
Attentional orienting towards emotion: P2 and N400 ERP effects
Upload
independentCategory
view
1download
0
1996 88: 4183-4194
C CerlettiV Evangelista, S Manarini, S Rotondo, N Martelli, R Polischuk, JL McGregor, G de Gaetano and P-selectin and the beta 2 integrin CD11b/CD18conditions: evidence of adhesion cascade and cross talk between Platelet/polymorphonuclear leukocyte interaction in dynamic
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
reserved.Copyright 2011 by The American Society of Hematology; all rights900, Washington DC 20036.weekly by the American Society of Hematology, 2021 L St, NW, Suite Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

Platelet/Polymorphonuclear Leukocyte Interaction in Dynamic Conditions: Evidence of Adhesion Cascade and Cross Talk Between P-Selectin and the
p2 Integrin CDllbICD18
By Virgilio Evangelista, Stefano Manarini, Serenella Rotondo, Nicola Martelli, Roman Polischuk, John L. McGregor, Giovanni de Gaetano, and Chiara Cerletti
Adhesion between platelets and polymorphonuclear leuko- cytes (PMN) is a key event in thrombosis and inflammation. Double color fluorescence-activated cell sorter (FACS) analy- sis was used t o determine the extent and kinetics of adhe- sion of thrombin-activated platelets to resting or activated PMN when mixed cell populations were incubated in dy- namic conditions. Activated platelets bound very rapidly t o PMN. Mixed cell conjugates reached a maximum at 1 minute and were reversible within 10 minutes. PlateWPMN adhe- sion required both CB2+ and Mgz+ and was markedly in- creased by the presence of Mn2+. The latter made mixed cell conjugates stable up t o 10 minutes. Adhesion of platelets required metabolic activity of PMN and was abolished by tyrosine kinase inhibitors. Furthermore, adhesion of plate- lets t o PMN resulted in binding of a monoclonal antibody (MoAb 24) known as p2 integrins "activation reporter." When PMN were activated by exogenous stimuli, the adhe- sion of platelets was markedly increased: fMLP induced a rapid and transient effect, while PMA resulted in a slower, but stable, increase in mixed conjugates formation. The hy- pothesis that activated PMN p2 integrins are able to bind a
HE BROAD RANGE of interactions between platelets and polymorphonuclear leukocytes (PMN) in vitro in-
cludes reciprocal modulatory effects, as well as metabolic cooperation possibly playing an important role in inflamma- tion and
In addition to functional and metabolic interaction, plate- lets and PMN physically interact4 at the membrane level creating optimal conditions for the exchange of soluble me- diators' and intermediate metabolites.6 Moreover, the mem- brane receptors engaged during cell-cell adhesion may them- selves transduce functional signals.','
Recent studies demonstrated that P-selectin expressed on the surface of activated platelets sustains a main adhesive mechanism between platelets and PMN?" possibly recog- nizing different ligands on PMN membrane (see Varki" for review). P-selectin binding to PMN does not require PMN activation."." PMN interaction with P-selectin at physio- logic flow rates has been modeled in vitro on P-selectin- containing phospholipid bilayersI4 and has been shown to have rapid association and dissociation rates." According to this model P-selectidcounter-receptor interaction sustains the rolling of PMN.
Although many studies on plateletiPMN adhesion have been reported, only a few of them were performed in dy- namic conditions, showing P-selectin-dependent PMN roll- ing on immobilized platelet^.'^." Yeo et all7 suggested that an unknown, PMN activation-dependent mechanism is nec- essary, in addition to P-selectin, for a stable adhesion to occur. Indeed, P-selectin is not the only possible PMN recep- tor on platelets: fibrinogen expressed on platelet surface might also function as a PMN binding receptor." On the other hand, intercellular adhesion molecule-2 (ICAM-2) has also lately been demonstrated on platelets.lg
T
Blood, Vol 88, No 11 (December l), 1996: pp 4183-4194
counter-receptor on platelets was directly demonstrated by the increase of mixed cell conjugates following PMN treat- ment with KIM127 and KIM185, two anti-CD18 antibodies able t o induce the active conformation of p2 integrins. Con- sistently, two other anti-CD18, as well as an anti-CDllb in- hibitory antibody abolished platelet/PMN adhesion. PMN p2 integrin activation was not the only mechanism for activated platelet/PMN adhesion to occur: indeed, this phenomenon could also be inhibited by two anti-P-selectin antibodies. Resting platelets did not adhere to resting PMN, but mark- edly adhered t o fMLP- or PMA-activated PMN. Resting plate- let/fMLP-activated PMN adhesion was abolished by anti- CD18 antibodies, but not by anti-P-selectin antibodies. In conclusion, activated platelet/PMN interaction can be mod- eled as an adhesion cascade involving a P-selectin-depen- dent recognition step and a functional signal. The latter pro- ceeds through tyrosine kinase activation and enables a 82 integrin-dependent adhesion to a not yet identified counter- receptor constitutively expressed on platelet surface. 0 1996 by The American Society of Hematology.
On the endothelium, the selectin-dependent rolling is fol- lowed by tight adhesion of PMN that requires the additional binding of p2 integrins to ICAMs and probably other uniden- tified ligands on endothelial membrane. The double-step model of PMN-endothelial cell adhesion has been docu- mented by experiments performed both in vitro and in vivo. To become competent to bind the counter-receptor, p2 inte- grins require functional upregulation, which always follows PMN activation by inflammatory stimuli, operating in solu- tion or bound on cell membrane (see Carlos and Harlan" for review).
