
Prevention of acute graft-versus-host disease by humanized anti-CD26 monoclonal antibody
15
Prevention of acute graft-versus-host disease by humanized anti-CD26 monoclonal antibody Ryo Hatano, 1 Kei Ohnuma, 1 Junpei Yamamoto, 1 Nam H. Dang, 2 Taketo Yamada 3 and Chikao Morimoto 1 1 Department of Therapy Development and Inno- vation for Immune disorders and Cancers, Grad- uate School of Medicine, Juntendo University, Tokyo, Japan, 2 Division of Hematology/Oncology, University of Florida, Gainesville, FL, USA, and 3 Department of Pathology, Keio University School of Medicine, Tokyo, Japan Received 22 January 2013; accepted for publication 3 April 2013 Correspondence: Dr Kei Ohnuma, Department of Therapy Development and Innovation for Immune disorders and Cancers, Graduate School of Medicine, Juntendo University, 2-1-1, Hongo, Bunkyo-ku, Tokyo 113-8421, Japan. E-mail: [email protected] Summary CD26 (DPP4) is a T cell costimulatory molecule as well as T cell activation marker, and CD26 + T cells are accumulated in inflamed tissues, such as rheumatoid synovitis and autoimmune thyroiditis. In the present study, we found accumulation of CD26 + T cells in graft-versus-host disease (GVHD) target organs. To expand our in vitro findings to an in vivo system, we examined CD26-dependent organ injury in a xenogeneic GVHD (x-GVHD) murine model. Following intraperitoneal injection of human peripheral blood mononuclear cells into non-obese diabetic severe combined immu- nodeficiency/c c / mice (hu-PBL-NOG mice), the mice exhibited the onset of GVHD symptoms associated with the presence of CD26 high human lym- phocytes in the peripheral blood and GVHD target tissues. Administration of humanized anti-human CD26 monoclonal antibody (mAb) decreased x-GVHD severity and prolonged survival in hu-PBL-NOG mice without loss of engraftment of human T cells, while increasing doses of CTLA4- immunoglobulin fusion protein diminished engraftment of human lymphocytes. Importantly, anti-CD26 mAb treatment preserved the graft- versus-leukaemia effects in studies using cotransplantation of P815 murine leukaemic cells. In addition, CD26 + lymphocytes infiltrated the GVHD patients’ target tissues. Altogether, our data indicate a role for CD26 in the regulation of GVHD and point to CD26 as a novel target for therapeutic intervention in this disease. Keywords: CD26 (DPP4), graft-versus-host disease, T cell costimulation, haematopoietic stem cell transplantation. Graft-versus-host disease (GVHD) is a severe complication and major cause of morbidity and mortality following alloge- neic haematopoietic stem cell transplantation (alloHSCT) (Pasquini et al, 2010; Giralt, 2012). GVHD results from an immunobiological attack on target recipient organs by donor allogeneic T cells that are transferred along with the allograft (Shlomchik, 2007; Blazar et al, 2012). The pivotal role of donor-derived T cells in acute GVHD (aGVHD) is supported by the complete abrogation of GVHD following T cell deple- tion from the graft (Giralt, 2012; Tsirigotis et al, 2012). This approach remains the most effective in preventing aGVHD. However, complications such as graft rejection, loss of graft- versus-leukaemia (GVL) effect or increased opportunistic infections can occur. Although new approaches are being developed in the clinical setting for the prevention or treatment of GVHD (Blazar et al, 2012), in-depth under- standing of the precise cellular mechanisms of human GVHD is necessary for more effective and less immunocompromis- ing strategies to improve the clinical outcome of alloHSCT. The important role of costimulatory pathways in trans- plant biology has been established (Briones et al, 2011). The most extensively studied pathways involve interactions between CD28 and the B7 molecules CD80 and CD86 (Rudd et al, 2009). Initial studies focused on the in vivo blockade of interactions between CD28 or cytotoxic T lymphocyte anti- gen 4 (CTLA4) and their B7 ligands, CD80 and CD86, using a CTLA4-immunoglobulin fusion protein (CTLA4-Ig) or B7-specific antibodies (Lenschow et al, 1992; Saito, 1998). While CD28 is widely expressed on T cells, the CD28 CD8 + T cell population is a well-recognized subset of na € ıve or ª 2013 John Wiley & Sons Ltd First published online 21 May 2013 British Journal of Haematology, 2013, 162, 263–277 doi:10.1111/bjh.12378 research paper
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Prevention of acute graft-versus-host disease by humanized anti-CD26 monoclonal antibody

Prevention of acute graft-versus-host disease by humanizedanti-CD26 monoclonal antibody
Ryo Hatano,1 Kei Ohnuma,1
Junpei Yamamoto,1 Nam H. Dang,2
Taketo Yamada3 and Chikao Morimoto1
1Department of Therapy Development and Inno-
vation for Immune disorders and Cancers, Grad-
uate School of Medicine, Juntendo University,
Tokyo, Japan, 2Division of Hematology/Oncology,
University of Florida, Gainesville, FL, USA, and3Department of Pathology, Keio University
School of Medicine, Tokyo, Japan
Received 22 January 2013; accepted for
publication 3 April 2013
Correspondence: Dr Kei Ohnuma, Department
of Therapy Development and Innovation for
Immune disorders and Cancers, Graduate
School of Medicine, Juntendo University,
2-1-1, Hongo, Bunkyo-ku, Tokyo 113-8421,
Japan.
E-mail: [email protected]
Summary
CD26 (DPP4) is a T cell costimulatory molecule as well as T cell activation
marker, and CD26+ T cells are accumulated in inflamed tissues, such as
rheumatoid synovitis and autoimmune thyroiditis. In the present study, we
found accumulation of CD26+ T cells in graft-versus-host disease (GVHD)
target organs. To expand our in vitro findings to an in vivo system, we
examined CD26-dependent organ injury in a xenogeneic GVHD (x-GVHD)
murine model. Following intraperitoneal injection of human peripheral
blood mononuclear cells into non-obese diabetic severe combined immu-
nodeficiency/cc�/� mice (hu-PBL-NOG mice), the mice exhibited the onset
of GVHD symptoms associated with the presence of CD26high human lym-
phocytes in the peripheral blood and GVHD target tissues. Administration
of humanized anti-human CD26 monoclonal antibody (mAb) decreased
x-GVHD severity and prolonged survival in hu-PBL-NOG mice without
loss of engraftment of human T cells, while increasing doses of
CTLA4- immunoglobulin fusion protein diminished engraftment of human
lymphocytes. Importantly, anti-CD26 mAb treatment preserved the graft-
versus-leukaemia effects in studies using cotransplantation of P815 murine
leukaemic cells. In addition, CD26+ lymphocytes infiltrated the GVHD
patients’ target tissues. Altogether, our data indicate a role for CD26 in the
regulation of GVHD and point to CD26 as a novel target for therapeutic
intervention in this disease.
Keywords: CD26 (DPP4), graft-versus-host disease, T cell costimulation,
haematopoietic stem cell transplantation.
Graft-versus-host disease (GVHD) is a severe complication
and major cause of morbidity and mortality following alloge-
neic haematopoietic stem cell transplantation (alloHSCT)
(Pasquini et al, 2010; Giralt, 2012). GVHD results from an
immunobiological attack on target recipient organs by donor
allogeneic T cells that are transferred along with the allograft
(Shlomchik, 2007; Blazar et al, 2012). The pivotal role of
donor-derived T cells in acute GVHD (aGVHD) is supported
by the complete abrogation of GVHD following T cell deple-
tion from the graft (Giralt, 2012; Tsirigotis et al, 2012). This
approach remains the most effective in preventing aGVHD.
However, complications such as graft rejection, loss of graft-
versus-leukaemia (GVL) effect or increased opportunistic
infections can occur. Although new approaches are being
developed in the clinical setting for the prevention or
treatment of GVHD (Blazar et al, 2012), in-depth under-
standing of the precise cellular mechanisms of human GVHD
is necessary for more effective and less immunocompromis-
ing strategies to improve the clinical outcome of alloHSCT.
The important role of costimulatory pathways in trans-
plant biology has been established (Briones et al, 2011). The
most extensively studied pathways involve interactions
between CD28 and the B7 molecules CD80 and CD86 (Rudd
et al, 2009). Initial studies focused on the in vivo blockade of
interactions between CD28 or cytotoxic T lymphocyte anti-
gen 4 (CTLA4) and their B7 ligands, CD80 and CD86, using
a CTLA4-immunoglobulin fusion protein (CTLA4-Ig) or
B7-specific antibodies (Lenschow et al, 1992; Saito, 1998).
While CD28 is widely expressed on T cells, the CD28�CD8+
T cell population is a well-recognized subset of na€ıve or
ª 2013 John Wiley & Sons Ltd First published online 21 May 2013British Journal of Haematology, 2013, 162, 263–277 doi:10.1111/bjh.12378
research paper

highly-antigen experienced (late-differentiated) cells (Rudd
et al, 2009; Strioga et al, 2011). Meanwhile, blockade of
CD28 pathways may lead to profound immunosuppression
(Blazar et al, 1997).
CD26 (DPP4) is also associated with T cell signal trans-
duction processes as a costimulatory molecule, as well as
being a marker of T cell activation (Fox et al, 1984; Morim-
oto et al, 1989; Dang et al, 1990; Tanaka et al, 1992; Morim-
oto & Schlossman, 1998; Ohnuma et al, 2008). In fact,
patients with autoimmune diseases, such as multiple sclero-
sis, Grave disease, and rheumatoid arthritis, have been found
to have increased numbers of CD4+CD26+ T cells in
inflamed tissues as well as in their peripheral blood (Hafler
et al, 1985; Eguchi et al, 1989; Gerli et al, 1996; Mizokami
et al, 1996), with enhancement of CD26 expression in these
autoimmune diseases correlating with disease severity
(Eguchi et al, 1989; Muscat et al, 1994; Gerli et al, 1996). In
addition, we previously demonstrated that T cells migrating
through endothelial cell monolayers in vitro express
high levels of CD26 (Masuyama et al, 1992). These findings
imply that CD26+ T cells play an important role in the
inflammation process and subsequent tissue damage in such
diseases, and suggest that CD26+ T cells appear to be effector
T cells. However, little is known about the effectiveness of
CD26-targeting therapy on aGVHD.
