
Posttranslational Regulation of IRF-4 Activity by the Immunophilin FKBP52
12
Immunity, Vol. 12, 129–140, February, 2000, Copyright 2000 by Cell Press Posttranslational Regulation of IRF-4 Activity by the Immunophilin FKBP52 (DBD). However, IRF-4 was exclusively found in B lym- phocytes associated with PU.1, an Ets family member involved in lymphoid and myeloid lineage development. Yae ¨ l Mamane, Sonia Sharma, Louisa Petropoulos, Rongtuan Lin, and John Hiscott* Terry Fox Molecular Oncology Group The IRF-4.PU.1 heterodimer formed a transcriptional ac- tivating complex. Independently, another group cloned Lady Davis Institute for Medical Research and Departments of Microbiology and Immunology, IRF-4 as lymphoid-specific IRF (LSIRF), which was ex- pressed at all stages of B cell development and in mature Medicine, and Oncology McGill University T cells. LSIRF expression was upregulated by receptor- cross-linking stimuli such as plant lectins, CD3, or IgM. Montreal, Canada H3T 1E2 In contrast to B cells, IRF-4 in T cells was able to bind autonomously to the ISRE of the MHC class I promoter (Matsuyama et al., 1995). A third group isolated a human Summary equivalent of IRF-4 from an adult T cell leukemia (ATL) cell line, called IFN consensus sequence-binding protein Interferon regulatory factor-4 (IRF-4) plays an impor- in adult T cell leukemia cell lines or activated T cells tant role in immunoregulatory gene expression in B and (ICSAT) (Yamagata et al., 1996). The human T cell leuke- T lymphocytes and is also highly expressed in human T mia virus-I (HTLV-I) is the etiologic agent of adult T cell cell leukemia virus type 1 infected cells. In this study, leukemia (ATL), an aggressive and fatal leukemia of we characterize a novel interaction between IRF-4 and CD4 1 T lymphocytes (Poiez et al., 1980; Yoshida et al., the FK506-binding protein 52 (FKBP52), a 59 kDa 1982) and is also associated with a neurological demye- member of the immunophilin family with peptidyl-prolyl linating disease, tropical spastic paraparesis (Gessain isomerase activity (PPIase). IRF-4-FKBP52 association et al., 1985; Daenkes et al., 1990). The oncogenic poten- inhibited IRF4-PU.1 binding to the immunoglobulin tial of HTLV-I resides in the 353–amino acid (aa), 40 kDa light chain enhancer E l2–4 as well as IRF-4-PU.1 trans- viral Tax oncoprotein, a positive regulator of viral gene activation, effects that were dependent on functional transcription. ICSAT/IRF-4 was shown to be expressed PPIase activity. FKBP52 association also resulted in a exclusively in a restricted subset of lymphocytes: only structural modification of IRF-4, detectable by immu- T cells treated with phorbol myristate acetate (PMA) noblot analysis and by IRF-4 partial proteolysis. These or infected with HTLV-1 produced IRF-4. Jurkat cells results demonstrate a novel posttranslational mecha- transiently transfected with the HTLV-1 tax gene also nism of transcriptional control, mediated through the expressed IRF-4, indicating that Tax may function as an interaction of an immunophilin with a transcriptional indirect activator of the IRF-4 promoter (Yamagata et regulator. al., 1996). IRF-4 (Pip/LSIRF/ICSAT) is structurally similar to another member of the IRF family, IRF-8 (formerly Introduction called ICSBP), and expression is not inducible by IFN. However, human IRF-4 possessed a very different func- The interferon regulatory factors (IRFs) are a family of tion compared to its murine counterpart; while IRF-4- transcription factors involved in the early host response PU.1 functioned as a transactivator complex, ICSAT to pathogens, immunomodulation, and hematopoietic exerted an IRF-2 and ICSBP-like repressive effect on development (Nguyen et al., 1997; Stark et al., 1998; IFN- and IRF-1-induced gene activation. Mamane et al., 1999). The best characterized members In this paper, a novel physical and functional inter- of this family, IRF-1 and IRF-2, were originally identified action between IRF-4 and FK506-binding protein 52 through studies on the transcriptional regulation of the (FKBP52), an immunophilin with peptidyl-prolyl iso- human IFNb gene (Fujita et al., 1988; Miyamoto et al., merase (PPIase) and chaperone-like activities, has been 1988). This family now includes seven additional mem- characterized. FKBP52 inhibited IRF-4 DNA binding and bers: IRF-3 through IRF-9 (Mamane et al., 1999), as well transactivation functions by a posttranslational modifi- as virally encoded IRF homologs in the human herpes cation of IRF-4 that was dependent on the PPIase activ- virus 8 (HHV-8) genome (Moore et al., 1996; Burysek et ity of FKBP52. These results suggest that FKBP52 in- al., 1999). Structurally, all IRF family members share a duces a conformational change in IRF-4 by cis-trans tryptophan repeat motif in the N-terminal DNA-binding prolyl isomerization that interferes with IRF-4 DNA bind- domain and homology with the Myb oncoproteins. ing and transcriptional activity, thus demonstrating a One of the recently discovered members of the IRF novel posttranslational mechanism controlling gene family, IRF-4, was cloned from three independent transcription. sources. First cloned as PU.1 interacting partner (Pip), which binds to the murine immunoglobulin (Ig) light Results chain enhancer E l2–4 (Eisenbeis et al., 1993, 1995), IRF-4 possesses an IRF-like N-terminal DNA-binding domain In Vivo Interaction of IRF-4 and FKBP52 To identify potential partners of IRF-4, a yeast two-hybrid analysis was performed; since full-length IRF-4 pos- * To whom correspondence should be addressed (e-mail: mijh@ musica.mcgill.ca). sessed intrinsic transcriptional activity, the bait utilized
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Posttranslational Regulation of IRF-4 Activity by the Immunophilin FKBP52

Immunity, Vol. 12, 129–140, February, 2000, Copyright 2000 by Cell Press
Posttranslational Regulation of IRF-4Activity by the Immunophilin FKBP52
(DBD). However, IRF-4 was exclusively found in B lym-phocytes associated with PU.1, an Ets family memberinvolved in lymphoid and myeloid lineage development.
Yael Mamane, Sonia Sharma,Louisa Petropoulos, Rongtuan Lin,and John Hiscott*Terry Fox Molecular Oncology Group The IRF-4.PU.1 heterodimer formed a transcriptional ac-
tivating complex. Independently, another group clonedLady Davis Institute for Medical Research andDepartments of Microbiology and Immunology, IRF-4 as lymphoid-specific IRF (LSIRF), which was ex-
pressed at all stages of B cell development and in matureMedicine, and OncologyMcGill University T cells. LSIRF expression was upregulated by receptor-
cross-linking stimuli such as plant lectins, CD3, or IgM.Montreal, Canada H3T 1E2In contrast to B cells, IRF-4 in T cells was able to bindautonomously to the ISRE of the MHC class I promoter(Matsuyama et al., 1995). A third group isolated a humanSummaryequivalent of IRF-4 from an adult T cell leukemia (ATL)cell line, called IFN consensus sequence-binding proteinInterferon regulatory factor-4 (IRF-4) plays an impor-in adult T cell leukemia cell lines or activated T cellstant role in immunoregulatory gene expression in B and(ICSAT) (Yamagata et al., 1996). The human T cell leuke-T lymphocytes and is also highly expressed in human Tmia virus-I (HTLV-I) is the etiologic agent of adult T cellcell leukemia virus type 1 infected cells. In this study,leukemia (ATL), an aggressive and fatal leukemia ofwe characterize a novel interaction between IRF-4 andCD41 T lymphocytes (Poiez et al., 1980; Yoshida et al.,the FK506-binding protein 52 (FKBP52), a 59 kDa1982) and is also associated with a neurological demye-member of the immunophilin family with peptidyl-prolyllinating disease, tropical spastic paraparesis (Gessainisomerase activity (PPIase). IRF-4-FKBP52 associationet al., 1985; Daenkes et al., 1990). The oncogenic poten-inhibited IRF4-PU.1 binding to the immunoglobulintial of HTLV-I resides in the 353–amino acid (aa), 40 kDalight chain enhancer El2–4 as well as IRF-4-PU.1 trans-viral Tax oncoprotein, a positive regulator of viral geneactivation, effects that were dependent on functionaltranscription. ICSAT/IRF-4 was shown to be expressedPPIase activity. FKBP52 association also resulted in aexclusively in a restricted subset of lymphocytes: onlystructural modification of IRF-4, detectable by immu-T cells treated with phorbol myristate acetate (PMA)noblot analysis and by IRF-4 partial proteolysis. Theseor infected with HTLV-1 produced IRF-4. Jurkat cellsresults demonstrate a novel posttranslational mecha-transiently transfected with the HTLV-1 tax gene alsonism of transcriptional control, mediated through theexpressed IRF-4, indicating that Tax may function as aninteraction of an immunophilin with a transcriptionalindirect activator of the IRF-4 promoter (Yamagata etregulator.al., 1996). IRF-4 (Pip/LSIRF/ICSAT) is structurally similarto another member of the IRF family, IRF-8 (formerly
