
Properties of transcription factors regulating interleukin-2 gene transcription through the NFAT...
11
1994 84: 2612-2621 A Mouzaki and D Rungger treated naive and memory T-helper cells transcription through the NFAT binding site in untreated or drug- Properties of transcription factors regulating interleukin-2 gene http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: reserved. Copyright 2011 by The American Society of Hematology; all rights 900, Washington DC 20036. weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.org From
Transcript of Properties of transcription factors regulating interleukin-2 gene transcription through the NFAT...

1994 84: 2612-2621
A Mouzaki and D Rungger treated naive and memory T-helper cellstranscription through the NFAT binding site in untreated or drug- Properties of transcription factors regulating interleukin-2 gene
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rights900, Washington DC 20036.weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

Properties of Transcription Factors Regulating Interleukin-2 Gene Transcription Through the NFAT Binding Site in Untreated or
Drug-Treated Naive and Memory T-Helper Cells
By Athanasia Mouzaki and Duri Rungger
Combining in vitro DNA binding studies and functional tran- scription assays in the Xenopus oocyte, we have tested the presence and functional state of transcription factors con- trolling the interleukin-2 (IL-2) promoter through the NFAT binding site. In naive T-helper cells, the IL-2 gene is repressed by a silencer. After first mitogenic stimulation, this silencer becomes undetectable while an activator is newly synthe- sized. In resting memory cells, the activator has low DNA- binding affinity and is located in the cytoplasm. However, no silencer is formed. Upon renewed cellular activation, this pre-existing activator is again targeted to the nucleus and regains function in promoting transcription. Cyclosporin A
HE PRODUCTION O F interleukin-2 (IL-2) is con- trolled mainly at the transcriptional level’ and the cis-
acting sequences critical for its regulation are located within 320 bp upstream the cap site.’ The transcription factors (TFs) that bind to distinct sites of the IL-2 promoter in T cells include the lymphotropic Octamer, AP-1, and NFkB factors, as well as the cyclosporin A (CsAf- and FK506-sensitive factor NFAT- 1 .3”9 The binding site of NFAT- 1 is a purine- rich (Pu) element present twice in the human promoter at positions -130 (proximal) and -276 (distal). It has been reported that an NFAT- I subunit is present constitutively in the cytoplasm of unstimulated T-cell lines and is tissue- specifi~.’~’~~’’ Upon cellular stimulation, this factor, recently characterized for a mouse T-cell clone,’” is targeted to the nucleus, where it is reported to interact with a newly synthe- sized nuclear subunit to form the positive TF NFAT-I .”,’‘’ Treatment with the immunosuppressive drugs CsA or FK506 blocks early events in IL-2 transcription.” It has been sug- gested that the drugs bind to cytoplasmic prolyl cis-tmrrs isomerases (CsA to cyclophilin and FK506 to FKBP) and that the resulting complexes may then interact with the phos- phatase calcineurin.” The activation of calcineurin, in re- sponse to a T-cell receptor (TCR)-induced increase in intra- cellular calcium, is thought to fuel the translocation of the cytosolic NFAT-1 subunit to the nucleus, where it forms complexes with the nuclear subunit that is synthesized in response to TCR-induced protein kinase C activation.”-*“ According to Flanagan et al,9 the drugs block the nuclear
T
From the Division of Haematology, Department of Medicine, Ge- neva UniversiQ Hospital, Geneva; and Station de Zoologie expiri- mentule, University of Geneva, ChEne-Bougeries, Switzerland.
Submitted November 17, 1993; uccepted June 16, 1994. Supported by grants of the Swiss National Foundation .for Scien-
ti& Research. Address reprint requests to Duri Rungger, PhD, Station de Zoolo-
gie exp&rimentale, University of Geneva, 154 route de Malagnou, CH-1224 ChEne-Bougeries, Switzerland.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1994 by The American Socieiy of Hematology. 0006-4971/94/8408-0022$3.00/0
2612
and FK506 act on two distinct levels of the IL-2 control mech- anism. They prevent nuclear transport and reactivation of the preformed activator in memory cells and, in naive cells, they render the silencer resistant to displacement by the activator. DNA-binding of silencer and activator from T- helper, and NFAT-1 from Jurkat cells, requires the same three G residues, but cross-linking analyses show differ- ences in their constituent subunits. Supershift experiments show that the activator contains fra-2 and junD, whereas the silencer reacts with none of the antibodies tested. 0 1994 by The American Society of Hematology.
targeting of NFAT-1, and recently it was reported that, through binding to calcineurin, the drugs render this phos- phatase inactive, thus preventing the (de)phosphorylation of the cytoplasmic subunit of NFAT-I needed for its nuclear translocation.”.” The NFAT-l nuclear subunit was shown to contain members of the oncogenic protein families offi).s and juri." Interestingly, the members of fos and jun that participate in the NFAT-I multimer formation seem to vary with the cell type. The nuclear NFAT-l subunit in the leu- kaemia Jurkat was shown to contain c-jun or junD, fos, and, to a lesser extent,,frc-I,” whereas in primary human T cells it was shown to contain ajunB-frtl-l heterodimer.”
It has also been suggested that in certain cellular activity phases, for instance, when the cells are resting” or are aner- gic,’” the IL-2 gene may be downregulated by a transcrip- tional silencer(s). A silencer has been identified that binds to an element ( - 110 to - 101 ) situated between an Octamer 1 and,fos-jun binding sites in Jurkat and Raji B cells.“’
We studied the presence and function of TFs that control IL-2 transcription in primary T cells by DNA-protein binding studies and a functional trans-activation assay in Xenopus oocytes. The latter involves microinjection of a target gene and putative regulatory factors into the oocyte, to assess whether transcription is upregulated or do~nregulated.“.’~ The IL-2 promoter, like most injected genes, exhibits a basal activity in oocytes. Addition of proteins extracted from total resting T cells isolated from mouse spleen2’~*‘ or human peripheral blood”’.’’ silences the basal activity of the full- length (2-kb) IL-2 promoter, independent of whether it is linked to the IL-2 gene including introns and 3‘ flanking sequences or to a CAT coding sequence. Proteins from resting T cells also silence the basal activity of a minimal promoter controlled by 4 copies of the NFAT element. Proteins from resting T cells and the NFAT probe give a fast-migrating complex in bandshifts, the presence of which is strictly correlated with functional repressor activity in the oocyte system. Proteins from activated primary T cells and the NFAT probe give rise to a slower migrating complex. Such extracts only slightly enhance (about 1.5-fold) basal activity, but activation can clearly be monitored in a situation in which previously silenced genes are fully derepressed by the addition of activator.’x
The basal activities of all other minimal IL-2 promoters
Blood, Vol 84, No 8 (October 15). 1994: pp 2612-2621
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

IL-2 REPRESSOR AND ACTIVATOR 2613
tested exhibited constitutive expression and were not re- pressed by the addition of resting T-cell extracts, promoters controlled either by the NFkF3, Octamer, or AP-1 binding sites of the E-2 promoter, as well as the herpes simplex virus thymidine kinase gene or the W40 early promoter. Proteins extracted from mouse kidney, liver, brain, or resting human peripheral blood B cells or monocytes or from the tumor cell lines Jurkat or EL4 had no effect on the basal activities of all the genes tested.32
In the present report, we studied in more detail the pres- ence and function of the factors binding to the NFAT recog- nition site in naive and memory T-helper (Th) cell popula- tions during the process of cellular induction, and show that only in naive cells is the activator newly synthesized. We also show that in naive T cells CsA or FK506 stabilize the silencer and prevent derepression of the gene by functional activator, whereas in memory cells, the drugs inhibit the nuclear targeting of the activator as well as modifications needed for its function in promoting IL-2 transcription.
