0.52 in C2 group and 0.65 in the C3 group. The valuesignificantly (P < 0.01) increased to 0.68, 0.70, 0.79 and 0.85 inED1eED2 groups respectively after 2 days of challenging. The valuesfurther increased in 10th day after challenging. The same mannerwas reflected in the 60 dpv also (Fig. 3a and b). The intracellularsuperoxide anion (O2
�) production was significantly (P < 0.0001)enhanced by the herbal immunostimulants during 60 dpv. Afterchallenging, the O2
� concentration gradually increased from in ED1to ED4 groups. The maximum value recorded was 0.085 at ED4 dietfed group after 10 days of challenging (Fig. 4).
4. Discussion
Crustaceans do not possess an adaptive immune response withimmunoglobulins; however, recently, “quasi-immune response”
Table 3bHaematological changes in the haemolymph of P. monodon juvenile fed different herbal
Days Treatments
C1 C2
C THC OXY C THC
2 163.6a � 0.47 29.6a � 0.47 1.15a � 0.205 158.6b � 1.24 32.0b � 0.814 155.3a � 0.47 23.0a � 0.81 0.94a � 0.020 160.3b � 0.94 27.0b � 0.818 e e e 160.3a � 0.94 29.6a � 1.6910 e e e 140.3a � 1.24 33.3a � 0.47
C e Coagulase activity (s); THC e Total haemocyte count (�106 cells ml�1); OH e oxyMeans with the same superscripts (aee) do not differ from each other (P < 0.05).
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
has been reported in kuruma shrimp (M. japonicus) that helps forsurvival with natural or experimental white spot syndrome virus(WSSV) infections and has made resistance against challenge withWSSV. In this study, efficacy of vaccines is made of inactivatedWSSV with or without immunostimulants [11]. The resistanceagainstWSSV appeared in 3weeks after the virus exposure andwasmaintained for about 1 month, which was endorsed by theappearance of virus neutralizing factor(s) in the haemolymph of“immunized” kuruma shrimp [7].
In the present study, the combined effect of herbal extracts andformalin-inactivated WSSV vaccination helped to improve theprolonged survival. The treatment helped to reduce themortality at50% after 60 dpv challenging with WSSV. It indicates that, theimmunostimulation induces the quasi-immune response to resistvirus. Simultaneously the growth parameters such as weight gainwere improved much. The percentage of weight gain was 41e50%from the control group to experimental groups after 30 dpv and60 dpv treatments, respectively. P. monodon were fed pelletscoated with two WSSV envelope proteins, VP19 and VP28, andthen these shrimp were challenged 3, 7 and 21 days later, and theresults showed significantly higher survival at 3 and 7 days(relative survival, 64% and 77%, respectively) but it was reduced at21 day (relative survival, 29%) [18]. The marine Fish Turbot andFlounder were vaccinated with haemolysin gene HL1 displayedyeasts and challenged with Vibrio harveyi SF-1 cell suspension.Surprisingly the Flounders and Turbot gained the survival of 62.5and 57.1% respectively after post challenge [19]. Rout et al. [20]vaccinated black tiger shrimp, P. monodonwith WSSV genes, VP15,VP28, VP35 and VP281 individually through DNA vaccine vectorpVAX1. The non-vaccinated control group survived only 14% whilethe immunized groups survived (P < 0.001) around 70% interest-ingly, after WSSV challenge. DNA construct containing pcDNA 3.1-WSSV VP28 gene encapsulated in chitosan nanoparticles wasinvestigated in P. monodon by Rajeshkumar et al. [21]. The vacci-nated shrimp showed that significant relative survival of 85% after7 days challenged with WSSV, whereas 100% mortality wasobserved in the control shrimp. Vaccine formulations often requireplant based immunoadjuvants for increased immunological effi-ciency and better vaccination schedules [22]. Patwardhan et al. [23]studied the immunoadjuvant and immunostimulatory effects inmammalian system by themedicinal plants,W. somnifera, Tinospora
extracts diets after 60th day of post-vaccination (dpv).
C3 ED1
OXY C THC OXY C
1.176a � 0.009 154.0b � 4.05 29.4ab � 0.56 1.06a � 0.05 158.3b � 0.470.97b � 0.004 145.0c � 3.36 25.6ab � 1.41 0.95b � 0.08 155.3a � 1.240.98a � 0.009 140.8b � 1.42 28.3a � 1.70 0.96a � 0.04 140.0b � 0.8160.99a � 0.004 137.0a � 1.74 34.0a � 2.18 1.07a � 0.07 120.6b � 0.94
haemocyanin (mmol l�1).
