
COVER SHEET for SEC FORM 20-IS SEC Registration Number 3 4 ...

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
PROOF COVER SHEET
Author(s): Amr S. Deaf, Ian C. Harding and John E. A. Marshall
Article title: Cretaceous (Albian–early? Santonian) palynology and stratigraphy of the Abu Tunis 1xborehole, northern Western Desert, Egypt
Article no: 828662
Enclosures: 1) Query sheet2) Article proofs
Journal Acronym: TPAL
Dear Author,
1. Please check these proofs carefully. It is the responsibility of the corresponding author to check these and approveor amend them. A second proof is not normally provided. Taylor & Francis cannot be held responsible for uncor-rected errors, even if introduced during the production process. Once your corrections have been added to the arti-cle, it will be considered ready for publication
Please limit changes at this stage to the correction of errors. You should not make insignificant changes, improveprose style, add new material, or delete existing material at this stage. Making a large number of small, non-essentialcorrections can lead to errors being introduced. We therefore reserve the right not to make such corrections.
For detailed guidance on how to check your proofs, please see http://journalauthors.tandf.co.uk/production/checkingproofs.asp.
2. Please review the table of contributors below and confirm that the first and last names are structured correctly andthat the authors are listed in the correct order of contribution. This check is to ensure that your name will appear cor-rectly online and when the article is indexed.
Sequence Prefix Given name(s) Surname Suffix
1 Amr S. Deaf
2 Ian C. Harding
3 John E. A. Marshall

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
Queries are marked in the margins of the proofs.
AUTHOR QUERIES
General query: You have warranted that you have secured the necessary written permission from the appropriatecopyright owner for the reproduction of any text, illustration, or other material in your article.(Please see http://journalauthors.tandf.co.uk/preparation/permission.asp.) Please check that any required acknowl-edgements have been included to reflect this.
Q1. AU: Please confirm that you are referring to Figure 1 here.
Q2. AU: The reference [Said 1990] is cited in the text but is not listed in the references list. Please either delete in-
text citation or provide full reference details following journal style [hyperlink to relevant style-sheet].
Q3. AU: Please spell out what “WEPCO” means on first use in the text.
Q4. AU: The reference [Stockmarr 1971] is cited in the text but is not listed in the references list. Please either
delete in-text citation or provide full reference details following journal style [hyperlink to relevant style-sheet].
Q5. AU: Please indicate which figure you are referring to here.Q6. AU: The reference [Schrank 2001] is cited in the text but is not listed in the references list. Please either delete
in-text citation or provide full reference details following journal style [hyperlink to relevant style-sheet].
Q7. AU: Please spell out what “NOCS” means.
Q8. AU: Please provide a short biography of each author.
Q9. AU: The reference [Brideaux 1971] is listed in the references list but is not cited in the text. Please either cite
the reference or remove it from the references list.
Q10. AU: Please provide missing publisher for the [Norton 1967] references list entry.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
How to make corrections to your proofs using Adobe Acrobat
Taylor & Francis now offer you a choice of options to help you make corrections to your proofs. Your PDF proof
file has been enabled so that you can edit the proof directly using Adobe Acrobat. This is the simplest and best way
for you to ensure that your corrections will be incorporated. If you wish to do this, please follow these instructions:
1. Save the file to your hard disk.
2. Check which version of Adobe Acrobat you have on your computer. You can do this by clicking on the “Help”
tab, and then “About.”
If Adobe Reader is not installed, you can get the latest version free from http://get.adobe.com/reader/.
� If you have Adobe Reader 8 (or a later version), go to “Tools”/ “Comments & Markup”/ “Show Comments &
Markup.”
� If you have Acrobat Professional 7, go to “Tools”/ “Commenting”/ “Show Commenting Toolbar.”
3. Click “Text Edits.” You can then select any text and delete it, replace it, or insert new text as you need to. If you
need to include new sections of text, it is also possible to add a comment to the proofs. To do this, use the StickyNote tool in the task bar. Please also see our FAQs here: http://journalauthors.tandf.co.uk/production/index.asp.
4. Make sure that you save the file when you close the document before uploading it to CATS using the “Upload
File” button on the online correction form. A full list of the comments and edits you have made can be viewed by
clicking on the “Comments” tab in the bottom left-hand corner of the PDF.
If you prefer, you can make your corrections using the CATS online correction form.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
5
10
15
20
25
30
30
35
40
45
50
55
60
65
Cretaceous (Albian–early? Santonian) palynology and stratigraphy of the Abu Tunis 1x borehole,northern Western Desert, Egypt
Amr S. Deafa,b*, Ian C. Hardinga and John E. A. Marshalla
aSchool of Ocean & Earth Science, National Oceanography Centre, Southampton (NOCS), University of Southampton, EuropeanWay, Southampton, SO14 3ZH, UK; bGeology Department, Faculty of Science, Assiut University, Assiut, 71516, Egypt
A palynological analysis has been conducted on the middle-upper Cretaceous sediments of the Abu Tunis 1xborehole, in the northern Western Desert, Egypt. The recovered palynomorphs have been analysed both qualita-tively and quantitatively and permit a refinement of the original stratigraphy with the identification of four time-rock units – these have been divided into four informal sporomorph and one dinoflagellate cyst palynozones. Thesebiozones are, from oldest to youngest: the Afropollis jardinus-Tricolporopollenites-Elaterosporites klaszii AssemblageZone (early–mid Albian), the Elaterosporites verrucatus-Sofrepites legouxae-Cretacaeiporites Assemblage Zone (lateAlbian–early Cenomanian), the Sofrepites legouxae Partial Range Zone (early–?mid Cenomanian), the Proteaciditescf. africaensis Total Range Zone (mid–late Cenomanian), and the Canningia senonica Total Range Zone (early?Santonian). A barren interzone has been identified just below the youngest palynozone, and this may be related tothe unfavourable lithology (i.e., limestone and dolostone).
The absolute abundance spore and pollen data represents the first quantitative description of an EgyptianAlbian–Cenomanian palynoflora, a flora that is characteristic of the Albian–Cenomanian Elaterate PhytogeographicProvince. The early Santonian palynoflora is exclusively marine phytoplankton; terrestrial palynomorphsrepresentative of the Senonian Palmae Province are completely absent. The quantitative and semi-quantitativedistributions of Afropollis jardinus have been compared with similar semi-quantitative distributions of this speciesfrom other wells in the north Western Desert of Egypt, and this permitted the identification of a mid Albian–earlyCenomanian Afropollis jardinus ‘acme’ as an important local biostratigraphic event in the mid Cretaceous.
Keywords: Cretaceous; palynology; stratigraphy; Western Desert; Egypt
1. Introduction
Egypt is currently located in the subtropical arid zone
and thus all its exposed sediments have been subjected
to extensive and deep weathering and are therefore paly-nologically barren. Most of the palynological research
conducted on Egyptian Cretaceous successions has been
based on deep borehole samples taken from exploratory
boreholes (e.g., Abdelmalik et al. 1981; Penny 1986;
Omran et al. 1990; Schrank & Ibrahim 1995; Mahmoud
& Deaf 2007; El Soughier et al. 2010), which were
drilled during hydrocarbon exploration activity in the
northern part of the EgyptianWestern Desert.The middle Cretaceous (Albian–Cenomanian) mixed
clastic-carbonate rocks represented by the Kharita,
Bahariya and lower Abu Roash formations of the north-
ern basinal area of Egypt were mainly deposited in very
shallow marine (brackish to coastal) to inner neritic open
marine conditions (Hantar 1990; Kerdany & Cherif
1990). These conditions were unfavourable for the prolif-
eration of planktonic forams and calcareous nannofos-sils and, thus, no independent age control is available, at
least for the Kharita succession. The upper Cretaceous
interval is generally composed of pre-Campanian (Turo-
nian–Santonian) middle to upper shelf deposits (upper
Abu Roash and Khoman ‘B’ formations), and of deeper
upper to middle slope deposits (Khoman ‘A’ Formation)
for the Campanian–Maastrichtian interval, which is
mainly represented by thick carbonate successions(Hantar 1990; Kerdany & Cherif 1990). Most of the
independently calibrated palynological work has been
produced as unpublished internal reports by oil explora-
tion companies. Thus the planktonic foraminifera-cali-
brated palynological work of Schrank and Ibrahim
(1995) and that of Abdel-Kireem et al. (1996) on upper
Cretaceous (Cenomanian–Maastrichtian) samples from
the Kahraman-1 and Abu Gharadig-1 boreholes in thenorthern Western Desert of Egypt is one of the few
attempts to provide micropalaeontologically calibrated
palynological work. Our own attempts to provide an
independent calibration using calcareous nannofossils
has proven unsuccessful: smear slides made from 10 sam-
ples of the carbonate succession of the Abu Tunis 1x
borehole revealed only very rare specimens which were
considered as of no biostratigraphic significance.
*Corresponding authors. Email: [email protected]; [email protected]
� 2013 AASP – The Palynological Society
Palynology, 2013
Vol. 00, No. 00, 1–27, http://dx.doi.org/10.1080/01916122.2013.828662

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
70
75
80
85
90
95
100
105
110
Thus in most of the Egyptian palynological work,
palynostratigraphers have correlated different palyno-
morph assemblages with similar ones from other palaeo-
geographically related areas in order to date theEgyptian Cretaceous successions. As a result, several
informal, yet sometimes concept-incompatible (i.e., acme
versus total range) palynological zonal schemes (e.g., El-
Shamma 1991; Aboul Ela & Mahrous 1992) have been
proposed for different parts of the Cretaceous record of
Egypt. The informal zones proposed by Schrank and
Ibrahim (1995) represent the most complete palynologi-
cal zonal scheme for the Egyptian middle–upper Creta-ceous sedimentary sequence. Our zonal scheme uses
index taxa that have been recorded in the same phyto-
geographic province and have micropalaeontologically
calibrated age ranges. By using the same definitions as
Schrank and Ibrahim (1995), an attempt has been made
to develop a unified zonal scheme for the middle–upper
Cretaceous of the northernWestern Desert.
The goals of this project were to: (i) integrate lithos-tratigraphic and biostratigraphic schemes for the Abu
Tunis 1x borehole; (ii) propose an informal unified paly-
nozonal scheme for the middle–upper Cretaceous of the
northern Western Desert; (iii) correlate biostratigraphic
results with those from other palaeogeographically
related areas, such as the African and South American
phytogeographic provinces for sporomorphs, and the
Tethyan Realm for dinoflagellate cysts; (iv) provide thefirst quantitative description of the Egyptian Albian–
Cenomanian palynoflora, and (v) complete a taxonomic
study of all of the recorded taxa, especially of the strati-
graphically significant forms.
2. Geological setting and lithostratigraphic history
The north Western Desert is a vast area representing
the western part of a tectonically active structural
province referred to as ‘the unstable shelf’ by Said
(1962; Figure 1 Q1). Tectonism has played a major role in
the development and closure of several small rift basins
in this area. The oldest of these basins was developedin the late Palaeozoic, most probably during Late
Permian times, but most of these basins developed
around the earliest Cretaceous (Guiraud 1998;
Guiraud et al. 2001), with late Santonian folding caus-
ing inversion in most of the basins (Guiraud 1998).
The Western Desert is characterized by a northward
dipping plain comprising Eocene and Miocene
Figure 1. Simplified structural map of Egypt showing the location of the Abu Tunis 1x borehole (modified after Kerdany andCherif, 1990).
B=w in print; colour online
2 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
115
120
125
130
135
140
145
150
155
160
165
170
175
180
185
190
195
200
205
210
215
carbonates, covered by Pliocene and Quaternary sedi-
ments along the Egyptian Mediterranean coast and the
Nile Delta areas (Hantar 1990).
Deposition of the upper lower Cretaceous sedimentsoccurred during a regressive phase, and fluvial processes
deposited the Albian Kharita Formation, which domi-
nates a large part of the Western Desert (Said 1990).Q2 In
contrast, the upper Cretaceous sediments indicate a
major transgressive phase. During the late Cenoma-
nian, regional subsidence related to Neotethyan rifting
took place across the northern African margin, and a
marine transgression covered the entire northernAfrican plate. As a result, transitional fluvial to shallow
marine deposits of the Bahariya Formation accumu-
lated in the Western Desert. During the Turonian,
marine conditions generally persisted across most of the
Western Desert, where a thick carbonate succession of
the the Abu Roash Formation was deposited in the
extreme northern Western Desert (Said 1990), where
the Abu Tunis 1x borehole is located. However, the lateTuronian Laramide tectonic event caused uplift and
basin inversion in some of the basins in the northern
Western Desert (Said 1990), and probably affected at
least the central part of the Matruh Basin in which the
Abu Tunis 1x borehole is located. However, by Conia-
cian times most of the northern Western Desert became
inundated by another marine transgression, during
which carbonates of the Khoman ‘B’ Formation weredeposited (Said 1990). By the late Santonian, right lat-
eral movement between Africa and Laurasia resulted in
northwest-directed compressive forces across the north-
eastern margin of the African plate (Meshref 1990;
Guiraud et al. 2001). These compressive forces in turn
resulted in a series of N-W folds (i.e., the Syrian Arc
System) associated with thrust faults across the north-
ern Western Desert (Meshref 1990; Guiraud et al.2001). As a result, the Santonian witnessed another
regression; continuing deposition of the lower Khoman
‘B’ Formation in the extreme north gave way south-
ward to marine clastic sedimentation across theWestern
Desert. During Campanian–Maastrichtian time, exten-
sion and subsidence dominated the northern Eastern
African margin (Guiraud & Bosworth 1999; Guiraud
et al. 2001), and most of the Western Desert was againcovered by deep marine waters, resulting in deposition
of a thick sequence of chalky limestone known as the
Khoman ‘A’ Formation (Said 1990).
