
Production and biochemical characterization of a type B ferulic acid esterase from Streptomyces...
11
Research Article Production and Biochemical Characterization of a High Maltotetraose (G4) Producing Amylase from Pseudomonas stutzeri AS22 Hana Maalej, Hanen Ben Ayed, Olfa Ghorbel-Bellaaj, Moncef Nasri, and Noomen Hmidet Laboratoire de G´ enie Enzymatique et de Microbiologie, Ecole Nationale d’Ing´ enieurs de Sfax, BP 1173, 3038 Sfax, Tunisia Correspondence should be addressed to Noomen Hmidet; hmidet [email protected] Received 24 February 2014; Accepted 22 April 2014; Published 26 May 2014 Academic Editor: Sofiane Ghorbel Copyright © 2014 Hana Maalej et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Amylase production and biochemical characterization of the crude enzyme preparation from Pseudomonas stutzeri AS22 were evaluated. e highest -amylase production was achieved aſter 24 hours of incubation in a culture medium containing 10g/L potato starch and 5 g/L yeast extract, with initial pH 8.0 at 30 ∘ C under continuous agitation at 200 rpm. e optimum temperature and pH for the crude -amylase activity were 60 ∘ C and 8.0, respectively. e effect of different salts was evaluated and it was found that both -amylase production and activity were Ca 2+ -dependent. e amylolytic preparation was found to catalyze exceptionally the formation of very high levels of maltotetraose from starch (98%, w/w) in the complete absence of glucose since the initial stages of starch hydrolysis (15 min) and hence would have a potential application in the manufacturing of maltotetraose syrups. 1. Introduction Maltotetraose, known as G4, is an oligosaccharide of 4 units of -D-glucopyranose linked by -(1–4) bond. is compound has a considerable specific interest like its use as a substrate of amylases to study their mode of action and as a highly sensitive substrate for detection of specific -amylase activity when coupled with a chromogenic compound [1]; it also finds potential applications in the food industries due to its properties [2, 3]. In fact, it can be used in baking due to its high moisture retention power which serves to prevent retrogradation of starch ingredient [4]. Maltotetraose is also being tested for its use as a food additive to improve the texture or to reduce the sweetness of foods without affecting their inherent taste and flavor [5, 6]. Besides the above properties, G4 syrup, considered as a partially undigested and unabsorbed substrate in the small intestine, has shown a prebiotic effect by selectively promot- ing the growth and/or activity of beneficial bacteria, once it reaches the colon. In fact it has been demonstrated through in vitro and in vivo experiments that oligosaccharides were utilized by bifidobacteria classified as beneficial intestinal bacteria, but they were not utilized by Escherichia coli or the Clostridium species which were unfavourable for their producing putrefactive substances (protein degrading) in the digestive tract. Consequently, the ingestion of G4 syrup could improve intestinal flora and suppress the formation of putrefactive products [7]. However, the preparation of maltooligosaccharides with a specific degree of polymerisation (DP) in larger amounts is so expensive. erefore, the discovery of microbial enzymes that produce from starch maltooligosaccharides of a specific length has made it possible to produce with good yield various maltooligosaccharides [8]. Microbial amylases are produced mainly from cultures of Aspergillus, Bacillus, Streptomyces, and Pseudomonas species [9]. Amylolytic activity is one of phenotypic characteristics of Pseudomonas stutzeri species [10], especially G4--amylases that have been subject of intense biochemical research such as purification and biochemical characterization [11]. In the view of advantages offered by the use of the amylolytic preparation instead of the purified enzyme, such as to avoid enzyme purification procedures that are expen- sive and time-consuming, we report in the present study Hindawi Publishing Corporation BioMed Research International Volume 2014, Article ID 156438, 11 pages http://dx.doi.org/10.1155/2014/156438
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Production and biochemical characterization of a type B ferulic acid esterase from Streptomyces...

Research ArticleProduction and Biochemical Characterization ofa High Maltotetraose (G4) Producing Amylase fromPseudomonas stutzeri AS22
Hana Maalej Hanen Ben Ayed Olfa Ghorbel-Bellaaj Moncef Nasri and Noomen Hmidet
Laboratoire de Genie Enzymatique et de Microbiologie Ecole Nationale drsquoIngenieurs de Sfax BP 1173 3038 Sfax Tunisia
Correspondence should be addressed to Noomen Hmidet hmidet noomenyahoofr
Received 24 February 2014 Accepted 22 April 2014 Published 26 May 2014
Academic Editor Sofiane Ghorbel
Copyright copy 2014 Hana Maalej et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Amylase production and biochemical characterization of the crude enzyme preparation from Pseudomonas stutzeri AS22 wereevaluated The highest 120572-amylase production was achieved after 24 hours of incubation in a culture medium containing 10 gLpotato starch and 5 gL yeast extract with initial pH 80 at 30∘C under continuous agitation at 200 rpmThe optimum temperatureand pH for the crude 120572-amylase activity were 60∘C and 80 respectively The effect of different salts was evaluated and it was foundthat both 120572-amylase production and activity were Ca2+-dependentThe amylolytic preparation was found to catalyze exceptionallythe formation of very high levels of maltotetraose from starch (98 ww) in the complete absence of glucose since the initial stagesof starch hydrolysis (15min) and hence would have a potential application in the manufacturing of maltotetraose syrups
1 Introduction
Maltotetraose known as G4 is an oligosaccharide of 4units of 120572-D-glucopyranose linked by 120572-(1ndash4) bond Thiscompound has a considerable specific interest like its use as asubstrate of amylases to study their mode of action and as ahighly sensitive substrate for detection of specific 120572-amylaseactivity when coupled with a chromogenic compound [1] italso finds potential applications in the food industries dueto its properties [2 3] In fact it can be used in baking dueto its high moisture retention power which serves to preventretrogradation of starch ingredient [4] Maltotetraose is alsobeing tested for its use as a food additive to improve thetexture or to reduce the sweetness of foods without affectingtheir inherent taste and flavor [5 6]
Besides the above properties G4 syrup considered as apartially undigested and unabsorbed substrate in the smallintestine has shown a prebiotic effect by selectively promot-ing the growth andor activity of beneficial bacteria once itreaches the colon In fact it has been demonstrated throughin vitro and in vivo experiments that oligosaccharides wereutilized by bifidobacteria classified as beneficial intestinal
bacteria but they were not utilized by Escherichia coli orthe Clostridium species which were unfavourable for theirproducing putrefactive substances (protein degrading) inthe digestive tract Consequently the ingestion of G4 syrupcould improve intestinal flora and suppress the formation ofputrefactive products [7]
However the preparation of maltooligosaccharides witha specific degree of polymerisation (DP) in larger amounts isso expensive Therefore the discovery of microbial enzymesthat produce from starch maltooligosaccharides of a specificlength has made it possible to produce with good yieldvarious maltooligosaccharides [8]
Microbial amylases are produced mainly from cultures ofAspergillus Bacillus Streptomyces and Pseudomonas species[9] Amylolytic activity is one of phenotypic characteristics ofPseudomonas stutzeri species [10] especially G4-120572-amylasesthat have been subject of intense biochemical research suchas purification and biochemical characterization [11]
In the view of advantages offered by the use of theamylolytic preparation instead of the purified enzyme suchas to avoid enzyme purification procedures that are expen-sive and time-consuming we report in the present study
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 156438 11 pageshttpdxdoiorg1011552014156438
2 BioMed Research International
the isolation and characterization of a novel G4-120572-amylaseproducing bacteriaPseudomonas stutzeriAS22 fromTunisiansoil samples Conditions of 120572-amylase production were opti-mized to achieve high enzyme production and the amylolyticpreparation was characterized
2 Materials and Methods
21 Strain Isolation and Identification Various soil sampleswere collected in the region of Sfax (Tunisia) and differentmicroorganisms were screened for their amylolytic activityusing nutrient agar plates containing soluble starch (1 wv)(Figure 1(a)) The strain having the highest activity wasidentified as Pseudomonas stutzeri by using the phylogeneticanalysis based on the 16S rDNA sequence analysis GenomicDNA for the PCR template was isolated from bacterial cellsgrown in Luria-Bertani (LB) media overnight by theWizardGenomic DNA Purification Kit from Promega and amplifiedusing the universal oligonucleotide primers (Bio BasicInc) 16SF (51015840GCTAACTAACGTGCCAGCAG) and 16SR(51015840CCCGGGATCCAAGCTTAAGGAGGTGATCCAGCC)Nucleotide sequence of the amplified 16S rDNA gene regionwas compared with those available in the GenBank databaseby using the BLAST method The BLAST result showed thatthe 16S rDNA sequence of the isolated strain AS22 has 99sequence similarity with the strain Pseudomonas stutzeri
22 Medium Composition and Culture Condition Inoculawere routinely grown in Luria-Bertani (LB) broth mediumcomposed of (gL) peptone 100 yeast extract 50 and NaCl50 [12] and the initial pH was adjusted to 70
The basal liquid culture medium used for 120572-amylaseproduction by the P stutzeri AS22 strain was composed of(gL) carbon source 10 ammonium sulphate as nitrogensource 1 MgSO
4(7 H2O) 01 K
2HPO414 KH
2PO407 and
NaCl 05 The medium was adjusted to pH 80 Media wereautoclaved at 121∘C for 20min
The strain was cultivated in 250mL conical flasks con-taining 25mL medium inoculated at initial OD of 0016and maintained for 24 h at 37∘C and 200 rpm The cultureswere centrifuged at 13000 rpm for 15min and the cell-freesupernatants were evaluated for their amylolytic activity
23 120572-Amylase Activity Assay 120572-Amylase activity was mea-sured by the determination of reducing sugars releasedduring starch hydrolysis by the dinitrosalicylic acid (DNS)method [13] The reaction mixture containing 05mL ofappropriately diluted enzyme and 05mL of 10 (wv)soluble potato starch (Sigma) in 100mMTris-HCl buffer (pH80) was incubated at 60∘C for 10min After that 3mL ofDNS reagent was added to the reaction volume boiled for10min and mixed with 20mL distilled water To determinethe activity the absorbance was measured at 550 nm and oneunit (U) of 120572-amylase activity was defined as the amountof enzyme that released 1 120583mol of reducing end groups perminute under the assay conditions
24 120572-Amylase Localization The 24 h culture broth (25mL)ofP stutzeriAS22was centrifuged at 13000timesg for 10min andthe supernatant was considered as the extracellular fractionThe cell pellet was washed twice with distilled water andsuspended in 5mL of Sodium Chloride-Tris-EDTA (STE)buffer containing 10mM Tris-HCl (pH 80) 100mM NaCland 1mM EDTA Then lysozyme was added to a finalconcentration of 200120583gmL and the mixture was incubatedat 0∘C for 1 h After centrifugation at 8000timesg for 25min thesupernatant was considered as the periplasmic fraction and5mL of Tris-HCl buffer (100mM pH 80) was added Theobtained mixture was sonicated twice for 3min each timeto disrupt cells The homogenate was finally centrifuged andits supernatant was considered as the intracellular fractionFinally the obtained three fractions were tested for theiramylolytic activity
25 Optimization of 120572-Amylase Production
251 Effects of Different Carbon Sources The effects of differ-ent carbon sources (glucose lactose maltose potato starchwheat starch and maize starch) on 120572-amylase production bythe P stutzeri strain were examined at a concentration of 1keeping constant the rest of the media composition The bestof these carbon sources was further optimized in the range of025ndash2 (wv)
252 Effects of Different Nitrogen Sources To investigate theeffects of different nitrogen sources on120572-amylase productionammonium sulphate in the basal medium containing 10 gLpotato starch was replaced with different organic (yeastextract casein pastone and soya peptone) and inorganic(ammonium sulphate and ammonium chloride) compoundsas nitrogen source at a concentration of 01 (wv) keepingconstant the rest of the media composition The concentra-tion of the selected nitrogen source was further optimized inthe range of 01ndash1 (wv)
253 Effects of Temperature Agitation Initial pH Saltsand Incubation Time on 120572-Amylase Production The effectof incubation temperature on 120572-amylase production wasinvestigated by incubating the media for 24 h at differenttemperatures (25 30 37 and 45∘C) in an automatic incubator
The effect of agitation on enzyme production was alsodetermined by incubating the inoculated culture flasks inan automatic mechanical shaker for 24 hours at 150 200and 250 rpm and then checking for extracellular 120572-amylaseproduction
To investigate the effect of pH on enzyme production theinitial pH of the medium was adjusted from 60 to 120
KH2PO4(07 gL) K
2HPO4(14 gL) NaCl (05 gL)
MgSO4 andCaCl
2were incorporated into growthmedium to
study the effects ofmineral sources on 120572-amylase productionThe basal medium containing 10 and 5 gL potato starch andyeast extract respectively was used as control To study theeffect of CaCl
2andMgSO
4 these chemicals were added to the
medium so that their final concentration ranged from 01 to
BioMed Research International 3
(a) (b)
Figure 1 Amylolytic (a) and proteolytic (b) activities of the isolated strain AS22 The amylolytic and proteolytic activities of the strainAS22 were evaluated by puncture inoculation of the strain inside nutrient agar medium containing soluble starch (1 wv) (a) and casein(b) respectively After incubation at 37∘C for 24 h to detect the amylolytic activity the plates were flooded with iodine solution at roomtemperature
08 gL Amylolytic activity was determined after incubationfor 24 h at 30∘C and 200 rpm agitation speed
For maximal 120572-amylase production fermentation period(0ndash240 h) was also optimized under the optimized fermenta-tion conditions Fermentation was performed at 30∘C in anautomatic incubator and samples were prepared at differentintervals (continuously for 10 days) for enzyme assay
26 Biochemical Properties of the Crude120572-Amylase Preparation
261 Effect of Temperature on Activity and Stability Theeffect of temperature on 120572-amylase activity was studied from20 to 90∘C Thermal stability was examined by incubatingthe enzyme preparation for 60min at different temperaturesranging from 30 to 70∘C Aliquots were withdrawn at desiredtime intervals and the remaining activitywasmeasured underenzyme assay conditions The nonheated enzyme was takenas 100
