
Ponomarenko E.V., D. Crossland and J. Loo 2013. Reconstructing species composition at the time of...
14
Proceedings of the Fourth International Meeting of Anthracology Brussels, 8–13 September 2008 Royal Belgian Institute of Natural Sciences Edited by Freddy Damblon BAR International Series 2486 2013
Transcript of Ponomarenko E.V., D. Crossland and J. Loo 2013. Reconstructing species composition at the time of...

Proceedings of the Fourth International Meeting of
Anthracology Brussels, 8–13 September 2008
Royal Belgian Institute of Natural Sciences
Edited by
Freddy Damblon
BAR International Series 24862013

Published by
ArchaeopressPublishers of British Archaeological ReportsGordon House276 Banbury RoadOxford OX2 [email protected]
BAR S2486
Proceedings of the Fourth International Meeting of Anthracology: Brussels, 8–13 September 2008, Royal Belgian Institute of Natural Sciences
© Archaeopress and the individual authors 2013
ISBN 978 1 4073 1100 5
Cover image: Hunters in Snow (1565) Pieter Bruegel the Elder © Kunsthistorisches Museum Wien (Austria)
Printed in England by Information Press, Oxford
All BAR titles are available from:
Hadrian Books Ltd122 Banbury RoadOxfordOX2 7BPEnglandwww.hadrianbooks.co.uk
The current BAR catalogue with details of all titles in print, prices and means of payment is available free from Hadrian Books or may be downloaded from www.archaeopress.com

203
RECONSTRUCTING SPECIES COMPOSITION AT THE TIME OF LAND CLEARANCE: TWO APPROACHES COMPARED
Elena PONOMARENKO Ecosystem Archaeology Services, 1139 Agincourt Rd., Ottawa ON, K2C2H8, Canada
Donna CROSSLAND Kejimkujik National Park and National Historic Site, Maitland Bridge, Annapolis County, NS, B0T 1B0, Canada
Judy LOO Atlantic Forestry Centre, 1350 Regent Street South, Fredericton, New Brunswick, Canada
Abstract: Botanical identification of charcoal from plough horizons in abandoned farmlands was applied to reconstruct pre-agricultural species composition of Acadian forest in Kouchibouguac National Park (Canadian Maritime Lowlands). Results of the reconstruction were compared to archival descriptions of the forest ca 1800, just prior to intensive anthropogenic modification. Both methods detected a high diversity of tree species including species that are currently not common on the landscape, such as Ulmus and Ostrya. Both methods indicated that forests in the area were mixed and dominated by mid- to late successional species (Picea spp., Tsuga canadensis, Betula spp., Acer spp., Fagus grandifolia, Abies balsamea and Pinus strobus), whereas early successional species, such as Populus spp., Pinus resinosa, Larix laricina and Quercus rubra were present at low levels. According to both methods, Pinus banksiana – a fire-dependant species that is common and widespread in the modern landscape, was absent at the time of land clearance. Reconstructed occurrences of species were generally similar using either method, but anthracological research indicated lower occurrences of some coniferous canopy species, such as Tsuga canadensis, Abies balsamifera and Picea sp., and higher occurrences of species that fill-in subordinate canopy positions, particularly Thuja, in charcoal assemblages in relation to witness trees results. Comparison of the two methods confirmed applicability of the analysis of charcoal assemblages for the site-oriented reconstruction of tree species composition at the time of land clearance.
Key words: charcoal botanical analysis, historical forest composition, ecosystem dynamics, disturbance return interval, vegetation management, paleoecology, anthracology, witness trees
INTRODUCTION
Many Canadian national parks and national historic sites feature former farmlands that are slowly returning to a forested state. Restoration of forest in such areas requires knowledge of the historical species composition, i.e. species composition prior to land clearance. European colonization of the Atlantic coast of Canada began in the 1700s, and during the last 300 years coastal forests have been progressively modified by burning, logging, and introduced forest pests. As a result, modern forests in the area do not necessarily reflect either a potential species composition or species composition at the time of land clearance. In some areas of the Maritimes, the composition of early forests at the time of agricultural colonization can be reconstructed using surveyor plan drawings and descriptions from diaries of the pioneers and early travelers. In other areas, clearing occurred within several years, before any systematic descriptions of the initial forest cover were documented. According to the historical descriptions of the methods of forest clearing for farming in the Maritimes, trees were first logged; their trunks and large branches collected for a variety of uses, and the logging slash burned (Johnston 1851). Logging slash was often piled around stumps of cut trees to hasten disintegration. Burning of the logging
slash produced ash and charcoal associated with the site clearance event. Once ploughing began, the charcoal was incorporated into the soil and re-distributed within the plough layer (Johnston 1851). This study explored the possibility of reconstructing pre-agricultural forest cover using botanical identification of soil charcoal from plough layers of modern and abandoned farmlands. Results of the charcoal analysis on the forest composition at the time of land clearance were compared with the analysis of archival data, (using witness trees and early forest descriptions), to validate both approaches.
OBJECTS AND METHODS OF THE COMPARATIVE STUDY
Study area
The study area is situated on the east coast of New Brunswick, Canada, within the ‘Eastern Lowlands’ ecoregion (Rowe 1972; DNRE 1996) (Figure 1). This region is defined by low elevation (approximately three-quarters of the land is less than 15 m above sea level) and limited topographic variation. Gentle moraines, a few eskers and kames, and short, steep banks of streams and meandering rivers dissect the landscape in an east-west

PROCEEDINGS OF THE FOURTH INTERNATIONAL MEETING OF ANTHRACOLOGY
204
Figure 1. Location of the Eastern Lowlands Ecoregion, archival study area, and Kouchibouguac National Park (anthracological study area)
pattern to empty into the Northumberland Strait (Wang and Rees 1983; Beach 1988). Poor drainage is a prominent feature, resulting in large raised peat bogs over approximately 1/5 of park surface (Beach 1988). Local drainage patterns are discontinuous, often changing rapidly over distances of only a few meters. Kouchibouguac National Park (KNP) is a 239 km2
protected area of the Eastern Lowlands that conserves over 125 km2 of Acadian forest.