From the "G. Biuozero" Laboratory of Platelet and Leukocyte Pharmacology, Laboratory of Tumor and Vascular Cell Biology, Unit of Morphology, Istituto di Ricerche Farmacologiche Mario Negri, Consorzio Mario Negri Sud, Santa Maria Imbaro, Italy; and INSERM Unite 33 1, Lyon, France.
Submitted May 7, 1996; accepted July 22, 1996. Supported in part by the Italian National Research Council: Con-
venzione CNR-Consorzio Mario Negri Sud, Progetto Finalizzato FATMA Contract No. 95.00951.41, bilateral project (V.E. and J.McG.), Contract No. 95.01714.CT04, and CNR-INSERM agree- ment (C.C. and J.McC. J, No. 132.22.1.
R.P. is on leave of absence from the Department of Anatomy, State Medical Institute, Ivanovo, Russia.
Address reprint requests to Virgilio Evangelista, MD, "G. Bizzoz- ero" Laboratory of Platelet and Leukocyte Pharmacology, Consor- zio Mario Negri Sud, 66030 Santa Maria Imbaro, Italy.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact. 0 1996 by The American Society of Hematology. 0006-4971/96/8811-0010$3.00/0
4183
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

4184 EVANGELISTA ET AL
At variance with endothelial cells, platelets may interact with PMN in suspension: the formation of mixed platelet/ PMN aggregates in circulating blood has been reported in experimental animal models.2’*22 This situation is mimicked in vitro by the formation of mixed cell aggregates, showing very tight membrane to membrane heterotypic adhesion, when mixed cell populations were incubated in dynamic conditions in a ~uvette.’~
The aim of the present study was to investigate the mecha- nisms and the adhesive molecules mediating platelet/PMN adhesion in cell suspensions subjected to a high-speed rota- tory motion. This condition highlights high-affinity adhesive cell-cell interaction not easily disrupted at moderate or high shear forces and increases the frequency of cell-cell colli- sions.
MATERIALS AND METHODS
Chernicnls. Hydroethidine (HE) was purchased from Molecular Probes Europe (Leiden, The Netherlands); 2’, 7’-bis-(2-carboxy- ethyl)-5(6)-carboxy-fluorescein uiacetoxy methyl ester (BCECF- AM), fluorescein isothiocyanate (FITC) conjugate antimouse IgG (whole molecule), n-formyl-methyl-leucyl-phenylalanine (fMLP), prostaglandin El (WE,), phorbol 12-myristate, 13 acetate (PMA), N-2 hydroxyethyl piperazine-N I-2-ethanesulfonic acid (HEPES) ethylene glycol-bis (b-aminoethyl ether)-N. N. N’, N‘,-tetraacetic acid (EGTA), thrombin from human plasma (2,000 National Insti- tutes of Health [NIH] unitslmg of protein), geninstein were pur- chased from Sigma Chemical CO (St Louis, MO), tyrphostin AG 879 from LC Laboratories (Inalco, Milano, Italy), deionized water from Merck (Milano, Italy). Paraformaldehyde (PFA) was purchased from Fluka (Milano, Italy). Dextran T500 and Ficoll-Hypaque were from Pharmacia Fine Chemicals (Uppsala, Sweden), Aldrich Chim- ica srl (Milano, Italy); purified human fibrinogen from Kabi Diagnos- tics (Stockholm, Sweden). Ro 31-8220 was a gift of Roche Products Ltd (Welwyn Garden City, UK). fMLP was dissolved in dimethyl sulfoxide (DMSO) at concentrations of 100 mom, stored at -20°C. and diluted in isotonic saline just before use. Thrombin was dissolved in saline at concentrations of 50 U/mL, and stored at -20°C until use. BCECF-AM and HE were dissolved in DMSO at concentrations of l mg/mL and 8 mg/mL, respectively, stored at -20°C and used within 4 weeks.
Antibodies and peptides. The anti-P-selectin monoclonal anti- bodies (MoAbs), ACll and AC1.2: were kindly provided by Dr Bruce Furie (New England Medical Center Hospital, Boston, MA), P2024 by one of us (J.L. McG.) and GA@ was purchased from Becton Dickinson (Milano, Italy) together with the mouse Ig G1 control (clone X 40). P18, recognizing IIbIIIa and CI,&?~ was from J.L. McG. The anti-CDlla, TSl122;’ and the anti-CD18, TS1/18*’ were purified from mouse ascites or hybridoma cell (American Type Culture Collection, Rockville, MD) supemantant using protein G- sepharose affinity column. The anti-CD18, IB4;* and 60.32’ were kindly provided by Dr Samuel D. Wright (The Rockefeller Univer- sity, New York, NY) and Dr John Harlan (University of Washington, School of Medicine, Seattle, WA), respectively. The antibodies KIM127 and KIM1U3’ directed against CD18, both stimulating CD1 %dependent adhesion, were kindly provided by Dr Martyn Rob- inson (Celltech Therapeutics Ltd, Berkshire UK). LM2/1 and OKMl, both against CD1 Ib, were kindly provided by Dr Dario C. Altieri (Yale University, New Haven, CT). while MoAb 24;’ recognizing an activation-dependent epitope of all CI subunits of CD1 K D 1 8 complex was a kind gift of Dr Nancy Hogg (Imperial Cancer Research Fund, London, UK). LPM19c, an anti-CD1 Ib, was
kindly provided by Dr Karen Pulford (University of Oxford, Oxford, UK). Fibrinogen-derived peptides: Arg-Gly-Asp-Ser (RGDS) was purchased from Sigma; P1, duplicating the fibrinogen -y chain se- quence G’go-Vzoz, was kindly provided together with the control peptide by Dr Dario C . Altieri. P1 specifically prevents fibrinogen binding to CDllb.32
Preparation of PMN leukocytes. Blood was collected from healthy volunteers who had not received any medication for at least 2 weeks. Approval was obtained from the Institutional Review Board for these studies. Volunteers were informed that blood samples were obtained for research purposes and that their privacy would be pro- tected. Nine parts of blood were anticoagulated with one part of 3.8% trisodium citrate. Platelet-rich plasma (PRP) was prepared by centrifugation of whole blood at 200g for 15 minutes. PMN, isolated from the remaining blood by Dextran sedimentation followed by Ficoll-Hypaque gradient and hypotonic lysis of erythrocytes, were washed and resuspended in HEPES-Tyrode buffer (pH 7.4) con- taining 129 mmol/L NaCl, 9.9 mmoK NaHCO,, 2.8 mmol/L KCl, 0.8 mmollL, KH,PO,, 0.8 mmol/L MgCl,-6H20, 5.6 mmol/L dex- trose, 10 mmolfL HEPES, and 1 mmol/L CaCI,. Cellular suspensions contained 95% of PMN and an average of 1 platelet/30 PMN.