262 Effect of pH on Activity and Stability The effect of pHon 120572-amylase activity was evaluated for the pH range of 30ndash130 at 60∘C For pH stability measurement the enzyme wasincubated at 30∘C for 1 h in different buffers and the residualactivity was determined under the enzyme assay conditionsThe following buffer systems were used 100mM glycine-HCl buffer pH 30-40 100mM acetate buffer pH 40ndash60100mM Tris-HCl buffer pH 70-80 100mM glycine-NaOHbuffer pH 90ndash110 and 100mMNa
2HPO4-NaOH buffer pH
120 and 130
263 Effect of Metal Ions Surfactants and Enzyme InhibitorsThe influence of various metal ions (5mM) on 120572-amylaseactivity was investigated using CaCl
2 ZnCl
2 FeCl
2 HgCl
2
BaCl2 MnCl
2 MgCl
2 CuCl
2 CdCl
2 NaCl and KCl Activity
in the absence of any additives was taken as 100The effects of some surfactants (Triton X-100 Tween 20
Tween 80 and SDS) and enzyme inhibitors (phenylmethyl-sulfonyl fluoride (PMSF) 120573-mercaptoethanol and ethylene-diaminetetraacetic acid (EDTA)) on 120572-amylase stability werestudied by preincubating the amylolytic preparation for30min at room temperature The remaining enzyme activitywas measured under enzyme assay conditions The activityof enzyme incubated under similar conditions without anyadditive was taken as 100
27 Analysis of End Products Starch Hydrolysis Starch solu-tion (1 wv 50mL) was prepared using Tris-HCl buffer(100mM pH 80) The P stutzeri AS22 crude 120572-amylase(06U) was added into 50mL of starch solution Reactionmixture was incubated at 60∘C Aliquots were withdrawnat various time intervals and boiled for 10min to stopthe reaction and samples were then centrifuged at 13000 gfor 10min and passed through 022 120583m filter Samples thusobtained were analyzed by thin-layer chromatography (TLC)and gel permeation chromatography (GPC) on Bio-Gel P2
TLC was performed on silica gel 60 (20 times 20 cmMerck Germany) with a mobile phase composed of chlo-roformacetic acidwater (60 70 10 vvv) The spots werevisualized by spraying TLC plates withH
2SO4ethanol (5 95
vv) followed by heating at 120∘C for 10minGPC was performed on a Bio-Gel P2 column (15 times
200 cm) eluted with water at a rate of 30mLh The differentoligosaccharides in the range of G1 to G7 were fractionatedand the percentages of different products were determined
28 Visualisation of 120572-Amylase Activity by Zymography 120572-Amylase activity staining was done by layering the SDS-PAGE gel on a thin 2 agarose-1 soluble potato starch gel
4 BioMed Research International
incubated as a sandwich for 60min at 50∘CUpon staining theagarose gel with iodine solution at room temperature proteinbands with amylolytic activity became visible as a white bandagainst a dark blue background
3 Results and Discussion
31 120572-Amylase Production by P stutzeri AS22 The produc-tion of amylases enzymes by microorganisms is significantlyaffected by physical and chemical parameters of the medium[14 15] In this regard appropriate media components andsuitable conditions must be attained for optimal productionof the required products
311 Effects of Different Carbon Sources on 120572-Amylase Produc-tion Because amylase synthesis is known to be induced bystarch or its hydrolytic products [16ndash18] P stutzeri AS22 wasgrown in the basal medium supplemented with starch fromvarious natural sources (potato wheat andmaize starch) andother carbohydrates including lactose maltose and glucosefor assessing their effects on the production of amylase(Table 1)
The highest amylase activity (08UmL) was producedon potato starch followed by maltose (05UmL) Howeveramylase production was significantly low when the strainwas grown on lactose (0055UmL) and glucose (01 UmL)This study falls in line with previous studies on P stutzeriamylase production in which enzyme was induced usingstarch amylodextrin or maltose while glucose was foundto inhibit amylase production [1 19ndash21] In contrast to ourresults glucose was found to be the best carbon source foramylase production by Pseudomonas sp IMD 353 (13UmL)while the amylolytic activity decreased to 2 and 3UmLwhenmaltose and starch were used respectively as sole carbonsources in the same conditions [22]
Since potato starchwas the best carbon source for amylasesynthesis the effect of its concentration (025ndash2) on theamylase production was studied in media containing 01ammonium sulphate as nitrogen source It was observedthat the increase in concentration of potato starch increasesamylase production and maximum activity (075UmL) wasobtained in the presence of 1 substrate (data not shown)However further increase (15 and 2) of potato starchconcentration resulted in rapid decrease of enzyme produc-tion although biomass remained nearly constant (decreasedslightly) This may be explained by the degradation duringthe fermentation of starch by 120572-amylases resulting in theaccumulation of high quantities of reducing sugar which ledto an enhancement of sugar concentration and therefore tocatabolite repression of 120572-amylase synthesis [17]
312 Effects of Various Nitrogen Sources on 120572-Amylase Pro-duction In general both organic and inorganic nitrogensources were used efficiently in the growth medium for thebiosynthesis of 120572-amylase
In the present study various nitrogen sources at a con-centration of 01 were evaluated using an optimum potatostarch concentration of 1 (Table 2) Maximum activity was
Table 1 Effects of different carbon sources on the production of 120572-amylase by P stutzeri AS22
Carbon sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)Lactose 749 01 0055Glucose 569 08 01Maltose 662 10 05Potato starch 652 0 8 08Wheat starch 669 104 01Maize starch 659 05 017Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) carbon source 10 ammonium sulphate 1 K2HPO4 14KH2PO4 07 MgSO4 01 and NaCl 05 and that were inoculated at initial ODof 0016 and adjusted to pH 80
Table 2 Effects of different nitrogen sources supplemented to thepotato starch on the production of 120572-amylase by P stutzeri AS22
Nitrogen sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)NH4Cl 66 094 05(NH4)2SO4 65 09 07Soya peptone 79 321 097Casein 785 161 10Pastone 778 308 15Yeast extract 787 356 25Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 K2HPO4 14 KH2PO4 07MgSO4 01NaCl 05 and different nitrogen sources (1 gL) and were adjusted to pH 80
obtained when yeast extract was used (25UmL) Additionof yeast extract to the medium increased the production of120572-amylase activity by more than three times over mediumwith ammonium sulphate as nitrogen source Compared tothe organic nitrogen sources which support good growthand extracellular 120572-amylase production inorganic nitrogensources like ammonium sulphate and ammonium chlorideare not efficient for amylase productionThese results corrob-orate well with previous studies showing that organic nitro-gen sources were preferred for the 120572-amylase productionThe stimulatory effect of these complex compounds may beattributed to their carbohydrate and protein compositions aswell as to the trace of minerals and ions that could be presentand which enhance the enzyme secretion [18] Yeast extracthas been also used as nitrogen source in the culture mediumfor amylase production alone in the case of P stutzeri NRRLB-3389 [19] or in combination with other nitrogen sourcessuch as polypeptone in the case ofPseudomonas strainMS300[23] and yeatex for Pseudomonas sp IMD 353 [22] Otherorganic nitrogen sources have been also reported to support120572-amylase production by P stutzeriMO-19 such as corn steepliquor and peptone [20] and polypeptone in the case of Psaccharophila IAM 1504 [24]
To investigate the best concentration on 120572-amylase pro-duction yeast extract (range 1ndash10 gL) was added to basal
BioMed Research International 5
Table 3 Effect of different initial pH values of fermentationmediumon the production of 120572-amylase by P stutzeri AS22
Initial pH value Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)60 473 39 070 72 49 3880 80 56 5990 855 55 585100 89 55 58110 922 56 56120 932 54 54Cultivation was performed for 24 h at 30∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 yeast extract 5 K2HPO4 14 KH2PO407 MgSO4 01 and NaCl 05 and were adjusted to different initial pH values
medium containing potato starch (10 gL) Maximum amy-lase production by P stutzeri AS22 was obtained with 5 gLyeast extract reaching 395UmL and further addition ofyeast extract decreased the level of amylase (data not shown)
313 Effects of Temperature Agitation Initial pH Values andSalts on120572-Amylase Production Theeffects of temperature (25to 45∘C) agitation (150 200 and 250 rpm) and initial pH (60to 120) on 120572-amylase production by P stutzeri AS22 werestudied in optimizedmedium containing 10 gL potato starchand 5 gL yeast extract
Optimum level of 120572-amylase production (6UmL) wasachieved at 30∘C (data not shown) This result falls in linewith previous studies on Pseudomonas 120572-amylases in whichmost of them are reported to be produced at 25 [23] and 30∘C[20 24] Amylase production seems to be very sensitive tohigher temperatures Indeed at 37∘C the amylolytic activitywas 40 lower than that at 30∘C and at 45∘C no 120572-amylaseactivity was detected
Among the tested agitation speeds the maximum amy-lase activity (61 UmL) was found at 200 rpm (data notshown)
The pH of the production medium strongly affects manyenzymatic processes and transport of compounds across thecell membrane [18 25] In this study the effect of pH on 120572-amylase production by P stutzeri AS22 was investigated withvarying pHs (60ndash120) The results revealed that the pH ofthe medium influences the 120572-amylase production as well asthe strain growth (Table 3) which were more favorable atalkaline pH 80ndash120 than the neutral one In contrast acidicconditions (pH 60) seem to affect the 120572-amylase productioneven when the stain might be able to grow From this resultit has been understood that amylase productionmay occur atpH 60 but since the final pH was lowered to 45 no amylaseactivity was detected after 24 h of incubation because of theinstability of the enzyme toward acidic conditions Most ofthe earlier studies revealed neutral or alkaline initial pH for120572-amylase production by Pseudomonas species [20 22 24] Onthe other hand Lalucat et al [10] reported that none of thePseudomonas stutzeri strains tolerate acidic conditions andthey do not grow at pH 45
In an attempt to increase the 120572-amylase production by Pstutzeri the effect of some salts including K
2HPO4(14 gL)
KH2PO4(07 gL) NaCl (05 gL) MgSO
4(01 gL) and
CaCl2(01 gL) was investigated Results (Table 4) revealed
that even though the growth rate was lower in medium sup-plemented with phosphate ions (as K
2HPO4and KH
2PO4)
production of 120572-amylase was enhanced as compared to themediumwithout salts addition (none) It is also interesting tonote that while amylase production was slightly affected bythe addition in the culture medium of the monovalent cationNa+ (in the form of NaCl) divalent cations (01 gL) seem toenhance moderately or effectively the amylase production byP stutzeri AS22 in the case of Mg2+ and Ca2+ respectivelyAs mentioned by Gupta et al [18] differences in the min-eral requirements of amylase producing bacteria have beenreported and whereas most of previous studies indicated thataddition of CaCl
2to the fermentationmedium is known to be
essential for amylase production [9 26 27] Ca-independentamylase production has also been reported [28 29] Incomparison with the control medium when CaCl
2(04 gL)
was added individually as a mineral source to the mediumamylase production was enhanced from 59 to 66UmLHowever by increasing the CaCl
2concentration from 08 to
12 gL decline in enzyme production by approximately 30was observed This finding is in accordance with previousstudies showing inhibition of amylase production by theexcess of some minerals such Ca2+ [30] and Mg2+ ions [31]
Maximum amylase production (68UmL) occurredwhen CaCl
2(04 gL) was supplemented to the control
medium containing potato starch 10 gL yeast extract 5 gLMgSO
4(7 H2O) 01 gL K
2HPO414 gL KH
2PO407 gL
and NaCl 05 gL
314 Pattern of 120572-Amylase Production by P stutzeri AS22P stutzeri AS22 was grown aerobically in the optimizedmedium (potato starch 10 gL yeast extract 5 gL MgSO
4(7
H2O) 01 gL K
2HPO414 gL KH
2PO407 gL and NaCl
05 and CaCl204 gL) for 240 h at 30∘C The pattern of 120572-
amylase production and bacterial growth was followed withtime (Figure 2) Amylase production was initiated during theexponential growth phase of the strain reached 675UmLduring the stationary phase at 24 h and remained constanttill 72 h suggesting that the enzyme production is growthassociated and induced by the presence of starch hydrolysisproducts in the mediumThese results are in agreement withthe reports of Nakada et al [20] and Fogarty et al [22] onthe relationship between pattern of cell growth and120572-amylaseproduction
However the level of 120572-amylase increased slightly from72 h to 96 h and thereafter it increasedmore rapidly from 192to 216 h as the bacterial cells approached their death phaseSince the increase of 120572-amylase activity was concomitantwith the cell optical density decrease the amylase activityincrease may be the result of bacterial cell dying and releaseof intracellular or periplasmic 120572-amylases in the culturemedium Therefore attempts to confirm this statementresulted in the localization of 120572-amylase activity after 24 hof incubation It was found that P Stutzeri AS22 produced
6 BioMed Research International
Table 4 Effect of supplementation of the culture medium with various salts on the production of 120572-amylase by P stutzeri AS22
Chemicals Concentration (gL) Final pH value Biomass (OD600
) 120572-Amylase activity (UmL)None 72 477 35Control 78 545 59NaCl 05 732 49 38K2HPO4 14 76 257 519KH2PO4 07 77 201 59
MgSO4
01 708 543 41304 70 635 44208 73 573 324
CaCl2
01 69 715 60104 67 63 6608 65 619 6512 63 62 46
Control + CaCl2 (04 gL) 72 62 68Cultivation was performed for 24 h at 30∘C with shaking at 200 rpm in media that consisted of (gL) potato starch 10 yeast extract 5 and different salts andwere adjusted to pH 80 Control potato starch 10 yeast extract 5 K2HPO4 14 KH2PO4 07 MgSO4 01 and NaCl 05 gL
0
1
2
3
4
5
6
7
0
2
4
6
8
10
12
14
16
0 24 48 72 96 120 144 168 192 216
Cel
l gro
wth
(OD
600
nm)
Am
ylas
e act
ivity
(Um
L)
Time (h)Amylase activity (UmL)Growth (OD 600 nm)
Figure 2 Pattern of growth and 120572-amylase production of P stutzeriAS22 strain Culture was conducted in media consisting of (gL)potato starch 10 yeast extract 5 MgSO
4
01 K2
HPO4
14 KH2
PO4
07 CaCl2
04 gL and NaCl 05 gL Incubation was carried out at30∘C in a rotary shaker with stirring at 200 rpm
a dominant extracellular 120572-amylase as well as intracellularand periplasmic 120572-amylases which constitute approximately10 and 3 of the extracellular fraction respectively
