
kwazulu-natal situational overview - KZN Planning Commission
Upload
independentCategory
view
3download
0
Pa
SIAa
b
c
d
e
f
g
a
ARRAA
KPPPHhEA
1
mor[
g
RT
0d
Peptides 31 (2010) 2123–2129
Contents lists available at ScienceDirect
Peptides
journa l homepage: www.e lsev ier .com/ locate /pept ides
ost-natal stress-induced endocrine and metabolic alterations in mice atdulthood involve different pro-opiomelanocortin-derived peptides�
tefano Loizzoa,∗, Gabriele Campanab, Stefano Vellaa, Andrea Fortunaa, Gabriella Galiettaa,rene Guarinoa, Loredana Costaa, Anna Capassoc, Paolo Renzid, Giovanni V. Frajesee,f, Flavia Franconig,lberto Loizzoa, Santi Spampinatob
Department of Therapeutic Research and Medicines Evaluation, Istituto Superiore di Sanità, Roma, ItalyDepartment of Pharmacology, University of Bologna, Bologna, ItalyDepartment of Pharmacological Sciences, University of Salerno, Salerno, ItalyDepartment of Psychology, University of Roma “La Sapienza”, Roma, ItalyFaculty of Motor and Adaptive Sciences, University of Cassino, ItalyInternal Medicine, University of Roma Tor Vergata, ItalyCenter for Biotechnology, Development and Biodiversity Research, University of Sassari, Italy
r t i c l e i n f o
rticle history:eceived 27 October 2009eceived in revised form 1 August 2010ccepted 1 August 2010vailable online 19 August 2010
eywords:sychological stressain stressro-opiomelanocortinypothalamus–pituitary–adrenal (HPA)ormonesndogenous opioid system
a b s t r a c t
In previous investigations we added a physical stress (mild pain) to the “classical” post-natal psycho-logical stress in male mice, and we found that this combination produced a series of dysmetabolic signsvery similar to mild human type-2 diabetes. Here, for the first time we demonstrate that within this dia-betes model at least two groups of signs depend on the unbalance of two different endogenous systems.Newborn male mice were daily exposed to stressful procedures for 21 days (brief mother separationplus sham injection). Other groups underwent the same procedure, and also received naloxone (Na) toblock �–� endogenous receptors, or a phosphorothioate antisense oligonucleotide (AS) directed againstpro-opiomelanocortin (POMC)-mRNA [to block adrenocorticotropin (ACTH)- and POMC-derived opioidpeptides]. Adult mice which received only post-natal stress increased body weight (+7.5%), abdominaloverweight (+74%), fasting glycemia (+43%), plasma corticosterone (+110%), plasma (+169%) and pitu-itary (+153%) ACTH levels. Conversely, hypothalamic ACTH and corticotropin-releasing hormone (CRH)
ntisense oligonucleotide (AS)were reduced (−70% and −75%, respectively). Neonatal AS administration reverted all parameters tocontrol values. Neonatal naloxone had little or no influence on glucose, corticosterone, ACTH, CRH levels,whereas it prevented body overweight and abdominal overweight. We conclude that, within this type-2diabetes model in male mice at least two endocrino-neurohumoral systems are damaged, one concerningthe opioid system, and the other concerning HPA hormones. The use of the two drugs was of primary
te thies” f
importance to demonstradefined as “separate entit
. Introduction
Brain development and subtle hormonal feedback loop for-
ation in mammals continue after birth, and early-life eventsccurring during crucial time windows can influence the matu-ation of the nervous system and the functions under its control14]. This phenomenon, dubbed “early programming”, has received
� This paper is dedicated to the memory of Maria Sabina Pignatelli–Sissi, whoreatly inspired and influenced our investigations and our ethics.∗ Corresponding author at: Istituto Superiore di Sanità, Department of Therapeuticesearch and Medicines Evaluation, Viale Regina Elena 299, 00161 Roma, Italy.el.: +39 06 49902882; fax: +39 06 49902014.
E-mail address: [email protected] (S. Loizzo).
196-9781/$ – see front matter © 2010 Elsevier Inc. All rights reserved.oi:10.1016/j.peptides.2010.08.001
is statement, and to demonstrate that these two groups of signs could beollowing our complex post-natal stress model.
© 2010 Elsevier Inc. All rights reserved.
a great deal of attention from researchers because it could con-tribute to a number of adulthood diseases [10,19,23]. In humans,strong correlations have been identified between birth weight,plasma cortisol and the development of type-2 diabetes melli-tus (DM), and it has been proposed that programming of thehypothalamus–pituitary–adrenal (HPA) axis in utero is linked toinsulin resistance and type-2 DM in adult humans [23]. Studieshave focused on the contribution of stressful post-natal proceduresinvolving the HPA axis to behavioral alterations in adult rodents[18,23]. It is now emerging that stressful post-natal events applied
to rodents during the lactation period may also contribute to thedevelopment of metabolic programming alterations in adulthood.Previously, we reported that daily neonatal manipulations innon-genetically susceptible male mice could program mild over-weight in adulthood, preceded and accompanied by metabolic and

2 ides 31
heanstsabfmtOmamsp(ocat
ffcadawasoi
tai�ea
booegpao(atthwm
2
2
o
124 S. Loizzo et al. / Pept
ormonal alterations similar to those found in experimental mod-ls of mild type-2 DM and metabolic syndrome [8]. Moreover, wepplied a combination of repeated mild psychological plus mildociceptive stimuli (i.e., brief maternal deprivation plus a shamubcutaneous injection). This stressful procedure was applied forhe entire 3 weeks lactation period, when at least two differenttructures (the opioid receptors and the glucocorticoid receptors)re in their critical period to gain functional maturation in therain [21,27], and trigger the dysmetabolic features [18]. There-ore, we hypothesized that metabolic alterations in our adult
ale mice may involve both the endogenous opioid system andhe hormones of the HPA axis and/or their receptors unbalance.pioid–HPA interactions are presumably dependent on the stressechanisms themselves. Stress mechanisms can be summarized
s follows: stress induces release of corticotropin-releasing hor-one (CRH) from the hypothalamus, which in turn stimulates the
ynthesis and release of pro-opiomelanocortin (POMC) from theituitary. In turn, POMC produces adrenocorticotropin hormoneACTH) and the endogenous opioid �-endorphin, together withther peptides, by cleavage mechanisms. Finally, ACTH stimulatesorticosterone release from the adrenal gland. Thus, �-endorphinnd the ACTH–corticosterone complex may be considered impor-ant stress chain promoters [1,20,25,28].