Introduction called ICSBP), and expression is not inducible by IFN.However, human IRF-4 possessed a very different func-
The interferon regulatory factors (IRFs) are a family of tion compared to its murine counterpart; while IRF-4-transcription factors involved in the early host response PU.1 functioned as a transactivator complex, ICSATto pathogens, immunomodulation, and hematopoietic exerted an IRF-2 and ICSBP-like repressive effect ondevelopment (Nguyen et al., 1997; Stark et al., 1998; IFN- and IRF-1-induced gene activation.Mamane et al., 1999). The best characterized members In this paper, a novel physical and functional inter-of this family, IRF-1 and IRF-2, were originally identified action between IRF-4 and FK506-binding protein 52through studies on the transcriptional regulation of the (FKBP52), an immunophilin with peptidyl-prolyl iso-human IFNb gene (Fujita et al., 1988; Miyamoto et al., merase (PPIase) and chaperone-like activities, has been1988). This family now includes seven additional mem- characterized. FKBP52 inhibited IRF-4 DNA binding andbers: IRF-3 through IRF-9 (Mamane et al., 1999), as well transactivation functions by a posttranslational modifi-as virally encoded IRF homologs in the human herpes cation of IRF-4 that was dependent on the PPIase activ-virus 8 (HHV-8) genome (Moore et al., 1996; Burysek et ity of FKBP52. These results suggest that FKBP52 in-al., 1999). Structurally, all IRF family members share a duces a conformational change in IRF-4 by cis-transtryptophan repeat motif in the N-terminal DNA-binding prolyl isomerization that interferes with IRF-4 DNA bind-domain and homology with the Myb oncoproteins. ing and transcriptional activity, thus demonstrating a
One of the recently discovered members of the IRF novel posttranslational mechanism controlling genefamily, IRF-4, was cloned from three independent transcription.sources. First cloned as PU.1 interacting partner (Pip),which binds to the murine immunoglobulin (Ig) light Resultschain enhancer El2–4 (Eisenbeis et al., 1993, 1995), IRF-4possesses an IRF-like N-terminal DNA-binding domain In Vivo Interaction of IRF-4 and FKBP52
To identify potential partners of IRF-4, a yeast two-hybridanalysis was performed; since full-length IRF-4 pos-* To whom correspondence should be addressed (e-mail: mijh@
musica.mcgill.ca). sessed intrinsic transcriptional activity, the bait utilized

Immunity130
Figure 1. Interactions between IRF-4 and FKBP52
(A) Schematic representation of FKBP52. FKBP52 possesses three functional domains (brackets): FKBP-like domain 1 with the PPIase activity(aa 1–148), FKBP-like domain 2 with an ATP/GTP binding site (aa 148–267), and three tetratricopeptide repeats (aa 267–389), which mediateprotein–protein interactions.(B–D) The proline-rich region of IRF-4 (aa 150–237) interacts with the tetratricopeptide repeats of FKBP52 (aa 233–459). COS-7 cells weretransfected with Myc-tagged FKBP52 and FLAG- or HA-tagged IRF-4 constructs (15 mg each), and whole-cell extracts (WCE) (300 mg) were

FKBP52 Inhibits IRF-4 Transcriptional Activity131
was IRF-4 aa 150–410 comprising a proline-rich domain this domain alone was not stably expressed as a taggedpeptide and therefore was not tested directly, hence theand the C-terminal IRF association domain (IAD). Among
the 106 clones screened, 10 clones were positive for parentheses in Figure 1E (Y. M., unpublished data).Interestingly, with the inclusion of additional C-termi-mating and were subsequently sequenced for identifica-
tion. Two of these clones corresponded to full-length nal amino acids beyond aa 410, FKBP52 did not associ-ate with IRF-4 (aa 1–439), IRF-4 (aa 1–419) (Figure 1D,cDNA of the human immunophilin FKBP52, a member
of the immunophilin family of proteins with peptidyl- lanes 2 and 4), or with full-length IRF-4 (Y. M., unpub-lished data); similarly, IRF-4 did not interact with theprolyl isomerase (PPIase) activity and chaperone-like
functions (Czar et al., 1994; Radanyi et al., 1994; Perrot- N- or C-terminal parts of FKBP52 (Y. M., unpublisheddata). The conformation of IRF-4 may be inaccessible toApplanat et al., 1995). FKBP52 contains three “FKBP-
like” domains: domain 1 possesses features required FKBP52 because of structural constraints imposed bythe C-terminal autoinhibition element, which folds backfor PPIase activity and FK506 binding, domain 2 con-
tains GTP- and ATP-binding sites, and domain 3 con- on the N-terminal DNA-binding domain (Eisenbeis et al.,1995; Brass et al., 1996, 1999).tains the tetratricopeptide repeats (TPR) that are respon-
sible for the association with HSP90 (Figure 1A) (Radanyiet al., 1994; Kay, 1996).
In Vivo Interaction between IRF-4 and FKBP52To determine which domain of FKBP52 interacted
in HTLV-1-Transformed Cells and in B Cellswith IRF-4, FKBP52 was truncated to an N-terminal frag-
IRF-4 has been previously shown to be expressed in allment (N-FKBP52 [aa 1–243]) containing domains 1 and
Tax-expressing HTLV-I-transformed cells as well as in2 and a C-terminal fragment (DN-FKBP52 [aa 233–459])
B cells (Eisenbeis et al., 1995; Matsuyama et al., 1995;containing the TPR domains, and both forms were sub-
Brass et al., 1996; Yamagata et al., 1996). To investigatecloned into a Myc-tagged vector. In order to confirm
IRF-4-FKBP52 interactions, three HTLV-I-transformed Tthe interaction between FKBP52 and IRF-4, Myc-tagged
cell lines, C8166, MT-2, and MT-4; one HTLV-II-trans-forms of FKBP52 and FLAG-tagged IRF-4 (aa 150–410)
formed T cell line MO-T; and five B lymphoid cell lineswere cotransfected into COS-7 cells. After immunopre-
were examined. C8166, MT-2, and MO-T expressed highcipitation of Myc-FKBP52 from cell extracts with anti-
amounts of IRF-4 whereas MT-4, which does not ex-Myc antibody, immunoblot analysis using a-FLAG anti-
press Tax (Jeang et al., 1997), contained no IRF-4 (Figurebody revealed that IRF-4 (aa 150–410) coprecipitated
2A, upper panel, lanes 1–3). The levels of FKBP52 werewith full-length FKBP52 (Figure 1B, lane 2) as well as
approximately equal in all the T cell lines (Figure 2A,the C-terminal fragment (Figure 1B, lane 4) but did not
lower panel). Coprecipitation assays using an a-FKBP52coimmunoprecipitate with the N-terminal 1–243 aa frag-
antibody revealed that endogenous IRF-4 interactedment (Figure 1B, lane 6), indicating TPR-mediated pro-
with endogenous FKBP52 in C8166, MT-2, and MO-Ttein–protein interactions between FKBP52 and IRF-4.
but not in the MT-4 cell line (Figure 2A, middle panel,The lower panel of Figure 1B revealed that Myc-tagged
lanes 1–3). This interaction was also examined in B cells:forms of FKBP52 were expressed and immunoprecipi-
two murine cell lines, A20 and 70Z/3; and three humantated with anti-Myc antibody.
cell lines, KR-12, MC/CAR, and Namalwa cells. Variableamounts of IRF-4 and PU.1 were detected in all B cell
Mapping the IRF-4 Interaction Domainslines (Figure 2B, upper panel, lanes 1–5) but not in Jurkat
To localize the region of IRF-4 interacting with the im-T cells (Figure 2B, upper panel, lane 6). IRF-4-FKBP52
munophilin, deletion mutants of IRF4 and full-lengthinteraction was detected in all B cells examined but not
FKBP52 were cotransfected into COS-7 cells and ana-in Jurkat T cells (Figure 2B, middle panel). The amount
lyzed by coimmunoprecipitation. FKBP52 coprecipi-of IRF-4 interacting with FKBP52 did vary amongst the
tated with IRF-4 (aa 1–410), IRF-4 (aa 1–340), IRF-4 (aacell lines, possibly due to differences in IRF-4 expression
1–237), IRF-4 (aa 150–340), and IRF-4 (aa 150–410) (Fig-level in the different cell types.
