
Presence and distribution of ghrelin-immunopositive cells in the chicken gastrointestinal tract
7
Acta histochemica 107 (2005) 3—9 Presence and distribution of ghrelin- immunopositive cells in the chicken gastrointestinal tract Simona Neglia, Nadia Arcamone, Vincenzo Esposito, Giuliana Gargiulo, Paolo de Girolamo Department ofBiological Structures, Functions and Technologies, University of Naples ‘‘Federico II’’, Via Veterinaria 1, I-80137 Naples, Italy Received 16 June 2004; received in revised form 15 October 2004; accepted 17 December 2004 Summary The presence and distribution patterns of ghrelin, a gastric acylated peptide, were studied in the entire gastrointestinal tract of the chicken (Gallus domesticus) using the peroxidase–antiperoxidase immunohistochemical method, western blot analysis and a specific antibody against the C-terminal region of rat ghrelin. Ghrelin- immunopositive cells were observed in the mucosal layer of all segments examined. The largest numbers of ghrelin-positive cells were located at the base of lobuli of the proventriculus gland, along villi of the intestines and in crypts of the duodenum. Lower numbers of ghrelin-immunostained cells were located in crypts of jejunum and ileum and only few ghrelin-immunostained cells were detected at the base of crypts of the large intestine. Closed and open types of cells were observed in all segments. Western blot analysis confirmed the presence of ghrelin-like protein in the entire chicken gastrointestinal tract. The anatomical distribution patterns and the morphological characteristics of chicken ghrelin-positive cells suggest that they are endocrine cells. Furthermore, it is concluded that ghrelin shows a high degree of preservation during evolution from non-mammalian vertebrates to mammals. & 2005 Elsevier GmbH. All rights reserved. Introduction Growth-hormone secretagogues (GHSs) are small synthetic peptides and non-peptidyl molecules that stimulate growth hormone (GH) release both in vivo and in vitro from the anterior pitituary gland via a G-protein-coupled receptor (GHS-R; Bowers, 1998). Recently, a novel GHS-R ligand was isolated and ARTICLE IN PRESS www.elsevier.de/acthis KEYWORDS Ghrelin; Chicken; Gastrointestinal tract; Immunohistochemis- try; Western blot 0065-1281/$ - see front matter & 2005 Elsevier GmbH. All rights reserved. doi:10.1016/j.acthis.2004.12.001 Corresponding author. Tel.: +39081 2536114; fax: +39081 2536097. E-mail address: [email protected] (P. de Girolamo).
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Presence and distribution of ghrelin-immunopositive cells in the chicken gastrointestinal tract

ARTICLE IN PRESS
Acta histochemica 107 (2005) 3—9
KEYWORDGhrelin;Chicken;Gastrointetract;Immunohistry;Western b
0065-1281/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/acthis
Presence and distribution of ghrelin-immunopositive cells in the chickengastrointestinal tract
Simona Neglia, Nadia Arcamone, Vincenzo Esposito, Giuliana Gargiulo,Paolo de Girolamo�
Department of Biological Structures, Functions and Technologies, University of Naples ‘‘Federico II’’, Via Veterinaria 1,I-80137 Naples, Italy
Received 16 June 2004; received in revised form 15 October 2004; accepted 17 December 2004
S
stinal
tochemis-
lot
ee front matter & 200acthis.2004.12.001
ing author. Tel.: +39 0ess: [email protected]
SummaryThe presence and distribution patterns of ghrelin, a gastric acylated peptide, werestudied in the entire gastrointestinal tract of the chicken (Gallus domesticus) usingthe peroxidase–antiperoxidase immunohistochemical method, western blot analysisand a specific antibody against the C-terminal region of rat ghrelin. Ghrelin-immunopositive cells were observed in the mucosal layer of all segments examined.The largest numbers of ghrelin-positive cells were located at the base of lobuli of theproventriculus gland, along villi of the intestines and in crypts of the duodenum.Lower numbers of ghrelin-immunostained cells were located in crypts of jejunum andileum and only few ghrelin-immunostained cells were detected at the base of crypts ofthe large intestine. Closed and open types of cells were observed in all segments.Western blot analysis confirmed the presence of ghrelin-like protein in the entirechicken gastrointestinal tract. The anatomical distribution patterns and themorphological characteristics of chicken ghrelin-positive cells suggest that they areendocrine cells. Furthermore, it is concluded that ghrelin shows a high degree ofpreservation during evolution from non-mammalian vertebrates to mammals.& 2005 Elsevier GmbH. All rights reserved.