The Acadian Forest Region covers most of the area of the Maritime Provinces of Canada (Loucks 1962; Rowe 1972), and comprises approximately 35 tree species forming a high variety of associations and community types. Red and black spruce (Picea rubens Sarg., P. mariana (Mill.) BSP.) and their hybrids, balsam fir (Abies balsamea (L.) Mill.), red maple (Acer rubrum L.), Eastern hemlock (Tsuga canadensis (L.) Carr.) and white pine (Pinus strobus L.) comprise a distinctive association on the landscape (Loucks 1962). Another prominent
species, eastern white cedar (Thuja occidentalis L.), is considered an edaphic climax species on heavy soils with impeded drainage (Dryade 1979). KNP features the largest protected eastern cedar stand east of Ontario (Desloges 1980). Eastern larch (Larix laricina (DuRoi) K. Koch) and black spruce predominate on expansive bog lands (Loucks 1962).
The Acadian Forest Region was the earliest Canadian forest type to be exploited for timber resources. European settlement began in earnest in the study area in 1820-1830, and local forests were subjected to increased resource harvesting from that time onwards. Contemporary forests of the study area exhibit extensive stands of intolerant pioneer species with trembling aspen (Populus tremuloides Michx.), white birch (Betula papyrifera Marsh.), grey birch (B. populifolia Marsh.), white spruce (Picea glauca (Moench) Voss), larch, and green alder (Alnus viridis subsp. crispa (Aiton) Turrill) (Loucks 1962; DNRE 1996). Widespread fires appear to

E. PONOMARENKO ET AL.: RECONSTRUCTING SPECIES COMPOSITION AT THE TIME OF LAND CLEARANCE: TWO APPROACHES COMPARED
205
have favored coniferous species, particularly black spruce and jack pine (Pinus banksiana Lamb.) (Rowe 1972).
Park vegetation currently comprises 26 tree species, forming 37 forest community types based on tree species composition, understory vegetation and site condition (Beach 1988). Black spruce, red spruce, balsam fir, trembling aspen, and white cedar, in various associations, account for approximately 70% of species on forested land (Desloges 1980). Narrow bands of better drained soil feature tolerant hardwood growth, including sugar maple (Acer saccharum Marsh.), beech (Fagus grandifolia Ehrh.), and small pockets of white ash (Fraxinus americana L.). These species, along with yellow birch (Betula alleghaniensis Britt.), are minor components, and have probably been greatly reduced in frequency, as the better-drained landscapes were favored for human settlement and agriculture (Desloges 1980).
Anthracological research of historical forests
Prior to European colonization, the inland area of Kouchibouguac National Park was used mainly as wintering grounds by the Mi’kmaq, a non-agricultural First Nations people (Leonard 1996). Starting from the late 1700s, the area has been logged and most of the moderately to well-drained sites were cleared for farmsteads. Once cleared, fields were continuously cultivated until 1969 when the area was acquired for the national park. Many of the abandoned fields were still open during the time of research, after 50 years of abandonment, due to a slow rate of natural re-forestation.
Thirty abandoned fields were examined in KNP over a mosaic of sites with varied drainage conditions: coastal plain, watershed bogs, swales, sand ridges, and well to moderately drained river banks. During land clearing, mostly well- to moderately drained sites were selected for cultivation, but due to the high complexity of the landscape each field included wetter and dryer spots, with wetter sites requiring drainage into dugout ditches prior to cultivation.
A single field was used as the sampling unit, with the size of individual fields varying from 3 to 20 ha. For purposes of this study, we assumed that the long-term ploughing averaged out the initial variations in the species composition within the forest patch that has been cleared for farming. Ploughing causes not only vertical mixing and incorporation of charcoal into the ground mass of the plough layer, but also charcoal fragmentation and long-distance lateral transportation of fragments as suggested by the studies of potsherds in archaeological sites. A single plowing action moves a particle only for 0.5-2.0 meters (Van Oost et al. 2000), but repetition of such action over many years leads to a significant re-distribution of charcoal particles. The distance and extent of such movement can be estimated by research on the distribution of ceramic fragments in archaeological sites: long-term field experiments and surficial archaeological surveys show that the dispersal may reach ~100 meters after 200 years of plowing (Yorston et al. 1990).
According to our observations on fields that were recently cleared from forests, piles of logging slash could be distributed approximately 20 m from one another. We assumed that if several logging slash burn sites were located within a field during land clearance, they would provide a sufficient mixing of combustion products throughout the field.
Local soils are developed on late Pleistocene-early Holocene deposits. Gray Luvisol and Luvic Gleysol are formed in minor areas of marine clay deposits, and Humo-Ferric Podzols are common on sand deposits of various genesis and thickness (Wang and Rees 1983). In most of our study sites, loamy glacial deposits are overlain by a veneer of fluvioglacial or aeolian sand from 0.3 m to over 2 meters deep. As a result, plough layers are predominantly sand, which allowed for extraction of charcoal by dry sieving in field.
Two sampling approaches were used:
– In each site, charcoal was collected from a trench wall (7 x 1.5 m) so that the whole perimeter of the trench was represented in the charcoal assemblage.
– Charcoal was sampled from test pits in catenas running across each field to ensure that data was representative for each field. The test pits were of a uniform size, 50 x 50 x 25 cm, and were spaced at 20 m intervals. The soil mass from the plough layer within the 50 cm2 squares was sifted through a sieve with 2 mm openings, and all charcoal fragments larger than 2 mm were collected for additional identification. Number of the sampled charcoal fragments and their size varied substantially within the catenas: in some test pits, we were able to collect only several charcoal fragments less than 4 mm in diameter, whereas in others, charcoal was abundant and up to 1 cm in diameter. It was realized later that those test pits rich in charcoal were located in close proximity to logging slash fireplaces. We were not able to process all the charcoal sampled within the catenas, as the amount of material by far exceeded the available time and budget. At this stage, identifications were limited to 5 test pits within each field.
The number of charcoal fragments identified for each field site had to meet statistical requirements for representativeness, but was constrained by the fact that charcoal identification is a time consuming procedure: it was not feasible to do more than 50 identifications per sample unit due to time and budget constraints. We identified a minimum of 30 fragments for sites with a low species composition (1-3 taxa), and 50 fragments for sites with a higher species diversity. A total of approximately 1300 charcoal fragments were identified.
