
Characterization and phylogenetic analysis of cellulase producing Streptomyces noboritoensis SPKC1
7
Characterization and phylogenetic analysis of biodemulsifier-producing bacteria Xiang-Feng Huang * , Wei Guan, Jia Liu, Li-Jun Lu, Jing-Cheng Xu * , Qi Zhou College of Environmental Science and Engineering, State Key Laboratory of Pollution Control and Resource Reuse, Tongji University, Shanghai 20092, China article info Article history: Received 5 May 2009 Received in revised form 28 July 2009 Accepted 31 July 2009 Available online 31 August 2009 Keywords: Biodemulsifier-producing bacteria Demulsification Phylogenetic analysis Biosurfactant abstract Based on demulsification performance, twenty biodemulsifier-producing strains were isolated from various environmental sources. Five of them achieved nearly or over 90% of emulsion breaking ratio within 24 h. With the aid of biochemical and physiological tests and 16S rDNA analysis, these isolates were classified into eleven genera, in which six genera (Brevibacillus sp., Dietzia sp., Ochrobactrum sp., Pusillimonas sp., Sphingopyxis sp. and Achromobacter sp.) were firstly reported as demulsifying strains. Moreover, with data in this study and other literatures, a phylogenic tree was constructed, showing a rich diversity of demulsifying bacteria. Half of these bacteria belong to Actinobacterale order, which is famous for hydrocarbon degradation and biosurfactant biosynthesis. However, some strains in the same genera differed remarkably in demulsifying capability, surface properties and biochemical and physio- logical characteristics. This implied the biosynthesis and composition of biodemulsifier were more complicated than expected. Ó 2009 Elsevier Ltd. All rights reserved. 1. Introduction Demulsifier is largely consumed in the treatment of emulsions generated in the recovery and processing of crude petroleum (Das, 2001). As water content in these emulsions increases, demul- sification becomes more problematic. Most emulsions produced in oilfields are W/O type and almost can be destabilized by chemi- cally-synthesized demulsifier, which is composed of refractory or- ganic polymers. Due to this feature, the separated water from demulsification should be treated properly before discharge to avoid hazardous risk on ambient ecosystem. This thereby greatly limits the application of chemical demulsifiers. Recently, biodemulsifier has attracted more and more attention in research due to their low toxicity and easy biodegradability (Amezcua-Vega et al., 2007). With the introduction of some spe- cific functional groups, biodemulsifiers are able to perform well under extreme conditions and surpass chemical demulsifiers in a wide range of applications. (Desai and Banat, 1997). Up to now several biodemulsifier-producing strains have been successfully isolated. Extensive studies were conducted particu- larly on Nocardia amarae (Cairns et al., 1982), Torulopsis bombicola (Duvnjak and Kosaric, 1987), Bacillus sp. (Janiyani et al., 1994) and Micrococcus sp. (Das, 2001). However, these studies mainly concen- trated on the isolation of demulsifying strains, the optimization of cultivation conditions and evaluation of demulsifying capability. Little light has been shed on the sourcing, distribution and characterization of demulsifying strains. On the contrary, biosur- factant-producing bacteria have already been studied regarding their distribution in nature and microbial diversity. They were found widely present in petroleum-polluted or undisturbed envi- ronment (Bodour et al., 2003). With the aid of molecular biological analysis means, their phylogenic evolution was revealed. It clearly indicated that the distribution of biosurfactant-producing bacteria covered three divisions of Eubacteria phylum and two divisions of Archaea phylum (Bodour et al., 2003). The results provided a roadmap to the future research, especially in isolation and identi- fication of new biosurfactant-producing strains as well as the produced biosurfactants. Although a couple of biosurfactant-pro- ducing strains isolated from diesel-contaminated soils exhibited promising potential in environment remediation (Bento et al., 2005), it has not been clearly identified which species of biosurfac- tant-producing bacteria are able to break emulsions. As a special kind of biosurfactant-producing bacteria, demulsifying strains were not screened solely by surface activity as biosurfactant-pro- ducing bacteria, but according to their demulsification ability. This definitely results in difference of phylogenetic scope between these two groups of bacteria. For further exploration of biodemul- sifier-producing bacteria, it is necessary and urgent to study their distribution in environment and microbial diversity so as to improve the screening efficiency of biodemulsifier-producing bacteria, to direct genetic modification and to optimize the biosyn- thesis of demulsifier. The aim of this paper is to study the microbial diversity and phylogenic relationship of biodemulsifier-producing bacteria. Based on previous work (Huang et al., 2009), twenty biodemulsifi- er-producing strains were identified by 16S rDNA sequencing 0960-8524/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.biortech.2009.07.086 * Corresponding authors. Tel./fax: +86 021 65982592. E-mail addresses: [email protected], [email protected] (X.-F. Huang), [email protected] (J.-C. Xu). Bioresource Technology 101 (2010) 317–323 Contents lists available at ScienceDirect Bioresource Technology journal homepage: www.elsevier.com/locate/biortech
Transcript of Characterization and phylogenetic analysis of cellulase producing Streptomyces noboritoensis SPKC1

Bioresource Technology 101 (2010) 317–323
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Characterization and phylogenetic analysis of biodemulsifier-producing bacteria
Xiang-Feng Huang *, Wei Guan, Jia Liu, Li-Jun Lu, Jing-Cheng Xu *, Qi ZhouCollege of Environmental Science and Engineering, State Key Laboratory of Pollution Control and Resource Reuse, Tongji University, Shanghai 20092, China
a r t i c l e i n f o
Article history:Received 5 May 2009Received in revised form 28 July 2009Accepted 31 July 2009Available online 31 August 2009
Keywords:Biodemulsifier-producing bacteriaDemulsificationPhylogenetic analysisBiosurfactant
0960-8524/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.biortech.2009.07.086
* Corresponding authors. Tel./fax: +86 021 6598259E-mail addresses: [email protected], ly008150@
[email protected] (J.-C. Xu).
