
USAID Has Contract Termination Guidance That Aligns With ...
Upload
independentCategory
view
0download
0
Molecular Cell
Article
An Allosteric Path to Transcription TerminationVitaly Epshtein,1 Christopher J. Cardinale,1 Andrei E. Ruckenstein,2,3 Sergei Borukhov,4 and Evgeny Nudler1,*1Department of Biochemistry, New York University School of Medicine, New York, NY 10016, USA2BioMaPS Institute for Quantitative Biology3Department of PhysicsRutgers University, Piscataway, NJ 08854, USA4Department of Cell Biology, University of Medicine and Dentistry of New Jersey, Stratford, NJ 08084, USA
*Correspondence: [email protected]
DOI 10.1016/j.molcel.2007.10.011
SUMMARY
Transcription termination signals in bacteriaoccur in RNA as a strong hairpin followed bya stretch of U residues at the 30 terminus. Torelease the transcript, RNA polymerase (RNAP)is thought to translocate forward without RNAsynthesis. Here we provide genetic andbiochemical evidence supporting an alternativemodel in which extensive conformationalchanges across the enzyme lead to terminationwithout forward translocation. In this model,flexible parts of the RNA exit channel (zipper,flap, and zinc finger) assist the initial step ofhairpin folding (nucleation). The hairpin theninvades the RNAP main channel, causingRNA:DNA hybrid melting, structural changesof the catalytic site, and DNA-clamp opening in-duced by interaction with the G(trigger)-loop.Our results envision the elongation complexas a flexible structure, not a rigid body, and es-tablish basic principles of the termination path-way that are likely to be universal in prokaryoticand eukaryotic systems.
INTRODUCTION
Transcription elongation complexes (ECs) in all cellular or-
ganisms are extremely stable structures that proceed
through thousands of base pairs without losing contact
with the DNA template or nascent RNA. However, upon
encountering termination signals, ECs dissociate in a mat-
ter of seconds. Bacterial intrinsic terminators play multiple
roles in the cell. They separate operons into independent
transcription units, control the expression of downstream
genes within an operon, and ensure recycling of RNAP for
the next round of transcription (Friedman et al., 1987;
Condon et al., 1995; Weisberg and Gottesman, 1999;
Nudler and Mironov, 2004). The termination signal is de-
fined as a GC-rich inverted repeat immediately followed
by a stretch of at least four thymidine residues, resulting
in a strong hairpin fused to a U stretch at the 30 terminus
of the nascent RNA (Platt, 1986; Brendel et al., 1986). Pre-
Molecular C
vious biochemical analysis of the termination pathway un-
covered two major steps leading to dissociation of the EC.
Initially, the EC pauses at the point of termination. This
pause, which is driven by the presence of the U stretch,
provides sufficient time for a hairpin structure to form
(Gusarov and Nudler, 1999). Once formed, the hairpin per-
manently inactivates the EC, displaces RNA from the
RNA-binding channel, and unwinds the upstream half of
the hybrid (Gusarov and Nudler, 1999; Komissarova
et al., 2002). The EC then rapidly dissociates at physiolog-
ical salt concentrations, or remains in an irreversibly ‘‘trap-
ped’’ configuration at low ionic strength (Gusarov and
Nudler, 1999).
The mechanism by which the hairpin interacts with
RNAP and disrupts its key contacts with RNA and DNA
has not been resolved. RNA extraction is thought to
require forward translocation of the EC without RNA syn-
thesis (Yarnell and Roberts, 1999; Santangelo and Rob-
erts, 2004) (Figure 1A). Here we present data arguing for
a different (allosteric) mechanism, which implicates hair-
pin-mediated conformational changes of the RNAP
hybrid-binding site, the front DNA duplex-binding site
(DNA clamp), and the catalytic center as critical elements
of the termination process.
RESULTS AND DISCUSSION
The Front Element of the TerminatorA direct prediction of the forward translocation model is
that an AT- or GC-rich DNA segment in front of the termi-
nation site will either facilitate or suppress termination,
respectively. Indeed, forward translocation requires the
unwinding of downstream DNA, and a weak (AT-rich)
DNA duplex should favor this process (Tadigotla et al.,
2006; Vassylyev et al., 2007a). To discriminate between
forward translocation and allosteric models of termination
(Figure 1A), we first prepared a series of templates in
which a short DNA segment just downstream of the phage
l tR2 terminator was systematically modified (Figure 1B;
Figure 2). Our results show that the termination efficiency
(%T) was much greater for a GC-rich downstream duplex,
arguing against a simple forward translocation scenario
(Figure 1B). Such a dramatic effect indicates that the
downstream segment is an important element of the intrin-
sic terminator.
ell 28, 991–1001, December 28, 2007 ª2007 Elsevier Inc. 991

Molecular Cell
Mechanics of Transcription Termination
Figure 1. Termination Efficiency as a Function of Down-
stream DNA Duplex Strength
(A) Alternative models of termination. The forward translocation model
(right) postulates that the hairpin forces the EC to move forward with-
out RNA synthesis. Such movement should lead to downstream DNA
duplex unwinding and RNA extraction (Santangelo and Roberts, 2004).
The allosteric model (left) postulates that the hairpin does not induce
forward translocation but instead enters the RNA exit channel and
main channel to melt the RNA:DNA hybrid and destabilize the EC (Gu-
sarov and Nudler, 1999). Conformational change in the catalytic center
(diamond) caused by the invading hairpin leads to irreversible EC inac-
tivation.
(B) Effect of downstream DNA changes on termination. Hairpin-proxi-
mal sequences of the tR2 terminator are indicated (top left). The red
triangle shows the position of the catalytic center between the RNA
30 terminus and the downstream DNA duplex. The green U indicates
the termination site (U7 position). GC substitutions are shown in
blue; AT substitutions are in pink. The radioautogram shows termina-
tion and full-length (runoff) RNA products from standard chase reac-
tions (see the Experimental Procedures). The efficiency of termination
(%T) is indicated under each lane.
(C) Effect of downstream DNA changes on the termination rate of
stalled ECU7. tR2 templates used are shown below. The orange G indi-
cates the hybrid stabilizing substitution that prevents rapid hairpin fold-
ing and termination (Gusarov and Nudler, 1999). The initial immobilized
EC was walked to the termination site at position +68 (U7) followed by
washing with transcription buffer (TB) to remove unincorporated NTPs.
After the time intervals indicated (min), a 200 mM mixture of CTP, ATP,
and UTP (AT-rich template), or CTP, UTP, and GTP (GC-rich template)
was added to extend the U7 transcript to +77 or +80, respectively. %T
was calculated as a fraction of unchasable U7 transcripts. In the control
reaction (W, wash lanes), inactive transcripts were washed away with
TB after the chase, confirming their release during termination.
992 Molecular Cell 28, 991–1001, December 28, 2007 ª2007 El
This ‘‘front element’’ could influence termination by two
independent mechanisms. First, it could modulate paus-
ing at the site of termination, thereby altering the time win-
dow for hairpin formation (Gusarov and Nudler, 1999).