In the current study, we utilized non-obese diabetic severe
combined immunodeficiency (NOD/SCID)/cc�/� (NOG)
mice inoculated with human peripheral lymphocytes devel-
oped xenogeneic GVHD (x-GVHD) by human lymphocytes
(Ito et al, 2002). We demonstrated that donor-derived
human lymphocytes expressed high level of human CD26 in
the peripheral blood of hu-PBL-NOG mice developing
x-GVHD, and human CD26+ lymphocyte levels were
enriched in the liver of mice undergoing x-GVHD. We dem-
onstrated that a humanized anti-CD26 monoclonal antibody
(mAb) was effective against x-GVHD-associated pathology
and mortality of hu-PBL-NOG mice. As compared to the
widely used costimulation blocking agent CTLA4-Ig, both
increasing doses of anti-CD26 mAb and CTLA4-Ig clearly
prolonged survival of hu-PBL-NOG mice with less manifes-
tation of x-GVHD physical symptoms. However, increasing
doses of CTLA4-Ig decreased engraftment of human lym-
phocytes and lowered proliferation of human CD4+ T cells
in hu-PBL-NOG mice, while such immune impairment after
inoculation was not observed in hu-PBL-NOG mice treated
with increasing doses of anti-CD26 mAb. More importantly,
anti-CD26 mAb preserved the GVL effect in hu-PBL-NOG
transplanted with tumour cells. Finally, we found that
human CD26+ lymphocytes infiltrated GVHD target organs
obtained from patients with aGVHD. These results provide
proof of principle that human CD26+ T cells are involved in
the pathophysiology of GVHD, including infiltration of
GVHD target organs, and that humanized anti-CD26 mAb
appears to be a promising novel therapy for the clinical
control of GVHD.
Materials and methods
Preparation of human T cells
Human peripheral blood mononuclear cells (PBMCs) were
collected from healthy adult volunteers after the documented
informed consent and institutional review board approval
were obtained. Using the magnetic-activated cell sorting
(MACS) cell separation system (Miltenyi, Bergisch Gladbach,
Germany), CD3+, CD4+ or CD8+ T cells were purified from
PBMC. The purity of CD3+, CD4+ or CD8+ T cells were
� 99%, � 97% or � 95%, respectively, as confirmed by flow
cytometry.
Flow cytometry and antibodies
Human-specific antibodies used for flow cytometry: fluores-
cein isothiocyanate (FITC)-labelled CD8 (clone HIT8a), FITC-
labelled CD14 (clone M5E2), FITC-labelled CD19 (clone
HIB19), phycoerythrin (PE)-labelled CD16 (clone 3G8), PE-
labelled CD56 (clone B159), peridinin chlorophyll- cyanin 5�5(PerCP-Cy5�5)-labelled CD4 (clone SK3), allophycocyanin
(APC)-labelled CD3 (clone UCHT1), APC-labelled CD8
(clone RPA-T8), and APC-labelled CD45 (clone HI30) were
purchased from BD Biosciences (San Jose, CA, USA). PE-
labelled CD26 antibody (clone 5K78), which recognizes
human CD26 even in the presence of humanized anti-CD26
mAb, was purchased from Santa Cruz Biotechnology (Dallas,
TX, USA) (Fig S1). Humanized anti-CD26 mAb was gener-
ously provided by Y’s Therapeutics (Tokyo, Japan). The
humanized anti-CD26 mAb employed in this study was gener-
ated by utilizing the complementarity determining regions of
murine anti-human CD26 mAb, 14D10, which has no crossre-
activity to murine CD26 (Dong et al, 1998). CTLA4-Ig (abata-
cept) and human polyclonal IgG (venilon-I) were purchased
from Alfresa Corporation (Tokyo, Japan). This CTLA4-Ig is
composed of the Fc region of human immunoglobulin IgG1
fused to the extracellular domain of human CTLA4, which also
reacts to murine CD80/86 molecules (Lenschow et al, 1992),
and is able to block interaction between human T cells and
murine antigen presenting cells in mice. For detection of cellu-
lar proliferation in x-GVHD mice, human PBMC (7 9 106/
body), labelled with 4 lmol/l of carboxy fluorescein diacetate
succinimidyl ester (CFSE; Molecular Probes, Carlsbad, CA,
USA), were inoculated peritoneally into NOG mice. Spleen
cells from mice were harvested on day 7, acquisition was per-
formed using FACSCalibur and data were analysed using a
FLOWJO software (TreeStar, Ashland, OR, USA).
Mice and x-GVHD experiments
NOD/SCID/cc�/� NOD.Cg-Prkdcscid Il2rgtm1Sug/Jic mice
(NOG mice) were purchased from The Central Institute
for Experimental Animals (Kawasaki, Japan) and housed in a
specific pathogen-free facility in micro-isolator cages. Mice
R. Hatano et al
264 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277

were used at 8–12 weeks. The animal protocol was approved
by Institutional Animal Care and Use Committee.
In the in vivo x-GVHD model, NOG mice were injected
intraperitoneally with human PBMC (10 9 106). Mice were
assessed for survival daily and weighed thrice weekly. In
GVHD prevention experiments, after 1 day of inoculation
of human cells, mice were injected intraperitoneally with
humanized anti-CD26 mAb, CTLA4-Ig, or control human
IgG at indicated doses in 200 ll of sterile phosphate-
buffered saline (PBS), and then were injected thrice weekly
for a total of 10 doses. To document human lymphocyte-
associated peripheral T cell expansion, mice were bled (10–
40 ll), red blood cells lysed, and PBMC subsets were
enumerated by flow cytometry. In addition, sera were
collected on day 21, serum alanine transaminase (ALT)
activity was assayed using an ALT Activity Kit (BioVision,
Milpitas, CA, USA). In GVL experiments, NOG mice
received H2-Kd MHC background P815 mastocytoma cells
(2 9 104) on the day of transplantation in addition to
human PBMCs. Tumour volumes were measured along
three orthogonal areas (a, b, and c) by digital vernier
calipers (Mitutoyo Corporation, Kanagawa, Japan), and
calculated as tumour volume = abc/2 (mm3). Treatments
with humanized anti-CD26 mAb, CTLA4-Ig, or control
human IgG were conducted as described above. Recipients
were monitored for tumour-related morbidity and mortality,
and death with leukaemia was determined if autopsy
identified hepatosplenomegaly, or if histopathology showed
leukaemic cell infiltration in spleen or liver.
Immunohistochemistry (IHC) staining and pathologydata
CD26-expressing cells were quantified in adult patients after
undergoing alloHSCT for acute myeloid leukaemia (AML)
at the Keio University Hospital. All patients had biopsies
performed as diagnostic procedures for dermal or digestive
GVHD symptoms. Clinical grading of acute GVHD was
scored using the Consensus scoring system (Przepiorka
et al, 1995), and tissue specimens were scored using histo-
pathological grading of skin or gastrointestinal (GI) GVHD
(Sale et al, 1977; Washington & Jagasia, 2009). This study
was approved by the Keio University Hospital ethical
review board and the permission was obtained from
patients. Human tissues were fixed in 10% formalin con-
taining PBS, and embedded in paraffin. Serial 5-lm sec-
tions from each tissue were stained with haematoxylin and
eosin (H&E). IHC staining for CD3, CD4, CD8 or CD26
was performed as described previously (Aoe et al, 2012).
Histopathological findings were evaluated and verified inde-
pendently by two pathologists. Images were captured with
an Olympus digital camera DP21 attached to an Olympus
BX43 microscope using CELLSENS software (Olympus, Tokyo,
Japan). The number of CD26+ MNC was counted and pre-
sented as per square millimetre.
To identify human CD26-expressing cells in x-GVHD
mice, specimens from skin, colon, spleen and liver of NOG
mice transplanted with or without human PBMC were fixed
in 10% formalin containing PBS, and embedded in paraffin.
Serial 5-lm sections from each tissue were stained with
H&E. Images were captured as described above. Slides were
analysed by two independent pathologists who were blinded
to the pathological results when quoting the intensity of the
staining and number of MNCs stained.
Quantitative real-time reverse transcription polymerasechain reaction (qRT-PCR) assay
In experiments assessing expression of mRNA of effector mole-
cules, human CD4+ or CD8+ T cells were purified from human
PBMCs, spleen or liver of hu-PBL-NOG mice, using human
CD4 or CD8 microbeads (Miltenyi). Quantification of mRNA
was performed as described previously (Hatano et al, 2013).
Expression levels of mRNA were calculated on the basis of stan-
dard curves generated for each gene and hypoxanthine phos-
phoribosyltransferase 1 (HPRT1) mRNA was used as an
invariant control. Sequences of primers used in qRT-PCR anal-
ysis are shown in the Table SI.
Statistics
Data were analysed by the analysis of variance (ANOVA) test
for multiple comparison test or Student’s t test for two
group comparison. P values � 0�05 were considered statisti-
cally significant. Survival rates were analysed by log-rank test
using Kaplan–Meier method.
Study approval
Human study protocols were approved by the Ethics Commit-
tees at Juntendo University, at the Institute of Medical Science,
University of Tokyo, and at Keio University. Informed consent
was obtained from all patients and healthy volunteers. All
studies on human subjects were carried out according to the
principles set out in the Declaration of Helsinki. Animal experi-
ments were conducted following protocols approved by the
Animal Care and Use Committees at Juntendo University and
at the Institute of Medical Science, University of Tokyo.
Results
Infiltration of human CD26+ T cells in the liver GI tractand skin of murine recipients with x-GVHD
To determine whether human CD26+ T cells play a role in the
animal model of inflammatory diseases by human effector
lymphocytes, we employed a xenogeneic model of GVHD
resulting from the adoptive transfer of human PBMCs into
NOG mice (hu-PBL-NOG mice) (Ito et al, 2002). We first
conducted a pathological analysis of x-GVHD target organs,
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 265British Journal of Haematology, 2013, 162, 263–277

such as the skin, colon or liver, in hu-PBL-NOG mice.
Control mice without x-GVHD (treated with PBS alone)
showed no GVHD pathology in the liver, GI tract or skin
(Fig 1A), while the liver, colon and skin of hu-PBL-NOG
mice were infiltrated with human CD3+ mononuclear cells
(MNCs), with associated organ destruction (Fig 1B, panels a,
b, d, e, g, h). Moreover, human CD3+ MNCs reactive to anti-
human CD26 Ab were readily visible in all evaluated samples
in the liver, colon or skin of x-GVHD mice (Fig 1B, panels c,
f, i). The infiltrated CD26+ cells were confirmed to be human
CD3+ T cells by co-staining analysis with flow cytometry (Fig.
S2). In contrast to the hu-PBL-NOG mice, human CD3+ or
CD26+ MNCs were not detected in all evaluated samples of
the liver, colon, or skin of NOG mice without x-GVHD
(Fig 1A, panels b, c, e, f, h, j). These results suggest that
donor-derived human CD26+ cells play a role in the patho-
genesis of x-GVHD in our hu-PBL-NOG murine model.
Humanized anti-CD26 mAb treatment significantlyreduces x-GVHD-associated lethality and weight loss inmurine model
As shown above, CD26+ lymphocytes might function as effec-
tor cells and be an appropriate target of x-GVHD treatment.