ure 1C, lanes 2, 4, 6, 9, and 11), indicating that theinteraction domain of IRF-4 was located within theN-terminal 237 aa. FKBP52 did not coprecipitate with FKBP52 Inhibits IRF-4 Interaction with PU.1
and Subsequent Binding to the lB ElementIRF-4 (aa 1–150) (Figure 1D, lane 8) or with IRF-4 con-taining the internal deletion aa D150–340 (Figure 1D, IRF-4 was originally cloned as a murine transcription
factor recruited to composite elements within the immu-lane 6). The results of the domain interaction studiesare summarized schematically in Figure 1E. Based on noglobulin light chain gene enhancer El2–4 through spe-
cific interaction with PU.1, an Ets family member (Eisen-these results, the proline-rich region of IRF-4 (aa 150–237) appears to be the minimal region required for beis et al., 1993). We therefore investigated the effect
of FKBP52 on the formation of the IRF-4-PU.1 complex.FKBP52 interaction. However, despite repeated efforts,
immunoprecipitated (IP) with anti-Myc antibody 9E10 (amyc). Immunoprecipitated complexes were run on a 15% SDS-PAGE and immunoblotted(IB) with anti-FLAG antibody to detect FLAG-IRF-4 expression (upper panels, [B and C]), anti-HA antibody for IRF-4 (upper panel, [D]). Themembranes were reprobed with amyc to verify protein expression and successful IP (lower panels, [B–D]). WCE (30 mg) were used as input,and extract from untransfected COS-7 cells (300 mg) was immunoprecipitated with amyc as control IP.(E) Schematic summary of the IRF-4 interaction domains with full-length FKBP52. The 150–237 aa construct could not be stably expressedas a tagged peptide and therefore was not tested directly, hence the parentheses in Figure 1E. The different domains of IRF-4 are illustrated:DNA-binding domain with the tryptophan repeats seen in all IRF members (DBD; aa 1–150), the proline-rich region (Pro; aa 150–237), the IRFassociation domain (IAD; aa 237–410), and the C-terminal autoinhibition element (AIE; aa 410–450).

Immunity132
Figure 2. Coimmunoprecipitation Demonstrates Interaction between FKBP52 and Endogenous IRF-4 in HTLV-I-Transformed T Cells and BCells
(A) C8166, MT2, MT4, and MO-T WCE (500 mg) were immunoprecipitated with an a-FKBP52 antibody, and complexes were run on 10% SDS-PAGE and immunoblotted using a-IRF-4 antibody (middle panel) and a-FKBP52 antibody (lower panel). Input extracts consisted of 50 mg ofWCE (upper panel). The control IP was performed by incubating WCE (500 mg) with normal precleared mouse serum (data not shown).(B) A20, KR-12, MC/CAR, 70Z/3, Namalwa, and Jurkat WCE (500 mg) were immunoprecipitated with an a-FKBP52 antibody, and complexeswere run on 10% SDS-PAGE and immunoblotted using a-IRF-4 antibody (second panel) and a-FKBP52 antibody (fourth panel). Input extractsconsisted of 50 mg WCE (first and third panels). The control IP was performed by incubating WCE (500 mg) with normal precleared mouseserum (data not shown).
Nuclear extracts from COS-7 cells transfected with concentration of immunosuppressant was previouslyshown to abolish all PPIase activity (Pahl and Keller,IRF-4 and PU.1 were analyzed by EMSA using the lB
element, which includes partially overlapping binding 1992; Peattie et al., 1992; Leverson and Ness, 1998) buthad no effect on IRF-4 or PU.1 binding in the absencesites for IRF-4 and PU.1 (Eisenbeis et al., 1995; Brass
et al., 1996, 1999). As previously shown, IRF-4 alone did of FKBP52 (Y. M., unpublished data). This result indi-cated that formation of the ternary complex (IRF-4-PU.1-not bind to the lB element (Figure 3A, lane 1); FKBP52
likewise did not have intrinsic DNA binding activity (Fig- DNA) was inhibited due to the sequestration of IRF-4by FKBP52. Likewise, ascomycin had no effect on IRF-ure 3A, lane 2). In contrast, the PU.1-DNA complex was
detected in the absence of IRF-4 (Figure 3A, lane 4) and 4-FKBP52 immunoprecipitations when incubated withcell extracts from the cell lines described in Figure 2the complex was shifted with anti-PU.1 antibody (Figure
3A, lane 3). Addition of FKBP52 to the PU.1-containing (Y. M., unpublished data).extracts for different times did not affect the PU.1 DNAcomplex (Figure 3A, lanes 5–8). Nuclear extracts from FKBP52 Peptidyl-Prolyl Isomerase Activity Is Required
for Inhibition of IRF-4 DNA Bindingcells transfected with both IRF-4 and PU.1 expressionplasmids were also incubated with the lB element and To examine the effect of FKBP52 on IRF-4 DNA binding,
recombinant IRF-4 and extracts from C8166 cells weretwo complexes were identified—the PU.1 specific com-plex as well as a slower migrating complex containing analyzed by EMSA using the ISRE element of ISG-15.
Complexes were detected using the ISRE probe withboth PU.1 and IRF-4 (Figures 3A, lane 11, and 3B, lane3) as determined by supershift analysis with anti-PU.1 recombinant IRF-4 or C8166 nuclear extracts incubated
for different times (Figures 3C, lanes 1–4, and 3D, lanesand anti-IRF-4 antibody (Figure 3A, lanes 9 and 10, re-spectively). FKBP52 had no effect on PU.1-DNA binding 2–5, respectively). Although one complex was seen with
recombinant IRF-4 (Figure 3C, lanes 1–4), two com-(Figure 3A, lanes 5–8) but blocked IRF-4 binding to thelB element (Figures 3A, lanes 12–15, and 3B, lanes 4–7). plexes were detected with C8166 nuclear extracts (Fig-
ure 3D, lanes 2–5). The presence of IRF-4 in bothTo determine if functional PPIase activity was requiredfor FKBP52 inhibition of IRF-4-PU.1 DNA binding, asco- complexes was confirmed by supershift analysis with
anti-IRF-4 antibody (Figure 3D, lane 1). Recombinant IRF-4mycin (an ethyl analog of FK506, an immunosuppressantthat inhibits PPIase activity) was incubated with cell or C8166 extracts were also incubated with the ISRE
probe in the presence of GST-FKBP52 (Figures 3C, lanesextracts for different times. The presence of as little as10 mM ascomycin in the DNA binding assay restored 5–8, and 3D, lanes 6–9); although FKBP52 itself did not
form a complex (Figures 3C, lane 13, and 3D, lane 14),IRF-4-PU.1 binding to the lB probe (Figure 3B, lanes8–11), whereas DMSO (used to dilute the drug) had no it prevented IRF-4 from binding to DNA (Figures 3C,
lanes 5–8, and 3D, lanes 6–9, respectively). To determineeffect on IRF-4 binding (Y. M., unpublished data). This
FKBP52 Inhibits IRF-4 Transcriptional Activity133
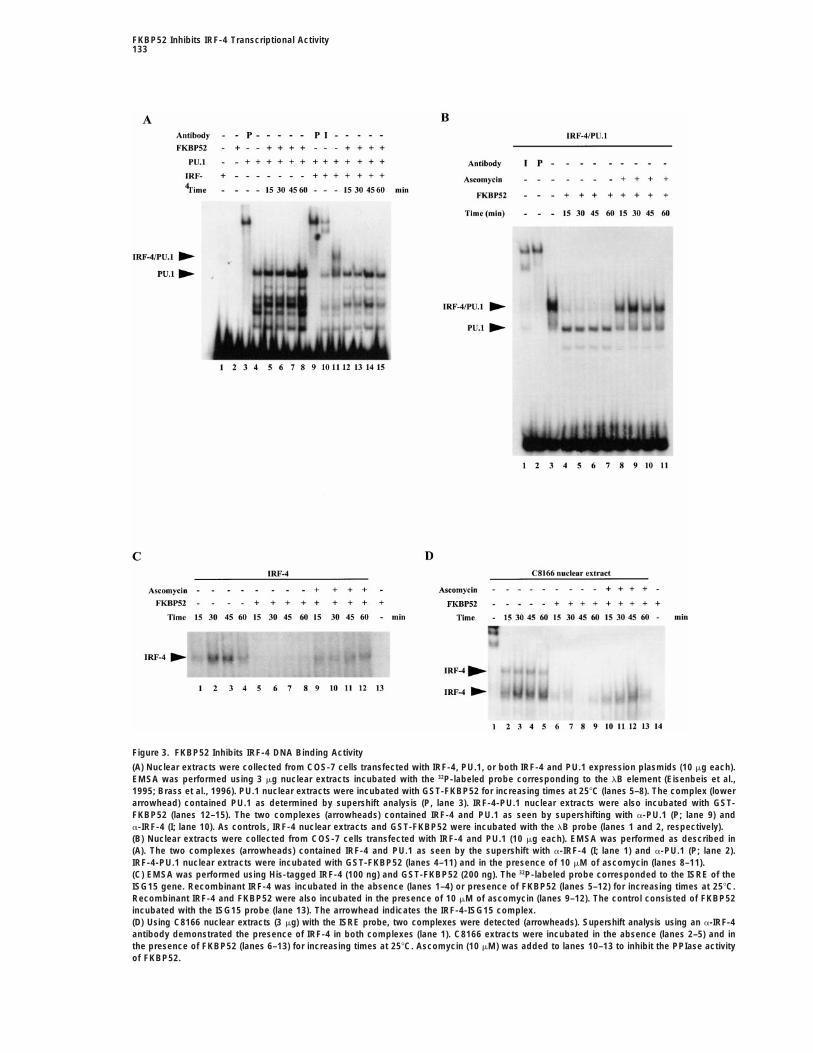
Figure 3. FKBP52 Inhibits IRF-4 DNA Binding Activity