Moreover, we have analyzed the importance of some nu- cleotides involved in binding of silencer and activator from Th cells, and compared them with the requirement for bind- ing of NFAT-l from Jurkat cells. In this respect, no striking differences exist because all factors recognize the same three G residues within their recognition site. Nevertheless, as judged from cross-linking experiments performed with the silencer, activator, and NFAT-1, each factor seems to consist of different subunits in addition to a common one found in all complexes. Finally, supershift experiments show that the activator contains ffa-2 and junD, whereas the silencer pro- teins cross-react with none of the antibodies tested.
MATERIALS AND METHODS
Cells and Proteins Total T cells were isolated by rosetting with sheep red blood cells
(SRBC) from peripheral blood mononuclear cells of adult donors or umbilical cord blood, and Th cells were purified by positive selection using immunomagnetic beads (Dynal AS, Oslo, Norway) coated with anti-CD4 monoclonal antibody (MoAb).” Phenotyping for naive or memory cell markers was performed by fluorescence-activated cell sorting (FACS) as described.,’ To activate the cells, they were stimu- lated in cell cultures with 10 ndmL phorbol myristate acetate (PMA) and l pmol/L ionomycine at various times in RPM1 1640 medium with 10% fetal calf serum (FCS) and standard supplements. IL-2- secreting, CD4+/CD45RO+ T-cell clones were derived from periph- eral blood T cells as de~cribed.~’ The clones were either cultured for 2 weeks in plain culture medium (resting memory cells tested for viability, IL-2 secretion, and mRNA synthesis) or stimulated with PMA and ionomycine. When required, the cells were cultured in the presence of 20 pdmL cycloheximide or 1 pglmL CsA or 20 ng/mL FK506. Jurkat cells were cultured and stimulated with PMA and ionomycin, as were the Th cells.
To produce protein extracts, total cells were lysed, sonicated, and salt adjusted to 300 mmol/L to extract proteins from chromatin, and the lysate was centrifuged at 100,OOOg for 1 hour and the superna- tants were dialyzed against KIB buffer (Mouzaki et alz8 and refer- ences therein). When indicated, we prepared separate fractions of nuclear and cytoplasmic protein^.^' Proteins were extracted from Jurkat cells using the method’, modified in that the cell lysate was centrifuged at 100,OOOg to sediment chromatin and the supernatant
was dialyzed against KIB buffer. Protein concentration was deter- mined using the Bradford assay (BioRad, Hercules, CA).
Oligonucleotide Probes The following oligonucleotide probes we used. Long Pu-probe
was used corresponding to the NFAT element and including an ets- like element (bold) and a cryptic AP-l site (italic): AAGAAAG- GAGGAAAAACTGZTCATAAGGA. To test the binding patterns of the NFAT-binding factors derived from naive or memory Th cells at different activity phases as well as from Jurkat cells, we used a short Pu probe (AAGAAAGGAGIG2AAAAACTG3T), from which half of the cryptic AP-l site is missing but the third G residue is conserved. In the mutant oligonucleotides G1, G2, and G3, the corresponding G residues G , , G 2 . and G,, respectively, were re- placed by a T.
For cross-linking experiments, a long Pu oligonucleotide and a 9mer (capitals) were hybridized and filled in (small letters) with Klenow polymerase in the presence of 100 mmoK dGTP, dCTP, N,-dUTP,’4 and 10 mmol/L of ’*Pa-dATP for 30 minutes at room temperature: AAGAAAGGAGGAAAAACTGTTTCATAAGGAa; uucuuuccuccuuuuugacaaaGTATTCCIT.
Electrophoretic Mobility Shift Assays (Bandshifts) These assays were performed as de~cribed.9~ Briefly, 5 pg of
protein (or as stated) was incubated with about 120 pg DNA radiola- beled short Pu probe (l0,OOO to 30,000 cpm) in the presence of 0.5 pg poly(dIdC) nonspecific competitor DNA per microgram of pro- tein in a total reaction volume of 15 /*L. After incubation for 15 minutes at room temperature, the samples were fractionated on 4% or 5% denaturing polyacrylamide gels at 10 V/cm for 1.5 or 2 hours, respectively. In competition experiments, a 50- to 500-fold excess of cold short Pu or G mutant probes was added to the reaction mixture before the addition of the proteins. To compare the binding affinities of factors shown in Fig 3c, the autoradiogram was scanned on an Apple “Onescanner” and analyzed by the NIH Image 1.40 program on a Macintosh IIc computer (Macintosh Computers, Inc, Cupertino, CA). For quantitative evaluation of the autoradiograms shown in Figs 5 and 6, low exposures were scanned using a comput- ing densitometer and the data analyzed using ImageQuant V3.0 soft- ware, both from Molecular Dynamics (Sunnyvale, CA).
UV cross-linking. The binding reactions (in Eppendorf tubes) were irradiated for 20 minutes on ice at a distance of 10 cm, using a UV transilluminator (254 nm, 7000 pW/cm*). Using N3.dUTP, 5% to 15% of total protein-DNA complex is ~ross-linked.’~ Cross- linked material was separated on 10% sodium dodecyl sulfate (SDS)- polyacrylamide gels with prestained molecular weight (Mr) markers and visualized by autoradiography.
Supershifl experiments. These experiments were performed by the addition of antibodies (2 pg each) to the bandshift reaction mixtures. The antibodies (Abs) used were rabbit polyclonal IgG (exceptions are noted), all purchased from Santa Cruz Biotechnol- ogy, Inc (Santa Cruz, CA). The Abs were specific to their antigen and, with the exception of ets-l, not cross-reactive with other closely related proteins. The Abs were directed against the AP-I transcrip- tion factors c-fos p62, fosB, fra-l, fra-2 p46, c-jun p39, junB p39, and junD p39; the NFkB transcription factor subunits p65 and p50; c-ref p75; ets-l p54 (cross-reactive with ets family proteins), and the tumor-suppressor protein p53 (mouse IgG2a).
Xenopus Oocyte Injection and Target Genes The Pu-CAT gene clone4 used for injections contains four copies
of the murine NFAT promoter element in front of a truncated (-81) HSV thymidine kinase promoter and a CAT-coding segment. In
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

2614
addition, we constructed a 3xPuAP-CAT clone with three copies of the corresponding human sequences AAGAAAGGAGGAAAA- ACTGTTTCATAG inserted into the blunted Xba I site upstream of the SP-I motif of PBL~-CAT.~' Orientation of the inserts was tested by cutting with Xba I and Xho 1. Similarly. a 3xPu-CAT clone was constructed in which the cryptic AP-I site was truncated but the G3 residue still present (AAGAAAGGAGGAAAAACTGT). The 3xAP-I-CAT clone contains three copies of a consensus AP-I site: AAGTTATGACTCACCG.'* As a control, pBL2-CAT was used.