Introduction
Growth-hormone secretagogues (GHSs) are smallsynthetic peptides and non-peptidyl molecules that
5 Elsevier GmbH. All rights rese
81 2536114; fax: +39 081 253609t (P. de Girolamo).
stimulate growth hormone (GH) release both in vivoand in vitro from the anterior pitituary gland via aG-protein-coupled receptor (GHS-R; Bowers, 1998).Recently, a novel GHS-R ligand was isolated and
rved.
7.

ARTICLE IN PRESS
S. Neglia et al.4
characterized in rat (Kojima et al., 1999) andhuman (Date et al., 2000) stomach and was calledghrelin. Ghrelin is a 28-amino-acid peptide in whichthe third serine residue (Ser3) shows an n-octanoylmodification, essential for its biological activity(Kojima et al., 1999). Accumulating evidencesuggests that, besides a potent GH-releasingaction, ghrelin regulates food intake, energybalance, body weight, gastric motility and acidsecretion, function of the endocrine pancreas andglucose metabolism (Kamegai et al., 2000; Masudaet al., 2000; Wren et al., 2000; Broglio et al., 2001;Date et al., 2001; Kamegai et al., 2001; Nakazatoet al., 2001). Ghrelin is mainly produced inthe stomach (Kojima et al., 1999; Ariyasu et al.,2001). Besides the gastroenteric tract, ghrelinis also expressed in various peripheral tissues(Mori et al., 2000; Gualillo et al., 2001; Volanteet al., 2002; Andreis et al., 2003) and in thearcuate nucleus (ARC) of the hypothalamus, anappetite regulating centre (Lu et al., 2002). In situhybridization and immunohistochemical studiesindicated that ghrelin-producing cells are a distincttype of endocrine cells, known as X/A-like cells,that are localized in the mucosal epitheliumof corpus and fundus of the stomach (Date et al.,2000; Dornonville de la Cour et al., 2001; Sakataet al., 2002).
Few studies have been carried out on ghrelinexpression in non-mammalian vertebrates. Re-cently, ghrelin was found in the brain and stomachof both frog (Galas et al., 2002) and chicken (Kaiyaet al., 2002). Chicken ghrelin consists of 26 aminoacids with n-octanoyl modification at Ser3 and hasthe same sequence in the N-terminal region as in asthe human and rat form. In fact, administration ofchicken ghrelin increases plasma GH levels in bothrats and chicks, and increases plasma corticoster-one levels in growing chicks, at a lower dose thanmammalian ghrelin (Kaiya et al., 2002).
In a previous study, Ahmed and Harvey (2002)showed ghrelin-immunostaining in the chickenhypothalamus with an antibody developed espe-cially for radioimmunoassays and raised in rabbitthat recognizes the C-terminal region of rat ghrelin(RAB-031-31; Phoenix Pharmaceuticals, Belmont,CA, USA). Recently, Wada et al. (2003) demon-strated ghrelin-positive cells in the chicken pro-ventriculus using an N-terminal region-recognizingantibody only. Because mammalian ghrelin is alsoexpressed in the intestines (Date et al., 2000), westudied distribution patterns of ghrelin in both theproventriculus and small and large intestines ofchicken using immunohistochemistry and westernblotting and a specific polyclonal antibody againstthe C-terminal region of rat ghrelin.