PMN were stained with the vital red fluorescent dye HE (20 pg/ 5.10’ cells/&) for 30 minutes at 4°C. Stained cells were washed three times in cold HEPES-Tyrode buffer, resuspended in the same cold medium and maintained at 4°C until the end of the experiment, which was usually performed within 1 hour after PMN preparation.
Energy-depleted PMN were prepared by washing and resus- pending the cells in a modified glucose-free HEPES-Tyrode con- taining 50 mmol/L 2-deoxyglucose, 0.05% NaN,, and 10 mmol/L glucono-&lactone to inhibit glycolysis, oxidative phosphorylation, and glycogen phosphorylase, respectively. The total adenosine tri- phosphate (ATP) content of treated PMN corresponded to about 20% of control, as evaluated in Triton-X-l00 lysed cells by luciferin- luciferase.
Preparation of platelets. Platelets were loaded with the green fluorescent dye BCECF by incubating PRF’ with 2 pg/mL of its membrane permeant acetoxymethyl ester (BCECF-AM) for 30 min- utes at 37°C. After loading, one third of the total PRP was diluted with one volume of 2% PFA and fixed for 1 hour at room temperature to obtain PFA-fixed platelets in resting state. The remaining two thirds of BCECF-loaded PRP was centrifuged at 1,lOOg for 15 min- utes in the presence of 2 pmol/L PGE, ; platelets were then washed in HEPES-Tyrode containing 2 pmo& PGE, and 5 mmoVL EGTA by further centrifugation at 1,lOOg for 10 minutes and resuspended in the same buffer with 1 mmol/L Ca2’ at a concentration of IO’ platelets/mL.
Three milliliters of platelet suspension was stimulated with throm- bin 0.5 U/mL for 2 minutes at room temperature, in static condition, immediately diluted to 50 mL with PFA (1% final concentration) and kept at room temperature for 1 hour.
After fixation, resting and thrombin-activated platelets were washed three times in HEPES-Tyrode buffer by centrifugation and resuspended in the same medium with 1 mmol/L Ca2+. Resuspending the platelet pellet by accurate pipetting allowed us to obtain a popula- tion of single platelets with less than 10% of small aggregates for both resting or thrombin-activated platelets as estimated by optical counting.
To minimize fibrinogen binding and platelet clumping, platelets were activated with thrombin in the presence of 800 pmoVL RGDS. Because no differences were observed with respect to the control condition, in the subsequent experiments platelets were activated in the absence of RGDS.
P-selectin expression in platelets fixed in PRP immediately after preparation was negligible as compared with nonexogenously acti-
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

CD1 1 WCD18 IN PLATELET/PMN LEUKOCYTE INTERACTION 4185
vated washed platelets: mean fluorescence intensity was, in fact, 9 versus 17 with 17% versus 65% of platelets expressing P-selectin, respectively. Virtually all thrombin-activated platelets showed a mean fluorescence intensity of 157 (mean of two different experi- ments).
The ability of BCECF-loaded platelets to respond to thrombin, evaluated in preliminary experiments as aggregation response, was similar to control platelets. After fixation, thrombin-activated plate- lets did not significantly aggregate under stimng in the absence of fibrinogen, but maintained the ability to aggregate in a RGDS- inhibitable manner in the presence of fibrinogen. The refractoriness to stimulation of PFA-fixed resting or thrombin-activated platelets was demonstrated by the inability of thrombin 0.5 U/mL or PMA 100 nmolk to induce further P-selectin expression.
Cytojluorimetric analysis of P-selectin expression on platelets and of Mac-I or MoAb 24 spec$c epitope on PMN. Fixed unstimulated or thrombin-activated platelets 10*/50 pL were incubated with or without AC1.2 (ascites, 1500 final dilution) for 30 minutes at room temperature, then washed twice in HEPES-Tyrode 1 mmoVL Ca2’ and resuspended in 50 pL of the same buffer. Samples were then incubated with FITC-conjugate antimouse IgG (whole molecule) for 30 minutes in the dark at room temperature. Samples were then washed twice and resuspended in 500 pL of HEPES-Tyrode buffer and analyzed by FACStar flow cytometer (FACStar, Becton Dickin- son, Milano, Italy) storing data in list mode files. Determination of the percentage of platelets expressing P-selectin labeling was performed using a threshold set obtained with platelets treated only with antimouse FITC-conjugate IgG. Platelet mean P-selectin label- ing was expressed as platelet mean fluorescence in arbitrary units.