a b s t r a c t
Based on demulsification performance, twenty biodemulsifier-producing strains were isolated fromvarious environmental sources. Five of them achieved nearly or over 90% of emulsion breaking ratiowithin 24 h. With the aid of biochemical and physiological tests and 16S rDNA analysis, these isolateswere classified into eleven genera, in which six genera (Brevibacillus sp., Dietzia sp., Ochrobactrum sp.,Pusillimonas sp., Sphingopyxis sp. and Achromobacter sp.) were firstly reported as demulsifying strains.Moreover, with data in this study and other literatures, a phylogenic tree was constructed, showing arich diversity of demulsifying bacteria. Half of these bacteria belong to Actinobacterale order, which isfamous for hydrocarbon degradation and biosurfactant biosynthesis. However, some strains in the samegenera differed remarkably in demulsifying capability, surface properties and biochemical and physio-logical characteristics. This implied the biosynthesis and composition of biodemulsifier were morecomplicated than expected.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
Demulsifier is largely consumed in the treatment of emulsionsgenerated in the recovery and processing of crude petroleum(Das, 2001). As water content in these emulsions increases, demul-sification becomes more problematic. Most emulsions produced inoilfields are W/O type and almost can be destabilized by chemi-cally-synthesized demulsifier, which is composed of refractory or-ganic polymers. Due to this feature, the separated water fromdemulsification should be treated properly before discharge toavoid hazardous risk on ambient ecosystem. This thereby greatlylimits the application of chemical demulsifiers.
Recently, biodemulsifier has attracted more and more attentionin research due to their low toxicity and easy biodegradability(Amezcua-Vega et al., 2007). With the introduction of some spe-cific functional groups, biodemulsifiers are able to perform wellunder extreme conditions and surpass chemical demulsifiers in awide range of applications. (Desai and Banat, 1997).
Up to now several biodemulsifier-producing strains have beensuccessfully isolated. Extensive studies were conducted particu-larly on Nocardia amarae (Cairns et al., 1982), Torulopsis bombicola(Duvnjak and Kosaric, 1987), Bacillus sp. (Janiyani et al., 1994) andMicrococcus sp. (Das, 2001). However, these studies mainly concen-trated on the isolation of demulsifying strains, the optimization ofcultivation conditions and evaluation of demulsifying capability.Little light has been shed on the sourcing, distribution and
ll rights reserved.
2.online.sh.cn (X.-F. Huang),
characterization of demulsifying strains. On the contrary, biosur-factant-producing bacteria have already been studied regardingtheir distribution in nature and microbial diversity. They werefound widely present in petroleum-polluted or undisturbed envi-ronment (Bodour et al., 2003). With the aid of molecular biologicalanalysis means, their phylogenic evolution was revealed. It clearlyindicated that the distribution of biosurfactant-producing bacteriacovered three divisions of Eubacteria phylum and two divisions ofArchaea phylum (Bodour et al., 2003). The results provided aroadmap to the future research, especially in isolation and identi-fication of new biosurfactant-producing strains as well as theproduced biosurfactants. Although a couple of biosurfactant-pro-ducing strains isolated from diesel-contaminated soils exhibitedpromising potential in environment remediation (Bento et al.,2005), it has not been clearly identified which species of biosurfac-tant-producing bacteria are able to break emulsions. As a specialkind of biosurfactant-producing bacteria, demulsifying strainswere not screened solely by surface activity as biosurfactant-pro-ducing bacteria, but according to their demulsification ability. Thisdefinitely results in difference of phylogenetic scope betweenthese two groups of bacteria. For further exploration of biodemul-sifier-producing bacteria, it is necessary and urgent to study theirdistribution in environment and microbial diversity so as toimprove the screening efficiency of biodemulsifier-producingbacteria, to direct genetic modification and to optimize the biosyn-thesis of demulsifier.
The aim of this paper is to study the microbial diversity andphylogenic relationship of biodemulsifier-producing bacteria.Based on previous work (Huang et al., 2009), twenty biodemulsifi-er-producing strains were identified by 16S rDNA sequencing
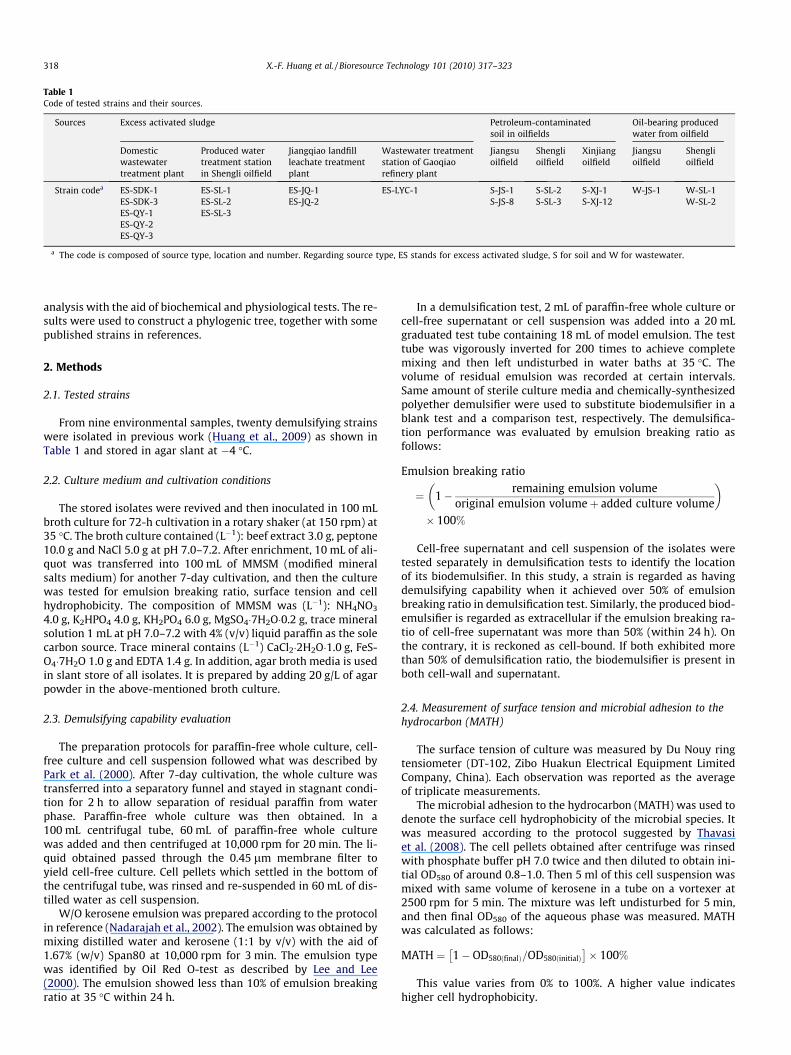
Table 1Code of tested strains and their sources.