3. Material and methods
Seventy-seven ditch cutting samples were collected
from between 7300 and 3500 ft (2225–1067 m) in theAbu Tunis 1x borehole. The borehole was drilled in the
Faghur area, at the centre of the Matruh Basin east of
the Faghur-Maamura High and north of the Umbarka
Subbasin: Lat. 31� 160 0800 N, and Long. 26� 500 4100 Eby WEPCO Q3(1968). A summary of the palynological
samples used with depths and total recovery of palyno-
morphs is provided in Appendix 1.Palynological processing involved acid digestion
using 36% hydrochloric acid (HCl) and 60% hydro-
fluoric acid (HF) (e.g., Phipps & Playford 1984; Green
2001). One tablet of a known quantity (12,542 grains/
tablet with V � 3.3%) of modern Lycopodium spores
(made by the Department of Quaternary Geology,
Lund University, batch no. 124961) was added, for
absolute abundance analysis, to each sample after thefirst HF decanting. Samples were boiled in 15 ml of
36% HCl for 1–2 minutes after being washed and
sieved through a 15-mm mesh to remove neo-formed
fluorides. Two permanent microscope slides were pre-
pared using Elvacite 2044 as a mounting medium.
Slides, residues and samples are stored in the Geologi-
cal Museum, Geology Department, Faculty of Science,
Assiut University, Egypt.Processed samples yielded well-preserved and
diverse assemblages of miospore-dominated palyno-
morphs and less frequent occurrences of lower-diver-
sity dinoflagellate cysts. Qualitative analysis was
carried out using an Olympus (BX41) transmitted-light
microscope (serial no. 8B25715) equipped with an
Infinity-1 digital camera for imaging. An alphabetical
list of the palynomorphs recorded in the Abu Tunis 1xis shown in Appendix 2. Quantitative analysis (grains/
gram) was determined by applying in the formula of
Stockmarr (1971) Q4as follows:
c ¼ Sc � Lt � t
Lc � wð1Þ
wherec ¼ concentration ¼ total number of specimens/
gram dried sediments
Sc ¼ number of specimens counted
Lt ¼ number of Lycopodium spores/tablet
t ¼ number of tablets added to the sample
Lc ¼ number of Lycopodium spores counted
w ¼ weight of dried sediments (g)
The first 250 grains were chosen for count as they pro-
vide a total maximum error of 7% in grain concentra-
tion according to the curve of Stockmarr (1971). To
detect any rare species, a further scan of the rest of
each mounted sample residue was also made.
4. Palynostratigraphy
4.1. Introductory comments
A careful review of Egyptian palynological literature
has shown that some of the previously proposed age
Palynology 3

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
220
225
230
235
240
245
250
255
260
265
270
275
280
285
290
295
300
305
310
315
320
assignments have been misinterpreted because they
refer to species ranges, which have no independent age
control, or to sedimentary sequences of doubtful ages
or to processing cuttings with their inevitable cavingseffect. Therefore, dating of the Abu Tunis 1x borehole
was accomplished using independently calibrated
age ranges of index taxa compiled from the literature
(Figure 2), in addition to correlation with accurately-
dated contemporaneous regional and interregional
palynofloral assemblages. Correlation with interconti-
nental palynofloral assemblages was made in
the context of the Albian–Cenomanian ElateratesPhytogeographic Province of Herngreen et al. (1996).
In terms of the dinoflagellate cyst biostratigraphy,
it is important to note that many of the independently
calibrated events in the Albian of the European Tethys
(Davey & Verdier 1973; Habib & Drugg 1983; Erba
et al. 1999; Torricelli 2006) cannot be recognized in the
southern Tethyan region (e.g., Libya and Egypt), pos-
sibly due to environmental exclusion.Schrank and Ibrahim (1995) argued for early late
Cretaceous (Turonian–Coniacian) diachroneity of the
biostratigraphic units in terms of their concept and char-
acteristics in northeast Africa andWest Africa. This was
based on these authors extending the range of Droseri-
dites senonicus down into the late Turonian of Egypt
based on independent foraminifera-based dating. Whilst
the biostratigraphic units are defined differently (i.e.,acme zone versus total range), the palynological species
stratigraphic ranges are not at variance. This late Turo-
nian lower limit of Droseridites senonicus has also been
recorded in the foraminifera-dated late Turonian of
northeast Nigeria (Lawal & Moullade 1986). The inter-
regional correlation of biostratigraphic ranges and/or
successive events of selected index angiosperm (e.g.,
Cretacaeiporites) and gymnosperm (e.g., Droseridites
senonicus) pollen between the foraminifera-dated Ceno-
manian–Coniacian sedimentary sequence of North and
West Africa (Senegal and Ivory Coast: Jardin�e &
Magloire 1965; Egypt: Schrank & Ibrahim 1995;
Ibrahim 1996) shows a close match between these two
regions (Figure 3) and does not support the proposed
stratigraphic diachroneity. A greater similarity between
the palynofloral assemblage compositions and species’stratigraphic ranges could be proposed in the light of
African plate movement during the Cretaceous. This
tectonic plate was moving anticlockwise towards Laura-
sia, as a result of the opening of the southern Atlantic
Ocean, bringing formerly tropical regions into a more
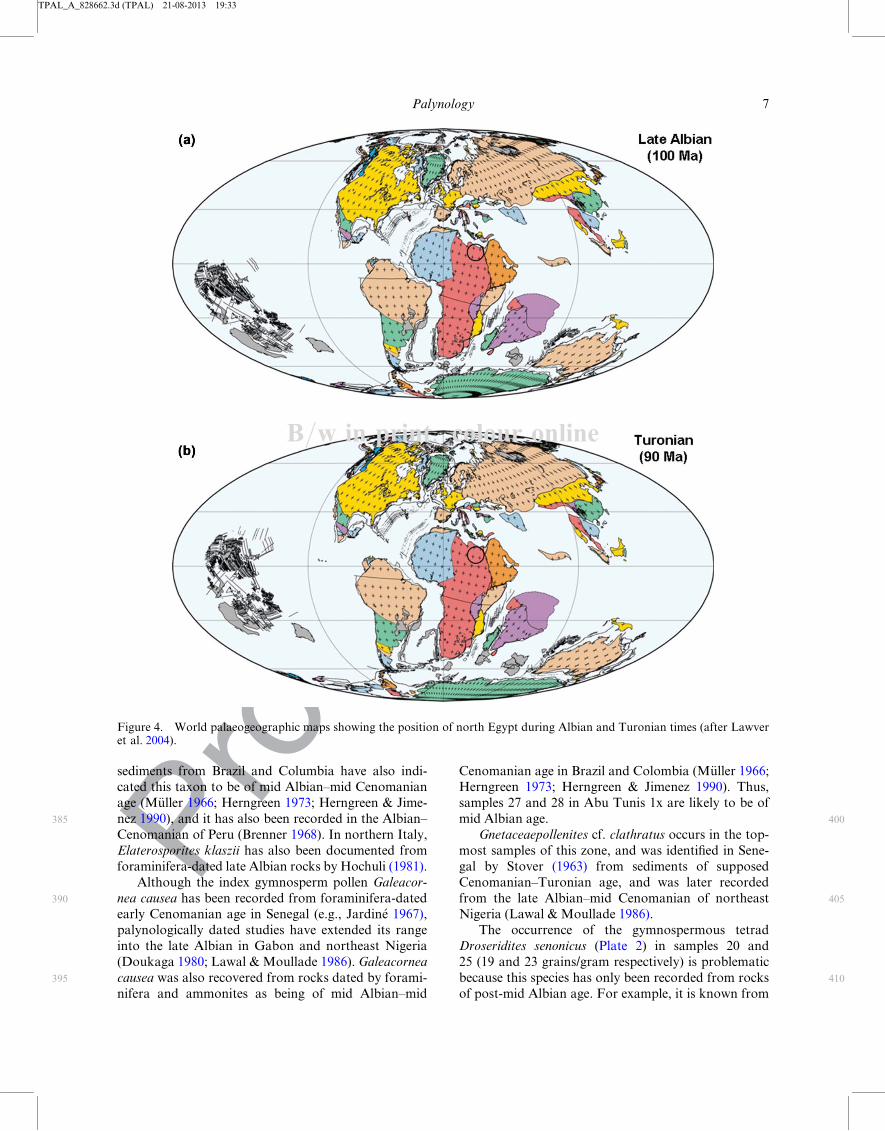
subtropical position by the late Cretaceous (Figure 4).
Despite the fact that Morocco is confined to the
Albian–Cenomanian Elaterate Phytogeographic Prov-ince, however, some differences in the palynofloral
assemblages have been noted. In the Agadir-Essaouira
Basin and in the southwestern Tarfaya Basin, the
Albian–Cenomanian palynoflora exhibits Albian–
Cenomanian Elaterate Province characteristics, but
contains some temperate pollen grains, for example
Alisporites, Podocarpidites, Vitreisporites and Cerebro-
pollenites (Bettar & M�eon 2001, 2006) which are char-
acteristic of the Cerebropollenites Province of
Herngreen et al. (1996). Therefore, the micropalaeon-
tologically dated palynological work carried out in
West Africa (Jardin�e and Magloire, 1965: Senegal and
the Ivory Coast; Boltenhagen 1980: Gabon; Lawal &
Moullade 1986: Nigeria) which shows pure Albian–
Cenomanian Elaterate Province characteristics will beemployed here for assigning dates in the mid and late
Cretaceous sample interval. The local, partly foramin-
iferally controlled post-Cenomanian palynological
work of Schrank and Ibrahim (1995) will also be
followed.
The spore and pollen grain ranges have been used
for biostratigraphic purposes, with dinoflagellate cysts
providing additional supporting evidence. Dinoflagel-late cyst taxa have been found to be facies-controlled
in the middle Cretaceous sediments (Thusu et al.
1988), and additionally they are extremely diluted by
an abundance of microforaminiferal test linings in the
upper Cretaceous carbonate sediments. However, the
dinoflagellate cysts are the only biostratigraphic tools
that can be used to date the upper part of the carbonate
succession. Biozonation and age assessment of thestudied samples was based on the vertical quantitative
distributions by highest occurrence (HO) of all taxa
recovered (Figure 5). Lowest occurrences (LO) of
index sporomorphs were used to define biostrati-
graphic units where possible, but HO were used when
suitable LO were lacking. The numbers given in paren-
theses after the names of the taxa refer to the position
of these species in the quantitative range chart(Figure 5).
4.2. Palynological zonation (PZ) and age assessments
PZ1: Afropollis jardinus-Tricolporopollenites-
Elaterosporites klaszii Assemblage Zone (early–mid
Albian)
Samples: This zone includes samples 1 to 28, and spansa depth from 7300 to 5950 ft (2225–1814 m).
Definition: From the LO of Afropollis jardinus (36) to
just below the LO of Sofrepites legouxae (26) and Ela-
terosporites verrucatus (27).