(A)
(B)
Non-x-GVHD (PBS alone)
x-GVHD (human PBMC inoculation)
H&E anti-human CD3Staining :
H&E anti-human CD3
Liver
Colon
Skin
anti-human CD26
anti-human CD26Staining :
Liver
Colon
Skin
(c)(b)(a)
(f)(e)(d)
(i)(h)(g)
(c)(b)(a)
(f)(e)(d)
(i)(h)(g)
Fig 1. NOD/SCID/cc�/� mice receiving human
peripheral blood mononuclear cells (hu-PBL-
NOG mice) develop manifestation of x-GVHD
in target organs. (A) Specimens were obtained
from control NOG mice [non-x-GVHD, phos-
phate-buffered saline (PBS) alone]. Representa-
tive serial sections of liver (panels a–c), colon(panels d–f) or skin (panels g–i) were stainedwith haematoxylin and eosin (H&E), with
anti-human CD3 polyclonal antibody (pAb),
or with anti-human CD26 pAb, counterstained
with haematoxylin (original magnification,
2009). Non-specific staining with anti-CD3
pAb was observed in a part of colonic mucosal
epithelia (panel e) and epidermal basal area
(panel h). Staining with anti-CD26 pAb was
observed in bile canaliculi of the liver (panel c),
colonic mucosal epithelia (panel f) and
epidermal basal layer (panel i), due to the cross-
reactivity of goat anti-human CD26 pAb against
to murine CD26 antigen. Similar results were
observed in independent experiments using
mice receiving PBS alone (n = 13). (B) The
specimens were obtained from hu-PBL-NOG
mice (x-GVHD, 4 weeks after inoculation of
human PBMCs). Representative serial sections
were stained by the same methods as in (A)
(original magnification, 2009). Human
CD3+CD26+ lymphocytes infiltrated the peri-
portal area of the liver (panels a–c), the crypticabscesses of the colon (panels d–f), and the
upper dermis with subepidermal clefting (panels
g–i). Similar results were observed in indepen-
dent experiments using mice receiving human
PBMCs (n = 20).
R. Hatano et al
266 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277
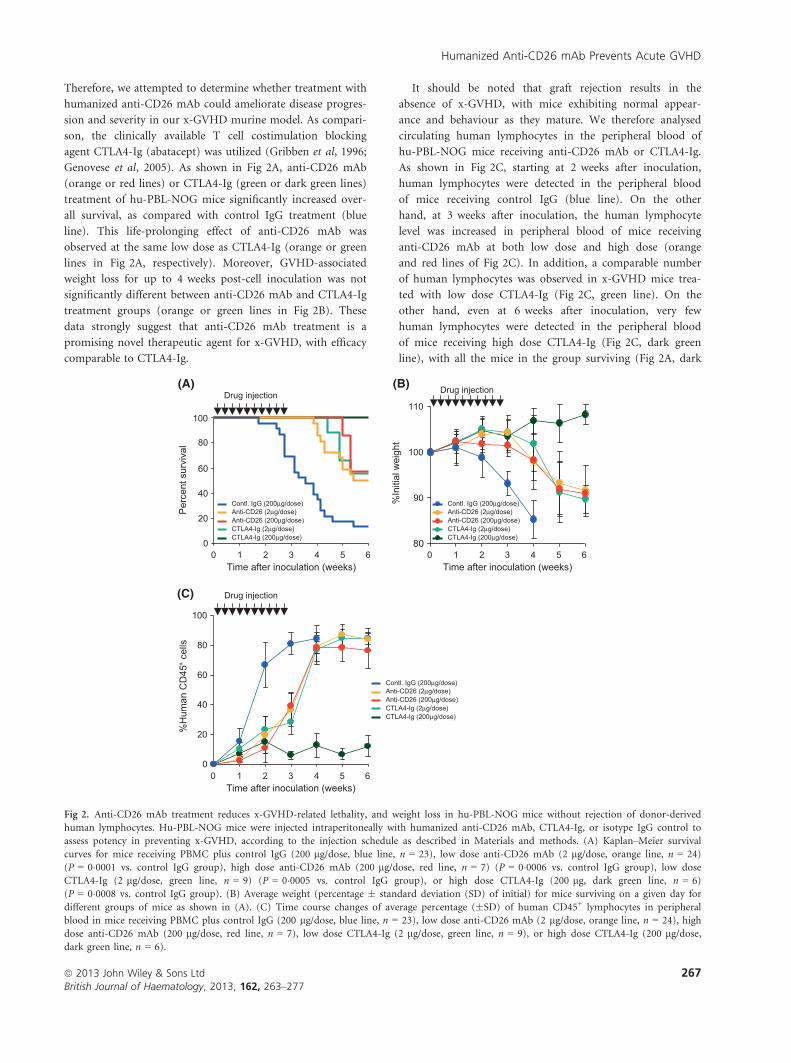
Therefore, we attempted to determine whether treatment with
humanized anti-CD26 mAb could ameliorate disease progres-
sion and severity in our x-GVHD murine model. As compari-
son, the clinically available T cell costimulation blocking
agent CTLA4-Ig (abatacept) was utilized (Gribben et al, 1996;
Genovese et al, 2005). As shown in Fig 2A, anti-CD26 mAb
(orange or red lines) or CTLA4-Ig (green or dark green lines)
treatment of hu-PBL-NOG mice significantly increased over-
all survival, as compared with control IgG treatment (blue
line). This life-prolonging effect of anti-CD26 mAb was
observed at the same low dose as CTLA4-Ig (orange or green
lines in Fig 2A, respectively). Moreover, GVHD-associated
weight loss for up to 4 weeks post-cell inoculation was not
significantly different between anti-CD26 mAb and CTLA4-Ig
treatment groups (orange or green lines in Fig 2B). These
data strongly suggest that anti-CD26 mAb treatment is a
promising novel therapeutic agent for x-GVHD, with efficacy
comparable to CTLA4-Ig.
It should be noted that graft rejection results in the
absence of x-GVHD, with mice exhibiting normal appear-
ance and behaviour as they mature. We therefore analysed
circulating human lymphocytes in the peripheral blood of
hu-PBL-NOG mice receiving anti-CD26 mAb or CTLA4-Ig.
As shown in Fig 2C, starting at 2 weeks after inoculation,
human lymphocytes were detected in the peripheral blood
of mice receiving control IgG (blue line). On the other
hand, at 3 weeks after inoculation, the human lymphocyte
level was increased in peripheral blood of mice receiving
anti-CD26 mAb at both low dose and high dose (orange
and red lines of Fig 2C). In addition, a comparable number
of human lymphocytes was observed in x-GVHD mice trea-
ted with low dose CTLA4-Ig (Fig 2C, green line). On the
other hand, even at 6 weeks after inoculation, very few
human lymphocytes were detected in the peripheral blood
of mice receiving high dose CTLA4-Ig (Fig 2C, dark green
line), with all the mice in the group surviving (Fig 2A, dark
Per
cent
sur
viva
l
(C)
%H
uman
CD
45+
cells
%In
itial
wei
ght
(B)
Time after inoculation (weeks)
Drug injection
Contl. IgG (200µg/dose)
CTLA4-Ig (200µg/dose)CTLA4-Ig (2µg/dose)Anti-CD26 (200µg/dose)Anti-CD26 (2µg/dose)
0
20
40
60
80
100
(A)
80
90
100
110
Time after inoculation (weeks)
Drug injection
Contl. IgG (200µg/dose)
CTLA4-Ig (200µg/dose)CTLA4-Ig (2µg/dose)Anti-CD26 (200µg/dose)Anti-CD26 (2µg/dose)
Contl. IgG (200µg/dose)
CTLA4-Ig (200µg/dose)CTLA4-Ig (2µg/dose)Anti-CD26 (200µg/dose)Anti-CD26 (2µg/dose)
0
20
40
60
80
100
0 1 2 3 4 5 6 0 1 2 3 4 5 6
0 1 2 3 4 5 6Time after inoculation (weeks)
Drug injection
Fig 2. Anti-CD26 mAb treatment reduces x-GVHD-related lethality, and weight loss in hu-PBL-NOG mice without rejection of donor-derived
human lymphocytes. Hu-PBL-NOG mice were injected intraperitoneally with humanized anti-CD26 mAb, CTLA4-Ig, or isotype IgG control to
assess potency in preventing x-GVHD, according to the injection schedule as described in Materials and methods. (A) Kaplan–Meier survival
curves for mice receiving PBMC plus control IgG (200 lg/dose, blue line, n = 23), low dose anti-CD26 mAb (2 lg/dose, orange line, n = 24)
(P = 0�0001 vs. control IgG group), high dose anti-CD26 mAb (200 lg/dose, red line, n = 7) (P = 0�0006 vs. control IgG group), low dose
CTLA4-Ig (2 lg/dose, green line, n = 9) (P = 0�0005 vs. control IgG group), or high dose CTLA4-Ig (200 lg, dark green line, n = 6)
(P = 0�0008 vs. control IgG group). (B) Average weight (percentage � standard deviation (SD) of initial) for mice surviving on a given day for
different groups of mice as shown in (A). (C) Time course changes of average percentage (�SD) of human CD45+ lymphocytes in peripheral
blood in mice receiving PBMC plus control IgG (200 lg/dose, blue line, n = 23), low dose anti-CD26 mAb (2 lg/dose, orange line, n = 24), high
dose anti-CD26 mAb (200 lg/dose, red line, n = 7), low dose CTLA4-Ig (2 lg/dose, green line, n = 9), or high dose CTLA4-Ig (200 lg/dose,dark green line, n = 6).
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 267British Journal of Haematology, 2013, 162, 263–277

green line) and showing no weight loss (Fig 2B, dark green
line). Therefore, in mice receiving high dose CTLA4-Ig,
most of the inoculated human PBMCs were rejected, which
is one of the main reasons all of the mice in this cohort
survived without developing x-GVHD. Taken together, these
data indicate that anti-CD26 mAb treatment for x-GVHD
was comparable with CTLA4-Ig treatment. Moreover, while
treatment with increasing doses of CTLA4-Ig resulted in the
absence of x-GVHD development, graft rejection did occur,
potentially analogous to findings in a clinical trial with a
new CTLA4-Ig agent, belatacept (Vincenti et al, 2010). On
the other hand, increasing doses of anti-CD26 mAb resulted
in no graft rejection in hu-PBL-NOG mice with prolonging
survival as compared to hu-PBL-NOG mice receiving
control IgG. Given that the effectiveness of anti-CD26 mAb
on x-GVHD was observed equally at low or high dose of
therapy, we conducted subsequent experiments at the 2 lg/dose level.
