
Possible mechanisms involved in the discrepancy of hepatic and aortic endothelial nitric oxide...
9
BASIC STUDIES Possible mechanisms involved in the discrepancy of hepatic and aortic endothelial nitric oxide synthases during the development of cirrhosis in rats Morvarid Shir Mohammadi 1,2 , Dominique Thabut 1,3 , Dominique Cazals-Hatem 4 , Arnaud Galbois 1,2 , Marika Rudler 1,2 , Dominique Bonnefont-Rousselot 5,6 , Richard Moreau 1,2 , Didier Lebrec 1,2 and Khalid A. Tazi 1,2 1 INSERM, U773, Centre de Recherche Biom ´ edical Bichat-Beaujon, Ho ˆ pital Beaujon, Clichy, France 2 Universit ´ e Denis Diderot-Paris 7, site Bichat, Paris, France 3 Service d’Hepato-Gastroenterologie, Ho ˆ pital de la Piti ´ e-Salpe ˆ trie ` re, Paris, France 4 Service d’Anatomie et de Cytologie Pathologique, Ho ˆ pital Beaujon, Clichy, France 5 Laboratoire des Lipides, Ho ˆ pital la Piti ´ e-Salpe ˆ trie ` re, Paris, France 6 Laboratoire de Biochimie M ´ etabolique (EA 3617), Facult ´ e de Pharmacie, Paris 5, Paris, France Keywords Akt – Cav-1 – cirrhosis – eNOS – HDL – portal hypertension – SR-BI Abbreviations BDL, bile duct ligated; Cav-1, caveolin-1; eNOS, endothelial nitric oxide synthase; HDL, high-density lipoprotein; SR-BI, scavenger receptor class B type I; TNF, tumour necrosis factor. Correspondence Didier Lebrec, INSERM, U773, Centre de Recherche Biom ´ edical Bichat-Beaujon CRB3, Hopital Beaujon, Clichy, France Tel: 133 40 87 55 09 Fax: 133 1 47 30 17 11 e-mail: [email protected] Received 9 July 2008 Accepted 11 September 2008 DOI:10.1111/j.1478-3231.2008.01909.x Abstract Background/Aim: In cirrhosis, systemic nitric oxide (NO) overproduction and hepatic NO hypoproduction lead to arterial vasodilatation and portal hypertension. The mechanisms involved in these alterations in endothelial NO synthase (eNOS)-derived NO production in hepatic and systemic vasculature remain unknown. The aim of this study was to evaluate the regulation of eNOS and its major modulators in the liver and aorta during the development of cirrhosis in rats. Methods: Activated eNOS and Akt and expressions, and caveolin-1 (Cav-1) and scavenger receptor class B type I (SR-BI) expressions were measured before and 1, 2, 3 and 4 weeks after bile duct ligation. Plasma high-density lipoprotein (HDL) levels were measured. Results: Activated aortic eNOS increased at week 1, whereas it began to decrease at week 3 in the liver. Aortic expression of Cav-1 decreased at week 3 while hepatic expression increased by four- fold. Activated aortic Akt increased progressively while in the liver it gradually decreased during the development of cirrhosis. HDL levels decreased during the first week and decreased thereafter. The hepatic expression of SR-BI decreased. Conclu- sion: This study shows that the modulation of Akt and Cav-1 is inverted in the liver and the aorta during the development of cirrhosis. In addition, decreased HDL levels may play a role in reduced hepatic eNOS activity. In cirrhosis, altered vascular nitric oxide (NO) production is involved in the pathogenesis of increased intrahepatic vascular resistance and abnormal systemic vasodilatation (1, 2) and evidence suggests that endothelial NO synthase (eNOS) is the major enzyme involved in altered NO overproduction (3–7). eNOS activity and NO release are regulated by post-transla- tional control mechanisms mainly by phosphorylation and protein–protein interaction with caveolin-1 (Cav-1) (8–10). Cav-1 is the main negative regulatory protein for eNOS (9). Recently, the upregulation of sinusoidal Cav-1 and a decreased eNOS activity have been shown in the cirrhotic rat model (2). Previous studies have shown that eNOS must be phosphory- lated to exhibit full functional capacity (11). Multiple protein kinases can phosphorylate eNOS (12). These kinases include the serine/threonine kinase Akt (protein kinase B). Agonists such as shear stress, vascular endothelial growth factor and high-density lipoprotein (HDL) activate eNOS through its phosphorylation by Akt activation (8, 10). In cirrhotic rats, eNOS and Akt activities are reduced in hepatic sinusoidal endothelial cells (2, 7). Another previous study in the same rat model showed that arterial vasodilatation in cirrhosis might be because of increased systemic Akt activity (6). Recent studies have demonstrated that circulating factor, in particular HDL, regulates signalling in caveolae and is a potent activator of eNOS via scavenger receptor class B type I (SR-BI) and Akt (13). On the other hand, SR-BI expression can be modulated by several extracellular factors such as HDL. Previous studies have shown that plasma HDL levels are reduced in cirrhosis (14). Although SR-BI expression is known to depend on plasma HDL levels, expression of these receptors and their effects on eNOS activity in cirrhosis have not yet been studied. Because Cav-1 regulates the interaction between HDL and SR-BI, HDL bind- ing to SR-BI may be affected by Cav-1 expression (13). In addition, in cirrhosis, the overproduction of the pro-inflamma- tory cytokine tumour necrosis factor-a (TNF-a) is involved (5, 15, 16). In patients with cirrhosis, the plasma TNF-a levels Liver International (2009) 692 c 2009 The Authors. Journal compilation c 2009 Blackwell Publishing Ltd Liver International ISSN 1478-3223
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Possible mechanisms involved in the discrepancy of hepatic and aortic endothelial nitric oxide...