A zymogram prepared using the cell-free culture super-natant at various time intervals (Figure 3) showed only onedominant protein band (PSA) in all the culture supernatantsTwominor amylases were also observed one active form (F1)was detected in 14ndash240 hr culture supernatants and the otherform (F2) was observed in 48ndash240 hr culture supernatantsThe change of PAGE pattern led us to suspect that F1 and F2are either produced from proteolytic degradation of PSA orlater secreted in the extracellular medium However since Pstutzeri AS22 lacks proteolytic activity (Figure 1(b)) and theintensity of the PSA band remains constant from beginningto 240 h this clearly indicates that F1 and F2 are not derived
14 h 24 h 36 h 48 h 72 h 96 h 240 h
PSA
F2F1
120 h 144 h 192 h
Figure 3 Changes in zymogram pattern for P stutzeri 120572-amylasesduring cultivation
fromPSAproteolysis but are later secreted in the extracellularmedium
Multiple forms of G4-amylase from Pseudomonas stutzeristrains have been reported Robyt and Ackerman [19]reported that P stutzeri NRRL B-3389 produced seven G4-amylases which differed in molecular mass and isoelectricpoint Strain MS300 was found to produce two major G4-amylases (amylases A and B) and two minor amylases(amylases C and D) [23] However the G4-amylase of PstutzeriMO-19 has been found to exist in two forms One ofthese is a 57 kDa protein (G4-1) the other is a 46 kDa protein(G4-2) which was derived by the limited proteolysis of G4-l[20]
32 Biochemical Characterization of Crude120572-Amylase Preparation
321 Effect of Temperature on 120572-Amylase Activity and Sta-bility The temperature activity profile shows that the crude120572-amylase was active at temperatures ranging from 20to 70∘C with an optimum activity at 60∘C (Figure 4(a))
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991

2 BioMed Research International
the isolation and characterization of a novel G4-120572-amylaseproducing bacteriaPseudomonas stutzeriAS22 fromTunisiansoil samples Conditions of 120572-amylase production were opti-mized to achieve high enzyme production and the amylolyticpreparation was characterized
2 Materials and Methods
21 Strain Isolation and Identification Various soil sampleswere collected in the region of Sfax (Tunisia) and differentmicroorganisms were screened for their amylolytic activityusing nutrient agar plates containing soluble starch (1 wv)(Figure 1(a)) The strain having the highest activity wasidentified as Pseudomonas stutzeri by using the phylogeneticanalysis based on the 16S rDNA sequence analysis GenomicDNA for the PCR template was isolated from bacterial cellsgrown in Luria-Bertani (LB) media overnight by theWizardGenomic DNA Purification Kit from Promega and amplifiedusing the universal oligonucleotide primers (Bio BasicInc) 16SF (51015840GCTAACTAACGTGCCAGCAG) and 16SR(51015840CCCGGGATCCAAGCTTAAGGAGGTGATCCAGCC)Nucleotide sequence of the amplified 16S rDNA gene regionwas compared with those available in the GenBank databaseby using the BLAST method The BLAST result showed thatthe 16S rDNA sequence of the isolated strain AS22 has 99sequence similarity with the strain Pseudomonas stutzeri
22 Medium Composition and Culture Condition Inoculawere routinely grown in Luria-Bertani (LB) broth mediumcomposed of (gL) peptone 100 yeast extract 50 and NaCl50 [12] and the initial pH was adjusted to 70
The basal liquid culture medium used for 120572-amylaseproduction by the P stutzeri AS22 strain was composed of(gL) carbon source 10 ammonium sulphate as nitrogensource 1 MgSO
4(7 H2O) 01 K
2HPO414 KH
2PO407 and
NaCl 05 The medium was adjusted to pH 80 Media wereautoclaved at 121∘C for 20min
The strain was cultivated in 250mL conical flasks con-taining 25mL medium inoculated at initial OD of 0016and maintained for 24 h at 37∘C and 200 rpm The cultureswere centrifuged at 13000 rpm for 15min and the cell-freesupernatants were evaluated for their amylolytic activity
23 120572-Amylase Activity Assay 120572-Amylase activity was mea-sured by the determination of reducing sugars releasedduring starch hydrolysis by the dinitrosalicylic acid (DNS)method [13] The reaction mixture containing 05mL ofappropriately diluted enzyme and 05mL of 10 (wv)soluble potato starch (Sigma) in 100mMTris-HCl buffer (pH80) was incubated at 60∘C for 10min After that 3mL ofDNS reagent was added to the reaction volume boiled for10min and mixed with 20mL distilled water To determinethe activity the absorbance was measured at 550 nm and oneunit (U) of 120572-amylase activity was defined as the amountof enzyme that released 1 120583mol of reducing end groups perminute under the assay conditions
24 120572-Amylase Localization The 24 h culture broth (25mL)ofP stutzeriAS22was centrifuged at 13000timesg for 10min andthe supernatant was considered as the extracellular fractionThe cell pellet was washed twice with distilled water andsuspended in 5mL of Sodium Chloride-Tris-EDTA (STE)buffer containing 10mM Tris-HCl (pH 80) 100mM NaCland 1mM EDTA Then lysozyme was added to a finalconcentration of 200120583gmL and the mixture was incubatedat 0∘C for 1 h After centrifugation at 8000timesg for 25min thesupernatant was considered as the periplasmic fraction and5mL of Tris-HCl buffer (100mM pH 80) was added Theobtained mixture was sonicated twice for 3min each timeto disrupt cells The homogenate was finally centrifuged andits supernatant was considered as the intracellular fractionFinally the obtained three fractions were tested for theiramylolytic activity
25 Optimization of 120572-Amylase Production
251 Effects of Different Carbon Sources The effects of differ-ent carbon sources (glucose lactose maltose potato starchwheat starch and maize starch) on 120572-amylase production bythe P stutzeri strain were examined at a concentration of 1keeping constant the rest of the media composition The bestof these carbon sources was further optimized in the range of025ndash2 (wv)
252 Effects of Different Nitrogen Sources To investigate theeffects of different nitrogen sources on120572-amylase productionammonium sulphate in the basal medium containing 10 gLpotato starch was replaced with different organic (yeastextract casein pastone and soya peptone) and inorganic(ammonium sulphate and ammonium chloride) compoundsas nitrogen source at a concentration of 01 (wv) keepingconstant the rest of the media composition The concentra-tion of the selected nitrogen source was further optimized inthe range of 01ndash1 (wv)
253 Effects of Temperature Agitation Initial pH Saltsand Incubation Time on 120572-Amylase Production The effectof incubation temperature on 120572-amylase production wasinvestigated by incubating the media for 24 h at differenttemperatures (25 30 37 and 45∘C) in an automatic incubator
The effect of agitation on enzyme production was alsodetermined by incubating the inoculated culture flasks inan automatic mechanical shaker for 24 hours at 150 200and 250 rpm and then checking for extracellular 120572-amylaseproduction
To investigate the effect of pH on enzyme production theinitial pH of the medium was adjusted from 60 to 120
KH2PO4(07 gL) K
2HPO4(14 gL) NaCl (05 gL)
MgSO4 andCaCl
2were incorporated into growthmedium to
study the effects ofmineral sources on 120572-amylase productionThe basal medium containing 10 and 5 gL potato starch andyeast extract respectively was used as control To study theeffect of CaCl
2andMgSO
4 these chemicals were added to the
medium so that their final concentration ranged from 01 to
BioMed Research International 3
(a) (b)
Figure 1 Amylolytic (a) and proteolytic (b) activities of the isolated strain AS22 The amylolytic and proteolytic activities of the strainAS22 were evaluated by puncture inoculation of the strain inside nutrient agar medium containing soluble starch (1 wv) (a) and casein(b) respectively After incubation at 37∘C for 24 h to detect the amylolytic activity the plates were flooded with iodine solution at roomtemperature
08 gL Amylolytic activity was determined after incubationfor 24 h at 30∘C and 200 rpm agitation speed
For maximal 120572-amylase production fermentation period(0ndash240 h) was also optimized under the optimized fermenta-tion conditions Fermentation was performed at 30∘C in anautomatic incubator and samples were prepared at differentintervals (continuously for 10 days) for enzyme assay
26 Biochemical Properties of the Crude120572-Amylase Preparation
261 Effect of Temperature on Activity and Stability Theeffect of temperature on 120572-amylase activity was studied from20 to 90∘C Thermal stability was examined by incubatingthe enzyme preparation for 60min at different temperaturesranging from 30 to 70∘C Aliquots were withdrawn at desiredtime intervals and the remaining activitywasmeasured underenzyme assay conditions The nonheated enzyme was takenas 100
262 Effect of pH on Activity and Stability The effect of pHon 120572-amylase activity was evaluated for the pH range of 30ndash130 at 60∘C For pH stability measurement the enzyme wasincubated at 30∘C for 1 h in different buffers and the residualactivity was determined under the enzyme assay conditionsThe following buffer systems were used 100mM glycine-HCl buffer pH 30-40 100mM acetate buffer pH 40ndash60100mM Tris-HCl buffer pH 70-80 100mM glycine-NaOHbuffer pH 90ndash110 and 100mMNa
2HPO4-NaOH buffer pH
120 and 130
263 Effect of Metal Ions Surfactants and Enzyme InhibitorsThe influence of various metal ions (5mM) on 120572-amylaseactivity was investigated using CaCl
2 ZnCl
2 FeCl
2 HgCl
2
BaCl2 MnCl
2 MgCl
2 CuCl
2 CdCl
2 NaCl and KCl Activity
in the absence of any additives was taken as 100The effects of some surfactants (Triton X-100 Tween 20
Tween 80 and SDS) and enzyme inhibitors (phenylmethyl-sulfonyl fluoride (PMSF) 120573-mercaptoethanol and ethylene-diaminetetraacetic acid (EDTA)) on 120572-amylase stability werestudied by preincubating the amylolytic preparation for30min at room temperature The remaining enzyme activitywas measured under enzyme assay conditions The activityof enzyme incubated under similar conditions without anyadditive was taken as 100
27 Analysis of End Products Starch Hydrolysis Starch solu-tion (1 wv 50mL) was prepared using Tris-HCl buffer(100mM pH 80) The P stutzeri AS22 crude 120572-amylase(06U) was added into 50mL of starch solution Reactionmixture was incubated at 60∘C Aliquots were withdrawnat various time intervals and boiled for 10min to stopthe reaction and samples were then centrifuged at 13000 gfor 10min and passed through 022 120583m filter Samples thusobtained were analyzed by thin-layer chromatography (TLC)and gel permeation chromatography (GPC) on Bio-Gel P2
TLC was performed on silica gel 60 (20 times 20 cmMerck Germany) with a mobile phase composed of chlo-roformacetic acidwater (60 70 10 vvv) The spots werevisualized by spraying TLC plates withH
2SO4ethanol (5 95
vv) followed by heating at 120∘C for 10minGPC was performed on a Bio-Gel P2 column (15 times
200 cm) eluted with water at a rate of 30mLh The differentoligosaccharides in the range of G1 to G7 were fractionatedand the percentages of different products were determined
28 Visualisation of 120572-Amylase Activity by Zymography 120572-Amylase activity staining was done by layering the SDS-PAGE gel on a thin 2 agarose-1 soluble potato starch gel
4 BioMed Research International
incubated as a sandwich for 60min at 50∘CUpon staining theagarose gel with iodine solution at room temperature proteinbands with amylolytic activity became visible as a white bandagainst a dark blue background
3 Results and Discussion
31 120572-Amylase Production by P stutzeri AS22 The produc-tion of amylases enzymes by microorganisms is significantlyaffected by physical and chemical parameters of the medium[14 15] In this regard appropriate media components andsuitable conditions must be attained for optimal productionof the required products
311 Effects of Different Carbon Sources on 120572-Amylase Produc-tion Because amylase synthesis is known to be induced bystarch or its hydrolytic products [16ndash18] P stutzeri AS22 wasgrown in the basal medium supplemented with starch fromvarious natural sources (potato wheat andmaize starch) andother carbohydrates including lactose maltose and glucosefor assessing their effects on the production of amylase(Table 1)
The highest amylase activity (08UmL) was producedon potato starch followed by maltose (05UmL) Howeveramylase production was significantly low when the strainwas grown on lactose (0055UmL) and glucose (01 UmL)This study falls in line with previous studies on P stutzeriamylase production in which enzyme was induced usingstarch amylodextrin or maltose while glucose was foundto inhibit amylase production [1 19ndash21] In contrast to ourresults glucose was found to be the best carbon source foramylase production by Pseudomonas sp IMD 353 (13UmL)while the amylolytic activity decreased to 2 and 3UmLwhenmaltose and starch were used respectively as sole carbonsources in the same conditions [22]
Since potato starchwas the best carbon source for amylasesynthesis the effect of its concentration (025ndash2) on theamylase production was studied in media containing 01ammonium sulphate as nitrogen source It was observedthat the increase in concentration of potato starch increasesamylase production and maximum activity (075UmL) wasobtained in the presence of 1 substrate (data not shown)However further increase (15 and 2) of potato starchconcentration resulted in rapid decrease of enzyme produc-tion although biomass remained nearly constant (decreasedslightly) This may be explained by the degradation duringthe fermentation of starch by 120572-amylases resulting in theaccumulation of high quantities of reducing sugar which ledto an enhancement of sugar concentration and therefore tocatabolite repression of 120572-amylase synthesis [17]
312 Effects of Various Nitrogen Sources on 120572-Amylase Pro-duction In general both organic and inorganic nitrogensources were used efficiently in the growth medium for thebiosynthesis of 120572-amylase
In the present study various nitrogen sources at a con-centration of 01 were evaluated using an optimum potatostarch concentration of 1 (Table 2) Maximum activity was
Table 1 Effects of different carbon sources on the production of 120572-amylase by P stutzeri AS22
Carbon sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)Lactose 749 01 0055Glucose 569 08 01Maltose 662 10 05Potato starch 652 0 8 08Wheat starch 669 104 01Maize starch 659 05 017Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) carbon source 10 ammonium sulphate 1 K2HPO4 14KH2PO4 07 MgSO4 01 and NaCl 05 and that were inoculated at initial ODof 0016 and adjusted to pH 80
Table 2 Effects of different nitrogen sources supplemented to thepotato starch on the production of 120572-amylase by P stutzeri AS22
Nitrogen sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)NH4Cl 66 094 05(NH4)2SO4 65 09 07Soya peptone 79 321 097Casein 785 161 10Pastone 778 308 15Yeast extract 787 356 25Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 K2HPO4 14 KH2PO4 07MgSO4 01NaCl 05 and different nitrogen sources (1 gL) and were adjusted to pH 80