Associated taxa: Verrucosisporites obscurilaesuratus (8),
Galeacornea causea (31), Ephedripites irregularis (33),
Rousea delicipollis (44), Triporites sp. (46), Retimonocol-
pites textus (49), Stellatopollis densiornatus (62),Afropol-lis aff. jardinus (66), Tetracolpites sp. (67), Senegalinium
aenigmaticum (85), Trichodinium castanea (87), Coroni-
fera oceanica (97), Oligosphaeridium albertense (105),
4 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
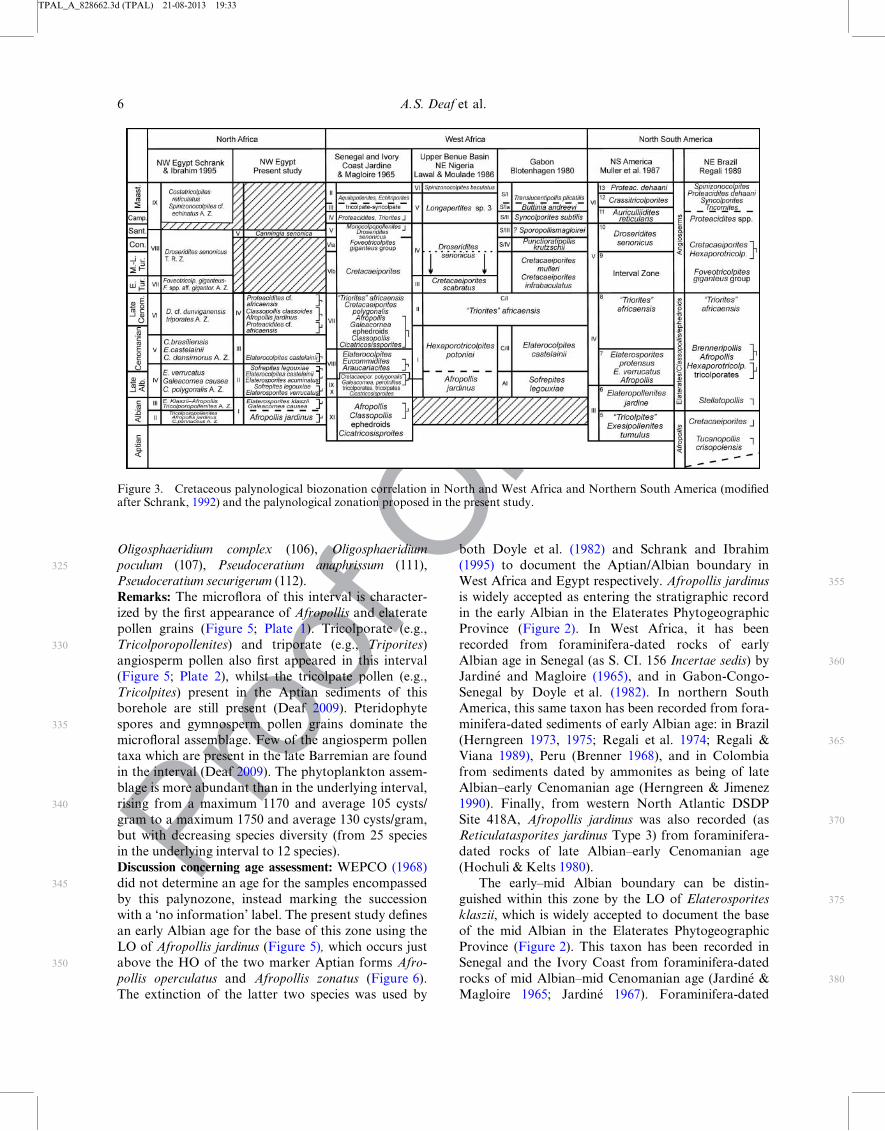
Figure 2. Compilation of the biostratigraphic range of most of the important Cretaceous marker species in different phytogeo-graphic provinces of North andWest Africa and north South America and the Tethyan Realm. Sources for African ranges: Doyleet al. 1982 (1); Hochuli 1981 (4, 11, 12); Hochuli and Kelts 1980 (1); Jan du Chene et al. (1978) (22, 25); Jardin�e 1967 (4, 5, 6, 8, 9,10, 11, 12, 14); Jardin�e and Magloire 1965 (1, 3, 4, 5, 6, 7, 8, 9, 13, 14, 15, 16, 17, 19, 21, 26); Lawal and Moullade 1986 (6, 7, 13,14, 15, 16, 17, 18, 22, 23, 24, 25, 26); Schrank and Ibrahim 1995 (2, 4, 6, 10, 11, 13, 16, 17, 20); (Tea-Yassi et al.) 1999 (18, 22, 23,24); Thusu and Van Der Eem (1985) (13). Sources for NS American ranges: Brenner 1968 (4, 6, 12); Herngreen 1973 (2, 3, 4, 5, 6,7, 8, 10, 11, 12, 13, 14); Herngreen 1975 (10); Herngreen and Due~naz Jimenez 1990 (12, 13, 14, 17, 19, 20, 21); Muller 1966 (1, 4,5, 12); Regali and Viana 1989 (2, 6); Regali et al. 1974 (1, 3, 5, 26). Sources for Tethyan ranges: Clark and Verdier 1967 (28, 30,31); Jan Du Ch�ene et al. 1978 (33); Roncaglia and Corradini 1997 (34); Schrank and Ibrahim 1995 (28, 29, 30, 31, 32, 33, 34);Torricelli and Amore 2003 (30, 31, 32, 34).
Palynology 5
TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
325
330
335
340
345
350
355
360
365
370
375
380
Oligosphaeridium complex (106), Oligosphaeridium
poculum (107), Pseudoceratium anaphrissum (111),Pseudoceratium securigerum (112).
Remarks: The microflora of this interval is character-
ized by the first appearance of Afropollis and elaterate
pollen grains (Figure 5; Plate 1). Tricolporate (e.g.,
Tricolporopollenites) and triporate (e.g., Triporites)
angiosperm pollen also first appeared in this interval
(Figure 5; Plate 2), whilst the tricolpate pollen (e.g.,
Tricolpites) present in the Aptian sediments of thisborehole are still present (Deaf 2009). Pteridophyte
spores and gymnosperm pollen grains dominate the
microfloral assemblage. Few of the angiosperm pollen
taxa which are present in the late Barremian are found
in the interval (Deaf 2009). The phytoplankton assem-
blage is more abundant than in the underlying interval,
rising from a maximum 1170 and average 105 cysts/
gram to a maximum 1750 and average 130 cysts/gram,but with decreasing species diversity (from 25 species
in the underlying interval to 12 species).
Discussion concerning age assessment: WEPCO (1968)
did not determine an age for the samples encompassed
by this palynozone, instead marking the succession
with a ‘no information’ label. The present study defines
an early Albian age for the base of this zone using the
LO of Afropollis jardinus (Figure 5), which occurs justabove the HO of the two marker Aptian forms Afro-
pollis operculatus and Afropollis zonatus (Figure 6).
The extinction of the latter two species was used by
both Doyle et al. (1982) and Schrank and Ibrahim
(1995) to document the Aptian/Albian boundary inWest Africa and Egypt respectively. Afropollis jardinus
is widely accepted as entering the stratigraphic record
in the early Albian in the Elaterates Phytogeographic
Province (Figure 2). In West Africa, it has been
recorded from foraminifera-dated rocks of early
Albian age in Senegal (as S. CI. 156 Incertae sedis) by
Jardin�e and Magloire (1965), and in Gabon-Congo-
Senegal by Doyle et al. (1982). In northern SouthAmerica, this same taxon has been recorded from fora-
minifera-dated sediments of early Albian age: in Brazil
(Herngreen 1973, 1975; Regali et al. 1974; Regali &
Viana 1989), Peru (Brenner 1968), and in Colombia
from sediments dated by ammonites as being of late
Albian–early Cenomanian age (Herngreen & Jimenez
1990). Finally, from western North Atlantic DSDP
Site 418A, Afropollis jardinus was also recorded (asReticulatasporites jardinus Type 3) from foraminifera-
dated rocks of late Albian–early Cenomanian age
(Hochuli & Kelts 1980).
The early–mid Albian boundary can be distin-
guished within this zone by the LO of Elaterosporites
klaszii, which is widely accepted to document the base
of the mid Albian in the Elaterates Phytogeographic
Province (Figure 2). This taxon has been recorded inSenegal and the Ivory Coast from foraminifera-dated
rocks of mid Albian–mid Cenomanian age (Jardin�e &
Magloire 1965; Jardin�e 1967). Foraminifera-dated
Figure 3. Cretaceous palynological biozonation correlation in North and West Africa and Northern South America (modifiedafter Schrank, 1992) and the palynological zonation proposed in the present study.
6 A.S. Deaf et al.
TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
sediments from Brazil and Columbia have also indi-cated this taxon to be of mid Albian–mid Cenomanian
age (M€uller 1966; Herngreen 1973; Herngreen & Jime-
nez 1990), and it has also been recorded in the Albian–
Cenomanian of Peru (Brenner 1968). In northern Italy,
Elaterosporites klaszii has also been documented from
foraminifera-dated late Albian rocks by Hochuli (1981).
Although the index gymnosperm pollen Galeacor-
nea causea has been recorded from foraminifera-datedearly Cenomanian age in Senegal (e.g., Jardin�e 1967),
palynologically dated studies have extended its range
into the late Albian in Gabon and northeast Nigeria
(Doukaga 1980; Lawal & Moullade 1986). Galeacornea
causea was also recovered from rocks dated by forami-
nifera and ammonites as being of mid Albian–mid
Cenomanian age in Brazil and Colombia (M€uller 1966;Herngreen 1973; Herngreen & Jimenez 1990). Thus,
samples 27 and 28 in Abu Tunis 1x are likely to be of
mid Albian age.
Gnetaceaepollenites cf. clathratus occurs in the top-
most samples of this zone, and was identified in Sene-
gal by Stover (1963) from sediments of supposed
Cenomanian–Turonian age, and was later recorded
from the late Albian–mid Cenomanian of northeastNigeria (Lawal & Moullade 1986).
The occurrence of the gymnospermous tetrad
Droseridites senonicus (Plate 2) in samples 20 and
25 (19 and 23 grains/gram respectively) is problematic
because this species has only been recorded from rocks
of post-mid Albian age. For example, it is known from
385
390
395
400
405
410
Figure 4. World palaeogeographic maps showing the position of north Egypt during Albian and Turonian times (after Lawveret al. 2004).
B=w in print; colour online
Palynology 7

TPAL_A_828662.3d (TPAL) 21-08-2013 19:33
foraminifera-controlled Coniacian–Santonian sequen-
ces in northeast Nigeria and the Angola Basin
(Morgan 1978; Lawal & Moullade 1986), and in Egypt
from rocks also dated by foraminifera as of lateTuronian–early Santonian age (Schrank & Ibrahim
1995). The older occurrences of this taxon in Abu
Tunis 1x may be due to caving.
Based on the data above, an early to mid Albian
age is justified for this sample interval. Taken
together, the early–mid Albian age and dominantly
sandstone lithology of samples 1 to 28 suggests thatthese samples represent the lower Kharita Formation
(Figure 6), a clastic Albian–Cenomanian unit com-
posed of fine- to coarse-grained sandstones with
415
420
425
Figure 5. Quantitative (grains/gram) distribution chart by highest appearance of the recovered palynomorphs of the Abu Tunis1x borehole.
8 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:34
subordinate shale and carbonate interbeds in the
north Western Desert (Hantar 1990; Kerdany &
Cherif 1990).Correlation: This zone resembles Sequence X and XI
(early–mid Albian) identified by Jardin�e and Magloire
(1965) in the Senegal Basin, and corresponds exactly to
Zone I (early–mid Albian) of Herngreen (1973) in the
Maranh~ao Basin, Brazil. There are similar zones
defined in the northern Western Desert of Egypt: Zone
I of Sultan and Aly (1986), in the WD-9-15-1 well and
Zones II–III of Schrank and Ibrahim (1995), in theKahraman-1 well. Zone 2 (mid Albian) of Ibrahim
(1996), in the Ghazalat-1 well, corresponds to the
upper part of the current zone. In contrast, the Albian
Zone PSII of Mahmoud and Moawad (2002) in the
Sanhur-1X well very possibly corresponds only to thelower part of the current zone (i.e., early Albian),
because it lacks the mid Albian index form Elaterospor-
ites klaszii.
PZ2: Elaterosporites verrucatus-Sofrepites legouxae-
Cretacaeiporites Assemblage Zone (late Albian–early
Cenomanian)
Samples: This zone includes samples 29 to 39, which
cover a depth from 5900 to 5050 ft (1798–1539 m).
430
435
440
445
Figure 5. Continued
B=w in print; colour online
Palynology 9

TPAL_A_828662.3d (TPAL) 21-08-2013 19:34
Definition: From the LO of Sofrepites legouxae (26)
and Elaterosporites verrucatus (27) to HO of Sofrepiteslegouxae (just above the HO of Elaterosporites
verrucatus).
Associated taxa: Classopollis classoides (19), Ephedri-
pites spp. (20), Araucariacites australis (24), Afropollis
jardinus (36), Tetraporopollenites sp. (39), Rousea deli-
cipollis (44), Tricolpites vulgaris (48), Retimonocolpites
textus (49), Rousea brenneri (50), Tricolpites parvus
(51), Triporopollenites spp. (53), Tricolpites micromunus
(54), Dichastopollenites ghazalatensis (55), Tricolpites
cf. crassimurus (56), Tricolpites sagax (58),
Retimonocolpites ghazalii (59), Stephanocolpites sp.
(60), Stellatopollis barghoornii (61), Stellatopollis den-
siornatus (62), Striatopollis cf. trochuensis (63) Tricho-
dinium castanea (87), Xiphophoridium alatum (95),
Cribroperidinium edwardsii (108).
Remarks: The microfloral assemblage of this interval is
characterised by elaterate pollen grains and a notice-
able increase in abundance of the tricolpate angio-
sperm pollen. The genus Afropollis also shows a
significant increase from a maximum 1500 (average�400) grains/gram in the underlying interval to a maxi-
mum of 5375 (average �3400) grains/gram. This
450
455
460
465
470
Figure 5. Continued
B=w in print; colour online
10 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:34
Plate 1. The sample/slide number, England Finder coordinates, and position of taxa on the quantitative range chart (numbers inparentheses) are indicated for all specimens; scale bar represents 20 mm. The microphotographs were all taken using plain trans-mitted light.
B=w in print; colour online
Palynology 11

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
Plate 2.
B=w in print; colour online
12 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
increase is accompanied by a decrease in diversity of
spore species, which are mainly represented by Deltoi-
dospora and Cicatricosisporites. The abundance of phy-
toplankton continues to decrease (maximum 244,average 100 cysts/gram) throughout the interval, but
with a similar low diversity (�14 species) to the under-
lying interval.
Discussion concerning age assessment: Sofrepites
legouxae is an index taxon in this interval (Figure 5;
Plate 1), which was found to range from the late Albian
to early Cenomanian in foraminifera-dated rocks of
Senegal by Jardin�e and Magloire (1965), Jardin�e(1967), and in Brazil by Herngreen (1973) and
Herngreen and Jimenez (1990). Elaterosporites
verrucatus is recorded in Senegal and the Ivory Coast
from foraminifera-dated mid Albian–early Cenoma-
nian sediments (Jardin�e & Magloire 1965; Jardin�e1967: Figure 2). However, in foraminifera- and ammo-nite-dated rocks from Brazil, this same taxon was
recorded from the latest mid to earliest late Albian
(Herngreen 1973; Regali et al. 1974; Herngreen &
Jimenez 1990), and a single occurrence has been
reported from the middle part of the ammonite-dated
late Albian of Columbia (Herngreen & Jimenez 1990).