Mac-l or MoAb 24 epitope expression on PMN was evaluated before and after mixing with platelets in standard conditions (except for the concentration of Mg” in the medium, which was 1.8 mmoU L). The interaction was stopped by addition of 1 volume of PFA 2%. Fixed PMN ( 2 X lO‘/lOO pL) were incubated with FITC-conju- gate anti-Mac-l antibody (Sigma), with an irrelevant FITC-conju- gate antimouse IgG (control) or MoAb 24 (hybridoma cell line super- natant 1:50 final dilution) for 30 minutes at room temperature. The latter was followed by FITC conjugate antimouse IgG. Samples were then washed twice and resuspended in HEPES-Tyrode buffer before flow cytometric analysis. Determination of the percentage of PMN labeling with MoAb 24 was performed using a threshold set obtained with PMN treated with the irrelevant FITC-labeled antibody. Label- ing with MoAb 24 and Mac-l was evaluated as mean fluorescence intensity in arbitrary units and reported as percentage of the value expressed by PMN alone.
Double color cytojluorimetric assay of platelet-PMN adhesion. Double color cytofluorimetric assay was used extensively to investi- gate cell-cell adhesion of mixed cell populations in suspension. In particular, it was used to study the mechanisms regulating homolo- gous PMN aggregat i~n,~~ as well as heterologous PMN-endothelial
The previously described meth~dology’~ was adapted to our ex- perimental conditions. Briefly, BCECF-loaded platelets ( 108/mL), HE-PMN (107/mL) were incubated in a final volume of 500 pL in siliconized glass tubes (internal diameter 6 mm; ChronoLog, Mascia Brunelli, Milano, Italy). The tubes were placed in an aggregometer (Platelet Ionized Calcium Aggregometer, PICA, ChronoLog, Mascia Brunelli) at 37°C with stimng (1,000 rpm) obtained by an iron bar (4 mm long) rotating under a magnetic field. Although the shear rate produced by this stirring speed cannot be precisely quantified, it should approximate 250 seconds”.36 The interaction was stopped at different times by addition of 1 volume of PFA 2%. Samples were kept at 4°C in the dark and analyzed by flow cytometly within 1 hour. When PMN were challenged with fMLP or PMA (both 100
or PMN-~latelet~~ adhesion.
nmoVL), stimuli were added to mixed cell populations preincubated at 37°C in stimng for 2 minutes (standard condition). In some experi- ments, PMN were activated with fMLP (100 nmol/L) 10 minutes before mixing with platelets.
For blocking studies, antibodies and peptides were added to the desired cell fraction for 10 minutes before coincubation. Protein kinase inhibitors or the diluent (DMSO) were preincubated with PMN 2 minutes before mixing with platelets.
Flow cytometry. The fluorescence on the leukocytes was ana- lyzed by a FACStar flow cytometer (Becton Dickinson) with an argon-ion laser operated at 400 mW output power. Forward and side scatter, as well as green (FITC) and red (PE) signals, were acquired by logarithmic amplification with excitation wavelength of 488 nm and emission wavelengths of 530130 nm and 585/42 nm, respec- tively. Electronic compensation was used to remove spectral overlap. Acquisition and processing of data from 5,000 cells was performed with a Consort 30 Program (Becton Dickinson) on a Hewlett Packard 300 computer (Milano, Italy). PMN were identified on the basis of forward and side scatter alone or in combination with the specific red fluorescent marker. Gating on events identified as PMN was performed to exclude single platelets. For each experiment, a sample in which resting or thrombin-activated platelets were mixed with PMN in the presence of 10 mmoVL EGTA was used to set a thresh- old on the green fluorescence scale (90% of events below the thresh- old) to identify PMN showing the platelet green marker fluorescence. The percentage of PMN showing the platelet marker, ie, above the threshold, represents the percentage of PMN binding platelets [PMN (+) %l.
Platelet adhesion to PMN was also quantified by evaluating the relative number of platelets bound to 100 PMN (PLT/100 PMN) in the following manner. For each experiment, a sample of resting and activated BCECF-labeled platelets was analyzed in the platelet size range using a narrow forward scatter gate. The mean fluorescence of the platelet population corresponds to the standard fluorescence of a single platelet. The mean platelet marker fluorescence of the positive PMN population divided by the standard fluorescence of a single platelet indicates the number of platelets per single PMN positive event. The latter value multiplied by the percentage of posi- tive PMN yielded a semiquantitative estimate of the number of platelets adherent to 100 PMN.
It is important to note that differences in the fluorescence intensity of the platelet marker between different experiments do not affect the determination of the percentage of positive PMN, as well as the relative number of platelets per 100 PMN.
Electron microscopy study. PFA-fixed platelets were incubated with PMN in standard conditions for different times in the absence or presence of fMLP or PMA (both 100 nmoVL). The reaction was stopped by addition of 1 volume of glutaraldehyde 2% and samples fixed for 1 hour at room temperature. After fixation, the supernatant was removed to eliminate unbound platelets. The pellet containing PMN with adherent platelets was processed for electron microscopy study. Briefly, the samples were postfixed in 1% OsOd for 1 hour, stained and blocked with 2% aqueous uranyl acetate (UA) in veronal- acetate buffer (pH 6.0) for 1 hour, dehydrated, and embedded in Epon 81 2. Ultrathin sections were stained with UA and lead citrate and examined in a Zeiss-109 electron microscope.