Sources Excess activated sludge Petroleum-contaminatedsoil in oilfields
Oil-bearing producedwater from oilfield
Domesticwastewatertreatment plant
Produced watertreatment stationin Shengli oilfield
Jiangqiao landfillleachate treatmentplant
Wastewater treatmentstation of Gaoqiaorefinery plant
Jiangsuoilfield
Shenglioilfield
Xinjiangoilfield
Jiangsuoilfield
Shenglioilfield
Strain codea ES-SDK-1 ES-SL-1 ES-JQ-1 ES-LYC-1 S-JS-1 S-SL-2 S-XJ-1 W-JS-1 W-SL-1ES-SDK-3 ES-SL-2 ES-JQ-2 S-JS-8 S-SL-3 S-XJ-12 W-SL-2ES-QY-1 ES-SL-3ES-QY-2ES-QY-3
a The code is composed of source type, location and number. Regarding source type, ES stands for excess activated sludge, S for soil and W for wastewater.
318 X.-F. Huang et al. / Bioresource Technology 101 (2010) 317–323
analysis with the aid of biochemical and physiological tests. The re-sults were used to construct a phylogenic tree, together with somepublished strains in references.
2. Methods
2.1. Tested strains
From nine environmental samples, twenty demulsifying strainswere isolated in previous work (Huang et al., 2009) as shown inTable 1 and stored in agar slant at �4 �C.
2.2. Culture medium and cultivation conditions
The stored isolates were revived and then inoculated in 100 mLbroth culture for 72-h cultivation in a rotary shaker (at 150 rpm) at35 �C. The broth culture contained (L�1): beef extract 3.0 g, peptone10.0 g and NaCl 5.0 g at pH 7.0–7.2. After enrichment, 10 mL of ali-quot was transferred into 100 mL of MMSM (modified mineralsalts medium) for another 7-day cultivation, and then the culturewas tested for emulsion breaking ratio, surface tension and cellhydrophobicity. The composition of MMSM was (L�1): NH4NO3
4.0 g, K2HPO4 4.0 g, KH2PO4 6.0 g, MgSO4�7H2O�0.2 g, trace mineralsolution 1 mL at pH 7.0–7.2 with 4% (v/v) liquid paraffin as the solecarbon source. Trace mineral contains (L�1) CaCl2�2H2O�1.0 g, FeS-O4�7H2O 1.0 g and EDTA 1.4 g. In addition, agar broth media is usedin slant store of all isolates. It is prepared by adding 20 g/L of agarpowder in the above-mentioned broth culture.
2.3. Demulsifying capability evaluation
The preparation protocols for paraffin-free whole culture, cell-free culture and cell suspension followed what was described byPark et al. (2000). After 7-day cultivation, the whole culture wastransferred into a separatory funnel and stayed in stagnant condi-tion for 2 h to allow separation of residual paraffin from waterphase. Paraffin-free whole culture was then obtained. In a100 mL centrifugal tube, 60 mL of paraffin-free whole culturewas added and then centrifuged at 10,000 rpm for 20 min. The li-quid obtained passed through the 0.45 lm membrane filter toyield cell-free culture. Cell pellets which settled in the bottom ofthe centrifugal tube, was rinsed and re-suspended in 60 mL of dis-tilled water as cell suspension.
W/O kerosene emulsion was prepared according to the protocolin reference (Nadarajah et al., 2002). The emulsion was obtained bymixing distilled water and kerosene (1:1 by v/v) with the aid of1.67% (w/v) Span80 at 10,000 rpm for 3 min. The emulsion typewas identified by Oil Red O-test as described by Lee and Lee(2000). The emulsion showed less than 10% of emulsion breakingratio at 35 �C within 24 h.
In a demulsification test, 2 mL of paraffin-free whole culture orcell-free supernatant or cell suspension was added into a 20 mLgraduated test tube containing 18 mL of model emulsion. The testtube was vigorously inverted for 200 times to achieve completemixing and then left undisturbed in water baths at 35 �C. Thevolume of residual emulsion was recorded at certain intervals.Same amount of sterile culture media and chemically-synthesizedpolyether demulsifier were used to substitute biodemulsifier in ablank test and a comparison test, respectively. The demulsifica-tion performance was evaluated by emulsion breaking ratio asfollows:
Emulsion breaking ratio
¼ 1� remaining emulsion volumeoriginal emulsion volumeþ added culture volume
� �
� 100%
Cell-free supernatant and cell suspension of the isolates weretested separately in demulsification tests to identify the locationof its biodemulsifier. In this study, a strain is regarded as havingdemulsifying capability when it achieved over 50% of emulsionbreaking ratio in demulsification test. Similarly, the produced biod-emulsifier is regarded as extracellular if the emulsion breaking ra-tio of cell-free supernatant was more than 50% (within 24 h). Onthe contrary, it is reckoned as cell-bound. If both exhibited morethan 50% of demulsification ratio, the biodemulsifier is present inboth cell-wall and supernatant.
2.4. Measurement of surface tension and microbial adhesion to thehydrocarbon (MATH)
The surface tension of culture was measured by Du Nouy ringtensiometer (DT-102, Zibo Huakun Electrical Equipment LimitedCompany, China). Each observation was reported as the averageof triplicate measurements.
The microbial adhesion to the hydrocarbon (MATH) was used todenote the surface cell hydrophobicity of the microbial species. Itwas measured according to the protocol suggested by Thavasiet al. (2008). The cell pellets obtained after centrifuge was rinsedwith phosphate buffer pH 7.0 twice and then diluted to obtain ini-tial OD580 of around 0.8–1.0. Then 5 ml of this cell suspension wasmixed with same volume of kerosene in a tube on a vortexer at2500 rpm for 5 min. The mixture was left undisturbed for 5 min,and then final OD580 of the aqueous phase was measured. MATHwas calculated as follows:
MATH ¼ 1� OD580ðfinalÞ=OD580ðinitialÞ� �
� 100%
This value varies from 0% to 100%. A higher value indicateshigher cell hydrophobicity.