Anti-CD26 mAb treatment decreases level of CD26high
lymphocytes in x-GVHD mice while preserving CD4+
T cell proliferation
We next examined human CD26 expression level in the
peripheral blood and spleen of hu-PBL-NOG mice receiving
anti-CD26 mAb or CTLA4-Ig. To avoid antigen masking by
crossreaction with humanized anti-CD26 mAb, we used an
anti-human CD26 mAb recognizing a separate epitope
distinct from the humanized anti-CD26 mAb, as described in
Materials and methods. As shown in Fig 3A, higher expres-
sion level of human CD26 on both CD4+ and CD8+ human
lymphocytes in the peripheral blood of hu-PBL-NOG mice
receiving control IgG (panels a and b, blue lines) or CTLA4-
Ig (panels a and b, green lines) was observed from 1 to
3 weeks after inoculation. On the other hand, in
hu-PBL-NOG mice receiving anti-CD26 mAb, neither CD4+
nor CD8+ T cells expressed CD26 from 1 to 3 weeks (Fig 3A,
panels a and b, red lines). At 3 weeks after inoculation, very
weak expression of human CD26 was observed on CD4+ or
CD8+ cells in the peripheral blood of hu-PBL-NOG mice
receiving anti-CD26 mAb (Fig 3A, panels a and b, red lines).
These findings suggest that circulation of donor-derived
human CD26high lymphocytes might be associated with
x-GVHD development in hu-PBL-NOG mice. To examine
this issue in more detail, we analysed human CD26 expres-
sion level on mouse spleen cells. CD26highCD4+ or
CD26highCD8+ human lymphocytes were predominantly
observed in the spleen of hu-PBL-NOG mice receiving con-
trol IgG (Fig 3B, panels a and b, blue lines) or CTLA4-Ig
(Fig 3B, panels a and b, green lines) at 2 weeks after inocula-
tion. On the other hand, CD26dullCD4+ or CD26dullCD8+
human lymphocytes were predominantly observed in the
spleen of hu-PBL-NOG mice receiving anti-CD26 mAb
(Fig 3B, panels a and b, red lines) at 2 weeks after inocula-
tion. Taken together, these data suggest that the decreased
number of CD26high effector T cells may be responsible for
the relative absence of x-GVHD development in mice receiv-
ing anti-CD26 mAb.
Given that T cell activation is strongly associated with
CD26high lymphocytes (Morimoto et al, 1989), we next
addressed issues regarding the immunological status of
CD26dull lymphocytes in hu-PBL-NOG mice receiving
anti-CD26 mAb. To further characterize the activity of donor-
derived human lymphocytes in hu-PBL-NOG mice, we per-
formed cell proliferation assays. For this purpose, NOG mice
were inoculated with CFSE-labelled human PBMCs, and
treated with control IgG, anti-CD26 mAb or CTLA4-Ig, and
spleens were harvested on day 7. The isolated splenocytes were
gated for human CD4+ or CD8+ cells, and CFSE intensity was
analysed for cell division as a reflection of proliferative activity.
The human CD4+ or CD8+ cell population in the spleen of
hu-PBL-NOG mice receiving control IgG contained increased
numbers of multiply dividing cells (Fig 3C, blue bars). These
data indicate that inoculated human lymphocytes proliferated
extensively in mice developing x-GVHD. On the other hand,
the proliferative activity of human CD4+ or CD8+ cells was
significantly decreased in hu-PBL-NOG mice receiving anti-
CD26 mAb or CTLA4-Ig, in contrast to control IgG (Fig 3C,
*, **). However, compared to hu-PBL-NOG mice receiving
CTLA4-Ig, human CD4+ cell proliferation was greater in
hu-PBL-NOG mice receiving anti-CD26-mAb (Fig 3C, ***),while proliferation of human CD8+ cells was equally
suppressed [Fig 3C, not significant (NS)]. Taken together with
the data above, these findings indicate that anti-CD26 mAb
had a therapeutic effect on x-GVHD-associated manifestations
that was comparable to CTLA4-Ig, and that a decreased level
of CD26high donor cells after inoculation may be associated
with a reduction in x-GVHD severity. In addition, as CD4+
cell proliferative activity in hu-PBL-NOG mice receiving
anti-CD26 mAb was greater than that in hu-PBL-NOG mice
receiving CTLA4-Ig, treatment with anti-CD26 mAb might
avoid unwanted immunosuppressive effects, such as loss of
GVL effect or early graft rejection.
Humanized anti-CD26 mAb treatment ameliorates tissuedamage and decreased production of proinflammatorycytokines in the liver of hu-PBL-NOG mice with GVHD
Given that CD26high effector lymphocytes have high capacity
for migration into inflamed tissues (Hafler et al, 1985;
Eguchi et al, 1989; Masuyama et al, 1992; Gerli et al, 1996),
we examined GVHD target organs of hu-PBL-NOG mice
for donor lymphocyte infiltration. As patients with aGVHD
can have life-threatening damage to the liver or GI tract
(Ferrara et al, 2009), our study focussed on the examination
of liver tissues of hu-PBL-NOG mice. In x-GVHD mice
receiving control IgG, liver damage was observed with infil-
tration of human CD4+ or CD8+ MNCs (Fig 4A, panels b,
f, j), which were not found in the liver of control NOG
mice receiving PBS alone (panels a, e, i of Fig 4A).
R. Hatano et al
268 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277

Moreover, slight inflammation of the portal duct areas in
the liver was observed in mice receiving CTLA4-Ig (Fig 4A,
panels d, h, l). On the other hand, infiltration of human T
cells in the liver was barely detected in mice receiving
anti-CD26 mAb (Fig 4A panels c, g, k). In addition to the
pathological changes seen in the liver, serum ALT activity
was examined as another method of detecting GVHD-asso-
ciated liver damage. Significant elevation of serum ALT
activity was observed in mice receiving control IgG or
CTLA4-Ig, while that of mice receiving anti-CD26 mAb was
found to be near normal levels (Fig 4B, *). These data
suggest that liver damage due to GVHD was suppressed in
(A)
MFI
of C
D26
(a) Human CD4+ T cells in PB (b) Human CD8+ T cells in PB
Human CD26
+ T cells in SP (B)
Rel
ativ
e ce
ll nu
mbe
r
Contl. IgG
CTLA4-Ig
Isotype contl.Human PBMC
Splenocytesin hu-PBL-NOG mice receiving:
(a) Human CD4 (b) Human CD8+ T cells in SP
(C)
Perc
enta
ge o
f CFS
E-ne
gativ
e (p
rolif
erat
ed) c
ells
Contl. IgGAnti-CD26CTLA4-Ig
Treatment:
Pre-inoculation cells:
CD4+ T cells CD8+ T cells0
20
40
60
80
100
Human T cells in spleen
NS
0
50
100
150
200
250
300
Time after inoculation (weeks)
Drug injection
0
30
60
90
120
150
180
0 1 2 3 4 5 6 0 1 2 3 4 5 6Time after inoculation (weeks)
MFI
of C
D26
Drug injection
Human CD26
CTLA4-Ig
Contl. IgGAnti-CD26
Anti-CD26
CTLA4-Ig
Contl. IgGAnti-CD26
100
80
60
40
20
0
100
80
60
40
20
0100 101 102 103 104 100 101 102 103 104
Fig 3. Anti-CD26 mAb treatment decreases CD26 expression of engrafted human lymphocytes in hu-PBL-NOG mice and decreases in vivo prolif-
eration of CD8+ T cells with preservation of CD4+ T cell proliferation. Hu-PBL-NOG mice receiving humanized anti-CD26 mAb, CTLA4-Ig, or
control IgG were bled every week. Red blood cells were then lysed, and PBMC subsets were analysed by flow cytometry. (A) Time course changes
of mean fluorescence intensity (MFI) of human CD26 of peripheral blood (PB) in NOG mice receiving human PBMC plus control IgG (2 lg/dose, blue line, n = 19), anti-CD26 mAb (2 lg/dose, red line, n = 24), or CTLA4-Ig (2 lg/dose, green line, n = 12). MFI of human CD26 expres-
sion gated for human CD4+ cells (panel a), or for human CD8+ cells (panel b) is shown as average � SD. (B) Representative histogram of
human CD26 expression on day 14 of spleen cells (SP) in hu-PBL-NOG mice receiving control IgG (2 lg/dose, blue line, n = 10), anti-CD26
mAb (2 lg/dose, red line, n = 10), or CTLA4-Ig (2 lg/dose, green line, n = 6). Panel (a) shows representative human CD26 expression gated for
human CD4+ cells in murine splenocytes, and panel (b) shows human CD26 expression gated for human CD8+ cells in splenocytes. Grey or
brown line shows histogram of isotype control Ig staining, or human CD26 expression of human PBMC before inoculation, respectively. Similar
results were observed in independent experiments using NOG mice receiving human PBMC plus control IgG (n = 19), anti-CD26 mAb (n = 24),
or CTLA4-Ig (n = 12). (C) To analyse in vivo cell proliferation in hu-PBL-NOG mice, CFSE-labelled human PBMC were inoculated intraperito-
neally into NOG mice, followed by injection of control IgG (2 lg/dose, blue bars, n = 7), anti-CD26 mAb (2 lg/dose, red bars, n = 8), or
CTLA4-Ig (2 lg/dose, green bars, n = 4). Spleen cells from mice were harvested on day 7, and acquisition was performed using flow cytometry.
Percentages of CFSE-negative, i.e., multiply dividing cells gated for human CD4+ or CD8+ cells, are shown as average � SD *, ** or *** indi-
cates P < 0�05, NS denotes not significant.
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 269British Journal of Haematology, 2013, 162, 263–277

hu-PBL-NOG mice receiving anti-CD26 mAb, and that
treatment with anti-CD26 mAb provided an advantage over
CTLA4-Ig in preventing GVHD at similarly low doses.