Charcoal fragments were broken manually and analyzed in reflected light. For botanical identification, existing manuals (e.g. Barefoot and Jenkins 1982) were consulted along with reference materials collected from the park. Charcoal was identified with confidence to the genus level, in some cases to a group of species within a genus. Other taxa were narrowed to the species level based on the species geographical ranges.

PROCEEDINGS OF THE FOURTH INTERNATIONAL MEETING OF ANTHRACOLOGY
206
EXPECTED ERRORS
Besides the charcoal associated with land clearance, it was possible that plough layers could contain both older and younger charcoal from other events. The former could originate from pre-agricultural forest fires and the latter could be charcoal from furnaces and woodstoves, sometimes added to the fields by local farmers as fertilizer. Although both components could be represented in the charcoal pool, we assumed that the charcoal associated with the land clearance would prevail due to the immense amount of logging slash produced during land clearing.
Charcoal fragments larger than 2 mm are rarely preserved in the surficial (0-25 cm) layer of soil following a forest fire for a longer duration than one, or at most two, tree generations. Shortly after a fire, charcoal fragments become buried deeper in the soil profile by pedoturbation (e.g., uprootings and animal activity) (Ponomarenko 1999), while the rest become progressively reduced in size and disaggregated by ploughing. Therefore, we assumed there would be a negligibly small proportion of old charcoal fragments found in the plough layers that exceeded 2 mm in diameter. However, if a fire had occurred shortly prior to the land clearance the plough layer could be contaminated by pre-clearance charcoal. Some morphological criteria were used to eliminate charcoal from furnaces from the analyzed assemblages: we removed the fragments that had a high percentage of soluble ash within charcoal fragments and no clay coating on the surface of the charcoal fragments, as charcoal within the plough layers is becomes progressively coated/impregnated by clay (Ponomarenko and Anderson 2000).
Archival records of historical forest composition
Archival documentation on early forest types has been analyzed for approximately the same area (Crossland 2006). Archival information consisted of both witness trees and early forest descriptions located at the Provincial Archives of New Brunswick. In the early 1800s, land surveyors divided the landscape into various-sized parcels, marking trees on the bounds of each plot. Location and species of such “witness trees” were recorded on surveyor plan drawings of land grants to private citizens from years 1805 to 1909. Pre-settlement forest descriptions were also reconstructed using 46 survey sketches, to produce 306 records of forest descriptions.
The analysis extended to lands beyond KNP in order to ensure an adequate witness tree sample size The study area was approximately 10 times greater than the area of Kouchibouguac National Park (2,420 km2 and 239 km2
correspondingly), extending both along the coast and into inland (Fig. 1). Witness trees were widely distributed throughout the study area, but were more concentrated on riparian zones. Witness tree densities varied from 0.3 to 2.6 trees/km2, averaging 1.1 tree/km2. The resulting witness tree database consisted of 2537 trees.
A total of ~2500 witness trees were used for the subsequent analysis and its comparison with results of botanical identification of ~1300 charcoal fragments. Information derived from both methods characterizes the Acadian forest ca 1800, just prior to heavy anthropogenic modification.
RESULTS AND THEIR INTERPRETATION
Age of identified charcoal fragments
Since most of the charcoal fragments originated from small branches and twigs, we assumed that the absolute age of the radiocarbon dated charcoal did not exceed several decades. However, some of charcoal fragments of shade-tolerant species, such as Fagus and Tsuga could be over 80 years old as estimated by the number of annual rings in the fragments that had an initial cylindrical shape preserved.
According to historical descriptions, the trees logged at the time of the land clearing greatly varied in size and, hence, in age (Crossland 2006). The age of the cleared forest patches also varied from young stands to old-growth forests. Ages of shade-tolerant species, such as Eastern hemlock and red spruce, can achieve 400 years, whereas, long-lived, light-demanding species, such as white pine and red oak can attain 250 years (Burns and Honkala 1990). Species composition at the time of land clearance reflects the conditions that existed at the time of establishing the forest communities, in some loci up to 400 years ago.
Direct radiocarbon dating of single charcoal fragments from the plough layer in three sites yielded 14C ages 264 ± 55 BP (BGS2525), 230 ± 40 BP (CAMS 86042), and 195 ± 35 BP (Beta -190069), the dates being consistent with the time of agricultural colonization of the area.
Taxa reconstructed
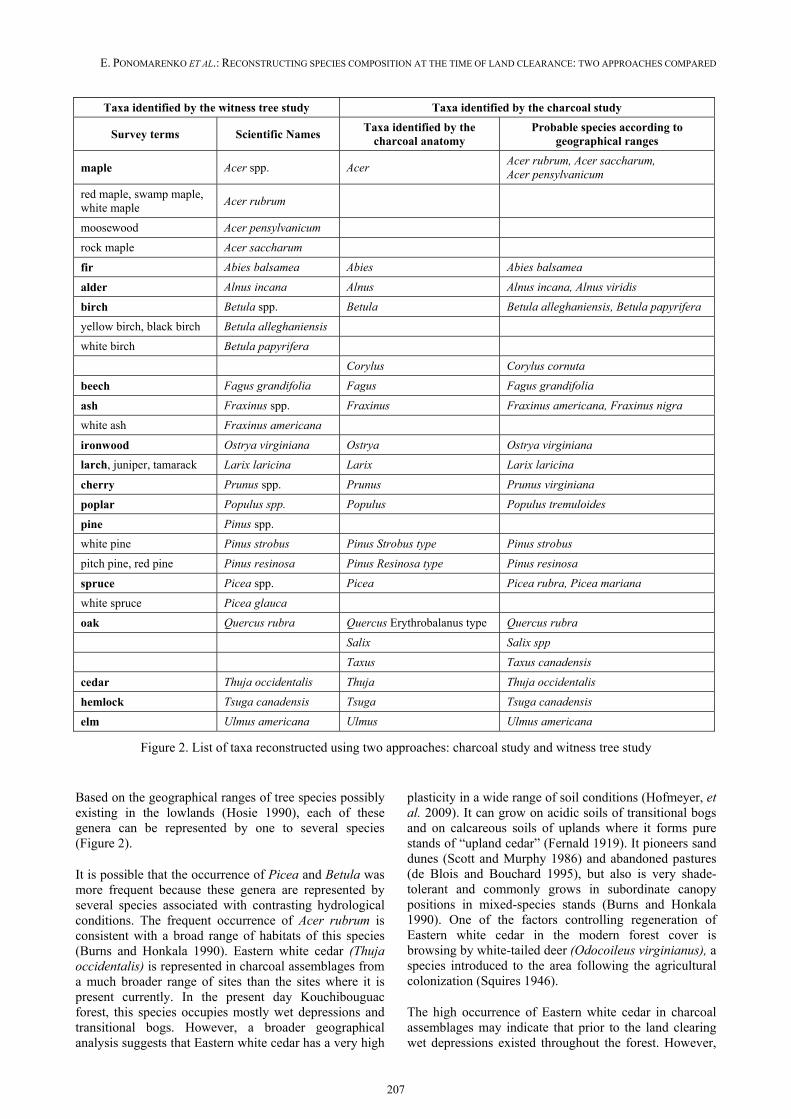
Charcoal analysis allowed for reconstruction of 20 taxa (Figure 2). The number of taxa recovered per site varied from 2 in drained cedar bogs to 10 in a well-drained riparian zone, revealing a high complexity of forest cover at the time of land clearance. More than 6 taxa were recorded in charcoal assemblages from ~50% of sites.