X.-F. Huang et al. / Bioresource Technology 101 (2010) 317–323 319
2.5. Biochemical and physiological characterization
The biochemical and physiological characterization of each iso-late followed Bergey’s Manual (the 9th edition) (Holt et al., 1994).For various species, different tests were conducted, including Gramstaining, acid-fast staining, spore staining, oxidase reaction, cata-lase reaction, glucose fermentation, acid production (from glucose,xylose, maltose and mannitol), nitrate reduction, hydrogen sulfideproduction, indole production, enzyme production (such asb-galactosidase, lysine-decarboxylase, ornithine decarboxylase,acetamidase, urease, arginine dihydrolase), MacConkey cultivation,citrate utilization and Voges Proskauer (VP) reaction.
2.6. PCR and sequencing identification of 16S rDNA and construction ofphylogenic tree
In the identification of 16S rDNA, DNA was extracted with DNAisolation kit (Omega). 16S rDNA encoding genes were amplified byPCR using universal primer 8F (50-AGAGTTTGATCCTGGCTCAG-3’)and 1492R (5’-GGTTACCTTGTTACGACTT-30). Each 50 lL of PCRmixture contained 1.5 lL template DNA, 0.25 lL Taq DNA poly-merase, 5 lL PCR buffer, 5 lL 2.5 mM dNTPs, 2 lL of each primer(6.5 lmol/L) and double distilled water. PCR program for amplifi-cation was initial denaturation at 94 �C for 3 min followed by 35cycles consisting of 94 �C for 30 s, 56 �C for 30 s and 72 �C for 80s, and a final extension at 72 �C for 10 min. The PCR product wasvisualized on agarose gel by electrophoresis. The purified PCRproduct was ligated to pMD18-T vector and then transferred intoEscherichia coli Top10 for cultivation. When colonies developed inagar plate, PCR identification was carried out with universal primerM13R(50-AGCGGATAACAATTTCACACAGGA-30) and M13F(50-CGCC-AGGGTTTTCCCAGTCACGAC-30) for T vectors in the same procedureas described above. The sequence of cloned product was analyzedby Invitrogen.
The 16S rDNA gene nucleotide sequence of each isolate wascompared with known sequences in GenBank using BLAST pro-gram. The sequence alignment was carried out with ClusterX(version 2.0). Sequences of isolates from this studies and otherreferences were used to construct a phylogenic trees with PAUP*
(version 4.0) by using neighbor-jointing method with Hasegawa-Kishino-Yano (HKY85) model (Bodour et al., 2003).
3. Results and discussion
3.1. Demulsifying capability and surface properties of tested strains
The paraffin-free culture of each isolate was examined for sur-face tension, cell hydrophobicity, location of produced biodemulsi-fier and emulsion breaking. The results were shown in Table 2.
All tested strains showed more than 50% of emulsion breakingratio, in which five strains (ES-QY-3, ES-SL-1, S-SL-2, S-XJ-1, S-XJ-12) exhibited nearly or over 90% of emulsion breaking ratio, muchhigher than the ratio achieved by the chemical demulsifier(73 ± 2%). The performance of the rest was either similar or inferiorto the chemical demulsifier. In previous studies, the productionand type of the produced biosurfactant was found closely relatedto carbon source and other cultivation conditions (Gogotov andKhodakov, 2008). Paraffin is the sole carbon source in this study.Various strains may have different preference in carbon sourcesand different utilization rate of paraffin. Thereby, the demulsifyingcapability of those inferior strains would be improved by changingthe carbon source or optimizing cultivation conditions.
Biodemulsifiers are a group of surface-active bio-products withboth hydrophilic and hydrophobic ends. For example, with aid ofthin layer chromatography and Fourier transform infrared
spectrometer, the biodemulsifier produced by S-XJ-1 was identi-fied as lipopeptide, which had amino-acid chain as hydrophilicend and hydrocarbon chain as hydrophobic end (Huang et al.,2009). A good demulsifier possesses three traits: good miscibilityin dispersed phase, good diffusivity to ensure a high enough diffu-sion flux to the interface and high surface activity to reduce thegradients of interfacial tension (Ivanov and Kralchevsky, 1997).The amphipathic molecule of biodemulsifier facilitates its adsorp-tion on oil/water interface and helps to alter the properties of theinterface. It was found in demulsification tests that the culturesof five highly-efficient strains quickly disperse into the emulsion,showing good miscibility. Moreover, their culture all displayedhigh surface activity with surface tension ranging from 30 to50 mN/m. In an emulsion, this feature enables the biodemulsifierto substitute emulsifier at the interface, which further leads to floc-culation and coalescence of dispersed globules and eventuallycomplete separation of two phases. Besides the amphipathic mol-ecule of biodemulsifer, high cell hydrophobicity can also contrib-ute to demulsification to some extent. For example, four of thefive strains exhibited a high MATH value, which indicated a highaffinity of the cells to kerosene.
The location of produced biodemulsifer determined the degreeof difficulty in its extraction and purification, which not only af-fected the in-depth investigation of biodemulsifier, but also con-tributed significantly to the production cost. As a result, researchadvances in a quicker pace for extracellular biodemulsifier, whichare readily-extractable. As shown in Table 2, nine strains producedextracellular biodemulsifier, eight produced cell-bound and threeproduced both. Moreover, highly-efficient biodemulsifier (withover 90% of emulsion breaking ratio) can be found in both extracel-lular and cell-bound form. Simultaneous production of extracellu-lar and cell-bound biosurfactants were also reported in otherresearch with hydrocarbon as substrate (Franzetti et al., 2008).By comparing the sources of these strains, it was found that mostof the strains isolated from activated sludge produced extracellularbiodemulsifiers, while the strains from petroleum-contaminatedsoils and waters mainly synthesized cell-bound biodemulsifiers.
In this study, the strains were isolated from samples taken fromseveral oilfields, which scattered over China, and wastewater treat-ment plants treating various wastewater. They represented a vari-ety of sources (as shown in Table 1). It is shown in Table 2 thatsurface properties of the tested isolates showed some correlationwith their sources. Seven strains with low surface tension and ninestrains with high MATH were isolated from petroleum-relatedsources, such as produced water of oilfield, activated sludge treat-ing produced water or oilfield soils. Moreover, four of the fivehighly-efficient demulsifying strains (except ES-QY-3) were alsoisolated from petroleum-related sources. This may be explainedas follows. In petroleum-contaminated environment, microbialdiversity decreases due to the natural selection and only strainscapable of utilizing bio-toxic hydrocarbons will survive and pre-dominate (Li et al., 2000). To facilitate the utilization of hydrocar-bons, these strains would either produce surface-active substancesor increase their affinity to oil by altering cell hydrophobicity.