Along with lymphocyte infiltration in the GVHD target
tissues, cytokine production level of donor-derived lympho-
cytes plays a role in tissue damages of the GVHD target
organs (Shlomchik, 2007; Ferrara et al, 2009; Blazar et al,
2012). We therefore analysed the mRNA level of effector
(A) NOG mice
Contl. IgG Anti-CD26 mAb CTLA4-Ig
hu-PBL-NOG mice
PBS
H&E
CD4staining
CD8staining
(b)(a) (c) (d)
(f)(e) (g) (h)
(k)(j)(i) (l)
Treatment :
(B)
1000
0
600
200
400
800
1200
Seru
m A
LT a
ctiv
ity (m
iu/m
l)
contl.IgG
Anit-CD26
CTLA4-Ig
Treatment:
CD8+ cells in Liver
(c)
(d) FASLG
(D)
(a) IL2
(c)
(d)
(b)
(C) CD4+ cells in Liver
Contl.IgG
Anti-CD26
CTLA4-IgTreatment:
Rel
ativ
e m
RN
A ex
pres
sion
00·20·40·60·81·0
00·20·40·60·81·0
00·20·40·60·81·0
00·20·40·60·81·0
0
0·5
1·0
1·5
2·0
00·20·40·60·81·0
00·20·40·60·81·0
00·20·40·60·81·0
(a) IL2
(b)
Contl.IgG
Anti-CD26
CTLA4-Ig
NS
NS
NS
IL17A
IL4 TNF
IFNG IFNG
R. Hatano et al
270 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277

cytokines of donor-derived human CD4+ or CD8+ cells in
the recipient liver. In hu-PBL-NOG mice receiving anti-
CD26 mAb, levels of IL2, IFNG, IL4 and IL17A of human
CD4+ cells isolated from the mouse liver decreased
compared to those of hu-PBL-NOG mice receiving control
IgG (in of Fig 4C, panels a–d, *). Similarly, treatment of
hu-PBL-NOG mice with CTLA4-Ig resulted in lower levels
of IL2, IFNG, IL4 and IL17A of human CD4+ cells isolated
from mouse liver as compared to treatment with control
IgG (Fig 4C, panels a–d, **). For human CD8+ cells
isolated from hu-PBL-NOG mice receiving anti-CD26 mAb,
levels of IFNG, TNF and FASLG levels were decreased as
compared to those of hu-PBL-NOG mice receiving control
IgG (Fig 4D, panels b–d, **), while slight elevation of IL2
was observed (Fig 4D, panel a, *). On the other hand,
CTLA4-Ig-treated hu-PBL-NOG mice showed lower levels of
IFNG and TNF as compared to control IgG-treated mice
(Fig 4D, panels b and c,***), while IL2 and FASLG produc-
tion was at similar levels in the two groups (Fig 4D, panels
a and d, NS). Taken together with the above data, these
results indicate that anti-CD26 treatment in hu-PBL-NOG
mice ameliorated liver GVHD by decreasing production of
proinflammatory cytokines of donor-derived human
lymphocytes as well as inhibiting lymphocyte infiltration in
the liver, comparable to CTLA4-Ig.
GVL effect is maintained in hu-PBL-NOG micereceiving anti-CD26 mAb
Given that aGVHD and GVL effects are immune reactions
that are highly linked to each other (Zorn et al, 2002; Wu &
Ritz, 2009), we evaluated the potential influence of anti-
CD26 mAb treatment on GVL effect. Prior to performing in
vivo GVL experiments, we conducted mixed lymphocyte
reaction assays to determine the cytotoxicity of human
T cells on P815 leukaemic cells, and demonstrated that
human CD3+ T cells exerted in vitro cytotoxic effect on P815
cells (Fig S3A). For the subsequent animal studies, mice were
injected subcutaneously in the flank with P815 leukaemic
cells (H2d) on day 0, followed by inoculation of human
PBMCs and GVHD prophylaxis with anti-CD26 mAb or
CTLA4-Ig. All NOG mice transplanted with P815 cells suc-
cumbed to leukaemia by 6 weeks of inoculation with pro-
gressive tumour enlargement (Fig 5A,B, grey lines; Fig S3B,
panel a). Mice transplanted with P815 cells along with
human PBMCs and control IgG showed minimal signs of
tumour growth in the inoculated region, but all mice died
around 4 weeks after inoculation (Fig 5A, blue line). Necrop-
sies revealed that these mice died due to x-GVHD as mani-
fested by the infiltration of human MNCs into the liver,
spleen and lung (data not shown), suggesting that GVL effect
was exerted in the presence of severe GVHD. On the other
hand, mice inoculated with P815 along with human PBMC
and anti-CD26 mAb exhibited enhanced survival rate with
minimal evidence of GVHD (Fig 5A, red line). Importantly,
mice in this group showed significantly slow initial tumour
growth (Fig 5B, red line; Fig S3B, panel b), suggesting the
preservation of GVL effect more than CTLA4-Ig treatment
(Fig 5A, B, green lines).
When competent T cells are transferred into the recipients,
allogeneic T cells have normal GVL capacity to eliminate
leukaemic cells within 2 weeks after transplantation, while
symptoms of GVHD become clinically obvious 3–4 weeks
after transplantation (Meguro, 2010). Therefore, to further
characterize the potential mechanisms involved in the preser-
vation of GVL effect observed in hu-PBL-NOG mice receiv-
ing anti-CD26 mAb, we examined the expression level of
effector cytokines of human CD8+ T cells isolated from the
spleens of hu-PBL-NOG mice at 2 weeks after transplanta-
tion. As shown in Fig 5C, the levels of IL2 (panel a), IFNG
(panel b), and FASLG (panel d) of human CD8+ T cells was
at comparable levels between hu-PBL-NOG mice receiving
anti-CD26 mAb (red bars) and control IgG (blue bars), and
the TNF level was increased in anti-CD26 mAb group
compared to control IgG group (Fig 5C, panel c, *). On the
other hand, CTLA4-Ig treatment suppressed the production
Fig 4. Anti-CD26 mAb treatment in hu-PBL-NOG mice reduces liver infiltration of human lymphocytes with reduction of proinflammatory
cytokines. (A) Representative examples of the liver on day 28 from NOG mice receiving PBS alone (negative control, panels a, e, i), or hu-
PBL-NOG mice receiving control IgG (panels b, f, j), low dose anti-CD26 mAb (panels c, g, k), or low dose CTLA4-Ig (panels d, h, l). Slides were
stained with H&E (panels a–d), anti-human CD4 (panels e–h), and anti-human CD8 (panels i–l). All slides are at 2009 magnification. Similar
results were observed in independent experiments using mice receiving PBS alone (n = 13), or hu-PBS-NOG mice receiving control IgG (n = 6),
anti-CD26 mAb (n = 14), or CTLA4-Ig (n = 10). (B) Serum alanine transaminase (ALT) activity on day 21 in hu-PBL-NOG mice receiving
control IgG (2 lg/dose, blue dots, n = 10), anti-CD26 mAb (2 lg/dose, red dots, n = 14), or CTLA4-Ig (2 lg/dose, green dots, n = 10). Each
dot indicates an individual value and horizontal bars indicate mean value. * indicates P < 0�05. NS denotes ‘not significant’. (C) Human CD4+ T
cells were isolated from the liver of hu-PBL-NOG mice receiving control IgG (Contl. IgG, blue bars, n = 6), anti-CD26 mAb (red bars, n = 6), or
CTLA4-Ig (green bars, n = 6) at 4 weeks after inoculation of human PBMCs. mRNA expression of human IL2 (panel a), IFNG (panel b), IL4
(panel c), or IL17A (panel d) was quantified by real-time RT-PCR, as described in Materials and methods. Each expression was normalized to
hypoxanthine phosphoribosyltransferase 1 (HPRT1) and relative expression levels compared with the sample of control IgG treatment (indicated
as dashed grey lines) were shown. * or ** indicates P < 0�05, compared to control IgG. (D) mRNA levels of human CD8+ T cells isolated from
the liver of hu-PBL-NOG mice receiving control IgG (Contl. IgG, blue bars, n = 6), anti-CD26 mAb (red bars, n = 6), or CTLA4-Ig (green bars,
n = 6) at 4 weeks after inoculation of human PBMCs are shown as in (C). mRNA expression of human IL2 (panel a), IFNG (panel b), TNF
(panel c), or FASLG (panel d) was quantified by real-time RT-PCR, as described in Materials and methods. The bar graphs are demonstrated as
in (C). *, ** or *** indicates P < 0�05, compared to control IgG. NS denotes ‘not significant’, compared to control IgG.
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 271British Journal of Haematology, 2013, 162, 263–277

of IL2, IFNG, TNF and FASLG of human CD8+ T cells in
hu-PBL-NOG spleen as compared to control IgG or anti-
CD26 mAb treatment (Fig 5C, ** and ***). These results
suggest that the GVL effect of cytotoxic effector function
occurring at the early time period prior to manifestation of
x-GVHD was preserved with the associated increase in the
production of IL2, IFNG, TNF or FASLG in hu-PBL-NOG
mice receiving anti-CD26 mAb. On the other hand, in
hu-PBL-NOG mice receiving CTLA4-Ig, proinflammatory
cytokines produced from donor-derived CD8+ cells sup-
pressed any potential GVL effect at the early time period
prior to x-GVHD development. These results suggest that
CTLA4-Ig treatment suppressed the development of GVL as
well as x-GVHD in hu-PBL-NOG mice, whereas effective
GVL function was maintained with anti-CD26 mAb therapy.
CD26+ MNCs infiltrate the skin or GI tract of alloHSCTpatients with acute GVHD
Although treatment with a murine antibody against human
CD26 exerted the therapeutic effect of controlling GVHD in
patients with steroid-resistant acute GVHD following
alloHSCT (Bacigalupo et al, 1985; de Meester et al, 1993),
the precise cellular basis of human GVHD, as well as the role
of CD26 in this process, has not yet been clearly delineated.
To address this issue, we examined human GVHD tissue
biopsies in alloHSCT recipients with or without active
GVHD. Although the liver is one of the GVHD targeting
organs, liver tissues are rarely obtained from patients with
active hepatic GVHD in the clinical setting due to the risks
of liver biopsies. Pathological diagnosis is typically conducted
(A)
Time after transplantation (weeks)
Per
cent
sur
viva
l(B)
Time after transplantation (weeks)
Tum
our v
olum
e (m
m3 )
(C)
60
1000
2000
3000
0
0·5
1·0
1·5
00·20·40·60·81·0
00·20·40·60·81·01·21·4
00·20·40·60·81·0
(a) IL2
(b) IFNG
(c) TNF
(d) FASLG
Contl.IgG
Anti-CD26
CTLA4-IgTreatment:
P815 transplanted into :
hu-PBL-NOG + CTLA4-Ig
hu-PBL-NOG + Contl. IgGhu-PBL-NOG + Anti-CD26
NOG
0
100
80
60
40
20
3 4 50 1 2 3 4 5 6
Drug injection
P815 transplanted into :
hu-PBL-NOG + CTLA4-Ighu-PBL-NOG + Anti-CD26NOG
NS
NS
NS
Rel
ativ
e m
RN
A ex
pres
sion
Fig 5. Anti-CD26 mAb treatment does not
eliminate GVL activity. P815 leukaemic cells
(2 9 104/body) were inoculated subcutane-
ously into the shaved flank of NOG mice. On
the next day, human PBMCs (1 9 107/body)
were inoculated intraperitoneally, followed by
treatment with humanized anti-CD26 mAb,
CTLA4-Ig, or control IgG (each, 2 lg/dose 9 10 doses). (A) Kaplan–Meier survival
curves for mice receiving P815 alone (grey line,
n = 4), P815 plus PBMC plus control IgG
(blue line, n = 3), P815 plus PBMC plus anti-
CD26 mAb (red line, n = 4) or P815 plus
PBMC plus CTLA4-Ig (green line, n = 3). (B)
Tumour volume of P815 transplanted into the
flank of NOG mice (grey line, n = 4), or hu-
PBL-NOG mice receiving anti-CD26 mAb (red
line, n = 4), or CTLA4-Ig (green line, n = 3).