Cretacaeiporites densimurus appears in the lower part
of this sample interval, and was first described bySchrank and Ibrahim (1995) from foraminifera-dated
rocks of early–mid Cenomanian age, and was later
475
480
485
490
495
500
Figure 6. The Abu Tunis 1x borehole with lithological column, sample positions, original age dating, key biostratigraphic eventsand ages deduced in the current work.
B=w in print; colour online
Palynology 13

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
recorded by Ibrahim (2002) from the late Albian–early
Cenomanian of Egypt. Single specimens of other spe-
cies of this genus have been found in the upper part of
this interval: Cretacaeiporites polygonalis, which has alate Albian–late Cenomanian range in Senegal (Jardin�e& Magloire 1965), and Cretacaeiporites mulleri, which
ranges from the late Albian up to the Santonian in Sen-
egal and northeast Nigeria (Jardin�e & Magloire 1965;
Lawal & Moullade 1986). In Brazil, Cretacaeiporites
mulleri was recorded as of late Albian–mid Cenoma-
nian age (Herngreen 1973).
Appearing throughout the upper part of this inter-val, the range of Elaterocolpites castelainii was used to
document the base of the late Albian and top of the mid
Cenomanian in the Elaterates Province in Senegal
(Jardin�e & Magloire 1965; Jardin�e 1967: FigureQ5 ) and
Brazil (Herngreen 1973; Herngreen & Jimenez 1990).
This taxon was also recorded by Hochuli (1981)
from the foraminifera-dated late Albian of southern
Switzerland. Other late Albian–early Cenomanianelaterate taxa present in this zone are Elaterosporites
acuminatus and Elaterosporites protensus (Figure 6).
Elaterosporites acuminatus was reported from the late
Albian–early Cenomanian of Senegal (Jardin�e 1967),
and occurs in the lower part of this interval. Elaterospor-
ites protensus is found in the same samples and was
recorded from foraminifera-dated rocks of mid Albian
to latest Albian/earliest Cenomanian age in Senegal, theIvory Coast (Jardin�e & Magloire 1965; Jardin�e 1967)
and Brazil (Herngreen 1973; Herngreen & Jimenez,
1990).
Afropollis kahramanensis was recorded by Schrank
and Ibrahim (1995) and Ibrahim (2002) from the fora-
minifera-dated early–mid Cenomanian of Egypt. The
former authors mentioned that the taxon described as
Pollen PO-304 by Lawal and Moullade (1986) from thesupposed late Albian–mid Cenomanian of northeast
Nigeria was identical to their new species; therefore, the
presence of Afropollis kahramanensis in this
interval does not contradict the proposed late Albian–
Cenomanian age. Foveotricolpites gigantoreticulatus
was recorded from foraminifera-dated rocks of
Turonian–Santonian age in Senegal and northeast
Nigeria (Jardin�e & Magloire 1965; Lawal & Moullade1986). However, Schrank and Ibrahim (1995) and
Schrank and Mahmoud (1998) documented an older
occurrence of this species in the palynologically dated
Albian–Cenomanian of Egypt. Thus, the presence of
Foveotricolpites gigantoreticulatus in the middle part of
this interval may not conflict with the proposed
late Albian–early Cenomanian age of the interval.
The rare presence of Triporites spp. in this intervalis in accordance with the late Albian–early Cenomanian,
as rare Triporites spp. were recorded from the early
Cenomanian of Senegal (Jardin�e &Magloire 1965).
The questionable occurrence of Droseridites bacu-
lites in this interval, which was described by Ibrahim
(1996) from the palynologically dated lower Turonian
of Egypt, may be attributed to caving.As for the dinocysts, Florentinia berran appears in
the lower part of the interval, a taxon that was recorded
from the Albian–early Cenomanian of the southern
Tethyan Realm in Morocco and northeast Libya
(Below 1982, 1984; Uwins & Batten 1988). Florentinia
laciniata and Florentinia mantellii first appear in this
interval and are known from ammonite-dated Aptian–
early Cenomanian sequences in the Tethyan Realm:they continue upward into the overlying intervals.
Based on the presence of the rare late Albian–early
Cenomanian elaterates and the marker angiosperm
forms mentioned above, a late Albian–early Cenoma-
nian age is postulated for the interval.
Samples from this part of Abu Tunis 1x were recog-
nized by WEPCO (1968) as ‘Cenomanian Clastics’.
However, the dominant sandstone composition of sam-ples 29–39 when combined with the late Albian–early
Cenomanian age proposed here suggests allocation of
these sediments to the upper Kharita Formation.
Correlation: The palynomorph assemblage recorded in
this zone is identical to that described in Zone II (late
Albian–early Cenomanian) of Herngreen (1973) from
the Maranh~ao Basin, Brazil. Similar late Albian–early
Cenomanian assemblages that share some of the elater-ate and angiosperm pollen grains (but not Sofrepites
legouxae) are those of sequences IX of Jardin�e and
Magloire (1965) in the Senegal Basin, and Subzone Ia
of Lawal and Moullade (1986) in the upper Benue
Basin, northeast Nigeria. The same applies to the
assemblage of intervals 3–4 in the Manndra 1 well and
intervals c–d in the Algerian Oued Melah 1 well of
Foucher et al. (1994). Similar late Albian–earlyCenomanian zones (but also lacking Sofrepites
legouxae) have also been documented from the Egyp-
tian northern Western Desert, for example, Zone III of
Aboul Ela and Mahrous (1992) in the East Tiba-1 well,
Zone IV of Schrank and Ibrahim (1995), in the
Kahraman-1 well, and assemblage ‘A’ of Ibrahim
(2002), in the Abu Gharadig-5 well. The upper part of
Zone II (mid/late Albian–early Cenomanian) of El-Beialy et al. (2011) in the Gebel Rissu-1 well is very
similar to the current palynozone, and this time
S. legouxae was recorded.
PZ3: Sofrepites legouxae Partial Range Zone (early–
?mid Cenomanian)
Samples: This zone spans samples 40 to 46 and a depthfrom 4950 to 4650 ft (1509–1417 m).
Definition: From the HO of Sofrepites legouxae (26) to
the LO of Proteacidites cf. africaensis (35).
505
510
515
520
525
530
535
540
545
550
555
560
565
570
575
580
585
590
595
600
605
14 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
Associated taxa: Deltoidospora spp. (1), Crybelosporites
pannuceus (2), Cicatricosisporites orbiculatus (4), Bal-
meiopsis limbatus (18), Classopollis classoides (19),
Ephedripites spp. (20), Elaterosporites klaszii (21),Araucariacites australis (24), Elaterocolpites castelainii
(25), Afropollis jardinus (36), Rousea delicipollis (44),
Cretacaeiporites mullerii (45), Triporites spp. (46), Rou-
sea cf. miculipollis (47), Florentinia mantellii (99), Flor-
entinia laciniata (103).
Remarks: Sporomorphs are represented by pterido-
phyte spores, pollen from xerophytic gymnosperms,
two elaterate, and five angiosperm pollen species. Thephytoplankton assemblage shows a slight increase in
abundance over the interval below (maximum 370,
average 120 cysts/gram) but with a consistently low
diversity (�10 species), and provides an important
characteristic for dating the interval.
Discussion concerning age assessment: Proteacidites cf.
africaensis, which is widely accepted to document the
base of the mid Cenomanian in the western part ofthe Elaterates Province (e.g., Jardin�e & Magloire
1965; Lawal & Moullade 1986) first appears in
the overlying interval (in Sample 47), and thus
delineates the lower boundary of the mid
Cenomanian. Consequently, the present interval may
be of early Cenomanian age.
Florentinia berran, which was found in sediments as
young as the early–mid Cenomanian in Egypt(Schrank & Ibrahim 1995; Ibrahim 2002), becomes
extinct in the lower part of the overlying interval, thus
favoring an early–mid Cenomanian age for this sample
interval. Interestingly, an exact match of the biostrati-
graphic events recorded herein: the highest occurrence
of Elaterocolpites castelainii, the occurrence of Elatero-
sporites klaszii, Afropollis jardinus, and rare triporate
pollen grains, with those recorded in the Zone V(early–?mid Cenomanian) of Schrank and Ibrahim
(1995) also supports an early–?mid Cenomanian age
for the current zone.
WEPCO (1968) referred to the interval from which
samples 40 to 46 come as ‘Cenomanian Clastics’. The
mixed, clastic-carbonate lithology of these samples
and the presumed early–?mid Cenomanian age indi-
cates this part of the sequence can now be identifiedas the lower Bahariya Formation (Figure 6), as the
lower part of this formation in the north Western
Desert is composed of sandstones with alternating
with shales and frequent limestone horizons. This
formation conformably overlies the Kharita
Formation and underlies the Abu Roash Formation
(Norton 1967).
Correlation: Regional comparisons of Egyptian (early–mid Cenomanian) palynozones reveal that Zone V of
Schrank and Ibrahim (1995) in the Kahraman-1 and
Abu Gharadiq-18 wells, Zone 3 of Ibrahim (1996) in
the Ghazalat-1 well and assemblage ‘B’ of Ibrahim
(2002) in the Abu Gharadiq-5 well can be correlated
with the present zone.
PZ4: Proteacidites cf. africaensis Total Range Zone(mid–late Cenomanian)
Samples: This zone includes samples 47 to 62 and cov-
ers a depth from 4600 to 3850 ft (1402–1173 m).
Definition: Total range of Proteacidites cf. africaensis
(35).
Associated taxa: Deltoidospora spp. (1), Crybelosporitespannuceus (2), Alisporites cf. grandis (17), Balmeiopsis
limbatus (18), Ephedripites spp. (20), Elaterosporites
klaszii (21), Retimonocolpites variplicatus (41), Tricol-
poropollenites sp. (43), Senegalinium aenigmaticum
(85), Trichodinium castanea (87), Surculosphaeridium
cf. longifurcatum (88), Xiphophoridium alatum (95),
Florentinia spp. (94), Florentinia mantellii (99).
Remarks: The palynofloral assemblage of this intervalshows characteristics similar to the underlying interval,
but with a continuing decrease in the abundance and
diversity of spores (maximum 48/average 24 grains/
gram, and only two species), the complete disappear-
ance of gymnosperm pollen grains at the end of the
interval, and the occurrence of nine angiosperm pollen
taxa. Phytoplankton increase in abundance (maximum
3160, average 140 cysts/gram), but have a very low spe-cies diversity (�8 species).
Discussion concerning age assessment: The inception of
Proteacidites cf. africaensis at the base of this interval
is taken here to mark the lower boundary of the mid
Cenomanian interval, as it was recorded in West
Africa, in Senegal and Nigeria (Jardin�e & Magloire
1965; Jardin�e 1967; Lawal & Moullade 1986) and
Brazil (Herngreen 1973), from foraminifera-datedrocks of mid–late Cenomanian age.
The occurrence of Afropollis jardinus in these sam-
ples requires explanation. The extinction of this species
has been shown to be diachronous across palaeotropi-
cal African regions, occurring either in the early or mid
Cenomanian, a character attributed by Doyle et al.
(1982) to palaeoclimatic influences. In the foraminif-
era-dated Gabon reference section for Afropollis spe-cies, Doyle et al. (1982) found that the abundance of
Afropollis jardinus declined in the early Cenomanian
Subzones C-XIIb and C-XIIc, and disappeared before
the appearance of Proteacidites africaensis (as Triorites
africaensis) in mid-late Cenomanian age sediments
(Jardin�e & Magloire 1965; Jardin�e 1967). However, in
Senegal Doyle et al. (1982) noted that Afropollis jardi-
nus diminishes in the early mid Cenomanian – laterthan in the Gabon section – and disappeared by the
end of the mid Cenomanian, contemporary with the
appearance of Proteacidites africaensis. Doyle et al.
610
615
620
625
630
635
640
645
650
655
660
665
670
675
680
685
690
695
700
705
710
715
Palynology 15

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
(1982) interpreted this persistence of Afropollis
jardinus through the mid Cenomanian as due to more
favourable (wetter) conditions. An interpretation that
was later supported by Schrank (2001)Q6 as he suggestedAfropollis thrived in humid coastal areas. The same
scenario could also apply to Egypt, which was at a
palaeolatitude similar to those of Senegal and
where Afropollis jardinus may have persisted into the
mid Cenomanian due to local humid coastal
conditions.
The HO of Afropollis jardinus is found at the top of
both the foraminifera-dated Zone V of Schrank andIbrahim (1995) and Assemblage Zone ‘B’ of Ibrahim
(2002), both of which are known to be of early–mid
Cenomanian age. Therefore, the HO of Afropollis jar-
dinus in sample 53 in Abu Tunis 1x is used to delineate
the upper boundary of the mid Cenomanian. Afropollis
kahramanensis, which was recorded in the foraminif-
era-dated lower–middle Cenomanian rocks of Egypt
(Schrank & Ibrahim 1995; Ibrahim 2002), has its HOat the same level as the HO of Afropollis jardinus. Ela-
terosporites klaszii has its highest occurrence in the
lower part of this interval, and is known to terminate
in the mid Cenomanian in the Elaterates Province
(Jardin�e & Magloire 1965; M€uller 1966; Jardin�e 1967;
Herngreen 1973; Herngreen & Jimenez 1990: Figure 2).