(A) Nuclear extracts were collected from COS-7 cells transfected with IRF-4, PU.1, or both IRF-4 and PU.1 expression plasmids (10 mg each).EMSA was performed using 3 mg nuclear extracts incubated with the 32P-labeled probe corresponding to the lB element (Eisenbeis et al.,1995; Brass et al., 1996). PU.1 nuclear extracts were incubated with GST-FKBP52 for increasing times at 258C (lanes 5–8). The complex (lowerarrowhead) contained PU.1 as determined by supershift analysis (P, lane 3). IRF-4-PU.1 nuclear extracts were also incubated with GST-FKBP52 (lanes 12–15). The two complexes (arrowheads) contained IRF-4 and PU.1 as seen by supershifting with a-PU.1 (P; lane 9) anda-IRF-4 (I; lane 10). As controls, IRF-4 nuclear extracts and GST-FKBP52 were incubated with the lB probe (lanes 1 and 2, respectively).(B) Nuclear extracts were collected from COS-7 cells transfected with IRF-4 and PU.1 (10 mg each). EMSA was performed as described in(A). The two complexes (arrowheads) contained IRF-4 and PU.1 as seen by the supershift with a-IRF-4 (I; lane 1) and a-PU.1 (P; lane 2).IRF-4-PU.1 nuclear extracts were incubated with GST-FKBP52 (lanes 4–11) and in the presence of 10 mM of ascomycin (lanes 8–11).(C) EMSA was performed using His-tagged IRF-4 (100 ng) and GST-FKBP52 (200 ng). The 32P-labeled probe corresponded to the ISRE of theISG15 gene. Recombinant IRF-4 was incubated in the absence (lanes 1–4) or presence of FKBP52 (lanes 5–12) for increasing times at 258C.Recombinant IRF-4 and FKBP52 were also incubated in the presence of 10 mM of ascomycin (lanes 9–12). The control consisted of FKBP52incubated with the ISG15 probe (lane 13). The arrowhead indicates the IRF-4-ISG15 complex.(D) Using C8166 nuclear extracts (3 mg) with the ISRE probe, two complexes were detected (arrowheads). Supershift analysis using an a-IRF-4antibody demonstrated the presence of IRF-4 in both complexes (lane 1). C8166 extracts were incubated in the absence (lanes 2–5) and inthe presence of FKBP52 (lanes 6–13) for increasing times at 258C. Ascomycin (10 mM) was added to lanes 10–13 to inhibit the PPIase activityof FKBP52.

Immunity134
Figure 4. Analysis of IRF-4 Transactivation Potential in the Presence of FKBP52
(A) COS-7 cells were transfected with 2 mg of each of the B4TK-CAT reporter (Eisenbeis et al., 1995; Brass et al., 1996) and the IRF-4 and/or PU.1 expressing plasmids, as well as increasing amounts of FKBP52 expressing plasmid (2, 5, 10, 15, 20, and 25 mg).(B) COS-7 cells were transfected as described in (A) and were treated with 2 mM of ascomycin for 16 hr.(C) COS-7 cells were transfected with 2 mg of each of the B4TK-CAT reporter and IRF-4 or PU.1 expressing plasmids, as well as increasingamounts of DN-FKBP52 (aa 233–459) expressing plasmid (2, 5, 10, 15, 20, and 25 mg).(D) COS-7 cells were transfected with 2 mg of each of the PRDII-CAT reporter and the NF-kB p65 expressing plasmid, as well as increasingamounts of FKBP52 expressing plasmid (2, 5, 10, 15, 20, and 25 mg). CAT activity was analyzed 48 hr posttransfection using 10 mg WCE for4 hr at 378C. CAT activity values correspond to an average of three to six independent experiments and were normalized to bGal expression.
if functional PPIase activity was required for FKBP52 FKBP52 Inhibits IRF-4 TransactivationPreviously, the IRF-4-PU.1 dimer was shown to stimu-inhibition of IRF-4 binding, recombinant IRF-4 or C8166
nuclear extracts were incubated with recombinant late the transcriptional activity of the El2–4 lB elementin a synergistic manner (Eisenbeis et al., 1995; Brass etFKBP52 and ascomycin (10 mM) for different times (Fig-
ures 3C, lanes 9–12, and 3D, lanes 10–13); again, the al., 1996). The ability of FKBP52 to interfere with IRF-4and IRF-4-PU.1 transactivation was next investigatedpresence of ascomycin in the DNA binding assay mix
restored IRF-4 binding to the ISRE probe. The nature using COS-7 cells transfected with IRF-4, PU.1, andincreasing amounts of FKBP52 expression plasmid. Us-of the upper IRF-4 complex, which is not restored by
ascomycin, is not clear (Figure 3D, lanes 2–5). This com- ing the B4TK-CAT construct with a tetramer of the El2–4
lB element driving the CAT reporter, a greater than 7- toplex may represent an IRF-4 dimer, IRF-4 and an uniden-tified host cellular protein, or a complex that is irrevers- 8-fold transactivation was produced by the synergistic
combination of IRF-4 and PU.1 (Figure 4A, lane 5), whileibly inhibited by FKBP52.

FKBP52 Inhibits IRF-4 Transcriptional Activity135
Figure 5. FKBP52 Induces a Conformational Change in IRF-4 and Protects from Proteolytic Degradation
(A) Recombinant His-tagged IRF-4 (100 ng) was incubated with GST-FKBP52 (200 ng) at 258C and 378C for different times. IRF-4 alone (lane1, upper panel) was incubated at 378C for 1 hr as control. The same experiment was repeated in the presence of 10 mM of ascomycin (lowerpanel). The control consisted of IRF-4 and 10 mM of ascomycin incubated at 378C for 1 hr (lane 1, lower panel). Samples were run on 8%SDS-PAGE, transferred to a membrane, and immunoblotted with anti-IRF-4 antibody.(B) In vitro translated 35S-labeled His-IRF-4 (1 ml) was preincubated with BSA (200 ng, lanes 1–8), GST-FKBP52 (200 ng, lanes 9–24), or GST-FKBP52 and ascomycin (1 mM, lanes 17–24) for 1 hr at 258C; IRF-4, IRF-4/FKBP52, and IRF-4/FKBP52/ascomycin reactions were subjectedto chymotrypsin digestion (2 ng per reaction) for different times at 258C. Digestion reactions were terminated by the addition of 1X SDS sampleloading buffer, loaded on 15% SDS-PAGE, and visualized by autoradiograghy. Controls (lanes 1 and 9) were incubated for 30 min at 258C inthe absence of chymotrypsin.
IRF-4 or PU.1 alone did not possess a strong transacti- unpublished data). Inhibition of IRF-4 was also IRF-4specific, based on the observation that increasingvation potential (Figure 4A, lanes 2 and 3, respectively).
FKBP52 itself had no effect on the B4TK-CAT (Figure FKBP52 did not affect NF-kB p65-mediated transactiva-tion of an NF-kB-containing CAT reporter plasmid (Fig-4A, lane 4), whereas increasing amounts of FKBP52 in
the presence of IRF-4 and PU.1 resulted in a dramatic ure 4D, lanes 4–9).decrease in IRF-4-PU.1 transactivation capacity (Figure4A, lanes 6–11). Next, transfected cells were incubated FKBP52 Induces a Conformational Change in IRF-4
Analysis of the expression pattern of IRF-4 in extractswith ascomycin (2 mM) for 16 hr; ascomycin completelyblocked the inhibition of IRF-4-PU.1 transactivation by from HTLV-1-infected cells revealed two closely migrat-
ing bands corresponding to IRF-4. The slower migratingFKBP52 (Figure 4B, lanes 6–11), whereas the drug hadno effect on IRF-4-PU.1 transactivation in the absence of form of IRF-4 did not appear to correspond to phosphor-
ylated IRF-4, since calf intestinal phosphatase treatmentFKBP52 (Figure 4B, lane 5). Similar results were obtainedwhen a truncated FKBP52 lacking the PPIase domain did not affect IRF-4 migration (Y. M., unpublished data).