3.2. Identification of tested strains and phylogenic study
All these isolates were chemoheterotrophs with four Gram-posi-tive and sixteen Gram-negative. The morphology of cells and colo-nies as well as their biochemical and physiological characteristicswere listed in Electronic Annex 1 in the online version of this article.
Due to intrinsic limitations, the biochemical and physiologicalfeatures can only provide preliminarily identification (Bizet et al.,1997). The final identification of strains was accomplished bycombing the alignment results of 16S rDNA sequence analysis withbiochemical and physiological characteristics.

Table 2Surface properties and demulsifying capability of tested isolatesa.
Source No. Strain code Paraffin-free culture MATH (%) Location of producedbiodemulsifierc
Emulsion breaking ratioin 24 h(%)b
Surface tension(mN/m)
Excess sludge 1 ES-JQ-1 65 ± 5 42.6 ± 0.4 44 ± 6 E2 ES-JQ-2 68 ± 1 45.8 ± 0.3 8 ± 6 E3 ES-LYC-1 70 ± 2 38.8 ± 0.5 54 ± 5 E & C4 ES-QY-1 69 ± 2 41.1 ± 0.2 68 ± 1 E5 ES-QY-2 50 ± 4 49.1 ± 0.9 28 ± 4 E6 ES-QY-3 98 ± 2 46.3 ± 0.3 17 ± 0 E7 ES-SDK-1 59 ± 4 49.3 ± 0.5 20 ± 1 E8 ES-SDK-3 62 ± 3 48.1 ± 0.4 0.3 ± 0 E9 ES-SL-1 99 ± 1 49.3 ± 0.3 57 ± 2 E10 ES-SL-2 58 ± 5 54.2 ± 1.7 9 ± 1 C11 ES-SL-3 67 ± 4 54.8 ± 2.2 65 ± 4 E
Petroleum-contaminatedsoil
12 S-JS-1 72 ± 2 30.3 ± 0.0 70 ± 6 C13 S-JS-8 65 ± 3 36.0 ± 1.9 70 ± 1 C14 S-SL-2 88 ± 3 34.6 ± 1.3 81 ± 2 E & C15 S-SL-3 50 ± 2 32.8 ± 0.2 50 ± 2 E & C16 S-XJ-1 91 ± 4 33.9 ± 1.1 62 ± 1 C17 S-XJ-12 100 ± 0 30.4 ± 0.2 46 ± 0 C
Oil-bearing produced waterfrom oilfield
18 W-JS-1 50 ± 2 53.7 ± 0.2 54 ± 4 C19 W-SL-1 56 ± 4 54.4 ± 0.5 26 ± 0 C20 W-SL-2 68 ± 3 50.5 ± 1.4 14 ± 4 C
a Data were reported as average of triplicate observations ± standard deviation.b The blank (with 2 ml of virgin culture instead of biodemulsifier) showed 3 ± 1% of emulsion braking ratio within 24 h.c As to the location of biodemulsifier, E stands for extracellular and C for cell-bound. It was determined according to what is described in Section 2.3. Emulsion breaking
ratios of cell-free supernatant and cell suspension are not shown here.
320 X.-F. Huang et al. / Bioresource Technology 101 (2010) 317–323
Strains ES-SDK-3 exhibited high homology (99%) with Castellan-iella sp. and Alcaligenes sp. Castellaniella sp. is capable of degradingnonylphenol ethoxylates (NPnEO) with low ethoxylation degree,which is particularly recalcitrant to biodegradation (Di Gioiaet al., 2008). But no reports can be found on the demulsifying capa-bility of this genus. Although it was reported as heterocyclicamine-degrading denitrifying bacteria, Alcaligenes sp. was alsofound showing surface activity (producing bioemulsan) (Toledoet al., 2008) or demulsifying capability (Nadarajah et al., 2002).The phylogenetic analysis (the phylogenic tree were not shownhere) indicated that ES-SDK-3 was closer to Alcaligenes sp. and thusit is probably a species of Alcaligenes sp.
Despite 98% of homology with both Castellaniella sp. and Alcalig-enes sp., strain ES-JQ-2 was also classified into the latter genus in asimilar way.
The highest homology obtained by strain S-JS-1 was 93% of sim-ilarity to Dietzia sp., which, together with Rhodococcus sp., belongsto Nocardioform Actinomycetes (Holt et al., 1994). Both showed sim-ilar features (Bizet et al., 1997). Previous studies showed that Rho-dococcus sp. was able to produce biosurfactant (Gogotov andKhodakov, 2008) and break emulsions (Das, 2001). Up to now, Diet-zia sp. was mainly isolated from petroleum-polluted soils and re-ported in degradation of oils (von der Weid et al., 2007). Becausethe biochemical and physiological features of strain S-JS-1 com-plied with those reported for Dietzia sp., it is probably a new spe-cies of Dietzia sp.
The rest seventeen strains all obtained single alignment resultwith more than 97% of similarity to the assigned genus. Their mor-phological, biochemical and physiological features were also con-sistent with the description in references. If 97% similarity of 16SrDNA gene sequence is defined as a good match standard, nineteenof the twenty tested strains can be identified into eleven genera asshow in Table 3.
3.3. Phylogenic analysis of biodemulsifier-producing bacteria
With the addition of the eleven genera found in this study, allthe known demulsifying bacteria can be classified into the follow-
ing genera, Nocardia sp.(Stewart et al., 1983), Corynebacteriumsp.(Stewart et al., 1983), Bacillus sp.(Janiyani et al., 1994), Rhodo-coccus sp.(Ma et al., 2006; Wilkinson and Cooper, 1985), Alcaligenessp.(Nadarajah et al., 2002), Pseudomonas sp.(Nadarajah et al.,2002), Streptomyces sp.(Park et al., 2000), Micrococcus sp.(Das,2001), Torulopsis bombicola (Duvnjak and Kosaric, 1987), Mycobac-terium sp.(Nadarajah et al., 2002), Acinetobacer sp.(Nadarajah et al.,2002), Alteromonas sp.(Nishimaki et al., 1999), Aeromonassp.(Nishimaki et al., 1999) and Arthrobacter sp.(Das, 2001). In orderto reveal the evolution of demulsifying bacteria, the 16S rDNA se-quence of all found strains were used to construct a phylogenictree with PAUP by using the neighbor-joining method.