Material and methods
Immunohistochemistry
Three-month-old chickens (Gallus domesticus) ofboth sexes were used in the present study. Thegastrointestinal tract was sectioned in small samplesof the following segments: proventriculus, ventricu-lus, duodenum, jejunum, ileum, caecum and colo-rectum. The tissues were immediately fixed inBouin’s solution for 24h, dehydrated in a gradedseries of alcohol, embedded in paraffin wax andtransverse 6–7-mm-thick serial sections were cut.After dewaxing, sections were processed for im-munohistochemistry using the peroxidase–antiper-oxidase (PAP) method according to Sternberger(1986). Endogenous peroxidase activity was blockedby treating the sections with 0.3% hydrogen per-oxide for 20min at room temperature (RT) and thenrinsed for 15min in 0.01M phosphate-bufferedsaline (PBS), pH 7.4. Sections were incubated for30min at RT with normal goat serum (dilution, 1:5;Jackson Immunoresearch Laboratories, West Grove,PA, USA) and then with primary antibody (18 h in amoist dark chamber at 4 1C). The primary antibody, arabbit antibody that recognizes the C-terminalregion of rat ghrelin and cross-reacts with chickenghrelin (H-031-31; Phoenix Pharmaceuticals), wasdiluted 1:750. After incubation, sections were rinsedin PBS for 15min and, incubated for 30min at RTwith goat anti-rabbit IgG (dilution, 1:50; VectorLaboratories, Burlingame, CA, USA). Subsequently,sections were rinsed in PBS for 15min and thenincubated for 30min at RT with PAP complex(dilution, 1:100; Dako, CA, USA). Peroxidase activitywas visualized with a solution of 10mg 3-30-diaminobenzidine tetrahydrocloride (DAB; Sigma,St. Louis, MO, USA) in 15ml 0.5M Tris buffer, pH7.6, containing 0.03% hydrogen peroxide.
Specificity of the immunohistochemical stainingreaction was tested as follows: (1) substitution ofeither the ghrelin antibody or the anti-rabbit IgG,or the PAP complex by PBS (negative controls); (2)application of rat stomach tissue known to bepositive for ghrelin (positive controls); (3) previousabsorption of the primary antibody with homolo-gous antigen (031-31; Phoenix Pharmaceuticals; upto 50 mg/ml antibody in the final dilution).
Sections were examined with a Aristoplan lightmicroscope (Leica, Wetzlar, Germany) equippedwith a DC300F digital camera (Leica).
Western blot analysis
Samples of all parts of the gastrointestinaltract were homogenized (1:2w/v) with a Potter

ARTICLE IN PRESS
Ghrelin in the chicken gastrointestinal tract 5
homogenizer in cold RIPA buffer [50mM Tris-HCl, pH8.0, 0.1% sodium dodecyl sulfate (SDS), 1% NP-40,150mM NaCl and 0.5% deoxycholic acid] containing1mM phenylmethylsulfonyl fluoride (Sigma) and100� inhibitor cocktail. Then, suspensions werecentrifuged at 14,000g for 15min at 4 1C and theprotein concentrations of the resulting pellets,divided in equal aliquots, were determined by theDC protein assay (Bio-Rad Laboratories, Hercules,CA, USA). Samples were separated by electrophor-esis on 10% SDS-polyacrylamide gels (NuPAGE Novex10% bis-Tris gels; Invitrogen, Carlsbad, CA, USA)and, then, gels were transferred onto nitrocellu-lose membranes (Millipore, Billerica, MA, USA).Membranes were blocked for 1 h at RT in PBS-Tweenbuffer (0.01M PBS and 0.05% Tween-20) containing5% non-fat milk and incubated for 1.5 h at RT withthe primary antibody, as described above, diluted1:1500 in PBS-Tween. After washing, membraneswere incubated for 1 h at RT with polyclonal goatanti-rabbit IgG conjugated with horseradish perox-idase (dilution, 1:2000; DAKO). Membranes wereincubated in chemoluminescence reagent (ECL;Amersham, Little Chalfont, England) to visualizeimmunocomplexes, and were exposed for 1–30 s tohyperfilm.
Results
Immunohistochemistry
Ghrelin-immunopositive cells (ghrelin-ic) wereobserved in mucosal layers of all gastrointestinaltract segments examined: proventriculus, duode-num, jejunum, ileum, caecum and colo-rectum.