Second, the front element could affect termination alloste-
rically via the DNA clamp, in a pause-independent way. To
examine the latter possibility, we artificially stalled the EC
at the termination site (ECU7) (Figure 1C), in which case the
role of pausing becomes irrelevant. ECU7 was prepared by
walking RNAP to the tR2 termination site (Nudler et al.,
2003). Instant termination of ECU7 was prevented through
a U3/G substitution within the U stretch of tR2 (Fig-
ure 1C). Because of competition between the hairpin
and hybrid (Gusarov and Nudler, 1999; Komissarova
et al., 2002), the hybrid-stabilizing point substitution
increased the half-life of ECU7 in the range of minutes un-
der normal salt conditions (150 mM KCl), so that the termi-
nation process could be accurately measured in real time
(Figure 1C). Similar to the situation described in Figure 1B,
the GC-rich and AT-rich front elements rendered the mod-
ified terminator either stronger or weaker, respectively
(Figure 1C and Figure 2). In the absence of the hairpin,
these downstream substitutions did not affect EC stability
(data not shown). Thus, the specific sequence of the front-
element DNA affects termination through both pause-
dependent and pause-independent effects. Moreover,
the results of substitutions in the front-element DNA are
inconsistent with intuitive predictions of the forward trans-
location model.
Mapping DNA-Clamp Interactions that InfluenceTerminationIn the absence of forward translocation, the results de-
scribed above must be due to an interaction between
front-element DNA and the RNAP clamp. To characterize
this interaction, we mapped key contacts of the clamp
with the front element by performing two parallel sets of
experiments, one on a series of templates with substitu-
tions in the front element and a second using mutations
in the DNA-clamp domain of RNAP. Measurements on
a series of 2 bp AT substitutions scanning the entire GC-
rich front element to position +12 (counting from +1 of
the catalytic center) showed that substitutions at positions
+5–7 had the greatest inhibitory effect on termination, in-
dicating that interactions between the clamp and these
bases are the most significant for the termination process
(Figure 2). However, other surrounding DNA contacts also
contribute to the destabilization of the complex at the
terminator because any 2 bp substitutions have only small
effects compared to those that occur after a complete
GC/AT substitution in the front element.
Guided by the structural model of the EC (Figure 6; see
the Experimental Procedures) (Korzheva et al., 2000;
Gnatt et al., 2001; Westover et al., 2004; Kettenberger
et al., 2004), we also made amino acid substitutions in
the part of the clamp predicted to contact the front
element of the terminator. Three out of four of these sub-
stitutions resulted in substantially increased termination
sevier Inc.

Molecular Cell
Mechanics of Transcription Termination
Figure 2. Effect of the Front Element and RNAP Changes on the Termination Rate of the Stalled EC
Modifications of the hairpin-proximal part of the tR2 terminator are colored. The orange G indicates the hybrid-stabilizing substitution that allows real-
time measurement of the termination rate for each RNAP/modified terminator pair. AT-rich changes of the front element are pink; GC-rich changes are
blue. The green U indicates the U7 termination site. The orange G indicates the hybrid-stabilizing substitution that prevents rapid hairpin folding and
termination (Gusarov and Nudler, 1999). The red diamond between the hybrid and the front DNA duplex indicates the position of the catalytic center.
For each modified terminator, the %T was determined as a fraction of unchasable and washable U7 transcript after 30 min of incubation. DT repre-
sents the change in %T compared to wild-type RNAP (WT) and the original front element (line 4). Clamp, b01146–1148 mutant; zipper, b050–52 mutant;
G1136S, G loop fast mutant; I1134V, G loop slow mutant. Numbers represent the average values from three independent experiments with a standard
deviation of less than 10%.
efficiency (Figure 3A). The strongest effect was caused by
b01146–1148 substitutions, which are closest to the +5/+6
positions on the DNA. These mutations also act in
a pause-independent manner, as they increased the ter-
mination rate of the stalled ECU7 (Figure 2). Furthermore,
the b01146–1148 mutant was less discriminating with
respect to the GC-rich or AT-rich front element of the ter-
minator than was the wild-type enzyme (Figure 3A). Taken
together, these results locate the key contacts between
the clamp and the front element of the terminator that
need to be broken for termination to occur. Breaking these
contacts via the folding hairpin can only be achieved allo-
sterically because of the spatial separation of these two
elements (see Figure 6), and can be the rate-limiting step
in the termination pathway. Because the lateral movement
of RNAP on DNA is not changed by clamp mutations (the
mutant RNAP has the same elongation properties as wild-
type, see Figure S1 available online), these mutations
could modulate termination not by facilitating or inhibiting
forward translocation, but by affecting DNA-clamp open-
ing and DNA and transcript release at the termination point
(see below). Notably, according to the recent structure of
T. thermophilus (Tth) EC (Vassylyev et al., 2007a) the
b0 clamp residue R1266 (E. coli b0 R1148) makes direct po-
lar contacts with the nontemplate DNA at positions +4 and
+5 (from the major groove). A second structural element of
the b0 clamp containing residues 103–108 (E. coli b0 115–
120), which we predicted is involved in interaction with
Molecular C
+5/+6 DNA, binds in the minor groove of the DNA at posi-
tions +4 to +8 (Vassylyev et al., 2007a). Two key residues
within this region, b0 K106 and V108 (E. coli K118 and
V120), make direct polar and hydrophobic contacts with
positions +5/+6 of the template DNA strand, respectively.
Therefore, Ala substitutions made immediately adjacent to
residues 115–117 are likely to affect interactions of
K118/V120 with positions +5/+6 DNA.
RNA 30 Terminus Interacts with the G(Trigger)-Loop during TerminationThe forward translocation model implies that the RNA 30
terminus is pulled from the catalytic center and shifted
toward the RNA exit channel (Santangelo and Roberts,
2004) (Figure 1A). To test this prediction directly, we
took advantage of a RNA crosslinkable probe to monitor
changes in protein-RNA interactions in the catalytic site
during termination. If forward translocation were to occur,
the 30 end of the RNA would shift upstream into the main
channel and contact the nearby rifampicin-binding region
of the b subunit. Without forward translocation, the 30 end
should remain in the catalytic site and contact the
b0 subunit (G[trigger]-loop/F-bridge domains) (Figure 4A).
The tR2 construct allowed us to incorporate the crosslink-
able reagent 4-thio-uridine-50-monophosphate (sU) spe-
cifically at the termination point (U7 position, Figure 4B).