The results are presented in two forms, as species frequencies and species abundance.
Mean tree species frequencies were calculated as a percent of sites where a species occurred. Eight genera dominated the area at the time of land clearance, occurring in more than 50% of the sites (in descending order): Picea (90%) > Thuja, Betula (~80%) > Acer (65%) > Tsuga, Pinus (Strobus type) (~50%), Abies, Fagus (~40%) (Figure 3). Less common were such taxa as Populus, Quercus (Erythrobalanus type), Pinus (Resinosa type), Larix, Ulmus, Alnus, Fraxinus, Taxus, Ostrya, and Prunus.
E. PONOMARENKO ET AL.: RECONSTRUCTING SPECIES COMPOSITION AT THE TIME OF LAND CLEARANCE: TWO APPROACHES COMPARED
207
Taxa identified by the witness tree study Taxa identified by the charcoal study
Survey terms Scientific Names Taxa identified by the
charcoal anatomy Probable species according to
geographical ranges
maple Acer spp. Acer Acer rubrum, Acer saccharum, Acer pensylvanicum
red maple, swamp maple, white maple
Acer rubrum
moosewood Acer pensylvanicum
rock maple Acer saccharum
fir Abies balsamea Abies Abies balsamea
alder Alnus incana Alnus Alnus incana, Alnus viridis
birch Betula spp. Betula Betula alleghaniensis, Betula papyrifera
yellow birch, black birch Betula alleghaniensis
white birch Betula papyrifera
Corylus Corylus cornuta
beech Fagus grandifolia Fagus Fagus grandifolia
ash Fraxinus spp. Fraxinus Fraxinus americana, Fraxinus nigra
white ash Fraxinus americana
ironwood Ostrya virginiana Ostrya Ostrya virginiana
larch, juniper, tamarack Larix laricina Larix Larix laricina
cherry Prunus spp. Prunus Prunus virginiana
poplar Populus spp. Populus Populus tremuloides
pine Pinus spp.
white pine Pinus strobus Pinus Strobus type Pinus strobus
pitch pine, red pine Pinus resinosa Pinus Resinosa type Pinus resinosa
spruce Picea spp. Picea Picea rubra, Picea mariana
white spruce Picea glauca
oak Quercus rubra Quercus Erythrobalanus type Quercus rubra
Salix Salix spp
Taxus Taxus canadensis
cedar Thuja occidentalis Thuja Thuja occidentalis
hemlock Tsuga canadensis Tsuga Tsuga canadensis
elm Ulmus americana Ulmus Ulmus americana
Figure 2. List of taxa reconstructed using two approaches: charcoal study and witness tree study
Based on the geographical ranges of tree species possibly existing in the lowlands (Hosie 1990), each of these genera can be represented by one to several species (Figure 2).
It is possible that the occurrence of Picea and Betula was more frequent because these genera are represented by several species associated with contrasting hydrological conditions. The frequent occurrence of Acer rubrum is consistent with a broad range of habitats of this species (Burns and Honkala 1990). Eastern white cedar (Thuja occidentalis) is represented in charcoal assemblages from a much broader range of sites than the sites where it is present currently. In the present day Kouchibouguac forest, this species occupies mostly wet depressions and transitional bogs. However, a broader geographical analysis suggests that Eastern white cedar has a very high
plasticity in a wide range of soil conditions (Hofmeyer, et al. 2009). It can grow on acidic soils of transitional bogs and on calcareous soils of uplands where it forms pure stands of “upland cedar” (Fernald 1919). It pioneers sand dunes (Scott and Murphy 1986) and abandoned pastures (de Blois and Bouchard 1995), but also is very shade-tolerant and commonly grows in subordinate canopy positions in mixed-species stands (Burns and Honkala 1990). One of the factors controlling regeneration of Eastern white cedar in the modern forest cover is browsing by white-tailed deer (Odocoileus virginianus), a species introduced to the area following the agricultural colonization (Squires 1946).
The high occurrence of Eastern white cedar in charcoal assemblages may indicate that prior to the land clearing wet depressions existed throughout the forest. However,

PROCEEDINGS OF THE FOURTH INTERNATIONAL MEETING OF ANTHRACOLOGY
208
Figure 3. Frequency (in gray) and abundance (in black) of various species in charcoal assemblages
it may also indicate that prior to the land clearance Eastern white cedar occupied a much wider range of edaphotops than currently.