RESULTS
Activated platelet/PMN adhesion: Description of the phenomenon. Figure 1 shows the contour graphs of PMN stained with HE (A), of resting (B) or activated (C) BCECF-loaded platelets. Figure 1D through G shows the contour graphs of gated HE-stained PMN after coincuba-
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

4186
lo00 1
- 10 100 1000
PLTI100 PMN=16.8 PMN(+)%=13.2
10 100 1000
GREEN FLUORESCENCE (PLT)
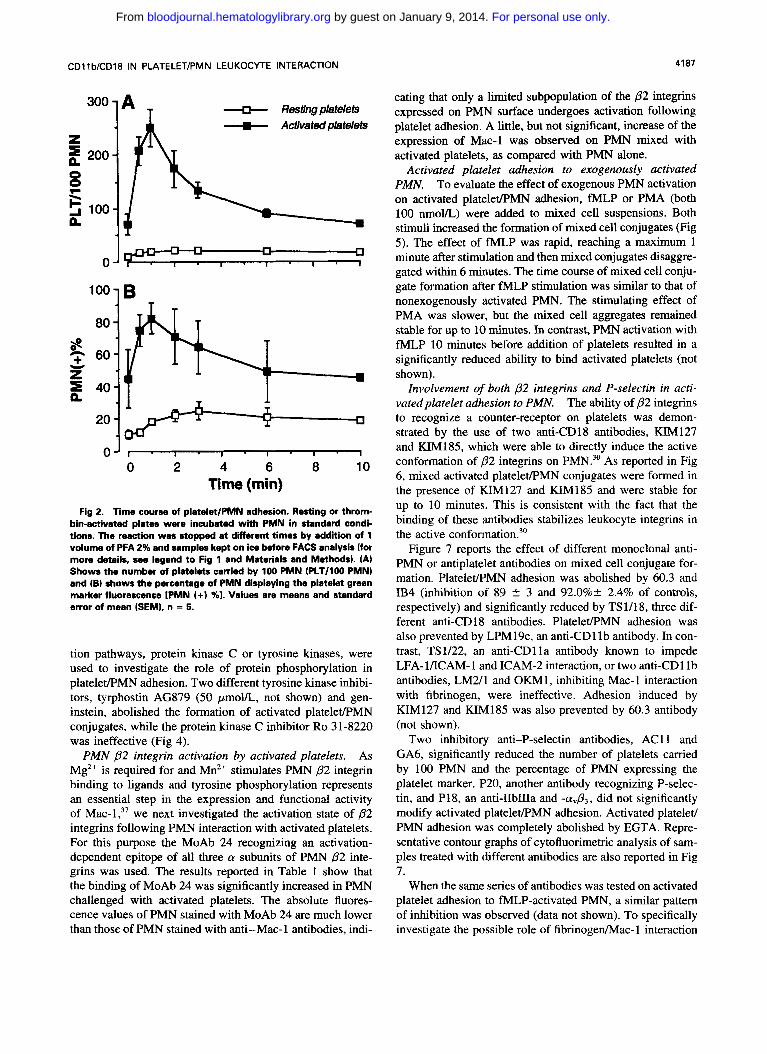
Fig 1. Representative contour graphs of BCECF-loaded platelets (PLT) and HE-loaded PMN. HE-loaded PMN (A), thrombin-activated (B), or resting (C) BCECF-loaded platelets, alone or combined at a plate1ets:PMN ratio of 101 ID through G) in the presence ID and El or absence (F and G) of 10 mmol/L EGTA, were incubated at 37°C at 1,000 rpm stirring for 1 minute. The reaction was stopped by 1% PFA and samples analyzed in a FACS scan flow cytometer (standard conditions). In mixed cell population (D through G) PMN were identi- fied on the basis of forward and side scatter. Gating on events identi- fied as PMN was performed to exclude single platelets from data acquisition. Five thousand PMN-gated events were measured for each sample. For each experiment, the sample in which cells were coincubated in the presence of EGTA was used to set a threshold on green fluorescence to identify PMN showing the platelet marker fluorescence. The percentage of PMN above the threshold represents the percentage of PMN binding platelets IPMN (+) %I. The Jemiquan- titative estimate of the mean number of platelets carried by 100 PMN IPLT/lOO PMN) was obtained by dividing the mean green fluores- cence intensity of the positive PMN population by the mean green fluorescence intensity of the platelet population and muitiplying the resulting value by the percentage of positive PMN (for details see Materials and Methods).
EVANGELISTA ET AL
tion with activated (D and F) or resting (E and G) platelets in the presence (D and E) or absence (F and G) of EGTA. The vertical line represents the threshold on the x-axis (green fluorescence) set on samples in which activated or resting platelets were mixed with PMN in the presence of 5 mmol/L EGTA for 1 minute; about 10% of the events identified as PMN (red fluorescent population) were on the right of the threshold in EGTA-treated samples. The positive PMN population carried no more than one platelet per PMN (D and E). When activated platelets were mixed with PMN in the absence of EGTA, 70% of the latter became positive and the calculated number of platelets carried by 100 PMN corresponded to 270 (F). A negligible increase of mixed cell conjugates formation could be ob- served in resting platelet/PMN population: 16% of PMN carried the platelet-dependent green marker with about I3 platelets per 100 PMN (G).
The formation of activated platelet/PMN mixed conju- gates rapidly increased following initiation of the stirring, reached the maximum around 1 minute and slowly reversed towards the basal level, resting platelet/PMN interaction be- ing negligible (Fig 2).