To prevent immediate dissociation of ECsU7, we prepared
the termination complex in low salt (5 mM KCl), thus
ell 28, 991–1001, December 28, 2007 ª2007 Elsevier Inc. 993
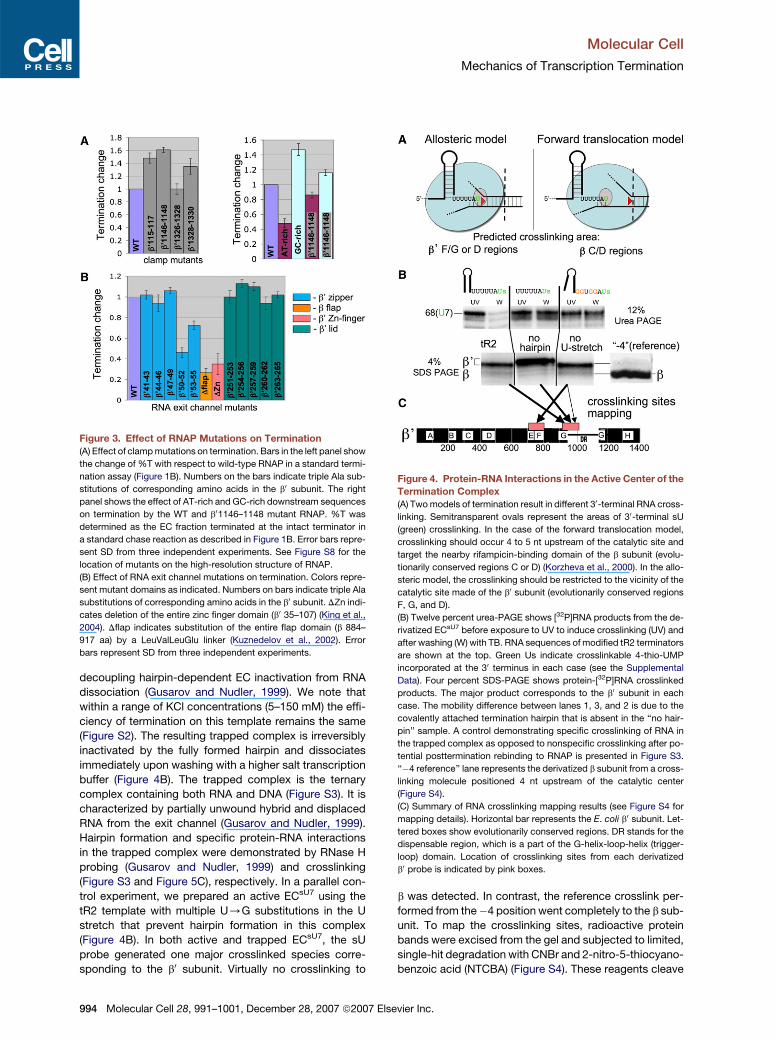
Molecular Cell
Mechanics of Transcription Termination
decoupling hairpin-dependent EC inactivation from RNA
dissociation (Gusarov and Nudler, 1999). We note that
within a range of KCl concentrations (5–150 mM) the effi-
ciency of termination on this template remains the same
(Figure S2). The resulting trapped complex is irreversibly
inactivated by the fully formed hairpin and dissociates
immediately upon washing with a higher salt transcription
buffer (Figure 4B). The trapped complex is the ternary
complex containing both RNA and DNA (Figure S3). It is
characterized by partially unwound hybrid and displaced
RNA from the exit channel (Gusarov and Nudler, 1999).
Hairpin formation and specific protein-RNA interactions
in the trapped complex were demonstrated by RNase H
probing (Gusarov and Nudler, 1999) and crosslinking
(Figure S3 and Figure 5C), respectively. In a parallel con-
trol experiment, we prepared an active ECsU7 using the
tR2 template with multiple U/G substitutions in the U
stretch that prevent hairpin formation in this complex
(Figure 4B). In both active and trapped ECsU7, the sU
probe generated one major crosslinked species corre-
sponding to the b0 subunit. Virtually no crosslinking to
Figure 3. Effect of RNAP Mutations on Termination
(A) Effect of clamp mutations on termination. Bars in the left panel show
the change of %T with respect to wild-type RNAP in a standard termi-
nation assay (Figure 1B). Numbers on the bars indicate triple Ala sub-
stitutions of corresponding amino acids in the b0 subunit. The right
panel shows the effect of AT-rich and GC-rich downstream sequences
on termination by the WT and b01146–1148 mutant RNAP. %T was
determined as the EC fraction terminated at the intact terminator in
a standard chase reaction as described in Figure 1B. Error bars repre-
sent SD from three independent experiments. See Figure S8 for the
location of mutants on the high-resolution structure of RNAP.
(B) Effect of RNA exit channel mutations on termination. Colors repre-
sent mutant domains as indicated. Numbers on bars indicate triple Ala
substitutions of corresponding amino acids in the b0 subunit. DZn indi-
cates deletion of the entire zinc finger domain (b0 35–107) (King et al.,
2004). Dflap indicates substitution of the entire flap domain (b 884–
917 aa) by a LeuValLeuGlu linker (Kuznedelov et al., 2002). Error
bars represent SD from three independent experiments.
994 Molecular Cell 28, 991–1001, December 28, 2007 ª2007 E
b was detected. In contrast, the reference crosslink per-
formed from the�4 position went completely to the b sub-
unit. To map the crosslinking sites, radioactive protein
bands were excised from the gel and subjected to limited,
single-hit degradation with CNBr and 2-nitro-5-thiocyano-
benzoic acid (NTCBA) (Figure S4). These reagents cleave
Figure 4. Protein-RNA Interactions in the Active Center of the
Termination Complex
(A) Two models of termination result in different 30-terminal RNA cross-
linking. Semitransparent ovals represent the areas of 30-terminal sU
(green) crosslinking. In the case of the forward translocation model,
crosslinking should occur 4 to 5 nt upstream of the catalytic site and
target the nearby rifampicin-binding domain of the b subunit (evolu-
tionarily conserved regions C or D) (Korzheva et al., 2000). In the allo-
steric model, the crosslinking should be restricted to the vicinity of the
catalytic site made of the b0 subunit (evolutionarily conserved regions
F, G, and D).
(B) Twelve percent urea-PAGE shows [32P]RNA products from the de-
rivatized ECsU7 before exposure to UV to induce crosslinking (UV) and
after washing (W) with TB. RNA sequences of modified tR2 terminators
are shown at the top. Green Us indicate crosslinkable 4-thio-UMP
incorporated at the 30 terminus in each case (see the Supplemental
Data). Four percent SDS-PAGE shows protein-[32P]RNA crosslinked
products. The major product corresponds to the b0 subunit in each
case. The mobility difference between lanes 1, 3, and 2 is due to the
covalently attached termination hairpin that is absent in the ‘‘no hair-
pin’’ sample. A control demonstrating specific crosslinking of RNA in
the trapped complex as opposed to nonspecific crosslinking after po-
tential posttermination rebinding to RNAP is presented in Figure S3.
‘‘�4 reference’’ lane represents the derivatized b subunit from a cross-
linking molecule positioned 4 nt upstream of the catalytic center
(Figure S4).
(C) Summary of RNA crosslinking mapping results (see Figure S4 for
mapping details). Horizontal bar represents the E. coli b0 subunit. Let-
tered boxes show evolutionarily conserved regions. DR stands for the
dispensable region, which is a part of the G-helix-loop-helix (trigger-
loop) domain. Location of crosslinking sites from each derivatized
b0 probe is indicated by pink boxes.
lsevier Inc.

Molecular Cell
Mechanics of Transcription Termination
the polypeptide chain next to Met and Cys residues, re-
spectively. Degradation patterns of the b0 subunit indicate
that the major crosslinks reside in the G(trigger)-loop and F
region in the termination (trapped) and active complexes,
respectively (Figure 4C and Figure S4). These results are
inconsistent with forward translocation of RNAP relative
to RNA at the terminator.