Functional assays in the Xenopus oocyte system are described in detail elsewhere (Rungger et al." Mouzaki et al,'x and references therein). Proteins (5 ng per injection per oocyte) were injected into the oocyte cytoplasm (groups of 1.5 oocytes). Two hours later, when the nuclear proteins had migrated to the germinal vesicle. I ng of DNA was injected intranuclearly, immediately followed by a second cytoplasmic or nuclear injection of proteins as indicated. Eighteen to 20 hours later, the activity of the target genes was measured by CAT assays as described.'* Usually. I O oocytes were lysed, and an aliquot corresponding to 1 oocyte was processed.
RESULTS
NFAT-Binding Factors in Naive Human T Cells Bandshifts with proteins from resting or stimulated naive
T cells isolated from cord blood and the NFAT probe (Fig 1 a, right-hand panel) show that the silencer can be detected until about 6 hours after addition of the mitogens. By about I O hours of mitogenic stimulation, the silencer disappears and the activator complex appears. Although the timing of the appearance of the activator vanes with different batches of cord blood, after performing several independent experi- ments we have seen that it appears at the earliest after 6 hours of stimulation with PMA and ionomycine; if the cells are stimulated with PMA alone or with phytohemagglutinin (PHA) and PMA, it can take longer than I O hours and as long as 24 hours to detect the activator in bandshifts (data not shown). It appears therefore that a combination of PMA and ionomycine is necessary for the synthesis of the activator from in vitro-stimulated cord blood-derived naive T cells (ie, strong calcium dependency). The activator is formed through de novo protein synthesis because it does not appear when the cells are stimulated in the presence of the transla- tional inhibitor, cycloheximide (or anisomycine; data not shown). On the other hand, the disappearance of the silencer on cellular stimulation is independent of translation. Band- shifts performed with a labeled NFAT element and proteins from resting or stimulated Th cells isolated from adult pe- ripheral blood are shown for comparison in Fig la (left panel).
Functional tests were performed in the Xenopus oocyte with the same protein extracts that were used for the band- shift assays (Fig I b). The results show that proteins extracted from resting naive or total T cells downregulate the basal transcription of a hybrid gene under the control of four NFAT promoter elements (4xPu-CAT). Extracts prepared between 2 to 6 hours after mitogenic stimulation of naive cells also downregulate the gene. Proteins extracted after a IO-hour stimulation of naive cells have lost their silencing activity. If the target gene is first silenced with protein ex- tracted from resting naive or total T cells, it cannot be dere- pressed by extracts from naive cells stimulated for 4 or 6
a
4- -
TO T4 b Pu-CAT
MOUZAKI AND RUNGGER
"
Q CO C2 C4 C 6 C l O C l r
B
0 CO C2 C4 C6 C10 TO TO TO TO CO C1yTOcx C4 C6ClOClO C10
Fig 1. De novo protein synthesis is required for the formation of the activator in naive Th cells. (a) Bandshifts with the NFAT probe and proteins extracted from TO, resting CD4' T cells isolated from human peripheral blood of adult donors; T4, 4-hour PMAlionomy- cine-stimulated TO; CO, resting CD4' cells isolated from cord blood; C2 through C10, 2- to 10-hour PMAlionomycine-stimulated cord blood cells; CIOcx, 10-hour PMAlionomycine-stimulated cord blood cells in t h e presence of cycloheximide. @, free probe. (b) Xenopus oocyte injections. CAT assays on oocytes injected first with proteins into their cytoplasm a s indicated in the first row, then with the Pu- CAT target gene into their nucleus and again into their cytoplasm, with the proteins indicated in the second row. Symbols as in la). 6, no protein injected.
hours in which the silencer is still present and the activator has not been synthesized. By contrast, I O hours after stimula- tion, the activator has been formed, and the resulting protein extracts fully derepress the silenced target promoter. When activator formation is blocked by cycloheximide, no such derepressing activity is obtained. In addition, the repressing activity of proteins from cells stimulated in the presence of cycloheximide is lost along with the disappearance of the silencer complex in the bandshift assay.
Effect of CA or FK506 on the Silencer
Addition of CsA or FKS06 to resting naive cells or at the beginning of their mitogenic stimulation results in an alteration of silencing activity. As shown in Fig 2, the re- pressing activity of proteins extracted from naive cells incu- bated for 4 hours with CsA in the absence of mitogen [CO""'"] or during mitogenic stimulation (C4"") silence basal transcription of the target gene. However, the silencing activity of these extracts cannot be overcome by the addition of proteins from activated memory cells (which contain only
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom
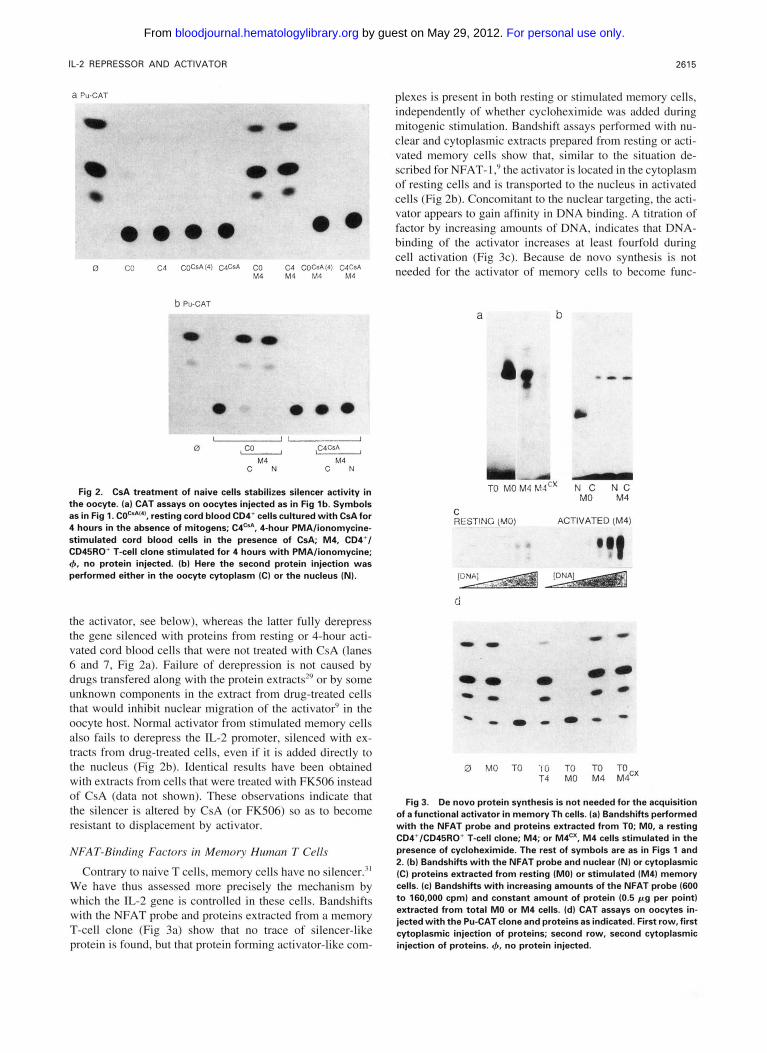
261 5 IL-2 REPRESSOR AND ACTIVATOR
a PU-CAT
Q , CO 1 C4CsA , M4
C N M4
C N
Fig 2. CsA treatment of naive cells stabilizes silencer activity in the oocyte. (a) CAT assays on oocytes injected as in Fig lb. Symbols as in Fig 1. COcd"', resting cord blood CD4' cells cultured with CsAfor 4 hours in the absence of mitogens; C4C'A, 4-hour PMAlionomycine- stimulated cord blood cells in the presence of CsA; M4, CD4'1 CD45RO' T-cell clone stimulated for 4 hours with PMAlionomycine; 4, no protein injected. (b) Here the second protein injection was performed either in the oocyte cytoplasm (C) or the nucleus (NI.