BAS IC STUDIES
Possiblemechanisms involved in the discrepancyof hepatic andaorticendothelial nitric oxide synthases during the developmentof cirrhosisin ratsMorvarid Shir Mohammadi1,2, Dominique Thabut1,3, Dominique Cazals-Hatem4, Arnaud Galbois1,2,Marika Rudler1,2, Dominique Bonnefont-Rousselot5,6, Richard Moreau1,2, Didier Lebrec1,2 andKhalid A. Tazi1,2
1 INSERM, U773, Centre de Recherche Biomedical Bichat-Beaujon, Hopital Beaujon, Clichy, France
2 Universite Denis Diderot-Paris 7, site Bichat, Paris, France
3 Service d’Hepato-Gastroenterologie, Hopital de la Pitie-Salpetriere, Paris, France
4 Service d’Anatomie et de Cytologie Pathologique, Hopital Beaujon, Clichy, France
5 Laboratoire des Lipides, Hopital la Pitie-Salpetriere, Paris, France
6 Laboratoire de Biochimie Metabolique (EA 3617), Faculte de Pharmacie, Paris 5, Paris, France
Keywords
Akt – Cav-1 – cirrhosis – eNOS – HDL – portal
hypertension – SR-BI
Abbreviations
BDL, bile duct ligated; Cav-1, caveolin-1;
eNOS, endothelial nitric oxide synthase; HDL,
high-density lipoprotein; SR-BI, scavenger
receptor class B type I; TNF, tumour necrosis
factor.
Correspondence
Didier Lebrec, INSERM, U773, Centre de
Recherche Biomedical Bichat-Beaujon CRB3,
Hopital Beaujon, Clichy, France
Tel: 133 40 87 55 09
Fax: 133 1 47 30 17 11
e-mail: [email protected]
Received 9 July 2008
Accepted 11 September 2008
DOI:10.1111/j.1478-3231.2008.01909.x
AbstractBackground/Aim: In cirrhosis, systemic nitric oxide (NO) overproduction and hepaticNO hypoproduction lead to arterial vasodilatation and portal hypertension. Themechanisms involved in these alterations in endothelial NO synthase (eNOS)-derivedNO production in hepatic and systemic vasculature remain unknown. The aim of thisstudy was to evaluate the regulation of eNOS and its major modulators in the liver andaorta during the development of cirrhosis in rats. Methods: Activated eNOS and Aktand expressions, and caveolin-1 (Cav-1) and scavenger receptor class B type I (SR-BI)expressions were measured before and 1, 2, 3 and 4 weeks after bile duct ligation.Plasma high-density lipoprotein (HDL) levels were measured. Results: Activated aorticeNOS increased at week 1, whereas it began to decrease at week 3 in the liver. Aorticexpression of Cav-1 decreased at week 3 while hepatic expression increased by four-fold. Activated aortic Akt increased progressively while in the liver it graduallydecreased during the development of cirrhosis. HDL levels decreased during the firstweek and decreased thereafter. The hepatic expression of SR-BI decreased. Conclu-sion: This study shows that the modulation of Akt and Cav-1 is inverted in the liverand the aorta during the development of cirrhosis. In addition, decreased HDL levelsmay play a role in reduced hepatic eNOS activity.
In cirrhosis, altered vascular nitric oxide (NO) production isinvolved in the pathogenesis of increased intrahepatic vascularresistance and abnormal systemic vasodilatation (1, 2) andevidence suggests that endothelial NO synthase (eNOS) is themajor enzyme involved in altered NO overproduction (3–7).eNOS activity and NO release are regulated by post-transla-tional control mechanisms mainly by phosphorylation andprotein–protein interaction with caveolin-1 (Cav-1) (8–10).Cav-1 is the main negative regulatory protein for eNOS (9).Recently, the upregulation of sinusoidal Cav-1 and a decreasedeNOS activity have been shown in the cirrhotic rat model (2).Previous studies have shown that eNOS must be phosphory-lated to exhibit full functional capacity (11). Multiple proteinkinases can phosphorylate eNOS (12). These kinases includethe serine/threonine kinase Akt (protein kinase B). Agonistssuch as shear stress, vascular endothelial growth factor andhigh-density lipoprotein (HDL) activate eNOS through itsphosphorylation by Akt activation (8, 10). In cirrhotic rats,
eNOS and Akt activities are reduced in hepatic sinusoidalendothelial cells (2, 7). Another previous study in the same ratmodel showed that arterial vasodilatation in cirrhosis might bebecause of increased systemic Akt activity (6). Recent studieshave demonstrated that circulating factor, in particular HDL,regulates signalling in caveolae and is a potent activator ofeNOS via scavenger receptor class B type I (SR-BI) and Akt(13). On the other hand, SR-BI expression can be modulated byseveral extracellular factors such as HDL. Previous studies haveshown that plasma HDL levels are reduced in cirrhosis (14).Although SR-BI expression is known to depend on plasma HDLlevels, expression of these receptors and their effects on eNOSactivity in cirrhosis have not yet been studied. Because Cav-1regulates the interaction between HDL and SR-BI, HDL bind-ing to SR-BI may be affected by Cav-1 expression (13). Inaddition, in cirrhosis, the overproduction of the pro-inflamma-tory cytokine tumour necrosis factor-a (TNF-a) is involved(5, 15, 16). In patients with cirrhosis, the plasma TNF-a levels
Liver International (2009)692 c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd
Liver International ISSN 1478-3223

progressively increased in relation to the severity of liverdysfunction (15). In cirrhotic rats, a recent study has shownthat the administration of reconstituted HDL reduces theeffects of endotoxin on TNF-a production, restores hepaticeNOS activity and lowers portal pressure (7). Because plasmaHDL levels are reduced and TNF-a is increased in cirrhosis,we hypothesized that this deregulation may play a role in thealteration of eNOS activity. To confirm this hypothesis, thepresent study investigated the regulation of eNOS and its majormodulators (Akt, Cav-1, TNF, HDL and SR-BI) in both the liverand the aorta during the development of cirrhosis in rats.