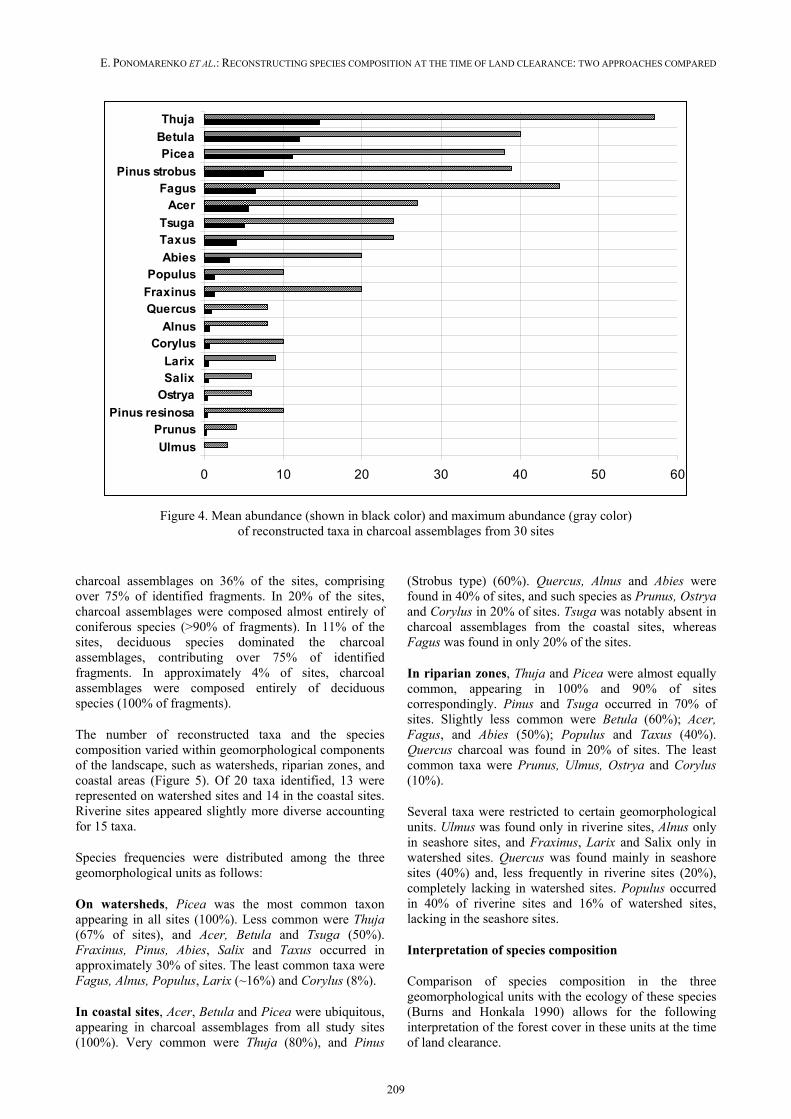
Species abundance has been estimated as a percentage of charcoal fragments belonging to species in assembla-ges from a particular site, and also as a mean percentage calculated for all study sites. To estimate the proportions of various arboreal species in each site, total percentage of identifiable components in each assemblage was assu-med to be 100%; non-identifiable components excluded.
It must be noted that in the sites with the highest species diversity recorded in charcoal assemblages, identification of even 50 fragments may be insufficient for the appropriate characterization of the percentage of each species. Moreover, our estimates of species abundance, (that were dependant on the estimates of species frequencies), are in turn reliant on less well known parameters, such as ashing coefficients (ash:charcoal ratio) of each species during initial combustion and differential preservation of charcoal from various species (different rates of resistance to mechanical weathering and grinding in the soil). Furthermore, proportions of charcoal represented between arboreal species may reflect varying proportions of crown wood versus trunk wood, as trunk wood does not greatly contribute to the charcoal assemblages associated with land clearance according to the descriptions of the land clearing process (Johnston 1851).
The mean percentage of fragments belonging to each species did not exceed 19%. This can be interpreted as
the absence of strongly dominant species over the studied range. Mean percentage of nine dominant taxa in decreasing order is as follows (Figure 4): Thuja (18%) > Betula, Picea (14%) > Pinus (Strobus type) (13%) > Tsuga, Acer, Fagus (7%) > Taxus (6%)> > Abies (4%).
In the sites where the maximum abundance of a species was recorded, the percentage of the species in charcoal assemblages was much higher than the mean abundance, reaching 70% for Thuja, 53% for Pinus (Strobus type), ~44% for Betula and Fagus, 38% for Picea, and ~32% for Tsuga and Acer. Other species such as Salix, Ulmus, Alnus, Prunus, Ostrya did not have high abundance on any studied sites.
It appears that the most common species (9 species recorded in most of the sites) were also the most abundant ones, whereas the uncommon species contributed a fairly low number of charcoal fragments in sites where they occurred (Figure 3). However, there are some inversions: e.g., Acer being a highly common species produced a relatively low proportion of charcoal fragments in the sites where it occurred. Contrarily, Pinus (Strobus type), being a less frequent species, contributed a relatively large proportion of fragments to the charcoal assemblages.
Spatial distribution and species associations
Tree species associations were highly variable over the landscape. In many sites (56%), the charcoal assemblages reflected a mixed forest composed of both deciduous and coniferous species. Coniferous species dominated
0 10 20 30 40 50 60 70 80 90 100
UlmusPinus resinosa
OstryaPrunus
LarixSalix
CorylusQuercusFraxinus
AlnusPopulus
TaxusFagusAbies
Pinus strobusTsuga
AcerBetulaThujaPicea
E. PONOMARENKO ET AL.: RECONSTRUCTING SPECIES COMPOSITION AT THE TIME OF LAND CLEARANCE: TWO APPROACHES COMPARED
209
Figure 4. Mean abundance (shown in black color) and maximum abundance (gray color) of reconstructed taxa in charcoal assemblages from 30 sites
charcoal assemblages on 36% of the sites, comprising over 75% of identified fragments. In 20% of the sites, charcoal assemblages were composed almost entirely of coniferous species (>90% of fragments). In 11% of the sites, deciduous species dominated the charcoal assemblages, contributing over 75% of identified fragments. In approximately 4% of sites, charcoal assemblages were composed entirely of deciduous species (100% of fragments).
The number of reconstructed taxa and the species composition varied within geomorphological components of the landscape, such as watersheds, riparian zones, and coastal areas (Figure 5). Of 20 taxa identified, 13 were represented on watershed sites and 14 in the coastal sites. Riverine sites appeared slightly more diverse accounting for 15 taxa.
Species frequencies were distributed among the three geomorphological units as follows:
On watersheds, Picea was the most common taxon appearing in all sites (100%). Less common were Thuja (67% of sites), and Acer, Betula and Tsuga (50%). Fraxinus, Pinus, Abies, Salix and Taxus occurred in approximately 30% of sites. The least common taxa were Fagus, Alnus, Populus, Larix (~16%) and Corylus (8%).