obtained when yeast extract was used (25UmL) Additionof yeast extract to the medium increased the production of120572-amylase activity by more than three times over mediumwith ammonium sulphate as nitrogen source Compared tothe organic nitrogen sources which support good growthand extracellular 120572-amylase production inorganic nitrogensources like ammonium sulphate and ammonium chlorideare not efficient for amylase productionThese results corrob-orate well with previous studies showing that organic nitro-gen sources were preferred for the 120572-amylase productionThe stimulatory effect of these complex compounds may beattributed to their carbohydrate and protein compositions aswell as to the trace of minerals and ions that could be presentand which enhance the enzyme secretion [18] Yeast extracthas been also used as nitrogen source in the culture mediumfor amylase production alone in the case of P stutzeri NRRLB-3389 [19] or in combination with other nitrogen sourcessuch as polypeptone in the case ofPseudomonas strainMS300[23] and yeatex for Pseudomonas sp IMD 353 [22] Otherorganic nitrogen sources have been also reported to support120572-amylase production by P stutzeriMO-19 such as corn steepliquor and peptone [20] and polypeptone in the case of Psaccharophila IAM 1504 [24]
To investigate the best concentration on 120572-amylase pro-duction yeast extract (range 1ndash10 gL) was added to basal
BioMed Research International 5
Table 3 Effect of different initial pH values of fermentationmediumon the production of 120572-amylase by P stutzeri AS22
Initial pH value Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)60 473 39 070 72 49 3880 80 56 5990 855 55 585100 89 55 58110 922 56 56120 932 54 54Cultivation was performed for 24 h at 30∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 yeast extract 5 K2HPO4 14 KH2PO407 MgSO4 01 and NaCl 05 and were adjusted to different initial pH values
medium containing potato starch (10 gL) Maximum amy-lase production by P stutzeri AS22 was obtained with 5 gLyeast extract reaching 395UmL and further addition ofyeast extract decreased the level of amylase (data not shown)
313 Effects of Temperature Agitation Initial pH Values andSalts on120572-Amylase Production Theeffects of temperature (25to 45∘C) agitation (150 200 and 250 rpm) and initial pH (60to 120) on 120572-amylase production by P stutzeri AS22 werestudied in optimizedmedium containing 10 gL potato starchand 5 gL yeast extract
Optimum level of 120572-amylase production (6UmL) wasachieved at 30∘C (data not shown) This result falls in linewith previous studies on Pseudomonas 120572-amylases in whichmost of them are reported to be produced at 25 [23] and 30∘C[20 24] Amylase production seems to be very sensitive tohigher temperatures Indeed at 37∘C the amylolytic activitywas 40 lower than that at 30∘C and at 45∘C no 120572-amylaseactivity was detected
Among the tested agitation speeds the maximum amy-lase activity (61 UmL) was found at 200 rpm (data notshown)
The pH of the production medium strongly affects manyenzymatic processes and transport of compounds across thecell membrane [18 25] In this study the effect of pH on 120572-amylase production by P stutzeri AS22 was investigated withvarying pHs (60ndash120) The results revealed that the pH ofthe medium influences the 120572-amylase production as well asthe strain growth (Table 3) which were more favorable atalkaline pH 80ndash120 than the neutral one In contrast acidicconditions (pH 60) seem to affect the 120572-amylase productioneven when the stain might be able to grow From this resultit has been understood that amylase productionmay occur atpH 60 but since the final pH was lowered to 45 no amylaseactivity was detected after 24 h of incubation because of theinstability of the enzyme toward acidic conditions Most ofthe earlier studies revealed neutral or alkaline initial pH for120572-amylase production by Pseudomonas species [20 22 24] Onthe other hand Lalucat et al [10] reported that none of thePseudomonas stutzeri strains tolerate acidic conditions andthey do not grow at pH 45
In an attempt to increase the 120572-amylase production by Pstutzeri the effect of some salts including K
2HPO4(14 gL)
KH2PO4(07 gL) NaCl (05 gL) MgSO
4(01 gL) and
CaCl2(01 gL) was investigated Results (Table 4) revealed
that even though the growth rate was lower in medium sup-plemented with phosphate ions (as K
2HPO4and KH
2PO4)
production of 120572-amylase was enhanced as compared to themediumwithout salts addition (none) It is also interesting tonote that while amylase production was slightly affected bythe addition in the culture medium of the monovalent cationNa+ (in the form of NaCl) divalent cations (01 gL) seem toenhance moderately or effectively the amylase production byP stutzeri AS22 in the case of Mg2+ and Ca2+ respectivelyAs mentioned by Gupta et al [18] differences in the min-eral requirements of amylase producing bacteria have beenreported and whereas most of previous studies indicated thataddition of CaCl
2to the fermentationmedium is known to be
essential for amylase production [9 26 27] Ca-independentamylase production has also been reported [28 29] Incomparison with the control medium when CaCl
2(04 gL)
was added individually as a mineral source to the mediumamylase production was enhanced from 59 to 66UmLHowever by increasing the CaCl
2concentration from 08 to
12 gL decline in enzyme production by approximately 30was observed This finding is in accordance with previousstudies showing inhibition of amylase production by theexcess of some minerals such Ca2+ [30] and Mg2+ ions [31]
Maximum amylase production (68UmL) occurredwhen CaCl
2(04 gL) was supplemented to the control
medium containing potato starch 10 gL yeast extract 5 gLMgSO
4(7 H2O) 01 gL K
2HPO414 gL KH
2PO407 gL
and NaCl 05 gL
314 Pattern of 120572-Amylase Production by P stutzeri AS22P stutzeri AS22 was grown aerobically in the optimizedmedium (potato starch 10 gL yeast extract 5 gL MgSO
4(7
H2O) 01 gL K
2HPO414 gL KH
2PO407 gL and NaCl
05 and CaCl204 gL) for 240 h at 30∘C The pattern of 120572-
amylase production and bacterial growth was followed withtime (Figure 2) Amylase production was initiated during theexponential growth phase of the strain reached 675UmLduring the stationary phase at 24 h and remained constanttill 72 h suggesting that the enzyme production is growthassociated and induced by the presence of starch hydrolysisproducts in the mediumThese results are in agreement withthe reports of Nakada et al [20] and Fogarty et al [22] onthe relationship between pattern of cell growth and120572-amylaseproduction
However the level of 120572-amylase increased slightly from72 h to 96 h and thereafter it increasedmore rapidly from 192to 216 h as the bacterial cells approached their death phaseSince the increase of 120572-amylase activity was concomitantwith the cell optical density decrease the amylase activityincrease may be the result of bacterial cell dying and releaseof intracellular or periplasmic 120572-amylases in the culturemedium Therefore attempts to confirm this statementresulted in the localization of 120572-amylase activity after 24 hof incubation It was found that P Stutzeri AS22 produced
6 BioMed Research International
Table 4 Effect of supplementation of the culture medium with various salts on the production of 120572-amylase by P stutzeri AS22
Chemicals Concentration (gL) Final pH value Biomass (OD600
) 120572-Amylase activity (UmL)None 72 477 35Control 78 545 59NaCl 05 732 49 38K2HPO4 14 76 257 519KH2PO4 07 77 201 59
MgSO4
01 708 543 41304 70 635 44208 73 573 324
CaCl2
01 69 715 60104 67 63 6608 65 619 6512 63 62 46
Control + CaCl2 (04 gL) 72 62 68Cultivation was performed for 24 h at 30∘C with shaking at 200 rpm in media that consisted of (gL) potato starch 10 yeast extract 5 and different salts andwere adjusted to pH 80 Control potato starch 10 yeast extract 5 K2HPO4 14 KH2PO4 07 MgSO4 01 and NaCl 05 gL
0
1
2
3
4
5
6
7
0
2
4
6
8
10
12
14
16
0 24 48 72 96 120 144 168 192 216
Cel
l gro
wth
(OD
600
nm)
Am
ylas
e act
ivity
(Um
L)
Time (h)Amylase activity (UmL)Growth (OD 600 nm)
Figure 2 Pattern of growth and 120572-amylase production of P stutzeriAS22 strain Culture was conducted in media consisting of (gL)potato starch 10 yeast extract 5 MgSO
4
01 K2
HPO4
14 KH2
PO4
07 CaCl2
04 gL and NaCl 05 gL Incubation was carried out at30∘C in a rotary shaker with stirring at 200 rpm
a dominant extracellular 120572-amylase as well as intracellularand periplasmic 120572-amylases which constitute approximately10 and 3 of the extracellular fraction respectively
A zymogram prepared using the cell-free culture super-natant at various time intervals (Figure 3) showed only onedominant protein band (PSA) in all the culture supernatantsTwominor amylases were also observed one active form (F1)was detected in 14ndash240 hr culture supernatants and the otherform (F2) was observed in 48ndash240 hr culture supernatantsThe change of PAGE pattern led us to suspect that F1 and F2are either produced from proteolytic degradation of PSA orlater secreted in the extracellular medium However since Pstutzeri AS22 lacks proteolytic activity (Figure 1(b)) and theintensity of the PSA band remains constant from beginningto 240 h this clearly indicates that F1 and F2 are not derived
14 h 24 h 36 h 48 h 72 h 96 h 240 h
PSA
F2F1
120 h 144 h 192 h
Figure 3 Changes in zymogram pattern for P stutzeri 120572-amylasesduring cultivation
fromPSAproteolysis but are later secreted in the extracellularmedium
Multiple forms of G4-amylase from Pseudomonas stutzeristrains have been reported Robyt and Ackerman [19]reported that P stutzeri NRRL B-3389 produced seven G4-amylases which differed in molecular mass and isoelectricpoint Strain MS300 was found to produce two major G4-amylases (amylases A and B) and two minor amylases(amylases C and D) [23] However the G4-amylase of PstutzeriMO-19 has been found to exist in two forms One ofthese is a 57 kDa protein (G4-1) the other is a 46 kDa protein(G4-2) which was derived by the limited proteolysis of G4-l[20]
32 Biochemical Characterization of Crude120572-Amylase Preparation
321 Effect of Temperature on 120572-Amylase Activity and Sta-bility The temperature activity profile shows that the crude120572-amylase was active at temperatures ranging from 20to 70∘C with an optimum activity at 60∘C (Figure 4(a))
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991
BioMed Research International 3
(a) (b)
Figure 1 Amylolytic (a) and proteolytic (b) activities of the isolated strain AS22 The amylolytic and proteolytic activities of the strainAS22 were evaluated by puncture inoculation of the strain inside nutrient agar medium containing soluble starch (1 wv) (a) and casein(b) respectively After incubation at 37∘C for 24 h to detect the amylolytic activity the plates were flooded with iodine solution at roomtemperature
08 gL Amylolytic activity was determined after incubationfor 24 h at 30∘C and 200 rpm agitation speed
For maximal 120572-amylase production fermentation period(0ndash240 h) was also optimized under the optimized fermenta-tion conditions Fermentation was performed at 30∘C in anautomatic incubator and samples were prepared at differentintervals (continuously for 10 days) for enzyme assay
26 Biochemical Properties of the Crude120572-Amylase Preparation
261 Effect of Temperature on Activity and Stability Theeffect of temperature on 120572-amylase activity was studied from20 to 90∘C Thermal stability was examined by incubatingthe enzyme preparation for 60min at different temperaturesranging from 30 to 70∘C Aliquots were withdrawn at desiredtime intervals and the remaining activitywasmeasured underenzyme assay conditions The nonheated enzyme was takenas 100
262 Effect of pH on Activity and Stability The effect of pHon 120572-amylase activity was evaluated for the pH range of 30ndash130 at 60∘C For pH stability measurement the enzyme wasincubated at 30∘C for 1 h in different buffers and the residualactivity was determined under the enzyme assay conditionsThe following buffer systems were used 100mM glycine-HCl buffer pH 30-40 100mM acetate buffer pH 40ndash60100mM Tris-HCl buffer pH 70-80 100mM glycine-NaOHbuffer pH 90ndash110 and 100mMNa
2HPO4-NaOH buffer pH
120 and 130
263 Effect of Metal Ions Surfactants and Enzyme InhibitorsThe influence of various metal ions (5mM) on 120572-amylaseactivity was investigated using CaCl
2 ZnCl
2 FeCl
2 HgCl
2
BaCl2 MnCl
2 MgCl
2 CuCl
2 CdCl
2 NaCl and KCl Activity
in the absence of any additives was taken as 100The effects of some surfactants (Triton X-100 Tween 20
Tween 80 and SDS) and enzyme inhibitors (phenylmethyl-sulfonyl fluoride (PMSF) 120573-mercaptoethanol and ethylene-diaminetetraacetic acid (EDTA)) on 120572-amylase stability werestudied by preincubating the amylolytic preparation for30min at room temperature The remaining enzyme activitywas measured under enzyme assay conditions The activityof enzyme incubated under similar conditions without anyadditive was taken as 100
27 Analysis of End Products Starch Hydrolysis Starch solu-tion (1 wv 50mL) was prepared using Tris-HCl buffer(100mM pH 80) The P stutzeri AS22 crude 120572-amylase(06U) was added into 50mL of starch solution Reactionmixture was incubated at 60∘C Aliquots were withdrawnat various time intervals and boiled for 10min to stopthe reaction and samples were then centrifuged at 13000 gfor 10min and passed through 022 120583m filter Samples thusobtained were analyzed by thin-layer chromatography (TLC)and gel permeation chromatography (GPC) on Bio-Gel P2
TLC was performed on silica gel 60 (20 times 20 cmMerck Germany) with a mobile phase composed of chlo-roformacetic acidwater (60 70 10 vvv) The spots werevisualized by spraying TLC plates withH
2SO4ethanol (5 95
vv) followed by heating at 120∘C for 10minGPC was performed on a Bio-Gel P2 column (15 times
200 cm) eluted with water at a rate of 30mLh The differentoligosaccharides in the range of G1 to G7 were fractionatedand the percentages of different products were determined
28 Visualisation of 120572-Amylase Activity by Zymography 120572-Amylase activity staining was done by layering the SDS-PAGE gel on a thin 2 agarose-1 soluble potato starch gel
4 BioMed Research International
incubated as a sandwich for 60min at 50∘CUpon staining theagarose gel with iodine solution at room temperature proteinbands with amylolytic activity became visible as a white bandagainst a dark blue background
3 Results and Discussion
31 120572-Amylase Production by P stutzeri AS22 The produc-tion of amylases enzymes by microorganisms is significantlyaffected by physical and chemical parameters of the medium[14 15] In this regard appropriate media components andsuitable conditions must be attained for optimal productionof the required products
311 Effects of Different Carbon Sources on 120572-Amylase Produc-tion Because amylase synthesis is known to be induced bystarch or its hydrolytic products [16ndash18] P stutzeri AS22 wasgrown in the basal medium supplemented with starch fromvarious natural sources (potato wheat andmaize starch) andother carbohydrates including lactose maltose and glucosefor assessing their effects on the production of amylase(Table 1)