G(Trigger)-Loop Motion Is Requiredfor TerminationRecent structural studies of ECs have revealed that the
flexible G(trigger)-loop can assume at least two conforma-
tions, ‘‘open’’ and ‘‘closed’’ (Gnatt et al., 2001; Ketten-
berger et al., 2004; Wang et al., 2006; Vassylyev et al.,
2007b). The open conformation appears to be necessary
for the RNA backtracking and transcript cleavage reac-
tions, whereas the closed conformation is required for
catalysis of RNA synthesis and, presumably, for formation
of the trapped EC and subsequent termination. The avail-
able biochemical data indicate that the RNA 30 terminus in
the EC can be at an interacting distance with (and can
crosslink to) the G(trigger)-loop region in two instances:
(1) in backtracked EC when G(trigger)-loop is in an open
conformation and the RNA 30 terminus enters the second-
ary channel; and (2) in the active EC when the G(trigger)-
loop is in a closed conformation and the RNA is in a pre-
translocated state (Borukhov et al., 1991; Markovtsov
et al., 1996; Bar-Nahum et al., 2005). Indeed, in the back-
tracked ECsU7, in the absence of a hairpin, such crosslink-
ing is detected (Figure 4C). In the active EC, the 30 cross-
link goes mostly to the F region of RNAP (Figure 4)
(Markovtsov et al., 1996; Bar-Nahum et al., 2005). How-
ever, the folded hairpin must inhibit backtracking of the
trapped complex, as evidenced by the higher resistance
of the trapped complex to GreB-mediated cleavage com-
pared with the backtracked ECU7 (Figure S5). Moreover,
backtracking at the termination site is incompatible with
termination (Figure S6). We conclude that, rather than
pushing RNAP forward, as proposed by the forward trans-
location model, the hairpin induces a conformational
change of the G(trigger)-loop from the open to closed
state, and that this results in G(trigger)-loop movement
toward the RNA 30 terminus. Consistent with this conclu-
sion, the following data directly implicate G(trigger)-loop
movement (folding) in the termination process. First,
a slowly elongating RNAP (exemplified by an I1134V
G[trigger]-loop mutant) terminates much more efficiently,
whereas a ‘‘fast’’ G1136S mutant is essentially resistant
to termination at physiological NTP concentrations (Bar-
Nahum et al., 2005). The fast (or ‘‘slow’’) G(trigger)-loop
mutants are either less (or more) responsive to pausing
signals, suggesting that the G(trigger)-loop acts on termi-
nation by modulating the termination pause, which in turn
affects the folding kinetics of the termination hairpin (Gu-
sarov and Nudler, 1999). Second, to examine whether
the G(trigger)-loop can also modulate termination inde-
pendently of pausing, we studied the effect of G1136S
and I1134V mutants on the termination rate of the EC arti-
Molecular
ficially stalled at the termination site (ECU7) (Figure 5A; Fig-
ure 2). The fast G1136S mutation significantly decreased
the rate of termination of ECU7, while the slow I1134V
mutation had the opposite effect (Figure 5A). This
pause-independent effect is the opposite of what one
would expect from the forward translocation model of ter-
mination, which implies that the preferentially forward
translocated fast RNAP (Bar-Nahum et al., 2005) should
terminate more efficiently.
Additional evidence documenting the relevance of
G(trigger)-loop motion to termination follows from target-
ing the G(trigger)-loop with specific monoclonal anti-
bodies (mAbs) (Rouby et al., 2002). If added to the active
EC, these mAbs inhibit elongation and pyrophosphoroly-
sis without causing backtracking (Figure S7), confirming
the critical role of G(trigger)-loop folding in the catalytic
process (Bar-Nahum et al., 2005; Wang et al., 2006;
Vassylyev et al., 2007b). To monitor the effect of mAbs
on termination, we compared the time course of dissocia-
tion of the terminating ECU7 in the presence or absence of
anti-G(trigger)-loop mAbs; these mAbs stabilized ECU7,
preventing RNA release (Figure 5B). Note that the EC
was stalled first at the termination point, followed by mea-
surement of the rate of RNA release in the presence of
Abs. These latter experiments demonstrate that G(trig-
ger)-loop movement is essential for pausing-independent
termination. Therefore, G(trigger)-loop movement must be
necessary for hairpin-mediated opening of the enzyme to
allow RNA and DNA release. These results also suggest
that mAbs inhibit both elongation and termination by bind-
ing to and stabilizing the G(trigger)-loop in an open confor-
mation, and/or by preventing its folding into a closed
conformation. After the trapped EC has formed, the
hairpin is likely to force the G(trigger)-loop to remain in
the closed conformation, thus blocking access of the
G(trigger)-loop to mAbs. This would explain why anti-
G(trigger)-loop mAbs inhibit formation of the trapped
complex, but not the release of RNA from the already
formed trapped EC (Figure 5B and Figure S7E).
Our previous crosslinking experiments demonstrated
direct contact between the upper portion of the hairpin
stem and the G(trigger)-loop domain in the trapped com-
plex (Gusarov and Nudler, 1999). Such contact could oc-
cur only if the hairpin invades the main channel of the en-
zyme reaching out toward the catalytic center and
downstream clamp (Figure 6). To verify that such a dra-
matic event indeed takes place, we placed a crosslinkable
sU at a single position within the hairpin loop. The modified
EC carrying sU in the head (loop) of the tR2 hairpin (�15
position), or mutant hairpin (control), was then walked to
the termination site (ECU7). After a brief exposure to UV
light to induce crosslinking, trapped ECU7 and active
ECU7 were denatured and subjected to 4% SDS-PAGE
to visualize b and b0 subunits carrying the crosslinked
radiolabeled RNA (Figure 5C). In both complexes, the effi-
ciency of RNA-protein crosslinking was comparable in low
(5 mM KCl) salt. However, at a higher salt concentration,
crosslinking in the trapped complex disappeared, while
Cell 28, 991–1001, December 28, 2007 ª2007 Elsevier Inc. 995

Molecular Cell
Mechanics of Transcription Termination
Figure 5. Role of G(Trigger)-Loop in Termination
(A) Effect of G(trigger)-loop mutations on the termination rate of stalled ECU7. The hairpin-proximal sequence of the modified tR2 is indicated (top).
Red triangle indicates position of the catalytic center between the RNA 30 terminus and the downstream DNA duplex. Green U indicates the termi-
nation site (U7 position), and orange G indicates the hybrid-stabilizing substitution. The initial wild-type, fast (G1136S), and slow (I1134V) EC (Bar-
Nahum et al., 2005) were immobilized on Co2+ beads and walked to the termination point followed by washing with TB to remove unincorporated
NTPs. After the indicated time intervals (min), a 200 mM mixture of CTP, UTP, and GTP was added to extend the +68 (U7) transcript to +72. Percent
(%) T was calculated as a fraction of unchasable U7 transcripts. In the control (W, wash lanes), the inactive transcripts were washed away with TB after
the chase, confirming their release during termination.
(B) Inhibition of termination by G(trigger)-loop targeting antibodies. ECU7 was prepared as in (A), followed by addition of anti-G loop 18B5 monoclonal
antibodies (mAb; see the Experimental Procedures). In the control, the same amount of an anti-NusA mAb was used. After the indicated time intervals,
aliquots were washed with TB (W lanes). Percent (%) T was calculated as a fraction of washed U7 transcripts.
(C) Direct contact between the G(trigger)-loop and the hairpin loop during termination. The crosslinking probe (sU) was incorporated into the �15
position in the hairpin loop. The top panel (4% SDS-PAGE) shows protein-[32P]RNA crosslinking products (derivatized b0 and b subunits) from trapped
(lanes 1 and 2), active (lanes 3 and 4), and binary (lane 5) complexes. To induce crosslinking, UV irradiation was performed in TB with the indicated
amount of salt. Lane 5 was overexposed to compensate for the low intensity of radioactive material. The lower panel shows mapping of RNA cross-
linking by single-hit cleavage at methionine (Met) and cysteine (Cys) residues with CNBr and NTCBA, respectively. Autoradiogram of gradient (6%–
14%) SDS-PAGE shows products of partial degradation of the major crosslinked adduct in the trapped complex (marked by the red star in top and
bottom panels). Schematics on the right show a summary of mapping results. Vertical bar represents the E. coli b0 subunit. Lettered boxes show
evolutionarily conserved regions. DR stands for the dispensable region, which is a part of the G-helix-loop-helix (trigger-loop) domain. The red
star indicates location of crosslinking.
it remained unaffected in control. This result demonstrates
the salt sensitivity of protein-RNA contacts in the trapped
complex, and confirms that all the observed crosslinking
originated from the trapped complex, not some stable
paused or arrested complexes. The major crosslink in
the trapped ECU7, corresponding to the top band on the
4% SDS-PAGE (upper panel), was mapped to the G(trig-
ger)-loop domain (lower panel). In contrast, the major
crosslink in the control active EC was mapped to the
N-terminal part of b0 subunit (data not shown; Nudler et al.,
1998), i.e., in the vicinity of the RNA exit channel. These re-
sults suggest that direct ‘‘pushing’’ against the G(trigger)-
loopbythehairpin in the trappedcomplex leads toachange
in G(trigger)-loop conformation, which in turn causes EC
inactivation and potentially destabilization (Figure 6).