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

Treatments
ED1 ED2 ED3 ED4
THC OXY C THC OXY C THC OXY C THC OXY
24.46a � 1.0 0.98a � 0.008 160.3b � 2.05 24.0ab � 0.81 1.04a � 0.04 161.6ab � 0.47 24.0ab � 0.81 0.98a � 0.008 160.3ab � 2.05 24.6ab � 0.47 1.06a � 0.0921.6b � 1.51 0.89ab � 0.014 155.0c � 1.63 23.0ab � 0.81 0.96b � 0.008 143.6d � 0.94 25.0c � 0.81 0.94b � 0.024 131.6e � 1.247 30.3d � 0.47 0.91ac � 0.012
e 1.1a � 0.081 143.6c � 1.24 32.3a � 0.41 1.00a � 0.004 132.0d � 0.816 34.0ab � 0.81 1.00a � 0.004 110.0e � 0.816 36.0c � 0.81 1.15a � 0.033e e 109.6a � 0.47 36.0a � 0.81 1.06a � 0.047 109.6a � 1.24 42.66b � 0.47 1.14a � 0.020 90.0c � 0.816 43.0c � 0.81 1.27b � 0.08
A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e10 7
cordifolia and Asparagus racemosus. Effective vaccination ofP. monodon and M. japonicus using inactivated Vibrio spp. has beenreportedbyseveral researchers [24,25]. Namikoshi et al. [11] noticedthat the protection of shrimp was enhanced by this dual adminis-tration (formalin-inactivatedWSSVþ Vibrio penaeicida) even on the30th dpv and the group gave significantly lowermortality (P< 0.05)than the control group. The present results revealed that, thedelivery of inactivated vaccine and immunostimulant treatedgroups had less survival compared to the dual administrated groups.
Indian traditional medicinal plants such as Aegle marmelos,Cynodon dactylon, Lantana camara, Momordica charantia and Phyl-lanthus amarus showed strong antiviral activity against WSSV inthe form of organic solvent extraction [9]. Direkbusarakom et al.[26] examined the ability of the ethanol extracts of Psidium guajavaleaves against various fish pathogenic viruses such as InfectiousHaematopoietic Necrosis Virus (IHNV) and infectious PancreaticNecrosis Virus (IPNV). Due to the antiviral compound nature, themethanolic extracts of the tested plants such as A. indica, C. dac-tylon, P. kurooa, W. somnifera and Z. officinalis effectively suppressedthe WSSV replications. Inactivated vaccines with immunostimu-lants administrated shrimps were achieved better haematologicalparameters compared to the control [27]. In the present study, thehaemolymph coagulated within 164 s in shrimp which was fed oncontrol diet. The heavy viral load in the haemolymph, may be thecause of late coagulation. The experimental diets help to decreasethe coagulation time around one time due to less viral load. Thesame manner was reflected in the THC and oxyhaemocyanin also.The decrease of THC which prevailed at the beginning of theexperiment, gradually increased and reached an optimal level(43 � 106 cells ml�1) after 8 days in the immunostimulant fed andvaccinated groups. Hennig et al. [28], studied that whenM. japonicus was fed on a commercial diet containing b-Glucan(BG) for 14 days and then infected with WSSV (formerly known asPRDV), the THCs of the BG-fed groups increased from 1.2 to 1.5times than those of the BG-free group before and after challenge.This was also supported by Chang et al. [29], and in this work theTHC was drastically reduced to 60% of its pre infection level by 24 hafter WSSV infection in P. monodon. In vivo experiments withM. japonicus demonstrated the presence of a quasi-immuneresponse, when WSSV survivors of both natural and experi-mental infections were rechallenged with WSSV and exhibiteda higher survival compared to naive shrimp. Moreover, haemo-lymphs of the survivors were able to neutralize virus preparationsup to two months after infection [30]. Roshan et al. [27], studied
Treatments
ED1 ED2 ED3
THC OXY C THC OXY C
32.6b � 0.47 1.166b � 0.012 160.0c � 0.816 30.6b � 0.47 1.133c � 0.047 161.6c �30.3c � 0.47 1.2c � 0.081 140.3c � 0.47 26.3bd � 0.47 1.06d � 0.094 140.3c �32.0b � 0.81 1.11a � 0.004 125.3c � 0.47 33.6b � 0.47 1.16a � 0.094 115.6d �40.0b � 0.81 1.13b � 0.008 114.6c � 0.47 39.0b � 0.81 1.17c � 0.01 102.3d �
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
the level of haemagglutinins changes in P. monodon, by theadministration of immunostimulants, beta-glucans and-or vibriobacterin with different mode of transmission. The study indicatesthat modulation of haemagglutinins may partly account for theimmunomodulatory activity of immunostimulants like beta-glucanor vibrio bacterins. Herbal extract, Stevia is responsible forimproved disease resistance, haematological and immunologicalparameters in grass prawn, P. monodon, juveniles against V. harveyiinfection [30].