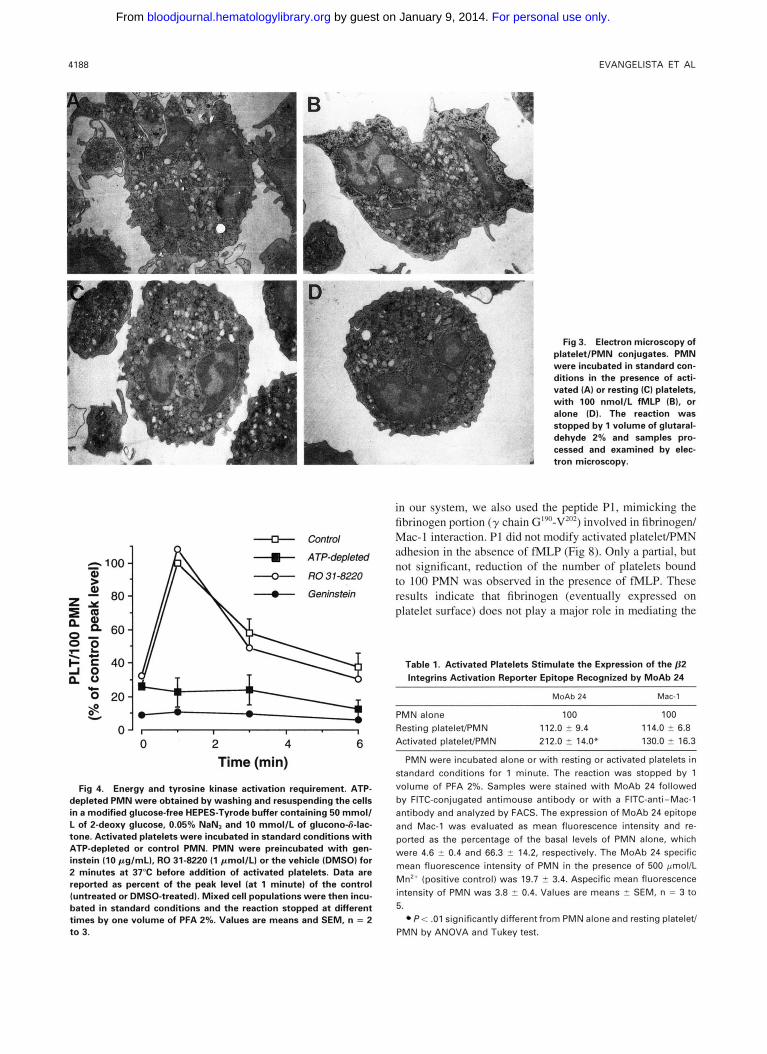
Electron microscopy examination (Fig 3) showed that PMN carrying activated platelets adherent on the surface (A) undergo morphological changes similar to those occurring in fMLP-activated PMN (B). In both conditions, PMN had irregular shape with a number of extended pseudopodes. In contrast, PMN mixed with resting platelets (C) were round in shape and without long protrusions, appearing morpholog- ically similar to PMN incubated in the absence of platelets (D). These results suggest that activated platelets trigger an activating signal in adherent PMN, resulting in a morpholog- ical change of these cells.
Calcium and magnesium requirement and stimulation by manganese. Activated platelet adhesion to PMN could be observed when both Ca” and Mgz+ (both 1 mmol/L) were present in the medium (data not shown). However, in the absence of Ca2’ or Mg2’, addition of Mn2’ ( I mmol/L) induced the formation of mixed cell conjugates (125.3 i 14.7 and 9.3 ? 1.3 platelets/100 PMN with 43.7 ? 7.0 and 7.8% ? 0.2% positive PMN, in the presence or absence of Mn”, respectively at 1 minute after initiation of the stirring). Mixed cell conjugates induced by Mn2’ steadily increased for up to 10 minutes, indicating that this cation may stabilize in an active conformation the adhesion molecule(s) involved. In these experiments, the combined addition of the different cations also showed that Mnz’-induced mixed cell conjugate formation was more rapid in the presence (273 ? 34 plate- lets/100 PMN with 68.9% ? 1.7% positive PMN at 1 minute) than in the absence of Ca2+ (see above). The presence of Mg2+ did not modify the effect of Mn”.
Energy and tyrosine kinase activation requirement h?: PMN. ATP-depleted PMN (see methods) were used to evaluate whether PMN activation was required for optimal adhesion. ATP-depleted PMN were virtually unable to inter- act with activated platelets (Fig 4), suggesting that a meta- bolic step is essential for adhesion.
Different inhibitors of the two main protein phosphoryla-
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom
CDllb/CD18 IN PLATELET/PMN LEUKOCYTE INTERACTION 41 81
z 200
c n 13 100
-0- Resting platelets
80 1 40 n
*O 1 o J I . I - I - I - I ' 1
0 2 4 6 8 10 Time (min)
Fig 2. Time course of platelet/PMN adhesion. Resting or throm- bin-activated plates were incubated with PMN in standard condi- tions. The reaction was stopped at ditferent times by addition of 1 volume of PFA 2% and samples kept on ice before FACS analysis (for more details, see legend to Fig l and Materials and Methods). (A) Shows the number of platelets carried by 100 PMN (PLT/lOO PMN) and (B) shows the percentage of PMN displaying the platelet green marker fluorescence IPMN I+) %l. Values are means and standard error of mean (SEMI, n = 5.
tion pathways, protein kinase C or tyrosine kinases, were used to investigate the role of protein phosphorylation in platelet/PMN adhesion. Two different tyrosine kinase inhibi- tors, tyrphostin AG879 (50 pmol/L, not shown) and gen- instein, abolished the formation of activated platelet/PMN conjugates, while the protein kinase C inhibitor Ro 3 1-8220 was ineffective (Fig 4).
PMN p2 integrin activation by activated platelets. As Mg2+ is required for and Mn2' stimulates PMN p 2 integrin binding to ligands and tyrosine phosphorylation represents an essential step in the expression and functional activity of Mac-1;' we next investigated the activation state of p 2 integrins following PMN interaction with activated platelets. For this purpose the MoAb 24 recognizing an activation- dependent epitope of all three a subunits of PMN p2 inte- grins was used. The results reported in Table 1 show that the binding of MoAb 24 was significantly increased in PMN challenged with activated platelets. The absolute fluores- cence values of PMN stained with MoAb 24 are much lower than those of PMN stained with anti-Mac-l antibodies, indi-
cating that only a limited subpopulation of the p2 integrins expressed on PMN surface undergoes activation following platelet adhesion. A little, but not significant, increase of the expression of Mac-l was observed on PMN mixed with activated platelets, as compared with PMN alone.