All mice receiving P815 plus PBMC plus con-
trol IgG (blue line of Fig 5A) died without
tumour formation, therefore tumour volume
of this group is not shown in the graph. *indicates P < 0�01 of anti-CD26 mAb group
versus P815 alone or control IgG groups. (C)
Human CD8+ T cells were isolated from the
spleen of hu-PBL-NOG mice receiving control
human IgG (Contl. IgG, n = 4), anti-CD26
mAb (n = 4), or CTLA4-Ig (n = 3) on day 14
after inoculation of human PBMCs. mRNA
expression of human IL2 (panel a), IFNG
(panel b), TNF (panel c), or FASLG (panel d)
was quantified by real-time RT-PCR. Each
expression was normalized to HPRT1 and rela-
tive expression levels compared with the sam-
ple of control IgG treatment (indicated dashed
grey lines) were shown. *, ** or *** indicates
P < 0�05. NS denotes ‘not significant’.
R. Hatano et al
272 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277

with endoscopic GI tract mucosal biopsy for GI GVHD
(Kreisel et al, 2011), as well as skin biopsy for the diagnosis
of skin GVHD (Sale et al, 1977). Therefore, we evaluated
specimens from skin biopsies and endoscopic samples of
upper GI (duodenal) mucosa harvested from patients with
active GVHD patients. Thirty-four adult patients (median
age, 38 years; range, 22–60 years) with AML who underwent
alloHSCT at the Keio University Hospital were included in
this study. GVHD prophylaxis was ciclosporin and metho-
trexate for related donor transplantation, or tacrolimus and
methotrexate for unrelated donor transplantation. The demo-
graphic profiles of the biopsied patients are shown in
Table I. All 34 patients had clinical skin GVHD (Stage I,
n = 16; Stage II, n = 11; Stage III, n = 7). Skin biopsies from
34 patients were analysed for CD26+ MNC infiltration. Stain-
ing of human skin biopsies for CD26 revealed that all
patients with active GVHD had CD26+ MNC infiltration in
the skin (Fig 6A, panels a–c). To determine that CD26+
MNCs are CD3+ T cells, co-staining of a representative skin
specimen revealed that CD3+CD26+ MNCs infiltrated the
skin of patients with aGVHD (Fig. S4). Fig 6B demonstrates
data correlating the absolute number (per square millimetre)
of CD26+ MNCs in the skin with the GVHD pathological
grade, and reveals that the level of CD26+ MNCs constitu-
tively infiltrating the skin of patients with aGVHD did not
correlate with disease severity. Furthermore, among patients
who had biopsies of the upper GI tract mucosa performed as
a diagnostic procedure for digestive symptoms, 32 patients
had clinical GI GVHD (Stage I, n = 10; Stage II, n = 14;
Stage III, n = 7; Stage IV, n = 1). Staining of human duode-
nal mucosal biopsies for CD26 revealed that all patients with
active GVHD had CD26+ MNC infiltration in the duodenal
mucosa (Fig 6C, panels a–c). In addition, the absolute num-
ber (per square millimetre) of CD26+ MNCs constitutively
infiltrating the duodenal mucosa of patients with aGVHD
did not correlate with disease severity as indicated by GVHD
pathologic grade (Fig 6D). However, in patients with severe
GVHD, CD26+ MNCs were found to infiltrate into the stro-
mal tissue of the duodenum, and were not only limited to
the surface mucosa (Fig 6C, panel c). Taken together, these
results strongly suggest that CD26+ MNCs play an important
role in the inflamed tissues of aGVHD patients, consistent
with previous data demonstrating that CD26+ lymphocytes
migrated to inflamed tissues with effector functions (Eguchi
et al, 1989; Gerli et al, 1996; Hafler et al, 1985; Mizokami
et al, 1996), suggesting that CD26 is a logical therapeutic
target in aGVHD.
Discussion
In the current study, we show that CD26+ T cells play an
important role in GVHD mediated by human lymphocytes,
and that anti-CD26 mAb is an effective treatment for GVHD
in hu-PBL-NOG mouse model. The effect of anti-CD26 mAb
on GVHD is shown to be comparable to the clinically avail-
able costimulation blocking agent, CTLA4-Ig (abatacept).
Importantly, anti-CD26 mAb therapy appears to be superior
to CTLA4-Ig treatment in that it preserves engraftment and
the GVL effect. Finally, our work demonstrates significant
infiltration of CD26+ MNCs in GVHD target tissues obtained
from patients with aGVHD.
Our present work indicates the infiltration of human
CD26+ lymphocytes in the x-GVHD lesions of hu-PBL-NOG
mice (Fig 1B) and the presence of human CD26high T cells
in the peripheral blood or spleen of hu-PBL-NOG mice
that developed x-GVHD (Fig 3A,B). When x-GVHD of
hu-PBL-NOG mice was diminished by anti-CD26 mAb treat-
ment, CD26high T cells were not detected in the peripheral
blood and spleen (Fig 3A,B). A potential explanation for this
finding may be internalization of surface CD26 expression on
the inoculated human lymphocytes induced by humanized
anti-CD26 mAb therapy (Dang et al, 1990). The decreased
CD26high lymphocyte level, with resultant decline in CD26-
mediated costimulation and migration, might contribute to
lowered xenogeneic activation and decreased tissue infiltra-
tion of GVHD target organs. In fact, a low level of lympho-
cyte infiltration in the liver was shown in hu-PBL-NOG mice
receiving a low dose of CTLA4-Ig (Fig 4A, panels d, h, l),
while lymphocyte infiltration in the liver was barely detected
Table I. Patients characteristics and clinical status at time of biopsy
for diagnosis of acute GVHD.
Disease (n)
AML 34
1st or 2nd CR 24
Advanced 10
Age (years)
Median 38
Range 22–60
Gender (n)
Male 20
Female 14
Allo-graft source (n)
BM 30
PB 1
CB 3
Conditioning (n)
Full 25
Reduced 9
Matching (n)
Related 8
Unrelated 26
*aGVHD clinical grade (n)
0 6
I 10
II 9
III 8
IV 1
*aGVHD, acute graft-versus-host disease; AML, acute myeloid leu-
kaemia; BM, bone marrow; CB, cord blood; CR, complete remission;
PB, peripheral blood.
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 273British Journal of Haematology, 2013, 162, 263–277

in hu-PBL-NOG mice receiving anti-CD26 mAb (Fig 4A,
panels c, g, k). These observations might result from the fact
that CTLA4 has no direct effect on lymphocyte migration,
whereas the CD26 molecule is strongly associated with
lymphocyte migration into inflamed tissues (Hafler et al,
1985; Eguchi et al, 1989; Masuyama et al, 1992; Gerli et al,
1996; Mizokami et al, 1996). It is also possible that anti-
CD26 mAb treatment could affect other functional program-
ming of inoculated human lymphocytes, such as cell cycle
regulation. We have previously reported that binding of anti-
CD26 mAb to CD26 on T cell surface inhibits human T cell
growth and proliferation in both CD26-transfected Jurkat
T-cell lines and human T cell clones by inducing G1/S arrest,
which is associated with enhancement of p21Cip1 (CDKN1A)
expression (Ohnuma et al, 2002). In this regard, the suppres-
sion of x-GVHD by anti-CD26 mAb shown here might result
from a direct effect on cell cycle inhibition mediated by anti-
CD26 mAb. In addition to the preventive effect on x-GVHD
of anti-CD26 mAb, we observed that systemic manifestations
of x-GVHD were greatly reduced by administration of anti-
CD26 mAb when x-GVHD symptoms developed after
4 weeks of human PBMCs inoculation (data not shown).
Taken together, our results indicate that anti-CD26 mAb
treatment appears to affect T cell functions of inoculated
human lymphocytes in hu-PBL-NOG mice.
Costimulatory pathways are necessary to induce T cell
proliferation, cytokine secretion and effector function follow-
ing antigen-mediated T cell receptor activation. Given that
CD28 is widely expressed on T cells and CD28�CD8+ T cells
are well-recognized as na€ıve or highly-antigen experienced
(late-differentiated) cells (Rudd et al, 2009; Strioga et al,
2011), blockade of CD28 pathway may lead to profound
suppression of T cell function. In fact, our current study
showed that T cell proliferation or cytokine production was
inhibited at a greater level by CTLA4-Ig than by anti-CD26
mAb (Figs 3C, 4C, 4D or 5C). In this regard, immunosup-
pression by CTLA4-Ig not only diminished the engraftment
of donor-derived human lymphocytes in NOG mice
Stage I Stage II Stage III
(C)
GI
Clinical GI GVHD
Stage I Stage II Stage III
(A)
Skin
Clinical Skin GVHD
(a) (b) (c)
(a) (b) (c)
Cells/mm 2(B)
(D)
CD
26+ M
NC
s
0
10
20
30
Skin GVHD (pathological grade)
Grade 0 Grade 1 Grade 2 Grade 3-4
Cells/mm2
CD
26+ M
NC
s
0
10
20
GI GVHD (pathological grade)
Grade 1 Grade 2 Grade 3-4Grade 0
Fig 6. CD26+ MNCs infiltrate into the skin or duodenal mucosa of alloHSCT patients with acute GVHD. Histopathological evaluation of
representative skin and duodenal mucosa samples biopsied from transplant patients and stained with anti-human CD26 polyclonal antibody,
counterstained with haematoxylin, and then analysed in a blinded fashion, as described in Materials and methods. (A) Representative examples of
skin obtained from patients with clinical Stage I (panel a), Stage II (panel b), or Stage III (panel c) of acute skin GVHD. Original magnification
1009. (B) Graphs comparing the relative amount of CD26+ MNCs in the skin with acute GVHD pathologic grade 0 (n = 6), 1 (n = 10), 2
(n = 9), or 3–4 (n = 9). Each dot indicates individual value. The horizontal lines in the scattergrams indicate each mean value, and error bars
indicate SD. (C) Representative examples of duodenal mucosa obtained from patients with gastrointestinal (GI) clinical Stage I (panel a), Stage II
(panel b), or Stage III (panel c) of acute GVHD. Original magnification 1009. CD26+ MNCs infiltrated locally in the mucosal epithelia, while for
patients with Stage III, increasing number of CD26+ MNCs was observed in the interstitial tissue as well as in the mucosal epithelia. (D) Graphs
comparing the relative amount of CD26+ MNCs in the duodenal mucosa with acute GVHD pathological grade 0 (n = 4), 1 (n = 11), 2 (n = 8),
or 3–4 (n = 9). Each dot indicates individual value. The horizontal lines in the scattergrams indicate each mean value, and error bars indicate
SD.
R. Hatano et al
274 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277

(Fig 2C), but also decreased the GVL effect in hu-PBL-NOG
mice (Fig 5A,B, green lines). These data strongly suggest
that while CTLA4-Ig exhibits strong suppressive effect on
x-GVHD, it also has several adverse effects, such as graft
rejection, inhibition of GVL reactivity or enhancement of
opportunistic infections.
