ANTIGENIC ANALYSIS OF GAMONTS OF HEPATOZOON CANIS PURIFIED FROM LEUKOCYTES
Upload
hms-harvardCategory
view
0download
0
THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1992 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 267, No. 8, Issue of March 15, pp. 5408-5415.1992 Printed in U.S.A.
Polyoma Virus Middle T Antigen-pp6OC'"" Complex Associates with Purified Phosphatidylinositol 3-Kinase in Vitro*
(Received for publication, September 19, 1991)
Kurt R. Auger$, Christopher L. CarpenterQ1, Steven E. ShoelsonII, Helen Piwnica-Worms, and Lewis C. Cantley From the Department of Physiology, Tufts University School of Medicine, Boston, Massachusetts 021 11, the SHematology- Oneolom Unit. Massachusetts General Hoswital. Boston. Massachusetts 02214, and the 11 Joslin Diabetes Center, Haruard Med ic i iSch i , Boston, Massachusetts 02i15 '
Reconstitution of the polyoma virus middle T antigen (mT)-pp60" complex and phosphatidylinositol 3-ki- nase (PtdIns 3-kinase) has been accomplished in vitro with immunopurified baculovirus-expressed mT- pp60""" and PtdIns 3-kinase purified from rat liver. Both the 110- and 86-kDa subunits of the PtdIns 3- kinase associated with the mT-pp60c-a" complex. The association of PtdIns 3-kinase with the mT-pp60""" complex was dependent on the protein-tyrosine kinase activity of pp6OC-"" as a kinase-inactive mutant (pp602g5c-ac) still complexed with mT, but the mT-
nase. The mT-pp60c-"" complex phosphorylated both subunits of PtdIns 3-kinase on tyrosine residues. The immunopurified mT-pp60C"" complex also associated with PtdIns 3-kinase activity from whole cell lysates, and this association was dependent upon the protein- tyrosine kinase activity of p~60"-~". Comparison of "S- labeled proteins from whole cell lysates which associ- ated with immunopurified mT-pp6O"" and mT-
major peptides dependent on protein-tyrosine kinase activity for association with the complex. In addition, a synthetic phosphopeptide (13-mer) containing se- quences conserved between the major tyrosine phos- phorylation site of murine polyoma virus mT, hamster polyoma virus mT, and the insulin receptor substrate (IRS-1) specifically blocked the association of the 85- and 110-kDa polypeptides with the mT-pp60c-am com- plex. The ability to block the association was depend- ent on the tyrosine phosphorylation of the peptide. Association of PtdIns 3-kinase activity was blocked concurrently. This is the first demonstration that the 110-kDa subunit of PtdIns 3-kinase can associate with mT-pp60""". This association in vitro is a step toward understanding protein-protein interactions important in the signal transduction pathway of oncogenic pro- teins.
PPf302g5c-s" complex was unable to bind PtdIns 3-ki-
PP60ae5c-a" revealed proteins of 110 and 85 kDa as the
* This work was supported in part by National Institutes of Health Grants GM 36624 and GM 41890 ( t o L. C. C. ) and CA 50767 ( t o H. P-W.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Supported in part by Training Grant F32 HL07437 from the National Institutes of Health. To whom correspondence should be addressed Dept. of Physiology, Tufts University School of Medicine, 136 Harrison Ave., Boston, MA 02111. Tel.: 617-956-6744; Fax: 617- 956-0445.
1 Supported by an American Heart Association-Parke Davis cli- nician scientist award.
Transformation by mouse polyoma virus requires middle T antigen, and this protein both regulates and is a substrate for protein kinases. A small fraction of the mT' molecules in fibroblasts is associated with the cellular proto-oncogenes pp6OC-"", pp62c-ye8, and p 5 P (1-4). In the case of pp6OC"", this association results in the activation of protein-tyrosine kinase activity (5-7). Transformation of fibroblasts by mT appears to require both activation of protein-tyrosine kinases and association with a phosphatidylinositol kinase (PtdIns kinase) (8-11). In addition, phosphorylation of tyrosine 315 of mT is critical for efficient transformation, PtdIns kinase associa- tion, and tumorigenesis (8, 9,12, 13).
The PtdIns kinase activity that associates with mT/pp- 60""" has been shown to associate with several activated protein-tyrosine kinases (8, 10, 14-19). The mechanism for association appears to require tyrosine phosphorylation and SH2 domains (reviewed in Refs. 20 and 21). PtdIns 3-kinase phosphorylates the 3-hydroxyl position of the inositol ring to produce phosphatidylinositol 3-phosphate (PtdIns-3-P) (16). In addition to phosphorylating phosphatidylinositol (PtdIns), the enzyme also phosphorylates phosphatidylinositol 4-phos- phate (PtdIns-4-P) and phosphatidylinositol 4,5-bisphos- phate (PtdIns-4,5-P2) to produce two recently characterized phospholipids, phosphatidylinositol 3,4-bisphosphate (PtdIns-3.4-Pz) and phosphatidylinositol 3,4,5-trisphosphate (PtdIns-3,4,5-P3) (22-24), respectively. The production of these more highly phosphorylated phospholipids is dependent on the activation of protein-tyrosine kinases, and, more im- portantly, the temporal production of these lipids makes them excellent candidates for important signaling molecules of cell division (25).
Phosphatidylinositol 3-kinase was recently purified to ho- mogeneity from rat liver cytosol (24) and was shown to be a heterodimer of 110- and 85-kDa subunits. The 85-kDa subunit is phosphorylated on tyrosine both in vivo in mT-transformed cells and upon co-precipitation with the mT-pp60'-s" complex in vitro (10). It has recently been demonstrated that tyrosine- phosphorylated mT reassociates with pp85 in vitro (26). The
* The abbreviations used are: mT, polyoma virus middle T antigen; mT-pp60""", polyoma virus middle T antigen-pp6O""" complex; PtdIns 3-kinase, phosphatidylinositol 3-(hydroxy)kinase; PtdIns, phosphatidylinositol; PtdIns-3-P, phosphatidylinositol 3-phosphate; PtdIns-4-P, phosphatidylinositol 4-phosphate; PtdIns-4,5-P2, phos- phatidylinositol 4,5-bisphosphate; PtdIns-3,4-P2, phosphatidylinosl- to1 3,4-bisphosphate; PtdIns-3,4,5-P3, phosphatidylinositol 3,4,5-tris- phosphate; HEPES, 4-(hydroxyethyl)-l-piperazineethanesulfonic acid; HPLC, high performance liquid chromatography; PAGE, poly- acrylamide gel electrophoresis; SDS, sodium dodecyl sulfate; MES, 4-morpholineethanesulfonic acid; DME, Dulbecco's modified Eagle's medium; PBS, phosphate-buffered saline; Fmoc, N-(g-fluorenyl) methoxycarbonyl.