We reported that a disruption of the corticosterone–ACTH–CRHeedback loop ensues in post-natally stressed male mice. This isollowed by enduring high plasma levels of both ACTH and corti-osterone with low hypothalamic ACTH and CRH from weaning,nd up to at least 69 days after the end of the stressful proce-ure [11]. Moreover, we have shown that behavioral and metaboliclterations including transitory nociceptive sensitivity, mild over-eight and immune system derangement observed in adulthood
fter post-natal stress are modulated by the endogenous opioidystem [16,24], whereas other alterations, including the responsef isolated organs to pharmacologic agents or electrical stimuli, arenfluenced chiefly by the HPA hormones [17].
The aim of this study was to investigate the contribution ofhese two components (the endogenous opioid system and the HPAxis) on metabolic (body weight, abdominal overweight and fast-ng glycemia) and hormonal parameters (plasma levels of ACTH,-endorphin and corticosterone, pituitary and hypothalamic lev-ls of ACTH) and on the hypothalamic content of CRH, which areltered by the stressful neonatal procedure used [18].
Groups of newborn male mice were daily stressed for 21 daysy a brief maternal deprivation plus subcutaneous (s.c.) injectionf saline. Other groups in addition were treated with the opi-id antagonist naloxone, with the aim of blocking the binding ofndogenous opioid peptides to � and � opioid receptors [16]. Otherroups of pups were treated with an antisense (AS) oligonucleotidehosphorothioate directed against POMC mRNA and thus acting asn antagonist for both ACTH–corticosterone- and POMC-derivedpioid peptide-mediated stress mechanisms, or with a mismatchMM) oligonucleotide phosphorothioate as control [31]. Metabolicnd hormonal parameters were evaluated 69 days after the lastreatment. We underline that only the use of the pharmacologicalools, naloxone and AS, allowed the demonstration of our workingypothesis, i.e., that there were two different bunches of signsithin the diabetes model, and these signs could be evidenced andodulated by specific drugs administration.
. Materials and methods
.1. Ethical approval
Experiments were carried out in accordance with the guidelinesf the Council of European Communities 86/609/EEC and protocols
(2010) 2123–2129
were approved by the Bioethical Committee of the Istituto Supe-riore di sanità (Roma, Italy) and by the Italian Ministry of Health.All efforts were made to minimize the number of animals used andtheir suffering.
2.2. General procedures
Animals were housed in environments controlled for light (12 hlight/dark cycle), temperature (20 ± 2 ◦C) and humidity (55 ± 5%)with water and food (Mucedola S.r.l., Settimo Milanese, Italy)available ad libitum; 1 g of food corresponded to 2.71 kcal. Theexperimental procedures were all performed in winter to avoidseasonal variation in receptor sensitivity [9]. Different series ofmultiparous pregnant female laboratory-born CD-1 mice (CharlesRiver Italia, Calco, Italy) were received at post-conception day 14,and the mice were housed one per cage. Pregnant females werechecked for litters twice daily (at 08:00 and 16:00). At about 12 hpost-natally the litters were all combined, randomly culled to sixmale pups per litter, and randomly cross-fostered. Each litter con-sisted of pups of the same weight to avoid interference determinedby the impairment of growth, which is an important determi-nant in the process of programming. To avoid manipulation ofcontrol mice, each complete litter received the same treatment,rather than each litter contributing to all treatments. Each litterwas randomly assigned to one of the following groups (the num-ber of experimental mice for each group is indicated in the figurecaptions and in Tables): (a) controls (C): pups were left undis-turbed with their mother, except for cage cleaning twice a week;(b) saline-treated (W): for 21 days, pups were removed (10 min)daily from the home cage and grouped in a container with freshbedding material, then each pup was weighed and given s.c. injec-tion under the skin of its back with sterile saline (1 ml/kg bodyweight) and returned to the home cage with their mother; (3)naloxone-treated (Na): pups were manipulated as in the vehi-cle group but received (−)naloxone hydrochloride (1 mg/kg bodyweight), instead of saline; (4) antisense-treated (AS): pups weremanipulated as in the saline-treated group and received the ASoligonucleotide (0.1 nmol/g body weight); (5) mismatch-treated(MM): pups were manipulated as in the saline-treated group andreceived MM oligonucleotide (0.1 nmol/g body weight). In a fewexperimental procedures, an additional group of pups that wereremoved daily from the home cage and grouped in a containerwith fresh bedding material and weighed (Wsp) was included;no injection was administered, and 10 min later the pups werereturned to the home cage with their mother. This latter groupwas added to study whether the “classic” stressful procedure con-sisting of brief daily maternal deprivation alone (manipulation)was able to induce metabolic changes in adulthood, and it wasconsidered as a positive control versus the saline-treated (W)group. At post-natal day 21, all animals were re-housed (three ani-mals from the same treated group per cage). Mice were weighedweekly and used for experimental procedures at the ages indicatedbelow.