To examine the relationship between the PPIase activity(DN-FKBP52) was cotransfected in increasing amounts;no decrease in transactivation potential was observed of FKBP52 and the distinct IRF-4 forms observed on
SDS-PAGE, recombinant IRF-4 and FKBP52 were incu-(Figure 4C, lanes 6–11). Using a different reporter con-struct, IRF-4 produced a moderate increase in ISG-15- bated at 258C and 378C for different times; recombinant
IRF-4 alone was detected as a unique form by immu-CAT activity (2- to 3-fold) (Y. M., unpublished data).Although FKBP52 itself had no effect on reporter gene noblot (I) (Figure 5A, lane 1). When incubated with
FKBP52, a second band (II) with a slightly faster migra-activity, increasing amounts of FKBP52 also produceda concentration-dependent inhibition of IRF-4 transacti- tion accumulated with time (Figure 5A, lanes 2–9). The
appearance of the faster migrating form was observedvation. FKBP52-mediated inhibition also required func-tional PPIase activity, since the use of ascomycin and as early as 5 min (Figure 5A, lanes 2 and 3), and by
45 min IRF-4 was completely converted into the fasterDN-FKBP52 cotransfection restored IRF-4 activity (Y. M.,

Immunity136
migrating form (Figure 5A, lanes 10 and 11). This effectwas specific to FKBP52, since IRF-4 alone remainedas a single form (Figure 5A, lane 1; data not shown).Furthermore, the appearance of the faster migratingform was completely inhibited in the presence of thePPIase inhibitor ascomycin (Figure 5A, lower panel,lanes 2–11). Thus, the appearance of the faster migratingform is dependent on functional PPIase activity ofFKBP52.
To further confirm the conformational alteration inIRF-4, partial proteolytic digestion of recombinant IRF-4was performed in the presence or absence of recombi-nant FKBP52 and ascomycin (Figure 5B, lanes 2–8, lanes10–16, and lanes 18–24). Partial proteolytic cleavage Figure 6. Ascomycin Induces IgLl mRNA Expression in the Pre-Bof 35S-labeled IRF-4 by chymotrypsin was blocked by Cell Line 70Z/3preincubation of IRF-4 with FKBP52 for 1 hr (Figure 5B, Three cell lines, A20, 70Z/3, and Jurkat T cells (1 3 107 cells each),lanes 10–16). Strikingly, the presence of 10 mM asco- were treated with ascomycin (2 mM) for different times. Total RNA
was extracted from these cells and RT–PCR was performed on 2mycin restored the partial proteolytic digest pattern andmg RNA using labeled IgLl (top panel), GAPDH (middle panel), andconfirmed the importance of the PPIase activity ofIRF-4 (bottom panel) specific primers. Due to high IgLl expressionFKBP52 in mediating proteolysis-resistant conformationin A20, the room temperature reaction was diluted 100-fold for theof IRF-4 (Figure 5B, lanes 18–24). Ascomycin had no PCR reaction. The room temperature reaction was also diluted 100-
effect on the proteolytic digest of IRF-4 alone, and simi- fold and 10-fold for GAPDH and IRF-4 amplification, respectively.lar results were obtained with trypsin proteolysis (Y. M., Cells not treated with ascomycin were incubated with DMSO (used
to dilute the drug) for 72 hr.unpublished data).
Ascomycin Induces the Expression of IgLl DNA binding required functional peptidyl-prolyl isom-in the Murine Pre-B Cell Line 70Z/3 erase (PPIase) activity of FKBP52, since the PPIase in-70Z/3, a mouse pre-B lymphoma cell line, has been hibitor ascomycin restored IRF-4 binding. Transactiva-used extensively as a model to study the transcriptional tion by IRF-4-PU.1 and IRF-4 was also abrogated byregulation of immunoglobulin genes (Paige et al., 1981; FKBP52 in a concentration-dependent manner, an effectKoromilas et al., 1995). 70Z/3 expresses both IRF-4 and that also required functional PPIase activity, since bothPU.1 at levels similar to those in differentiated B cells ascomycin and a mutant of FKBP52 (DN-FKBP52) lack-(Figure 2B, lanes 1–5; data not shown) but does not ing PPIase activity blocked the inhibitory effect. Incuba-express surface immunoglobulins. Furthermore, the tion with FKBP52 led to the appearance of a faster mi-PU.1/IRF-4 complex does not stimulate IgLl mRNA pro- grating IRF-4 band as visualized by immunoblot analysisduction in these cells. To determine if the PPIase activity and by partial proteolysis of IRF-4. Finally, incubationof FKBP52 was responsible for the absence of IRF-4/ of the pre-B cell line 70Z/3 with ascomycin resulted inPU.1 activity, the pre-B cell line 70Z/3 was treated with the expression of IgLl, an IRF-4-PU.1-regulated geneascomycin (2 mM) for 0–72 hr. Using an RT–PCR-based in B cells. Based on these observations, we concludeassay, IgLl expression was detected at 48 and 72 hr that FKBP52 alters the conformation of IRF-4 by cis-after ascomycin treatment (Figure 6, upper panel, lanes trans isomerization of proline residues; such a modifica-7 and 8); IgLl mRNA expression was also detected at tion appears to convert IRF-4 from a configuration capa-36 hr (Y. M., data not shown). The differentiated B cell ble of interaction with PU.1, DNA binding, and activationline A20 was used as a positive control that already of gene expression (IgLl) to a conformation that noexpressed high amounts of IgLl (Figure 6, upper panel, longer interacts with PU.1 or binds DNA. Ascomycinlane 1). Inhibition of PPIase activity also increased IgLl relieves the inhibition catalyzed by FKBP52 by abrogat-expression in these cells by 4-fold (Figure 6, upper panel, ing the PPIase activity of the immunophilin.lane 2), whereas no such increase was detected in Jurkat The inhibition of IRF-4-PU.1 DNA binding and trans-T cells treated with the drug (Figure 6, upper panel, lane activation potential by FKBP52 has important implica-9). Finally, ascomycin had no effect on GAPDH or IRF-4 tions for B cell development. The IRF-4-PU.1 complexmRNA expression levels (Figure 6, middle and bottom is essential for the expression of the IgLl and the Jpanels). chain genes. Both genes are required for the expression
of a productively rearranged immunoglobulin l gene inplasma cells (Brass et al., 1999). FKBP52 may thereforeDiscussionfunction as a negative regulator of immunoglobulin geneexpression during B cell differentiation, or alternativelyIn the present study, a novel interaction between IRF-4
and the immunophilin FKBP52 has been characterized. it may act to prevent expression of certain genes priorto the plasma cell stage. Inhibition of the transactivatingThe proline-rich region of IRF-4 (aa 150–237) and the
C-terminal tetratricopeptide repeats of FKBP52 were the complex IRF-4-PU.1 by FKBP52 suggests a new rolefor immunophilins in the regulation of B cell developmentminimal domains required for interaction. Association of
FKBP52 with IRF-4 inhibited IRF-4-PU.1 binding to the and maturation.Previously, the IRF-4-PU.1 dimer was shown to stimu-IgLl enhancer El2–4, as well as IRF-4 interaction with the
ISG-15 ISRE element. Furthermore, inhibition of IRF-4 late the transcriptional activity of the El2–4 lB element

FKBP52 Inhibits IRF-4 Transcriptional Activity137
Figure 7. Schematic Model of the Role ofFKBP52 in Regulating IRF-4 Function
(A) PPIase catalyzes cis-trans isomerizationat specific proline residues. Peptide bonds onthe amino side of proline residues are foundmainly in the trans-conformation. The ap-pearance of the cis-conformation is catalyzedby PPIase (upper panel). Amino acid preced-ing (A21) and following the proline residue (A2)are linked to polypeptide chains (adaptedfrom Kay, 1996). Residues within the proline-rich domain of IRF-4 (aa 150–237) that maybe subject to isomerization by FKBP52 areunderlined (lower panel).(B) Schematic model describing the potentialfunctions of FKBP52 in mediating nucleartransport/chaperone function, protein foldingby cis-trans isomerization, and regulation ofIRF-4 functions such as DNA binding andtransactivation. DBD, DNA-binding domain;IAD, IRF-association Domain; Pro, proline-rich region; closed diamond, tetratricopep-tide repeats.