5408

Polyoma Virus Middle T Antigen-ppGO"-"" Complex 5409
reassociation was done in solution and by blotting after SDS denaturation. Tyrosine phosphorylation of mT appeared to regulate the subcellular distribution of the 85-kDa protein (26). The reassociated pp85 from whole cell lysates is the same 85-kDa protein found in the purified PtdIns 3-kinase complex (24). In addition, the 85-kDa subunit of PtdIns 3- kinase has been cloned (27-29) and may function as the mediator for the interaction between the catalytic subunit of PtdIns 3-kinase and the activated platelet-derived growth factor (PDGF) receptor (27,28) and the mT-pp60c-"" complex (27).
In an attempt to better understand the importance of PtdIns 3-kinase and mT-pp60""" complex formation in the signal transduction pathway, we have reconstituted this com- plex in uitro using immunopurified mT-pp6O""" complex from overproducing insect cells and PtdIns 3-kinase purified from rat liver. When protein kinase assays were performed in vitro both subunits of PtdIns 3-kinase were phosphorylated on tyrosine residues. The reconstituted complex utilized PtdIns, PtdIns-4-P, and PtdIns-4,5-Pz as substrates to generate the three newly described phospholipids. These results are con- sistent with previous data obtained from in vivo studies (23). In addition, immunopurified rnT-pp6Oc-'" associated with PtdIns 3-kinase from a whole cell lysate after incubation in uitro, and this association was also dependent upon the pro- tein-tyrosine kinase activity of pp60""". The association of PtdIns 3-kinase and immunopurified mT/ppGO"-"" could be blocked by preincubating the whole cell lysate or the purified enzyme with a synthetic peptide derived from the mT se- quence. Tyrosine phosphorylation of the peptide was neces- sary to block association. The ability of the phosphorylated peptide to block association of PtdIns 3-kinase with the mT- pp6OC-"" complex provides additional evidence for the specific region of mT involved in binding PtdIns 3-kinase. This data supports the model we have recently proposed (20).
MATERIALS AND METHODS
Cell CuEture-NIH/3T3 and Rat-1 fibroblasts were maintained by standard cell culture techniques in DME supplemented to contain 10% calf serum. Spodoptera frugiperda (Sf9) cells were cultured and used as described by Piwnica-Worms et al. (30). Sf9 cells were seeded into 60-mm plates (3 X lo6 cells), allowed to attach, and then were coinfected with baculovirus-mT and bacu1ovirus-pp60"'"", or baculo- ~irus-pp60"~'" as described (30). Cell pellets were prepared 40 h postinfection as described below.
Preparation of Sf9 Cell Pellets, Lysates, and Immunoprecipitates- Sf9 cells were scrape-harvested from 60-mm cell culture plates in phosphate-buffered saline (PBS) and collected by centrifugation. The cell pellet was washed once with cold PBS and aliquoted to 1.5-ml microcentrifuge tubes and again separated by centrifugation. The supernatant was aspirated from the cells, and the pellets were quick- frozen in dry ice/ethanol and stored at -70 "C until the lysates were prepared.
Lysates were generated from the frozen pellets by the addition of 1.0 ml of standard lysis buffer (137 mM NaC1, 20 mM Tris-HC1 (pH 7.4), 1 mM MgC12, 1 mM CaClz, 10% glycerol, 1% Nonidet P-40, 150 p M vanadate, 0.1 mM phenylmethylsulfonyl fluoride, 1 pg/ml apro- tinin, and 1 pg/ml leupeptin) and incubated at 4 'C with constant rocking for 15 min. Lysates were cleared by centrifugation at 12,000 X g for 5 min at 4 "C. The mT-pp6OC-"" complex was purified by immunoprecipitation essentially as described (31). Briefly, anti-mT (a-mT) antiserum (kindly provided by D. Pallas and T. Roberts, Dana-Farber Cancer Institute, Harvard Medical School) were added to the lysates and incubated at 4 'C for 2 h with constant agitation. The immune complexes were collected on protein A-Sepharose (CL- 4B, Sigma) that had been prewashed in 1% bovine serum albumin and then stored in a 50% suspension with PBS. Immune complexes were washed 2 times with 1% Nonidet P-40 in PBS, 2 times with radioimmune precipitation buffer (20 mM HEPES (pH 7.5), 137 mM NaCl, 2 mM EDTA, 10% glycerol, 1% Nonidet P-40,0.1% SDS, 0.5% deoxycholate), twice with 0.5 M LiCl in 0.1 M Tris-HC1 (pH 7.4), and
finally 2 times with TNE (10 mM Tris-HC1 (pH 7.4), 100 mM NaC1, 1 mM EDTA). The immunoprecipitates were used immediately for in vitro phosphorylation and association assays.
Purification of Phosphatidylinositol 3-Kinase-PtdIns 3-kinase was purified to homogeneity from rat liver as previously described (24). The purified enzyme had a specific activity of approximately 500 nmol/mg/min when assayed as described below with PtdIns as the substrate. The enzyme was stored at 4 "C in 50 mM MES (pH 6.7), 100 mM KCl, 0.5 mM dithiothreitol, 1 pg/ml leupeptin and pepstatin A and used within 2 weeks of purification.
In Vitro Association of Phosphatidylinositol 3-Kinase with Immu- nopurified mT-pp6P-""-Washed mT-pp60c-a" immune complexes were incubated with varying amounts of purified PtdIns 3-kinase in 750 pl to 1 ml of standard lysis buffer (see above) a t 4 "C on a platform rocker. The typical incubation was for 2-3 h after which the com- plexes were collected by a brief centrifugation and washed twice with 1% Nonidet P-40 in PBS, twice with 0.5 M LiCl in 0.1 M Tris-HC1, and twice with TNE.
Mammalian whole cell lysates were prepared from confluent 10- cm cell culture plates. The monolayer was washed once with 4 "C PBS and lysed in 1.0 ml of standard lysis buffer. After 10 min of incubation on a rocker platform at 4 'C, the cells were scrape- harvested, and lysates were cleared by centrifugation at 12,000 X g for 5 min at 4 "C. Lysates were used immediately for association experiments with the immunopurified mT-pp6O'"" complex.
Peptides were mixed with whole cell lysates or the purified PtdIns 3-kinase at a final concentration of 50 p~ and incubated at 4 "C on a rocker platform for 30 min. The mixture was then incubated with the immunoprecipitated mT-pp6O""" complexes as described above.
Phosphatidylinositol Kinase Assays and Protein Kinase Assays- Phosphatidylinositol kinase assays were performed directly on the immune complex as described (8, 22). Sonicated phospholipids were added to the washed beads, and the reaction was initiated with the addition of 50 p~ ATP, 5-25 pCi of [y-32P]ATP (3000 Ci/mmol), 5 mM MgClz in 20 mM HEPES (pH 7.5). Reactions were incubated for 5 min at room temperature. The 50-p1 reactions were stopped by the addition of 80 pl of 1 M HCl and 1 p1 of 500 mM EDTA. The lipids were extracted with 160 p1 of methanol/chloroform (l:l), and the organic layer was collected for analysis.