The phylogenic tree (as shown in Fig. 1) indicated a rich diver-sity of demulsifying bacteria, which covered three divisions (Prote-obacteria, Firmicutes and Actinobacteria) of Eubacteria andAscomycota division of Eukarya. It is worth noting that the classifi-cation of some strains published long time ago has been updateddue to recent advances in bacterial taxonomy. For example, N.amarae ATCC72808 (Stewart et al., 1983) has been re-classifiedinto Gordonia amarae according to the analysis result of small-sub-unit ribosomal DNA sequence (Ruimy et al., 1994). Another species,Rhodococcus aurantiacus, which was re-classified into Gordona aur-antiaca, was then grouped into Tsukamurella inchonensis and hasrecently been changed to Tsukamurella paurometabola by Kattaret al. (2001). Other examples are Rhodococcus rubropertinctus re-classified into Gordonia rubripertinctu (Rainey et al., 1995); Coryne-bacterium fascians re-grouped into Rhodococcus fascians in 1984(Rainey et al., 1995); a demulsifying yeast, Torulopsis bombicolaATCC22214 (Duvnjak and Kosaric, 1987) re-classified into Candidabombicola (Odds et al., 1997). As a result, demulsifying strains areexpanded to three genera (Gordonia sp., Tsukamurella sp. and Can-dida sp.), which still belong to the four divisions mentioned above.
The isolates in this study were evenly distributed among Prote-obacteria, Firmicutes and Actinobacteria division. So far no demulsi-fying strain was found in Fungus. Except five genera (including twoof Rhodococcus sp., two of Bacillus sp., three of Alcaligenes sp., two ofGordonia sp. and one of Pseudomonas sp.) were already reported inprevious biodemulsifier studies, the remaining six genera of this

Table 3Identification of the twenty tested strains with 16S rDNA gene analysis.
No. Strain code GenBankRegister No.
16S rDNA gene sequence Genera
Nearest relative in GenBank Similarity (%)
1 ES-JQ-1 FJ529026 Brevibacillus agri strain NCHU1002(AY319301) 99 Brevibacillus sp.2 ES-JQ-2 FJ529027 Alcaligenes sp. strain Ic4(DQ421393) 98 Alcaligenes sp.3 ES-LYC-1 FJ529028 Gordonia sp. strain D2(DQ787430) 99 Gordonia sp.4 ES-QY-1 FJ529029 Dietzia sp. strain ES18(AY028326) 100 Dietzia sp.5 ES-QY-2 FJ529030 Ochrobactrum intermedium clone kl-2(EU816698) 99 Ochrobactrum sp.6 ES-QY-3 FJ529031 Pusillimonas terrae strain BN9T(DQ466075) 98 Pusillimonas sp.7 ES-SDK-1 FJ529037 Sphingopyxis granuli strain Kw07(AY563034) 99 Sphingopyxis sp.8 ES-SDK-3 FJ531636 Alcaligenes sp. strain ESPY2(A-III)(EF205261) 99 Alcaligenes sp.9 ES-SL-1 FJ529038 Brevibacillus borstelensis strain MH301(DQ350830) 99 Brevibacillus sp.10 ES-SL-2 FJ529039 Bacillus badius strain NBRC 15713(AB271748) 99 Bacillus sp.11 ES-SL-3 FJ529040 Brevibacillus borstelensis strain T2–1(AB215102) 99 Brevibacillus sp.12 S-JS-1 FJ529032 Dietzia natronolimnaea strain LL 51(DQ821754) 93 Dietzia sp.13 S-JS-8 FJ529034 Pseudomonas sp. strain BFXJ-8(EU013945) 99 Pseudomonas sp.14 S-SL-2 FJ529022 Rhodococcus sp. strain F12(EU697084) 99 Rhodococcus sp.15 S-SL-3 FJ529024 Rhodococcus sp. strain E33(AY114109) 100 Rhodococcus sp.16 S-XJ-1 EF117974 Alcaligenes sp. strain mp-2(AY331576) 99 Alcaligenes sp.17 S-XJ-12 FJ529035 Dietzia sp. strain ES18(AY028326) 100 Dietzia sp.18 W-JS-1 FJ529036 Gordonia sp. strain D2(DQ787430) 99 Gordonia sp.19 W-SL-1 FJ529041 Achromobacter sp. strain EP17(AM398226) 99 Achromobacter sp.20 W-SL-2 FJ529042 Bacillus sp. strain BSi20511(EF673289) 99 Bacillus sp.
Fig. 1. Phylogenic tree was based on gene sequences, showing the position of 20 demulsifying strains found in this study and published demulsifying strains from literatures.The unrooted tree was created by using the neighbor-joining method. The bar indicates 0.05 substitutions per nucleotide position.
X.-F. Huang et al. / Bioresource Technology 101 (2010) 317–323 321
study were firstly reported as demulsifying strains. They wereOchrobactrum sp., Pusillimonas sp., Sphingopyxis sp. and Achromo-bacter sp. of Proteobacteria division; Brevibacillus sp. of Firmicutesdivision and Dietzia sp. of Actinobacteria division. In addition, thisstudy has contributed five demulsifying strains (ES-JQ-2, ES-QY-3, ES-SDK-3, S-XJ-1, W-SL-1) to Betaproteobacteria class. Thisgreatly increased the quantity of demulsifying strains in this class.
As shown by the phylogenic tree, more than ten demulsifyinggenera were found in Actinobacteria class of Actinobacteria division(according to LPSN and NCBI taxonomy system). In this study, fiveof seven isolates in this division (ES-LYC-1, S-JS-1, S-SL-2, S-SL-3,S-XJ-12) were able to reduce surface tension below 40 mN/m; six(ES-LYC-1, ES-QY-1, S-JS-1, S-SL-2, S-SL-3, W-JS-1) showed more
than 50% of MATH and two achieved approximately 90% ofemulsion breaking ratio. Compared with strains in the other twodivisions, most of the strains in Actinobacteria division showedhigher surface activity and cell hydrophobicity, indicating promis-ing potential in biodegradation of hydrocarbon and production ofbiosurfactant. As a result, strains in Actinobacteridae – Actinobacte-rale order also exhibited other valuable capabilities such asbiosynthesis of antibiotic and useful metabolism byproducts,which can be applied in biological treatment of wastewater or solidwastes (Goodfellow et al., 1996). For instance, Mycobacterium sp.,Corynebacterium sp. and Brevibacterium sp. were able to produceextracellular lipopeptide biosurfactants (Haferburg et al., 1986)and a Rhodococcus wratislaviensis strain BN38 was capable of

322 X.-F. Huang et al. / Bioresource Technology 101 (2010) 317–323
producing glycolipid biosurfactants (Tuleva et al., 2008). Somestrains, such as Mycobacterium, Rhodococcus, Dietzia and Gordoniaalkanivorans, exhibited ability of degrading hydrocarbons (Britoet al., 2006; Ta-Chen et al., 2008).