Materials and methods
Animals/protocols
Male Sprague–Dawley rats weighing between 225 and 250 g(Charles River Laboratories, Saint-Aubin-les-Elbeuf, France)were divided into two groups. The first group included ratsthat underwent bile duct ligation (BDL) as described previously(17). Briefly, under ketamine (100 mg/kg bw, intraperitoneally)anaesthesia, the common bile duct was exposed by medianlaparotomy and occluded by double ligature with a non-resorbable (silk 3-0) suture. The common bile duct was thensectioned and resected between the two ligatures and theabdominal incision was closed. Studies were performed 1, 2, 3and 4 weeks after BDL to allow the development of secondarybiliary cirrhosis. Cirrhosis was confirmed by macroscopic liverhistology examination. There were 31 rats in this group (BDL).
A second group included normal rats (control rats). Studieswere performed in cirrhotic and control rats weighing between300 and 380 g. Protocols performed in this laboratory wereapproved by the French Agriculture Office in accordance withEuropean legislation for research involving animals.
In this study, animals were sacrificed at weeks 1, 2, 3 and 4after ligation. Control rats were sacrificed by decapitationimmediately after they had adapted to the laboratory environ-ment. Immediately after sacrifice, blood was obtained from theneck, collected into heparinized tubes, centrifuged (3000g,10 min, at 37 1C) and stored at � 80 1C. Blood samples wereobtained to measure plasma levels of TNF-a, cholesterol, HDLand aspartate aminotransferase (AST) and alanine aminotrans-ferase (ALT) activities. Liver sections and the thoracic aortawere removed and flash frozen in liquid nitrogen. Both organswere stored at � 80 1C.
A third group of four control and 13 cirrhotic rats receivedreconstituted HDL (rHDL) (80 mg/kg, intravenously) as de-scribed previously (7). Rats were sacrificed by decapitation after4 h. Immediately after sacrifice, blood samples were taken; theliver section and the thoracic aorta were removed as describedabove. Hepatic and aortic eNOS phosphorylations with proteinexpressions and Akt phosphorylation were measured. PlasmaTNF-a concentration was also measured.
Serum transaminase activities
Serum aspartate transaminase and alanine transaminase activ-ities were measured with an Lx20 Beckman automatic analyzer(Beckman Coulter, Villepinte, France).
Serum cholesterol and high-density lipoprotein levels
Total serum cholesterol concentrations were determined enzyma-tically with a Konelabs kit (Thermoclinical Labsystems, Cergy-
Pontoise, France) on a Konelab 30i analyzers. HDL cholesterolwas determined using a phosphotungstic acid/MgCl2 reagent(Boehringer, Mannheim, Germany) to precipitate the apoB-containing lipoproteins (18, 19) and cholesterol was measured inthe supernatant, as described earlier in serum.
Determination of plasma tumour necrosis factor-aconcentrations
Plasma TNF-a levels were measured with a commerciallyavailable solid-phase enzyme-linked immunosorbent assay ac-cording to the protocol supplied by the manufacturer (R&DSystems, Minneapolis, MN, USA). Standards and samples wereincubated with a TNF-a antibody-coated 96-well microplate. Anenzyme-linked polyclonal antibody specific for rat TNF-a wasthen added after washing. The intensity of the colour wasmeasured in a microplate reader (Molecular Devices, Sunnyvale,CA, USA).
Liver histology
Liver samples for histological examination were obtained fromall rats. Liver tissues were fixed in 10% buffered formalin,embedded in paraffin, sectioned and stained with haematein–eosin and Masson trichromic stain. The proportion of hepato-cyte fibrosis was assessed by semiquantitative evaluation. F1corresponded to minimum non-extensive portal fibrosis; F2corresponded to extensive portal fibrosis; F3 correspondedto septal extensive portal fibrosis; and F4 corresponded tocirrhosis (20).