the activator, see below), whereas the latter fully derepress the gene silenced with proteins from resting or 4-hour acti- vated cord blood cells that were not treated with CsA (lanes 6 and 7, Fig 2a). Failure of derepression is not caused by drugs transfered along with the protein extracts'" or by some unknown components in the extract from drug-treated cells that would inhibit nuclear migration of the activator' in the oocyte host. Normal activator from stimulated memory cells also fails to derepress the IL-2 promoter, silenced with ex- tracts from drug-treated cells, even if it is added directly to the nucleus (Fig 2b). Identical results have been obtained with extracts from cells that were treated with FKS06 instead of CsA (data not shown). These observations indicate that the silencer is altered by CsA (or FKS06) so as to become resistant to displacement by activator.
NFAT-Binding Factors in Memo97 Human T Cells
Contrary to naive T cells, memory cells have no silencer." We have thus assessed more precisely the mechanism by which the IL-2 gene is controlled in these cells. Bandshifts with the NFAT probe and proteins extracted from a memory T-cell clone (Fig 3a) show that no trace of silencer-like protein is found, but that protein forming activator-like com-
plexes is present in both resting or stimulated memory cells, independently of whether cycloheximide was added during mitogenic stimulation. Bandshift assays performed with nu- clear and cytoplasmic extracts prepared from resting or acti- vated memory cells show that, similar to the situation de- scribed for NFAT-I: the activator is located in the cytoplasm of resting cells and is transported to the nucleus in activated cells (Fig 2b). Concomitant to the nuclear targeting, the acti- vator appears to gain affinity in DNA binding. A titration of factor by increasing amounts of DNA, indicates that DNA- binding of the activator increases at least fourfold during cell activation (Fig 3c). Because de novo synthesis is not needed for the activator of memory cells to become func-
a b
"a - TU rv;o M< ~ * : 3 ~ ~ N c N c
MO M4
C RESTING (MO) ACTIVATED (M4)
0 MO TO 'IO TO TO TO T4 MO M4 M4"
Fig 3. De novo protein synthesis is not needed for the acquisition of a functional activator in memory Th cells. (a) Bandshifts performed with the NFAT probe and proteins extracted from TO; MO, a resting CD4'/CD45RO' T-cell clone; M4; or M4". M4 cells stimulated in the presence of cycloheximide. The rest of symbols are as in Figs 1 and 2. (b) Bandshifts with the NFAT probe and nuclear (NI or cytoplasmic (C) proteins extracted from resting (MO) or stimulated (M41 memory cells. (c) Bandshifts with increasing amounts of the NFAT probe (600 t o 160,000 cpml and constant amount of protein (0.5 p g per point) extracted from total MO or M4 cells. (d) CAT assays on oocytes in- jected with the Pu-CAT clone and proteins as indicated. First row, first cytoplasmic injection of proteins; second row, second cytoplasmic injection of proteins. 4, no protein injected.
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

2616 MOUZAKI AND RUNGGER
tional, this increase is likely to be caused by an increased DNA-binding affinity. The functional capacity of the activa- tor closely reflects this alteration in affinity (Fig 3d). Proteins from resting memory cells, although they have no silencing activity on the IL-2 target promoter, are not capable of dere- pressing genes that were previously silenced with a dose of proteins from resting Th cells purified from adult peripheral blood. Extracts from activated memory cells, on the other hand, fully reactivate silenced genes, independently of whether the donor cells were stimulated in the absence or presence of cycloheximide (Fig 3d). These observations sug- gest that, in vivo, once acquired during a first mitogenic stimulation, the activator is maintained. In memory cells, IL- 2 regulation now proceeds in the absence of a silencer, through postranslational modifications of the activator that determine its cellular location, DNA-binding affinity, and its function in transcriptional activation of the IL-2 promoter.
Efiect of CsA or FK506 on Actiwtor
The addition of CsA or FKS06 to memory cells at the beginning of their mitogenic stimulation causes effects that closely resemble those described for NFAT- 1 ." As verified by bandshifts performed with nuclear and cytoplasmic pro- teins extracted from memory cells that were activated in the presence of either drug (shown here for CsA), nuclear targeting of the preformed activator is inhibited (Fig 4b). Proteins from drug-treated memory cells did not affect the basal activity of the Pu-CAT gene in the oocyte (Fig 4e).
The differences observed between the factors binding to the NFAT recognition site in naive and memory T cells have been shown in this report using naive (cord blood-derived) and memory T cells (clone). We performed an additional series of experiments aiming to directly compare these fac- tors with factors from total T cells isolated from adult periph- eral blood that contains a mixture of naive and memory cells (Mouzaki et a13' and references therein). To this end, we isolated cytoplasmic proteins from total resting T cells and then extracted proteins from chromatin of the same cells with salt washes and performed bandshift assays with the NFAT probe. In the same experiment, we also used proteins extracted from stimulated T cells. The resulting bandshifts are shown in Fig 4a and confirm that ( 1 ) in resting cells the silencer is found only in the nucleus and (2) the activator only in the cytoplasm, and that (3) the nuclear and cyto- plasmic activator from ex vivo T cells forms complexes of the same size in bandshifts as cytoplasmic or nuclear activa- tor isolated from pure memory CD4' cells. The cytoplasmic form of activator isolated from adult peripheral blood cells does not affect the basal activity of the Pu-CAT gene in the oocyte (Fig 4c) and it is incapable of derepressing silenced genes regardless of whether i t is injected into the oocyte nucleus or cytoplasm (Fig 4d).
Binding Requirements and Subunit Composi~ion of the Silencw and Activator
The silencer and activator of Th cells and NFAT-I of Jurkat cells all act through the same recognition sequence. To sort out whether differences exist in the residues recog-
a b
- 0
e PU-CA?
C3 CO MO MdCra MO M4 M4"' 0
Fig 4. CsA treatment of memory cells blocks nuclear targeting of the activator. (a) Bandshifts with the NFAT probe and proteins extracted from total peripheral blood T cells. C, cytoplasmic extract from resting cells (TO); CS, cytoplasmic extract from TO tested for DNA binding in the presence of 300 mmol/L salt; TO, proteins ex- tracted from chromatin of resting T cells after the removal of the cytoplasmic fraction; T4, total protein extract from 4-hour stimulated T cells. (b) Bandshifts with the NFAT probe and nuclear IN) or cyto- plasmic (C) proteins isolated from a memory CD4* cell clone stimu- lated for 4 hours in the absence (M41 or presence of CsA IM4C'A). +, free probe. (c) CAT assays performed on lysates from oocytes injected into their cytoplasm with proteins extracted from resting total T cells from peripheral blood (TO, first row) or cytoplasmic extract from the same cells (TO', first row), followed by nuclear injection of the Pu- CAT gene. Second row, second injection of proteins into the oocyte cytoplasm. T8, total protein extracted from 8-hour stimulated T cells. Id) CAT assays on oocytes injected first with proteins from resting total T cell extracts (TO) into their cytoplasm, as indicated in the first row, then with the Pu-CAT target gene into their nucleus and, again with cytoplasmic extract from TO cells (TO'), either into the cytoplasm (C) or into the nucleus (NI, as indicated in the second row. le) CAT assays on oocytes injected first with proteins from resting naive (cord blood TI cells (CO) or resting memory cell clone (MO) or activated memory cell clone in the absence IM4) or presence of CsA (M4C'A) into their cytoplasm, then with the Pu-CAT target gene into their nucleus. Duplicates denote protein preparations from different batches of cord blood or memory cell clones.