Our protocol included purchasing pregnant mice from a breeder.This procedure was adopted several years ago by our group, andhas also been used by other investigators under similar condi-tions [37]. According to Shoener et al. [30], it is not likely thatmaternal stress resulting from transportation during the first fewdays of pregnancy affects fetal HPA axis programming. Our exper-iments support and extend this assertion, as our control mice didnot develop any metabolic, or hormonal alterations described for
pre-natal and post-natal stress. The data obtained from the con-trol mice were of the same order of magnitude as those fromrodents born of mothers that were not transferred during theirpregnancy, as previously described by us and other investigators[4,11,18]. We cannot rule out that the stress of travel might have
ides 31
ienm(
2
Iog(5sifwmoirct5
2
waaawTt
2
dovdbwa
2
T1t&t
2I
mtfa
the glucose levels to control values (Fig. 1C). Conversely, nalox-one treatment did not reduce the glycemia induced by the stressfulprocedures. Glucose levels in the Wsp mice were similar to thoseof controls [F(5,138) = 37.13 (ANOVA); P < 0.001].
Table 1Nociceptive threshold (in seconds) evaluated by the tail-flick assay in 30-days-oldmice exposed for 21 days after birth to the stressful procedures described in thetext.
C W Na AS Wsp MM
1.5 ± 0.2 6.3 ± 0.9* 1.8 ± 0.4 2.3 ± 0.6 2.0 ± 0.2 6.1 ± 0.8*
S. Loizzo et al. / Pept
nterfered somehow with the post-natal manipulation, even if ourxperimental design we could not find evidence of this possibility;o metabolic alterations were ever recorded in control mice or inice that underwent the stress of brief maternal deprivation only
Wsp).
.3. Drug treatment
(−)Naloxone hydrochloride was purchased from Sigma–Aldrichtalia (Milano, Italy). The dose (1 mg/kg) was indicated as the weightf the base compound and was chosen based on previous investi-ations [24]. AS and MM were produced by EUROBIO LaboratoriesLes Ulis Cedex, France). The 21-base sequence of AS-POMC was′-TCTGGCTCTTCTCGGAGGTCA-3′, and it reduces the synthesis ofome cleavage POMC-derived hormones (�-endorphin and ACTH)n in vivo and in vitro models [17,31]. To avoid the potential con-ounding factor of a huge decrease in the activity of the HPA axis,e selected an AS dose that was able to reduce the increased hor-one levels induced by handling, but not to below the basal level
f the POMC-derived molecules found in control mice [11]. Theres no apparent toxic effect produced by various doses of AS afterepeated administration in neonate and adult mice, which we hadonfirmed in previous studies [11,17]. The 21-base sequence ofhe MM phosphorothioate oligonucleotide, used as control, was:′-TGTGCCTCTTTCCGGTGGACA-3′.
.4. Evaluation of nociceptive threshold: tail-flick test
At post-natal day 30, the animals underwent a tail-flick test,hich mainly evaluates pain mediated by the spinal mechanism,
nd was performed according to Loizzo et al. [16]. Briefly, radi-nt heat was focused on a spot 15 mm from the tip of the tail,nd the intensity of the light beam was adjusted to obtain a tail-ithdrawal reaction time of about 1–2 s in the intact control animal.
he measurements were terminated at the first response or whenhe latency exceeded the cut-off time of 10 s.
.5. Abdominal overweight evaluation
After the animals were killed at 90 days of life, the two epi-idymal fat pads were removed for measurement of abdominalverweight. Each fat pad was dissected distally to the major bloodessel, and included the tissue around the testis epididymus anductus deferens. The tissues were washed in warm saline, thenlotted, and both pads were weighed together so that the total weteight could be determined for each animal. Data were computed
s absolute weight in g, and as percent of body weight.
.6. Glycemia testing
At the age of 90 days, the mice underwent testing for glycemia.he test was performed in the morning in groups of animals, after2 h fasting, with one drop of blood put on test strips that werehen inserted into a gluconometer (One-Touch EuroFlash; JohnsonJohnson Co). Each test was performed twice, and the mean of the
wo measures was recorded.
.7. Measurement of immunoreactive (IR)-ACTH,R-corticosterone, IR-ˇ-endorphin and IR-CRH
At 90 days of age, between 08:00 and 12:00 h, the three ani-als in each cage were rapidly guillotined at the same time by
hree independent investigators to minimize possible stress; noasting animals were used for these analyses. Plasma was collectednd stored at −80 ◦C until assayed. The pituitary and hypothalamus
(2010) 2123–2129 2125
were dissected on dry ice and immediately stored at −80 ◦C untilassayed.
The IR-corticosterone, IR-ACTH and IR-�-endorphin-like mate-rial was assayed using radioimmunassay kits (ICN, Costa Mesa, USAand Peninsula Laboratories Inc., Belmont, CA, USA, respectively). IR-CRH was extracted from the hypothalamus as described [11]. Theday of assay tissues were extracted in 0.1 mM acetic acid (90 ◦C)[31] and used throughout. All assays were measured in duplicatein the same day, according to previous described techniques [11].