The highest amylase activity (08UmL) was producedon potato starch followed by maltose (05UmL) Howeveramylase production was significantly low when the strainwas grown on lactose (0055UmL) and glucose (01 UmL)This study falls in line with previous studies on P stutzeriamylase production in which enzyme was induced usingstarch amylodextrin or maltose while glucose was foundto inhibit amylase production [1 19ndash21] In contrast to ourresults glucose was found to be the best carbon source foramylase production by Pseudomonas sp IMD 353 (13UmL)while the amylolytic activity decreased to 2 and 3UmLwhenmaltose and starch were used respectively as sole carbonsources in the same conditions [22]
Since potato starchwas the best carbon source for amylasesynthesis the effect of its concentration (025ndash2) on theamylase production was studied in media containing 01ammonium sulphate as nitrogen source It was observedthat the increase in concentration of potato starch increasesamylase production and maximum activity (075UmL) wasobtained in the presence of 1 substrate (data not shown)However further increase (15 and 2) of potato starchconcentration resulted in rapid decrease of enzyme produc-tion although biomass remained nearly constant (decreasedslightly) This may be explained by the degradation duringthe fermentation of starch by 120572-amylases resulting in theaccumulation of high quantities of reducing sugar which ledto an enhancement of sugar concentration and therefore tocatabolite repression of 120572-amylase synthesis [17]
312 Effects of Various Nitrogen Sources on 120572-Amylase Pro-duction In general both organic and inorganic nitrogensources were used efficiently in the growth medium for thebiosynthesis of 120572-amylase
In the present study various nitrogen sources at a con-centration of 01 were evaluated using an optimum potatostarch concentration of 1 (Table 2) Maximum activity was
Table 1 Effects of different carbon sources on the production of 120572-amylase by P stutzeri AS22
Carbon sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)Lactose 749 01 0055Glucose 569 08 01Maltose 662 10 05Potato starch 652 0 8 08Wheat starch 669 104 01Maize starch 659 05 017Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) carbon source 10 ammonium sulphate 1 K2HPO4 14KH2PO4 07 MgSO4 01 and NaCl 05 and that were inoculated at initial ODof 0016 and adjusted to pH 80
Table 2 Effects of different nitrogen sources supplemented to thepotato starch on the production of 120572-amylase by P stutzeri AS22
Nitrogen sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)NH4Cl 66 094 05(NH4)2SO4 65 09 07Soya peptone 79 321 097Casein 785 161 10Pastone 778 308 15Yeast extract 787 356 25Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 K2HPO4 14 KH2PO4 07MgSO4 01NaCl 05 and different nitrogen sources (1 gL) and were adjusted to pH 80
obtained when yeast extract was used (25UmL) Additionof yeast extract to the medium increased the production of120572-amylase activity by more than three times over mediumwith ammonium sulphate as nitrogen source Compared tothe organic nitrogen sources which support good growthand extracellular 120572-amylase production inorganic nitrogensources like ammonium sulphate and ammonium chlorideare not efficient for amylase productionThese results corrob-orate well with previous studies showing that organic nitro-gen sources were preferred for the 120572-amylase productionThe stimulatory effect of these complex compounds may beattributed to their carbohydrate and protein compositions aswell as to the trace of minerals and ions that could be presentand which enhance the enzyme secretion [18] Yeast extracthas been also used as nitrogen source in the culture mediumfor amylase production alone in the case of P stutzeri NRRLB-3389 [19] or in combination with other nitrogen sourcessuch as polypeptone in the case ofPseudomonas strainMS300[23] and yeatex for Pseudomonas sp IMD 353 [22] Otherorganic nitrogen sources have been also reported to support120572-amylase production by P stutzeriMO-19 such as corn steepliquor and peptone [20] and polypeptone in the case of Psaccharophila IAM 1504 [24]
To investigate the best concentration on 120572-amylase pro-duction yeast extract (range 1ndash10 gL) was added to basal
BioMed Research International 5
Table 3 Effect of different initial pH values of fermentationmediumon the production of 120572-amylase by P stutzeri AS22
Initial pH value Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)60 473 39 070 72 49 3880 80 56 5990 855 55 585100 89 55 58110 922 56 56120 932 54 54Cultivation was performed for 24 h at 30∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 yeast extract 5 K2HPO4 14 KH2PO407 MgSO4 01 and NaCl 05 and were adjusted to different initial pH values
medium containing potato starch (10 gL) Maximum amy-lase production by P stutzeri AS22 was obtained with 5 gLyeast extract reaching 395UmL and further addition ofyeast extract decreased the level of amylase (data not shown)
313 Effects of Temperature Agitation Initial pH Values andSalts on120572-Amylase Production Theeffects of temperature (25to 45∘C) agitation (150 200 and 250 rpm) and initial pH (60to 120) on 120572-amylase production by P stutzeri AS22 werestudied in optimizedmedium containing 10 gL potato starchand 5 gL yeast extract
Optimum level of 120572-amylase production (6UmL) wasachieved at 30∘C (data not shown) This result falls in linewith previous studies on Pseudomonas 120572-amylases in whichmost of them are reported to be produced at 25 [23] and 30∘C[20 24] Amylase production seems to be very sensitive tohigher temperatures Indeed at 37∘C the amylolytic activitywas 40 lower than that at 30∘C and at 45∘C no 120572-amylaseactivity was detected
Among the tested agitation speeds the maximum amy-lase activity (61 UmL) was found at 200 rpm (data notshown)
The pH of the production medium strongly affects manyenzymatic processes and transport of compounds across thecell membrane [18 25] In this study the effect of pH on 120572-amylase production by P stutzeri AS22 was investigated withvarying pHs (60ndash120) The results revealed that the pH ofthe medium influences the 120572-amylase production as well asthe strain growth (Table 3) which were more favorable atalkaline pH 80ndash120 than the neutral one In contrast acidicconditions (pH 60) seem to affect the 120572-amylase productioneven when the stain might be able to grow From this resultit has been understood that amylase productionmay occur atpH 60 but since the final pH was lowered to 45 no amylaseactivity was detected after 24 h of incubation because of theinstability of the enzyme toward acidic conditions Most ofthe earlier studies revealed neutral or alkaline initial pH for120572-amylase production by Pseudomonas species [20 22 24] Onthe other hand Lalucat et al [10] reported that none of thePseudomonas stutzeri strains tolerate acidic conditions andthey do not grow at pH 45
In an attempt to increase the 120572-amylase production by Pstutzeri the effect of some salts including K
2HPO4(14 gL)
KH2PO4(07 gL) NaCl (05 gL) MgSO
4(01 gL) and
CaCl2(01 gL) was investigated Results (Table 4) revealed
that even though the growth rate was lower in medium sup-plemented with phosphate ions (as K
2HPO4and KH
2PO4)
production of 120572-amylase was enhanced as compared to themediumwithout salts addition (none) It is also interesting tonote that while amylase production was slightly affected bythe addition in the culture medium of the monovalent cationNa+ (in the form of NaCl) divalent cations (01 gL) seem toenhance moderately or effectively the amylase production byP stutzeri AS22 in the case of Mg2+ and Ca2+ respectivelyAs mentioned by Gupta et al [18] differences in the min-eral requirements of amylase producing bacteria have beenreported and whereas most of previous studies indicated thataddition of CaCl
2to the fermentationmedium is known to be
essential for amylase production [9 26 27] Ca-independentamylase production has also been reported [28 29] Incomparison with the control medium when CaCl
2(04 gL)
was added individually as a mineral source to the mediumamylase production was enhanced from 59 to 66UmLHowever by increasing the CaCl
2concentration from 08 to
12 gL decline in enzyme production by approximately 30was observed This finding is in accordance with previousstudies showing inhibition of amylase production by theexcess of some minerals such Ca2+ [30] and Mg2+ ions [31]
Maximum amylase production (68UmL) occurredwhen CaCl
2(04 gL) was supplemented to the control
medium containing potato starch 10 gL yeast extract 5 gLMgSO
4(7 H2O) 01 gL K
2HPO414 gL KH
2PO407 gL
and NaCl 05 gL
314 Pattern of 120572-Amylase Production by P stutzeri AS22P stutzeri AS22 was grown aerobically in the optimizedmedium (potato starch 10 gL yeast extract 5 gL MgSO
4(7
H2O) 01 gL K
2HPO414 gL KH
2PO407 gL and NaCl
05 and CaCl204 gL) for 240 h at 30∘C The pattern of 120572-
amylase production and bacterial growth was followed withtime (Figure 2) Amylase production was initiated during theexponential growth phase of the strain reached 675UmLduring the stationary phase at 24 h and remained constanttill 72 h suggesting that the enzyme production is growthassociated and induced by the presence of starch hydrolysisproducts in the mediumThese results are in agreement withthe reports of Nakada et al [20] and Fogarty et al [22] onthe relationship between pattern of cell growth and120572-amylaseproduction
However the level of 120572-amylase increased slightly from72 h to 96 h and thereafter it increasedmore rapidly from 192to 216 h as the bacterial cells approached their death phaseSince the increase of 120572-amylase activity was concomitantwith the cell optical density decrease the amylase activityincrease may be the result of bacterial cell dying and releaseof intracellular or periplasmic 120572-amylases in the culturemedium Therefore attempts to confirm this statementresulted in the localization of 120572-amylase activity after 24 hof incubation It was found that P Stutzeri AS22 produced
6 BioMed Research International
Table 4 Effect of supplementation of the culture medium with various salts on the production of 120572-amylase by P stutzeri AS22
Chemicals Concentration (gL) Final pH value Biomass (OD600
) 120572-Amylase activity (UmL)None 72 477 35Control 78 545 59NaCl 05 732 49 38K2HPO4 14 76 257 519KH2PO4 07 77 201 59
MgSO4
01 708 543 41304 70 635 44208 73 573 324
CaCl2
01 69 715 60104 67 63 6608 65 619 6512 63 62 46
Control + CaCl2 (04 gL) 72 62 68Cultivation was performed for 24 h at 30∘C with shaking at 200 rpm in media that consisted of (gL) potato starch 10 yeast extract 5 and different salts andwere adjusted to pH 80 Control potato starch 10 yeast extract 5 K2HPO4 14 KH2PO4 07 MgSO4 01 and NaCl 05 gL
0
1
2
3
4
5
6
7
0
2
4
6
8
10
12
14
16
0 24 48 72 96 120 144 168 192 216
Cel
l gro
wth
(OD
600
nm)
Am
ylas
e act
ivity
(Um
L)
Time (h)Amylase activity (UmL)Growth (OD 600 nm)
Figure 2 Pattern of growth and 120572-amylase production of P stutzeriAS22 strain Culture was conducted in media consisting of (gL)potato starch 10 yeast extract 5 MgSO
4
01 K2
HPO4
14 KH2
PO4
07 CaCl2
04 gL and NaCl 05 gL Incubation was carried out at30∘C in a rotary shaker with stirring at 200 rpm
a dominant extracellular 120572-amylase as well as intracellularand periplasmic 120572-amylases which constitute approximately10 and 3 of the extracellular fraction respectively
A zymogram prepared using the cell-free culture super-natant at various time intervals (Figure 3) showed only onedominant protein band (PSA) in all the culture supernatantsTwominor amylases were also observed one active form (F1)was detected in 14ndash240 hr culture supernatants and the otherform (F2) was observed in 48ndash240 hr culture supernatantsThe change of PAGE pattern led us to suspect that F1 and F2are either produced from proteolytic degradation of PSA orlater secreted in the extracellular medium However since Pstutzeri AS22 lacks proteolytic activity (Figure 1(b)) and theintensity of the PSA band remains constant from beginningto 240 h this clearly indicates that F1 and F2 are not derived
14 h 24 h 36 h 48 h 72 h 96 h 240 h
PSA
F2F1
120 h 144 h 192 h
Figure 3 Changes in zymogram pattern for P stutzeri 120572-amylasesduring cultivation
fromPSAproteolysis but are later secreted in the extracellularmedium
Multiple forms of G4-amylase from Pseudomonas stutzeristrains have been reported Robyt and Ackerman [19]reported that P stutzeri NRRL B-3389 produced seven G4-amylases which differed in molecular mass and isoelectricpoint Strain MS300 was found to produce two major G4-amylases (amylases A and B) and two minor amylases(amylases C and D) [23] However the G4-amylase of PstutzeriMO-19 has been found to exist in two forms One ofthese is a 57 kDa protein (G4-1) the other is a 46 kDa protein(G4-2) which was derived by the limited proteolysis of G4-l[20]
32 Biochemical Characterization of Crude120572-Amylase Preparation
321 Effect of Temperature on 120572-Amylase Activity and Sta-bility The temperature activity profile shows that the crude120572-amylase was active at temperatures ranging from 20to 70∘C with an optimum activity at 60∘C (Figure 4(a))
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991
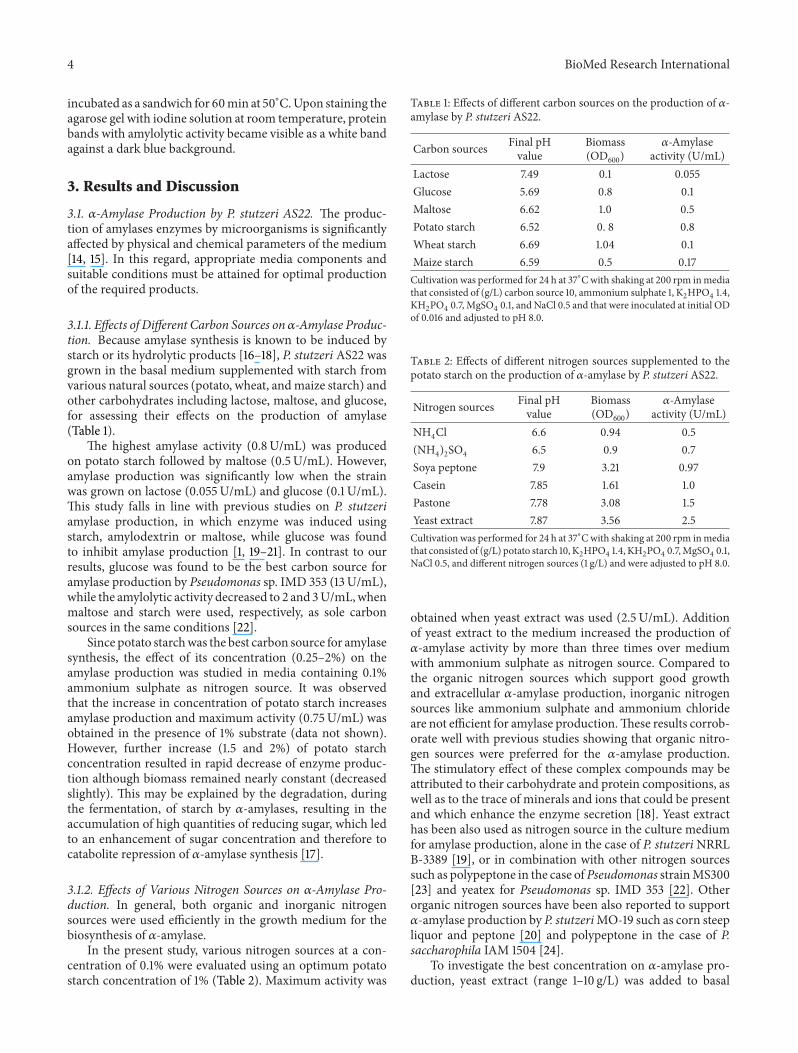
4 BioMed Research International
incubated as a sandwich for 60min at 50∘CUpon staining theagarose gel with iodine solution at room temperature proteinbands with amylolytic activity became visible as a white bandagainst a dark blue background
3 Results and Discussion
31 120572-Amylase Production by P stutzeri AS22 The produc-tion of amylases enzymes by microorganisms is significantlyaffected by physical and chemical parameters of the medium[14 15] In this regard appropriate media components andsuitable conditions must be attained for optimal productionof the required products
311 Effects of Different Carbon Sources on 120572-Amylase Produc-tion Because amylase synthesis is known to be induced bystarch or its hydrolytic products [16ndash18] P stutzeri AS22 wasgrown in the basal medium supplemented with starch fromvarious natural sources (potato wheat andmaize starch) andother carbohydrates including lactose maltose and glucosefor assessing their effects on the production of amylase(Table 1)
The highest amylase activity (08UmL) was producedon potato starch followed by maltose (05UmL) Howeveramylase production was significantly low when the strainwas grown on lactose (0055UmL) and glucose (01 UmL)This study falls in line with previous studies on P stutzeriamylase production in which enzyme was induced usingstarch amylodextrin or maltose while glucose was foundto inhibit amylase production [1 19ndash21] In contrast to ourresults glucose was found to be the best carbon source foramylase production by Pseudomonas sp IMD 353 (13UmL)while the amylolytic activity decreased to 2 and 3UmLwhenmaltose and starch were used respectively as sole carbonsources in the same conditions [22]