Protein kinase assays were also performed directly on the immune complex with or without added PtdIns 3-kinase. The reaction was done in 20 mM HEPES (pH 7.4), 10 mM MgClZ, 1-5 p~ ATP, and typically 10-50 pCi of [T-~'P]ATP (3000 Ci/mmol) per assay. Exper- iments for the time course of phosphorylation of PtdIns 3-kinase by the mT-pp60""" complex were done with 100 p~ ATP. To analyze the phosphorylation of purified PtdIns 3-kinase, the purified enzyme was added directly to the washed immune complex prior to the protein kinase reaction. The reaction was stopped by the addition of 2 X SDS gel loading buffer, and samples were boiled for 5 min prior to SDS- PAGE.
Phosphoamino Acid Analysis-Phosphoamino acid analysis of the 3ZP-labeled proteins from the SDS gel was performed as described (32). Briefly, radioactive gel slices were excised after autoradiography, fixed in 30% methanol for several hours, and dried by rotoevapora- tion. The gel slices were treated with tosylphenylalanyl chloromethyl ketone-trypsin (Worthington) in 50 mM ammonium bicarbonate at 37 "C overnight. The supernatants were collected and dried and washed with successively decreasing volumes of H20 (eg. 1 ml, 500 pl, 300 pl, 200 pl, 100 pl). The tryptic fragments were hydrolyzed with 6 M HCl at 100 "C for 90 min. Samples were then dried and washed with HzO. The samples were dissolved and analyzed by electropho- resis in pyridine/acetic acid/water (1:10189, v/v/v) for 90 min at 800 V with nonradioactive standards. The standards were visualized by spraying with 0.2% ninhydrin in acetone, and radiolabeled samples were visualized by autoradiography.
Two-dimensional analysis was also performed as described (32). Electrophoretic separation in the first dimension was done in formic acid/acetic acid/HzO (25:78:897, v/v/v). Plates were thoroughly dried and then developed by ascending chromatography for the second dimension in a 2-propanol/HC1/HzO (7015:15, v/v/v) solvent system. Standards and samples were visualized as described above.
Tran3'S-label of NIH/3T3 Cells, Rat-1 Cells, and Sf9 Cells-Rodent cells were grown to approximately 80-90% confluence in 10-cm plates in DME supplemented to contain 10% calf serum. The culture me- dium was removed and replaced with 5 ml of DME minus methionine (GIBCO) that contained 500 pCi of Tran%-label (ICN, >lo00 Ci/ mmol) supplemented with one-third of the amount of unlabeled methionine normally used. The cells were cultured for 12-48 h and

5410 Polyoma Virus Middle T Antigen-pp6WsK Complex
then harvested as follows. Radioactive culture medium was aspirated, and the cell monolayer was washed 3 times with ice-cold PBS. The cells were lysed by the addition of 1 ml of standard lysis buffer (see above) and incubated on a rocker platform for 15-20 min at 4 "C. The lysate was harvested with the aid of a cell scraper and cleared of insoluble material by centrifugation at 12,000 X g for 5 min at 4 "C. The lysates were used immediately for association assays.
Sf9 cells were labeled at 38-40 h postinfection with Tran=S-label as described (30).
Peptide Synthesis-The phosphopeptide RENEpYMPMAPQIH corresponding to the hamster mT sequence (33) was synthesized essentially as described (34). Ne-Fmoc-L-phosphotyrosine (Fmoc- Tyr(P)) was prepared by treating L-phosphotyrosine (Sigma) with N- (9-fluorenylmethoxycarbony1)succinimide (Aldrich) (35). Solid-phase peptide synthesis was conducted on a Milligen/Biosearch 9600 syn- thesizer using His(Trt)-PAC resin (Milligen/Biosearch) a t a substi- tution level of 0.37 mmol/g. Acylation reactions with standard side chain-protected amino acids (Arg(Pmc), Glu(t-butyl)) were conducted with 4 eq each of Na-Fmoc amino acid, l-hydroxybenzotriazole, and diisopropylcarbodiimide in dimethyl formamide for 1 h at 22 "C; the peptide bond-forming reaction with Fmoc-Tyr(P) was conducted manually for 2 h with 3 eq of each reagent. Asn and Gln were incorporated as the respective pentafluorophenyl esters. N"-Fmoc deprotections were carried out with a mixture of piperidine, toluene, and dimethyl formamide (30:35:35, v/v/v) for 7 min. The peptide was cleaved from the resin, and side chain-protecting groups were removed by treatment with a mixture of trifluoroacetic acid, thioanisole, ethanedithiol, and anisole (905:3:2, by volume) for 2 h a t 4 "C. The cleaved product was precipitated in diethyl ether, desalted on a column (2.6 X 100 cm) of Bio-Gel P-2, equilibrated in 3 M acetic acid, and lyophilized. The major product was purified by preparative HPLC (Waters Prep 4000) on a Dynamax-3OOA 12-pm C8 column (41.4 X 250 mm) equipped with a matched guard column; peptides were eluted with a mobile phase composed of acetonitrile in 0.05% aqueous trifluoroacetic acid (80 ml/min). Amino acid analysis (N/D, 1.0; Q/ E, 3.0; P, 2.0; A, 1.0; M, 2.0; I, 1.0; Y, 1.0; H, 0.9; R, 0.9) and fast- atom bombardment mass spectrometry (molecular ion a t m/z 1699.3) were as predicted.
Thin Layer Chromatography, Deacylatwn of Phospholipids, and HPLC Anulysis-Intact phospholipids were analyzed on oxalate- treated silica gel 60 plates (E. Merck) in a solvent system of 1- propanol, 2 M acetic acid (6535, v/v). For experiments in which only PtdInsP and/or PtdInsP, were analyzed, the faster chloroform, meth- anol, 2.2 M NHzOH (9:7:2, v/v/v) solvent system was utilized. Phos- pholipids were deacylated and analyzed by HPLC as previously described (31). 3H-Labeled phosphoinositides and inositol standards were obtained from Du Pont-New England Nuclear, and the lipids were deacylated by the same method used for the 32P-labeled samples. The standards and samples were co-injected with the nonradioactive nucleotides ADP and ATP for each HPLC analysis.
RESULTS
Association of Immunopurified mT-pp6OC-'" with Purified Phosphutidylinositol 3-Kinase Is Dependent on Protein-Tyro- sine Kinase Activity-PtdIns 3-kinase associates with the mT- pp60'-"" complex in vivo and can be immunoprecipitated from mT-transformed cells using antibodies against mT or pp60'-
(8,9, 16). The mT-pp6O"" complex has been generated in insect cells by co-infection with recombinant baculoviruses encoding mT and pp60""" genes (30). This complex has been shown to have protein-tyrosine kinase activity. Both mT and pp60'.'" are phosphorylated on tyrosine in the co-infected cells, and the purified complex has tyrosine kinase activity toward exogenous substrates. A kinase-defective complex, mT-pp60zgk.3m, has also been characterized.' The pp60295c~s" mutant protein associates with mT but does not have protein tyrosine kinase activity. The mT-pp6O""" and mT/pp60'95c~"m complexes were purified from infected Sf3 cells using a-mT antibodies conjugated to protein A-Sepharose beads. The purified complexes lacked PtdIns 3-kinase activity, although a small amount of PtdInsP kinase activity was detected (Figs. 1 and 3).