In coastal sites, Acer, Betula and Picea were ubiquitous, appearing in charcoal assemblages from all study sites (100%). Very common were Thuja (80%), and Pinus
(Strobus type) (60%). Quercus, Alnus and Abies were found in 40% of sites, and such species as Prunus, Ostrya and Corylus in 20% of sites. Tsuga was notably absent in charcoal assemblages from the coastal sites, whereas Fagus was found in only 20% of the sites.
In riparian zones, Thuja and Picea were almost equally common, appearing in 100% and 90% of sites correspondingly. Pinus and Tsuga occurred in 70% of sites. Slightly less common were Betula (60%); Acer, Fagus, and Abies (50%); Populus and Taxus (40%). Quercus charcoal was found in 20% of sites. The least common taxa were Prunus, Ulmus, Ostrya and Corylus (10%).
Several taxa were restricted to certain geomorphological units. Ulmus was found only in riverine sites, Alnus only in seashore sites, and Fraxinus, Larix and Salix only in watershed sites. Quercus was found mainly in seashore sites (40%) and, less frequently in riverine sites (20%), completely lacking in watershed sites. Populus occurred in 40% of riverine sites and 16% of watershed sites, lacking in the seashore sites.
Interpretation of species composition
Comparison of species composition in the three geomorphological units with the ecology of these species (Burns and Honkala 1990) allows for the following interpretation of the forest cover in these units at the time of land clearance.
0 10 20 30 40 50 60
Ulmus
PrunusPinus resinosa
OstryaSalixLarix
CorylusAlnus
QuercusFraxinus
PopulusAbies
TaxusTsuga
AcerFagus
Pinus strobus
PiceaBetula
Thuja

PROCEEDINGS OF THE FOURTH INTERNATIONAL MEETING OF ANTHRACOLOGY
210
Figure 5. Tree species frequencies of tree species in charcoal assemblages from three geomorphological zones of Kouchibouguac National Park
On watersheds, the canopy layer was commonly composed of spruce (most probably, red spruce in well-drained sites and black spruce in sites of poorer drainage) with a proportion of Eastern hemlock, balsam fir, and deciduous species: red maple and birch (presumably mainly yellow birch, given its association with late-
successional forest complexes). White pine was part of the canopy layer in more localized stands, and American beech occurred only in well-drained sites. Canada yew was a common understory shrub in moderately to well-drained sites, and beaked hazel grew in more open, light-canopy forests on well-drained soils.

E. PONOMARENKO ET AL.: RECONSTRUCTING SPECIES COMPOSITION AT THE TIME OF LAND CLEARANCE: TWO APPROACHES COMPARED
211
The currently drained bogs that existed at the time of land clearance were occupied by tamarack and Eastern white cedar, whereas margins of bogs, small creeks and beaver ponds were a suitable habitat for white pine, quaking aspen and gray alder. Wet depressions provided a niche for black ash and Eastern white cedar. Finally, various willow species could inhabit any of the above-mentioned wetlands.
Given poor drainage conditions on watersheds, the frequent occurrence of Fraxinus could be at least partially due to presence of black ash (Fraxinus nigra). However, its occurrence in watershed sites is higher than the proportions of any other wetland species, therefore it is possible that both black and white ash (Fraxinus americana) were present in the landscape at the time of the land clearance.
In coastal sites with sufficient drainage, the canopy layer could be composed of white pine, red oak, red maple, American beech, balsam fir, and species of birch and spruce. Some of these forests must have been early successional (or recently disturbed) to allow for reproduction of such understory species as pin cherry (Prunus pensylvanica), beaked hazel (Corylus cornuta) and possibly green alder (Alnus viridis). Eastern white cedar and possibly gray alder (Alnus incana) inhabited more poorly drained coastal sites.
In riparian sites, canopy layer could be composed of white pine, Eastern hemlock, red oak, red maple, American beech, balsam fir, and species of spruce (most probably red spruce (Picea rubens) and birch (both yellow and white birch). Some riparian forests had to be early successional (or recently disturbed) to allow for reproduction of such species as quaking aspen (Populus tremuloides), pin cherry, American elm, and beaked hazel. Although the riparian zone is generally well-drained, there were numerous loci of poorer drainage inhabited by Eastern white cedar.
Frequency of natural disturbances
The composition and spatial distribution of reconstructed taxa allow for some interpretation of natural disturbances at the time of land clearance.
The species composition at the time of land clearance was dominated by mid- to late successional, shade-tolerant species. Reproduction of such species is governed by small-scale disturbances or gap dynamics (Pickett and White 1985). Supporting this observation was the high frequency and high abundance of Eastern white cedar in most of the study sites. This species is usually absent in recently disturbed sites.
In a few sites, light-demanding species were a part of the forest cover, appearing either in the canopy (white pine, red oak, quaking aspen, American elm, and species of ash) or in understorey – e.g., pin cherry, species of alder, and beaked hazel.
Reproduction of light-demanding, early successional species requires periodic disturbances of a larger scale, such as fires, windstorms, flooding, and insect infestations. Therefore, the presence of light-demanding species in the charcoal assemblages associated with the time of land clearance indicates the occurrence of such disturbances, and the proportion of sites where such species have been found reflects frequency of the disturbances. Such disturbances had to closely precede the establishment of the light-demanding species in the site. Hence, the maximum longevity of the species provides us with an estimate of the maximum time since the last disturbances in the site.
Long-lived, light-demanding canopy species, such as red pine, white pine and red oak can achieve age of 250-300 years in the area, therefore their presence in the landscape/site may indicate the occurrence of some disturbances (mainly fire) as much as ~250-300 years prior to the land clearance event. Red pine, a species directly associated with fire for reproduction, was found in only one site. White pine was recorded in 45% of study sites, while both pine and oak species were recorded in 20% of the sites. Presumably, these numbers correlate with the frequency of large-scale disturbances during 250-300 years preceding the land clearance.