Activated platelet adhesion to exogenously activated PMN. To evaluate the effect of exogenous PMN activation on activated platelet/PMN adhesion, N L P or PMA (both 100 nmol/L) were added to mixed cell suspensions. Both stimuli increased the formation of mixed cell conjugates (Fig 5). The effect of fMLP was rapid, reaching a maximum l minute after stimulation and then mixed conjugates disaggre- gated within 6 minutes. The time course of mixed cell conju- gate formation after N L P stimulation was similar to that of nonexogenously activated PMN. The stimulating effect of PMA was slower, but the mixed cell aggregates remained stable for up to 10 minutes. In contrast, PMN activation with N L P 10 minutes before addition of platelets resulted in a significantly reduced ability to bind activated platelets (not shown).
Involvement of both p2 integrins and P-selectin in acti- vatedplatelet adhesion to PMN. The ability of p2 integrins to recognize a counter-receptor on platelets was demon- strated by the use of two anti-CD18 antibodies, KIM127 and KIM185, which were able to directly induce the active conformation of p2 integrins on PMN.30 As reported in Fig 6, mixed activated platelet/PMN conjugates were formed in the presence of KIM127 and KIM185 and were stable for up to 10 minutes. This is consistent with the fact that the binding of these antibodies stabilizes leukocyte integrins in the active conformation?'
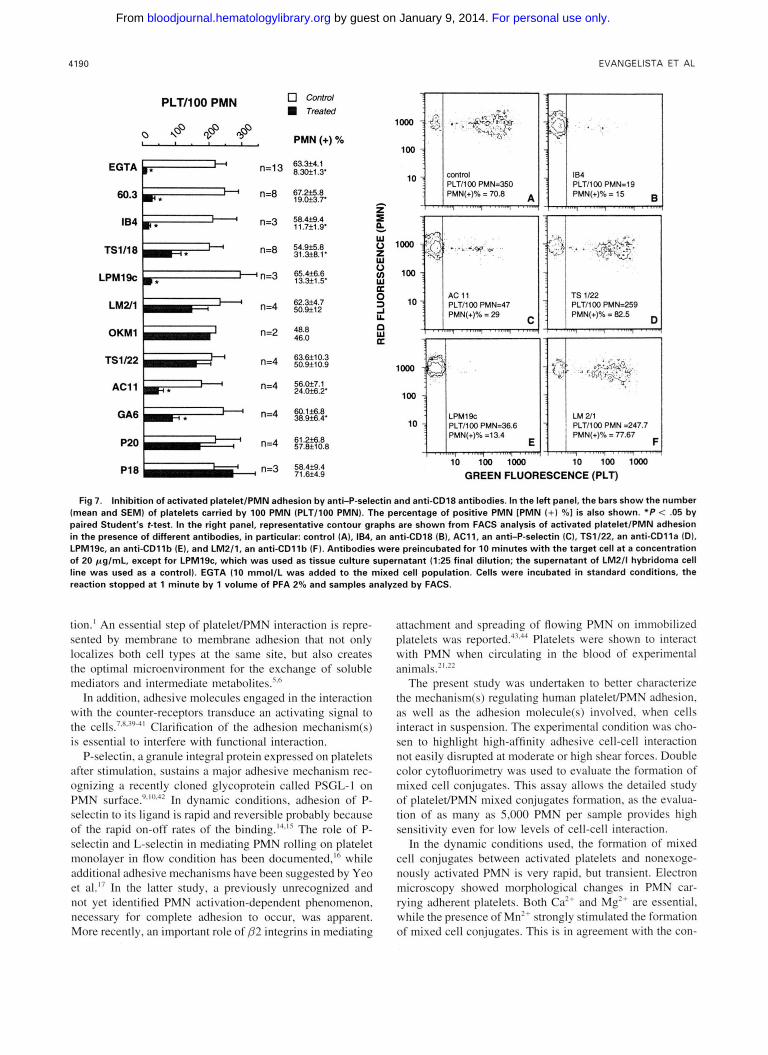
Figure 7 reports the effect of different monoclonal anti- PMN or antiplatelet antibodies on mixed cell conjugate for- mation. PlateleeMN adhesion was abolished by 60.3 and IB4 (inhibition of 89 2 3 and 92.0%+ 2.4% of controls, respectively) and significantly reduced by TS1/18, three dif- ferent anti-CD 18 antibodies. PlateleWMN adhesion was also prevented by LPM19c, an anti-CD1 l b antibody. In con- trast, TS1/22, an anti-CD1 l a antibody known to impede LFA-l/ICAM-1 and ICAM-2 interaction, or two anti-CD1 lb antibodies, LM2/1 and OKM1, inhibiting Mac-l interaction with fibrinogen, were ineffective. Adhesion induced by KIM127 and KIM185 was also prevented by 60.3 antibody (not shown).
Two inhibitory anti-P-selectin antibodies, AC11 and GA6, significantly reduced the number of platelets carried by 100 PMN and the percentage of PMN expressing the platelet marker. P20, another antibody recognizing P-selec- tin, and P18, an anti-IIbIIIa and -a&, did not significantly modify activated platelet/PMN adhesion. Activated platelet/ PMN adhesion was completely abolished by EGTA. Repre- sentative contour graphs of cytofluorimetric analysis of sam- ples treated with different antibodies are also reported in Fig 7.