Haemocytes play an important role in cellular defence, a lowernumber of circulating haemocytes in crustaceans reduces resis-tance to pathogens [14]. Yoganandhan et al. [31] observed signifi-cant differences of the coagulative time and THC between theWSSV and non-infected Penaeus indicus shrimps. The haemocytecount varies among crustacean species and is known to be affectedby a variety of factors such as infection and environmental stress[32]. Maeda et al. [33], have observed a decline in total haemocytecount in shrimp infected with penaeid rod-shaped DNA virus. Thedecrease in THC in infected animals was caused by haemocyticaccumulation at the site of injection for wound healing andphagocytosis of foreign bodies [34]. Haemocyanin represents80e95% of total protein in the haemolymph of crustacean. It hasbeen affected by the moulting cycle, nutritional conditions andstress [15].
The present study revealed that, the protein and glucose levelwas increased in the haemolymph on the earlier days of infectiondue to heavy viral load, especially in the C1 group. After herbaltreatment, the total protein level was reduced to 30% in theexperimental groups due to reduction of viral load. Lo et al. [35],reported that, high concentrations of protein and amino acids in thehaemolymph of crustaceans are due to heavy viral load. Anotherpossibility of the increase in protein in haemolymph of infectedshrimp is that baculoviruses encode a variety of proteases and otherenzymes that ‘melt’ the tissues [36], and that the proteins of the‘melted’ cells (muscle and hepatopancreas) would be incorporatedinto the shrimp haemolymph. Harwood et al. [37] reported thepresence of an abundant viral protein in the haemolymph ofManduca sexta larvae infected with polydnavirus.
It was evident that the total carbohydrate and glucose levelsincreased in haemolymph and decreased in muscle and hepato-pancreas of WSSV infected shrimps when compared with healthyshrimps [31]. Generally, the glucose level increases in infected orstressed animals. The possibility of high levels of glucose and totalcarbohydrate in haemolymph might be due to the transport of
ED4
THC OXY C THC OXY
0.94 34.0c � 0.81 1.15a � 0.016 159.0c � 1.63 31.0bd � 0.816 1.18a � 0.0080.47 30.3c � 0.47 1.1d � 0.141 130.3d � 0.47 30.0c � 0.81 1.03d � 0.0470.94 35.0c � 0.81 1.14a � 0.012 105.3e � 1.247 34.6c � 0.47 1.16b � 0.0081.24 41.3c � 0.94 1.26d � 0.008 90.3e � 0.94 44.3d � 0.94 1.23d � 0.01
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

Fig. 3. (a) Pro Phenoloxidase (ProPO) activity of inactivated vaccine with immunostimulant treated P. monodons’ haemocytes after 30th dpv. (b) Pro Phenoloxidase (ProPO) activityof inactivated vaccine with immunostimulant treated P. monodons’ haemocytes after 60th dpv.
A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e108
glucose and carbohydrate from hepatopancreas and muscle tohaemolymph. Hall and Van Ham [38], reported a significant eleva-tion of haemolymph glucose in P. monodon in stressed condition.