* H. Piwnica-Worms, unpublished observations.
c - s r c m T / p p 6 0 m T / p p 6 0
2 Y S c - s r e
P t d I n s 3 - k i n a s e : + +
Ptd
Ptd
Ptd I
n s P
n s q
n s q
1 2 3 4 FIG. 1. Analysis of a phosphatidylinositol kinase reaction
after association of purified rat liver PtdIns 3-kinase with baculovirus-expressed, immunopurified mT-pp6Pm and mT-pp60296'~"". The mT complexes were immunopurified as de- scribed and incubated with or without purified PtdIns 3-kinase at 4 "C for 2 h in standard lysis buffer. Phosphatidylinositol kinase assays were done with a mixture of phosphatidylserine, PtdIns, PtdIns-4-P, and PtdIns-4,5-P2 as lipid substrates (1:1:1:2). Immuno- purified, baculovirus-expressed mT-pp6O""" (lanes 1 and 2) and mT- pp60295E."" (lanes 3 and 4 ) were used. Purified PtdIns 3-kinase was added (lunes 2 and 4 ) or was omitted (lanes 1 and 3). The migration of PtdInsP, PMInsP2, and PtdInsPs standards is indicated.
In order to test for in vitro association between the mT-pp- 60'"" complex and PtdIns 3-kinase, purified rat liver PtdIns 3-kinase (24) was incubated with the baculovirus-expressed mT-ppGO'~". After incubation at 4 "C, the beads containing the complex were washed and then assayed for lipid kinase activity. As shown in Fig. 1, PtdIns, PtdIns-4-P, and PtdIns- 4,5-P2 kinase activities associated with the mT-pp60'-3m com- plex when purified PtdIns 3-kinase was added. However, very little PtdIns kinase activity associated with the protein ki- nase-deficient mT-pp6029k-"" complex (Fig. 1, lane 4) . This is in spite of there being equivalent amounts of complex in each case (data not shown). Thus, the in vitro association of PtdIns 3-kinase with the mT-pp6O"" complex appears to be depend- ent on the protein-tyrosine kinase activity of pp60""". These results are consistent with previous in vivo findings that mT must associate with and activate pp60"'"" in order for PtdIns kinase association (9, 13).
The results in Fig. 1 demonstrated that immunopurified mT-pp60c-"" and purified PtdIns 3-kinase associate in vitro. Association was also demonstrated by adding purified PtdIns 3-kinase directly to the insect cell lysate and incubating the mixture at 4 "C. After incubation, a-mT antiserum was added, and the immune complexes were collected on protein A- Sepharose and then washed. Lipid kinase assays demon-

Polyoma Virus Middle T Antigen-ppGO"-"" Complex 5411
strated that purified PtdIns 3-kinase associated with the mT- pp60"'"" complex in the crude insect cell lysate (data not shown). A monoclonal antibody (EC10) directed against pp60"'" also immunoprecipitated the mT-pp6O""" complex and associated lipid kinase activity (data not shown).
Phosphorylation and Phosphoamino Acid Analysis of the Phosphatidylinositol 3-Kinase-Purified PtdIns 3-kinase is a heterodimer of 85- and 110-kDa proteins (24). The 85-kDa subunit has been shown to be phosphorylated on tyrosine in uiuo in mT-transformed cells (10, 24). In order to determine if one or both of the subunits were phosphorylated on tyrosine residues in uitro by mT-ppGO"'", protein kinase reactions were performed directly with the immunoprecipitated complex. PAGE of the proteins after the phosphorylation reaction established that both the 110- and 85-kDa polypeptides were phosphorylated (see Fig. 2A) . As seen in Fig. 2A, more label was incorporated into the 85-kDa subunit than into the 110- kDa subunit. This result reflects preferential phosphorylation of the 85-kDa subunit as PtdIns 3-kinase purified from rat liver has equimolar amounts of the 85- and 110-kDa subunits (24). The purified enzyme has two distinct but related 110- kDa subunits with slightly different mobilities on SDS- PAGE. 35S-Labeling studies of cultured cells (discussed below) indicated that the difference in phosphorylation between the 85- and 110-kDa subunits is not due to differential immuno- precipitation or release from the immune complex, as similar amounts of labeled protein remained in the complex. For both the 110- and 85-kDa polypeptides the phosphorylation was saturable and reached a plateau after 30 min (data not shown). The stoichiometry of phosphorylation was calculated to be 1.2 mol of phosphate/mol of 85-kDa protein added to the assay. The stoichiometry was 0.38 mol of phosphate per mol of 110-kDa protein.
Phosphoamino acid analysis demonstrated that both the 85- and 110-kDa subunits were phosphorylated on tyrosine residues, as expected for phosphorylation by mT-ppGO""" (Fig. 2, B and C).
Complex Formation of mT-pp6@" with Phosphatidylinosi- to1 3-Kinase from Whole Cell Lysates-The immunopurified mT-pp60""" also associated with PtdIns 3-kinase from fibro- blast lysates. NIH/3T3 or Rat-1 lysates were prepared from confluent cells in standard lysis buffer as described under "Materials and Methods." Immunopurified mT-pp60""" com- plexes were added to the whole cell lysates and incubated at 4 'C. The protein A-Sepharose-conjugated mT-pp60c-"" com- plex was then washed with detergent and salt and assayed for associated phospholipid kinase activity. The resultant sam- ples were extracted and deacylated for analysis by HPLC anion exchange chromatography. As depicted in Fig. 3, PtdIns, PtdIns-4-P, and PtdIns-4,5-P2 kinase activities were present. The HPLC migration positions of the three products were consistent with the structure PtdIns-3-P, PtdIns-3,4-P2, and PtdIns-3,4,5-P3.
Immunopurified mT-pp60295e-"" (the kinase-inactive mu- tant) was unable to associate with the endogenous PtdIns 3- kinase activity of fibroblast whole cell lysates as would be expected from the results with purified PtdIns 3-kinase (data not shown). A small amount of PtdIns-4,5-P2 was generated in the immunoprecipitates from both mT-pp6O""" and mT- pp60206c~"" complexes (Fig. 3B). This is consistent with the results in Fig. 1 in which some PtdInsP kinase activity was found associated with both of these complexes in the absence of added PtdIns 3-kinase. Thus, the ability of the mT-pp- 60""" to associate with PtdIns 3-kinase from whole cell lysates was also dependent upon the protein-tyrosine kinase activity of pp60""".
The amount of PtdIns 3-kinase activity that associated with the mT-pp6O"-"" complex was dependent on the amount of lysate in the incubation. Titration of Rat-1 lysates against a fixed amount of mT-pp6O""" complex revealed the immune complex was in excess, and lysate from approximately 2.5 X IO6 cells represented 50% saturation (data not shown).