The presence of Cretacaeiporites densimurus is also
consistent, as it has its uppermost occurrence at thetop of the mid Cenomanian in Egypt (Schrank &
Ibrahim 1995; Ibrahim 2002). The late Cenomanian
age of the upper part of this zone is inferred from the
upward continuation of Proteacidites cf. africaensis
and from the very rare occurrence (and later complete
disappearance) of the gymnosperm pollen Classopollis
spp.. In Senegal, Jardin�e and Magloire (1965) recorded
high abundances of Classopollis spp. (up to 80%) fromthe Barremian to the mid Cenomanian, which then
declined rapidly and became extinct by the end of the
late Cenomanian. Similarly, Schrank and Ibrahim
(1995) and Ibrahim (2002) recorded rare final occur-
rences of Classopollis in the middle Cenomanian rocks
of Egypt.
A mid–late Cenomanian age is therefore assigned
to this interval based on the presence of the indexgymnosperm and angiosperm taxa mentioned above.
The mid to late Cenomanian age combined with the
dominantly carbonate lithology of the samples in this
palynozone suggests the unit should be identified as
the upper Bahariya to lower Abu Roash formations.
WEPCO (1968) did not recognise such subdivisions
in their original study of Abu Tunis 1x, but the
new assignments are consistent with the known litho-logical and age characteristics of the Abu Roash
Formation, a limestone sequence with interbeds of
shale and sandstone known to range in age from late
Cenomanian to Turonian (Hantar 1990). The Abu
Roash is conformably underlain by the Bahariya
Formation and overlain by the Khoman ‘B’
Formation.Correlation: The characteristics of this palynozone
strongly resemble the mid–late Cenomanian Sequence
VII of Jardin�e and Magloire (1965) in the Senegal
Basin and the ‘Triorites’ africaensis Zone (II) of Lawal
and Moullade (1986), upper Benue Basin, northeast
Nigeria. Zone III (late Cenomanian) of Herngreen
(1973), Brazil, only equates to the upper part of the
present zone. Other zones similar to the current onehave been identified in the northern Western Desert:
Zone VI of Schrank and Ibrahim (1995) in the Kahra-
man-1 and Abu Gharadiq-18 wells, and Zone 4 of
Ibrahim (1996) in the Ghazalat-1 well, both zones hav-
ing been allocated a mid?–late Cenomanian age. Zone
I (late Cenomanian) of El Beialy et al. (2011) in the
Gebel Rissu-1 well could also correspond to the cur-
rent zone, as the latter authors have equated their zoneto the aforementioned Zone VI of Schrank and
Ibrahim (1995). This could be further supported by the
mid Cenomanian Elaterosporites klaszii and Cretacaei-
porites densimurus which occur in the current zone as
they do in the lower part (samples 3–5) of Zone I of El
Beialy et al. (2011).
PZ5: Canningia senonica Total Range Zone (early?
Santonian)
Samples: This zone includes samples 73 to 74 and cov-
ers a depth from 3250 to 3200 ft (991–975 m). The two
overlying samples, 75 and 76, are barren. Sample 77 is
separated from 75 and 76 by an unconformity surface
(identified in the well log by WEPCO 1968) and exhib-
its very poor dinoflagellate recovery and was deemedbarren (Figure 5).
Definition: Total range of Canningia senonica (79).
Associated taxa: Alisporites cf. grandis (17), Spiniferites
spp. (77), Exochosphaeridium bifidum (80), Dinogym-
nium spp. (82), Senegalinium aenigmaticum (85), Tri-
chodinium castanea (87), Surculosphaeridium cf.
longifurcatum (88), Downiesphaeridium sp. (92).
Remarks: Palynological assemblages from this zone aredominated by phytoplankton, with only one terrestrial
palynomorph being present: the gymnosperm Alispor-
ites cf. grandis.
Discussion concerning age assessment: Canningia senon-
ica (Plate 3) appears in both samples 73 and 74
(Figure 5), a species characteristic of the Santonian–
Maastrichtian. Canningia senonica has been recorded
from the ammonite-dated late Santonian of the Isle ofWight, southern England (Clarke & Verdier 1967),
from the foraminifera-dated early Santonian–late
Maastrichtian of Egypt (Schrank & Ibrahim 1995),
720
725
730
735
740
745
750
755
760
765
770
775
780
785
790
795
800
805
810
815
820
825
16 A.S. Deaf et al.
TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
Plate 3.
B=w in print; colour online
Palynology 17

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
and from the nannoplankton-dated late Campanian of
Italy (Torricelli & Amore 2003).
This interval contains a few long-ranging species
of pre-Coniacian to Maastrichtian age, such asChlamydophorella discreta which appears first in this
interval, a species which has its highest occurrence in
the late Santonian of the Isle of Wight (Clarke &
Verdier 1967), and ranges in Egypt from the early
Cenomanian to the Turonian (Schrank & Ibrahim
1995). Isabelidinium acuminatum was recorded (as
Deflandrea acuminata) from the ammonite-dated late
Santonian of the Isle of Wight (Clarke & Verdier1967), and from the foraminifera-dated Coniacian–
Santonian of Algeria (Foucher et al. 1994). Dinogym-
nium denticulatum, recorded from the ammonite-dated
Late Santonian of the Isle of Wight (Clarke & Verdier
1967), from the foraminifera-dated sequences of
Coniacian–Maastrichtian age in Egypt (Schrank &
Ibrahim 1995), and of Coniacian–Santonian age in
Algeria (Foucher et al. 1994). Eucladinium gamban-
gense, which was identified by Cookson and Eisenack
(1970) from the Senonian rocks of Australia, occurs
at the base of this interval.
There are no index angiosperm pollen taxa that one
might expect for the Turonian, such as Foveotricolpites
giganteus or Foveotricolpites gigantoreticulatus, which
are characteristic of the early Turonian–Santonian in
West Africa (e.g., Jardin�e & Magloire 1965; Lawal &Moullade 1986; Schrank & Ibrahim 1995). Nor is Dro-
seridites senonicus present, diagnostic of the late
Turonian–early Santonian (Lawal & Moullade 1986;
Salard-Cheboldaeff 1990; Schrank & Ibrahim 1995;
Ibrahim 1996).
Thus, based on the presence of the early Santonian
index form Canningia senonica and the complete
absence of index Turonian sporomorphs mentionedabove, an early Santonian age is proposed for this
interval. The WEPCO report (1968) did not allocate an
age to this part of the borehole succession.
The proposed ages and carbonate lithology of sam-
ples 73 and 74 suggest this part of the sequence can be
assigned to the lower Khoman ‘B’ Formation, which is
easily recognized in north Egypt by its white chalk/
chalky limestone lithology. This unit unconformablyoverlies the Abu Roash Formation in the Abu Tunis
1x borehole (as is the case in many basins of the north
Western Desert), and conformably underlies the
Khoman ‘A’ Formation. The Khoman ‘B’ Formation
has been allocated a Coniacian–Santonian age (Hantar
1990; Kerdany & Cherif 1990).
Correlation: The dinoflagellate Zone 8 (early
Santonian) of Schrank and Ibrahim (1995), in theKahraman-1 well in the northern Western Desert
of Egypt, is the only zone that correlates to the
palynozone 5.
5. Discussion of the Abu Tunis 1x palynostratigraphy
Semi-quantitative frequencies are closed scales in
which measured variables depend on each other, and
thus reflect a false picture of real quantitative distribu-tions of variables measured. Thus, this section deals
with the quantitative (grains/g of sediments) distribu-
tions but combined with semi-quantitative to explore
effect of data closure on semi-quantitative distribu-
tions, and to enable a wider comparison of quantitative
data with its counterparts (i.e., semi-quantitative) in
regional and interregional contexts.
5.1. Albian–Cenomanian Elaterates PhytogeographicProvince
5.1.1. Quantitative characteristics of the Albian–
Cenomanian palynoflora from the present study
The palynoflora of the Abu Tunis 1x exhibits a remark-
able similarity to that of the Albian–Cenomanian Ela-
terates Phytogeographic Province of Herngreen et al.
(1996). Features diagnostic for the province are repre-
sented here by the appearance and later increased
abundances of Afropollis jardinus (0.4–54%, avg. 15%;4–7003, avg. 1219 grains/g), and occurrence of Crybelo-
sporites (0.4–3.6%, avg. 2%; 4–523, avg. 118 grains/g).
Other typical criteria for the recognition of this prov-
ince found in the Abu Tunis 1x borehole are the
appearance and diversification of gymnospermous ela-
terate pollen grains: Elaterosporites (0.4–7.6%, avg.
39%; 4–854, avg. 262 grains/g), Elaterocolpites (0.4%;
40–52 and avg. 46 grains/g), Galeacornea (0.8%;26 grains/g), and Sofrepites(0.4–2.4%, avg. 1%; 21–465,
avg. 146 grains/g), and the presence of Cretacaeiporites
(0.4–1.6%, avg. 0.9%; 19–174, avg. 75 grains/g). The
diversification of Tricolpites (0.4–3.2%, avg. 1.6%; 25–
319, avg. 90 grains/g), Tricolporites (0.4%; 12–58, avg.
35 grains/g), and Triporites (0.4–1.2%, avg. 0.6%; 12–
119, avg. 43 grains/g) accompanied with a drop in
abundance of smooth trilete spores from (17–87%, avg.53%; 1332–40762, avg. 7663 grains/g) in the late Albian
to (0.8–38%, avg. 11; 9–2195, avg. 639 grains/g) in the
Cenomanian are important features. The complete
absence of bi- and tri-saccate gymnospermous pollen
grains, and the abundance (0.4–5.6%, avg. 0.2%; 5–
1394, avg. 270 grains/g) and later disappearance of
Classopollis in the mid–late Cenomanian are also diag-
nostic features.
5.1.2. Regional mid Albian–early Cenomanian acme of
Afropollis jardinus
The genus Afropollis jardinus, as mentioned in Section4.2.4, shows different biostratigraphic and semi-quan-
titative distributions between northeast Africa and
West Africa that have been interpreted to be the result
830
835
840
845
850
855
860
865
870
875
880
885
890
895
900
905
910
915
920
925
930
18 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:35
of palaeoecological factors. Palynological studies of
this taxon in Egypt have so far been semi-quantitative,
and no attempt has been made to explore the possible
quantitative significance of the species on a regionalscale. Quantitatively, the temporal abundance of Afro-
pollis jardinus (Figure 7) shows very low concentra-
tions (� 33–738, avg. 350 grains/g; 0.4–17%, avg. 4%)
in the lower Albian of the Abu Tunis 1x well until the
middle Albian–lower Cenomanian marine sediments,
where a peak abundance is recorded of 1400–7003 and
average 2900 grains/g (24–62%, avg. 34%). In an adja-
cent basin, about 51 km to the southwest of AbuTunis, Schrank and Ibrahim (1995) recorded a simi-
larly high relative abundance acme (31–61%, avg. 40%)
in the mid Albian–early Cenomanian of the
Kahraman-1 well. The slightly higher percentages of
the species in the mid Albian of the Kahraman-1 well
may potentially be attributed to more favourablelithologies, as there is a greater shale content in this
part of the well when compared to Abu Tunis 1x. Simi-
larly, about 190 km to the southeast of Abu Tunis 1x,
the same mid Albian–early Cenomanian acme (11–
64%, avg. 34%) of Afropollis jardinus was recorded
from the Bed 1–1 well (El-Beialy 1994). Consistency
between quantitative and semi-quantitative values
through the Afropollis jardinus acme as recorded frommarine sediments of three wells in three different basins
supports the contention made by Schrank (2001) that
this acme event may have been controlled by locally
humid conditions associated with proximity to shallow
marine depositional environments, as the Afropollis-
parent plant is thought to have thrived in humid
coastal plans. Interregionally, similar mid Albian–early
Cenomanian (10–40%, avg. 14%) peaks of Afropollis
jardinus have been documented in Senegal (Doyle et al.
1982). In Gabon, the appearance of the Afropollis jar-
dinus acme in the early–mid Albian differs from those
recorded in Egypt and Senegal, and may be due to dif-
ferences in palaeogeographic position and geological
history of the basins (Doyle et al. 1982). In this light,
acmes of Afropollis are only of biostratigraphic rele-
vance on a regional – i.e., not interregional – scale.We believe that the acme event of Afropollis jardi-
nus in the north Western Desert of Egypt is an impor-
tant regional mid Albian–early Cenomanian bioevent.
5.2. Senonian palynoflora from the present study
The early Santonian palynoflora of the Abu Tunis 1x
borehole is exclusively represented by marine phyto-planktons and completely lacks any terrestrial palyno-
morphs that would be representative for the Senonian
Palmae Province of Herngreen et al. (1996).
5.3. Late Cretaceous uplift of the Faghur area
An unconformity surface is identified by WEPCO
(1968) in the composite log of the Abu Tunis 1x bore-hole separating the palynomorph-barren interval
assigned by WEPCO (1968) as of Turonian age from
the overlying dinoflagellate-identified lower Khoman
‘B’ (early? Santonian) Formation (Figure 6). This may
indicate a short time cessation in deposition and ero-
sion, which may be related to the Turonian uplift that
affected most of the Western Desert basins (e.g.,
Kerdany & Cherif 1990). By the early Santonian, theFaghur area had become marine, based on the marine
palynomorphs recorded in the Abu Tunis 1x borehole
sediments.
935
940
945
950
955
960
965
970
975
980
985
990
995
Figure 7. Quantitative and semi-quantitative vertical distri-butions of Afropollis jardinus in the Abu Tunis 1x boreholecompared to semi-quantitative distributions in Egyptianwells and Senegal and Gabon Albian–Cenomanian sections.