Our previous studies showed that CD26-mediated costi-
mulation in human CD4+ T cells exerted an effect on
production of TH1 type proinflammatory cytokines such as
IL2 or IFNG (Morimoto & Schlossman, 1998), and that
soluble anti-CD26 mAb treatment decreased IL2 production
of human CD4+ T cells (Ohnuma et al, 2002). Our present
data indicate that anti-CD26 mAb treatment suppressed the
generation of IL2 and IFNG of donor-derived human CD4+
cells in the liver of hu-PBL-NOG mice (Fig 4C, panels a, b),
and that this suppression might result in the inhibition of x-
GVHD. In addition to TH1 type cytokines, the TH2 type
cytokine IL4 was found to be abundant in GVHD liver
lesions in human studies, and a high level of IL4 production
in the GVHD liver tissue appears to contribute to its patho-
genesis (Lai et al, 2012). In agreement with previous studies
of patients with aGVHD (Lai et al, 2012), we observed the
reduction of IL4 production of donor-derived human CD4+
T cells in the liver of hu-PBL-NOG mice receiving
anti-CD26 mAb or CTLA4-Ig (Fig 4C, panel c). Recently, it
was reported that CD26high T cells contain TH17 cells, and
that CD26high TH17 cells are enriched in the inflamed tissue
of patients with hepatitis and inflammatory bowel disease
(Bengsch et al, 2012). We observed decrease in the produc-
tion of IL17A in hu-PBL-NOG mice treated with anti-CD26
mAb or CTLA4-Ig (Fig 4C, panel d). Our present data on
cytokine production suggest that, due to the paradoxical and
variable effects resulting from targeting TH1-, TH2- and
TH17-type cytokines, such approaches alone may not be suf-
ficient to fully treat or prevent GVHD in the clinical setting,
but they may serve as useful adjunct therapeutic strategies to
reduce GVHD-associated tissue injuries.
Recently, it has been reported that CD8+ T cells specific for
influenza express high levels of CD26, and that this subset
could produce IL2 following stimulation, while CD8+ T cells
specific for chronic infection with persistent antigens, such as
cytomegalovirus (CMV), Epstain-Barr virus (EBV) or human
immunedeficiency virus (HIV), lack CD26 expression (Ibegbu
et al, 2009). The authors concluded that high CD26 expres-
sion among CD8+ T cells may be a marker of effective long-
term memory T cell formation and that the lack of CD26
expression among CD8+ T cells in CMV, EBV or HIV infec-
tion may indicate defective memory T cells (Ibegbu et al,
2009). It is therefore a concern that anti-CD26 mAb therapy
may potentially cause increased infection. However, CD8+ T
cells in hu-PBL-NOG mice treated with anti-CD26 mAb
exhibited increased IL2 production (Fig 4D, panel a; Fig 5C,
panel a). These results strongly suggest that IL2-producing
CD8+ T cells persist despite humanized anti-CD26 mAb
therapy, which may lead to the formation of effective memory
T cells against potential opportunistic infections following
alloHSCT. In addition, IL2 is necessary for the proliferation of
regulatory T cells (Treg), which can ameliorate GVHD
(Koreth et al, 2011). Therefore, our data that IL2-producing
CD8+ T cells still remain following humanized anti-CD26
mAb therapy may indicate the existence of immune tolerance
via Treg proliferation in the GVHD target organs.
Results from murine GVHD models have not always
translated to human disease and discrepancies between mur-
ine and human samples have been reported. While the
CD26 amino acid sequence has 85% amino acid identity
with the mouse CD26 (Marguet et al, 1992), mouse CD26
exerts no costimulatory function in mouse T cells, and is
not an activation marker of T cells nor an adenosine deami-
nase-binding protein (Marguet et al, 1992; Yan et al, 2003).
On the other hand, caveolin-1 (CAV1), a costimulatory
ligand for CD26 in humans, has 95% amino acid identity
with the mouse caveolin-1 (Engelman et al, 1998), and the
binding regions of the mouse caveolin-1 for human CD26
are well conserved. Therefore, human T cells were activated
even in NOG mice via CD26-caveolin-1 interaction. Another
important point to consider when interpreting our data is
the fact that, in our studies, mice were analysed at a time
when they were moribund, in contrast to the samples
obtained from patients that, for the most part, were obtained
at the time of GVHD diagnosis or shortly thereafter. More-
over, first-line therapies for clinical GVHD, such as metho-
trexate or calcineurin inhibitors, may have an effect on T
cell costimulatory signalling, cytokine production or expres-
sion of cell surface molecules.
Specific inhibition of DPP4 enzyme via Diprotin A
enhanced bone marrow engraftment in certain murine BMT
models (Christopherson et al, 2004). Although DPP4 enzy-
matic activity is required for CD26-mediated T cell costimu-
lation (Morimoto & Schlossman, 1998), it is unclear whether
DPP4 enzyme inhibition has an effect on GVHD develop-
ment. Regarding this point, we note that the humanized
anti-CD26 mAb used in the present study does not inhibit
DPP4 enzymatic activity of human CD26 (data not shown).
In conclusion, CD26-mediated T cell activation appears to
play a significant role in human GVHD. As full suppression
of x-GVHD with interventional therapies is currently a
difficult challenge, our data demonstrating that control of
x-GVHD can be achieved by modulating CD26high T cells
with anti-CD26 mAb are potentially important clinically.
Our work also suggests that anti-CD26 mAb treatment may
be a novel therapeutic approach for GVHD in the future.
Acknowledgements
The authors thank Ms Kaoru Komoriya for excellent assis-
tance with animal husbandry. This work was supported by
Grant-in-Aid of The Ministry of Education, Science, Sports
(K.O. and C.M.) and Culture, Ministry of Health, Labour,
and Welfare, Japan (C.M.).
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 275British Journal of Haematology, 2013, 162, 263–277

Authorship contributions
Contributions: R.H., and J.Y. performed the experiments,
interpreted the data and assisted with the paper, K.O. and
C.M. designed the research, interpreted the data and wrote
the paper, N.H.D. interpreted the data, assisted with the
paper, and proof-read the manuscript, and T.Y. provided the
clinical data and samples, performed the experiments, analy-
sed pathological results, interpreted the data and assisted
with the paper.
Conflict-of-interest disclosure
The authors declare no competing financial interests.
Supporting Information
Additional Supporting Information may be found in the
online version of this article:
Fig S1. Mouse anti-human CD26 mAb clone 5K78
recognizes human CD26 even in the presence of humanized
anti-CD26 mAb.
Fig S2. Human CD3+CD26+ T cells infiltrated the liver of
hu-PBL-NOG mice developing x-GVHD.
Fig S3. GVL experiments in hu-PBL-NOG mice.
Fig S4. CD3+CD26+ MNCs infiltrate the skin of alloHSCT
patients with acute GVHD.
Table SI. The primer sequences for RT-PCR.
References
Aoe, K., Amatya, V.J., Fujimoto, N., Ohnuma, K.,
Hosono, O., Hiraki, A., Fujii, M., Yamada, T.,
Dang, N.H., Takeshima, Y., Inai, K., Kishimoto,
T. & Morimoto, C. (2012) CD26 overexpression
is associated with prolonged survival and
enhanced chemosensitivity in malignant pleural
mesothelioma. Clinical Cancer Research, 18,
1447–1456.
Bacigalupo, A., Corte, G., Ramarli, D., van Lint,
M.T., Frassoni, F. & Marmont, A. (1985) Intra-
venous monoclonal antibody (BT 5/9) for the
treatment of acute graft-versus-host disease. Acta
Haematologica, 73, 185–186.
Bengsch, B., Seigel, B., Flecken, T., Wolanski, J.,
Blum, H.E. & Thimme, R. (2012) Human Th17
cells express high levels of enzymatically active
dipeptidylpeptidase IV (CD26). Journal of
Immunology, 188, 5438–5447.
Blazar, B.R., Taylor, P.A., Boyer, M.W., Panoskalt-
sis-Mortari, A., Allison, J.P. & Vallera, D.A.
(1997) CD28/B7 interactions are required for
sustaining the graft-versus-leukemia effect of
delayed post-bone marrow transplantation
splenocyte infusion in murine recipients of mye-
loid or lymphoid leukemia cells. Journal of
Immunology, 159, 3460–3473.
Blazar, B.R., Murphy, W.J. & Abedi, M. (2012)
Advances in graft-versus-host disease biology and
therapy. Nature Reviews Immunology, 12, 443–458.
Briones, J., Novelli, S. & Sierra, J. (2011) T-cell
costimulatory molecules in acute-graft-versus
host disease: therapeutic implications. Bone
Marrow Research, 2011, 976793.
Christopherson, K.W. II, Hangoc, G., Mantel, C.R.
& Broxmeyer, H.E. (2004) Modulation of hema-
topoietic stem cell homing and engraftment by
CD26. Science, 305, 1000–1003.
Dang, N.H., Torimoto, Y., Sugita, K., Daley, J.F.,
Schow, P., Prado, C., Schlossman, S.F. & Mor-
imoto, C. (1990) Cell surface modulation of
CD26 by anti-1F7 monoclonal antibody.
Analysis of surface expression and human T cell
activation. Journal of Immunology, 145,
3963–3971.
Dong, R.P., Tachibana, K., Hegen, M., Scharpe, S.,
Cho, D., Schlossman, S.F. & Morimoto, C.
(1998) Correlation of the epitopes defined by
anti-CD26 mAbs and CD26 function. Molecular
Immunology, 35, 13–21.
Eguchi, K., Ueki, Y., Shimomura, C., Otsubo, T.,
Nakao, H., Migita, K., Kawakami, A., Matsuna-
ga, M., Tezuka, H., Ishikawa, N., Ito, K. &
Nagataki, S. (1989) Increment in the Ta1+ cells
in the peripheral blood and thyroid tissue of
patients with Graves’ disease. Journal of Immu-
nology, 142, 4233–4240.
Engelman, J.A., Zhang, X., Galbiati, F., Volonte,
D., Sotgia, F., Pestell, R.G., Minetti, C., Scherer,
P.E., Okamoto, T. & Lisanti, M.P. (1998)
Molecular genetics of the caveolin gene family:
implications for human cancers, diabetes,
Alzheimer disease, and muscular dystrophy.
American Journal of Human Genetics, 63,
1578–1587.
Ferrara, J.L., Levine, J.E., Reddy, P. & Holler, E.
(2009) Graft-versus-host disease. Lancet, 373,
1550–1561.
Fox, D.A., Hussey, R.E., Fitzgerald, K.A., Acuto,
O., Poole, C., Palley, L., Daley, J.F., Schlossman,
S.F. & Reinherz, E.L. (1984) Ta1, a novel 105
KD human T cell activation antigen defined by
a monoclonal antibody. Journal of Immunology,
133, 1250–1256.