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

IL-2 REPRESSOR AND ACTIVATOR 2617
nized by the different factors, we first narrowed down the sequences required for correct regulation of the IL-2 gene. To this end, we constructed the 3xPuAP-CAT clone with the complete Pu-rich motif and the cryptic AP-I site, the 3xPu-CAT in which the AP- I motif is truncated, and a 3xAP- l-CAT clone with a consensus AP-I motif. These three clones were used for transcription assays in the Xenopus oocyte (data not shown). Both, the human 3xPuAP-CAT and 3xPu-CAT clones are regulated as the original murine 4xPu- CAT clone used above, ie, they are repressed by injection of proteins from resting naive Th cells and derepressed by the later addition of proteins from stimulated T cells. By contrast, the 3xAP-I-CAT, like the pBL2-CAT clone used a control, displays basal activity, but is not regulated by silencer- or activator-containing extracts. Thus, the presence of the complete cryptic AP- I site does not seem to be neces- sary for correct IL-2 regulation and a canonical AP-I site cannot assume this function alone.
Binding of proteins extracted from stimulated total periph- eral blood T cells to the NFAT probe has been shown to involve three G residues," two of which lay within the Pu- rich motif and the third in the cryptic Ap-l site. We tested the importance of these three residues also for the binding of silencer and activator from Th cells. We first tested the binding of a wild-type Pu probe and of the GI , G2. and G3 mutant probes with increasing amounts of silencer from resting cells and from resting and FKS06-treated cells or of cytoplasmic or nuclear activator from memory cells. The raw data for the cytoplasmic activator and the FKS06-treated silencer are shown in Fig 5a. The autoradiograms were scanned and the data from all experiments, normalized to maximal binding of the wild-type Pu probe by the respective protein, are summarized in Fig 5b. The results show that all proteins bind the wild-type Pu probe and about half as efficiently also retard the G3 mutant. By contrast, the GI mutation is hardly recognized by any of the proteins and recognition of the G2 mutant is very weak also.
To evaluate more critically binding affinity, we performed competition experiments with these probes. The same pro- teins were incubated with hot wild-type Pu probe in the absence or presence of a 50- or 500-fold excess of cold wild-type Pu or mutant GI , G2, or G3 probes. The data, normalized to Pu-binding in the absence of competitor, are shown in Fig 6. In agreement with the above binding studies, the Pu element competes efficiently with its binding and the G3 mutant is capable of competing with the wild-type probe, although only at a high excess. Moreover, it appears that the third G residue is more important for NFAT-I binding than for binding of silencer. Despite such slight variations, there is no decisive difference emerging between the binding prop- erties of the silencer and activator from Th cells and NFAT- 1 from Jurkat cells.
These results indicate that the different factors may share a common or similar subunit(s), eventually involved in DNA binding. To test whether common subunits exist between the silencer and activator, we performed mixing experiments of extracts from resting and activated Th cells. One of these experiments is shown in Fig 7a. Under no experimental con- ditions were intermediate bands formed between the two
a Cytoplasmic Activator '-""m .-._ " ~" .
Pu 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 0 1 2 3 4 I l G2 I ~3
,,o 1 Cyt. Activator Nucl. Activator NFAT-1
0.8 0.6 1 0.4 1 ,i 0.2
/ ' ~
1 0.0 i &
I . 0 1 2 3 4 0 1 2 3 4 3 1 2 3 4 p g
Fig 5. TF binding to G-mutated NFAT probes. Increasing amounts (0 to 4 mg) of proteins derived from resting cord blood T cells (si- lencer), FK5064reated cord blood cells cytoplasmic proteins from resting peripheral blood T cells (Cyt. activator), nuclear proteins from activated T cells (Nucl. activator), and proteins from activated Jurkat cells (NFAT-1) were used for bandshii assays with a fixed amount (150 pg) of wild-type NFAT-binding site (Pu) or mu- tants in which the first (GI), second (G2). or third (G31 G residue" was substituted by a T residue. (a) Examples of autoradiograms of bandshifts obtained with cytoplasmic activator and ~ i l e n c e r , f " ~ ~ re- spectively. (bl Quantitative data obtained by scanning of the autora- diograms from all experiments. The values are normalized to binding of wild-type Pu probe by 4 p g of the respective protein extract.
major retarded complexes nor any larger complexes indica- tive of interactions between silencer and activator subunits.
Cross-linking studies are another possibility to test differ- ences between factors binding to the same recognition motif. To perform such experiments, we used Azido-dUTP," which strongly enhances efficiency of UV cross-linking of proteins to DNA. A long Pu probe was produced in which the T residues of the strand complementary to the purine-rich re- gion were replaced by such N3.dUTPs. Analysis of cross- linked complexes on denaturing SDS-PAGE (Fig 7b) shows several bands for each of the Pu-specific factors tested. The silencer, activator, and NFAT-I extracts yield a common complex (open circle) with a size of about 40 kD, including the 30-bp-long cross-linked probe. About 10 kD (single strand) has to be deduced from all values. The common band could be a general DNA-binding protein (in view of its size, probably histone HI). In addition, several complexes of different sizes are formed by each protein preparation (black dots). The silencer forms two more bands (44 and 46 kD), the activator two larger bands (S3 and 62 kD), and NFAT-I a whole array of complexes (43, SO, 58, 67, and 105 to 130 kD). Deducing the probe, the main constituents
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom
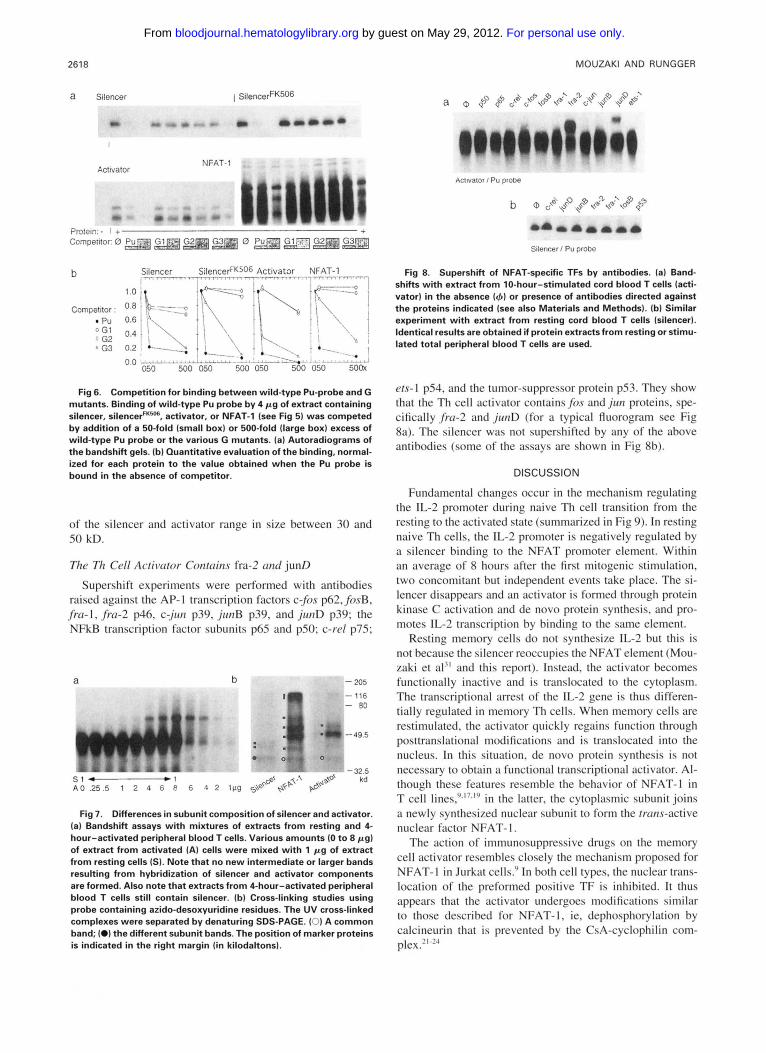
2618 MOUZAKI AND RUNGGER
a Silencer
Activator NFAT-1
Fig 6. Competition for binding between wild-type Pu-probe and G mutants. Binding of wild-type Pu probe by 4 p g of extract containing silencer, silencerFK50B, activator, or NFAT-1 (see Fig 51 was competed by addition of a 50-fold (small box) or 500-fold (large box) excess of wild-type Pu probe or the various G mutants. (a) Autoradiograms of the bandshift gels. lb) Quantitative evaluation of the binding, normal- ized for each protein to the value obtained when the Pu probe is bound in the absence of commtitor.