Palynology 19

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
5.4. A Unified mid-Late Cretaceous palynologicalzonal scheme for the northern Western Desert
The zonal scheme devised for the Abu Tunis 1x bore-
hole suggests the use of unified biozones of those of
Schrank and Ibrahim (1995) and our zones, which are
at least applicable to the Egyptian Albian–middle Cen-
omanian rocks of the northern Western Desert. This is
because the Albian–Cenomanian diagnostic taxa have
been widely recorded in Egypt and have calibrated
stratigraphic ranges across the Albian–CenomanianElaterates Phytogeographic Province (PZ1-4). There
are some difficulties when dealing with the upper Ceno-
manian rocks. This may be due to change in lithology
and depositional environment from clastic shallow
marine (as in the Kahraman-1 well, Schrank & Ibrahim
1995) to carbonate inner neritic environment towards
the north (as in the Abu Tunis 1x well). More difficul-
ties arise when dealing with Turonian and youngerrocks. Palynomorph recovery from post Cenomanian–
Maastrichtian carbonates is rather low due to deeper
neritic marine environment, regional late Cretaceous
tectonism and a global late Cretaceous decline in gen-
era and species diversity of palynomorphs. In order to
draw a complete and unified mid-late Cretaceous zonal
scheme for the Egyptian northern Western Desert, it is
suggested that further palynological (preferably agecontrolled) work on successions of adjacent and less
tectonically affected basins that were slightly close to
palaeovegetation cover is needed.
6. Systematic palynology
A. Elaterate pollen
Genus: Elaterosporites Jardin�e 1967Type species: Elaterosporites verrucatus (Jardin�e andMagloire) Jardin�e 1967, p. 244, pl. 2, figs. E-G, pl. 3,fig. G.Elaterosporites verrucatus (Jardin�e and Magloire)Jardin�e 1967Plate 1, figs. 6, 8
Description: Grains with ellipsoidal plano-convex to
sub-hemispherical body with three U-shaped horns
implanted on the convex distal face. Proximal face flator depressed, bounded by 3–8 mm wide annular ring
structure parallel to the equator ending with protrud-
ing ends parallel to the long axis. These three U-shaped
appendages with their middle part running parallel to
the longest axis; one is placed over the distal pole, the
other two are placed laterally. The appendages are
solid and of uniform diameter. Exine is granulate to
verrucate. Main body length 38–64 mm and breadth25–45 mm.
Dimension: Maximum main body length (2 specimens)
40 (52.5) 65 mm, breadth undetermined due to grain
orientation, annular ring 3–8 mm wide; appendage
length 33 (36.5) 40 mm, width 3.5 (4.5) 5.5 mm.
Remark: Elaterosporites verrucatus can be distin-
guished from Elaterosporites acuminatus (Plate 1, fig-ures 7, 14) by its 3–8 (here also 3–8) mm wide annular
ring structure and granulate to verrucate exine (here
verrucae 1–3.5 mm wide) versus 8–12 (here 6–8) mm
wide annular ring and densely and uniformly packed
exine with spines 4–6 (here 4–5) mm high and 2.5–3
(here 2.5–3.5) mm wide.
Elaterosporites protensus (Stover) Jardin�e 1967Plate 1, fig. 9
Remark: Stover (1963) distinguished Elaterosporites
protensus from Elaterosporites acuminatus by its larger
size (Elaterosporites acuminatus: 52 � 28 mm), greater
ring width, and appendages ending with round tips
rather than with sharply tipped ends as in the latterspecies. Here dimensions of Elaterosporites protensus
are: maximum main body length (1 specimen) 65 mm,
breadth 35 mm, annular ring 7–9.5 mm wide; append-
age length and width undetermined due to grain orien-
tation. Dimensions of Elaterosporites acuminatus are:
maximum main body length (2 specimens) 39 (44)
49 mm, breadth 27 (28) 29 mm, annular ring 6–8 mm
wide; appendage length 32 (36) 40 mm.
B. Pollen tetrad
Genus: Droseridites Cookson 1947 ex Potoni�e 1960
Type species: Droseridites spinosus (Cookson) Potoni�e1960, p. 137–139.
Droseridites senonicus Jardin�e and Magloire, 1965Plate 2, fig. 16
Remark: The present specimens show greater tetrad
diameters than that of the original specimens (12–
19 mm) described by Jardin�e and Magloire (1965).
Maximum tetrad diameter (3 specimens) 19 (21)22 mm.
7. Conclusions
(1) This work provides the first quantitative distri-
butions of Egyptian post-Aptian palynofloral
assemblages, which, combined with semi-quan-
titative data, enables a wider comparison ofassemblages on regional and interregional
scales. The Albian–Cenomanian palynoflora
recovered from the Abu Tunis 1x
1000
1005
1010
1015
1020
1025
1030
1035
1040
1045
1050
1055
1060
1065
1070
1075
1080
1085
1090
20 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
borehole shows characteristics of the Albian–
Cenomanian Elaterates Phytogeographic Prov-
ince, whereas the early Santonian palynoflora
is exclusively represented by marine phyto-plankton and completely lacks any terrestrial
palynomorphs representative of the Senonian
Palmae Province.
(2) The qualitative and quantitative distributions
of index palynomorph taxa of the Abu Tunis
1x borehole have enabled the recognition of
four Cretaceous lithostratigraphic units, which
permit new age assignments to sections of theborehole undated by the drilling company.
These four units have been subdivided into five
palynological zones. A barren interzone is
recorded just below the uppermost biozone. An
absence of Turonian–Coniacian palynomorphs
may be related to the unfavourable limestone
and dolostone lithologies. An unconformity
surface located between the barren interval andthe overlying lower Santonian may indicate a
short time cessation in deposition and erosion,
which may be related to the Turonian uplift
and basin inversion of the Faghur area, which
has affected most of the northern basins of
Egypt.
(3) Correlation between the biozones defined for
the Abu Tunis 1x borehole with those ofSchrank and Ibrahim (1995) shows that the
new scheme is applicable at least to Albian–
middle Cenomanian succession in the Western
Desert. However, there are some difficulties
correlating the upper Cenomanian parts of the
successions, which are due to the change in
depositional environments from clastic shallow
marine to carbonate-rich open marine environ-ments in the more northern basins. Further pal-
ynological (preferably independently age
controlled) work on successions in adjacent
and less tectonically affected basins may permit
the development of a complete late Cenoma-
nian–Maastrichtian palynozonal scheme for
the Egyptian northern Western Desert.
(4) A major synchronicity between the North andWest African late Cretaceous biostratigraphic
units is suggested here, in contrast to the argu-
ment made by Schrank and Ibrahim (1995).
The synchronicity is based on occurrence of
foraminifera-calibrated biostratigraphic events
in both the North and West African sequences.
The cause of the synchronicity is strongly sug-
gested to be the continuous mid–late Cenoma-nian anticlockwise north-northeast African
plate movement towards Laurasia as a
response to the breakup of Western
Gondwana. This resulted in North and West
African countries being brought to a similar
palaeosubtropical position and thus under sim-
ilar palaeoclimate, whereby these regionsshould have similar palynofloral compositions
of similar age ranges.
(5) The quantitative vertical distribution of Afro-
pollis jardinus recorded from the Abu Tunis 1x
borehole sediments shows a mid Albian–early
Cenomanian acme event that can be correlated
with similar semi-quantitatively identified
acmes of the species in other wells in the northWestern Desert of Egypt, permitting the recog-
nition of this acme event as an important
regional stratigraphic marker.
Acknowledgements
A.S. Deaf gratefully thanks the Egyptian Government forgenerously funding him a PhD scholarship at University ofSouthampton, UK. The authors wish to thank the EgyptianGeneral Petroleum Corporation for providing well logs andsamples of the Abu Tunis 1x borehole. Thanks are also dueto Mr. Shir Akbari at NOCS, Q7University of Southampton,for his help in the laboratory.
Author biography Q8
References
Abdel-Kireem MR, Schrank E, Samir AM, Ibrahim, MIA.1996. Cretaceous palaeoecology, palaeogeography andpalaeoclimatology of the northern Western Desert,Egypt. J Afr Earth Sci. 22(1):93–112.
Abdelmalik WM, Aboul Ela NA, El-Shamma AG. 1981.Upper Jurassic-lower Cretaceous microflora from thenorth Western Desert, Egypt. Neues Jahrbuch f€ur Geolo-gie und Pal€aontologie – Abhandlungen. 162:244–263.
Aboul Ela NM, Mahrous HAR. 1992. Albian-Cenomanianmiospores from the subsurface of the north Western Des-ert, Egypt. Neues Jahrbuch f€ur Geologie undPal€aontologie Monatschefte. 10(1):595–613.
Below R. 1982. Dinoflagellate cysts from Valanginian tolower Hauterivian sections near Ait Hamouch, Morocco.Revista Espa~nola de Micropaleontologia. 14:23–52.
Below R. 1984. Aptian to Cenomanian dinoflagellate cystsfrom the Mazagan Plateau, northwest Africa (Site 545and Site 547, Deep Sea Drilling Project Leg 79). InitialReports of the Deep Sea Drilling Project. 79(Nov):621–649.
Bettar I, M�eon H. 2001. Palynological study of the middle/upper Albian transition in the Tarfaya Basin (southwestof Morocco) and some new data about the African-SouthAmerican Province. Revue de Micropal�eontologie.44:107–123.
Bettar I, M�eon H. 2006. La palynoflore continentale del’Albien du Bassin d’Agadir-Essaouira (Maroc). Revuede Pal�eobiologie. 25:593–631.
Boltenhagen E. 1980. Palynologie du Cr�etac�e sup�erieur duGabon. M�emoires de la Section des Sciences. 7:11–191.
1095
1100
1105
1110
1115
1120
1125
1130
1135
1140
1145
1150
1155
1160
1165
1170
1175
1180
1185
1190
1195
1200
Palynology 21

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
Brenner GJ. 1968. Middle Cretaceous spores and from north-eastern Peru. Pollen et Spores. 10:341–382.
Brideaux WW. 1971. Palynology of the lower ColoradoGroup, central Alberta, Canada. 1. Introductoryremarks, geology and microplankton studies. Palaeon-tographica Abteilung B. 135:53–114.Q9
Clarke RFA, Verdier J-P. 1967. An investigation of micro-plankton assemblages from the Chalk of the Isle ofWight, England. Verhandelingen der Koninklijke Neder-landse Akademie van Wetenschappen, Afdeeling Natuur-kunde, Eerste Reeks. 24(3):1–96.
Cookson IC, Eisenack A. 1970. Cretaceous microplanktonfrom the Eucla Basin, Western Australia. Proceedings ofthe Royal Society of Victoria. 83(2):137–157.
Davey RJ, Verdier JP. 1973. An investigation of microplank-ton assemblages from latest Albian (Vraconian) sedi-ments. Revista Espa~nola de Micropaleontologia. 5:173–212.
Deaf AS. 2009. Palynology, palynofacies and hydrocarbonpotential of the Cretaceous rocks of northern Egypt.Unpublished Ph.D. thesis, Southampton, U.K.: Univer-sity of Southampton p. 348.
Doukaga, M. 1980. Etude palynologique dans le Cr�etac�emoyen du bassin sedimentaire du Gabon. Lille, France:Universit�e des Sciences et Technologies de Lille; p. 174.
Doyle JA, Jardin�e S, Doerenkamp A. 1982. Afropollis, a newgenus of early angiosperm pollen, with notes on the Cre-taceous palynostratigraphy and paleoenvironments ofnorthern Gondwana. Bulletin des Centres de RecherchesExploration-Production Elf Aquitaine. 6:39–117.
El-Beialy SY. 1994. Palynostratigraphy and palynofaciesanalysis of some subsurface Cretaceous formations in theBadr El Dein (Bed 1-1) borehole, north Western Desert,Egypt. Neues Jahrbuch f€ur Geologie und Pal€aontologie -Abhandlungen. 192:133–149.
El-Beialy SY, El-Soughier M, Abdel Mohsen S, El Atfy H.2011. Palynostratigraphy and paleoenvironmental signifi-cance of the Cretaceous succession in the Gebel Rissu-1well, north Western Desert, Egypt. J Afr Earth Sci.59:215–226.
El-Shamma AE. 1991. Palynostratigraphic zonation of theBahariya Formation in El-Salam no. 17 well, WesternDesert, Egypt. Egyptian Journal of Geology. 35:261–273.
El Soughier MI, Mahmoud MS, and Li J. 2010. Palynologyand palynofacies of the Lower Cretaceous succession ofthe Matruh2-1X borehole, northwestern Egypt. RevistaEspa~nola de Micropaleontologia. 42(1):37–58.
Erba E, Channell JET, ClapsM, Jones C, Larson R, Opdyke B,Premoli Silva I, Riva A, Salvini G, Torricelli S. 1999. Inte-grated stratigraphy of the Cismon Apticore (SouthernAlps, Italy): a “reference” section for the Barremian-Aptianinterval at low latitudes. J Foramin Res. 29(4):371–391.
Foucher J-C, Pons D, Mami L, Bellier J-P. 1994. Premierinventaire de la microflore Cr�etac�ee (dinokystes, sporeset pollen) du Sud-Est constantinois (Alg�erie): Cons�equen-ces biostratigraphiques. Compte rendu hebdomadaire desseances de l’Academie des Sciences Paris. 318:1563–1570.
Green OR. 2001. A Manual of Practical Laboratory andField Techniques in Palaeobiology. Dordrecht: KluwerAcademic Publishers; p. 538.
Guiraud R. 1998. Mesozoic rifting and basin inversion alongthe northern African Tethyan margin: an overview. In:Macgregor DS, Moody RT, Clark-Lowes DD, editors.Petroleum Geology of North Africa. London: GeologicalSociety; p. 217–229.