In the present experiment, the combined parameters likevaccination and immunostimulation helped to increase the proPOactivity and O2
� production that seem to act as a promoter of shrimpimmune system against the WSSV infection. proPO activatingsystem stimulates several cellular defence reactions, includingphagocytosis, nodule formation, encapsulation, and haemocytelocomotion [39]. Activated haemocytes also produce extra bacteri-cidal substances, such as H2O2 and superoxide anion (O2
�) that mayincrease disease resistance [17]. The present study revealed that, the
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
immunostimulation along with the vaccination helped to activatethe immune system. The experimental results revealed that, thehigher concentrations of herbal extracts in the diets (ED3 & ED4)induced the secretion of Prophenol Oxidase (PO) enzymes. This wasalready supported by Citarasu et al. [4] when delivering herbalimmunostimulants to the WSSV infected shrimps. Phagocytes havea unique membranous enzyme, NADPH oxidase, capable of one-electron reduction of molecular oxygen into superoxide anion(O2
�) during a process known as the respiratory burst. Being the firstproduct released during the respiratory burst, O2
� concentration iswidely accepted as an accurate parameter quantifying the intensityof a respiratory burst [40]. Chang et al. [41] found that, the THC of
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
0.1
C1 C2 C3 ED1 ED2 ED3 ED4
Treatments
OD
630
2nd Day 4th Day 8th Day 10th Day
Fig. 4. Intracellular superoxide anion (O2�) production of inactivated vaccine with immunostimulant treated P. monodons’ after 60th dpv.
A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e10 9
the BG-0.2, 1 and 2% diet fed P. monodon started their recovery after6 days of challenge and so did the O2
� production and the phagocyticactivity. Tinocordin based commercial immunostimulant, Immu-mod, on the growth and immune response of P. monodon throughfeeds against the Vibrio anguillarum challenge and the shrimpsgrown with immumod added diet at 0.1% showed maximumimmunity enhancement in terms of higher shrimp survival rate of83.33% whereas in control it was only 16.67%. Immunity enhance-ment was higher (P > 0.01) in shrimps grown at all concentrationsof Immumod added diets than the control [42]. Brewers yeast b-glucans significantly (P < 0.05) enhanced phenoloxidase (PO)activity of black tiger shrimp haemolymph as compared to controlswithout glucan [43].
The herbals active principles studied here are having the anti-viral and immunostimulating characteristics which offer bettergrowth, immunostimulation and act as antiviral during the dualadministration against the WSSV infection. The practicing ofimmunostimulation with herbal active principles is also a low cost,eco-friendly and proved to evolve “quasi-immune response”.Hence, the application will be very effective in aquaculture opera-tions against the viral infections.
Acknowledgements
One of the authors Dr. T. Citarasu gratefully acknowledges theUniversity Grant Commission (UGC), Government of India, for itsfinancial support, in the form of a research grant (No. 37-271 (SR)dtd 01.02.2010).
References
[1] Sahul Hameed AS, Anilkumar M, Stephen Raj ML, Jayaraman K. Studies on thepathogenicity of systemic ectodermal and mesodermol baculovirus and itsdetection in shrimp by immunological methods. Aquaculture 1998;160:31e45.
[2] Citarasu T. Herbal biomedicines e a new opportunity to aquaculture industry.Aquacult Int 2010;18:403e14.
[3] Smith P, Hiney MP, Samuelsen OB. Bacterial resistance to antimicrobial agentused in fish farming: a critical evaluation of method and meaning. Ann RevFish Dis 1994;4:273e313.
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
[4] Citarasu T, Sivaram V, Immanuel G, Rout N, Murugan V. Influence of selectedIndian immunostimulant herbs against white spot syndrome virus (WSSV)infection in black tiger shrimp, Penaeus monodon with reference to haema-tological, biochemical and immunological changes. Fish Shellfish Immunol2006;21:372e84.
[5] Venegas CA, Nonaka L, Mushiake K, Nishizawa T, Muroga K. Quasi-immuneresponse of Penaeus japonicus to penaeid rod-shaped DNA virus (PRDV). DisAquat Org 2000;42:3e89.
[6] Itami T, Yan Y, Takahashi Y. Efficacy of vaccination against vibriosis in culturedkuruma prawns Penaeus japonicus. J Aquat Anim Health 1999;1:234e42.
[7] Wu JL, Nishioka T, Mori K, Nishizwa T, Muroga KA. Time course study on theresistance of Penaeus japonicus induced by artificial infection with white spotsubdrome virus. Fish Shellfish Immunol 2002;13:391e403.
[8] Nadkarni KM. Indian materia medica with ayurvedic, unani, tibbi, sidha,allopathic, homeopathi, naturopathi and home remedies appendices andindexes, vol-I and II. New Delhi: Ram Pintograph (India); 1995.
[9] Balasubramanian G, Sarathi M, Rajesh Kumar S, Sahul Hameed AS. Screeningthe antiviral activity of Indian medicinal plants against white spot syndromevirus in shrimp. Aquaculture 2007;263:15e9.