In this study, six of eleven genera contained more than twostrains. Surprisingly, the strains in the same genera were distinctin many aspects. For example, both ES-LYC-1 and W-JS-1 wereidentified as Gordonia sp., and both showed 99% of homology in16S rDNA analysis. However, they gave opposite response in gramstaining, oxidase reaction and hydrogen sulfide production.Regarding demulsification performance, W-JS-1 performed farpoorer than ES-LYC-1 (in Table 2). The surface tension of their cul-tures also differed remarkably. Strain ES-LYC-1 produced bothextracellular and cell-bound biodemulsifier, while W-JS-1 onlyproduced cell-bound biodemulsifier. Distinctions were also ob-served in other genera, such as ES-QY-1 and S-XJ-12 of Dietziasp., even though these two strains showed 100% of homology intheir 16S rDNA sequence. As reported in previous study, thehomology mixture of biosurfactant produced by strains in thesame genera may differed slightly in the composition or chemicalstructures, which would have great consequence on its surfaceactivity (Franzetti et al., 2008). Similar to biosurfactant, our previ-ous work also showed that produced biodemulsifer was a mixtureof homologue (data not shown). Moreover, it was found that twostrains displayed different surface activity, even though therewas a difference of only 1 bp in the 1466 bp of their 16S rRNAsequence (Bodour et al., 2003). This suggested that more sophisti-cated molecular approaches, such as repetitive extragenicpalindromic (REP)-PCR fingerprint should be used to further differ-entiate these strains.
Comparing the sources of each isolate, it can be seen thateleven strains belonging to nine genera were isolated from excessactivated sludge; three isolates classified into three genera werefrom oil-bearing wastewater and six isolates grouped into fourgenera were from petroleum-contaminated soils. Moreover,strains isolated from activated sludge and oil-bearing wastewatercovered Proteobacteria, Actinobacteria and Firmicutes divisionwhile strains from petroleum-polluted soils only distributed inthe former two divisions. Compared with the other two sources,strain from activated sludge showed a richer microbial diversity.This may be explained by rich composition of treated wastewa-ters, especially diverse substrates and abundant nutrients, whichprovides a favorable environment for the survival of diversemicrobes.
4. Conclusions
In this study, twenty demulsifying strains were isolated fromenvironmental sources. Five of them were highly-efficient indemulsification, showing nearly or over 90% of emulsion breakingratio. By combining the results of this study with previous findings,a phylogenic tree of demulsifying bacteria was created. It showedthat all the known demulsifying strains were classified into elevengenera and distributed among Proteobacteria, Firmicutes andActinobacteria division in Eubacteria and Ascomycota division inEucaryote. The findings of this study added six new genera to biod-emulsifier-producing bacteria. Strains isolated from activatedsludge showed a wider diversity, including nine genera of threedivisions.
Acknowledgements
The authors would like to express their gratitude to Science andTechnology Commission of Shanghai Municipality (Grant No.06DZ22007), and Science and Technology Agency of Xinjiang Uy-
gur Autonomous Region (Grant No. 200891115) for the financialsupport.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.biortech.2009.07.086.
References
Amezcua-Vega, C., Poggi-Varaldo, H.M., Esparza-Garcia, F., Rios-Leal, E., Rodriguez-Vazquez, R., 2007. Effect of culture conditions on fatty acids composition of abiosurfactant produced by Candida ingens and changes of surface tension ofculture media. Bioresource Technology 98, 237–240.
Bento, F.M., Camargo, F.A.D., Okeke, B.C., Frankenberger, W.T., 2005. Diversity ofbiosurfactant producing microorganisms isolated from soils contaminated withdiesel oil. Microbiological Research 160, 249–255.
Bizet, C., Barreau, C., Harmant, C., Nowakowski, M., Pietfroid, A., 1997. Identificationof Rhodococcus, Gordona and Dietzia species using carbon source utilization tests(‘‘Biotype-100” strips). Research in Microbiology 148, 799–809.
Bodour, A.A., Drees, K.P., Maier, R.M., 2003. Distribution of biosurfactant-producingbacteria in undisturbed and contaminated arid southwestern soils. Applied andEnvironmental Microbiology 69, 3280–3287.
Brito, E.M., Guyoneaud, R., Goni-Urriza, M., Ranchou-Peyruse, A., Verbaere, A.,Crapez, M.A.C., Wasserman, J.C.A., Duran, R., 2006. Characterization ofhydrocarbonoclastic bacterial communities from mangrove sediments inGuanabara Bay, Brazil. Research in Microbiology 157, 752–762.
Cairns, W.L., Cooper, D.G., Zajic, J.E., Wood, J.M., Kosaric, N., 1982. Characterizationof Nocardia amarae as a potent biological coalescing agent of water–oilemulsions. Applied Environmental Microbiology 43, 362–366.
Das, M., 2001. Characterization of de-emulsification capabilities of a Micrococcusspecies. Bioresource Technology 79, 15–22.
Desai, J.D., Banat, I.M., 1997. Microbial production of surfactants and theircommercial potential. Microbiology and Molecular Biology Reviews 61, 47–64.
Di Gioia, D., Michelles, A., Pierini, M., Bogialli, S., Fava, F., Barberio, C., 2008. Selectionand characterization of aerobic bacteria capable of degrading commercialmixtures of low-ethoxylated nonylphenols. Journal of Applied Microbiology104, 231–242.
Duvnjak, Z., Kosaric, N., 1987. De-emulsification of petroleum water-in-oilemulsions by selected bacterial and yeast cells. Biotechnology Letters 9, 39–42.
Franzetti, A., Bestetti, G., Caredda, P., La Colla, P., Tamburini, E., 2008. Surface-activecompounds and their role in the access to hydrocarbons in Gordonia strains.FEMS Microbiology Ecology 63, 238–248.