Since potato starchwas the best carbon source for amylasesynthesis the effect of its concentration (025ndash2) on theamylase production was studied in media containing 01ammonium sulphate as nitrogen source It was observedthat the increase in concentration of potato starch increasesamylase production and maximum activity (075UmL) wasobtained in the presence of 1 substrate (data not shown)However further increase (15 and 2) of potato starchconcentration resulted in rapid decrease of enzyme produc-tion although biomass remained nearly constant (decreasedslightly) This may be explained by the degradation duringthe fermentation of starch by 120572-amylases resulting in theaccumulation of high quantities of reducing sugar which ledto an enhancement of sugar concentration and therefore tocatabolite repression of 120572-amylase synthesis [17]
312 Effects of Various Nitrogen Sources on 120572-Amylase Pro-duction In general both organic and inorganic nitrogensources were used efficiently in the growth medium for thebiosynthesis of 120572-amylase
In the present study various nitrogen sources at a con-centration of 01 were evaluated using an optimum potatostarch concentration of 1 (Table 2) Maximum activity was
Table 1 Effects of different carbon sources on the production of 120572-amylase by P stutzeri AS22
Carbon sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)Lactose 749 01 0055Glucose 569 08 01Maltose 662 10 05Potato starch 652 0 8 08Wheat starch 669 104 01Maize starch 659 05 017Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) carbon source 10 ammonium sulphate 1 K2HPO4 14KH2PO4 07 MgSO4 01 and NaCl 05 and that were inoculated at initial ODof 0016 and adjusted to pH 80
Table 2 Effects of different nitrogen sources supplemented to thepotato starch on the production of 120572-amylase by P stutzeri AS22
Nitrogen sources Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)NH4Cl 66 094 05(NH4)2SO4 65 09 07Soya peptone 79 321 097Casein 785 161 10Pastone 778 308 15Yeast extract 787 356 25Cultivation was performed for 24 h at 37∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 K2HPO4 14 KH2PO4 07MgSO4 01NaCl 05 and different nitrogen sources (1 gL) and were adjusted to pH 80
obtained when yeast extract was used (25UmL) Additionof yeast extract to the medium increased the production of120572-amylase activity by more than three times over mediumwith ammonium sulphate as nitrogen source Compared tothe organic nitrogen sources which support good growthand extracellular 120572-amylase production inorganic nitrogensources like ammonium sulphate and ammonium chlorideare not efficient for amylase productionThese results corrob-orate well with previous studies showing that organic nitro-gen sources were preferred for the 120572-amylase productionThe stimulatory effect of these complex compounds may beattributed to their carbohydrate and protein compositions aswell as to the trace of minerals and ions that could be presentand which enhance the enzyme secretion [18] Yeast extracthas been also used as nitrogen source in the culture mediumfor amylase production alone in the case of P stutzeri NRRLB-3389 [19] or in combination with other nitrogen sourcessuch as polypeptone in the case ofPseudomonas strainMS300[23] and yeatex for Pseudomonas sp IMD 353 [22] Otherorganic nitrogen sources have been also reported to support120572-amylase production by P stutzeriMO-19 such as corn steepliquor and peptone [20] and polypeptone in the case of Psaccharophila IAM 1504 [24]
To investigate the best concentration on 120572-amylase pro-duction yeast extract (range 1ndash10 gL) was added to basal
BioMed Research International 5
Table 3 Effect of different initial pH values of fermentationmediumon the production of 120572-amylase by P stutzeri AS22
Initial pH value Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)60 473 39 070 72 49 3880 80 56 5990 855 55 585100 89 55 58110 922 56 56120 932 54 54Cultivation was performed for 24 h at 30∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 yeast extract 5 K2HPO4 14 KH2PO407 MgSO4 01 and NaCl 05 and were adjusted to different initial pH values
medium containing potato starch (10 gL) Maximum amy-lase production by P stutzeri AS22 was obtained with 5 gLyeast extract reaching 395UmL and further addition ofyeast extract decreased the level of amylase (data not shown)
313 Effects of Temperature Agitation Initial pH Values andSalts on120572-Amylase Production Theeffects of temperature (25to 45∘C) agitation (150 200 and 250 rpm) and initial pH (60to 120) on 120572-amylase production by P stutzeri AS22 werestudied in optimizedmedium containing 10 gL potato starchand 5 gL yeast extract
Optimum level of 120572-amylase production (6UmL) wasachieved at 30∘C (data not shown) This result falls in linewith previous studies on Pseudomonas 120572-amylases in whichmost of them are reported to be produced at 25 [23] and 30∘C[20 24] Amylase production seems to be very sensitive tohigher temperatures Indeed at 37∘C the amylolytic activitywas 40 lower than that at 30∘C and at 45∘C no 120572-amylaseactivity was detected
Among the tested agitation speeds the maximum amy-lase activity (61 UmL) was found at 200 rpm (data notshown)
The pH of the production medium strongly affects manyenzymatic processes and transport of compounds across thecell membrane [18 25] In this study the effect of pH on 120572-amylase production by P stutzeri AS22 was investigated withvarying pHs (60ndash120) The results revealed that the pH ofthe medium influences the 120572-amylase production as well asthe strain growth (Table 3) which were more favorable atalkaline pH 80ndash120 than the neutral one In contrast acidicconditions (pH 60) seem to affect the 120572-amylase productioneven when the stain might be able to grow From this resultit has been understood that amylase productionmay occur atpH 60 but since the final pH was lowered to 45 no amylaseactivity was detected after 24 h of incubation because of theinstability of the enzyme toward acidic conditions Most ofthe earlier studies revealed neutral or alkaline initial pH for120572-amylase production by Pseudomonas species [20 22 24] Onthe other hand Lalucat et al [10] reported that none of thePseudomonas stutzeri strains tolerate acidic conditions andthey do not grow at pH 45
In an attempt to increase the 120572-amylase production by Pstutzeri the effect of some salts including K
2HPO4(14 gL)
KH2PO4(07 gL) NaCl (05 gL) MgSO
4(01 gL) and
CaCl2(01 gL) was investigated Results (Table 4) revealed
that even though the growth rate was lower in medium sup-plemented with phosphate ions (as K
2HPO4and KH
2PO4)
production of 120572-amylase was enhanced as compared to themediumwithout salts addition (none) It is also interesting tonote that while amylase production was slightly affected bythe addition in the culture medium of the monovalent cationNa+ (in the form of NaCl) divalent cations (01 gL) seem toenhance moderately or effectively the amylase production byP stutzeri AS22 in the case of Mg2+ and Ca2+ respectivelyAs mentioned by Gupta et al [18] differences in the min-eral requirements of amylase producing bacteria have beenreported and whereas most of previous studies indicated thataddition of CaCl
2to the fermentationmedium is known to be
essential for amylase production [9 26 27] Ca-independentamylase production has also been reported [28 29] Incomparison with the control medium when CaCl
2(04 gL)
was added individually as a mineral source to the mediumamylase production was enhanced from 59 to 66UmLHowever by increasing the CaCl
2concentration from 08 to
12 gL decline in enzyme production by approximately 30was observed This finding is in accordance with previousstudies showing inhibition of amylase production by theexcess of some minerals such Ca2+ [30] and Mg2+ ions [31]
Maximum amylase production (68UmL) occurredwhen CaCl
2(04 gL) was supplemented to the control
medium containing potato starch 10 gL yeast extract 5 gLMgSO
4(7 H2O) 01 gL K
2HPO414 gL KH
2PO407 gL
and NaCl 05 gL
314 Pattern of 120572-Amylase Production by P stutzeri AS22P stutzeri AS22 was grown aerobically in the optimizedmedium (potato starch 10 gL yeast extract 5 gL MgSO
4(7
H2O) 01 gL K
2HPO414 gL KH
2PO407 gL and NaCl
05 and CaCl204 gL) for 240 h at 30∘C The pattern of 120572-
amylase production and bacterial growth was followed withtime (Figure 2) Amylase production was initiated during theexponential growth phase of the strain reached 675UmLduring the stationary phase at 24 h and remained constanttill 72 h suggesting that the enzyme production is growthassociated and induced by the presence of starch hydrolysisproducts in the mediumThese results are in agreement withthe reports of Nakada et al [20] and Fogarty et al [22] onthe relationship between pattern of cell growth and120572-amylaseproduction
However the level of 120572-amylase increased slightly from72 h to 96 h and thereafter it increasedmore rapidly from 192to 216 h as the bacterial cells approached their death phaseSince the increase of 120572-amylase activity was concomitantwith the cell optical density decrease the amylase activityincrease may be the result of bacterial cell dying and releaseof intracellular or periplasmic 120572-amylases in the culturemedium Therefore attempts to confirm this statementresulted in the localization of 120572-amylase activity after 24 hof incubation It was found that P Stutzeri AS22 produced
6 BioMed Research International
Table 4 Effect of supplementation of the culture medium with various salts on the production of 120572-amylase by P stutzeri AS22
Chemicals Concentration (gL) Final pH value Biomass (OD600
) 120572-Amylase activity (UmL)None 72 477 35Control 78 545 59NaCl 05 732 49 38K2HPO4 14 76 257 519KH2PO4 07 77 201 59
MgSO4
01 708 543 41304 70 635 44208 73 573 324
CaCl2
01 69 715 60104 67 63 6608 65 619 6512 63 62 46
Control + CaCl2 (04 gL) 72 62 68Cultivation was performed for 24 h at 30∘C with shaking at 200 rpm in media that consisted of (gL) potato starch 10 yeast extract 5 and different salts andwere adjusted to pH 80 Control potato starch 10 yeast extract 5 K2HPO4 14 KH2PO4 07 MgSO4 01 and NaCl 05 gL
0
1
2
3
4
5
6
7
0
2
4
6
8
10
12
14
16
0 24 48 72 96 120 144 168 192 216
Cel
l gro
wth
(OD
600
nm)
Am
ylas
e act
ivity
(Um
L)
Time (h)Amylase activity (UmL)Growth (OD 600 nm)
Figure 2 Pattern of growth and 120572-amylase production of P stutzeriAS22 strain Culture was conducted in media consisting of (gL)potato starch 10 yeast extract 5 MgSO
4
01 K2
HPO4
14 KH2
PO4
07 CaCl2
04 gL and NaCl 05 gL Incubation was carried out at30∘C in a rotary shaker with stirring at 200 rpm
a dominant extracellular 120572-amylase as well as intracellularand periplasmic 120572-amylases which constitute approximately10 and 3 of the extracellular fraction respectively
A zymogram prepared using the cell-free culture super-natant at various time intervals (Figure 3) showed only onedominant protein band (PSA) in all the culture supernatantsTwominor amylases were also observed one active form (F1)was detected in 14ndash240 hr culture supernatants and the otherform (F2) was observed in 48ndash240 hr culture supernatantsThe change of PAGE pattern led us to suspect that F1 and F2are either produced from proteolytic degradation of PSA orlater secreted in the extracellular medium However since Pstutzeri AS22 lacks proteolytic activity (Figure 1(b)) and theintensity of the PSA band remains constant from beginningto 240 h this clearly indicates that F1 and F2 are not derived
14 h 24 h 36 h 48 h 72 h 96 h 240 h
PSA
F2F1
120 h 144 h 192 h
Figure 3 Changes in zymogram pattern for P stutzeri 120572-amylasesduring cultivation
fromPSAproteolysis but are later secreted in the extracellularmedium
Multiple forms of G4-amylase from Pseudomonas stutzeristrains have been reported Robyt and Ackerman [19]reported that P stutzeri NRRL B-3389 produced seven G4-amylases which differed in molecular mass and isoelectricpoint Strain MS300 was found to produce two major G4-amylases (amylases A and B) and two minor amylases(amylases C and D) [23] However the G4-amylase of PstutzeriMO-19 has been found to exist in two forms One ofthese is a 57 kDa protein (G4-1) the other is a 46 kDa protein(G4-2) which was derived by the limited proteolysis of G4-l[20]
32 Biochemical Characterization of Crude120572-Amylase Preparation
321 Effect of Temperature on 120572-Amylase Activity and Sta-bility The temperature activity profile shows that the crude120572-amylase was active at temperatures ranging from 20to 70∘C with an optimum activity at 60∘C (Figure 4(a))
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991

BioMed Research International 5
Table 3 Effect of different initial pH values of fermentationmediumon the production of 120572-amylase by P stutzeri AS22
Initial pH value Final pHvalue
Biomass(OD600
)120572-Amylase
activity (UmL)60 473 39 070 72 49 3880 80 56 5990 855 55 585100 89 55 58110 922 56 56120 932 54 54Cultivation was performed for 24 h at 30∘Cwith shaking at 200 rpm inmediathat consisted of (gL) potato starch 10 yeast extract 5 K2HPO4 14 KH2PO407 MgSO4 01 and NaCl 05 and were adjusted to different initial pH values
medium containing potato starch (10 gL) Maximum amy-lase production by P stutzeri AS22 was obtained with 5 gLyeast extract reaching 395UmL and further addition ofyeast extract decreased the level of amylase (data not shown)
313 Effects of Temperature Agitation Initial pH Values andSalts on120572-Amylase Production Theeffects of temperature (25to 45∘C) agitation (150 200 and 250 rpm) and initial pH (60to 120) on 120572-amylase production by P stutzeri AS22 werestudied in optimizedmedium containing 10 gL potato starchand 5 gL yeast extract
Optimum level of 120572-amylase production (6UmL) wasachieved at 30∘C (data not shown) This result falls in linewith previous studies on Pseudomonas 120572-amylases in whichmost of them are reported to be produced at 25 [23] and 30∘C[20 24] Amylase production seems to be very sensitive tohigher temperatures Indeed at 37∘C the amylolytic activitywas 40 lower than that at 30∘C and at 45∘C no 120572-amylaseactivity was detected
Among the tested agitation speeds the maximum amy-lase activity (61 UmL) was found at 200 rpm (data notshown)
The pH of the production medium strongly affects manyenzymatic processes and transport of compounds across thecell membrane [18 25] In this study the effect of pH on 120572-amylase production by P stutzeri AS22 was investigated withvarying pHs (60ndash120) The results revealed that the pH ofthe medium influences the 120572-amylase production as well asthe strain growth (Table 3) which were more favorable atalkaline pH 80ndash120 than the neutral one In contrast acidicconditions (pH 60) seem to affect the 120572-amylase productioneven when the stain might be able to grow From this resultit has been understood that amylase productionmay occur atpH 60 but since the final pH was lowered to 45 no amylaseactivity was detected after 24 h of incubation because of theinstability of the enzyme toward acidic conditions Most ofthe earlier studies revealed neutral or alkaline initial pH for120572-amylase production by Pseudomonas species [20 22 24] Onthe other hand Lalucat et al [10] reported that none of thePseudomonas stutzeri strains tolerate acidic conditions andthey do not grow at pH 45