in a synergistic manner (Eisenbeis et al., 1995; Brass et autoinhibition of DNA binding and ternary complex for-mation (Brass et al., 1996). Consistent with these ob-al., 1996). A model of IRF-4-PU.1 complex activation
proposed that in the absence of PU.1, IRF-4 was in a servations, FKBP52 did not immunoprecipitate withfull-length IRF-4; only after removal of the C-terminalclosed conformation unable to bind to the El2–4 lB ele-
ment. Following interaction of IRF-4 with PU.1 and DNA, autoinhibition domain (aa 411–450) did IRF-4 interactwith FKBP52. IRF-4 appears to be modified by FKBP52IRF-4 underwent a conformational change that swiveled
the regulatory domain (aa 170–450) away from its own through proline isomerization to maintain a closed non–DNA binding conformation, which is relieved throughDBD (aa 1–150) and into direct contact with the PEST
region of PU.1. Furthermore, deletion of the C-terminal the intermolecular interaction with PU.1. As describedpreviously (Eisenbeis et al., 1995; Brass et al., 1996,segment generated a form of IRF-4 with PU.1-indepen-
dent binding activity (Brass et al., 1996, 1999). Ortiz 1999), IRF-4 may be inaccessible to FKBP52 becauseof structural constraints imposed by the C-terminal au-et al. further delimited the IRF-4 interaction domain to
residues 245–422 (Ortiz et al., 1999). Site-directed muta- toinhibition element.Several physiological roles have been ascribed to im-genesis of conserved amino acids within two predicted
a helices confirmed the importance of these residues munophilins: (1) binding and sequestration of cal-cineurin, (2) protein folding and assembly, (3) proteinfor IRF-4-PU.1 DNA binding and transactivation. These
two a helices are also highly conserved amongst the trafficking, (4) direct regulation of protein activity, and5) chaperone-like activity (Kay, 1996). Immunophilins areIRF family members and could therefore be involved in
heterodimerization with other transcription factors (Ortiz peptidyl-prolyl isomerases (PPIases) involved in catalyz-ing cis-trans isomerization of proline residues withinet al., 1999). To analyze the function of the IRF-4-PU.1
dimer in vivo, a chimeric repressor was engineered by proteins. cis prolines are important to protein structuralintegrity because they introduce bends within proteins.fusing PU.1 and IRF-4 DNA-binding domains through a
flexible POU domain. This fused dimer strongly re- During protein synthesis, the peptide bonds on theamino side of proline residues are found in the openpressed expression of a rearranged immunoglobulin
gene (IgLl) (Brass et al., 1999). trans-conformation (Figure 7A). When the three-dimen-sional structure was determined, 15% of these bondsDeletion analysis of IRF-4 also revealed the presence
of a C-terminal segment (aa 410–450) important for were in the alternate cis-conformation (Kay, 1996;

Immunity138
Marks, 1996). Enzymes with peptidyl-prolyl isomerase observed at 4–5 weeks of age, but with time IRF-42/2
activity such as FKBP52 catalyze the cis-trans intercon- mice gradually exhibited severe lymphadenopathy andversion, a modification that would otherwise occur defects in both B and T cell activation (Mittrucker et al.,slowly. Immunophilins target specific proline residues 1997). Early T cell events such as calcium influx andpreceded by a bulky hydrophobic residue for cis-trans expression of the T cell activation markers CD25 andisomerization (Kay, 1996; Marks, 1996; Hunter, 1998). CD69 remained normal in IRF-42/2 T cells, indicating thatWithin IRF-4, several clustered prolines preceded by IRF-4 may function at later stages of T cell activation,hydrophobic residues are available for peptidyl-prolyl possibly at the level of IL-2 production and/or IL-2 re-isomerization by FKBP52 (Figure 7A). In the model illus- sponse. The unexpected convergence of transcriptiontrated in Figure 7B, we propose that FKBP52 can act factors and immunophilins suggests a novel posttrans-as a chaperone that escorts IRF-4 into the nucleus. lational role for immunophilins in the regulation of geneDuring transport or once in the nucleus, FKBP52 would transcription and cell growth in lymphocytes.alter IRF-4 conformation through proline isomerization,thereby sequestering IRF-4. The interaction with the Experimental Proceduresproper transcriptional partner such as PU.1 would allevi-
Yeast Two-Hybrid Screenate this inhibitory effect and permit DNA binding andThe fragment containing the proline-rich and IRF association do-transactivation of target genes such as IgLl (Figure 7B).mains (aa 150–410) of IRF-4 was PCR amplified with primer AOne of the first members of the immunophilin family,(59-ATCAGAATTCCCGGGAGTGTACAGGATTGTTCC-39) and primer B
cyclophilin A, was shown to be the primary target of (59-ATCAGTCGACGGATCCTGAGGGTCTGGAAACTCC-39) and sub-cyclosporin A, a clinically used immunosuppressive cloned into the pAS2-1 vector (GAL4 1–147 DNA-binding domain,drug that blocks the calcineurin-dependent activation TRP1; Clontech) in frame with the DNA-binding domain of GAL4
and sequenced. The yeast strain was cotransformed by the lithiumof NF-AT in T cells (Emmel et al., 1989; Mattila et al.,acetate permeabilization method with the pAS2-1-IRF-4 (150–410)1990; Morris, 1994; Mattila, 1996). FKBP52 (HBI, p59,vector and with an EBV-transformed B cell cDNA library cloned intoHSP56) was originally discovered in association withthe pACT vector (Clontech). Positive yeast clones were selected for
unliganded steroid receptor complexed with heat shock prototrophy for leucine, tryptophan, and histidine and screened forproteins HSP70 and HSP90 and involved in cytoplasmic- b-galactosidase expression. Subsequent two-hybrid assays wereto-nuclear shuttling (Peattie et al., 1992; Perrot-Applanat carried out by a mating assay between the Y187 and Y190. Diploid
cells from the mating assay were selected for Trp-, Leu-, and His-et al., 1995). Recently, another member of the immu-selective medium and screened by replica plating for their ability tonophilin family, the human cyclophilin Cyp-40, wasproduce for b-galactosidase. Plasmid DNA was isolated from yeastshown to regulate the transcriptional activity of c-Myband transformed in E. coli DH5a by electroporation. The constructstranscription factor (Gonda, 1998; Gonda et al., 1989)were sequenced using 59PACT primer (59-TACCACTACAATGGATG-39).
by interfering with the capacity of c-Myb to bind DNA;interestingly, the oncogenic form of v-Myb evaded this Protein Expression and Purificationinhibition of DNA binding (Leverson and Ness, 1998). The GST-FKBP52 fusion protein was expressed and isolated fromPoint mutations and deletions in v-Myb that contribute E. coli BL21 following a 3 hr induction with 1 mM IPTG (Pharmacia)
at 308C. Bacterial extracts in PBS containing 1% Triton X-100 wereto its oncogenic potential also abrogate the cyclophilin-incubated with glutathione Sepharose beads (Pharmacia) for 20 minmediated inhibition of DNA binding of v-Myb (Lipsick,at room temperature and the fusion protein was eluted as previously1996; Gonda, 1998). By analogy with c-Myb and Cyp-described (Lin et al., 1999) The batches were 80%–90% pure with
40, IRF-4 interacts with an immunophilin that inhibits its a concentration of 33–100 ng/ml. The IRF-4 fusion protein was ex-DNA binding activity and transactivation potential in a pressed as a poly-histidine tagged protein, isolated from E. coliPPIase-dependent fashion. BL21 following a 3 hr induction with 1 mM IPTG (Pharmacia) at 308C.