Proteins Associated with Baculouirus Expressed mT-pp- 6@-"" from Whole Cell Lysates-To define the specific proteins which associated with the protein-kinase active complex NIH/3T3 cells were labeled with Tran3'S-label, and whole cell lysates were prepared. Immunopurified protein A-conju- gated mT-pp60c-"" and mT-pp60295e-"" were added to separate radiolabeled lysates and incubated at 4 "C for 3 h. The com- plexes were washed with Nonidet P-40, LiC1, and TNE. The associated proteins were analyzed by SDS-PAGE, and the results are shown in Fig. 4. Although a number of radiolabeled peptides associated with both the protein-kinase active (lane I ) and -kinase inactive (lane 2) complexes, the most striking difference was observed in the 85- and the 110-kDa regions as indicated by the arrows (Fig. 4). A band at 173 kDa also preferentially associated with the active mT-ppGO"'" complex. These results are analogous to the results obtained with the purified PtdIns 3-kinase, in that both the 110-kDa doublet and the 85-kDa polypeptide associated with the mT-pp60c-a" complex in uitro.
Peptides That Block the Association of PtdIns 3-Kinase with the mT-pp6@-'" Complex-Data presented demonstrate that tyrosine kinase activity in the mT-pp60c-"" complex is re- quired for association of PtdIns 3-kinase. The major site of tyrosine phosphorylation on murine mT by pp60"-"" is tyrosine 315 (36). Amino acid sequence in this region of mT is con- served in various protein-tyrosine kinases that have been observed to associate with PtdIns 3-kinase (20). The in uitro association was further characterized, and the importance of tyrosine phosphorylation was directly tested. Competition experiments were done with a phosphorylated and unphos- phorylated version of a synthetic 13-amino acid peptide based on the region of hamster mT. This sequence is related to murine mT at tyrosine 315 (see Table I).
Preincubation of the phosphorylated peptide with whole cell lysates blocked association of PtdIns 3-kinase with the immunopurified mT-pp6O""" complex (Fig. 5, lanes 5 and 6). Preincubation of the unphosphorylated version of the peptide with cell lysates did not affect the association (Fig. 5, lanes 3 and 4 ) . The phosphorylated version of the peptide could also block the association of PtdIns 3-kinase activity with the immunopurified mT-ppGO"-"" when purified PtdIns 3-kinase was used for the association assay (data not shown).
Proteins that were prevented from associating with the mT-pp6O"-"" complex by the phosphorylated peptide were also determined. Whole cell lysates from Tran35S-labeled Rat-1 cells were preincubated in the absence of peptide, with phos- phorylated peptide or unphosphorylated peptide, and then association with exogenously added mT/pp60c-"" was moni- tored. Fig. 6 shows that proteins of similar molecular weight to the subunits of purified PtdIns 3-kinase (a doublet around 110 kDa and an 85-kDa band) associated with the kinase active complex (lane 2) but not with the protein kinase- deficient complex (lane 1 ). The unphosphorylated peptide did not affect this association (compare lanes 3 and 4 with lanes 1 and 2). However, the phosphorylated peptide specifically blocked three proteins (lane 6, see arrows) from associating with the kinase active mT-pp6O"-"" complex. Other, higher molecular weight proteins that associated in a protein kinase- dependent manner (compare lanes 1 and 2 or lanes 3 and 4 )

5412
A 1 2
Polyoma Virus Middle T Antigen-ppGV-"" Complex
B C
. . . . .. . I ...; . .
FIG. 2. Phosphorylation of purified PtdIns 3-kinase by the immunopurified mT-pp60"'" complex. A protein kinase reaction was performed using immunopurified mT-pp60""" complex in the presence or ahsence of purified PtdIns 3-kinase. The protein kinase reaction was initiated with the addition of [y-""I]ATP and stopped after 10 min with the addition of 2 X gel loading buffer (see "Materials and Methods"). The samples were hoiled for 5 min and analyzed on a 6% polyacrylamide gel. A, autoradiography of gel-separated proteins. L a n p 1 is m7" pp60""" in the absence of PtdIns 3-kinase, and lane 2 is in the presence of PtdIns %kinase. Arrows indicate the position of the 110-kDa, 85-kDa, pp60"" (BO-kDa), and mT (58-kDa) proteins. H , two-dimensional phosphoamino acid analysis of the 85-kDa hand from the in vitro kinase reaction was performed as descrihed under "Materials and Methods." P-SER, phosphoserine; P-THR, phosphothreonine; P-TYR, phosphotyrosine. C, one-dimensional phosphoamino acid analysis of the 110-kDa proteins from the in vitro kinase reaction.
were not blocked by the phosphorylated peptide (lanes 5 and 6).
DISCUSSION
We have reconstituted purified PtdIns 3-kinase with recom- binant, baculovirus-expressed, immunopurified mT-pp6O""" in vitro. The PtdIns 3-kinase associated with protein-tyrosine kinase active mT-ppGO'."" but failed to associate with an inactive mutant complex. ATP was not required during the precipitation, indicating that phosphorylation of PtdIns 3- kinase itself was not needed for tight complex formation. Previous work has shown that mT is phosphorylated on Tyr when co-expressed with pp6W."" in insect cells (30). The ability of the wild-type mT-pp6O""" complex but not the mutant to associate with PtdIns 3-kinase (Fig. 1) thus sug- gests that tyrosine phosphorylation of the mT-pp6O"'" com- plex is needed for the association. This result is consistent with the observation that mT must be phosphorylated on Tyr in order to blot the 85-kDa subunit of PtdIns 3-kinase using the Western blot procedure (24, 26).
Consistent with this idea, a phosphotyrosine-containing peptide based on a region highly conserved between hamster and murine polyoma m T (Table I) blocked association of PtdIns 3-kinase activity with mT-pp60c-"". The non-phos- phorylated peptide had no effect on the association. The phosphotyrosine of this sequence is analogous to Tyr-315 of murine mT, the major site of tyrosine phosphorylation in vivo (36). A mutation of Tyr-315 to Phe reduced (but did not eliminate) association with PtdIns 3-kinase i n vivo (8), and a n antibody against this region preferentially immunoprecip- itates the fraction of mT that does not associate with PtdIns 3-kinase (13). Mutations in the region of Tyr-315 also com- promise the transforming ability of polyoma virus without affecting the tyrosine kinase activity of the mT-pp60""" com- plex, indicating that this region is critical for in viuo targeting of the protein-tyrosine kinase (20).
As pointed out previously, receptor-type tyrosine kinases that bind PtdIns 3-kinase have short stretches of sequences with similarity to the Tyr-315 region of mT (20). In the PDGF receptor, the sequences around Tyr-740 and Tyr-751 obey these rules (Table I), and the latter site is known to be phosphorylated in response to PDGF binding (37). Both Tyr- 751 and Tyr-740 have been implicated in binding PtdIns 9- kinase (37-40), and peptides phosphorylated at Tyr-740 or Tyr-751 have been shown to block association of purified PDGF receptor with PtdIns %kinase from cell lysates (40). Thus, our results with mT-pp6O""" are analogous to the results with the PDGF receptor and extend these studies to show that purified PtdIns 3-kinase will complex with mT- pp6OC."" without the need of other cellular factors.