2.8. Statistical evaluation
Analysis of variance (ANOVA) (one factor and 4–5 levels,depending on number of groups) and Dunnett’s test were used toestimate the significance level of the differences between groups.The accepted level of significance (two-tailed) was P < 0.05.
3. Results
3.1. Evaluation of nociceptive threshold
As expected, the nociceptive threshold of 30-days-old mice, 9days after the last stressful section, was higher in the W- and MM-treated mice compared with controls; the increase was preventedby naloxone and AS treatment [F(5,66) = 14.6 (ANOVA); P < 0.001]. Inthe Wsp group, which underwent maternal deprivation alone, nopain sensitivity alterations were recorded (Table 1).
3.2. Body weight and epididymal fat pads
As expected, at 90 days of age (69 days after the last stress-ful procedure), the body weight of W- and MM-treated mice wassignificantly heavier than that of controls (P < 0.01). This increasewas prevented by naloxone and AS administered during the post-natal period [F(5,66) = 3.91 (ANOVA); P < 0.01]. Wsp mice had bodyweights close to those of controls (Fig. 1A). The weight of the epi-didymal fat pads was consistently enhanced in W- and MM-treatedmice versus controls (P < 0.001) (Fig. 1B), and both naloxone and ASwere able to prevent this increase. The Wsp mice did not showany consistent variations versus controls [F(5,66) = 6.36 (ANOVA);P < 0.001].
3.3. Glycemia
Basal fasting glucose levels were consistently enhanced in W-and MM-treated mice (P < 0.01). AS prevented this effect, reducing
Newborn mice were randomly assigned to one of the following groups, as inFig. 1: controls (C); 10 min mother-deprived & saline-treated (W); 10 min mother-deprived & naloxone-treated (Na); 10 min mother-deprived & antisense-treated(AS); 10 min mother-deprived weighed and uninjected (Wsp); 10 min mother-deprived & mismatch-treated (MM). Values are the mean ± S.E.M. of 12 mice/group.
* P < 0.001 vs. C, Na, AS and Wsp.
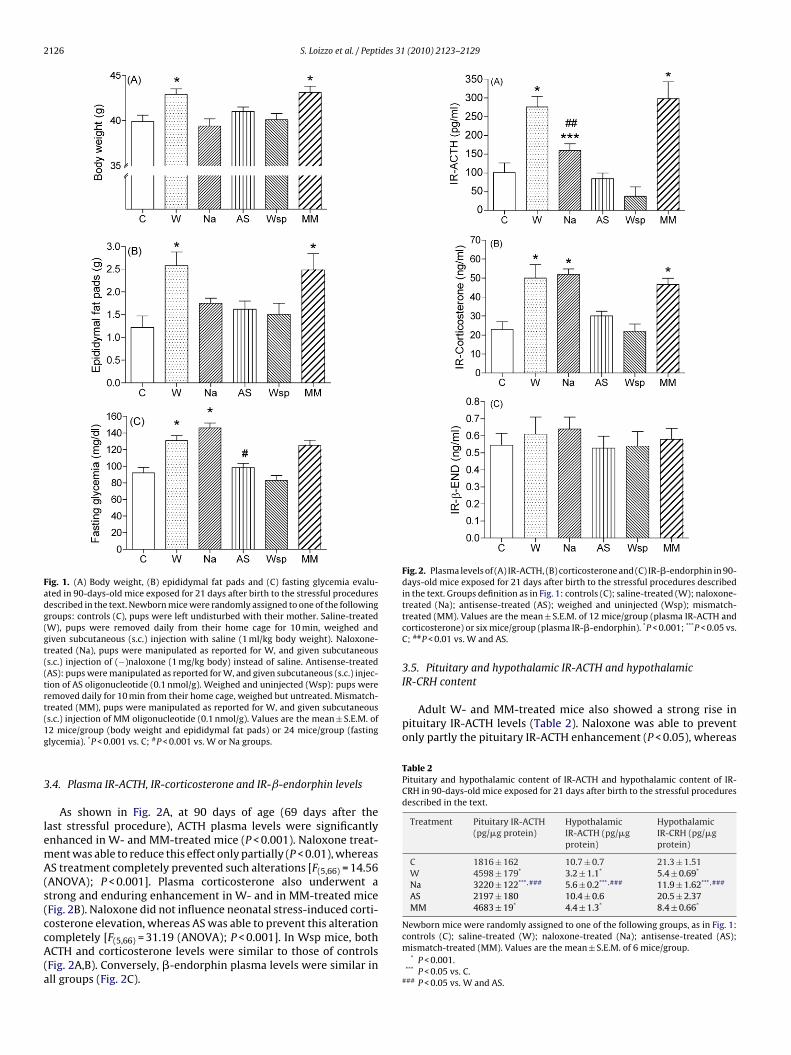
2126 S. Loizzo et al. / Peptides 31 (2010) 2123–2129
Fig. 1. (A) Body weight, (B) epididymal fat pads and (C) fasting glycemia evalu-ated in 90-days-old mice exposed for 21 days after birth to the stressful proceduresdescribed in the text. Newborn mice were randomly assigned to one of the followinggroups: controls (C), pups were left undisturbed with their mother. Saline-treated(W), pups were removed daily from their home cage for 10 min, weighed andgiven subcutaneous (s.c.) injection with saline (1 ml/kg body weight). Naloxone-treated (Na), pups were manipulated as reported for W, and given subcutaneous(s.c.) injection of (−)naloxone (1 mg/kg body) instead of saline. Antisense-treated(AS): pups were manipulated as reported for W, and given subcutaneous (s.c.) injec-tion of AS oligonucleotide (0.1 nmol/g). Weighed and uninjected (Wsp): pups wereremoved daily for 10 min from their home cage, weighed but untreated. Mismatch-treated (MM), pups were manipulated as reported for W, and given subcutaneous(1g
3
lemA(s(ccA(a
Fig. 2. Plasma levels of (A) IR-ACTH, (B) corticosterone and (C) IR-�-endorphin in 90-days-old mice exposed for 21 days after birth to the stressful procedures describedin the text. Groups definition as in Fig. 1: controls (C); saline-treated (W); naloxone-
IR-CRH content
Adult W- and MM-treated mice also showed a strong rise inpituitary IR-ACTH levels (Table 2). Naloxone was able to preventonly partly the pituitary IR-ACTH enhancement (P < 0.05), whereas
Table 2Pituitary and hypothalamic content of IR-ACTH and hypothalamic content of IR-CRH in 90-days-old mice exposed for 21 days after birth to the stressful proceduresdescribed in the text.