In an attempt to increase the 120572-amylase production by Pstutzeri the effect of some salts including K
2HPO4(14 gL)
KH2PO4(07 gL) NaCl (05 gL) MgSO
4(01 gL) and
CaCl2(01 gL) was investigated Results (Table 4) revealed
that even though the growth rate was lower in medium sup-plemented with phosphate ions (as K
2HPO4and KH
2PO4)
production of 120572-amylase was enhanced as compared to themediumwithout salts addition (none) It is also interesting tonote that while amylase production was slightly affected bythe addition in the culture medium of the monovalent cationNa+ (in the form of NaCl) divalent cations (01 gL) seem toenhance moderately or effectively the amylase production byP stutzeri AS22 in the case of Mg2+ and Ca2+ respectivelyAs mentioned by Gupta et al [18] differences in the min-eral requirements of amylase producing bacteria have beenreported and whereas most of previous studies indicated thataddition of CaCl
2to the fermentationmedium is known to be
essential for amylase production [9 26 27] Ca-independentamylase production has also been reported [28 29] Incomparison with the control medium when CaCl
2(04 gL)
was added individually as a mineral source to the mediumamylase production was enhanced from 59 to 66UmLHowever by increasing the CaCl
2concentration from 08 to
12 gL decline in enzyme production by approximately 30was observed This finding is in accordance with previousstudies showing inhibition of amylase production by theexcess of some minerals such Ca2+ [30] and Mg2+ ions [31]
Maximum amylase production (68UmL) occurredwhen CaCl
2(04 gL) was supplemented to the control
medium containing potato starch 10 gL yeast extract 5 gLMgSO
4(7 H2O) 01 gL K
2HPO414 gL KH
2PO407 gL
and NaCl 05 gL
314 Pattern of 120572-Amylase Production by P stutzeri AS22P stutzeri AS22 was grown aerobically in the optimizedmedium (potato starch 10 gL yeast extract 5 gL MgSO
4(7
H2O) 01 gL K
2HPO414 gL KH
2PO407 gL and NaCl
05 and CaCl204 gL) for 240 h at 30∘C The pattern of 120572-
amylase production and bacterial growth was followed withtime (Figure 2) Amylase production was initiated during theexponential growth phase of the strain reached 675UmLduring the stationary phase at 24 h and remained constanttill 72 h suggesting that the enzyme production is growthassociated and induced by the presence of starch hydrolysisproducts in the mediumThese results are in agreement withthe reports of Nakada et al [20] and Fogarty et al [22] onthe relationship between pattern of cell growth and120572-amylaseproduction
However the level of 120572-amylase increased slightly from72 h to 96 h and thereafter it increasedmore rapidly from 192to 216 h as the bacterial cells approached their death phaseSince the increase of 120572-amylase activity was concomitantwith the cell optical density decrease the amylase activityincrease may be the result of bacterial cell dying and releaseof intracellular or periplasmic 120572-amylases in the culturemedium Therefore attempts to confirm this statementresulted in the localization of 120572-amylase activity after 24 hof incubation It was found that P Stutzeri AS22 produced
6 BioMed Research International
Table 4 Effect of supplementation of the culture medium with various salts on the production of 120572-amylase by P stutzeri AS22
Chemicals Concentration (gL) Final pH value Biomass (OD600
) 120572-Amylase activity (UmL)None 72 477 35Control 78 545 59NaCl 05 732 49 38K2HPO4 14 76 257 519KH2PO4 07 77 201 59
MgSO4
01 708 543 41304 70 635 44208 73 573 324
CaCl2
01 69 715 60104 67 63 6608 65 619 6512 63 62 46
Control + CaCl2 (04 gL) 72 62 68Cultivation was performed for 24 h at 30∘C with shaking at 200 rpm in media that consisted of (gL) potato starch 10 yeast extract 5 and different salts andwere adjusted to pH 80 Control potato starch 10 yeast extract 5 K2HPO4 14 KH2PO4 07 MgSO4 01 and NaCl 05 gL
0
1
2
3
4
5
6
7
0
2
4
6
8
10
12
14
16
0 24 48 72 96 120 144 168 192 216
Cel
l gro
wth
(OD
600
nm)
Am
ylas
e act
ivity
(Um
L)
Time (h)Amylase activity (UmL)Growth (OD 600 nm)
Figure 2 Pattern of growth and 120572-amylase production of P stutzeriAS22 strain Culture was conducted in media consisting of (gL)potato starch 10 yeast extract 5 MgSO
4
01 K2
HPO4
14 KH2
PO4
07 CaCl2
04 gL and NaCl 05 gL Incubation was carried out at30∘C in a rotary shaker with stirring at 200 rpm
a dominant extracellular 120572-amylase as well as intracellularand periplasmic 120572-amylases which constitute approximately10 and 3 of the extracellular fraction respectively
A zymogram prepared using the cell-free culture super-natant at various time intervals (Figure 3) showed only onedominant protein band (PSA) in all the culture supernatantsTwominor amylases were also observed one active form (F1)was detected in 14ndash240 hr culture supernatants and the otherform (F2) was observed in 48ndash240 hr culture supernatantsThe change of PAGE pattern led us to suspect that F1 and F2are either produced from proteolytic degradation of PSA orlater secreted in the extracellular medium However since Pstutzeri AS22 lacks proteolytic activity (Figure 1(b)) and theintensity of the PSA band remains constant from beginningto 240 h this clearly indicates that F1 and F2 are not derived
14 h 24 h 36 h 48 h 72 h 96 h 240 h
PSA
F2F1
120 h 144 h 192 h
Figure 3 Changes in zymogram pattern for P stutzeri 120572-amylasesduring cultivation
fromPSAproteolysis but are later secreted in the extracellularmedium
Multiple forms of G4-amylase from Pseudomonas stutzeristrains have been reported Robyt and Ackerman [19]reported that P stutzeri NRRL B-3389 produced seven G4-amylases which differed in molecular mass and isoelectricpoint Strain MS300 was found to produce two major G4-amylases (amylases A and B) and two minor amylases(amylases C and D) [23] However the G4-amylase of PstutzeriMO-19 has been found to exist in two forms One ofthese is a 57 kDa protein (G4-1) the other is a 46 kDa protein(G4-2) which was derived by the limited proteolysis of G4-l[20]
32 Biochemical Characterization of Crude120572-Amylase Preparation
321 Effect of Temperature on 120572-Amylase Activity and Sta-bility The temperature activity profile shows that the crude120572-amylase was active at temperatures ranging from 20to 70∘C with an optimum activity at 60∘C (Figure 4(a))
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991
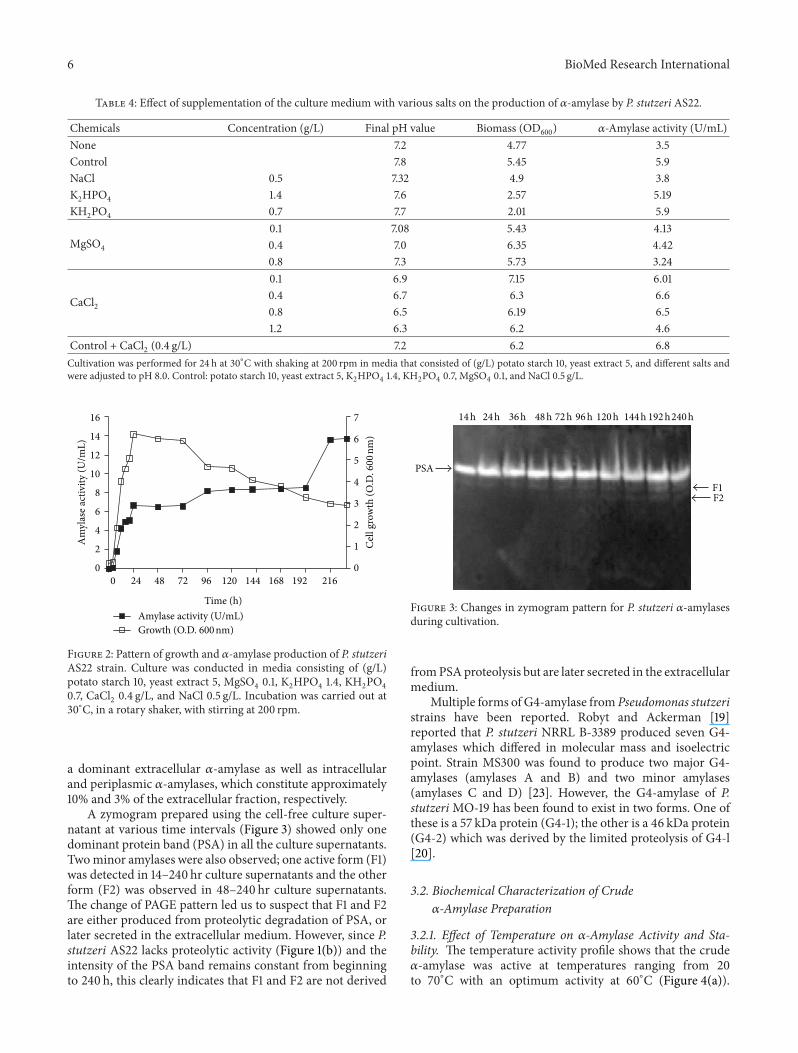
6 BioMed Research International
Table 4 Effect of supplementation of the culture medium with various salts on the production of 120572-amylase by P stutzeri AS22
Chemicals Concentration (gL) Final pH value Biomass (OD600
) 120572-Amylase activity (UmL)None 72 477 35Control 78 545 59NaCl 05 732 49 38K2HPO4 14 76 257 519KH2PO4 07 77 201 59
MgSO4
01 708 543 41304 70 635 44208 73 573 324
CaCl2
01 69 715 60104 67 63 6608 65 619 6512 63 62 46
Control + CaCl2 (04 gL) 72 62 68Cultivation was performed for 24 h at 30∘C with shaking at 200 rpm in media that consisted of (gL) potato starch 10 yeast extract 5 and different salts andwere adjusted to pH 80 Control potato starch 10 yeast extract 5 K2HPO4 14 KH2PO4 07 MgSO4 01 and NaCl 05 gL
0
1
2
3
4
5
6
7
0
2
4
6
8
10
12
14
16
0 24 48 72 96 120 144 168 192 216
Cel
l gro
wth
(OD
600
nm)
Am
ylas
e act
ivity
(Um
L)
Time (h)Amylase activity (UmL)Growth (OD 600 nm)
Figure 2 Pattern of growth and 120572-amylase production of P stutzeriAS22 strain Culture was conducted in media consisting of (gL)potato starch 10 yeast extract 5 MgSO
4
01 K2
HPO4
14 KH2
PO4
07 CaCl2
04 gL and NaCl 05 gL Incubation was carried out at30∘C in a rotary shaker with stirring at 200 rpm
a dominant extracellular 120572-amylase as well as intracellularand periplasmic 120572-amylases which constitute approximately10 and 3 of the extracellular fraction respectively
A zymogram prepared using the cell-free culture super-natant at various time intervals (Figure 3) showed only onedominant protein band (PSA) in all the culture supernatantsTwominor amylases were also observed one active form (F1)was detected in 14ndash240 hr culture supernatants and the otherform (F2) was observed in 48ndash240 hr culture supernatantsThe change of PAGE pattern led us to suspect that F1 and F2are either produced from proteolytic degradation of PSA orlater secreted in the extracellular medium However since Pstutzeri AS22 lacks proteolytic activity (Figure 1(b)) and theintensity of the PSA band remains constant from beginningto 240 h this clearly indicates that F1 and F2 are not derived
14 h 24 h 36 h 48 h 72 h 96 h 240 h
PSA
F2F1
120 h 144 h 192 h
Figure 3 Changes in zymogram pattern for P stutzeri 120572-amylasesduring cultivation
fromPSAproteolysis but are later secreted in the extracellularmedium
Multiple forms of G4-amylase from Pseudomonas stutzeristrains have been reported Robyt and Ackerman [19]reported that P stutzeri NRRL B-3389 produced seven G4-amylases which differed in molecular mass and isoelectricpoint Strain MS300 was found to produce two major G4-amylases (amylases A and B) and two minor amylases(amylases C and D) [23] However the G4-amylase of PstutzeriMO-19 has been found to exist in two forms One ofthese is a 57 kDa protein (G4-1) the other is a 46 kDa protein(G4-2) which was derived by the limited proteolysis of G4-l[20]
32 Biochemical Characterization of Crude120572-Amylase Preparation
321 Effect of Temperature on 120572-Amylase Activity and Sta-bility The temperature activity profile shows that the crude120572-amylase was active at temperatures ranging from 20to 70∘C with an optimum activity at 60∘C (Figure 4(a))
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991
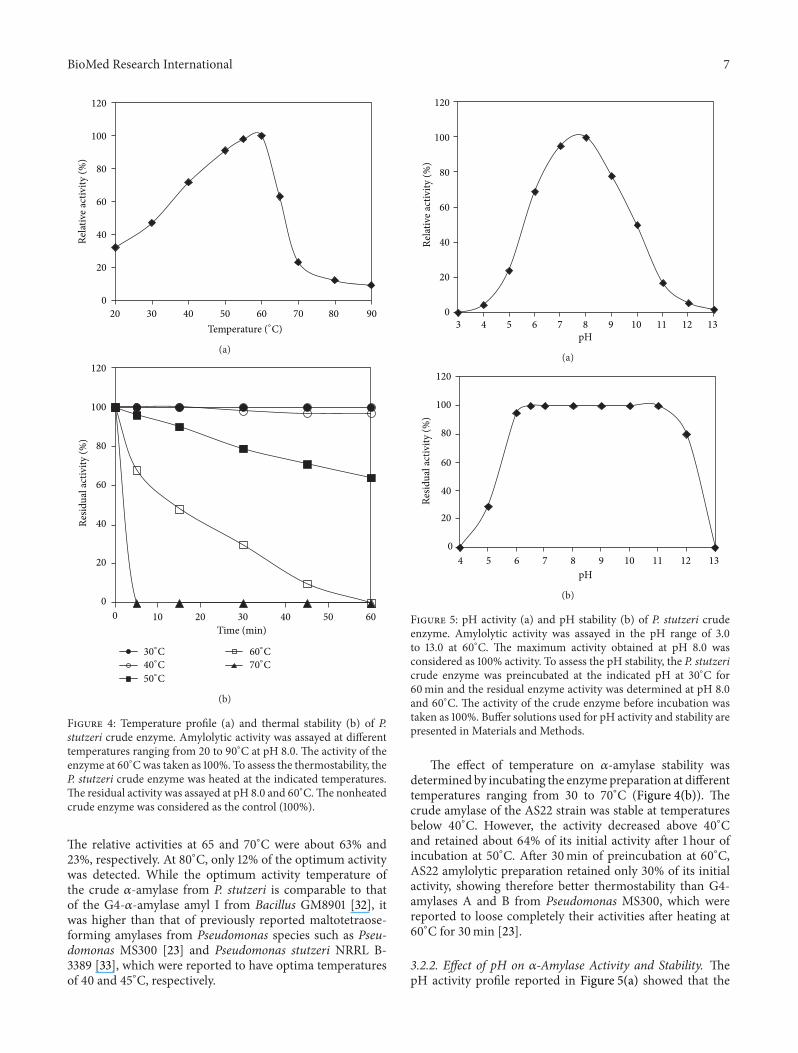
BioMed Research International 7
0
20
40
60
80
100
120
20 30 40 50 60 70 80 90
Rela
tive a
ctiv
ity (
)
Temperature (∘C)
(a)
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Resid
ual a
ctiv
ity (
)
Time (min)
30∘C
40∘C
60∘C
70∘C
50∘C
(b)
Figure 4 Temperature profile (a) and thermal stability (b) of Pstutzeri crude enzyme Amylolytic activity was assayed at differenttemperatures ranging from 20 to 90∘C at pH 80 The activity of theenzyme at 60∘Cwas taken as 100 To assess the thermostability theP stutzeri crude enzyme was heated at the indicated temperaturesThe residual activity was assayed at pH 80 and 60∘CThe nonheatedcrude enzyme was considered as the control (100)
The relative activities at 65 and 70∘C were about 63 and23 respectively At 80∘C only 12 of the optimum activitywas detected While the optimum activity temperature ofthe crude 120572-amylase from P stutzeri is comparable to thatof the G4-120572-amylase amyl I from Bacillus GM8901 [32] itwas higher than that of previously reported maltotetraose-forming amylases from Pseudomonas species such as Pseu-domonas MS300 [23] and Pseudomonas stutzeri NRRL B-3389 [33] which were reported to have optima temperaturesof 40 and 45∘C respectively
0
20
40
60
80
100
120
3 4 5 6 7 8 9 10 11 12 13
Rela
tive a
ctiv
ity (
)
pH
(a)
0
20
40
60
80
100
120
4 5 6 7 8 9 10 11 12 13
Resid
ual a
ctiv
ity (
)
pH
(b)
Figure 5 pH activity (a) and pH stability (b) of P stutzeri crudeenzyme Amylolytic activity was assayed in the pH range of 30to 130 at 60∘C The maximum activity obtained at pH 80 wasconsidered as 100 activity To assess the pH stability the P stutzericrude enzyme was preincubated at the indicated pH at 30∘C for60min and the residual enzyme activity was determined at pH 80and 60∘C The activity of the crude enzyme before incubation wastaken as 100 Buffer solutions used for pH activity and stability arepresented in Materials and Methods
The effect of temperature on 120572-amylase stability wasdetermined by incubating the enzymepreparation at differenttemperatures ranging from 30 to 70∘C (Figure 4(b)) Thecrude amylase of the AS22 strain was stable at temperaturesbelow 40∘C However the activity decreased above 40∘Cand retained about 64 of its initial activity after 1 hour ofincubation at 50∘C After 30min of preincubation at 60∘CAS22 amylolytic preparation retained only 30 of its initialactivity showing therefore better thermostability than G4-amylases A and B from Pseudomonas MS300 which werereported to loose completely their activities after heating at60∘C for 30min [23]
322 Effect of pH on 120572-Amylase Activity and Stability ThepH activity profile reported in Figure 5(a) showed that the
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991
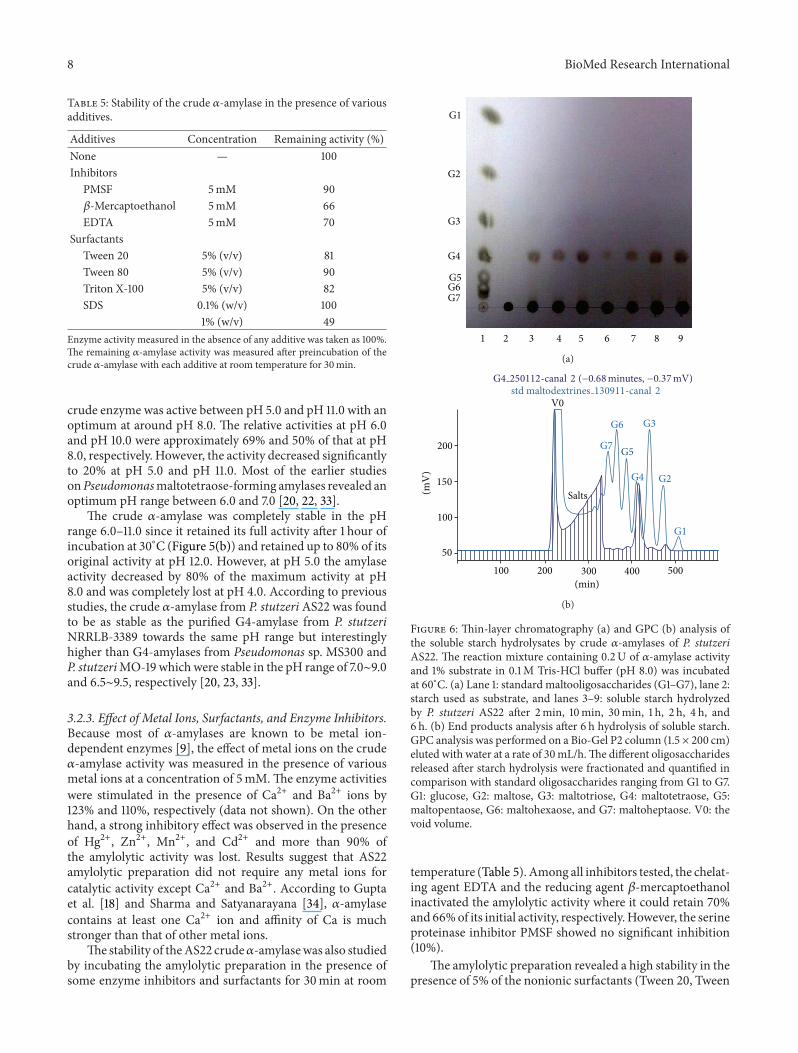
8 BioMed Research International
Table 5 Stability of the crude 120572-amylase in the presence of variousadditives
Additives Concentration Remaining activity ()None mdash 100Inhibitors
PMSF 5mM 90120573-Mercaptoethanol 5mM 66EDTA 5mM 70
SurfactantsTween 20 5 (vv) 81Tween 80 5 (vv) 90Triton X-100 5 (vv) 82SDS 01 (wv) 100
1 (wv) 49Enzyme activity measured in the absence of any additive was taken as 100The remaining 120572-amylase activity was measured after preincubation of thecrude 120572-amylase with each additive at room temperature for 30min
crude enzyme was active between pH 50 and pH 110 with anoptimum at around pH 80 The relative activities at pH 60and pH 100 were approximately 69 and 50 of that at pH80 respectively However the activity decreased significantlyto 20 at pH 50 and pH 110 Most of the earlier studiesonPseudomonasmaltotetraose-forming amylases revealed anoptimum pH range between 60 and 70 [20 22 33]
The crude 120572-amylase was completely stable in the pHrange 60ndash110 since it retained its full activity after 1 hour ofincubation at 30∘C (Figure 5(b)) and retained up to 80 of itsoriginal activity at pH 120 However at pH 50 the amylaseactivity decreased by 80 of the maximum activity at pH80 and was completely lost at pH 40 According to previousstudies the crude 120572-amylase from P stutzeri AS22 was foundto be as stable as the purified G4-amylase from P stutzeriNRRLB-3389 towards the same pH range but interestinglyhigher than G4-amylases from Pseudomonas sp MS300 andP stutzeriMO-19 which were stable in the pH range of 70sim90and 65sim95 respectively [20 23 33]
323 Effect of Metal Ions Surfactants and Enzyme InhibitorsBecause most of 120572-amylases are known to be metal ion-dependent enzymes [9] the effect of metal ions on the crude120572-amylase activity was measured in the presence of variousmetal ions at a concentration of 5mMThe enzyme activitieswere stimulated in the presence of Ca2+ and Ba2+ ions by123 and 110 respectively (data not shown) On the otherhand a strong inhibitory effect was observed in the presenceof Hg2+ Zn2+ Mn2+ and Cd2+ and more than 90 ofthe amylolytic activity was lost Results suggest that AS22amylolytic preparation did not require any metal ions forcatalytic activity except Ca2+ and Ba2+ According to Guptaet al [18] and Sharma and Satyanarayana [34] 120572-amylasecontains at least one Ca2+ ion and affinity of Ca is muchstronger than that of other metal ions
The stability of theAS22 crude120572-amylasewas also studiedby incubating the amylolytic preparation in the presence ofsome enzyme inhibitors and surfactants for 30min at room
G1
G2
G3
G4
G5G6G7
1 2 3 4 5 6 7 8 9
(a)
Salts
V0
G1
G2
G5
G6
G7
G4
G3
100 200 300 400 500(min)