Genovese, M.C., Becker, J.C., Schiff, M., Luggen,
M., Sherrer, Y., Kremer, J., Birbara, C., Box, J.,
Natarajan, K., Nuamah, I., Li, T., Aranda, R.,
Hagerty, D.T. & Dougados, M. (2005) Abatacept
for rheumatoid arthritis refractory to tumor
necrosis factor alpha inhibition. New England
Journal of Medicine, 353, 1114–1123.
Gerli, R., Muscat, C., Bertotto, A., Bistoni, O.,
Agea, E., Tognellini, R., Fiorucci, G., Cesarotti,
M. & Bombardieri, S. (1996) CD26 surface
molecule involvement in T cell activation and
lymphokine synthesis in rheumatoid and other
inflammatory synovitis. Clinical Immunology
and Immunopathology, 80, 31–37.
Giralt, S. (2012) Graft-versus-host disease: have we
solved the problem? Journal of Clinical Oncology,
30, 3160–3161.
Gribben, J.G., Guinan, E.C., Boussiotis, V.A., Ke,
X.Y., Linsley, L., Sieff, C., Gray, G.S., Freeman,
G.J. & Nadler, L.M. (1996) Complete blockade
of B7 family-mediated costimulation is necessary
to induce human alloantigen-specific anergy: a
method to ameliorate graft-versus-host disease
and extend the donor pool. Blood, 87, 4887–
4893.
Hafler, D.A., Fox, D.A., Manning, M.E., Schloss-
man, S.F., Reinherz, E.L. & Weiner, H.L. (1985)
In vivo activated T lymphocytes in the periph-
eral blood and cerebrospinal fluid of patients
with multiple sclerosis. New England Journal of
Medicine, 312, 1405–1411.
Hatano, R., Ohnuma, K., Yamamoto, J., Dang,
N.H. & Morimoto, C. (2013) CD26-mediated
co-stimulation in human CD8+ T cells provokes
effector function via pro-inflammatory cytokine
production. Immunology, 138, 165–172.
Ibegbu, C.C., Xu, Y.X., Fillos, D., Radziewicz, H.,
Grakoui, A. & Kourtis, A.P. (2009) Differential
expression of CD26 on virus-specific CD8+ T
cells during active, latent and resolved infection.
Immunology, 126, 346–353.
Ito, M., Hiramatsu, H., Kobayashi, K., Suzue, K.,
Kawahata, M., Hioki, K., Ueyama, Y., Koyanagi,
Y., Sugamura, K., Tsuji, K., Heike, T. & Nakah-
ata, T. (2002) NOD/SCID/ccnull mouse: an
excellent recipient mouse model for engraftment
of human cells. Blood, 100, 3175–3182.
Koreth, J., Matsuoka, K., Kim, H.T., McDonough,
S.M., Bindra, B., Alyea, E.P. 3rd, Armand, P.,
Cutler, C., Ho, V.T., Treister, N.S., Bienfang,
D.C., Prasad, S., Tzachanis, D., Joyce, R.M.,
Avigan, D.E., Antin, J.H., Ritz, J. & Soiffer, R.J.
(2011) Interleukin-2 and regulatory T cells in
graft-versus-host disease. New England Journal of
Medicine, 365, 2055–2066.
Kreisel, W., Dahlberg, M., Bertz, H., Harder, J.,
Potthoff, K., Deibert, P., Schmitt-Graeff, A. &
Finke, J. (2011) Endoscopic diagnosis of acute
intestinal GVHD following allogeneic hematopoi-
etic SCT: a retrospective analysis in 175 patients.
Bone Marrow Transplantation, 47, 430–438.
Lai, H.Y., Chou, T.Y., Tzeng, C.H. & Lee, O.K.
(2012) Cytokine profiles in various graft-versus-
R. Hatano et al
276 ª 2013 John Wiley & Sons LtdBritish Journal of Haematology, 2013, 162, 263–277

host disease target organs following hematopoietic
stem cell transplantation. Cell Transplantation, 21,
2033–2045.
Lenschow, D.J., Zeng, Y., Thistlethwaite, J.R.,
Montag, A., Brady, W., Gibson, M.G., Linsley,
P.S. & Bluestone, J.A. (1992) Long-term survival
of xenogeneic pancreatic islet grafts induced by
CTLA4Ig. Science, 257, 789–792.
Marguet, D., Bernard, A.M., Vivier, I., Darmoul,
D., Naquet, P. & Pierres, M. (1992) cDNA clon-
ing for mouse thymocyte-activating molecule. A
multifunctional ecto-dipeptidyl peptidase IV
(CD26) included in a subgroup of serine prote-
ases. Journal of Biological Chemistry, 267, 2200–
2208.
Masuyama, J., Berman, J.S., Cruikshank, W.W.,
Morimoto, C. & Center, D.M. (1992) Evidence
for recent as well as long term activation of T
cells migrating through endothelial cell mono-
layers in vitro. Journal of Immunology, 148,
1367–1374.
de Meester, I., Scharpe, S., Vanham, G., Bosmans,
E., Heyligen, H., Vanhoof, G. & Corte, G.
(1993) Antibody binding profile of purified and
cell-bound CD26. Designation of BT5/9 and
TA5.9 to the CD26 cluster. Immunobiology, 188,
145–158.
Meguro, A. (2010) Lack of IL-21 signal attenuates
graft-versus-leukemia effect in the absence of
CD8 T-cells. Bone Marrow Transplantation, 46,
1557–1565.
Mizokami, A., Eguchi, K., Kawakami, A., Ida, H.,
Kawabe, Y., Tsukada, T., Aoyagi, T., Maeda, K.,
Morimoto, C. & Nagataki, S. (1996) Increased
population of high fluorescence 1F7 (CD26)
antigen on T cells in synovial fluid of patients
with rheumatoid arthritis. Journal of Rheumatol-
ogy, 23, 2022–2026.
Morimoto, C. & Schlossman, S.F. (1998) The
structure and function of CD26 in the T-cell
immune response. Immunological Reviews, 161,
55–70.
Morimoto, C., Torimoto, Y., Levinson, G., Rudd,
C.E., Schrieber, M., Dang, N.H., Letvin, N.L. &
Schlossman, S.F. (1989) 1F7, a novel cell surface
molecule, involved in helper function of CD4
cells. Journal of Immunology, 143, 3430–3439.
Muscat, C., Bertotto, A., Agea, E., Bistoni, O.,
Ercolani, R., Tognellini, R., Spinozzi, F., Cesar-
otti, M. & Gerli, R. (1994) Expression and func-
tional role of 1F7 (CD26) antigen on peripheral
blood and synovial fluid T cells in rheumatoid
arthritis patients. Clinical and Experimental
Immunology, 98, 252–256.
Ohnuma, K., Ishii, T., Iwata, S., Hosono, O.,
Kawasaki, H., Uchiyama, M., Tanaka, H.,
Yamochi, T., Dang, N.H. & Morimoto, C.
(2002) G1/S cell cycle arrest provoked in human
T cells by antibody to CD26. Immunology, 107,
325–333.
Ohnuma, K., Dang, N.H. & Morimoto, C. (2008)
Revisiting an old acquaintance: CD26 and its
molecular mechanisms in T cell function. Trends
in Immunology, 29, 295–301.
Pasquini, M.C., Wang, Z., Horowitz, M.M. & Gale,
R.P. (2010) 2010 report from the Center for
International Blood and Marrow Transplant
Research (CIBMTR): current uses and outcomes
of hematopoietic cell transplants for blood and
bone marrow disorders. Clinical Transplants,
2010, 87–105.
Przepiorka, D., Weisdorf, D., Martin, P., Klinge-
mann, H.G., Beatty, P., Hows, J. & Thomas,
E.D. (1995) 1994 consensus conference on acute
GVHD grading. Bone Marrow Transplantation,
15, 825–828.
Rudd, C.E., Taylor, A. & Schneider, H. (2009)
CD28 and CTLA-4 coreceptor expression and
signal transduction. Immunological Reviews, 229,
12–26.
Saito, K. (1998) Involvement of CD40 ligand-
CD40 and CTLA4-B7 pathways in murine acute
graft-versus-host disease induced by allogeneic T
cells lacking CD28. J. Immunol., 160, 4225–4231.
Sale, G.E., Lerner, K.G., Barker, E.A., Shulman,
H.M. & Thomas, E.D. (1977) The skin biopsy in
the diagnosis of acute graft-versus-host disease in
man. American Journal of Pathology, 89, 621–636.
Shlomchik, W.D. (2007) Graft-versus-host disease.
Nature Reviews Immunology, 7, 340–352.
Strioga, M., Pasukoniene, V. & Characiejus, D.
(2011) CD8+CD28- and CD8+CD57+ T cells and
their role in health and disease. Immunology,
134, 17–32.
Tanaka, T., Camerini, D., Seed, B., Torimoto, Y.,
Dang, N.H., Kameoka, J., Dahlberg, H.N.,
Schlossman, S.F. & Morimoto, C. (1992)
Cloning and functional expression of the T cell
activation antigen CD26. Journal of Immunology,
149, 481–486.
Tsirigotis, P., Or, R., Resnick, I.B. & Shapira, M.Y.
(2012) Immunotherapeutic approaches to
improve graft-versus-tumor effect and reduce
graft-versus-host disease. Immunotherapy, 4,
407–424.
Vincenti, F., Charpentier, B., Vanrenterghem, Y.,
Rostaing, L., Bresnahan, B., Darji, P., Massari,
P., Mondragon-Ramirez, G.A., Agarwal, M., Di
Russo, G., Lin, C.S., Garg, P. & Larsen, C.P.
(2010) A phase III study of belatacept-based
immunosuppression regimens versus cyclospor-
ine in renal transplant recipients (BENEFIT
study). American Journal of Transplantation, 10,
535–546.
Washington, K. & Jagasia, M. (2009) Pathology of
graft-versus-host disease in the gastrointestinal
tract. Human Pathology, 40, 909–917.
Wu, C.J. & Ritz, J. (2009) Revealing tumor immu-
nity after hematopoietic stem cell transplanta-
tion. Clinical Cancer Research, 15, 4515–4517.
Yan, S., Marguet, D., Dobers, J., Reutter, W. &
Fan, H. (2003) Deficiency of CD26 results in a
change of cytokine and immunoglobulin secre-
tion after stimulation by pokeweed mitogen.
European Journal of Immunology, 33,
1519–1527.
Zorn, E., Wang, K.S., Hochberg, E.P., Canning, C.,
Alyea, E.P., Soiffer, R.J. & Ritz, J. (2002)
Infusion of CD4+ donor lymphocytes induces
the expansion of CD8+ donor T cells with
cytolytic activity directed against recipient
hematopoietic cells. Clinical Cancer Research, 8,
2052–2060.
Humanized Anti-CD26 mAb Prevents Acute GVHD
ª 2013 John Wiley & Sons Ltd 277British Journal of Haematology, 2013, 162, 263–277




