Western blotting
The thoracic aorta and liver tissues were homogenized in a lysisbuffer (500 and 600 ml, respectively) containing 320 mmol/Lsucrose, 50 mmol/L Tris HCl, 1 mmol/L ethylenediaminetetraa-cetic acid, 1 mmol/L dithiothreitol, 1% Nonidet P-40, 0.1%sodium dodecyl sulphate (SDS), 10 mg/ml leupeptin, 10 mg/mltrypsin inhibitor, 5 mg/ml aprotinin, 100mg/ml phenylmethyl-suphonyl fluoride and a protease inhibitor cocktail tablet(Roche Diagnostics, Mannheim, Germany), pH 7.5. Sampleswere sonicated three times for 20 s. Homogenates were centri-fuged at 12 000g for 5 min at 4 1C. The supernatant was used todetermine Cav-1, SR-BI, phosphorylated Akt, eNOS and phos-phorylated eNOS proteins. Total protein concentrations werequantified using the Bio-Rad protein determination assay (Bio-Rad Laboratories Ltd, Hemel Hempstead, UK) (21), and analiquot of homogenate was suspended in SDS sample buffer(2% SDS, 2% b-mercaptoethanol and 10% glycerol in300 mmol/L Tris-Cl, pH 6.8). Equal amounts of total protein(5–15 mg) were then subjected to SDS-polyacrylamide gelelectrophoresis. Gels were transferred to nitrocellulose. Nitro-cellulose filters were blocked by incubation in 5% non-fat drymilk, 1% bovine serum albumin, 0.1 mol/L NaCl, 0.01 mol/LTris-base (pH 7.5) and 0.1% Tween-20 overnight at 4 1C. Blotswere incubated for 1 h with specific antibodies that recognizeCav-1, phosphorylated Akt at Ser 473 (Santa Cruz Biotechnol-ogy Inc., Santa Cruz, CA, USA), SR-BI (Novus Biologicals Inc.,Littleton, CO, USA), eNOS and phosphorylated eNOS at Ser1177 (Cell Signalling Technology, Beverly, MA, USA) in thesame buffer. The same membrane was reprobed with theantibody to b-actin (Sigma, St Louis, MO, USA), which wasused as an internal control for protein loading. Blots werewashed in 0.1 mol/L NaCl, 0.01 mol/L Tris-base (pH 7.5) and
Liver International (2009)c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd 693
Shir Mohammadi et al. Discrepancy of hepatic and aortic endothelial NOS

0.1% Tween 20, and incubated for 1 h at 25 1C with a horse-radish peroxidase-conjugated secondary antibody (Santa CruzBiotechnology Inc.). Blots were again washed in 0.1 mol/LNaCl, 0.01 mol/L Tris-base (pH 7.5) and 0.1% Tween-20 andimmunoreactivity was assessed using an enhanced chemilumi-nescent Western blot detection system according to the manufac-turer’s instructions (Amersham International, Buckinghamshire,UK). The immunoreactive bands were quantified by videodensitometry. Values are given as arbitrary units. Results areexpressed as percentages of control values.
Phosphorylation of Akt and endothelial nitric oxidesynthase
The activities of Akt and eNOS were assessed by measuring theexpression of phosphorylated Akt at Ser 473 and phosphory-lated eNOS at Ser 1177 respectively (5).
Immunohistochemistry
Caveolin-1 immunohistochemistry was determined with aBMK iVIEW DAB Paraffine V.1s kit using the Module bench-mark IHC/ISH analyzers (Ventana, Strasbourg, France).Immunostaining was performed on a formalin-fixed paraffin-embedded section of liver tissue. The antibody used was anti-Cav-1 (Santa Cruz Biotechnology Inc.), at a dilution of 1/100.
Statistical analysis
Values are mean� standard error of the mean. Two-wayanalysis of variance with repeated measures and Student’s t-testfor paired and unpaired data were used when appropriate.P value o 0.05 was considered significant.
Results
Serum transaminase activities during cirrhosisdevelopment
Serum AST and ALT activities increased during the develop-ment of cirrhosis and were significantly higher in cirrhotic ratsthan in controls (Table 1). AST activity began to increase onweek 1 after the induction of cirrhosis, whereas ALT activityincreased 4 weeks later.
Plasma cholesterol and high-density lipoprotein levelsduring the development of cirrhosis
Plasma cholesterol levels were higher in cirrhotic than incontrol rats. In contrast, plasma HDL levels were reducedduring the development of cirrhosis (Table 2). Plasma HDLlevels decreased as of the first week after induction of cirrhosis
and decreased gradually during the development of cirrhosis(Table 2).
Plasma tumour necrosis factor-a levels during thedevelopment of cirrhosis
Plasma TNF-a levels started to increase significantly the firstweek after induction of cirrhosis (from 5.2� 2.4 to51.0� 33.1 pg/ml, Po 0.05) and TNF-a levels reached78.1� 13.3 pg/ml in cirrhotic rats (4 weeks after the inductionof cirrhosis).
Liver histology during the development of cirrhosis
Liver histology showed four different degrees of fibrosis in BDLrats (F1, F2, F3 and F4). The first week after the induction ofcirrhosis, F1 and F2 fibrosis was seen. Two weeks after theinduction of cirrhosis, F2 and F3 were found, 3 weeks after F2,F3 and F4 and 4 weeks after F3 and F4 (Fig. 1).