50
100
150
200
(mV
)
G4 250112-canal 2 (minus068minutes minus037mV)std maltodextrines 130911-canal 2
(b)
Figure 6 Thin-layer chromatography (a) and GPC (b) analysis ofthe soluble starch hydrolysates by crude 120572-amylases of P stutzeriAS22 The reaction mixture containing 02U of 120572-amylase activityand 1 substrate in 01M Tris-HCl buffer (pH 80) was incubatedat 60∘C (a) Lane 1 standard maltooligosaccharides (G1ndashG7) lane 2starch used as substrate and lanes 3ndash9 soluble starch hydrolyzedby P stutzeri AS22 after 2min 10min 30min 1 h 2 h 4 h and6 h (b) End products analysis after 6 h hydrolysis of soluble starchGPC analysis was performed on a Bio-Gel P2 column (15 times 200 cm)eluted with water at a rate of 30mLhThe different oligosaccharidesreleased after starch hydrolysis were fractionated and quantified incomparison with standard oligosaccharides ranging from G1 to G7G1 glucose G2 maltose G3 maltotriose G4 maltotetraose G5maltopentaose G6 maltohexaose and G7 maltoheptaose V0 thevoid volume
temperature (Table 5) Among all inhibitors tested the chelat-ing agent EDTA and the reducing agent 120573-mercaptoethanolinactivated the amylolytic activity where it could retain 70and 66of its initial activity respectivelyHowever the serineproteinase inhibitor PMSF showed no significant inhibition(10)
The amylolytic preparation revealed a high stability in thepresence of 5 of the nonionic surfactants (Tween 20 Tween
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991
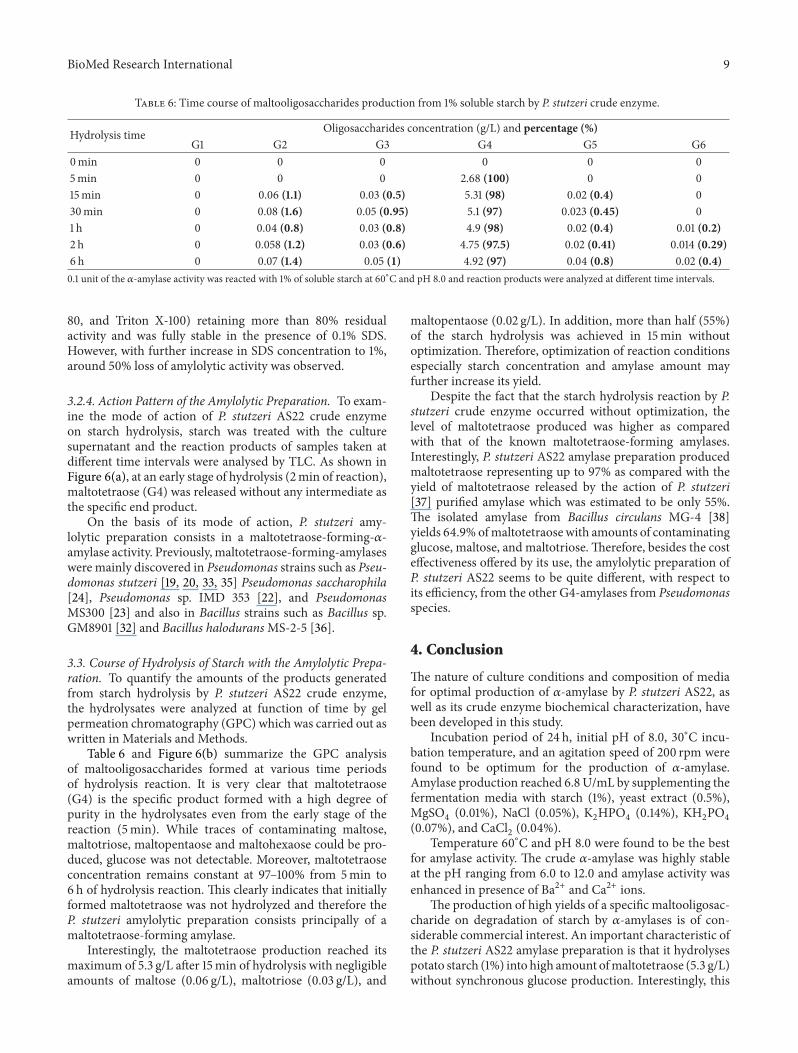
BioMed Research International 9
Table 6 Time course of maltooligosaccharides production from 1 soluble starch by P stutzeri crude enzyme
Hydrolysis time Oligosaccharides concentration (gL) and percentage ()G1 G2 G3 G4 G5 G6
0min 0 0 0 0 0 05min 0 0 0 268 (100) 0 015min 0 006 (11) 003 (05) 531 (98) 002 (04) 030min 0 008 (16) 005 (095) 51 (97) 0023 (045) 01 h 0 004 (08) 003 (08) 49 (98) 002 (04) 001 (02)2 h 0 0058 (12) 003 (06) 475 (975) 002 (041) 0014 (029)6 h 0 007 (14) 005 (1) 492 (97) 004 (08) 002 (04)01 unit of the 120572-amylase activity was reacted with 1 of soluble starch at 60∘C and pH 80 and reaction products were analyzed at different time intervals
80 and Triton X-100) retaining more than 80 residualactivity and was fully stable in the presence of 01 SDSHowever with further increase in SDS concentration to 1around 50 loss of amylolytic activity was observed
324 Action Pattern of the Amylolytic Preparation To exam-ine the mode of action of P stutzeri AS22 crude enzymeon starch hydrolysis starch was treated with the culturesupernatant and the reaction products of samples taken atdifferent time intervals were analysed by TLC As shown inFigure 6(a) at an early stage of hydrolysis (2min of reaction)maltotetraose (G4) was released without any intermediate asthe specific end product
On the basis of its mode of action P stutzeri amy-lolytic preparation consists in a maltotetraose-forming-120572-amylase activity Previously maltotetraose-forming-amylaseswere mainly discovered in Pseudomonas strains such as Pseu-domonas stutzeri [19 20 33 35] Pseudomonas saccharophila[24] Pseudomonas sp IMD 353 [22] and PseudomonasMS300 [23] and also in Bacillus strains such as Bacillus spGM8901 [32] and Bacillus haloduransMS-2-5 [36]
33 Course of Hydrolysis of Starch with the Amylolytic Prepa-ration To quantify the amounts of the products generatedfrom starch hydrolysis by P stutzeri AS22 crude enzymethe hydrolysates were analyzed at function of time by gelpermeation chromatography (GPC) which was carried out aswritten in Materials and Methods
Table 6 and Figure 6(b) summarize the GPC analysisof maltooligosaccharides formed at various time periodsof hydrolysis reaction It is very clear that maltotetraose(G4) is the specific product formed with a high degree ofpurity in the hydrolysates even from the early stage of thereaction (5min) While traces of contaminating maltosemaltotriose maltopentaose and maltohexaose could be pro-duced glucose was not detectable Moreover maltotetraoseconcentration remains constant at 97ndash100 from 5min to6 h of hydrolysis reaction This clearly indicates that initiallyformed maltotetraose was not hydrolyzed and therefore theP stutzeri amylolytic preparation consists principally of amaltotetraose-forming amylase
Interestingly the maltotetraose production reached itsmaximum of 53 gL after 15min of hydrolysis with negligibleamounts of maltose (006 gL) maltotriose (003 gL) and
maltopentaose (002 gL) In addition more than half (55)of the starch hydrolysis was achieved in 15min withoutoptimization Therefore optimization of reaction conditionsespecially starch concentration and amylase amount mayfurther increase its yield
Despite the fact that the starch hydrolysis reaction by Pstutzeri crude enzyme occurred without optimization thelevel of maltotetraose produced was higher as comparedwith that of the known maltotetraose-forming amylasesInterestingly P stutzeri AS22 amylase preparation producedmaltotetraose representing up to 97 as compared with theyield of maltotetraose released by the action of P stutzeri[37] purified amylase which was estimated to be only 55The isolated amylase from Bacillus circulans MG-4 [38]yields 649 ofmaltotetraose with amounts of contaminatingglucose maltose and maltotrioseTherefore besides the costeffectiveness offered by its use the amylolytic preparation ofP stutzeri AS22 seems to be quite different with respect toits efficiency from the other G4-amylases from Pseudomonasspecies
4 Conclusion
The nature of culture conditions and composition of mediafor optimal production of 120572-amylase by P stutzeri AS22 aswell as its crude enzyme biochemical characterization havebeen developed in this study
Incubation period of 24 h initial pH of 80 30∘C incu-bation temperature and an agitation speed of 200 rpm werefound to be optimum for the production of 120572-amylaseAmylase production reached 68UmL by supplementing thefermentation media with starch (1) yeast extract (05)MgSO
4(001) NaCl (005) K
2HPO4(014) KH
2PO4
(007) and CaCl2(004)
Temperature 60∘C and pH 80 were found to be the bestfor amylase activity The crude 120572-amylase was highly stableat the pH ranging from 60 to 120 and amylase activity wasenhanced in presence of Ba2+ and Ca2+ ions
The production of high yields of a specific maltooligosac-charide on degradation of starch by 120572-amylases is of con-siderable commercial interest An important characteristic ofthe P stutzeri AS22 amylase preparation is that it hydrolysespotato starch (1) into high amount ofmaltotetraose (53 gL)without synchronous glucose production Interestingly this
10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991

10 BioMed Research International
amylase preparation producing maltotetraose on a higheryield (98) as compared with that of previously reported G4-amylases seems to be promising in the manufacture of highmaltotetraose syrups from starch
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Alain Heyraud and Laurine Buon(CERMAV CNRS Grenoble FR) for providing us with theGPC analysis
References
[1] M Fujita K Torigoe T Nakada et al ldquoCloning and nucleotidesequence of the gene (amyP) formaltotetraose-forming amylasefrom Pseudomonas stutzeriMO-19rdquo Journal of Bacteriology vol171 no 3 pp 1333ndash1339 1989
[2] H Barreteau C Delattre and P Michaud ldquoProduction ofoligosaccharides as promising new food additive generationrdquoFood Technology and Biotechnology vol 44 no 3 pp 323ndash3332006
[3] T Nakakuki ldquoPresent status and future of functional oligosac-charide development in JapanrdquoPure andApplied Chemistry vol74 no 7 pp 1245ndash1251 2002
[4] H R Palacios P B Schwarz and B L DrsquoAppolonia ldquoEffect of120572-amylases from different sources on the retrogradation andrecrystallization of concentrated wheat starch gels relationshipto bread stalingrdquo Journal of Agricultural and Food Chemistryvol 52 no 19 pp 5978ndash5986 2004
[5] M Rivero-Urgell and A Santamaria-Orleans ldquoOligosaccha-rides application in infant foodrdquo Early Human Developmentvol 65 no 2 pp S43ndashS52 2001
[6] P V Aiyer ldquoAmylases and their applicationsrdquo African Journal ofBiotechnology vol 4 no 13 pp 1525ndash1529 2005
[7] J MalabenduM Chiranjit S Saptadip et al ldquoSalt-independentthermophilic 120572-amylase from Bacillus megaterium VUMB109an efficacy testing for preparation of maltooligosaccharidesrdquoIndustrial Crops and Products vol 41 pp 386ndash390 2013
[8] L Duedahl-Olesen K Matthias Kragh and W ZimmermannldquoPurification and characterisation of a malto-oligosaccharide-forming amylase active at high pH from Bacillus clausii BT-21rdquoCarbohydrate Research vol 329 no 1 pp 97ndash107 2000
[9] A Pandey P Nigam C R Soccol V T Soccol D Singh andRMohan ldquoAdvances inmicrobial amylasesrdquo Biotechnology andApplied Biochemistry vol 31 no 2 pp 135ndash152 2000
[10] J Lalucat A Bennasar R Bosch E Garcıa-Valdes and N JPalleroni ldquoBiology of Pseudomonas stutzerirdquo Microbiology andMolecular Biology Reviews vol 70 no 2 pp 510ndash547 2006
[11] H Maalej N Hmidet O Ghorbel-Bellaaj and M NasrildquoPurification and biochemical characterization of a detergentstable 120572-amylase from Pseudomonas stutzeri AS22rdquo Biotechnol-ogy and Bioprocess Engineering vol 18 no 5 pp 878ndash887 2013
[12] J H Miller Experiments in Molecular Genetics Cold springHarbor Laboratory Cold Spring Harbor NY USA 3rd edition1972
[13] G L Miller ldquoUse of dinitrosalicyIic acid reagent for determina-tion of reducing sugarrdquo Analytical Chemistry vol 31 no 3 pp426ndash428 1959
[14] A Sharma and T Satyanarayana ldquoOptimization of mediumcomponents and cultural variables for enhanced production ofacidic high maltose-forming and Ca2+-independent 120572-amylaseby Bacillus acidicolardquo Journal of Bioscience and Bioengineeringvol 111 no 5 pp 550ndash553 2011
[15] K R Babu and T Satyanarayana ldquoParametric optimizationof extracellular 120572-amylase production by thermophilic Bacilluscoagulansrdquo Folia Microbiologica vol 38 no 1 pp 77ndash80 1993
[16] K Tonomura H Suzuki N Nakamura K Kuraya and OTanabe ldquoOn the inducers of 120572-amylase formation inAspergillusoryzaerdquo Agricultural and Biological Chemistry vol 25 no 1 pp1ndash6 1961
[17] R Morkeberg M Carlsen and J Nielsen ldquoInduction andrepression of 120572-amylase production in batch and continuouscultures of Aspergillus oryraerdquoMicrobiology vol 141 no 10 pp2449ndash2454 1995
[18] R Gupta P Gigras H Mohapatra V K Goswami and BChauhan ldquoMicrobial 120572-amylases a biotechnological perspec-tiverdquo Process Biochemistry vol 38 no 11 pp 1599ndash1616 2003
[19] J F Robyt and R J Ackerman ldquoIsolation purification andcharacterization of a maltotetraose-producing amylase fromPseudomonas stutzerirdquo Archives of Biochemistry and Biophysicsvol 145 no 1 pp 105ndash114 1971
[20] T Nakada M Kubota S Sakai and Y Tsujisaka ldquoPurificationand characterization of two forms of maltotetraose-formingamylase from Pseudomonas stutzerirdquoAgricultural and biologicalchemistry vol 54 no 3 pp 737ndash743 1990
[21] W M Fogarty and C T Kelly Industrial Enzymes and Biotech-nology Elsevier Applied Science New York NY USA 2ndedition 1990 W M Fogarty C T Kelly Eds
[22] W M Fogarty C T Kelly A C Bourke and E M DoyleldquoExtracellular maltotetraose-forming amylase of Pseudomonassp IMD 353rdquo Biotechnology Letters vol 16 no 5 pp 473ndash4781994
[23] H Kobayashi Y Takaki K Kobata H Takami and AInoue ldquoCharacterization of 120572-maltotetraohydrolase producedby Pseudomonas sp MS300 isolated from the deepest site of theMariana trenchrdquo JAMSTEC Journal of Deep Sea Research vol16 pp 109ndash116 2000
[24] S Kobayashi H Okemoto K Hara H Hashimoto andK Yamasato ldquoPreparation and some properties of a novelmaltotetraose-forming enzyme of Pseudomonas saccharophilardquoJournal of the Japanese Society of Starch Science vol 38 no 1 pp27ndash36 1991
[25] K Kathiresan and S Manivannan ldquo120572-Amylase productionby Penicillium fellutanum isolated from mangrove rhizospheresoilrdquoAfrican Journal of Biotechnology vol 5 no 10 pp 829ndash8322006
[26] A Rameshkumar and T Sivasudha ldquoOptimization of nutri-tional constitute for enhanced alpha amylase production usingby solid state fermentation technologyrdquo International Journal ofMicrobiological Research vol 2 no 2 pp 143ndash148 2011
[27] S Sivaramakrishnan D Gangadharan K M NampoothiriC R Soccol and A Pandey ldquo120572-amylases from microbialsourcesmdashan overview on recent developmentsrdquo Food Technol-ogy and Biotechnology vol 44 no 2 pp 173ndash184 2006
[28] RMalhotra SMNoorwez and T Satyanarayana ldquoProductionand partial characterization of thermostable and calcium-independent 120572-amylase of an extreme thermophile Bacillus
BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991

BioMed Research International 11
thermooleovoransNP54rdquoLetters inAppliedMicrobiology vol 31no 5 pp 378ndash384 2000
[29] G Subramanian S Ayyadurai T Sharma S A Singh MRele and L S Kumar ldquoStudies on maltoheaose (G6) producingalkaline amylase fromanovel alkalophilic Streptomyces speciesrdquoThe IIOAB Journal vol 3 no 3 pp 15ndash30 2012
[30] Z Konsoula and M Liakopoulou-Kyriakides ldquoCo-productionof 120572-amylase and 120573-galactosidase by Bacillus subtilis in complexorganic substratesrdquo Bioresource Technology vol 98 no 1 pp150ndash157 2007
[31] Sudha ldquoEffect of different concentrations ofmetal ions on alphaamylase production by Bacillus amyloliquefaciensrdquo Research inBiotechnology vol 3 pp 67ndash71 2012
[32] T U Kim B G Gu J Y Jeong S M Byun and Y CShin ldquoPurification and characterization of a maltotetraose-forming alkaline120572- amylase from an alkalophilicBacillus strainGM8901rdquo Applied and Environmental Microbiology vol 61 no8 pp 3105ndash3112 1995
[33] Y Sakano Y Kashiwagi and T Kobayashi ldquoPurification andproperties of an exo-120572-amylase from Pseudomonas stutzerirdquoAgricultural and Biological Chemistry vol 46 no 3 pp 639ndash646 1982
[34] A Sharma and T Satyanarayana ldquoMicrobial acid-stable 120572-amylases characteristics genetic engineering and applicationsrdquoProcess Biochemistry vol 48 no 2 pp 201ndash211 2013
[35] M J Lee and M J Chung ldquoStudies on the exo-maltotetraohydrolase ofPseudomonas stutzeri IAM 12097mdashpartII characteristics of exo-maltotetraohydrolaserdquo Journal of theKorean Chemical Society vol 27 pp 271ndash273 1984
[36] S Murakami K Nagasaki H Nishimoto et al ldquoPurifica-tion and characterization of five alkaline thermotolerant andmaltotetraose-producing 120572-amylases from Bacillus haloduransMS-2-5 and production of recombinant enzymes in Escherichiacolirdquo Enzyme andMicrobial Technology vol 43 no 4-5 pp 321ndash328 2008
[37] J Schmidt and M John ldquoStarch metabolism in Pseudomonasstutzeri I Studies onmaltotetraose-forming amylaserdquo Biochim-ica et Biophysica Acta vol 566 no 1 pp 88ndash99 1979
[38] Y Takasaki H Shinohara M Tsuruhisa S Hayashi and KImada ldquoMaltotetraose-producing amylase from Bacillus spMG-4rdquo Agricultural and Biological Chemistry vol 55 no 7 pp1715ndash1720 1991




