[10] Boonyaratpalin M. Nutritional requirements of grouper Epinephelus. In: Theproceedings of grouper culture. Thailand: National Institute of CoastalAquaculture. Department of Fisheries; 1993. p. 50e5.
[11] Namikoshi A, Wu JL, Yamashita T, Nishizawa T, Nishioka T, Arimoto M, et al.Vaccination trial with Penaeus japonicus to induce resistance to white spotsyndrome virus. Aquculture 2003;229(1e4):25e35.
[12] Malik CP, Singh MB. Plant enzymology and histoenzymology. New Delhi:Kalyani Publishers; 1980. p. 278.
[13] Sachdev KN. Clinical pathology and bacteriology. New Delhi: Jaypee Brother;1983. 133 pp.
[14] Le Moullac G, Le Groumellec M, Ansquer D, Froissard S, Aquacop Levy P.Haematological and phenoloxidase activity changes in the shrimp Penaeusstylirostris in relation with the moult cycle: protection against vibriosis. FishShellfish Immunol 1997;7:227e34.
[15] Hagerman L. Haemocyanin concentration of juvenile lobster (Homarus gam-marus) in relation to molting cycle and feeding conditions. Mar Biol 1983;77:11e7.
[16] Söderhäll K. Beta-1,3 glucan enhancement of protease activity in crayfishhaemocyte lysate. Comp Biochem Physiol 1983;74:221e4.
[17] Song YL, Hsieh YT. Immunostimulation of tiger shrimp (Penaeus monodon)haemocytes for generation of microbiocidal substances: analysis of reactiveoxygen species. Dev Comp Immunol 1994;18:201e9.
[18] Witteveldt J, Cifuentes CC, Vlak JM, Van Hulten MCW. Protection of Penaeusmonodon against white spot syndrome virus by oral vaccination. J Virol 2004;78:2057e61.
[19] Zhu K, Chi Z, Li J, Zhang F, Li M, Yasoda HN, et al. The surface display ofhaemolysin from Vibrio harveyi on yeast cells and their potential applicationsas live vaccine in marine fish. Vaccine 2006;24:6046e52.
[20] Rout N, Kumar S, Jaganmohan S, Murugan V. DNA vaccines encoding viralenvelope proteins confer protective immunity against WSSV in black tigershrimp. Vaccine 2007;25:2778e86.
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029

A. Yogeeswaran et al. / Fish & Shellfish Immunology xxx (2012) 1e1010
[21] Rajeshkumar S, Venkatesan C, Sarathi M, Sarathbabu V, Thomas J, AnverBasha K, et al. Oral delivery of DNA construct using chitosan nanoparticles toprotect the shrimp from white spot syndrome virus (WSSV). Fish ShellfishImmunol 2009;26:429e37.
[22] Ruszala MV, Lin YI, Durr FE, Wang BS. Low molecular weight immunopo-tentiators. Int J Immunopharmacol. 64, 497e10.
[23] Patwardhan BK, Kalbag D, Patki PS, Nagsampagi A. Search of immunomodu-latory agents: a review. Indian Drugs 1990;28:56e63.
[24] Kou GH, Chen SN, Huang SL. Studies on bacterial infection and vaccinationtrials for culture Penaeus monodon in Taiwan. In: Diseases of fish and shellfish,abstract of the 4th EAFP International conference, 24e28 September 1989,Santiago de Compostella, Spain; 1989, p. 94.
[25] Teunissen OSP, Faber R, Booms GHR, Latscha T, Boon JH. Influence of vacci-nation on vibriosis resistance of the giant black tiger shrimp Penaeus monodon(Fabricius). Aquaculture 1998;164:359e66.
[26] Direkbusarakom S, Herunsalee A, Boonyaratpalin S, Danayadol Y,Aekpanithanpong U. Effect of Phyllanthus spp against yellow head baculovi-rus infection in black tiger shrimp, Penaeus monodon. In: shariff M, Arthur JR,Subasinghe RP, editors. Disease in Asian aquaculture II. Manila: Fish HealthSection. Asian Fisheries Society; 1993. p. 81e8.
[27] Roshan P, Khushiramani R, Karunasagar I, Karunasagar I. Effect of immunos-timulants on the haemolymph haemagglutinins of tiger shrimp Penaeusmonodon. Aquacult Res 2008;39(12):1339e45.
[28] Hennig O, Itami T, Maeda M, Kondo M, Natsukari Y, Takahashi Y. Analyses ofhaemolymph immunoparameters in kuruma shrimp infected with penaeidrod-shaped DNA virus. Fish Pathol 1998;33:389e93.