Treatment Pituitary IR-ACTH(pg/�g protein)
HypothalamicIR-ACTH (pg/�gprotein)
HypothalamicIR-CRH (pg/�gprotein)
C 1816 ± 162 10.7 ± 0.7 21.3 ± 1.51W 4598 ± 179* 3.2 ± 1.1* 5.4 ± 0.69*
Na 3220 ± 122*** ,### 5.6 ± 0.2*** ,### 11.9 ± 1.62*** ,###
AS 2197 ± 180 10.4 ± 0.6 20.5 ± 2.37MM 4683 ± 19* 4.4 ± 1.3* 8.4 ± 0.66*
Newborn mice were randomly assigned to one of the following groups, as in Fig. 1:
s.c.) injection of MM oligonucleotide (0.1 nmol/g). Values are the mean ± S.E.M. of2 mice/group (body weight and epididymal fat pads) or 24 mice/group (fastinglycemia). *P < 0.001 vs. C; #P < 0.001 vs. W or Na groups.
.4. Plasma IR-ACTH, IR-corticosterone and IR-ˇ-endorphin levels
As shown in Fig. 2A, at 90 days of age (69 days after theast stressful procedure), ACTH plasma levels were significantlynhanced in W- and MM-treated mice (P < 0.001). Naloxone treat-ent was able to reduce this effect only partially (P < 0.01), whereasS treatment completely prevented such alterations [F(5,66) = 14.56
ANOVA); P < 0.001]. Plasma corticosterone also underwent atrong and enduring enhancement in W- and in MM-treated miceFig. 2B). Naloxone did not influence neonatal stress-induced corti-osterone elevation, whereas AS was able to prevent this alteration
ompletely [F(5,66) = 31.19 (ANOVA); P < 0.001]. In Wsp mice, bothCTH and corticosterone levels were similar to those of controlsFig. 2A,B). Conversely, �-endorphin plasma levels were similar inll groups (Fig. 2C).
treated (Na); antisense-treated (AS); weighed and uninjected (Wsp); mismatch-treated (MM). Values are the mean ± S.E.M. of 12 mice/group (plasma IR-ACTH andcorticosterone) or six mice/group (plasma IR-�-endorphin). *P < 0.001; ***P < 0.05 vs.C; ##P < 0.01 vs. W and AS.
3.5. Pituitary and hypothalamic IR-ACTH and hypothalamic
controls (C); saline-treated (W); naloxone-treated (Na); antisense-treated (AS);mismatch-treated (MM). Values are the mean ± S.E.M. of 6 mice/group.
* P < 0.001.*** P < 0.05 vs. C.
### P < 0.05 vs. W and AS.

ides 31
tPhtftIit(
4
cb3isswetwwetat
aitictaa[
ewpisnto[eadbewtcm
m[�mpt
S. Loizzo et al. / Pept
he AS treatment completely prevented it [F(4,25) = 82.8 (ANOVA);< 0.001]. Conversely, W- and MM-treated mice showed very lowypothalamic IR-ACTH and IR-CRH levels at 90 days of age. Respec-ively, for hypothalamic IR-ACTH [F(4,25) = 15.9 (ANOVA); P < 0.001];or hypothalamic IR-CRH, [F (4,25) = 22.3 (ANOVA); P < 0.001]. ASreatment was able to restore the hypothalamic IR-ACTH andR-CRH levels to control values whereas naloxone treatmentnduced only a partial elevation of both hypothalamic peptideso intermediate values between controls and W-treated miceTable 2).
. Discussion
The present data confirm that mild psychological stressful pro-edures that are of short duration (10 min maternal deprivation)ut repeated daily, plus mild pain (s.c. saline injection), applied forweeks during the lactation period induce in male mice an endur-
ng disruption of the POMC–HPA hormone feedback mechanisms,hort-term disruption of the naloxone-sensitive endogenous opioidystem, and disruption of body and brain metabolic programming,ith long-term changes in several metabolic and hormonal param-
ters in the body and brain. Under our experimental conditions,he combination of the two procedures during the lactation periodas necessary to attain long-term metabolic/hormonal alterations,hereas maternal deprivation alone produced few or no metabolic
ffects. Conversely, this latter stressful model appears to be “pro-ective” for some learning capabilities shown by rats at a very oldge, and is accompanied by corticosterone levels that are lowerhan in controls [22].