In addition to receptor-type tyrosine kinases, the major substrate of the insulin receptor tyrosine kinase (IRS-1) con- tains several repeats of sequences similar to the Tyr-915 region of polyoma mT (41) (Table I). This protein is not a tyrosine kinase but associates with PtdIns .?-kinase in insulin- stimulated (but not unstimulated) cells (41). IRS-1 has two regions of sequence that are quite similar to the hamster mT phosphopeptide used for competition studies in this paper (Table I). Further work is needed to determine the sequence specificity of the tyrosine-phosphorylated peptide domains that bind PtdIns 3-kinase and what proteins utilize these domains for recruitment of this enzyme.
The results presented here provide the first evidence that the 110-kDa subunit of PtdIns %kinase associates with the mT-ppGO'."" complex. The 110-kDa subunit was not phos- phorylated as well as the 85-kDa subunit in the reconstituted complex (Fig. 2) and was not previously detected in "P(7:-- labeled proteins associated with mT immunoprecipitates from polyoma-infected cells (10). However, using [,""S]methionine- labeled cells, the major proteins that associated with baculo- virus-expressed mT-pp6O""" complex and failed to associate with the kinase-defective mutant complex migrated at the
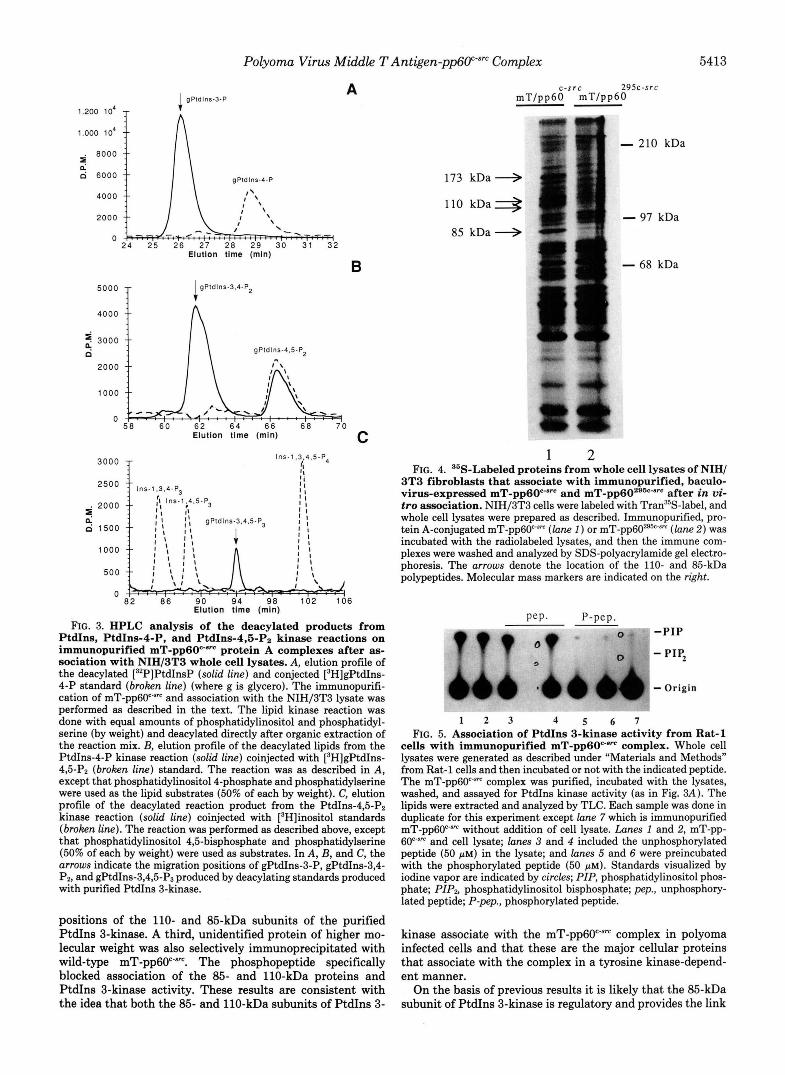
Polyoma Virus Middle TAntigen-pp6O""" Complex 5413
1.200 lo4
1.000 10'
g so00
d 6000
4000
2000
0
A 1 gPldlns-3-P
5000 T
Elutlon t h e (rnln)
B IgPldlns-3.4-P2
4000 f
1000
0 58 60 62 64 66 68 70
Elutlon tlme (rnln) C
3000 T lns - l .3 .4 ,S-P4
* 2ooo $ 1500
1000
500
0 8'2 86 90 9 4 9 8 102 106
Elution tlrne (mln)
FIG. 3. HPLC analysis of the deacylated products from PtdIns, PtdIns-4-P, and PtdI11s-4,Fi-p~ kinase reactions on immunopurified mT-pp60c-"" protein A complexes after as- sociation with NIH/3T3 whole cell lysates. A, elution profile of the deacylated [32P]PtdInsP (solid line) and conjected [3H]gPtdIns- 4-P standard (broken line) (where g is glycero). The immunopurifi- cation of mT-pp6O""" and association with the NIH/3T3 lysate was performed as described in the text. The lipid kinase reaction was done with equal amounts of phosphatidylinositol and phosphatidyl- serine (by weight) and deacylated directly after organic extraction of the reaction mix. E, elution profile of the deacylated lipids from the PtdIns-4-P kinase reaction (solid line) coinjected with [3H]gPtdIns- 4,5-Pz (broken line) standard. The reaction was as described in A, except that phosphatidylinositol 4-phosphate and phosphatidylserine were used as the lipid substrates (50% of each by weight). C, elution profile of the deacylated reaction product from the PtdIns-4,5-Pz kinase reaction (solid line) coinjected with [3H]inositol standards (broken line). The reaction was performed as described above, except that phosphatidylinositol 4,5-bisphosphate and phosphatidylserine (50% of each by weight) were used as substrates. In A, B, and C, the arrows indicate the migration positions of gPtdIns-3-P, gPtdIns-3,4- Pz, and gPtdIns-3,4,5-P3 produced by deacylating standards produced with purified PtdIns 3-kinase.
positions of the 110- and 85-kDa subunits of the purified PtdIns 3-kinase. A third, unidentified protein of higher mo- lecular weight was also selectively immunoprecipitated with wild-type mT-pp6O"'"". The phosphopeptide specifically blocked association of the 85- and 110-kDa proteins and PtdIns 3-kinase activity. These results are consistent with the idea that both the 85- and 110-kDa subunits of PtdIns 3-
c-src 295c-src m T / p p 6 0 m T / p p 6 0
173 kDa + 110 k D a 3
85 kDa 4
- 210 kDa
- 97 kDa
- 68 kDa
1 2 FIG. 4. "S-Labeled proteins from whole cell lysates of NIH/
3T3 fibroblasts that associate with immunopurified, baculo- virus-expressed mT-pp60"'"" and mT-pp60286"~"~ after in vi- tro association. NIH/3T3 cells were labeled with Tran35S-label, and whole cell lysates were prepared as described. Immunopurified, pro- tein A-conjugated mT-pp60""" ( l a n e 1 ) or mT-pp60z9k-sm (lane 2 ) was incubated with the radiolabeled lysates, and then the immune com- plexes were washed and analyzed by SDS-polyacrylamide gel electro- phoresis. The arrows denote the location of the 110- and 85-kDa polypeptides. Molecular mass markers are indicated on the right.