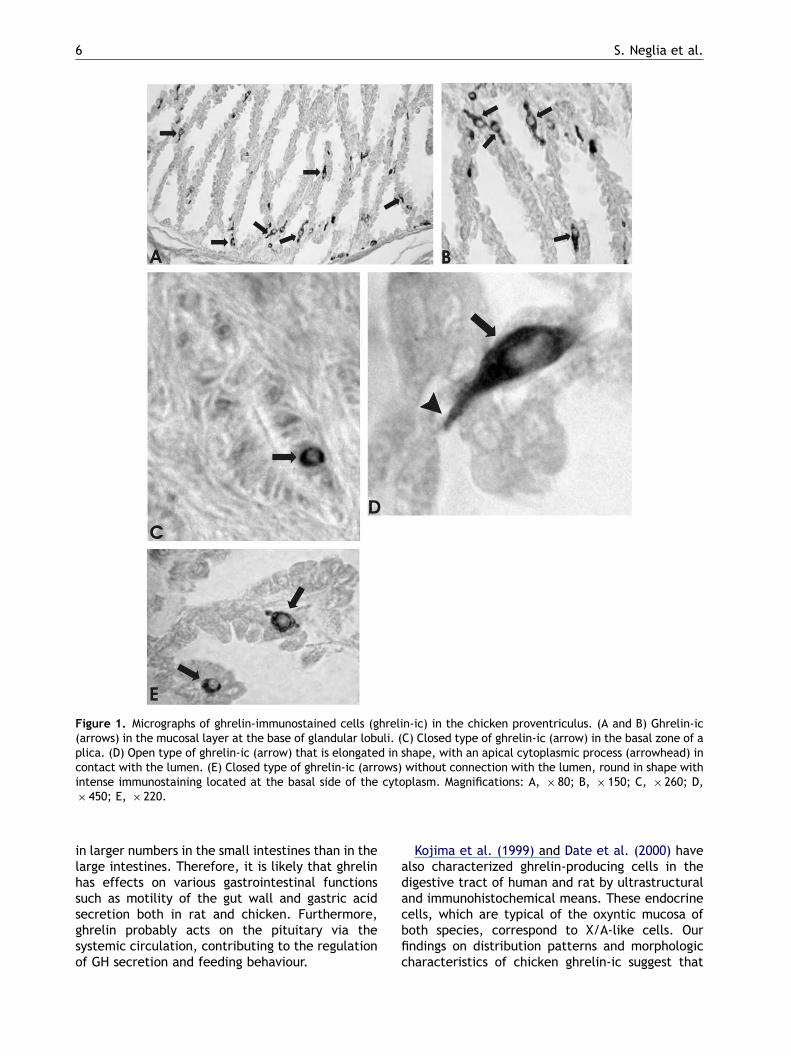
High numbers of strongly stained ghrelin-ic werefound at the base of glandular lobuli of theproventriculus (Fig. 1A and B). In contrast, fewghrelin-ic were observed in the basal zone ofproventriculus plicae (Fig. 1C), which showed anintense immunostaining that was restricted to thebasal region of the cytoplasm. These cells had asmall round shape and were considered to be of theclosed cell type (Fig. 1D). Few ghrelin-ic were ofthe open cell type that showed an elongated shapeand apical cytoplasmic processes in contact withthe lumen (Fig. 1E).