It was assumed that the sites lacking long-lived, light-demanding canopy species were not affected by disturbances within the maximum lifetime of these species; whereas those sites that featured these species were affected by disturbances within the species longevity interval. Therefore, it was possible to calculate the disturbance return interval as a division of the indicative species longevity on the fraction of sites where those species were present.
The maximal disturbance return interval calculated using the frequency of red oak – the species that requires canopy replacing disturbances, such as fire and wind, is 1250 years (=250 / 0.2, where 250 is species longevity and 0.2 is the fraction of sites with this species).
Red oak was always found in association with white pine, but white pine occupied a wider range of sites in association with a wide range of species. It may indicate that reproduction of white pine in the area is governed by a wider spectrum of factors than the reproduction of red oak. Both red oak and white pine (where they dominate forest stands) are considered to be largely dependent on fires for their reproduction (Burns and Honkala 1990). At the same time, our observations on white pine reproduction in the area indicate that white pine can also be established on margins of bogs, creeks and beaver ponds, replacing deciduous vegetation that is selectively consumed by beaver. The width of the “feeding zone” cleared by beaver foraging can reach 60 meters (Rossell et al. 2005, Johnston and Naiman 1990). Therefore, calculating the canopy replacing disturbance return interval based on white pine frequency would probably underestimate the real disturbance return interval and can be used as a minimal assessment. Thus, minimal estimate

PROCEEDINGS OF THE FOURTH INTERNATIONAL MEETING OF ANTHRACOLOGY
212
is 555 years (=250 / 0.45, where 250 is species longevity and 0.45 is a fraction of sites with this species). Combining these two estimates, one can conclude that during the period between 450 and 200 years ago the canopy replacing disturbance return interval for the studied area was between approximately 1250 and 550 years.
Such estimates were calculated on the assumption that all sites are equally prone to disturbances.
However, a proportion of the “disturbance-dependant species” varied among geomorphological units. Red oak and white pine were much more common in riparian and coastal sites than in watersheds, with red oak completely absent on watersheds. Red oak was found in ~70% of coastal sites, but only 30 % of riverine sites. Assuming that all sites within the same geomorphological unit are equally prone to disturbances, the minimal and maximal estimates will be 555/833 years in riparian zone, 417/625 years in coastal sites, and 1000 to over 3012 years on watersheds.
Short-lived species, such as quaking aspen, American elm, gray alder, willow species, and black ash require the appearance of openings in the canopy at least once every 50-80 years. All these species are associated with wetlands that are very numerous in the area. Periodic (decadal to centennial) pulsation of wetland margins is typical for the area and can be caused by a number of factors, such as changes in atmospheric precipitation, changes in ground water regimes, and beaver activity. Such pulsation can provide new habitats for reproduction of the short-lived wetland species with a periodicity sufficient for their maintenance in the landscape.
Comparison with historical records
Results of the charcoal analysis have been compared with the analysis of archival descriptions of the forest composition (Crossland 2006). Results of botanical identification of ~1300 charcoal fragments from 30 sites were used for comparison with results of the analysis of ~2500 witness trees. The archival data represented a much higher sample of trees, as each of the analyzed charcoal fragments did not represent a single tree. Also, the archival data represented a much larger study area, as the majority of witness trees were located outside of KNP.
Each research method detected a high diversity of species in 19th century forests (Figure 2), including tree species that are generally not common on the landscape, such as Ulmus americana and Ostrya virginiana.
Archival analyses revealed presence of 20 taxa in pre-clearance forests: 18 of them were assigned to certain species, whereas 2 could be determined only to genus level.
Charcoal study captured 20 taxa: it recorded all tree genera that were represented in the witness tree record,
and the method allowed some genera to be consistently identified to the species level, e.g. Pinus strobus was separated from P. resinosa that was found in only one study site. Witness tree results also confirmed minor amounts of Pinus resinosa (a frequency of 0.6% in the landscape). According to both methods, Pinus banksiana, a fire-dependant species that is common and widespread in the modern landscape, was absent at the time of land clearance. Species of Picea, Betula, and Acer were not distinguished through charcoal study, nor were they consistently discerned in the witness tree record, so further quantitative and comparative analyses were confined to the genus level.
Both methods revealed the presence of shrub or understorey species, such as Alnus and Prunus. Charcoal analysis identified three additional shrub genera that were not detected in the witness tree record: Taxus canadensis, Corylus cornuta, and Salix spp. An understorey tree species discerned in the witness tree record, but not through charcoal analysis was Acer pensylvanicum.
Both methods indicated that historic forests in the area were mixed and comprised of mid- to late successional species (Picea sp, Tsuga canadensis, Betula sp, Acer sp, Fagus grandifolia, Abies balsamea and Pinus strobus) in the majority of sites. Early successional and shade-intolerant species, such as Populus sp., Pinus resinosa and Quercus rubra were present at very low levels according to both methods.
Occurrence of taxa reconstructed by the analysis of charcoal assemblages have been compared with the occurrence of taxa calculated using witness trees records. For this comparison we excluded shrub species (Salix, Corylus and Taxus) from the analysis since they were not generally used as witness trees and compared only 16 tree genera common for both data sets. To make the two sets of data comparable, we calculated the occurrence of each species in charcoal assemblages as n/(N1 + N2 +…+ Nx), where n is the number of sites where a species occurred, and N is the total number of species recorded at each study site. In total there were 173 tree species occurrences in charcoal assemblages. These data were compared with a total 306 occurrence records of witness trees, also expressed as a percent of total tree species occurrences.
The occurrences of species calculated by these two methods were remarkably similar (Figure 6), given the differences in sampling distribution and sample sizes. Both methods revealed the same eight most frequent taxa, with Picea being the most frequent taxon. Occurrences of Betula, Acer, Fagus, Pinus (Strobus type) and Fraxinus were very similar in either method. Smaller trees and shrubby species, such as Thuja occidentalis, Ostrya virginiana, Prunus pensylvanica, Populus spp., and Alnus spp. were represented in the witness tree data at much lower occurrences than indicated by charcoal study. Results differed, however, among coniferous species such as Tsuga canadensis, Abies balsamea and Picea sp., which had much higher occurrences in witness tree

E. PONOMARENKO ET AL.: RECONSTRUCTING SPECIES COMPOSITION AT THE TIME OF LAND CLEARANCE: TWO APPROACHES COMPARED
213
Figure 6. Comparison of tree species occurrences as estimated by the studies of charcoal assemblages and witness tree records
results in relation to charcoal assemblages. Under-representation of smaller understorey tree species and over-representation of coniferous canopy species in archival records may be explained by a bias towards selecting larger trees during surveying.