One of the original sources of IRF-4 was adult T cell Bacterial extracts in PBS containing 1% Triton X-100 were incubatedwith His-Bind resin (Novagen) for 20 min at room temperature. Highleukemia cell lines (Yamagata et al., 1996). The specificpurity (95%) and concentration (100 ng/ml) were obtained.detection of high levels of IRF-4 expression in HTLV-1-
infected cells was unexpected, since IRF-4 expressionCAT Assay, Western Blot Analysis, and Coimmunoprecipitationin activated T cells was often difficult to detect (Yama-The HTLV-1-infected cell lines C8166, MT-2, MT-4, and Jurkat Tgata et al., 1996). Interestingly, HTLV-1 Tax expression cells were cultured in RPMI 1640 media (CLT), the MO-T cell line in
appears to correlate with IRF-4 expression and may be Iscove’s media (CLT) supplemented with 10% fetal bovine serum,acting in an indirect manner to stimulate gene expres- and antibiotics. The B cell lines were purchased from ATCC andsion through the interaction with host transcription cultured according to their directives. The A20 cell line was cultured
in RPMI 1640/10% fetal bovine serum to express high amounts offactors (Hiscott et al., 1995). The role of IRF-4 per sesurface immunoglobulins (ATCC). All CAT assays and coimmuno-in HTLV-1-induced leukemogenesis remains unclear.precipitations were carried in monkey embryonic kidney COS-7 cellsHowever, the relationship between IRF-4 expressiongrown in Dulbecco’s MEM media (GIBCO–BRL) supplemented with
and oncogenicity is highlighted by the observation that 10% fetal bovine serum and antibiotics. Subconfluent cells werein some patients with multiple myeloma, a chromosomal transfected with 2–15 mg of CsCl purified plasmid by lipofectaminetranslocation—t(6;14)(p25;q32)—juxtaposes the immu- (CLT) method. The reporter plasmids contained the IRF-4-PU.1-noglobulin heavy chain (IgH) locus to MUM1 (multiple responsive elements (B4TK-CAT) (Eisenbeis et al., 1995) or the inter-
feron-stimulated gene 15 (ISG15) promoter (Lin et al., 1998) linkedmyeloma 1); the MUM1 locus at 6p25 is virtually identicalto the CAT reporter gene as well as the IRF-4-, PU.1-, DN-FKBP52-to IRF-4. This chromosomal translocation involving IRF-4(aa 233–459), and FKBP52-expressing plasmids. Sixteen hoursmay thus contribute to leukemogenesis, since MUM1/post-transfection, transfected COS-7 cells were treated with asco-
IRF-4 has oncogenic activity in vitro (Iida et al., 1997). mycin (2 mM) or DMSO. At 48 hr after the transfection, total proteinA potential candidate gene that may be modulated in extracts were prepared and 10 mg were used for CAT assay. CAT
HTLV-1-infected cells by IRF-4 is the IL-2 gene. In IRF- activities represent an average of CAT values obtained from three tosix independent experiments. For Western blot analysis, equivalent4-deficient mice, a normal T and B cell distribution was

FKBP52 Inhibits IRF-4 Transcriptional Activity139
amounts of whole cell extracts (20–50 mg) or of recombinant proteins Acknowledgments(50–100 ng) were subjected to 15% SDS-PAGE and then transferredto Hybond transfer membrane (Pharmacia). The membrane was The authors wish to thank Dr. Harinder Singh at the Howard Hughes
Institute, University of Chicago, for IRF-4, the deletions constructsprobed overnight at 48C with different antibodies: anti-ICSAT/IRF-4antibody (Santa Cruz), anti-HA antibody (Boehringer Manheim), anti- of IRF-4, and the B4TK-CAT reporter; Dr. Antonis Koromilas, Lady
Davis Institute, McGill University, for the 70Z/3 cell line; Dr. MatthewMyc antibody 9E10, anti-Tax antibody (NIH), anti-FKBP52 antibody(Biomol), anti-PU.1 antibody (Santa Cruz), or anti-FLAG (Sigma) anti- Fenton, Boston University, for the PU.1 construct; and Dr. Steve
Michnick at Universite de Montreal for many helpful discussions andbody at a dilution of 1:1000.For coimmunoprecipitations, COS-7 cells (15 cm plate at 70%– suggestions. We also thank the members of the Molecular Oncology
Group, Lady Davis Institute, McGill University, for helpful discus-80% confluency) were transfected by the lipofectamine method(CLT) with 15 mg expression plasmids. Whole-cell extracts from sions. This research was supported by grants from the National
Cancer Institute with funds from the Canadian Cancer Society andCOS-7, HTLV-transformed cells, and B cell lines (500 mg) were lysedwith TNN (50 mM Tris–HCl, 200 mM NaCl, 0.5% NP-40). Immuno- the Medical Research Council of Canada. Y. M. was supported by
a MRC studentship, R. L. was supported in part by a Fraser Monatprecipitating antibodies such as a-MYC 9E10 or a-FKBP52 werecross-linked to 300 ml Sepharose G or Sepharose A beads with McPherson Fellowship from McGill University, and J. H. was sup-
ported by an MRC Senior Scientist award.dymethyl pimelimidate (Sigma). The extracts were incubated with30 ml protein Sepharose beads cross-linked with antibody for 2–3hr at 48C. Immunocomplexes were eluted by boiling the beads andseparated by SDS-PAGE. Received May 27, 1999; revised December 29, 1999.
Electrophoretic Mobility Shift Assay ReferencesNuclear extracts from C8166 or COS-7 transfected cells (3 mg) wereprepared as previously described (Dignam et al., 1983), or 200 ng Brass, A., Kehrli, E., Eisenbeis, C., Storb, U., and Singh, H. (1996).recombinant protein were assayed for IRF-4 binding in gel shift Pip, a lymphoid restricted IRF, contains a regulatory domain that isanalysis using 32P-labeled oligonucleotide corresponding to the important for autoinhibition and ternary complex formation with theISRE of the ISG-15 gene (59-GATCGGGAAAGGGAAACCGAAACT Ets factor PU.1. Genes Dev. 10, 2335–2347.GAAGCC-39) and the lB element (Eisenbeis et al., 1995). Binding Brass, A.L, Zhu, A.Q., and Singh, H. (1999). Assembly requirementsreactions and electrophoresis were performed as previously de- of PU.1/Pip (IRF-4) activator complexes—inhibiting function in vivoscribed (Lin et al., 1999). In some experiments, whole-cell extracts using fused dimers. EMBO J. 18, 977–991.and recombinant protein were incubated with 10 mM of ascomycin
Burysek, L., Yeow, W.S., Luyova, B., Schafer, S.L., Huang, Y.Q.,(Calbiochem) diluted in DMSO, an ethyl analog of FK506. DMSO orand Pitha, P.M. (1999). Functional analysis of human herpesvirusascomycin had no effect on IRF-4 DNA binding (data not shown).8-encoded viral interferon regulatory factor 1 and its associationwith cellular interferon regulatory factors and p300. J. Virol. 73,7334–7342.Partial Proteolysis of IRF-4Czar, M.J., Owens-Grillo, J.K., Yem, A.W., Leach, K.L., Deibel, M.R.,In vitro translated IRF-4 was produced using the T7 polymeraseWeish, M.J., and Pratt, W.B. (1994). The hsp56 immunophilin compo-TnT system (Promega) with 35S-labeled mix containing cysteine andnent of the untransformed steroid receptor complex is localizedmethionine (ProRedivue, Amersham). Purity of labeled protein wasboth to the microtubules in the cytoplasm and in the same nonran-verified by SDS-PAGE and autoradiography. Labeled IRF-4 (1 ml perdom regions within the nucleus as the steroids receptors. Mol. Endo-reaction) was preincubated with GST-FKBP52, ascomycin (1 mM),crinol. 8, 1731–1741.or BSA (200 ng) for 1 hr at room temperature. Chymotrypsin was
added at different time points and the proteolytic reactions were D’Addario, M., Wainberg, M.A., and Hiscott, J. (1992). Activation ofstopped by the addition of 1X SDS-loading buffer. The control with cytokine genes in HIV-1 infected myelomonoblastic cells by phorbolno chymotrypsin was incubated for 30 min at room temperature. ester and tumor necrosis factor. J. Immunol. 148, 1222–1229.Ascomycin was also preincubated with IRF-4 alone and had no Daenkes, S., Nightingale, S., Cruickshank, J.K., and Bangham, C.R.effect on proteolysis (data not shown). Samples were loaded on (1990). Sequence variants of HTLV-I from patients with tropical spas-15% SDS gels, enhanced with Amplify solution (Dupont), dried, and tic paraparesis and adult T cell leukemia do not distinguish neurolog-exposed to film. ical from leukemic isolates. J. Virol. 64, 1278–1282.
Dignam, J.D., Lebovitz, R.M., and Roeder, R.G. (1983). Accuratetranscription initiation by RNA polymerase II in a soluble extractRT–PCR Analysis and IgLl mRNA Expressionfrom isolated mammalian nuclei. Nucleic Acids Res. 11, 1475–1489.Total RNA isolated from 70Z/3 and A20 cells was treated with 1 UEisenbeis, C., Singh, H., and Storb, U. (1993). PU.1 is a componentRNase-free DNase (RQ1 DNase, Promega Biotech) for 30 min atof a multiprotein complex which binds an essential site in the murine378C and ethanol-precipitated. Reverse transcription was performedimmunoglobulin l2–4 enhancer. Mol. Cell. Biol. 13, 6452–6461.with 2 mg of total RNA and 0.2 pmol of random hexamers using
200 U of Moloney murine leukemia virus reverse transcriptase (Life Eisenbeis, C.F., Singh, H., and Storb, U. (1995). Pip, a novel IRFTechnologies) in buffer containing 50 mM Tris–HCl (pH 8.3), 75 mM family member, is a lymphoid-specific, PU.1-dependent transcrip-KCl, 10 mM dithiothreitol, 3 mM MgCl2, 500 nM dNTP, 0.1 mg/ml tional activator. Genes Dev. 9, 1377–1387.bovine serum albumin, and 272.5 U/ml RNase inhibitors (Amersham Emmel, E.A., Verweij, C.L., Durand, D.B., Higgins, K.M., Lacy, E., andPharmacia Biotech). PCR assays were performed using 5 ml and 1 Crabtree, G.R. (1989). Cyclosporin A specifically inhibits function ofml of room temperature product for IgLl (undiluted) and GAPDH nuclear proteins involved in T cell activation. Science 246, 1617–(100-fold diluted) as well as IRF-4 (10-fold diluted) mRNA, in 100 1620.mM Tris–HCl (pH 8.4), 500 mM KCl, 15 mM MgCl2, 200 mM dNTP,
Fujita, T., Sakakibara, J., Sudo, Y., Miyamoto, M., Kimura, Y., and15 pmol of g32P-labeled primers, and 1.25 U of Taq DNA polymeraseTaniguchi, T. (1988). Evidence for a nuclear factor(s), IRF-1, mediat-(Amersham Pharmacia Biotech). Nucleotide sequence of primersing induction and silencing properties to human IFN-b gene regula-used for IgLl were 59-ATGGCCTGGACTTCACTTA-39 and 59-TCTtory elements. EMBO J. 7, 3397–3405.TGGGCTGGCTTAGGA-39, and for IRF-4, 59-CGCTCTTCAAGGCTTGessain, A., Barin, F., Vernant, J.C., Gout, O., Maurs, L., Calender,GGG-39 and 59-GGTCGAGGGGTGGCATCA-39. The PCR reactionA., and de The, G. (1985). Antibodies to human T-lymphotropic virusmixture was subjected to 25 cycles of denaturation for 30 s at 958C,type-I in patients with tropical spastic paraparesis. Lancet 2,annealing for 30 s at 628C, and polymerization for 1 min at 728C.407–410.PCR products were analyzed on a 5% denaturing polyacrylamide gel.