of the silencer and activator range in size between 30 and S0 kD.
The Th Cell Actiwtor Contcrins fra-2 and junD
Supershift experiments were performed with antibodies raised against the AP-I transcription factors c$)s p62, fosB, .fra-l. .fra-2 p46, c-jun p39. jlrnB p39, and junD p39; the NFkB transcription factor subunits p65 and p50; c-re1 p75;
- 205
- 116 - BO
-49.5
Fig 7. Differences in subunit composition of silencer and activator. (a) Bandshift assays with mixtures of extracts from resting and 4- hour-activated peripheral blood T cells. Various amounts (0 to 8 pg) of extract from activated (A) cells were mixed with 1 p g of extract from resting cells (S) . Note that no new intermediate or larger bands resulting from hybridization of silencer and activator components are formed. Also note that extracts from 4-hour-activated peripheral blood T cells still contain silencer. (b) Cross-linking studies using probe containing azido-desoxyuridine residues. The UV cross-linked complexes were separated by denaturing SDS-PAGE. (0) A common band; (0) the different subunit bands. The position of marker proteins is indicated in the right margin (in kilodaltonsl.
AcUvalOr I Pu probe
Fig 8. Supershift of NFAT-specific TFs by antibodies. (a) Band- shifts with extract from 10-hour-stimulated cord blood T cells lacti- vator) in the absence (4) or presence of antibodies directed against the proteins indicated (see also Materials and Methods). (bl Similar experiment with extract from resting cord blood T cells (silencer). Identical results are obtained if protein extracts from resting or stimu- lated total peripheral blood T cells are used.
ets-l pS4, and the tumor-suppressor protein pS3. They show that the Th cell activator containsfos andjtm proteins, spe- cifically ,fra-2 and jtrnD (for a typical fluorogram see Fig 8a). The silencer was not supershifted by any of the above antibodies (some of the assays are shown in Fig 8b).
DISCUSSION
Fundamental changes occur in the mechanism regulating the IL-2 promoter during naive Th cell transition from the resting to the activated state (summarized in Fig 9). In resting naive Th cells, the IL-2 promoter is negatively regulated by a silencer binding to the NFAT promoter element. Within an average of 8 hours after the first mitogenic stimulation, two concomitant but independent events take place. The si- lencer disappears and an activator is formed through protein kinase C activation and de novo protein synthesis, and pro- motes IL-2 transcription by binding to the same element.
Resting memory cells do not synthesize IL-2 but this is not because the silencer reoccupies the NFAT element (Mou- zaki et al" and this report). Instead, the activator becomes functionally inactive and is translocated to the cytoplasm. The transcriptional arrest of the IL-2 gene is thus differen- tially regulated in memory Th cells. When memory cells are restimulated, the activator quickly regains function through posttranslational modifications and is translocated into the nucleus. In this situation, de novo protein synthesis is not necessary to obtain a functional transcriptional activator. Al- though these features resemble the behavior of NFAT-I in T cell line^,".^'."' in the latter, the cytoplasmic subunit joins a newly synthesized nuclear subunit to form the trcms-active nuclear factor NFAT- 1.
The action of immunosuppressive drugs on the memory cell activator resembles closely the mechanism proposed for NFAT-I in Jurkat cells." In both cell types. the nuclear trans- location of the preformed positive TF is inhibited. It thus appears that the activator undergoes modifications similar to those described for NFAT-I, ie, dephosphorylation by calcineurin that is prevented by the CsA-cyclophilin com- Plex."-?"
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

IL-2 REPRESSOR AND ACTIVATOR 2619
PRIMARY T CELLS NAIVE MEMORY resting 4h-activated 1 Oh-activated rested 20' reactivated
SILENCER DISAPPEARANCE ACTIVATOR SYNTHESIS INACTIVATION~ACTIVATION
TUMOR T CELL LINE JURKAT
unstimulated stimulated
and a comparison with the proposed mechanism for Jurkat cells.' This figure is commented on in the dis- CYTOPLASMIC*NUCLEAR NF-AT cussion. NUCLEAR SUBUNIT SYNTHESIS
The activator is not the only target of CsA action. When resting naive cells are cultured with CsA or FK506, or if either drug is present during their mitogenic stimulation, the silencer seems to be altered as well. Such protein extracts exhibit a repression that, contrary to that caused by normal silencer (extracted from resting, untreated naive cells), can- not be overcome by activator injected into the oocyte. Evi- dence that CsA treatment has a direct effect on silencer stability and does not merely inhibit nuclear transport of injected activator is provided by three observations. ( I ) Di- rect injection of drugs into the oocyte has no effect on the activity of silencer or a~tivator. '~ (2) Nuclear injection of the activator also fails to derepress the IL-2 promoter (this report). (3) Irreversible silencing is also obtained with pro- teins extracted from cells cultured with CsA without being exposed to mitogenic stimulation (this report). Thus, the maintenance of a given phosphorylated or dephosphorylated state of the silencer protein would in this case have to be a continuous process. Because this process is affected by CsA and FKS06, it may depend on calcineurin action as well.