Pep. P-pep. " -7 -PIP
- PIP,
b - Origin
1 2 3 4 5 6 7 FIG. 5. Association of PtdIns 3-kinase activity from Rat-1
cells with immunopurified mT-pp60c-"" complex. Whole cell lysates were generated as described under "Materials and Methods" from Rat-1 cells and then incubated or not with the indicated peptide. The mT-pp6F" complex was purified, incubated with the lysates, washed, and assayed for PtdIns kinase activity (as in Fig. 3A). The lipids were extracted and analyzed by TLC. Each sample was done in duplicate for this experiment except lane 7 which is immunopurified mT-pp60c'sm without addition of cell lysate. Lanes I and 2, mT-pp- 60""" and cell lysate; lanes 3 and 4 included the unphosphorylated peptide (50 p ~ ) in the lysate; and lanes 5 and 6 were preincubated with the phosphorylated peptide (50 p ~ ) . Standards visualized by iodine vapor are indicated by circles; PIP, phosphatidylinositol phos- phate; PIP, phosphatidylinositol bisphosphate; pep., unphosphory- lated peptide; P-pep., phosphorylated peptide.
kinase associate with the mT-pp6O"'" complex in polyoma infected cells and that these are the major cellular proteins that associate with the complex in a tyrosine kinase-depend- ent manner.
On the basis of previous results it is likely that the 85-kDa subunit of PtdIns 3-kinase is regulatory and provides the link

5414 Polyoma Virus Middle T Antigen-ppGO"-"' Complex
1 n T / p p 6 6 " ~ ' - + - + - + m T / p p 6 0 + - + - + - Pep. P-pep. - . - - - + +
295c-src
- - + + "
173 kDa 4
110 kDa E.
85 kDa +
1 2 3 4 5 6
FIG. 6. Association of "S-labeled proteins from Rat-1 whole cell lysates with mT-pp6Wam or mT/pp60296c"m with or without preincubation with peptide. Rat-1 cells were labeled with Tran3'S-label, and lysates were prepared as described. The lysates were incubated with phosphorylated peptide (lanes 5 and 6), unphos- phorylated peptide (lanes 3 and 4) , or no peptide (lanes 1 and 2). The samples were then incubated with immunopurified mT-pp-
immune complexes were collected and analyzed by SDS-PAGE on a 6% gel. The upper double arrows denote the location of the 110-kDa proteins, and the lower arrow indicates the location of the 85-kDa protein.
6029b8m (lanes I, 3, and 5) or mT-pp60c-am (lanes 2, 4, and 6). The
TABLE I Sequence of the hamster mT peptide and similar sequences from
proteins known to associate with PtdIns 3-kinase The hamster mT peptide sequence was synthesized and used for
experiments as described in the text. The analogous sequence in the PDGF receptor has been shown to be involved in the binding of PtdIns 3-kinase after receptor phosphorylation. The specific sites of tyrosine phosphorylation in hamster mT and rat IRS-1 have not been mapped (indicated by *). Y, phosphorylated tyrosine; the number in parentheses indicates the location of the Tyr in the protein sequence.
Protein Seauence
Hamster mT (298*) RENEYMPMAPQIH Murine mT (315) EEEEYMPMEDLYL Rat IRS-1 (608*) TDDGYMPMSPGVA Rat IRS-1 (628*) GNGDYMPMSPKSV Rat IRS-1 (lolo*) APVSYADmTGIA Human PDGF Rec.j3 (751) ESVDWPMLDMKG Human PDGF Rec.6 (740) SDGGYMDMSKDES
between mT and the 110-kDa subunit. The 110-kDa subunit is probably the catalytic subunit of P a n s 3-kinase (42). The 85-kDa protein has two -80-amino acid stretches of homology to the amino-terminal non-catalytic domain of pp60'"" (SH- 2 domain) (27-29). SH-2 domains have been shown to bind to tyrosine-phosphorylated regions of target proteins to form 'tight complexes both in vivo and in vitro (43) (reviewed in Refs. 20 and 21). Tyrosine-phosphorylated mT specifically blots the 85-kDa (but not the 110-kDa) subunit of PtdIns 3- kinase using a Western blot procedure (24, 26). The regions of the 85-kDa subunit required for the association with mT and with the 110-kDa subunit are under investigation.
Finally, reconstitution of purified PtdIns 3-kinase with mT- pp60""" did not have a major effect on substrate specificity. The purified enzyme utilizes three substrates in vitro (PtdIns, PtdIns-4-P, and PtdIns-4,5-Pz), and the relative activities
toward these substrates are very sensitive to assay conditions (24). However, association of PtdIns 3-kinase from whole cell lysates with the mT-pp6OC*'" did change the substrate speci- ficity from using only PtdIns to utilizing PtdIns, PtdIns-4-P, and PtdIns-4,5-Pz (24): Cells transformed with polyoma mT or stimulated with PDGF have increased levels of PtdIns-3,4- Pz and PtdIns-3,4,5-P3 but little change in PtdIns-3-P sug- gesting that transformation by polyoma virus or by activation with PDGF enhances phosphorylation of PtdIns-4-P and/or PtdIns-4,5-Pz by PtdIns 3-kinase (22, 23, 44). Using our standard assay conditions the ratio of products formed when a mixture of all three substrates was presented was similar whether pure enzyme or reconstituted mT-pp6O""-PtdIns 3- kinase complex was used. The ratio was also similar to that found in mT immunoprecipitates from mT-transformed cells (23). The factors that affect in vivo specificity of this enzyme remain to be determined.
Acknowledgments-We thank B. C. Duckworth and R. Kapeller for help in purifying PtdIns 3-kinase and J. Falk and M. S. Atkisson for their excellence in culture of the Sf9 cells. We also thank Brian S. Schaffhausen for helpful discussions and critical reading of the manuscript.