Gogotov, I.N., Khodakov, R.S., 2008. Surfactant production by the Rhodococcuserythropolis sH-5 bacterium grown on various carbon sources. AppliedBiochemistry and Microbiology 44, 186–191.
Goodfellow, M., Davenport, R., Stainsby, F.M., Curtis, T.P., 1996. Actinomycetediversity associated with foaming in activated sludge plants. Journal ofIndustrial Microbiology 17, 268–280.
Haferburg, D., Hommel, R., Claus, R., Kleber, H.-P., 1986. Extracellular microbiallipids as biosurfactants. In: Fiechter, A. (Ed.), Advances in BiochemicalEngineering/Biotechnology. Springer-Verlag, Berlin, Heidelberg, pp. 53–93.
Holt, J.G., Krieg, N.R., Sneath, P.H.A., Staley, J.T., Williams, S.T., 1994. Bergey’s Manualof Determinative Bacteriology, ninth ed. Lippincott, Williams & Wilkins,Philadelphia, PA 19106, USA.
Huang, X.F., Liu, J., Lu, L.J., Wen, Y., Xu, J.C., Yang, D.H., Zhou, Q., 2009. Evaluation ofscreening methods for demulsifying bacteria and characterization oflipopeptide bio-demulsifier produced by Alcaligenes sp. BioresourceTechnology 100, 1358–1365.
Ivanov, I.B., Kralchevsky, P.A., 1997. Stability of emulsions under equilibrium anddynamic conditions. Colloids and Surfaces A: Physicochemical and EngineeringAspects 128, 155–175.
Janiyani, K.L., Purohit, H.J., Shanker, R., Khanna, P., 1994. De-emulsification of oil-in-water emulsions by Bacillus-subtilis. World Journal of Microbiology andBiotechnology 10, 452–456.
Kattar, M.M., Cookson, B.T., Carlson, L.C., Stiglich, S.K., Schwartz, M.A., Nguyen, T.T.,Daza, R., Wallis, C.K., Yarfitz, S.L., Coyle, M.B., 2001. Tsukamurella strandjordaesp. nov., a proposed new species causing sepsis. Journal of Clinical Microbiology39, 1467–1476.
Lee, J.C., Lee, K.Y., 2000. Emulsification using environmental compatible emulsifiersand de-emulsification using DC field and immobilized Nocardia amarae.Biotechnology Letters 22, 1157–1163.
Li, G.H., Zhang, X., Huang, W., 2000. Characteristics of degrading microorganismdistribution in polluted soil with petroleum hydrocarbons. Huanjing Kexue 21,61–64.
Ma, T., Ling, F.-L., Xi, Y.-W., Liu, R.-L., 2006. Performance of demulsification byRhodococcus sp.PR-1. Huanjing Kexue 27, 1191–1196.
Nadarajah, N., Singh, A., Ward, O.P., 2002. Evaluation of a mixed bacterial culture forde-emulsification of water-in-petroleum oil emulsions. World Journal ofMicrobiology and Biotechnology 18, 435–440.
Nishimaki, F., Takahashi, N., Tsuchida, T., Watanabe, K., Hino, S., 1999.Microorganisms, demulsifiers and processes for breaking an emulsion. In:

X.-F. Huang et al. / Bioresource Technology 101 (2010) 317–323 323
FreePatentsOnline/PatentStorm (Ed.), US Patent Office. Tonen Corporation(Tokyo, JP), USA.
Odds, F.C., Rinaldi, M.G., Cooper, C.R., Fothergill, A., Pasarell, L., McGinnis, M.R.,1997. Candida and Torulopsis: a blinded evaluation of use of pseudohyphaformation as basis for identification of medically important yeasts. Journal ofClinical Microbiology 35, 313–316.
Park, S.H., Lee, J.H., Ko, S.H., Lee, D.S., Lee, H.K., 2000. Demulsification of oil-in-wateremulsions by aerial spores of a Streptomyces sp. Biotechnology Letters 22, 1389–1395.
Rainey, F.A., Burghardt, J., Kroppenstedt, R.M., Klatte, S., Stackebrandt, E., 1995.Phylogenetic analysis of the genera Rhodococcus and Nocardia and evidence forthe evolutionary origin of the genus Nocardia from within the radiation ofRhodococcus Species. Microbiology-UK 141, 523–528.
Ruimy, R., Boiron, P., Boivin, V., Christen, R., 1994. A Phylogeny of the genusNocardia deduced from the analysis of small-subunit ribosomal DNA-sequences, including transfer of Nocardia-amarae to the genus Gordona asGordona-amarae comb-nov. FEMS Microbiology Letters 123, 261–267.
Stewart, A.L., Gray, N.C.C., Cairns, W.L., Kosaric, N., 1983. Bacteria-induced de-emulsification of water-in-oil petroleum emulsions. Biotechnology Letters 5,725–730.
Ta-Chen, L., Chang, J.S., Young, C.C., 2008. Exopolysaccharides produced by Gordoniaalkanivorans enhance bacterial degradation activity for diesel. BiotechnologyLetters 30, 1201–1206.
Thavasi, R., Jayalakshmi, S., Balasubramanian, T., Banat, I.M., 2008. Production andcharacterization of a glycolipid biosurfactant from Bacillus megaterium usingeconomically cheaper sources. World Journal of Microbiology andBiotechnology 24, 917–925.
Toledo, F.L., Gonzalez-Lopez, J., Calvo, C., 2008. Production of bioemulsifier byBacillus subtilis, Alcaligenes faecalis and Enterobacter species in liquid culture.Bioresource Technology 99, 8470–8475.
Tuleva, B., Christova, N., Cohen, R., Stoev, G., Stoineva, I., 2008. Production andstructural elucidation of trehalose tetraesters (biosurfactants) from a novelalkanothrophic Rhodococcus wratislaviensis strain. Journal of AppliedMicrobiology 104, 1703–1710.
von der Weid, I., Marques, J.M., Cunha, C.D., Lippi, R.K., dos Santos, S.C.C., Rosado,A.S., Lins, U., Seldin, L., 2007. Identification and biodegradation potential of anovel strain of Dietzia cinnamea isolated from a petroleum-contaminatedtropical soil. Systematic and Applied Microbiology 30, 331–339.
Wilkinson, M.A., Cooper, D.G., 1985. Testing of microbial demulsifiers with heavycrude emulsions. Biotechnology Letters 7, 406–408.




