An important question concerns the relationship between silencer and activator on one side, and activator and NFAT- l derived from tumor cell lines, such as the leukaemia Jurkat, on the other. As far as the contacted residues within the recognition sequence are concerned, no striking differences could be observed. The silencer and activator both bind less efficiently to mutant oligodesoxynucleotides in which any of the three G residues contacted by a protein from stimulated peripheral blood T cells" was substituted by a T residue. Moreover, no apparent difference exists in this respect in the binding pattern between normal silencer and silencer from FK506-treated cells, nor between inactive cytoplasmic and functional nuclear activator. If ever, the third G residue located in the noncanonical AP-l site seems less important for silencer binding than for binding of activator and NFAT-
SYMBOLS
a SILENCER
@ INACTIVE ACTIVATOR
@ ACTIVE ACTIVATOR
@ CYTOPLASMIC SUBUNIT
NUCLEAR SUBUNIT
ACTIVE NF-AT
1. Interestingly, the noncanonical AP- 1 site (TGTTTCA) adjacent the Pu-rich motif is not necessary for IL-2 gene regulation by any of these TFs when the G residue remains in place (ie, a Pu-element missing the TTTCA end is func- tional).
However, marked differences exist in the subunit compo- sition of the silencer and activator. The cross-linking studies show that each factor contains two subunits that clearly differ in size. Whether the common band corresponds to a common subunit or is due to cross-linking of general DNA-binding proteins (probably histone HI) present in the incubation mix- ture cannot be decided yet. The presence of two bands may be due to the cross-linking of one or the other constituents of a heterodimer. Cross-linking of both components would be less efficient and could not be detected. The presence of one slightly larger band in the activator analysis, as judged from its size, may rather be due to an additional single com- ponent than to a dimer. Although in the presence of the cross-linked probe, molecular weight is difficult to estimate, the apparent size of these subunits lies between 30 and SO kD and is compatible with the identification of some subunits (see below).
Moreover, the different subunits of the two factors seem not able to form hybrid complexes when the respective pro- tein extracts are mixed. This might be due to the fact that each factor always exists as dimer or multimer, and that no free subunits are available that could form such hybrids. However, we assume, that the subunits of the different TFs are entirely different and are not capable of interacting with each other.
Additional evidence that the constituents of the silencer and activator are different has been obtained by supershift experiments. The activator contains fru-2 and junD (rather thanfru- 1 and junB).'* By contrast, the silencer still escapes identification. None of the numerous antibodies directed
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

2620 MOUZAKI AND RUNGGER
against known TFs were capable of supershifting the silencer complex. The main silencer components are thus different from those of the activator. Purification and molecular char- acterization of the silencer is underway and will clarify this question.
The behavior of memory T-helper cell activator resembles in its nuclear-cytoplasmic shuttle, and also in its response to immunosuppressive drugs, the well-characterized behav- ior of NFAT-l from tumor cell lines (see introduction for references). Also, the subunit composition of the activator, including fru-2 and junD, as determined by our supershift experiments, is similar to that of the Jurkat NFAT-1, the AP-I component of which was reported to include junD, c- jun, or fos.” However, in our cross-linking gels, NFAT- 1 contains many components and some large proteins are involved as could be expected from data published by oth- ers .~7 .~9 ,2n We had no antibody to test for the presence of the 120 kD component corresponding to NFATp,’” but, in the cross-linking experiment, the activator yields a relatively weak signal corresponding to a protein of this size.
An important difference between NFAT-l and activator resides in the fact that formation of activator in memory cells is independent of de novo protein synthesis (this report), whereas a nuclear subunit of NFAT-l has to be newly syn- thesized upon Jurkat cell activation.’,’ In addition, the two complexes are functionally different in that NFAT-l is not capable of derepressing silenced IL-2 promoters, as we have shown earlier.” Differences appear also in bandshift experi- ments on the level of nondenatured complexes. In general, NFAT-l (isolated from two batches of Jurkat cells using the same method we use to extract activator from ex vivo T cells) shows larger c~mplexes . ’~
Such differences may not be too surprising, because NFAT-like elements are found in the promoters of many cytokine and viral genes and proteins binding to such motifs can be derived from different cell types such as T cells, B cells, monocytes, mast cells, and NK cell^.^^^')^^' The resulting protein complexes seem to have a common component, NFATp, which, like the memory cell activator, may act alone4’ or in conjunction with several other proteins that vary depending on the promoter and cell type.“ In addition, the distal NFAT element of the IL-2 promoter was shown to form identical complexes in vitro when incubated with protein extracts from T or B cells, but, upon transfection, the NFATd element was inactive in B cells.42 It thus seems that NFAT-I is a collective term for a variety of NFAT- binding multimers.
Conclusively, IL-2 regulation is different in Th cells and in tumor cell lines. The main differences reside in the exis- tence of an IL-2-specific repressor in resting naive Th cells that is not found in other lymphoid cells, primary or immor- talized. As far as the activator is concerned, it is formed through de novo protein synthesis only during first mitogenic stimulation of naive Th cells, whereas later on its activation and deactivation depends on posttranslational modifications, and no subunit has to be formed each time through de novo protein synthesis. Moreover, the effect of immunosuppres- sive drugs is not identical in different types of T cells either. Whereas the effect on the Th cell activator and NFAT-I is
similar and consists in blocking of modifications of the pre- existing cytoplasmic activator or NFATp, in naive Th cells the drugs also affect the functional state of the silencer that becomes resistant to displacement. These observations may be pertinent for our understanding of 1L-2 gene regulation and evaluating drug effects in vivo.
ACKNOWLEDGMENT
We thank the amiable nurses of the Maternity Unit of the Univer- sity Hospital (Geneva) for supplying us with cord blood. Dr 0. Hagenbuchle (ISREC, Epalinges, Switzerland) for his advise on the cross-linking experiments and for sharing with us his extremely limited supply of N1.dUTP, Prof E. Serfling (Wurzburg, Germany) for the Pu-CAT clone, L. Muster for constructing the clones, C. Drijard for excellent technical assistance. and S. Chraiti for photog- raphy. We also thank Sandoz (Basel, Switzerland) for the gift of CsA and Pfizer Central Research (Groton, Switzerland) for the gift of FKS06.