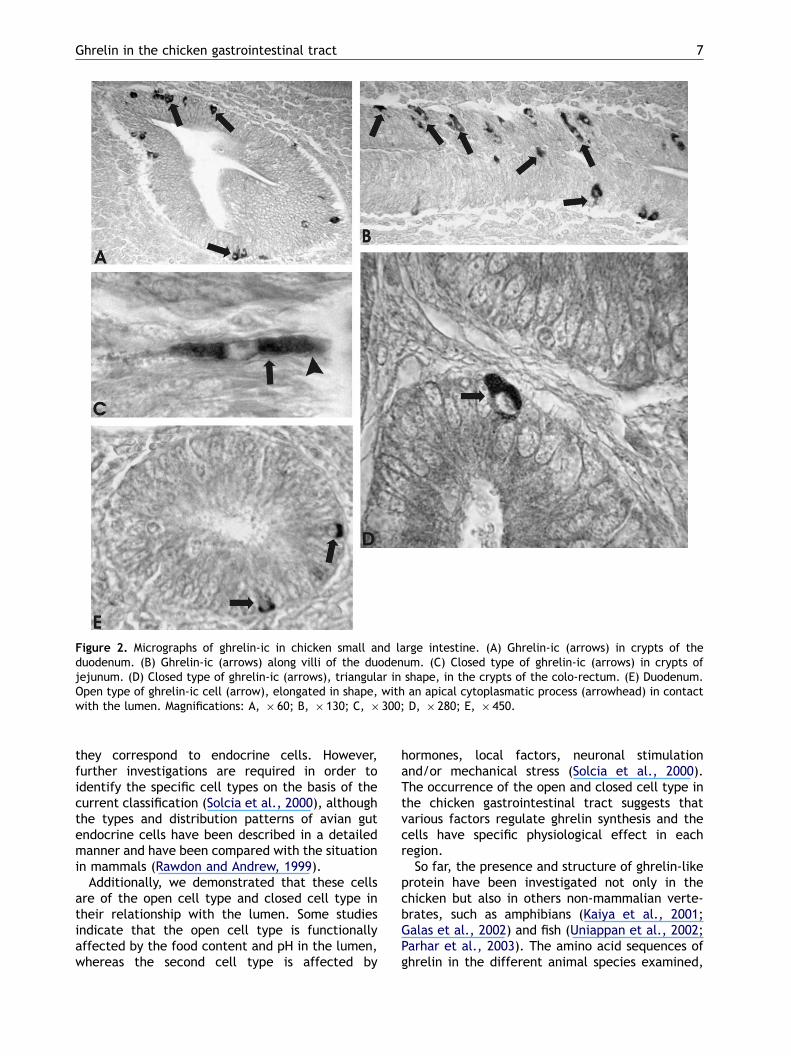
Many ghrelin-ic were found to be scattered inepithelium of crypts (Fig. 2A) and along villi of theduodenum (Fig. 2B). Closed and open cell types(Fig. 2C) were both observed in this intestinalsegment as well. Fewer ghrelin-ic were found inepithelia of crypts of the jejunum and ileum andgenerally they appeared to be of the closed cell
type (Fig. 2D). Very few ghrelin-ic, and especiallyof the closed cell type and triangular in shape,were detected in crypts of the large intestine(Fig. 2E).
Negative controls and sections stained with pre-absorbed antibodies did not show any specificstaining. Specificity of immunostaining was con-firmed by the positive controls (data not shown).
Western blot analysis
Ghrelin-like protein was detected by westernblot analysis of homogenates of all segments of thechicken gastrointestinal tract which was in agree-ment with our immunohistochemical data. Theghrelin-like protein that was detected had anestimated molecular mass of approximately44–46 kDa (Fig. 3).
Discussion
In the present study, we demonstrated theoccurrence and distribution patterns of ghrelin-likeprotein in the entire chicken gastrointestinal tractusing western blot analysis and peroxidase–anti-peroxidase immunohistochemistry respectively.Our findings are not in agreement with a recentstudy of Ahmed and Harvey (2002) who foundghrelin immunostaining to be present in thechicken brain only and not in the gastrointestinaltract. This discrepancy may be due to a differencein antibodies used in the two studies. In fact, bothantibodies recognize the C-terminal region ofghrelin, but we used an antibody especially devel-oped for immunohistochemistry, whereas Ahmedand Harvey (2002) used an antibody that wasdeveloped especially for radioimmunoassays. Thepresence of ghrelin-ic in the chicken gastrointest-inal tract suggests that this peptide may act as agastrointestinal hormone besides its function as aneuropeptide (Ahmed and Harvey, 2002).