[29] Chang CF, Su MS, Chen HY, Lo CF, Kou GH, Liao IC. Effect of dietary b-1,3-glucan on resistance to white spot syndrome virus (WSSV) in postlarval andjuvenile Penaeus monodon. Dis Aquat Organ 1999;36:163e8.
[30] Leano EM, Xi YC, Liao IC. Effects of stevia extract on growth, non-specificimmune response and disease resistance of grass prawn, Penaeus monodon(Fabricius), Juveniles. J Fish Soc Taiwan 2007;34:165e75.
[31] Yoganandhan K, Thirupathi S, Sahul Hameed AS. Biochemical, physiologicaland hematological changes in white spot syndrome virus infected shrimp,Penaeus indicus. Aquaculture 2002;62:1e11.
[32] Persson M, Cerenius L, Soderhall K. The influence of haemocytes number onthe resistance of the freshwater crayfish Pacifastacus leniusculus Dana, to theparasitic fungus Aphanomyces astaci. J Fish Dis 1987;10:471e7.
Please cite this article in press as: Yogeeswaran A, et al., Protection of Pvaccine with herbal immunostimulants, Fish & Shellfish Immunology (20
[33] Maeda M, Itami T, Kondo M, Henning O, Takahashi Y, Hirono I, et al. Char-acteristics of penaeied rod shaped DNA virus of kuruma shrimp. Newapproaches to viral disease of aquatic animals. NRIA International Workshop.Nansei, Mie, Japan: National Research Institute of Aquaculture; 1997. pp.218e228.
[34] Ratcliffe NA, Rowley AF. Role of haemocytes in defense against biologicalagents. In: Gupta AP, editor. Insect haemocytes development forms, functionsand techniques. Cambridge, UK: CambridgeUniversity Press; 1979. p. 331e414.
[35] Lo CF, Ho CH, Chen CH, Liu KF, Chiu YL, Yeh PY. Detection and tissue tropism ofwhite spot syndrome baculovirus (WSBV) in captured brooders of Penaeusmonodon with a special emphasis on reproductive organ. Dis Aquat Organ1997;30:53e72.
[36] Beckage NE. Interactions of viruses with invertebrate cells. In: Soderhall K,Iwanaga S, Vasta GR, editors. New directions in invertebrate immunology.New York: SOS Publication; 1996. p. 375e99.
[37] Harwood SH, Grosovsky AJ, Cowles EA, Davis JW, Beckage NE. An abundantlyexpressed haemolymph glycoprotein isolated from newly parasitised Man-duca sexta larvae in polydnavirus gene product. Virology 1994;205:393e405.
[38] Hall MR, Van Ham EH. The effect of different types of stress on blood glucosein the giant tiger prawn Penaeus monodon. J World Aquacult Soc 1998;29:290e9.
[39] Johansson MW, Keyser P, Sritunyalucksana K, Soderhall K. Crustacean hae-mocytes and haematopoiesis. Aquaculture 2000;191:45e52.
[40] Secombes CJ. Isolation of salmonid macrophages and analysis of their killingactivity. In: Stolen JS, Fletcher TC, Anderson DP, Robertson BS, VanMuiswinkel WB, editors. Techniques in fish immunology. Fair Haven, NJ: SOSPublications; 1990. p. 137e54.
[41] Chang CF, Su MS, Chen HY, Liao IC. Dietary-1,3-glucan effectively improvesimmunity and survival of Penaeus monodon challenged with white spotsyndrome virus. Fish Shellfish Immunol 2003;15:297e310.
[42] Felix N, Jeyaseelan MJP, Kirubakaran CJW, George MR. Administration ofa commercial immunostimulant, immumod as a feed supplement enhancesimmunity and growth of postlarvae of tiger shrimp Penaeus monodon. In:Proceedings of the national seminar on new frontiers in marine bioscienceresearch, January 22e23, 2004, pp. 343e47.
[43] Suphantharika M, Khunrae P, Thanardkit P, Verduyn C. Preparation of spentbrewer s yeast beta-glucanswith a potential application as an immuno-stimulantfor black tiger shrimp, Penaeus monodon. Bioresour Technol 2003;88:55e60.
enaeus monodon against white spot syndrome virus by inactivated12), doi:10.1016/j.fsi.2012.02.029




