Aortic and hepatic endothelial nitric oxide synthaseexpressions and activity during the development ofcirrhosis
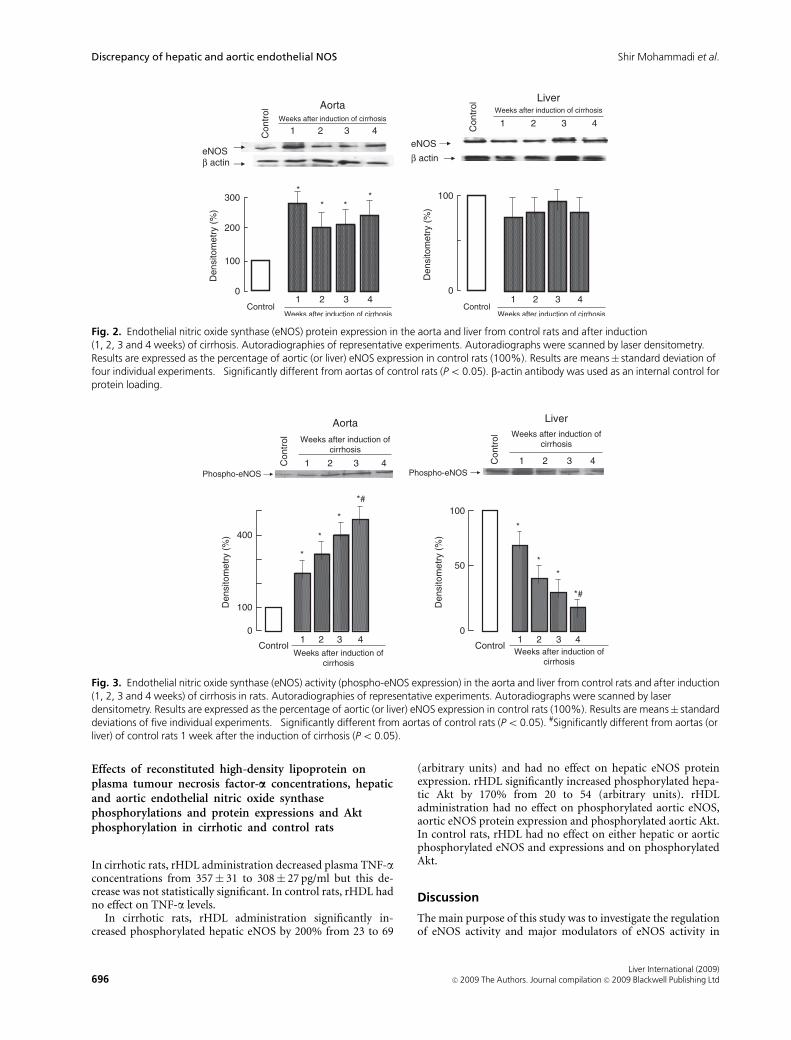
In cirrhotic rats, aortic eNOS expression was overexpressed inthe first week after induction of cirrhosis and remained un-changed up to week 4. Hepatic eNOS expression was un-changed during the development of cirrhosis (Fig. 2).
Aortic phosphorylated eNOS at serine-1177 (activatedeNOS) started to increase by approximately two-fold the firstweek after the induction of cirrhosis and gradually increasedduring the development of cirrhosis. In contrast, hepatic eNOSactivity began to decrease at the same time and continued todecrease up to week 4 (Fig. 3).
Table 1. Serum aspartate aminotransferase and alanineaminotransferase activities during the development of cirrhosis
Control
Weeks after induction of cirrhosis
1 2 3 4
AST 193�24 346� 54 273� 21 386� 58� 396� 48�
ALT 56�2 77 (12) 71 (8) 79 (13) 120 (29)�
Results are expressed (UI/L) as means� standard error of the means.�Significantly different from control rats (Po 0.05). n = 5 rats in each
group.
ALT, alanine aminotransferase; AST, aspartate aminotransferase.
Table 2. Plasma cholesterol and high-density lipoprotein levels during the development of cirrhosis
Control
Weeks after induction of cirrhosis
1 2 3 4
HDL 0.47�0.04 0.26� 0.02� 0.25� 0.03� 0.24� 0.11� 0.16� 0.08�
Cholesterol 0.65�0.03 0.82� 0.05� 0.75� 0.17 0.69� 0.01 0.78� 0.05�
Results are expressed (g/L) as means� standard error of the means.�Significantly different from control rats (Po 0.05). n = 5 rats in each group.
HDL, high-density lipoprotein.
Liver International (2009)694 c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd
Discrepancy of hepatic and aortic endothelial NOS Shir Mohammadi et al.

Phosphorylated aortic and hepatic Akt during cirrhosisdevelopment
Aortic phosphorylated Akt (activated Akt) started to increase byapproximately 1.5-fold the first week after the induction ofcirrhosis and gradually increased during the development ofcirrhosis. In contrast, phosphorylated Akt decreased by approxi-mately 60% in the liver the second week after the induction ofcirrhosis and continued to decrease up to week 4 (Fig. 4).
Aortic and hepatic caveolin-1 expressions during thedevelopment of cirrhosis
Aortic Cav-1 expression began to decrease the third week afterthe induction of cirrhosis, while hepatic Cav-1 expressionincreased by four-fold after the induction of cirrhosis (Fig. 5).
Immunohistochemistry of hepatic caveolin-1 during thedevelopment of cirrhosis
Low levels of Cav-1 were detected in the portal vessels but not inthe sinusoids of control rats, while Cav-1 was evident in theportal vasculature and was detected predominantly in thesinusoids in cirrhotic rats (F1–F4) (Fig. 6).
Aortic and hepatic scavenger receptor class B type Iexpressions
Aortic SR-BI expression was unchanged during the develop-ment of cirrhosis, whereas hepatic SR-BI expression signifi-cantly decreased in the third week after the induction ofcirrhosis compared with controls (Fig. 7).
A C
B D
E
Fig. 1. Rat liver tissue during the development of cirrhosis. Haematein–eosin staining (A, B, C, D, E) �250. (A) Control rat;(B) non-extensive portal fibrosis, F1; (C) extensive portal fibrosis, F2; (D) septal fibrosis, F3; and (E) cirrhosis, F4. Arrows show theportal fibrosis.
Liver International (2009)c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd 695
Shir Mohammadi et al. Discrepancy of hepatic and aortic endothelial NOS
Effects of reconstituted high-density lipoprotein onplasma tumour necrosis factor-a concentrations, hepaticand aortic endothelial nitric oxide synthasephosphorylations and protein expressions and Aktphosphorylation in cirrhotic and control rats
In cirrhotic rats, rHDL administration decreased plasma TNF-aconcentrations from 357� 31 to 308� 27 pg/ml but this de-crease was not statistically significant. In control rats, rHDL hadno effect on TNF-a levels.