In a separate control experiment, we addressed the
possibility that G(trigger)-loop-RNA crosslinking origi-
996 Molecular Cell 28, 991–1001, December 28, 2007 ª2007 E
nated not from the trapped complex but from a nonspe-
cific binary complex (Kashlev and Komissarova, 2002).
We isolated sU-containing RNA from the trapped complex
and then added it to RNAP core in low salt to generate a bi-
nary complex, followed by UV irradiation. The resulting
crosslink was different from that obtained in the trapped
complex. First, the intensity of crosslinking was �103
lower than in the case of the trapped complex (the gel
was overexposed to allow for visualization of crosslinking
bands; Figure 5C, last lane). Second, the pattern of cross-
linking adducts was different, with the major (middle) band
corresponding to a different region or regions of the
b0 subunit that could not be precisely mapped due to
low intensity and poor specificity. Thus, G(trigger)-loop-
RNA hairpin crosslinks are specific to the trapped com-
plex, again confirming its mechanistically relevant, inter-
mediate state in the process of termination.
lsevier Inc.

Molecular Cell
Mechanics of Transcription Termination
In summary, these results show that the G(trigger)-loop
domain participates in each step of the termination path-
way. It controls pausing at the termination point mediated
by the U stretch (Bar-Nahum et al., 2005); it also directly
interacts with the folded hairpin and participates in EC in-
activation and dissociation. The latter is a key component
of our proposed allosteric mechanism.
Allosteric Mechanism of Intrinsic TerminationOur results indicate that formation of the termination
hairpin does not lead to a forward translocation of the EC
(see also the Supplemental Data). The alternative allos-
teric model we propose relies on the intrinsic structural
plasticity of RNAP to accommodate the hairpin inside
the RNA exit and main channels. Significant conforma-
tional changes across the enzyme, therefore, must occur
in order to cause the rapid and simultaneous release of
RNA and DNA. The structures of yeast and bacterial EC
(Gnatt et al., 2001; Westover et al., 2004; Kettenberger
et al., 2004; Wang et al., 2006; Vassylyev et al., 2007a) in
combination with the genetic and biochemical information
presented here allow us to define the following structural
model of intrinsic termination.
Hairpin Nucleation Phase
Termination occurs within a short distance (8 ± 1 bp) of the
hairpin base, setting a firm constraint on the positioning of
the hairpin within the enzyme. In our model, different mov-
ing parts of the RNAP play specific roles in termination: the
highly flexible domains of the RNA exit channel (flap, zip-
per, and lid), which is shaped as a narrow funnel, are likely
to move slightly apart to accommodate the upper portion
of the hairpin (Figure 6B). In addition, the flap, zipper, and
zinc finger (N-terminal zinc-binding-domain of b0) may act
as chaperones for the nucleation of the RNA hairpin and/or
its stabilization. Indeed, deletion of the flap and zinc finger,
as well as multiple point substitutions in the zipper do-
main, strongly inhibit termination in a pause-independent
manner (Figure 3B and Figure 2; King et al., 2004). The
active role played by these elements beyond merely spa-
tially accommodating the termination hairpin within the
exit channel is further supported by a series of genetic ex-
periments, the results of which are presented in Figure 3B.
These indicate that only those mutations in the RNA exit
channel that are predicted to be closest to the upper
portion of the folding hairpin have an inhibitory effect on
termination. Substitutions at the distal side of the zipper
(Figure 3B), substitutions in or deletion of the lid domain
(Figure 3B; Toulokhonov and Landick, 2006), or deletion
of the flap tip (b 900–909) (Toulokhonov et al., 2001)
have virtually no effect on termination. Thus, highly spe-
cific interactions between the RNA exit channel domains
and the head of the hairpin may facilitate initial RNA fold-
ing. Also, these interactions may allosterically induce fold-
ing of G(trigger)-loop, thus causing RNAP to pause in the
pretranslocated state (Toulokhonov et al., 2007), which
would prepare the EC for the next (irreversible) phase of
termination. Hairpin nucleation is likely to be the most sen-
sitive step in the termination process, hence the strong
Molecular C
negative effect of mutations. Specific and nonspecific
promoter-proximal alternative RNA secondary structures
and termination/antitermination factors should readily in-
terfere with hairpin formation at this point (Goliger et al.,
1989; Telesnitsky and Chamberlin, 1989; Rees et al.,
1997; King et al., 2004; Gusarov and Nudler, 2001; Miro-
nov et al., 2002).
Hairpin Incursion Phase
Two other elements that play crucial roles in our model are
the G(trigger)-loop and the DNA clamp, both of which are
implicated in the final stages of the termination process
(Figures 6C and 6D). Our structural model suggests that,
once the hairpin grows beyond 4 to 5 bp, it invades the
main channel with an inevitable melting of the hybrid (Gu-
sarov and Nudler, 1999) (Figure 6C). Melting of the up-
stream portion of the hybrid was demonstrated directly
in our previous crosslinking experiments. Growth of the
hairpin stem to 7 to 8 bp pushes the hairpin further within
the main channel. Due to a natural tendency of the RNA
hairpin to coil (e.g., Cilley and Williamson, 2003), the
head of the hairpin bends around the nontemplate strand,
and reaches toward and directly contacts the G(trigger)-
loop in the open conformation, pushing it to assume
the closed conformation. This scenario is directly sup-
ported by our crosslinking experiments at the terminator
(Figure 5C and Gusarov and Nudler, 1999). This feature
of our model is the most realistic explanation of how the
head of the hairpin (the loop and upper portion of the
stem) can reach the G(trigger)-loop. It is consistent with
the ability of dsRNA to interact with the main channel in
a mode distinct from that of the RNA:DNA hybrid (Ketten-
berger et al., 2006). An alternative (but less plausible)
explanation, which is also consistent with our G(trigger)-
loop-RNA crosslinking data, is that the hairpin stem
replaces the downstream DNA in the clamp, thus destabi-
lizing the EC. A direct contact between the G(trigger)-loop
and hairpin has two consequences. First, because the
G(trigger)-loop is a part of the catalytic site, its hairpin-in-
duced folding into the closed state can inhibit substrate
binding (Bar-Nahum et al., 2005; Wang et al., 2006; Vassy-
lyev et al., 2007b), and results in an irreversible trapping of
RNAP in a pretranslocated position (trapped complex).