These data are in line with different DM models in rodentsnd primates: adults in general show overweight, with increasesn plasma lipids and abdominal lipid storage, presence of hyper-rophic fat cells, fasting hyperglycemia and hyperinsulinemia,ncrease in production of some cytokines, and higher caloric effi-iency [3,13,16,18]. In type-2 diabetic models and in patients withype-2 DM, the HPA axis appears to be chronically stimulated, prob-bly as a result of disruption of negative feedback mechanismsnd HPA axis hormones, including increased plasma cortisol levels3,5,11,15,33].
Naloxone fully prevented elevation of nociceptive threshold,nhancement of body weight and induction of abdominal over-eight (epididymal fat pads) caused by the neonatal stressfulrocedure used. The endogenous opioid system is a determin-
ng factor for the modulation of pain sensitivity. We previouslyhowed that the stressful neonatal procedure used is accompa-ied by enhancement of plasma �-endorphin levels at the end ofhe manipulation period [11] and by enhancement of the thresh-ld to pain sensitivity 10 days later, up to the age of 45–50 days7,24]. Conversely, as reported here, at 90 days of age, plasma �-ndorphin is of the same order of magnitude in all groups (Fig. 2C)nd, at this age, male mice stressed during the neonatal periodo not show alterations in pain sensitivity [8,24]. A relationshipetween pain sensitivity alteration and �-endorphin plasma lev-ls is apparent, although other endogenous systems may interfereith pain sensitivity. For instance, Ploj and Nylander [26] showed
hat manipulation early in life induces persistent neurochemicalhanges in the cerebral �-opioid receptor system of young adultale rats.As regards metabolic alterations sensitive to naloxone treat-
ent, these findings agree with previous data of Tabarin et al.
32], who demonstrated reduced food conversion efficiency in-opioid receptor knockout mice versus wild-type mice. Further-ore, we confirmed that naloxone treatment during the lactatingeriod is able to reduce the food conversion efficiency to con-rol levels (see Supplementary data, Table S-2). Interestingly, AS
(2010) 2123–2129 2127
treatment, capable of blocking POMC-derived peptide synthesisand release, was effective in preventing neonatal stress-inducedmetabolic alterations. In agreement with these data, it has beenshown that increased glucocorticoid levels may promote vis-ceral obesity in adipocytes by overexpression of the enzyme11-�-hydroxysteroid-dehydrogenase-1 [36]. Thus, it seems likelythat there is a noteworthy interaction between the effectsmodulated by opioids and those modulated by corticosteroids[25].
In our mice, exposure to post-natal stress is followed by endur-ing elevation of plasma IR-ACTH and IR-corticosterone and ofpituitary IR-ACTH levels, and by enduring reduction in hypotha-lamic IR-ACTH and IR-CRH levels, thus indicating a breakdownof the pituitary feedback mechanisms, whereas the hypothala-mic feedback mechanisms apparently still continue to operate.All hormonal and behavioral–metabolic alterations studied in ourexperiments were prevented by AS treatment, blocking the pro-duction of POMC-derived peptides in a specific post-natal temporalwindow in mice exposed to a mild stressful procedure; these hor-monal alterations in turn contribute to the previously describedmetabolic alterations. By contrast, post-natal naloxone only par-tially prevented stress-induced alterations in plasma, pituitary andhypothalamic IR-ACTH and hypothalamic IR-CRH, whereas plasmaIR-corticosterone levels remained similar to those measured instressed mice treated with the vehicle. Higher glucocorticoidplasma levels may impair glucose tolerance, and hyperglycemia iscommon in patients with and without diabetes after glucocorticoidadministration [6]. Hyperglycemia can also be induced throughepidural glucocorticoid administration [12]. Therefore, preventionof hyperglycemia after plasma corticosterone normalization in AS-treated stressed mice was tested. Conversely, stressed mice treatedwith naloxone showed high plasma IR-corticosterone levels andhyperglycemia; these data strongly confirm the interplay betweenplasma glucocorticoid levels and glucose metabolism. Interest-ingly, mice stressed during the lactation period had overweight,abdominal overweight and fasting hyperglycemia, as it happensin type-2 DM in humans, whereas mice stressed and treated withnaloxone had normal weight, normal abdominal weight but stillhyperglycemia.
The data from the present study confirm and extend thatmetabolic derangements may arise, at least partly, following eventsoccurring early in life (programming), and that a combination ofmild psychological and nociceptive stimuli are essential to pro-duce these features in our mice. We show here that the enduringeffects induced by stressful procedures in mammals are modulatedby the type and duration of stress. Repeated psychological stress(brief maternal deprivation) induce long-term consequences differ-ent from those induced by repeated psychological stress plus painstress, and that these different long-term consequences have theirroots in different endogenous brain systems (e.g., opioid and/orACTH–corticosteroid systems).
We found a reliable increase in adult body weight in miceexposed post-natally to the complex stress procedure used. How-ever, differences between experimental protocols and animalspecies may be the reason for the different results obtainedby different investigators [2]. For example, other investigatorsfound alteration in pain sensitivity and in pharmacologic effectsof morphine in adult female rats after long (180 min) but notbrief (10–15 min) daily maternal separation. [35]. Others [34]found lower body weight and higher plasma ACTH but lowercortisol levels in adult boars after repeated brief neonatal mater-
nal deprivation. Finally, other investigators [13] using a modelof early-life stress (variable foraging demand) found that suchstress during a crucial period of neural development in macaquescan result in the peri-pubertal emergence of obesity and insulinresistance. Several questions remain unanswered, and further
2 ides 31
eedus
smmadooTtriStwbbipre
A
oHsLHgm(ibR
mataFa
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
128 S. Loizzo et al. / Pept
xperiments are needed to clarify the role exerted by differ-nt stress mechanisms and procedures, through (presumably)ifferent receptors and endogenous neurochemical structurespset, on different metabolic–endocrine–psychological signs andymptoms.