Recently, Wada et al. (2003) demonstratedghrelin immunostaining in the mucosal layer ofthe chicken proventriculus. In agreement withthese results, we observed the largest number ofghrelin-ic to be present in the chicken proventri-culus which corresponds to the gastric fundus inmammals. Somewhat lower numbers of ghrelin-icwere found in jejunum, ileum and large intestine.Our findings agree with distribution patternsreported in human and rat digestive tract. In fact,Kojima et al. (1999) and Date et al. (2000) foundghrelin-ic to be mainly located in the oxynticmucosa of gastric fundus. These cells were present
ARTICLE IN PRESS
Figure 1. Micrographs of ghrelin-immunostained cells (ghrelin-ic) in the chicken proventriculus. (A and B) Ghrelin-ic(arrows) in the mucosal layer at the base of glandular lobuli. (C) Closed type of ghrelin-ic (arrow) in the basal zone of aplica. (D) Open type of ghrelin-ic (arrow) that is elongated in shape, with an apical cytoplasmic process (arrowhead) incontact with the lumen. (E) Closed type of ghrelin-ic (arrows) without connection with the lumen, round in shape withintense immunostaining located at the basal side of the cytoplasm. Magnifications: A, � 80; B, � 150; C, � 260; D,� 450; E, � 220.
S. Neglia et al.6
in larger numbers in the small intestines than in thelarge intestines. Therefore, it is likely that ghrelinhas effects on various gastrointestinal functionssuch as motility of the gut wall and gastric acidsecretion both in rat and chicken. Furthermore,ghrelin probably acts on the pituitary via thesystemic circulation, contributing to the regulationof GH secretion and feeding behaviour.
Kojima et al. (1999) and Date et al. (2000) havealso characterized ghrelin-producing cells in thedigestive tract of human and rat by ultrastructuraland immunohistochemical means. These endocrinecells, which are typical of the oxyntic mucosa ofboth species, correspond to X/A-like cells. Ourfindings on distribution patterns and morphologiccharacteristics of chicken ghrelin-ic suggest that
ARTICLE IN PRESS
Figure 2. Micrographs of ghrelin-ic in chicken small and large intestine. (A) Ghrelin-ic (arrows) in crypts of theduodenum. (B) Ghrelin-ic (arrows) along villi of the duodenum. (C) Closed type of ghrelin-ic (arrows) in crypts ofjejunum. (D) Closed type of ghrelin-ic (arrows), triangular in shape, in the crypts of the colo-rectum. (E) Duodenum.Open type of ghrelin-ic cell (arrow), elongated in shape, with an apical cytoplasmatic process (arrowhead) in contactwith the lumen. Magnifications: A, � 60; B, � 130; C, � 300; D, � 280; E, � 450.
Ghrelin in the chicken gastrointestinal tract 7
they correspond to endocrine cells. However,further investigations are required in order toidentify the specific cell types on the basis of thecurrent classification (Solcia et al., 2000), althoughthe types and distribution patterns of avian gutendocrine cells have been described in a detailedmanner and have been compared with the situationin mammals (Rawdon and Andrew, 1999).
Additionally, we demonstrated that these cellsare of the open cell type and closed cell type intheir relationship with the lumen. Some studiesindicate that the open cell type is functionallyaffected by the food content and pH in the lumen,whereas the second cell type is affected by
hormones, local factors, neuronal stimulationand/or mechanical stress (Solcia et al., 2000).The occurrence of the open and closed cell type inthe chicken gastrointestinal tract suggests thatvarious factors regulate ghrelin synthesis and thecells have specific physiological effect in eachregion.
So far, the presence and structure of ghrelin-likeprotein have been investigated not only in thechicken but also in others non-mammalian verte-brates, such as amphibians (Kaiya et al., 2001;Galas et al., 2002) and fish (Uniappan et al., 2002;Parhar et al., 2003). The amino acid sequences ofghrelin in the different animal species examined,

ARTICLE IN PRESS
Figure 3. Western blot analysis of ghrelin in extracts ofchicken gastrointestinal tract. Lane 1, protein markers;lane 2, homogenate of gastrointestinal tract showing astrongly stained band at approximately 44–46 kDa.
S. Neglia et al.8
show a high preservation of the ‘‘active core’’(GSSF), essential for its molecular biological activ-ity, in tilapia, eel and chicken (Kaiya et al., 2002;2003a, b), but not bullfrog (GLTF; Kaiya et al.,2001) and goldfish (GTSF; Uniappan et al., 2002).Interestingly, these first four amino acids arepreserved in human and rat as well (Kojima etal., 1999). These findings show that ghrelin isphylogenetically highly preserved from non-mam-malian species to mammals not only in its aminoacid sequence but also in its distribution patterns.