In cirrhotic rats, rHDL administration significantly in-creased phosphorylated hepatic eNOS by 200% from 23 to 69
(arbitrary units) and had no effect on hepatic eNOS proteinexpression. rHDL significantly increased phosphorylated hepa-tic Akt by 170% from 20 to 54 (arbitrary units). rHDLadministration had no effect on phosphorylated aortic eNOS,aortic eNOS protein expression and phosphorylated aortic Akt.In control rats, rHDL had no effect on either hepatic or aorticphosphorylated eNOS and expressions and on phosphorylatedAkt.
Discussion
The main purpose of this study was to investigate the regulationof eNOS activity and major modulators of eNOS activity in
43214321
1 2 3 4 21 3 4Control Control
0
100
200
300*
* * *
Aorta
Den
sito
met
ry (
%)
0
100
Den
sito
met
ry (
%)
Con
trol
Con
trol
eNOSeNOS
β actin β actin
Liver
Fig. 2. Endothelial nitric oxide synthase (eNOS) protein expression in the aorta and liver from control rats and after induction(1, 2, 3 and 4 weeks) of cirrhosis. Autoradiographies of representative experiments. Autoradiographs were scanned by laser densitometry.Results are expressed as the percentage of aortic (or liver) eNOS expression in control rats (100%). Results are means� standard deviation offour individual experiments. �Significantly different from aortas of control rats (Po 0.05). b-actin antibody was used as an internal control forprotein loading.
Weeks after induction ofcirrhosis
1 2 3 4
Weeks after induction ofcirrhosis
1 2 3 4
LiverAorta
100
400
100
50
0
Den
sito
met
ry (
%)
Den
sito
met
ry (
%)
0
*#
*
*
*
*
**
*#
1 2 3 4 1 2 3 4Control
Weeks after induction ofcirrhosis
ControlWeeks after induction of
cirrhosis
Con
trol
Con
trol
Phospho-eNOS Phospho-eNOS
Fig. 3. Endothelial nitric oxide synthase (eNOS) activity (phospho-eNOS expression) in the aorta and liver from control rats and after induction(1, 2, 3 and 4 weeks) of cirrhosis in rats. Autoradiographies of representative experiments. Autoradiographs were scanned by laserdensitometry. Results are expressed as the percentage of aortic (or liver) eNOS expression in control rats (100%). Results are means� standarddeviations of five individual experiments. �Significantly different from aortas of control rats (Po 0.05). #Significantly different from aortas (orliver) of control rats 1 week after the induction of cirrhosis (Po0.05).
Liver International (2009)696 c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd
Discrepancy of hepatic and aortic endothelial NOS Shir Mohammadi et al.

both the liver and the aorta during the development of cirrhosisin rats. Modulation of Akt and Cav-1 (a major eNOS activatorand inhibitor respectively) was reversed in the liver and theaorta during the development of cirrhosis. In addition, TNF-aand HDL levels began to increase and decrease, respectively, 1week after the induction of cirrhosis and liver SR-BI expressionswere downregulated in cirrhotic rats. These results suggest that
decreased circulating HDL and SR-BI downregulation played arole in reduced hepatic eNOS activity probably through an Akt-and Cav-1-dependent signalling pathway. This study alsoshowed that aortic SR-BI expression remained unchangedduring the development of cirrhosis, suggesting that otherfactors such as TNF-a are involved in the upregulation of aorticeNOS and Akt in cirrhotic rats.
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
Aorta Liver
100
200
100
50
0
Den
sito
met
ry (
%)
Den
sito
met
ry (
%)
0
* #
*
*
* #
Con
trol
Control Control
Con
trol
* #* #
* #
1 2 3 41 2 3 4
1 2 3 4 1 2 3 4
Fig. 4. Akt activity (phospho-Akt expression) in the aorta and liver from control rats and after induction (1, 2, 3 and 4 weeks) of cirrhosis.Autoradiographies of representative experiments. Autoradiographs were scanned by laser densitometry. Results are expressed as thepercentage of aortic (or liver) endothelial nitric oxide synthase expression in control rats (100%). Results are means� standard deviations offive individual experiments. �Significantly different from aortas of control rats (Po 0.05). #Significantly different from aortas (or liver) of controlrats 1 week after the induction of cirrhosis (Po0.05).
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
LiverAorta
200
100
0
Den
sito
met
ry (
%)
Den
sito
met
ry (
%)
*
Con
trol
Control Control
Con
trol
*
0
100
Cav-1 Cav-1
50
*
*
1 2 3 4
1 2 3 4
Weeks after induction ofcirrhosis
1 2 3 4
1 2 3 4
Fig. 5. Caveolin-1 (Cav-1) protein expression in the aortas and liver from control rats and after the induction (1, 2, 3 and 4 weeks) of cirrhosisin rats. Autoradiographies of representative experiments. Autoradiographs were scanned by laser densitometry. Results are expressed as thepercentage of aortic (or liver) endothelial nitric oxide synthase (eNOS) expression in control rats (100%). Results are means� standarddeviations of five individual experiments. �Significantly different from aortas of control rats (Po 0.05).
Liver International (2009)c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd 697
Shir Mohammadi et al. Discrepancy of hepatic and aortic endothelial NOS

The results of this study demonstrate that in cirrhoticrats, aortic eNOS activity is upregulated within the first weekand increases progressively during the development of cirrho-sis, while hepatic eNOS activity begins to decrease at thesame time and continues to decrease until week 4. A recentstudy has shown that on the first day after portal vein ligationin rats, early upregulation of eNOS activity is an initial eventleading to NO overproduction and vasodilatation (6). Similaropposite regulation of eNOS has been observed in the liverand lungs of cirrhotic rats with the hepatopulmonary syn-drome (22). eNOS phosphorylation by Akt was shown to bea potential mechanism for the induction of eNOS activity.Another study showed that Akt could directly phosphor-ylate eNOS and increased eNOS activity, leading to NOoverproduction, and an NOS inhibitor blocked Akt-mediatedvasodilatation, suggesting that Akt causes NO-mediated vaso-dilatation (5).