Guiraud R, Bosworth W. 1999. Phanerozoic geodynamicevolution of north-eastern Africa and the north-westernArabian platform. Tectonophysics. 315(1–4):73–108.
Guiraud R, Issawi B, Bosworth W. 2001. Phanerozoic historyof Egypt and surrounding areas. In: Ziegler PA, CavazzaW, Robertson AHF, Crasquin-Soleau S, editors. Peri-Tethys Memoir 6: Peri-Tethyan Rift/Wrench Basins andPassive Margins. Paris: M�emoires du Mus�eum National�dHistoire Naturelle; p. 469–509.
Habib D, Drugg WS. 1983. Dinoflagellate Age of MiddleJurassic-Early Cretaceous Sediments in the Blake-Bahama Basin. Initial Reports of the Deep Sea DrillingProject. 76(Nov):623–638.
Hantar G. 1990. North Western Desert. In: Said R, editor.The Geology of Egypt. Rotterdam: Balkema; p. 293–319.
Herngreen GFW. 1973. Palynology of Albian-Cenomanianstrata of borehole 1-QS-1-MA, State of Maranh~ao,Brazil. Pollen et Spores. 15:515–555.
Herngreen GFW. 1975. Palynology of middle and upper Cre-taceous strata in Brazil. Mededelingen Rijks GeologischeDienst, N.S. 26:39–91.
Herngreen GFW, Jimenez HD. 1990. Dating of the CretaceousUne Formation, Colombia and the relationship with theAlbian-Cenomanian African-South American microfloralprovince. Rev Palaeobot Palyno. 66(3–4):345–359.
Herngreen GFW, Kedves M, Rovnina LV, Smirnova SB.1996. Cretaceous palynofloral provinces: a review. In:Jansonius J, Mcgregor DC. editors. Palynology: Princi-ples and Applications. Texas: American Association ofStratigraphic Palynologists Foundation; p. 1157–1188.
Hochuli PA. 1981. North Gondwanan floral elements inlower to middle Cretaceous sediments of the southernAlps (southern Switzerland, northern Italy). Rev Palaeo-bot Palyno. 35:337–358.
Hochuli PA, Kelts K. 1980. Palynology of middle Cretaceousblack clay facies from Deep Sea Drilling Project Sites 417and 418 of the western North Atlantic. Initial Reports ofthe Deep Sea Drilling Project. 51–53:897–935.
Ibrahim MIA. 1996. Aptian-Turonian palynology of theGhazalat-1 well (GTX-1), Qattara Depression, Egypt.Rev Palaeobot Palyno. 94:137–168.
Ibrahim MIA. 2002. Late Albian-middle Cenomanian paly-nofacies and palynostratigraphy, Abu Gharadig-5 well,Western Desert, Egypt. Cretaceous Res. 23:775–788.
Jan Du Chene R, De Klasz I, Archibong EE. 1978. Biostrati-graphic study of the borehole OJO-1, SW Nigeria, withspecial emphasis on the Cretaceous microflora. Revue deMicropal�eontologie. 2:123–139.
Jardin�e S. 1967. Spores �a expansions en forme d`elateres duCr�etac�e moyen d`Afrique occidentale. Rev PalaeobotPalyno. 1:235–258.
Jardin�e S, Magloire L. 1965. Palynologie et stratigraphie duCr�etac�e, des bassins de S�en�egal et d` Cote De`Ivoire.M�emoire Bureau Recherche G�eologie et Min�eralogie.32:187–245.
Kerdany MT, Cherif OH. 1990. Mesozoic. In: Said R, editor.The Geology of Egypt. Rotterdam: Balkema; p. 407–437.
Lawal O, Moullade M. 1986. Palynological biostratigraphyof Cretaceous sediments in the Upper Benue Basin, NENigeria. Revue de Micropal�eontologie. 29:61–83.
Lawver LA, Dalziel IWD, Gahagan LM, Kygar RM, HerberBD. 2004. PLATES 2009 Atlas of Plate Reconstructions(750 Ma to Present Day). University of Texas Institutefor Geophysics. [viewed 2013 May 12] Available from:http://www.ig.utexas.edu/research/projects/plates/.
1205
1210
1215
1220
1225
1230
1235
1240
1245
1250
1255
1260
1265
1270
1275
1280
1285
1290
1295
1300
1305
1310
1315
1320
1325
1330
22 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
Mahmoud MS, Deaf AS. 2007. Cretaceous palynology(spores, pollen and dinoflagellate cysts) of the Siqeifa 1-Xborehole, northern Egypt. Rivista Italiana di Paleontolo-gia e Stratigrafia. 113(2):203–221.
Mahmoud MS, Moawad AMM. 2002. Cretaceous palynol-ogy of the Sanhur-1X borehole, northern western Egypt.Revista Espa~nola de Micropaleontologia. 34:129–143.
Meshref WM. 1990. Tectonic framework. In: Said R, editor.The Geology of Egypt.Rotterdam: Balkema; p. 113–156.
Morgan R. 1978. Albian to Senonian palynology of Site 364,Angola Basin. Initial Rep Deep Sea. 40:915–951.
M€uller J. 1966. Montane pollen from the Tertiary of NWBorneo. Blumea. 4:231–235.
Norton P. 1967. Rock stratigraphic nomenclature of theWestern Desert. Internal Report. Cairo.Q10
Omran AM, Soliman HA, Mahmoud MS. 1990. Early Creta-ceous palynology of three boreholes from northernWesternDesert (Egypt). Rev Palaeobot Palyno. 66(3–4):293–312.
Penny JHJ. 1986. An early Cretaceous angiosperm pollenassemblages from Egypt. In: Batten DJ, Briggs DEG,editors. Studies in Palaeobotany and Palynology in Hon-our of N. F. Hughes. London: Special Papers in Palaeon-tology; p. 119–132.
Phipps D, Playford G. 1984. Laboratory techniques for extrac-tion of palynomorphs from sediments. Papers, Deptart-ment of Geology, University of Queensland. 11:1–23.
Regali MSP, Uesugui N, Santos AS. 1974. Palinologia dossedimentos Meso-Cenozoicos do Brasil (I & II). BoletimTecnico da Petrobras. 17:177–191, 263–301.
Regali MSP, Viana CF. 1989. Late Jurassic-early Cretaceousin Brazilian sedimentary basins: correlation with theinternational standard scale. Report. Rio de Janeiro:Petrobras.
Roncaglia L, Corradini D. 1997. Correlation of key dinofla-gellate events with calcareous nannoplankton and plank-tonic foraminiferal zones in the Solignano Formation(Maastrichtian, late Cretaceous),northern Apennines,Italy. Rev Palaeobot. Palynol. 97:177–196.
Said R. 1962. Tectonic framework of Egypt. In: Said R, editor.The Geology of Egypt.Amsterdam: Elsevier; p. 28–44.
Salard-Cheboldaeff M. 1990. Intertropical African palynos-tratigraphy from Cretaceous to Quaternary times. J AfrEarth Sci. 11: 1–24.
Schrank E. 1992. Nonmarine Cretaceous correlations inEgypt and northern Sudan; palynological and palaeobo-tanical evidence. Cretaceous Res. 13(4):351–368.
Schrank E, Ibrahim MIA. 1995. Cretaceous (Aptian-Maastrichtian) palynology of foraminifera-dated wells(KRM-1, AG-18) in north-western, Egypt. BerlinerGeowissenschaftliche Abhandlungen - Reihe A. 177(Reihe A):1–44.
Schrank E, Mahmoud MS. 1998. Palynology (pollen, sporesand dinoflagellates) and Cretaceous stratigraphy of theDakhla Oasis, central Egypt. J Afr Earth Sci. 26(2):167–193.
Stover LE. 1963. Some middle Cretaceous palynomorphsfromWest Africa. J Micropalaeontol. 9:85–94.
Sultan IZ, Aly SM. 1986. Palynological zonation of Albian-Cenomanian sediments in the northern part of the West-ern Desert, Egypt. Bulletin of the Faculty of Science,Alexandria University. 26:80–101.
Tea-Yassi J, Digbehi ZB, Yao KR, Boblai V. 1999. Study ofsome palynomorphs of Upper Cretaceous in Coted’Ivoire sedimentary “Offshore” basin. Biostratigraphicand paleoenvironmental implications. J Afr Earth Sci. 29(4):783–798.
Thusu B, Van Der Eem JGLA. 1985. Early Cretaceous (Neo-comian-Cenomanian) palynomorphs. J Micropalaeontol.4(1):131–150.
Thusu B, Van Der Eem JGLA, El-Mehdawi A, Bu-ArgoubF. 1988. Jurassic-early Cretaceous palynostratigraphyin north-east Libya. In: El-Aranuti A, Owens B, ThusuB, editors. Subsurface Palynostratigraphy of NE Libya.Benghazi: Garyounis University Publications; p. 171–213.
Torricelli S. 2006. Dinoflagellate cyst stratigraphy of theScisti a Fucoidi Formation (early Cretaceous) from Piob-bico, central Italy: calibrated events for the Albian of theTethyan Realm. Rivista Italiana di Paleontologia e Strat-igrafia. 112(1):95–111.
Torricelli S, Amore MR. 2003. Dinoflagellate cysts and cal-careous nannofossils from the upper Cretaceous Sara-ceno Formation (Calabria, Italy): implications about thehistory of the Liguride complex. Rivista Italiana di Pale-ontologia e Stratigrafia. 109(3):499–516.
Uwins PJR, Batten DJ. 1988. Early to mid-Cretaceous paly-nology of north-east Libya. In: El-Aranuti A, Owens B,Thusu B, editors. Subsurface Palynostratigraphy of NELibya. Benghazi: Garyounis University Publications;p. 215–258.
WEPCO. 1968. Final report and composite log of the AbuTunis 1x borehole. Report. Cairo: Wepco.
1335
1340
1345
1350
1355
1360
1365
1370
1375
1380
1380
1385
1390
1395
1400
1405
1410
1415
1420
Palynology 23
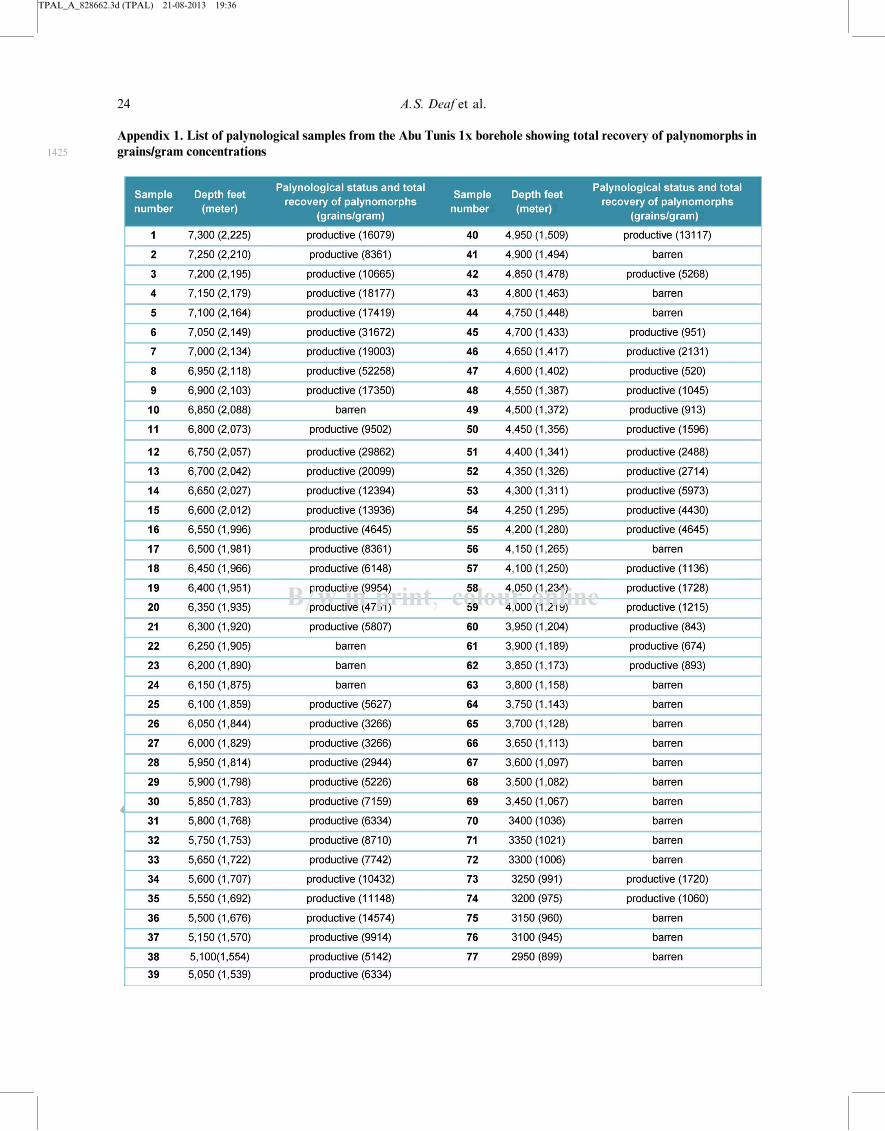
TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
Appendix 1. List of palynological samples from the Abu Tunis 1x borehole showing total recovery of palynomorphs in
grains/gram concentrations1425
B=w in print; colour online
24 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
Appendix 2. Alphabetic list of palynomorphs recorded in
the Abu Tunis 1x borehole
Spores
Balmeisporites cf. holodictyus Cookson & Dettmann, 1958 (9).Cicatricosisporites orbiculatus Singh, 1964 (4).Cicatricosisporites sinuosusHunt, 1985 (13).Cicatricosisporites spp. (7).Concavissimisporites spp. (11).Crybelosporites pannuceus (Brenner) Srivastava, 1977 (2).Deltoidospora halliiMiner, 1935 (5).Deltoidospora toralis (Leschik) Lund, 1977 (16).Deltoidospora spp. (1).Gleicheniidites senonicus Ross, 1949 (15).Todisporites minor Couper, 1958 (14).Trilobosporites laevigatus El-Beialy, 1994 (6).Triplanosporites sp. (3).Triporoletes reticulatus (Pocock) Playford, 1971 (12).Verrucosisproites obscurilaesuratus Pocock, 1962 (8).