References
Ahmed S, Harvey S. Ghrelin: a hypotalamic GH-releasingfactor in domestic fowl (Gallus domesticus). JEndocrinol 2002;172:117–25.
Andreis PG, Malendowicz LK, Trejter M, Neri G, SpinazziR, Rossi GP, Nussdorfer GG. Ghrelin and growthhormone secretagogue receptor are expressed in therat adrenal cortex:evidence that ghrelin stimulatesthe growth, but not the secretory activity of adrenalcells. FEBS Lett 2003;536:173–9.
Ariyasu H, Takaya K, Tagami T, Ogawa Y, Hosoda K,Akamizu T, Suda M, Koh T, Natsui K, Toyooka S,Shirakami G, Usui T, Shimatsu A, Doi K, Hosoda H,Kojima M, Kangawa K, Nakao K. Stomach is a majorsource of circulating ghrelin, and feeling statedetermines plasma ghrelin-like immunoreactivity inhumans. J Clin Endocrinol Metab 2001;86:4753–8.
Bowers CY. Growth hormone-releasing peptide (GHRP).Cell Mol Life Sci 1998;54:1316–29.
Broglio F, Arvat E, Benso A, Gottero C, Cuccioli G, PapottiM, van der Lely AJ, Deghenghi R, Ghigo E. Ghrelin, anatural GH secretagogue produced by the stomach,induces hyperglycemia and reduces insulin secretionin humans. J Clin Endocrinol Metab 2001;86:5083–6.
Date Y, Kojima M, Hosoda H, Sawaguchi A, Mondal MS,Suganuma T, Matsukura S, Kangawa K, Nakazato M.Ghrelin, a novel growth hormone-releasing acylatedpeptide, is synthesized in a distinct endocrine celltype in the gastrointestinal tracts of rats and human.Endocrinology 2000;141:4255–66.
Date Y, Nakazato M, Murakami N, Kojima M, Kangawa K,Matsukura S. Ghrelin acts in the central nervoussystem to stimulate gastric acid secretion. BiochemBiophys Res Commun 2001;280:904–7.
Dornonville de la Cour C, Bjorkqvist M, Sandvik AK, BakkeI, Zhao CM, Chen D, Hakanson R. A-like cells in the ratstomach contain ghrelin and do not operate undergastrin control. Regul Pept 2001;99:141–50.
Galas L, Chartrel N, Kojima M, Kangawa K, Vaudry H.Immunohistochemical localization and biochemicalcharacterization of ghrelin in the brain and stomachof the frog Rana esculenta. J Comp Neurol 2002;450:34–44.
Gualillo O, Caminos JE, Blanco M, Garcia-Caballero T,Kojima M, Kangawa K, Dieguez C, Casanueva FF.Ghrelin, a novel placental-derived hormone. Endocri-nology 2001;142:788–94.
Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H,Wakabayashi I. Central effect of ghrelin, an endogen-ous growth hormone secretagogue, on hypothalamicpeptide gene expression. Endocrinology 2000;141:4797–800.
Kamegai J, Tamura H, Shimizu T, Ishii S, Sugihara H,Wakabayashi I. Chronic central infusion of ghrelinincreases hypotalamic neuropeptide Y and agouti-related protein mRNA levels and body weight in rats.Diabetes 2001;50:2438–43.
Kaiya H, Kojima M, Hosoda H, Koda A, Yamamoto K,Kitajima Y, Matsumoto M, Minamitake Y, Kikuyama S,Kangawa K. Bullfrog ghrelin is modified by n-octanoicacid at the third threonine residue. J Biol Chem2001;276:40,441–8.
Kaiya H, Van Der Geyten S, Kojima M, Hosoda H, KitajimaY, Matsumoto M, Geelissen S, Darras VM, Kangawa K.Chicken ghrelin: purification, cDNA cloning and biolo-gical activity. Endocrinology 2002;143:3454–63.