We also studied the role of Akt in the changes in eNOS andfound that aortic Akt activity increased progressively after BDLuntil week 4, while hepatic Akt decreased. Akt is known to be animportant downstream target of PI3K. Furthermore, the PI3K/Akt pathway has been shown to directly activate eNOS byphosphorylation at Ser1177, and can be induced by variouscytokines such as TNF-a (5, 6). These results showed that TNF-a levels started to increase during the first week after theinduction of cirrhosis, suggesting that TNF-a may be involvedin the early upregulation of aortic eNOS via the Akt pathway. Inaddition, it has recently been demonstrated that the adminis-tration of the neutralizing antibody to TNF-a to cirrhotic ratsinduced a decrease in aortic Akt and eNOS activities (5).
The role of Cav-1, a major eNOS inhibitor, was also studiedin both the liver and the aorta. One previous study showedincreased Cav-1 expression in the liver vasculature after BDL,but aortic changes of Cav-1 have not been studied. In the
D
A
C
B
E
Fig. 6. Immunostaining for caveolin-1 (Cav-1) in hepatic vasculature in control rats and after cirrhosis induction. (A, B, C, D, E) � 250.(A) Control rats liver (B, C, D, E) after induction of cirrhosis (1, 2, 3 and 4 weeks respectively). Cav-1 is more predominant and detected insinusoids. Arrows show the Cav-1 marker.
Liver International (2009)698 c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd
Discrepancy of hepatic and aortic endothelial NOS Shir Mohammadi et al.

present study, we demonstrated for the first time that aorticCav-1 expression is decreased during the development ofcirrhosis in rats and this reduction was more marked after week3, suggesting that decreased Cav-1 expression may be impli-cated in aortic eNOS upregulation. In addition, we confirmedan association between increased hepatic Cav-1 and hypercho-lesterolaemia, which has already been shown in BDL rats (9).This was confirmed by measuring plasma cholesterol levels incirrhotic rats when hepatic Cav-1 expression was significantlyincreased at week 4 compared with controls. In addition, Cav-1was shown to be over-expressed in endothelial sinusoid cells incirrhotic rats compared with normal rats by immunohisto-chemistry, suggesting that vascular NO production in endothe-lial is decreased in cirrhotic liver.
This study also investigated the role of HDL and its principalreceptor, SR-BI, as an important stimulator of eNOS activity inboth the liver and the aorta. HDL is a potent activator of eNOSvia SR-BI and Akt (23) and SR-BI can be modulated bycirculating HDL and Cav-1 levels (13). Previous studies haveshown that the plasma HDL levels are modified in cirrhosis(7, 24) and reduced plasma HDL levels have been demonstratedwhen cirrhosis worsens. Our results showed that plasma HDLlevels decreased progressively during the development of cir-rhosis and that hepatic expression of SR-BI decreased signifi-cantly, while it did not change in the aorta. These results suggestthat in cirrhosis, decreased HDL levels and downregulationof SR-BI play a role in reduced hepatic eNOS activity prob-ably through downregulation of Akt and over-expression ofCav-1. Further studies are needed to determine the exact roleof HDL in eNOS alterations during cirrhosis, for example todetermine whether administration of HDL (to normalizecirculatory HDL levels) to rats on week 1 after the induction ofcirrhosis modulates both aortic and hepatic eNOS, systemicand hepatic haemodynamics. The results of administration ofrHDL in cirrhotic rats partly answer to this question, showingthat HDL increases hepatic eNOS activity by increasing Aktactivity (phosphorylated Akt) and has no effect on aortic eNOS
activity. In contrast, TNF-a may have opposite effects of HDLbecause HDL administration decreases plasma TNF-a concen-trations (7).
Finally, there is no clear explanation as to why eNOSregulation differs between the aorta and the liver in cirrhosis.This study suggests, however, two different mechanisms. AorticeNOS upregulation evidenced by increased activated eNOS andprotein expression mainly depend on shear stress. Shear stressactivates Akt as other kinases that are the main modulators ofactivity and expression of aortic eNOS (5). In contrast, hepaticAkt activity is reduced as eNOS activity while hepatic eNOSexpression is not altered. In addition, it might be hypothesizedthat the reduction of HDL, the main activator of Akt, plays amajor role in the reduction of hepatic Akt activity and eNOSactivity (7). This hypothesis suggests that shear stress is absentin the liver or has no effect in the liver.
In conclusion, this study shows that the modulation of Aktand Cav-1 (a major eNOS activator and inhibitor respectively)is inverted in the liver and aorta during the development ofcirrhosis. In addition, decreased HDL levels and SR-BI down-regulation may play an important role in reduced hepatic eNOSactivity.
References
1. Sogni P, Moreau R, Gadano A, Lebrec D. The role of nitric oxide in
the hyperdynamic circulatory syndrome associated with portal
hypertension. J Hepatol 1995; 23: 218–24.
2. Langer DA, Shah VH. Nitric oxide and portal hypertension: inter-
face of vasoreactivity and angiogenesis. J Hepatol 2006; 44: 209–16.
3. Wiest P, Groszmann RJ. The paradox of nitric oxide in cirrhosis and
portal hypertension: too much, not enough. Hepatology 2002; 35:
478–91.
4. Tazi KA, Barriere E, Moreau R, et al. Role of shear stress in aortic
eNOS upregulation in rats with biliary cirrhosis. Gastroenterology
2002; 122: 1869–77.