This notion is supported by crosslinking of the 30 end of
RNA to the G(trigger)-loop in the trapped complex (Fig-
ure 4) and with the fact that point mutations in the G(trig-
ger)-loop change the efficiency of termination by affecting
the rate at which RNAP enters the trapped state
(Figure 5A). Second, the forced motion of the G(trigger)-
loop, which constitutes the side wall of the DNA-binding
clamp, may induce the movement of adjacent structural
elements (such as b fork 2; Wang et al., 2006; Vassylyev
et al., 2007b) and result in simultaneous opening of the
clamp. In turn, opening of the clamp would remove all
major interactions between RNAP and nucleic acids
(Figure 6D), thus inducing simultaneous release of the
transcript and DNA. This explains why point mutations in
both the clamp and its downstream DNA target alter the
efficiency of termination, but not the lateral movement of
ell 28, 991–1001, December 28, 2007 ª2007 Elsevier Inc. 997

Molecular Cell
Mechanics of Transcription Termination
Figure 6. Structural Model of Intrinsic
Termination
RNAP structures with explanatory schemes
represent four possible stages of the termina-
tion process. Key structural elements are color
coded: gray (green), nontemplate (template)
strand of DNA; red, nascent RNA; yellow,
b flap; violet, b0 lid; blue, b0 zipper; green, zinc
finger; rose and gold, b lobes 1 and 2; aqua,
b rudder; dark blue, G(trigger)-loop; and teal,
DNA clamp and other parts of b0. Star, the cat-
alytic center.
(A) Paused EC. The first step involves a pause
at the end of the U stretch, the role of which is
to provide sufficient time for the hairpin to initi-
ate folding (Gusarov and Nudler, 1999).
(B) Hairpin nucleation. Nucleation of the hairpin
could be initiated by interactions of the nascent
RNAwith the zinc finger andflapdomains, which
constitute the edge of the exit channel. The exit
channel funnel may be wide enough to accom-
modate the hairpin head and a few base pairs
of the stem, but tocontinue its growth the hairpin
has to open the exit channel into a groove.
(C) Hairpin incursion. The hairpin stem grows to
4 to 5 bp and begins invading the primary chan-
nel, unwinding the upstream end of the hybrid
and initiating collapse of the upstream edge of
the transcription bubble, while simultaneously
pushing on the flexible b0 rudder and inducing
opening movement of b lobe 2. This would result
in widening of the main channel, which would al-
low the hairpin to move toward the downstream
DNA and the secondary channel of RNAP.
(D) Trapped complex. The hairpin stem grows
to its final size of 7 to 8 bp while its head bends
around the downstream edge of the transcrip-
tion bubble, clashing with the G(trigger)-loop.
This shortens the hybrid to about 3 bp, further
collapsing the bubble. The clashing of the hair-
pin with the G(trigger)-loop may force it to
move toward the active site and refold into
a closed conformation, which would lock the
enzyme in the pretranslocated state. Addition-
ally, the hairpin action results in distortion of
the hybrid, which could cause complete and
irreversible enzyme inactivation. At the same
time, G(trigger)-loop movement induces
DNA-binding-clamp opening, resulting in re-
lease of the nascent RNA and disengagement
of RNAP from the DNA template.
(E) Different view of the trapped complex with
G(trigger)-loop in unfolded conformation. The
view is obtained by rotation of (D) around
the vertical axis by �90� counterclockwise.
The head of the hairpin is shown at interacting
distance of �2 to 3 A from the G(trigger)-loop.
(F) Trapped complex with G(trigger)-loop re-
folded into a closed conformation and moved
away from the hairpin by �5–7 A. The view is
shown in the same orientation as (E).
RNAP on DNA during productive elongation (Figures 1 and
3 and Figure S1). Our model also explains the fact that
G(trigger)-loop-specific antibodies prevent RNA/DNA
release and thus abolish termination (Figure 5B). We
998 Molecular Cell 28, 991–1001, December 28, 2007 ª2007
note that free energy gained in the process of hairpin fold-
ing must be sufficient to induce all the conformational
changes involved in the above model and to overcome
potential repulsion between a few phosphates of the
Elsevier Inc.

Molecular Cell
Mechanics of Transcription Termination
downstream DNA and the hairpin stem located at a dis-
tance of �6 A from each other (see the Supplemental
Data).
Concluding RemarksHere we present experimental evidence implicating key
mobile parts of RNAP such as the DNA clamp, G(trig-
ger)-loop, and RNA exit channel domains in termination.
We also present direct crosslinking data (Figure 4 and
Figure S4) and several additional pieces of evidence
showing that no forward translocation occurs during
termination. Finally, we formulate a structural model of ter-
mination shown in Figure 6. This model is the result of
searching for a consistent structural representation in
which the hairpin invades the exit and main channels of
RNAP in the course of intrinsic termination, an element
that appears to be required by the present experimental
results. The specific scenario outlined above was reached
by computationally optimizing the corresponding struc-
tural pathway to involve the shortest sterical paths with
no clashes between the RNA phosphate backbone and
polypeptide, while minimizing short-range electrostatic
repulsion in each structural state along the pathway (see
the Experimental Procedures). The resulting model is con-
sistent with and complements all the genetic, biochemi-
cal, crosslinking, and biophysical evidence presented
here (see also the Supplemental Data) and elsewhere
(Gusarov and Nudler, 1999).
The structural organization and principles by which RNA
and DNA are retained in the EC are highly conserved
among all cellular RNA polymerases (Korzheva et al.,
2000; Gnatt et al., 2001; Kettenberger et al., 2004; Wang
et al., 2006; Vassylyev et al., 2007a), suggesting that the
allosteric path leading to termination is universal. Prokary-
otic and eukaryotic termination factors and intrinsic sig-
nals are likely to utilize the same strategy to rapidly disrupt
the extremely stable and processive elongation com-
plexes. The conceptual framework and methodology
developed in the present study will be useful in testing
this prediction.
EXPERIMENTAL PROCEDURES
DNA Templates, Proteins, and Plasmids
The T7-tR2 template was a 154 bp PCR amplified fragment carrying
the T7A1 promoter and tR2 terminator (Gusarov and Nudler, 2001).
Its transcribed sequence upstream of the terminator is 50-ATCGA-
GAGGGACACGGCGAATAGCCATCCCAATCGAACATCG-30. All other
templates were prepared from this template by PCR-mediated muta-
genesis. All modifications are shown in the figures. His6-tagged wild-
type, Zipper, Lid, and zinc finger mutant RNAP were purified and
immobilized on Talon Co2+ beads (Clontech) as described (Nudler
et al., 2003). A plasmid expressing the rpoC lacking the whole zinc fin-
ger was kindly provided by R. Weisberg (NIH, Bethesda). His6-tagged
Dflap RNAP was provided by K. Severinov (Rutgers University).
EcoRQ111 was expressed and purified as described (Wright et al.,
1989). GreB was purified and stored as described (Borukhov and
Goldfarb, 1996). Plasmid constructions were based on a clone of the
rpoC gene in the vector pBAD-myc/His-A (Bar-Nahum et al., 2005).
Complementary pairs of oligonucleotides (IDT) were prepared to en-
Molecular C
code mutations in the coding sequence of rpoC. Mutagenesis was per-
formed by overlap extension PCR (see the Supplemental Data).