REFERENCES
1 . Kronke M, Leonard WJ, Depper JM, Greene WC: Sequential expression of genes involved in human T lymphocyte growth and differentiation. J Exp Med 161: 1593, I985
2. Fujita T, Shibuya H, Ohashi T, Yamanishi K, Taniguchi T: Regulation of interleukin-2 gene: Functional DNA sequences in the 5’ flanking region for the gene expression in activated T lympho- cytes. Cell 46:403, 1986
3. Shaw JP, Utz PJ, Durand DB, Toole JJ, Emmel EA, Crabtree GR: Identification of a putative regulator of early T cell activation genes. Science 241:202, 1988
4. Serfling E, Barthelmas R, Pfeuffer 1, Schenk B, Zarius S, Swo- boda R, Mercurio F, Karin M: Ubiquitous and lymphocyte-specific factors are involved in the induction of the mouse interleukin-2 gene in T lymphocytes. EMBO J 8:46S, 1989
5. Shibuya H, Taniguchi T: Identification of multiple cis-elements and trcms-acting factors involved in the induced expression of human 1L-2 gene. Nucleic Acids Res 17:9173, 1989
6. Kamps MP: Corcol-an L. Lebowitz JH, Baltimore D: The pro- moter of the human interleukin-2 gene contains two Octamer-binding sites and is partially activated by the expression of Oct-2. Mol Cell Biol 105464, 1990
7. Verweij CL, Geerts M, Aarden LA: Activation of interleukin-2 gene transcription via the T-cell surface molecule CD28 is mediated through an NFkB-like response element. J Biol Chem 226:14179, 1991
X . Fraser JD, Irving BA, Crabtree GR, Weiss A: Regulation of interleukin-2 gene enhancer activity by the T cell accessory molecule CD28. Science 251:313. 1991
9. Flanagan WM, Corthesy B, Bram RJ, Crabtree GR: Nuclear association of a T-cell transcription factor blocked by FK-S06 and cyclosporin A. Nature 352303, 1991
10. Williams TM, Moolten D, Burlein J , Romano J . Bhaerman R, Godillot A, Mellon M, Rauscher FJ 111, Kant JA: Identitication of a zinc finger protein that inhibits IL-2 gene expression. Science 254:1791, 1991
11. Granelli-Piperno A, Nolan P: Nuclear transcription factors that bind to elements of the IL-2 promoter. J Immunol 147:2734. I991
12. Kang SM, Tran AC, Grilli M, Lenardo MJ: NFkB subunit regulation in non transformed CD4+ T lymphocytes. Science 2561452, 1992
13. Jain J, McCaffrey PG, Valge-Archer VE, Rao A: Nuclear factor of activated T cells contains Fos and Jun. Nature 356:801. 1992
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom

IL-2 REPRESSOR AND ACTIVATOR 2621
14. Chen D, Rothenberg EV: Molecular basis for developmental changes in interleukin-2 gene inducibility. Mol Cell Biol 13:228, 1993
15. Ghosh P, Tan TH, Rice NR, Sica A, Young HA: The interleu- kin-2 CD28-responsive complex contains at least three members of the NFkB family. Proc Natl Acad Sci USA 90:1696, 1993
16. Ullman KS, Northrop JP, Admon A, Crabtree GR: Jun family members are controlled by a calcium-regulated, cyclosporin A-sensi- tive signaling pathway in activated T-lymphocytes. Genes Dev 7: 188, 1993
17. Northrop JP, Ullman KS, Crabtree GR: Characterization of the nuclear and cytoplasmic components of the lymphoid-specific nuclear factor of activated T cells (NFAT) complex. J Biol Chem 2682917, 1993
18. Boise LH, Petryniak B, Mao X, June CH, Wang CY, Lindsten T, Bravo R, Kovary K, Leiden JM, Thompson CB: The NFAT-I DNA binding complex in activated T cells contains Fra-1 and JunB. Mol Cell Biol l3:19Il, 1993
19. Jain J, McCaffrey PG, Miner Z, Kerppola TM, Lambert JN, Verdine GL, Curran T, Rao A: The T-cell transcription factor NFATp is a substrate for calcineurin and interacts with Fos and Jun. Nature 365:352, 1993
20. McCaffrey PG, Luo C, Kerppola TK, Jain J, Badalian TM, Ho AM, Burgeon E, Lane WS, Lambert JN, Curran T, Verdine CL, Rao A, Hogan PG: Isolation of the cyclosporin-sensitive T cell transcription factor NFATp. Science 262:750, 1993
2 I . Bierer BE, Hollander G, Fruman D, Burakoff SJ: Cyclosporin A and FK506: Molecular mechanisms of immunosuppression and probes for transplantation biology. Curr Opin Immunol 5:763, 1993
22. Liu J, Farmer JD, Lane WS, Friedman J, Weissman I, Schreiber SL: Calcineurin is a common target of cyclophilin- cyclosporin A and FKBP-FK506 complexes. Cell 66:807, 1991
23. Clipstone NA, Crabtree GR: Identification of calcineurin as a key signaling enzyme in T-lymphocyte activation. Nature 357:695, 1992
24. O’Keefe SJ, Tamura J, Kincaid RL, Tocci MJ, O’Neill EA: FK-506- and CsA-sensitive activation of the interleukin-2 promoter by calcineurin. Nature 357:692, 1992
25. Nabel G, Gorka C, Baltimore D: T-cell-specific expression of interleukin-2: Evidence for a negative regulatory site. Proc Natl Acad Sci USA 852934, 1988
26. Go C, Miller J: Differential induction of transcription factors that regulate the interleukin 2 gene during anergy induction and restimulation. J Exp Med 175:1327, 1992
27. Rungger D, Muster L, Boeck R, Nichols A: Tissue-specific trans-activation of the rabbit-b-globin promoter in Xenopus oocytes. Differentiation 44:8, I990
28. Mouzaki A, Weil R, Muster L, Rungger D: Silencing and transactivation of the mouse IL-2 gene in Xenopus oocytes by pro- teins from resting and mitogen-induced primary T-lymphocytes. EMBO J 10:1399, 1991
29. Mouzaki A, Dai Y, Weil R, Rungger D: Cyclosporin A and FK506 prevent the derepression of the L 2 gene in mitogen-induced primary T lymphocytes. Cytokine 4: 151, 1992
30. Mouzaki A, Zubler RH, Doucet A, Rungger D: Trans-active factors controlling the IL-2 gene in adult human T-cell subsets. Mediators Inflammation 1:33, 1992
31. Mouzaki A, Rungger D, Tucci A, Doucet A, Zubler RH: Occurrence of a silencer of the interleukin-2 gene in naive but not in memory resting Th lymphocytes. Eur J Immunol 23:1469, 1993
32. Mouzaki A, Rungger D: Interleukin 2 gene regulation at the Pud (NFAT) promoter element: The complex situation in primary T lymphocytes. Mol Biol Hematopoiesis 3:475, 1994
33. Schreiber E, Matthias P, Muller MM, Schaffner W: Rapid detection of Octamer binding proteins with “mini extracts” prepared from a small number of cells. Nucleic Acids Res 17:6419, 1989
34. Evans RK, Haley BE: Synthesis and biological properties of 5-azido-2’-deoxyuridine 5’-triphosphate, a photoactive nucleotide suitable for making light-sensitive DNA. Biochemistry 26:269, 1987
35. Schreiber E, Matthias P, Muller MM, Schaffner W: Identifi- cation of a novel specific octamer binding protein (OTF-2B) by proteolytic clipping bandshift assay (PCBA). EMBO J 7:422 I , 1988
36. Roux E, Strubin M, Hagenbuchle 0, Wellauer PK: The cell- specific transcription factor PTFl contains two different subunits that interact with the DNA. Genes Dev 3:1613, 1989
37. Luckow B, Schutz G: CAT constructions with multiple unique restriction sites for the functional analysis of eukaryotic promoters and regulatory elements. Nucleic Acids Res 15:5490, 1987
38. Ryseck R-P, Bravo R: c-jun, junB and junD differ in their binding affinities to AP-l CRE consensus sequences: Effect of FOS proteins. Oncogene 6:533, 1991
39. Gorman CM, Moffat LF, Howard BH: Recombinant genomes express chloramphenicol acetyl transferase activity in mammalian cells. Mol Cell Biol 2:1044, 1982
40. Rao A: NFATp: A transcription factor required for the co- ordinate induction of several cytokine genes. Immunol Today 15:274, 1994
41. Rooney JW, Hodge MR. McCaffrey PG, Rao A, Glimcher LH: A common factor regulates both Thl- and Th2-specific cytokine gene expression. EMBO J 13:625, 1994
42. Yaseen NR, Maize1 AL, Wang F, Sharma S: Comparative analysis of NFAT (nuclear factor of activated T cells) complex in human T and B lymphocytes. J Biol Chem 268:14285, 1993
For personal use only. by guest on May 29, 2012. bloodjournal.hematologylibrary.orgFrom




