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
Weeks after induction ofcirrhosis
LiverAorta
1 2 3 4
1 2 3 4 1 2 3 4
100
50
0
Den
sito
met
ry (
%)
Den
sito
met
ry (
%)
Con
trol
Control Control
Con
trol
*
0
100
50 *
SR-BI SR-BI
Weeks after induction ofcirrhosis
1 2 3 4
Fig. 7. Scavenger receptor class B type I (SR-BI) protein expression in the aortas and liver from control rats and after induction(1, 2, 3 and 4 weeks) of cirrhosis in rats. Autoradiographies of representative experiments. Autoradiographs were scanned by laserdensitometry. Results are expressed as the percentage of aortic (or liver) endothelial nitric oxide synthase expression in control rats (100%).Results are means� standard deviations of five individual experiments. �Significantly different from aortas of control rats (Po 0.05).
Liver International (2009)c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd 699
Shir Mohammadi et al. Discrepancy of hepatic and aortic endothelial NOS

5. Tazi KA, Moreau R, Herve P, et al. Norfloxacin reduces aortic NO
synthases and proinflammatory cytokine up-regulation in cirrhotic
rats: role of Akt signaling. Gastroenterology 2005; 129: 303–14.
6. Iwakiri Y, Tsai MH, McCabe TJ, et al. Phosphorylation of eNOS
initiates excessive NO production in early phase of portal hyperten-
sion. Am J Physiol Heart Circ Physiol 2002; 10: H2084–90.
7. Thabut D, Tazi KA, Bonnefont-Rousselot D, et al. HDL adminis-
tration attenuates liver proinflammatory response, restores liver
eNOS and lowers portal pressure in cirrhotic rats. Hepatology 2007;
46: 1893–906.
8. Minshall RD, Sessa WC, Stan RV, Anderson RGW, Malik A.
Caveolin regulation of endothelial function. Am J Physiol Lung Cell
Mol Physiol 2003; 285: L1179–83.
9. Shah V, Cao S, Hendrickson H, Yao J, Katusic ZC. Regulation of hepatic
eNOS by caveolin and calmodulin after bile duct ligation in rats. Am J
Physiol Gastrointest Liver Physiol 2001; 280: 1209–16.
10. Shaul PW. Regulation of endothelial nitric oxide synthase: location,
location, location. Annu Rev Physiol 2002; 64: 749–74.
11. Liu S, Premont RT, Kontos CD, Zhu S, Rockey DC. A crucial role for
GRK2 in regulation of endothelial cell nitric oxide synthase function in
portal hypertension. Nat Med 2005; 11: 952–8.
12. Boo YC, Ho H. Flow-dependent regulation of endothelial nitric
oxide synthase: role of protein kinases. Am J Physiol Cell Physiol
2003; 285: C499–508.
13. Matveev S, Uittenbogaard A, van Der Westhuyzen D, Smart EJ.
Caveolin-1 negatively regulates SR-BI mediated selective uptake of
high-density lipoprotein-derived cholesteryl ester. Eur J Biochem
2001; 268: 5609–16.
14. Camps J, Simo JM, Guaita S, Ferre N, Joven J. Altered composition
of lipoproteins in liver cirrhosis compromises three homogeneous
methods for HDL-cholesterol. Clin Chem 1999; 45: 685–8.
15. Giron-Gonzalez JA, Martinez-Sierra C, Rodriguez-Ramos C, et al.
Implication of inflammation-related cytokines in the natural his-
tory of liver cirrhosis. Liver Int 2004; 24: 437–45.
16. Wong F, Bernardi M, Balk R, et al. Sepsis in cirrhosis: report on the
7th meeting of the International Ascites Club. Gut 2005; 54: 718–25.
17. Lebrec D. Animal models in portal hypertension. In: Okuda K,
Benhamou JP, eds. Portal Hypertension: Clinical and Physiological
Aspects. Tokyo: Springer-Verlag, 1991; 101–13.
18. Allain CC, Poon LS, Chan CS, Richmond W, Fu PC. Enzymatic
determination of total serum cholesterol. Clin Chem 1997; 20:
470–5.
19. Leglise D. Commission de standardisation de l’ARCOL Recom-
mandations pratiques pour le dosage du HDL-C apres precipitation
par le phosphotungstate de sodium et le chlorure de magnesium.
Inf Sci Biol 1996; 13: 2–6.
20. Pinzani M. Non invasive evaluation of hepatic fibrosis. Gut 2006;
55: 310–2.
21. Bradford MM. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding. Ann Biochem 1976; 72: 248–54.
22. Frossard JL, Schiffer E, Cikirikcioglu B, et al. Opposite regulation of
endothelial NO synthase by HSP90 and caveolin in liver and lungs
of rats with hepatopulmonary syndrome. Am J Gastrointest Liver
Physiol 2007; 293: G864–70.23. Morales-Ruiz M, Cejudo-Martin P, Fernandez-Varo G, et al. Transduc-
tion of the liver with activated Akt normalizes portal pressure in
cirrhotic rats. Gastroenterology 2003; 125: 522–31.
24. Habib A, Mihas AA, Abou-Assi SG, et al. High-density lipoprotein
cholesterol as an indicator of liver function and prognosis in
noncholestatic cirrhotics. Clin Gastroenterol Hepatol 2000; 3:
286–91.
Liver International (2009)700 c� 2009 The Authors. Journal compilation c� 2009 Blackwell Publishing Ltd
Discrepancy of hepatic and aortic endothelial NOS Shir Mohammadi et al.




