In conclusion, within our type-2 diabetes model naloxone-ensitive endogenous opioid system derangement after post-natalanipulation is sufficient to cause a number of significantetabolic and immune [16] alterations, while other metabolic
nd hormonal alterations being most sensitive to HPA hormoneerangement. We demonstrated such statement through the usef drugs which induce selective �–� opioid antagonism (nalox-ne) and opioid plus HPA-derived substances antagonism (AS).he present data suggest also that the endocrine/metabolic pat-erns found in stressed mice are similar to an acute stresseaction that is prolonged beyond the third month of life ands presumed, according to the original theory proposed byelye [29] to adapt the organism to a sudden physical reac-ion against an environmental insult. However, we wonderhether the prolonged endocrine–metabolic alterations triggered
y our model may evolve toward pathophysiological brain andody metabolic conditions, as a consequence of these endur-
ng changes on older age rodents, e.g., whether a prolongedlasma ACTH/corticosterone enhancement may result in increasedisk for cardiovascular damage and/or brain degenerative dis-ases.
cknowledgements
This investigation was supported in part by Istituto Superi-re di Sanita’ (ISS) through a ISS and US National Institutes ofealth joint research Project “Gender difference in seizure sen-
itivity: role of steroids and neuroactive steroids” to S Loizzo, Aoizzo and S Vella, in part by the ISS and US National Institutes ofealth joint research Project for Rare Diseases “Proposal for an inte-rated approach to rare diseases: a study between basic laboratoryodels and clinical epidemiology in amyotrophic lateral sclerosis
ALS)” to A Loizzo, S Vella and S Loizzo; in part by the Italian Min-stry of University and Research MIUR Project “Animal models andiological aspects of eating disorders” Prot. 2004068908 002 to Penzi.
We gratefully acknowledge the help given by “Friends for theemory of Maria Sabina Pignatelli–Sissi”. Patent pending for AS
nd its variants for the prevention and the treatment of PTSD, athe Ufficio Italiano Brevetti e Marchi, n. RM2007A000005. Thanksre due to Mrs Carla Campanella for the editing job, to Mr Stefanoidanza, Mr Adriano Urcioli and Mrs Antonella Romeo for valuablenimal care.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at doi:10.1016/j.peptides.2010.08.001.
eferences
[1] Akil HA, Morano MI. Stress. In: Bloom FE, Kupfer DJ, editors. Psychopharma-cology: the fourth generation of progress. New York: Raven Press; 1995. p.773–85.
[2] Anand KJS, Coskun V, Thrivikraman KV, Nemeroff CB, Plotsky PM. Long-term behavioral effects of repetitive pain in neonatal rat pups. Physiol Behav1999;66:627–37.
[3] Azuma K, Heilbronn L, Albu J, Smith SR, Ravussin E, Kelley DE. Adipose tissuedistribution in relation to insulin resistance of type 2 diabetes. Am J PhysiolEndocrinol Metab 2007;293:E435–42.
[4] Barbazanges A, Piazza PV, Le Moal M, Maccari S. Maternal glucocorticoidsecretion mediates long-term effects of prenatal stress. J Neurosci 1996;16:3943–9.
[
[
(2010) 2123–2129
[5] Bruehl H, Rueger M, Dziobek I, Sweat V, Tirsi A, Javier E, et al. Hypothalamic-pituitary-adrenal axis dysregulation and memory impairments in type 2diabetes. J Clin Endocrinol Metab 2007;92:2439–45.
[6] Clore JN, Thurby-Hay L. Glucocorticoid-induced hyperglycemia. Endocr Pract2009;15:469–74.
[7] d’Amore A, Marano G, Loizzo A. Reduced antinociceptive response to �-endorphin in adult mice after chronic neonatal handling. Physiol Behav1993;53:1025–7.
[8] d’Amore A, Mazzucchelli A, Loizzo A. Long-term changes induced by neonatalhandling in the nociceptive threshold and body weight in mice. Physiol Behav1995;57:1195–7.
[9] De Ceballos ML, De Felipe C. Circannual variation in opioid receptor sensitivityin mouse vas deferens. Eur J Pharmacol 1984;106:227–8.
10] De Giorgis GF, Nonnis E, Crocioni F, Gregori P, Rosini MP, Leuzzi V, et al. Evo-lution of daytime quiet sleep components in early treated phenylketonuricinfants. Brain Dev 1996;18:201–6.
11] Galietta G, Loizzo A, Loizzo S, Trombetta G, Spampinato S, Campana G, etal. Administration of antisense oligonucleotide against pro-opiomelanocortinprevents enduring hormonal alterations induced by neonatal handling in malemice. Eur J Pharmacol 2006;550:180–5.
12] Gonzalez P, Laker SR, Sullivan W, Harwood JE, Akuthota V. The effects of epidu-ral betamethasone on blood glucose in patients with diabetes mellitus. PM R2009;1:340–5.
13] Kaufman D, Banerji MA, Shorman I, Smith ELP, Coplan JD, Rosenblum LA, etal. Early-life stress and the development of obesity and insulin resistance injuvenile bonnet macaques. Diabetes 2007;56:1382–6.