Kaiya H, Kojima M, Hosoda H, Riley LG, Hirano T, GrauEG, Kangawa K. Identification of tilapia ghrelin and itseffects on growth hormone and prolactin release inthe tilapia, Oreochromis mossambicus. Comp BiochemPhysiol B. Biochem Mol Biol 2003a;135:421–9.
Kaiya H, Kojima M, Hosoda H, Riley LG, Hirano T, GrauEG, Kangawa K. Amidated fish ghrelin: purification,cDNA cloning in the Japanese eel and its biologicalactivity. J Endocrinol 2003b;176(3):415–23.
Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H,Kangawa K. Ghrelin is a growth-hormone-releasing

ARTICLE IN PRESS
Ghrelin in the chicken gastrointestinal tract 9
acylated peptide from stomach. Nature 1999;402:656–60.
Lu S, Guan JL, Wang QP, Uehara K, Yamada S, Goto N,Date Y, Nakazato M, Kojima M, Kangawa K, Shioda S.Immunocytochemical observation of ghrelin-contain-ing neurons in the rat arcuate nucleus. Neurosci Lett2002;321:157–60.
Masuda Y, Tanaka T, Inomata N, Ohnuma N, Tanaka S, ItohZ, Hosoda H, Kojima M, Kangawa K. Ghrelin stimulatesgastric acid secretion and motility in rats. BiochemBiophys Res Commun 2000;276:905–8.
Mori K, Yoshimoto A, Takaya K, Hosoda K, Ariyasu H,Yahata K, Mukoyama M, Sugawara A, Hosoda H, KojimaM, Kangawa K, Nakao K. Kidney produces a novelacylated peptide, ghrelin. FEBS Lett 2000;486:213–6.
Nakazato M, Murakami N, Date Y, Kojima M, Matsuo H,Kangawa K, Matsukura S. A role for ghrelin in thecentral regulation of feeding. Nature 2001;409:194–8.
Parhar IS, Sato H, Sakuma Y. Ghrelin gene fish ismodulated by sex and development. Biochem BiophysRes Commun 2003;305:169–75.
Rawdon BB, Andrew A. Gut endocrine cells in birds: anoverview, with particular reference to the chemistryof gut peptides and the distribution, ontogeny,embryonic origin and differentiation of the endocrinecells. Prog Histochem Cytochem 1999;34:3–82.
Sakata I, Nakamura K, Yamakazi M, Matsubara M, HayashiY, Kangawa K, Sakai T. Ghrelin-producing cells exist as
two types of cells, closed- and opened-type cells, inthe rat gastrointestinal tract. Peptides 2002;23:531–6.
Solcia E, Rindi G, Buffa R, Fiocca R, Capella C. Gastricendocrine cells: types, function and growth. RegulPept 2000;93:31–5.
Sternberger LA. Immunocytochemistry, 3rd ed. New York,NY, USA: Wiley; 1986.
Uniappan S, Lin X, Cervini L, Rivier J, Kaya H, Kangawa K,Peter RE. Goldfish ghrelin: molecular characterizationof the complementary deoxyribonucleic acid, partialgene structure and evidence for its stimulatory role infood intake. Endocrinology 2002;143:4143–6.
Volante M, Allia E, Gugliotta P, Funaro A, Broglio F,Deghenghi R, Muccioli G, Ghigo E, Papotti M. Expres-sion of ghrelin and of GHS receptor by pancreatic isletcells and related endocrine tumors. J Clin EndocrinolMetab 2002;279:E695–706.
Wada R, Sakata I, Kaiya H, Nakamura K, Hayashi Y,Kangawa K, Sakai T. Existence of ghrelin-immunopo-sitive and -expressing cells in the proventriculus of thehatching and adult chicken. Regul Pept 2003;111:123–8.
Wren AM, Small CJ, Ward HL, Murphy KG, Dakin CL, TaheriS, Kennedy AR, Roberts GH, Morgan DCA, Ghatei MA,Bloom SR. The novel hypothalamic peptide ghrelinstimulates food intake and growth hormone secretion.Endocrinology 2000;141:4325–8.




