Some of the discrepancies between the two methods may also reflect the differences in sample site locations: e.g., Quercus is over-represented and Tsuga underrepresented in charcoal assemblages in relation to witness tree data. The former species is most frequent and the latter is least frequent in coastal sites where a smaller portion of witness trees were present to compare to the charcoal research sites. Finally, some of the discrepancies may reflect differences in preservation of charcoal of different species, which requires additional study.
CONCLUSION
Comparison between the two methods confirmed applicability of the analysis of charcoal assemblages from plough layers for the reconstruction of tree species composition at the time of land clearance. This presents an opportunity of applying the proposed anthracological approach for reconstructing historical forest composition in agricultural areas that were previously forested.
References
BEACH, H. (Ed.) 1988. The Resources of Kouchibougu-ac National Park: Resource Description and Analysis. Kouchibouguac National Park, Envir. Canada.
BURNS, R.M. and HONKALA, B.H. tech. coords. 1990. Silvics of North America: 1. Conifers; 2. Hardwoods. Agriculture Handbook 654. U.S. Department of Agriculture, Forest Service, Washington, DC. 2, 877 pp.
CROSSLAND, D. 2006. Defining a forest reference condition for Kouchibouguac National Park and adjacent landscape in Eastern New Brunswick using four reconstructive approaches. Unpublished MScF thesis, University of New Brunswick.
De BLOIS, S. and BOUCHARD, A. 1995. Dynamics of Thuja occidentalis in an agricultural landscape of southern Quebec. J. Veg. Sci. 6(4), 531-542.
DESLOGES, C. 1980. Les ressources naturelles du Parc national Kouchibouguac. Parcs Canada, région de l’Atlantique. Halifax, Nova Scotia. 137 pp.
DNRE. 1996. An ecological land classification system of New Brunswick. Ecosystem Classification Working Group, Department of Natural Resources and Energy, Fredericton, NB.
DRYADE (Le Groupe) 1979. Kouchibouguac National Park forest vegetation. Parks Canada. Unpublished report. 186 pp.
DUCHESNE, L.C. and HAWKES, B.C. 2000. Fire in northern ecosystems. In Brown, J.K. and Smith, J.K. (Eds.) Wildland fire in ecosystems: Effects of fire on flora. USDA Forest Service Gen. Tech. Rep. RMRS-GTR-42, 2: 35-51.
FERNALD, M.L. 1919. Lithological factors limiting the ranges of Pinus banksiana andThuja occidentalis. Rhodora 21 41-67.
Charcoal Assemblages
0 5 10 15 20 25 30
Ulmus
Ostrya
Larix
Quercus
Fraxinus
Prunus
Alnus
Populus
Fagus
Abies
Tsuga
Pinus strobus
Acer
Betula
Thuja
Picea
Witness Trees
0 5 10 15 20 25 30
Ulmus
Ostrya
Larix
Quercus
Fraxinus
Prunus
Alnus
Populus
Fagus
Abies
Tsuga
Pinus strobus
Acer
Betula
Thuja
Picea
c

PROCEEDINGS OF THE FOURTH INTERNATIONAL MEETING OF ANTHRACOLOGY
214
HOFMEYER, P.V. 2008. Ecology and silviculture of northern white-cedar (Thuja occidentalis L.) in Maine. Ph.D. dissertation. Univ. of Maine, Orono, ME. 160 pp.
HOSIE, R.C. 1990. Native trees of Canada. Fitzhenry and Whiteside Ltd., Ont. 380 pp.
JOHNSTON, J.F.W. 1851. Notes on North America. Vol. I and II., C.C. Little and J. Brown, Boston. 512 pp.
LEONARD, K.J. 1996. Mi’kmaq culture during the Late Woodland and Early Historic periods. Thesis, U. Toronto, Dept. of Anthropology, 329 pp.
LOUCKS, O.L. 1962. A Forest Classification for the Maritime Provinces. Proceedings of the Nova Scotian Institute of Science. Vol. 25, Part 2.
PICKETT, S.T.A. and WHITE, P.S. 1985. The ecology of natural disturbance and patch dynamics. Academic Press, Inc. Toronto, ON. 472 pp.
PONOMARENKO E.V. and ANDERSON, D.W. 2000. The importance of charred organic matter in Black Chernozemic soils. Can.J.Soil Sci. 81, 285-297.
PONOMARENKO E.V. 1999. Methodological approaches to the analysis of the succession process in soil cover. In Smirnova, O.V., Shaposhnikov, E.S. (eds.). Forest succession in protected areas of Russia and problems of biodiversity conservation. Russian
Botanical Society. St-Petersburg. p. 34-57. (In Russian).
ROSSELL F., BOZSERT, O., COLLEN, P. and PARKER, H. 2005. Ecological impact of beavers Castor fiber and Castor canadensis and their ability to modify ecosystems. Mammal Review, 35, 3-4, 248-276.
ROWE, J.S. 1972. Forest Regions of Canada. Publication No. 1300. Ottawa: Canadian Forestry Service, Department of the Environment.
SCOTT, M.L., and MURPHY, P.G. 1986. Regeneration patterns of northern white-cedar, an old-growth forest dominant. Am. Midl. Nat. 117, 10-16.
SQUIRES, W.A. 1946. Changes in mammal populations in New Brunswick. Acadian Naturalist, 2:26-44.
Van OOST, K., GOVERS, G., Van MUYSEN, W. and QUINE, T.A. 2000. Modeling translocation and dispersion of soil constituents by tillage on sloping land. Soil Society of America Journal, 64, 1733-1739.
WANG, C. and REES, H.W. 1983. Soils of the Rogersville-Richibucto Region of New Brunswick. Agriculture Canada, 239 pp.
YORSTON, R.M., GAFFRNEY, V.L. and REYNOLDS, P.G. 1990. Simulation of artifact movement due to cultivation. Journal of Archaeological Science, 17, 67-83.




