REFERENCES
1. Cheng, S. H., Harvey, R., Espino, P. C., Semba, K., Yamamoto, T., Toyoshima, K., and Smith, A. E. (1988) EMBO J. 7,3845- 3855
2. Courtneidge, S. A., and Smith, A. E. (1983) Nature 303, 435- 439
3. Kornbluth, S., Sudol, M., and Hanafusa, H. (1987) Nature 325, 171-173
4. Kypta, R. M., Hemming, A., and Courtneidge, S. A. (1988) EMBO
5. Bolen, J. B., Thiele, C. J., Israel, M. A., Yonemoto, W., Lipsich, L. A., and Brugge, J. S. (1984) Cell 38,767-777
6. Courtneidge, S. A. (1985) EMBO J. 4,1471-1477 7. Raptis, L., Lamfrom, H., and Benjamin, T. L. (1985) Mol. Cell.
8. Whitman, M., Kaplan, D. R., Schaffhausen, B., Cantley, L., and Roberts, T. M. (1985) Nature 315,239-242
9. Kaplan, D. R., Whitman, M., Schaffhausen, B., Raptis, L., Gar- cea, R. L., Pallas, D., Roberts, T. M., and Cantley, L. (1986) Proc. Natl. Acud. Sci. U. S. A. 83,3624-3628
10. Kaplan, D. R., Whitman, M., Schaffhausen, B., Pallas, D. C., White, M., Cantley, L., and Roberts, T. M. (1987) Cell 50,
J. 7,3837-3844
BWl. 5,2476-2485
1021-1029 11. Courtneidge, S. A., and Heber, A. (1987) Cell 50,1031-1037 12. Carmichael, G., Schaffhausen, B. S., Mandel, G., Liang, T. J.,
and Benjamin, T. L. (1984) Proc. Natl. Acud. Sci. U. S. A. 81,
13. Talmage, D. A., Freund, R., Young, A. T., Dahl, J., Dawe, C. J., and Benjamin, T. L. (1989) Cell 59,55-65
14. Sugimoto, Y., Whitman, M., Cantley, L. C., and Erikson, R. L. (1984) Proc. Natl. Acud. Sci. U. S. A. 81, 2117-2121
15. Macara, I. G., Marinetti, G. V., and Balduzzi, P. C. (1984) Proc. Natl. Acud. Sci. U. S. A. 81,2728-2732
16. Whitman, M., Downes, C. P., Keeler, M., Keller, T., and Cantley, L. (1988) Nature 332,644-646
17. Varticovski, L., Drucker, B., Morrison, D., Cantley, L., and Rob- erts, T. (1989) Nature 342,699-702
18. Fukui, Y., Kornbluth, S., Jong, S.-m., Wang, L.-h., andHanafusa, H. (1989) Oncogene Res. 4,283-292
19. Ruderman, N. B., Kapeller, R., White, M. F., and Cantley, L. C. (1990) Proc. Natl. Acud. Sci. U. S. A. 87,1411-1415
20. Cantley, L. C., Auger, K. R., Carpenter, C., Duckworth, B.,
302 Graziani, A., Kapeller, R., and Soltoff, S. (1991) Cell 64, 281-
21. Koch, C. A., Anderson, D., Moran, M. F., Ellis, C., and Pawson, T. (1991) Science 252,668-674
22. Auger, K. R., Serunian, L. A., Soltoff, S. P., Libby, P., and Cantley, L. C. (1989) Cell 57, 167-175
679-683
K. R. Auger, unpublished data.

Polyoma Virus Middle T Antigen-pp6Psm Complex 5415 23. Serunian, L. A., Auger, K. R., Roberts, T., and Cantley, L. C.
(1990) J. Virol. 64, 4718-4725 24. Carpenter, C. L., Duckworth, B. C., Auger, K. R., Cohen, B.,
Schaffhausen, B. S., and Cantley, L. C. (1990) J. Biol. Chem.
25. Auger, K. R., and Cantley, L. C. (1991) Cancer Cells (Cold Spring Harbor) 3,263-270
26. Cohen, B., Yoakim, M., Piwnica-Worms, H., Roberts, T. M., and Schaffhausen, B. S. (1990) Proc. Natl. Acad. Sci. U. S. A. 87, 4458-4462
27. Otsu, M., Hiles, I., Gout, I., Fry, M. J., Ruiz-Larrea, F., Panayo- tou, G., Thompson, A., Dhand, R., Hsuan, J., Totty, N., Smith, A. D., Morgan, S. J., Courtneidge, S. A., Parker, P. J., and Waterfield, M. D. (1991) Cell 66,91-104
28. Escobedo, J. A., Navankasattusas, S., Kavanaugh, W. M., Milfay, D., Fried, V. A., and Williams, L. T. (1991) CeU 66, 75-82
29. Skolnik, E. Y., Margolis, B., Mohammadi, M., Lowenstein, E., Fischer, R., Drepps, A., Ullrich, A., and Schlessinger, J. (1991) Cell 66,83-90
30. Piwnica-Worms, H., Williams, N. G., Cheng, S. H., and Roberts, T. M. (1990) J. Virol. 64,61-68
31. Auger, K. R., Serunian, L. A., and Cantley, L. C. (1990) in Methods in Znositide Research (Irvine, R. F., ed) pp. 159-166, Raven Press, New York
32. Parker, L. L., Atherton-Fessler, S. E., Lee, M. S., Ogg, S., Falk,
266,19704-19711
J. L., Swenson, K. I., and Piwnica-Worms, H. (1991) EMEO J.
33. Delmas, V., Bastien, C., Scherneck, S., and Feunteun, J. (1985)
34. Chatterjee, S., Goldstein, B. J., Csermely, P., and Shoelson, S. E. (1992) in Peptides: Chemistry and Sio&~gy (Smith, J., and
35. Stewart, J. M., and Young, J. D. (1984) Solid Phase Peptide Rivier, J., eds) Escom, Leiden, in press
Synthesis, Pierce Chemical Co., Rockford, IL 36. Schaffhausen, B. S., and Benjamin, T. L. (1981) J. Virol. 40,
37. Kazlauskas, A., and Cooper, J. A. (1989) Cell 68,1121-1133 38. Kazlauskas, A., and Cooper, J. A. (1990) EMEO J. 9, 3279-3286 39. Heidaran, M. A., Pierce, J. H., Lombardi, D., Ruggiero, M.,
Gutkind, J. S., Matsui, T., and Aaronson, S. A. (1991) Mol. Cell. Eiol. 11, 134-142
40. Escobedo, J. A., Kaplan, D. R., Kavanaugh, W. M., Turck, C. W., and Williams, L. T. (1991) Mol. CeU. BioZ. 11,1125-1132
41. Sun, X. J., Rothenberg, P., Kahn, C. R., Backer, J. M., Araki, E., Wilden. P. A,. Cahill. D. A.. Goldstein. B. J.. and White. M. F.
10,1255-1263
EMBO J. 4,1279-1286
184-196
(1991) Nature 362,73-77 ’ 42. Shibasaki. F.. Homma. Y.. and Takenawa. T. (1991) J. Eiol. . . .
Chem. 266; 8108-8114 ’
Pawson, T. (1990) Science 260,979-982 43. Anderson, D., Koch, C. A., Grey, L., Ellis, C., Moran, M. F., and
44. Ulug, E. T., Hawkins, P. T., Hanley, M. R., and Courtneidge, S. A. (1990) J. V i d . 64,3895-3904
Copyright © 2022 FDOKUMEN