Primers for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) Gonda, T. (1998). The c-Myb oncoprotein. Int. J. Biochem. Cell. Biol.30, 517–551.were used as described previously (D’Addario et al., 1992).

Immunity140
Gonda, T.J., Ramsay, R.G., and Johnson, G.R. (1989). Murine my- protein-protein interactions. Proc. Natl. Acad. Sci. USA 96, 2740–2745.eloid cell lines derived by in vitro infection with recombinant c-myb
retroviruses express myb rearranged vector proviruses. EMBO J. 8, Pahl, A., and Keller, U. (1992). FK-506-binding proteins from Strepto-1767–1775. myces producing immunosuppressive macrolactones of the FK-506
type. J. Bacteriol. 174, 5888–5894.Hiscott, J., Petropoulos, L., and Lacoste, J. (1995). Molecular inter-actions between HTLV-1 Tax protein and the NF-kB/IkB transcrip- Paige, C.H., Kincade, P.W., and Ralph, P. (1981). Independent con-tion complex. Virology 214, 3–11. trol of the immunoglobulin heavy and light chain expression in a
murine pre-B-cell line. Science 292, 631–633.Hunter, T. (1998). Prolyl isomerases and nuclear function. Cell 92,141–143. Peattie, D.A., Harding, M.W., Fleming, M.A., DeCenzo, M.T., Lippke,
J.A., Livingston, G.D.J., and Benasutti, M. (1992). Expression andIida, S., Rao, P.H., Butler, M., Corradini, P., Boccadoro, M., Klein,B., Chaganti, R.S.K., and Dalla-Favera, R. (1997). Deregulation of characterization of human FKBP52, an immunophilin that associates
with 90 KDa heat shock protein and is a component of steroidMUM1/IRF-4 by chromosomal translocation in multiple myeloma.Nat. Gen. 17, 226–230. receptor complexes. Proc. Natl. Acad. Sci. USA 89, 10974–10978.
Perrot-Applanat, M., Cibert, C., Geraud, G., Renoir, J.-M., and Bau-Jeang, K.-T., Derse, D., Matocha, M., and Sharma, O. (1997). Expres-sion status of the Tax protein in HTLV-I-transformed MT4 cells: lieu, E.-E. (1995). The 59 kDa FK506-binding protein, a 90 kDa heat
shock protein binding immunophilin (FKBP59-HBI), is associatedrecall of MT4 cells distributed by the NIH AIDS research and refer-ence reagent program. J. Virol. 71, 6277–6278. with the nucleus, the cytoskeleton and the mitotic apparatus. J. Cell
Sci. 108, 2037–2051.Kay, J.E. (1996). Structure-function relationships in the FK506-bind-ing protein (FKBP) family of the peptidylprolyl cis-trans isomerases. Poiez, B.F., Ruscetti, F.W., Gazadar, P.A., Bunn, P.A., Minna, J.D.,
and Gallo, R.C. (1980). Detection and isolation of type C retrovirusBiochem. J. 314, 361–385.particles from fresh cultured lymphocytes of a patient with cutane-Koromilas, A.E., Cantin, C., Craig, A.W.B., Jagus, R., Hiscott, J., andous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 77, 7415–7419.Sonnenberg, N. (1995). The interferon-inducible protein kinase PKR
modulates the transcriptional activation of immunoglobulin k gene. Radanyi, C., Chambraud, B., and Baulieu, E.E. (1994). The ability ofthe immunophilin FKBP59-HBI to interact with the 90-kDa heatJ. Biol. Chem. 270, 25426–25434.shock protein is encoded by its tetratricopeptide repeat. Proc. Natl.Leverson, J.D., and Ness, S.A. (1998). Point mutations in v-MybAcad. Sci. USA 91, 11197–11201.disrupt a cyclophilin-catalyzed negative regulatory mechanism. Mol.
Cell 1, 203–211. Stark, G.R., Kerr, I.M., Williams, B.R.G., Silverman, R.H., andSchreiber, R.D. (1998). How cells respond to interferons. Annu. Rev.Lin, R., Heylbroeck, C., Pitha, P.M., and Hiscott, J. (1998). VirusBiochem. 67, 227–264.dependent phosphorylation of the IRF-3 transcription factor regu-
lates nuclear translocation, transactivation potential and protea- Yamagata, T., Nishida, J., Tanaka, T., Sakai, R., Mitani, K., Yoshida,M., Taniguchi, T., Yazaki, Y., and Hirai, H. (1996). A novel interferonsome mediated degradation. Mol. Cell. Biol. 18, 2986–2996.regulatory factor family transcription factor, ICSAT/Pip/LSIRF, thatLin, R., Mamane, Y., and Hiscott, J. (1999). Structural and functionalnegatively regulates the activity of interferon-regulated genes. Mol.analysis of interferon regulatory factor 3: localization of the transacti-Cell. Biol. 16, 1283–1294.vation and autoinhibitory domains. Mol. Cell. Biol. 19, 2465–2474.Yoshida, M., Miyoshi, I., and Hinuma, Y. (1982). Isolation and charac-Lipsick, J.S. (1996). One billion years of Myb. Oncogene 13, 223–235.terization of retrovirus from cell lines of human adult T-cell leukemiaMamane, Y., Heylbroeck, C., Genin, P., Algarte, M., Servant, M.,and its implication in the disease. Proc. Natl. Acad. Sci. 79, 2031–Lepage, C., DeLuca, C., Kwon, H., Lin, R., and Hiscott, J. (1999).2035.Interferon regulatory factors: the next generation. Gene 237, 1–14.
Marks, A.R. (1996). Cellular functions of immunophilins. Physiol.Rev. 76, 631–649.
Matsuyama, T., Grossman, A., Mittrucker, H.-W., Siderovski, D.P.,Kiefer, F., Kawakami, T., Richardson, C.D., Taniguchi, T., Yoshinaga,S.K., and Mak, T.W. (1995). Molecular cloning of LSIRF, a lymphoid-specific member of the interferon regulatory factor family that bindsthe interferon-stimulated response element (ISRE). Nucleic AcidsRes. 23, 2127–2136.
Mattila, P.S. (1996). The actions of cyclosporin A and FK506 on Tlymphocyte activation. Biochem. Soc. Trans. 24, 45–49.
Mattila, P.S., Ullman, K.S., Fiering, S., Emmel, E.A., McCutcheon,M., Crabtree, G.R., and Herzenberg, L.A. (1990). The actions ofcyclosporin A and FK506 suggest a novel step in the activation ofT lymphocytes. EMBO J. 9, 4425–4433.
Mittrucker, H.-W., Matsuyama, T., Grossman, A., Kundig, T.M., Pot-ter, J., Shahinian, A., Wakeham, A., Patterson, B., Ohashi, P.S., andMak, T.W. (1997). Requirement for the transcription factor LSIRF/IRF4 for mature B and T lymphocyte function. Science 275, 540–543.
Miyamoto, M., Fujita, T., Kimura, Y., Maruyama, M., Harada, H.,Sudo, Y., Miyata, T., and Taniguchi, T. (1988). Regulated expressionof a gene encoding a nuclear factor, IRF-1, that specifically bindsto the IFN-b gene regulatory elements. Cell 54, 903–913.
Moore, P.S., Boshoff, C., Weiss, R.A., and Chang, Y. (1996). Molecu-lar mimicry of human cytokine and cytokine response pathwaygenes by KSHV. Science 274, 1739–1744.
Morris, R. (1994). Modes of action of FK506, cyclosporin A, andrapamycin. Transplant. Proc. 26, 3272–3275.
Nguyen, H., Hiscott, J., and Pitha, P.M. (1997). The growing familyof IRF transcription factors. Cytokine Growth Factor Rev. 8, 293–312.
Ortiz, M.A., Light, J., Maki, R.A., and Assa-Munti, N. (1999). Mutationanalysis of the Pip interaction domain reveals critical residues for




