Gymnosperm pollen
Alisporites cf. grandis (Cookson) Dettmann, 1963 (17).Arucariacites australis Cookson ex Couper, 1953 (24).Balmeiopsis limbatus (Balme) Archangelsky, 1979 (18).Classopollis classoides Pflug, 1953 (19).Classopollis spp. (23).Cycadopites spp. (22).Elaterocolpites castelainii Jardin�e &Magloire, 1965 (25).Elaterosporites acuminatus (Stover) Jardin�e, 1967 (28).Elaterosporites klaszii (Jardin�e &Magloire) Jardin�e, 1967 (21).Elaterosporites protensus (Stover) Jardin�e, 1967 (29).Elaterosporites verrucatus (Jardin�e & Magloire) Jardin�e, 1967(27).Ephedripites irregularisHerngreen, 1973 (33).Ephedripites spp. (20).Eucommidites treodsonii (Erdtman) Potoni�e, 1958 (34).Galeacornea causea Stover, 1963 (31).Gnetaceaepollenites cf. clathratus Stover, 1964 (30).Inaperturopollenites undulatusWeyland & Greifeld, 1953 (32).Sofrepites legouxae Jardin�e, 1967 (26).
Angiosperm pollen
Afropollis jardinusDoyle et al., 1982 (36).Afropollis aff. jardinusDoyle et al., 1982 (66).Afropollis kahramanensis Ibrahim & Schrank 1995 (42).Cretacaeiporites densimurus Schrank & Ibrahim, 1995 (38).Cretacaeiporites mulleriiHerngreen, 1973 (45).Cretacaeiporites polygonalis (Jardin�e & Magloire) Herngreen,1973 (52).Dichastopollenites ghazalatensis Ibrahim, 1996 (55).Foveotricolpites gigantoreticulatus (Jardin�e & Magloire)Schrank, 1987a (57).Papillopollis vancampoaeKedves & Pittau, 1979 (40).Proteacidites cf. africaensis (Jardin�e & Magloire) Schrank &Ibrahim, 1995 (35).Retimonocolpites ghazalii Ibrahim, 2002 (59).Retimonocolpites variplicatus Schrank &Mahmoud, 1998 (41).Retimonocolpites textus (Norris) Singh, 1983 (49).Rousea brenneri Singh, 1983 (50).Rousea delicipollis Srivastava, 1977 (44).Rousea cf. miculipollis Srivastava, 1975 (47).Stellatopollis barghoorniiDoyle in Doyle et al., 1976 (61).Stellatopollis dejaxii Ibrahim, 2002 (64).
Stellatopollis densiornatus (Lima) Ward, 1986 (62).Stellatopollis doylei Ibrahim, 2002 (69).Stellatopollis limai Ibrahim, 2002 (65).Stellatopollis spp. (68).Stephanocolpites sp. (60).Striatopollis cf. trochuensis (Srivastava) Ward, 1986 (63).Tetracolpites sp. (67).Tetraporopollenites sp. (39).Tricolpites cf. crassimurus (Groot & Penny) Singh, 1971 (56).Tricolpites micromunus (Groot & Penny) Singh, 1971 (54).Tricolpites parvus Stanley, 1965 (51).Tricolpites sagaxNorris, 1967 (58).Tricolpites vulgaris (Pierce) Srivastava, 1969 (48).Tricolpites spp. (37).Tricolporopollenites sp. (43).Triporites spp. (46).Triporopollenites spp. (53).
Pollen tetrads
Cretacaeiporites densimurus Schrank & Ibrahim, 1995 (70).Droseridites baculites Ibrahim, 1996 (71).Droseridites senonicus Jardin�e & Magloire, 1965 (72).
Freshwater algae
Botryococcus sp. (75).Chomotriletes minor (Kedves) Pocock, 1970 (76).Fungal fruiting body (74).Ovoidites parvus (Cookson &Dettmann) Nakoman, 1966 (73).
Dinoflagellate cysts
Canningia senonica Clarke & Verdier, 1967 (79).Cannosphaeropsis utinensisWetzel, 1933 (96).Chatangiella madura Lentin & Williams, 1976 (81).Chlamydophorella discreta Clarke & Verdier, 1967 (91).Circulodinim distinctum (Deflandre & Cookson) Jansonius,1986 (100).Coronifera albertiiMillioud, 1969 (101).Coronifera oceanica Cookson & Eisenak, 1958 (97).Coronifera tubulosa Cookson & Eisenak, 1974 (104).Coronifera spp. (113).Cribroperidinium edwardsii (Cookson & Eisenack) Davey,1969 (108).Cribroperidenium sp. (110).Dinogymnium denticulatum (Alberti) Evitt et al., 1967 (89).Dinogymnium spp. (82).Downiesphaeridium sp. (92).Eucladinium gambangense (Cookson & Eisenack) Stover &Evitt, 1978 (93).Exochosphaeridium bifidum (Clarke & Verdier) Clarke et al.,1968 (80).Florentinia berran Below, 1982 (98).Florentinia laciniataDavey & Verdier, 1973 (103).Florentinia mantellii (Davey & Williams) Davey & Verdier,1973 (99).Florentinia spp. (94).Isabelidinium acuminatum (Cookson & Eisenack) Stover &Evitt, 1978 (90).Litosphaeridium siphoniphorum (Cookson & Eisenack) Davey&Williams, 1966 (86).Odontochitina operculata (Wetzel) Deflandre & Cookson,1955 (78).
1430
1435
1440
1445
1450
1455
1460
1465
1470
1475
1480
1485
1490
1495
1500
1505
1510
1515
1520
1525
1530
1535
1540
Palynology 25

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
Odontochitina costata Alberti, 1961 (83).Odontochitina porifera Cookson, 1956 (84).Oligosphaeridium albertense (Pocock) Davey & Williams,1969 (105).Oligosphaeridium complex (White) Davey & Williams, 1966(106).Oligosphaeridium poculum Jain, 1977 (107).Pseudoceratium anaphrissum (Sarjeant) Bint, 1986 (111).Pseudoceratium securigerum (Davey & Verdier) Bint, 1986(112).Pterodinium sp. (102).Senegalinium aenigmaticum (Boltenhagen) Lentin & Wil-liams, 1981 (85).Spinifereites spp. (77).Subtilisphaera senegalensis Jain & Millepied, 1973 (109).Surculosphaeridium cf. longifurcatum (Firtion) Davey et al.,1966 (88).Trichodinium castaneaDeflandre, 1935 (87).Xiphophoridium alatum (Cookson & Eisenack) Sarjeant, 1966(95).
Acritarchs
Baltisphaeridium spp. (114).Micrhystridium spp. (115).
Microforaminiferal test linings (116)
Albian angiosperm pollen grains
1–5. Afropollis jardinus Doyle et al. 1982, 1, sample 89/A,R13; 2, sample 110/A, L46/3; 3, sample 89/B, Q22; 4, sample93/A, N19/3; 5, sample 110/A, L46/3, (36).
Mid Albian elaterate gymnosperm pollen grains
6, 8. Elaterosporites verrucatus (Jardin�e & Magloire) Jardin�e1967, sample 88/B, M31/3, sample 93/A, J23/3, (27).
7, 14. Elaterosporites acuminatus (Stover) Jardin�e 1967,sample 93/A, G45/4, sample 91/A, H32, (28).
9. Elaterosporites protensus (Stover) Jardin�e 1967, sample90/A, U16, (29).
10. Elaterosporites klaszii (Jardin�e & Magloire) Jardin�e1967, sample 97/A, G13/4, (21).
Late Albian elaterate gymnosperm pollen grains
13, 15–17. Sofrepites legouxae Jardin�e 1967, 13, sample 93/A,M32/3; 15, sample 86/B, Q30; 16, sample 93/A, T23/2; 17,sample 95/A, H17/3, (26).
Late Albian–Mid Cenomanian pollen grains
11. Cretacaeiporites mullerii Herngreen 1973, sample 103/A,U25, (45).
12. Cretacaeiporites polygonalis (Jardin�e & Magloire)Herngreen 1973, 94/A, K32/3, (52).
18. Ephedripites irregularis Herngreen 1973, sample 77/B,C19/2, (33).
19. Gnetaceaepollenites cf. clathratus Stover 1964, sample83/A, K44/1, (30).
20. Galeacornea causea Stover 1963, sample 84/A, K46,(31).
21. Afropollis kahramanensis Ibrahim & Schrank 1995,sample 88/A, Q38/1, (42).
22. Foveotricolpites gigantoreticulatus (Jardin�e &Magloire) Schrank 1987, sample 92/A, L13/3, (57).
Albian–Cenomanian spore and pollen grains
1. Tetracolpites sp., sample 77/B, G28/2, (67).2. Tricolpites sagaxNorris 1967, sample 92/A, G29, (58).3. Striatopollis cf. trochuensis (Srivastava) Ward 1986,
sample 86/A, Q42/4, (63).4. Stephanocolpites sp., sample 91/A, Q23/3, (60).5. Tricolpites parvus Stanley 1965, sample 94/A, M36,
(51).6. Tricolpites cf. crassimurus (Groot & Penny) Singh
1971, sample 85/A, J20/4, (56).7. Rousea delicipollis Srivastava 1977, sample 71/A, D34/
2, (44).8. Tricolpites micromunus (Groot & Penny) Singh 1971,
sample 75/A, Q43/3, (54).9. Triporopollenites sp., sample 94/A, S45, (53).10. Rousea cf. miculipollis Srivastava 1975, sample 97/A,
P38/2, (47).11. Proteacidites cf. africaensis (Jardin�e & Magloire)
Schrank & Ibrahim 1995, sample 85/A, O42, (35).12. Tricolporopollenites sp., sample 93/B, W14/3, (43).15. Droseridites baculites Ibrahim 1996, sample 92/A,
Q40/4, (71).16. Droseridites senonicus Jardin�e & Magloire 1965, sam-
ple 77/B, P27/4, (72).17. Crybelosporites pannuceus (Brenner) Srivastava 1977,
sample 91/A, L44, (2).
Cretaceous spore and pollen grains
13, 14, 19–22. Retimonocolpites textus (Norris) Singh 1983, 13and 14, sample 94/A, N28/3; 19 and 20, sample 89/A, K40/1;21, sample 94/A, G31/4; 22, sample 94/A, G31/4, (49).
18. Dichastopollenites ghazalatensis Ibrahim 1996, sample88/B, X37/2, (55).
24. Retimonocolpites variplicatus Schrank & Mahmoud1998, sample 90/A, Q30, (41).
Fresh water algae
23. Botryococcus sp., sample 99/B, R42/4, (75).25. Fungal fruiting body, sample 109/B, N34/4, (74).
Santonian dinoflagellate cyst
1. Canningia senonica Clarke & Verdier 1967, sample 131/B,L22, (79).
Post-Turonian dinoflagellate cysts
2, 6, 15. Dinogymnium denticulatum (Alberti) Evitt et al.1967, sample 130/A, Q36, sample 130/A, P42, sample 130/B,N29/4, (89).
3, 7. Eucladinium gambangense (Cookson & Eisenack)Stover & Evitt 1978, sample 130/A, K28, sample 130/B, Q42/4,(93).
4, 13. Isabelidinium acuminatum (Cookson & Eisenack)Stover & Evitt 1978, sample 131/A, U27/2, sample 131/B,M34, (90).
1545
1550
1555
1560
1565
1570
1575
1580
1585
1590
1595
1600
1605
1610
1615
1620
1625
1630
1635
1640
1645
1650
26 A.S. Deaf et al.

TPAL_A_828662.3d (TPAL) 21-08-2013 19:36
9. Dinogymnium sp., sample 130/B, S45/1, (82).10. Dinogymnium sp., sample 131/B, U26/3, (82).12. Chatangiella madura Lentin & Williams 1976 sample
134/A, U26, (81).14. Exochosphaeridium bifidum (Clarke & Verdier) Clarke
et al. 1968 sample 133/A, C11/3, (80).17. Cannosphaeropsis utinensis Wetzel 1933, sample 117/
A, S33/3, (96).19. Odontochitina porifera Cookson 1956, sample 133/A,
L39/1, (84).
Cretaceous dinoflagellate cysts
5. Trichodinium castanea Deflandre 1935, sample 116/A, L12/4, (87).
8. Xiphophoridium alatum (Cookson & Eisenack) Sarjeant1966, sample 103/B, E43/4, (95).
11, 18. Senegalinium aenigmaticum (Boltenhagen) Lentin& Williams 1981, sample 110/A, G13/3, sample 109/A, S31/1,(85).
16. Odontochitina costata Alberti 1961, sample 134/A,V38/2, (83).
1655
1660
1665
1670
Palynology 27
Copyright © 2022 FDOKUMEN




