14] Keverne EB. Understanding well-being in the evolutionary context of braindevelopment. Philos Trans R Soc Lond B Biol Sci 2004;359:1349–58.
15] Lesage J, Del-Favero F, Leonhardt M, Louvart H, Maccari S, Vieau D, et al. Pre-natal stress induces intrauterine growth restriction and programmes glucoseintolerance and feeding behaviour disturbances in the aged rat. J Endocrinol2004;181:291–6.
16] Loizzo A, Loizzo S, Lopez L, d’Amore A, Renzi P, Spampinato S, et al. Nalox-one prevents cell-mediated immune alterations in adult mice followingrepeated mild stress in the neonatal period. Br J Pharmacol 2002;135:1219–26.
17] Loizzo A, Capasso A, Galietta G, Severini C, Campana G, Spampinato S. Vasdeferens response to selective opioid receptor agonists in adult mice isimpaired following postnatal repeated mild stress. Eur J Pharmacol 2003;458:201–5.
18] Loizzo A, Loizzo S, Galietta G, Caiola S, Spampinato S, Campana G, et al.Overweight and metabolic and hormonal parameter disruption are inducedin adult male mice by manipulations during lactation period. Pediatr Res2006;59:111–5.
19] Maggisano V, Chiarotti F, Botunac I, Campanella C, Galietta G, Loizzo A. Ado-lescence as possible critical temporal window for blood pressure short termmonitoring in boys and girls. Eur J Epidemiol 2005;20:517–24.
20] Mattews SG. Early programming of the hypothalamo–pituitary–adrenal axis.Trends Endocrinol Metab 2002;13:373–80.
21] McDowell J, Kitchen I. Development of opioid systems: peptides, receptors andpharmacology. Brain Res 1987;434:397–421.
22] Meaney MJ, Szyf M, Seckl JR. Epigenetic mechanisms of perinatal program-ming of hypothalamic–pituitary–adrenal function and health. Trends Mol Med2007;13:269–77.
23] O’Regan D, Welberg LL, Holmes MC, Seckl JR. Glucocorticoid programmingof pituitary–adrenal function: mechanisms and physiological consequences.Semin Neonatol 2001;6:319–29.
24] Pieretti S, d’Amore A, Loizzo A. Long-term changes induced by developmentalhandling on pain threshold: effects of morphine and naloxone. Behav Neurosci1991;105:215–8.
25] Pieretti S, Di Giannuario A, Domenici MR, Sagratella S, Capasso A, SorrentinoL, et al. Dexamethasone-induced selective inhibition of the central mu opioidreceptor: functional in vivo and in vitro evidence in rodents. Br J Pharmacol1994;113:1416–22.
26] Ploj K, Nylander I. Long-term effects on brain opioid and opioid receptor like-1 receptors after short periods of maternal separation in rats. Neurosci Lett2003;345:195–7.
27] Rosenfeld P, van Eekelen JA, Levine S, de Kloet ER. Ontogeny of corticosteroidreceptors in the brain. Cell Mol Neurobiol 1993;13:295–319.
28] Sandman CA, Kastin AJ, Schally AV, Kendall JW, Miller LH. Neuroendocrineresponses to physical and psychological stress. J Comp Physiol Psychol1973;84:386–90.
29] Selye H. Confusion and controversy in the stress field. J Human Stress1975;1:37–44.
30] Shoener JA, Baig R, Page KC. Prenatal exposure to dexamethasone alters hip-pocampal drive on hypothalamic-pituitary-adrenal axis activity in adult malerats. Am J Physiol Regul Integr Comp Physiol 2006;290:R1366–73.
31] Spampinato S, Canossa M, Carboni L, Campana G, Leanza G, Ferri S. Inhibition ofproopiomelanocortin expression by an oligodeoxynucleotide complementaryto beta-endorphin mRNA. Proc Natl Acad Sci USA 1994;91:8072–6.
32] Tabarin A, Diz-Chaves Y, Carmona Mdel C, Catargi B, Zorrilla EP, Roberts AJ, etal. Resistance to diet-induced obesity in mu-opioid receptor-deficient mice:evidence for a “thrifty gene”. Diabetes 2005;54:3510–6.
33] Tojo C, Takao T, Nishioka T, Numata Y, Suemaru S, Hashimoto K. Hypothalamic-pituitary-adrenal axis in WBN/Kob rats with non-insulin dependent diabetesmellitus. Endocr J 1996;43:233–9.

ides 31
[
[
S. Loizzo et al. / Pept
34] Weaver SA, Aherne FX, Meaney MJ, Schaefer AL, Dixon WT. Neonatal handlingpermanently alters hypothalamic–pituitary–adrenal axis function, behaviour,and body weight in boars. J Endocrinol 2000;164:349–59.
35] Weaver SA, Diorio J, Meaney MJ. Maternal separation leads to persistent reduc-tions in pain sensitivity in female rats. J Pain 2007;8:962–9.
[
[
(2010) 2123–2129 2129
36] Wolf G. Glucocorticoids in adipocytes stimulate visceral obesity. Nutr Rev2002;60:148–51.
37] Zou B, Golarai G, Connor JA, Tang AC. Neonatal exposure to a novel environmentenhances the effects of corticosterone on neuronal excitability and plasticityin adult Hippocampus. Develop Brain Res 2001;130:1–7.
Copyright © 2022 FDOKUMEN




