Transcription Reactions
All substrates were purified by ion exchange chromatography (Nudler
et al., 2003). The preparation of a start-up EC with a 32 nt 32P-labeled
RNA and solid phase transcription reactions (walking) were performed
as described (Nudler et al., 2003). Briefly, His6-RNAP (�2 pmol) was
mixed with a 2-fold molar excess of DNA in 20 ml of TB50 (10 mM
MgCl2, 40 mM Tris-HCl [pH 7.9], and 50 mM KCl) for 5 min at 37� fol-
lowed by addition of ApUpC (10 mM, Oligos Etc.) and GTP and ATP (25
mM) for 7 min. Next, 5 ml TB100-equilibrated Talon Co2+ affinity bead
suspension (Clontech) was added for 5 min at room temperature fol-
lowed by washing with 23 1.5 ml of TB1000 (1 M KCl) and 23 1.5 ml
of TB200 (200 mM KCl). To produce start-up EC32, ATP, GTP
(5 mM), and 1 ml of [a-32P]CTP (3000 Ci/mmol; NEN Life Science Prod-
ucts) were added for 5 min at room temperature followed by CTP
(5 mM) for a further 2 min. All chase reactions were performed at
25�C with 10 or 100 mM NTP (Pharmacia) for 5 min. The efficiency of
termination (%T) was calculated by dividing the amount of radioactivity
in a particular terminated band by the total radioactivity in the temi-
nated and all readthrough bands.
To measure the termination rate of the stalled EC, the start-up EC
was walked to template position +61 followed by washing with
TB500 (500 mM NaCl, 40 mM Tris HCl [pH 8.0], and 10 mM MgCl2).
Next, UTP, GTP, and ATP (5 mM) were added for 30 s at room temper-
ature to produce EC68, followed by washing with TB500 to initiate the
time course of termination. Aliquots were taken at the indicated time
intervals to tubes containing premixed ATP, GTP, and CTP, or GTP,
CTP, and UTP (100 mM final concentration) and incubated for 5 min
at room temperature. The reactions were stopped by addition of an
equal volume of stop buffer (SB: 8 M urea, TBE buffer, 20 mM EDTA,
0.125% bromophenol blue, and 0.125% xylene cyanol). The longest
time point sample in each experiment also served as a control for
RNA release: it was washed twice with TB500 to remove the transcript
from the terminated complexes before quenching. The roadblock
experiments were performed essentially as described (Nudler et al.,
1995).
For mAbs-mediated termination inhibition, slow terminating EC68
was obtained as described above, washed with TB500, and incubated
at room temperature either with 1 mg/10 ml of control mAbs or 1 mg/10 ml
of the anti-G(trigger)-loop mAbs 18B5 (provided by Jean-Paul Leo-
netti, Montpellier University, France). Aliquots were taken after 10
and 30 min and washed with TB500 at room temperature to determine
the amount of released RNA. The reaction was stopped by addition of
an equal volume of SB. For mAbs-mediated elongation inhibition the
startup EC20 was prepared on Co2+ beads (as above), washed with
TB200, and then chased with 10 mM NTPs for the indicated time inter-
vals in the presence of albumin or anti-G(trigger)-loop antibody 18B5
(1 mg/10 ml) before quenching with SB.
Crosslinking and Mapping of Crosslinked Sites
EC61 was obtained as described above and washed with TB200. UTP
(25 mM) was added for 10 min on ice. The resulting EC66 was washed
53 with TB. Incorporation of the 4-thio-uridine-50-triphosphate (40 mM;
TriLink) substrate analog along with ATP (5 mM) was performed at room
temperature for 10 min. The crosslink in the resulting trapped complex
was induced by UV irradiation at 308 nM for 10 min with a UV hand
lamp (Cole-Palmer 9815 series lamp) placed on top of an Eppendorf
tube held in ice. The samples were treated with imidazole at 150 mM
to remove the complexes from the Co2+ beads, and the supernatant
material was fractionated by SDS-PAGE followed by autoradiographic
visualization. Mapping of crosslinked sites was performed by limited
chemical degradation. Briefly, the b0 subunit carrying the radioactive
adduct was excised from the gel and eluted with 3 vols of 0.2% SDS
at 37�C for 1 hr. The eluate was freeze-dried with a SpeedVac and
redissolved in water to a final concentration of 1%–2% SDS. Protein
ell 28, 991–1001, December 28, 2007 ª2007 Elsevier Inc. 999

Molecular Cell
Mechanics of Transcription Termination
degradation effected by CNBr (at Met residues) and NTCBA (at Cys
residues) was performed according to Nudler et al. (1998). Gels con-
taining radioactive materials were scanned on a PhosphoImager.
Modeling of the Termination Complex
3D structural modeling of the termination complex was performed
starting from known structures of the yeast RNAP II ternary elongation
complex (Gnatt et al., 2001; Westover et al., 2004; Kettenberger et al.,
2004) and the Thermus thermophilus (Tth) holoenzyme (Vassylyev
et al., 2002), and from the model of the Thermus aquaticus (Taq) elon-
gation complex (Korzheva et al., 2000). Model hairpins were generated
using NMR data on the 24 nt-long RNA hairpin (Cilley and Williamson,
2003). The structures were docked and manipulated using the
Accelrys Viewer-Pro and MolSoft ICM-Pro software packages. Sin-
gle-stranded RNA segments in the complex were modeled using the
shortest sterically allowed paths; the optimization of the structure
was carried out by eliminating clashes between the RNA phosphate
backbone and the polypeptide chain and by minimizing short-range
electrostatic repulsion by inspection of the electrostatic charge distri-
bution. The elements of our structural model of termination, including
clamp-DNA interactions, G(trigger)-loop mobility, and the flexibility of
the RNA exit channel, are in a good agreement with the recently pub-
lished high-resolution crystal structureofTthEC (Vassylyev etal., 2007a).
Supplemental Data
Supplemental Data include supplemental text, nine figures, and Sup-
plemental References and can be found with this article online at
http://www.molecule.org/cgi/content/full/28/6/991/DC1/.
ACKNOWLEDGMENTS
We thank J.P. Lionetti, K. Severinov, and R. Weisberg for materials;
J. Lee for artwork; and P.H. von Hippel and N. Cowan for comments.
This work was supported by NIH grants R01 GM58750 and GM72814
(E.N.), GM54098 (S.B.), and P20 GM64375 (A.E.R.).
Received: March 31, 2007
Revised: July 18, 2007
Accepted: October 10, 2007
Published: December 27, 2007
REFERENCES
Bar-Nahum, G., Epshtein, V., Ruckenstein, A.E., Rafikov, R., Mustaev,
A., and Nudler, E. (2005). A ratchet mechanism of transcription elonga-
tion and its control. Cell 120, 183–193.
Borukhov, S., and Goldfarb, A. (1996). Purification and assay of
Escherichia coli transcript cleavage factors GreA and GreB. Methods
Enzymol. 274, 315–326.
Borukhov, S., Lee, J., and Goldfarb, A. (1991). Mapping of a contact for
the RNA 30 terminus in the largest subunit of RNA polymerase. J. Biol.
Chem. 266, 23932–23935.
Brendel, V., Hamm, G.H., and Trifonov, E.N. (1986). Terminators of
transcription with RNA polymerase from Escherichia coli: what they
look like and how to find them. J. Biomol. Struct. Dyn. 3, 705–723.
Cilley, C.D., and Williamson, J.R. (2003). Structural mimicry in the
phage phi21 N peptide-boxB RNA complex. RNA 9, 663–676.
Condon, C., Squires, C., and Squires, C.L. (1995). Control of rRNA
transcription in Escherichia coli. Microbiol. Rev. 59, 623–645.
Friedman, D.I., Imperiale, M.J., and Adhya, S.L. (1987). RNA 30 end for-
mation in the control of gene expression. Annu. Rev. Genet. 21,
453–488.
Gnatt, A.L., Cramer, P., Fu, J., Bushnell, D.A., and Kornberg, R.D.
(2001). Structural basis of transcription: an RNA polymerase II elonga-
tion complex at 3.3 A resolution. Science 292, 1876–1882.
1000 Molecular Cell 28, 991–1001, December 28, 2007 ª2007
Goliger, J.A., Yang, X.J., Guo, H.C., and Roberts, J.W. (1989). Early
transcribed sequences affect termination efficiency of Escherichia
coli RNA polymerase. J. Mol. Biol. 205, 331–341.
Gusarov, I., and Nudler, E. (1999). The mechanism of intrinsic tran-
scription termination. Mol. Cell 3, 495–504.
Gusarov, I., and Nudler, E. (2001). Control of intrinsic transcription ter-
mination by N and NusA: the basic mechanisms. Cell 107, 437–449.
Kashlev, M., and Komissarova, N. (2002). Transcription termination:
primary intermediates and secondary adducts. J. Biol. Chem. 277,
14501–14508.
Kettenberger, H., Armache, K.J., and Cramer, P. (2004). Complete
RNA polymerase II elongation complex structure and its interactions
with NTP and TFIIS. Mol. Cell 16, 955–965.
Kettenberger, H., Eisenfuhr, A., Brueckner, F., Theis, M., Famulok, M.,
and Cramer, P. (2006). Structure of an RNA polymerase II-RNA inhib-
itor complex elucidates transcription regulation by noncoding RNAs.
Nat. Struct. Mol. Biol. 13, 44–48.
King, R.A., Markov, D., Sen, R., Severinov, K., and Weisberg, R.A.
(2004). A conserved zinc binding domain in the largest subunit of
DNA-dependent RNA polymerase modulates intrinsic transcription
termination and antitermination but does not stabilize the elongation
complex. J. Mol. Biol. 342, 1143–1154.
Komissarova, N., Becker, J., Solter, S., Kireeva, M., and Kashlev, M.
(2002). Shortening of RNA:DNA hybrid in the elongation complex of
RNA polymerase is a prerequisite for transcription termination. Mol.
Cell 10, 1151–1162.
Korzheva, N., Mustaev, A., Kozlov, M., Malhotra, A., Nikiforov, V.,
Goldfarb, A., and Darst, S.A. (2000). A structural model of transcription
elongation. Science 289, 619–625.
Kuznedelov, K., Minakhin, L., Niedziela-Majka, A., Dove, S.L., Rogulja,
D., Nickels, B.E., Hochschild, A., Heyduk, T., and Severinov, K. (2002).
A role for interaction of the RNA polymerase flap domain with the
sigma subunit in promoter recognition. Science 295, 855–857.
Markovtsov, V., Mustaev, A., and Goldfarb, A. (1996). Protein-RNA in-
teractions in the active center of transcription elongation complex.
Proc. Natl. Acad. Sci. USA 93, 3221–3226.
Mironov, A.S., Gusarov, I., Rafikov, R., Lopez, L.E., Shatalin, K.,
Kreneva, R.A., Perumov, D.A., and Nudler, E. (2002). Sensing small
molecules by nascent RNA: a mechanism to control transcription in
bacteria. Cell 111, 747–756.
Nudler, E., and Mironov, A.S. (2004). The riboswitch control of bacterial
metabolism. Trends Biochem. Sci. 29, 11–17.
Nudler, E., Kashlev, M., Nikiforov, V., and Goldfarb, A. (1995). Coupling
between transcription termination and RNA polymerase inchworming.
Cell 81, 351–357.
Nudler, E., Gusarov, I., Avetissova, E., Kozlov, M., and Goldfarb, A.
(1998). Spatial organization of transcription elongation complex in
Escherichia coli. Science 281, 424–428.
Nudler, E., Gusarov, I., and Bar-Nahum, G. (2003). Methods of walking
with the RNA polymerase. Methods Enzymol. 371, 160–169.
Platt, T. (1986). Transcription termination and the regulation of gene
expression. Annu. Rev. Biochem. 55, 339–372.
Rees, W.A., Weitzel, S.E., Das, A., and von Hippel, P.H. (1997). Regu-
lation of the elongation-termination decision at intrinsic terminators by
antitermination protein N of phage lambda. J. Mol. Biol. 273, 797–813.
Rouby, J., Pugniere, M., Mani, J.C., Granier, C., Monmouton, P., Theu-
lier Saint Germain, S., and Leonetti, J.P. (2002). Characterization of
monoclonal antibodies against Escherichia coli core RNA polymerase.
Biochem. J. 361, 347–354.
Santangelo, T.J., and Roberts, J.W. (2004). Forward translocation is
the natural pathway of RNA release at an intrinsic terminator. Mol.
Cell 14, 117–126.
Elsevier Inc.

Molecular Cell
Mechanics of Transcription Termination
Tadigotla, V.R., O’Maoileidigh, D., Sengupta, M.A., Epshtein, V.,
Ebright, R.H., Nudler, E., and Ruckenstein, A.E. (2006). Thermodynam-
ics and kinetics based identification of transcriptional pauses. Proc.
Natl. Acad. Sci. USA 103, 4439–4444.
Telesnitsky, A.P., and Chamberlin, M.J. (1989). Sequences linked to
prokaryotic promoters can affect the efficiency of downstream termi-
nation sites. J. Mol. Biol. 205, 315–330.
Toulokhonov, I., and Landick, R. (2006). The role of the lid element in
transcription by E. coli RNA polymerase. J. Mol. Biol. 361, 644–658.
Toulokhonov, I., Artsimovitch, I., and Landick, R. (2001). Allosteric con-
trol of RNA polymerase by a site that contacts nascent RNA hairpins.
Science 292, 730–733.
Toulokhonov, I., Zhang, J., Palangat, M., and Landick, R. (2007). A
central role of the RNA polymerase trigger loop in active-site rear-
rangement during transcriptional pausing. Mol. Cell 27, 406–419.
Vassylyev, D.G., Sekine, S., Laptenko, O., Lee, J., Vassylyeva, M.N.,
Borukhov, S., and Yokoyama, S. (2002). Crystal structure of a bacterial
RNA polymerase holoenzyme at 2.6 A resolution. Nature 417,
712–719.
Molecular Ce
Vassylyev, D.G., Vassylyeva, M.N., Perederina, A., Tahirov, T.H., and
Artsimovitch, I. (2007a). Structural basis for transcription elongation
by bacterial RNA polymerase. Nature 448, 157–162.
Vassylyev, D.G., Vassylyeva, M.N., Zhang, J., Palangat, M., Artsimo-
vitch, I., and Landick, R. (2007b). Structural basis for substrate loading
in bacterial RNA polymerase. Nature 448, 163–168.
Wang, D., Bushnell, D.A., Westover, K.D., Kaplan, C.D., and Kornberg,
R.D. (2006). Structural basis of transcription: role of the trigger loop in
substrate specificity and catalysis. Cell 127, 941–954.
Weisberg, R.A., and Gottesman, M.E. (1999). Processive antitermina-
tion. J. Bacteriol. 181, 359–367.
Westover, K.D., Bushnell, D.A., and Kornberg, R.D. (2004). Structural
basis of transcription: nucleotide selection by rotation in the RNA poly-
merase II active center. Cell 119, 481–489.
Wright, D.J., King, K., and Modrich, P. (1989). The negative charge of
Glu-111 is required to activate the cleavage center of EcoRI endonu-
clease. J. Biol. Chem. 264, 11816–11821.
Yarnell, W.S., and Roberts, J.W. (1999). Mechanism of intrinsic tran-
scription termination and antitermination. Science 284, 611–615.
ll 28, 991–1001, December 28, 2007 ª2007 Elsevier Inc. 1001
Copyright © 2022 FDOKUMEN




