When the same series of antibodies was tested on activated platelet adhesion to NLP-activated PMN, a similar pattern of inhibition was observed (data not shown). To specifically investigate the possible role of fibrinogenhlac-l interaction
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom
4188 EVANGELISTA ET AL
Fig 3. Electron microscopy of platelet/PMN conjugates. PMN were incubated in standard con- ditions in the presence of acti- vated (A) or resting (C) platelets, with 100 nmol/L fMLP (B), or alone (D). The reaction was stopped by 1 volume of glutaral- dehyde 2% and samples pro- cessed and examined by elec- tron microscopy.
in our system, we also used the peptide P1, mimicking the fibrinogen portion ( y chain CJ'~-V'"') involved in fibrinogen/
Control Mac-l interaction. P1 did not modify activated platelet/PMN adhesion in the absence of fMLP (Fig 8). Only a partial, but not significant, reduction of the number of platelets bound
R03"822CJ to 100 PMN was observed in the presence of fMLP. These - Genhstein results indicate that fibrinogen (eventually expressed on platelet surface) does not play a major role in mediating the
\ T
T U v
0 2 4 6 l ime (min)
Fig 4. Energy and tyrosine kinase activation requirement. ATP- depleted PMN were obtained by washing and resuspending the cells in a modified glucose-free HEPES-Tyrode buffer containing 50 mmoll L of 2-deoxy glucose, 0.05% NaN, and 10 mmol/L of glucono4-lac- tone. Activated platelets were incubated in standard conditions with ATP-depleted or control PMN. PMN were preincubated with gen- instein ( l 0 pg/mL), R 0 31-8220 (1 pmol/L) or the vehicle (DMSO) for 2 minutes at 37°C before addition of activated platelets. Data are reported as percent of the peak level (at 1 minute) of the control (untreated or DMSO-treated]. Mixed cell populations were then incu- bated in standard conditions and the reaction stopped at different times by one volume of PFA 2%. Values are means and SEM, n = 2 t o 3.
Table 1. Activated Platelets Stimulate the Expression of the p2 lntegrins Activation Reporter Epitope Recognized by MoAb 24
MoAb 24 Mac-l
PMN alone 100 100
Resting platelet/PMN 112.0 2 9.4 114.0 -C 6.8
Activated platelet/PMN 212.0 z 14.0* 130.0 2 16.3
PMN were incubated alone or with resting or activated platelets in standard conditions for 1 minute. The reaction was stopped by 1 volume of PFA 2%. Samples were stained with MoAb 24 followed by FITC-conjugated antimouse antibody or with a FITC-anti-Mac-l antibody and analyzed by FACS. The expression of MoAb 24 epitope and Mac-l was evaluated as mean fluorescence intensity and re- ported as the percentage of the basal levels of PMN alone, which were 4.6 2 0.4 and 66.3 2 14.2, respectively. The MoAb 24 specific mean fluorescence intensity of PMN in the presence of 500 pmol/L Mn2' (positive control) was 19.7 f 3.4. Aspecific mean fluorescence intensity of PMN was 3.8 5 0.4. Values are means 2 SEM, n = 3 to 5.
P < .01 significantly different from PMN alone and resting platelet/ PMN by ANOVA and Tukey test.
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

CDllbKD18 IN PLATELET/PMN LEUKOCYTE INTERACTION 4189
l A
n
1 OJT I I I I
100
80
60
40
B
2ol 0 1 . , . , . , . , 0 2 4 6 8
Time (min)
Fig 5. PMN stimulation increases adhesion with activated plate- lets: different effects of fMLP and PMA. Activated platelets and PMN were incubated in standard conditions. The stimuli, fMLP or PMA (both 100 nmol/L), or the vehicle were added after 2 minutes of preincubation. The reaction was stopped before and at different times after stimulation by 1 volume of PFA 1% and samples analyzed by FACS. Data are means and SEM, n = 2 t o 5.
82 integrin-dependent platelet/PMN adhesion in our experi- mental conditions.
82 Integrin counter-receptor presence on resting plate- lets. Further experiments were performed to evaluate whether the 82 integrin counter-receptor on platelets requires platelet activation or is constitutively present on platelet membrane.
PFA-fixed or unfixed resting platelets did not significantly bind resting PMN. On the other hand, resting platelets were significantly bound to fMLP-activated PMN, although the number of platelets carried by PMN was five to six times lower with respect to activated platelets (Fig 9). Electron microscopy observation of these samples showed a tight membrane to membrane contact in the formation of mixed aggregates between activated PMN and fixed resting plate- lets, ruling out the possibility of a passive entrapment of platelets within PMN adherent to each other (not shown).
When PMA was used as a stimulus, PMN bound a number of fixed resting platelets similar to that of fixed activated platelets, showing also a similar time course (Fig 9). PMA
did not modify P-selectin expression of fixed resting platelets (not shown), excluding a role for P-selectin in this condition.
The formation of resting platelet/fMLP-activated PMN mixed conjugates was abolished by anti-CD18 antibodies, while it was unaffected by anti-P-selectin antibodies (Fig IO), reinforcing the concept that P-selectin did not play any role in this condition.
Treatment of PMN with the activating antibodies KIM 127 and KIM185 induced a platelet binding capacity for fixed, as well as unfixed, resting washed platelets indicating that the p2 integrin counter-receptor is constitutively present on the platelet surface (not shown). This result also allows us to exclude that leukocyte integrin recognizes platelet surface protein(s) after PFA fixation.
DISCUSSION
The interaction between platelets and PMN at the site of vascular damage or inflammatory reaction may promote thrombosis and inflammation. In vitro studies demonstrated a complex cross talk between platelets and PMN resulting in reciprocal functional modulation3* and metabolic coopera-
800
z h 600
0
- c o n f r d - L M a l KIM 127
i" l"
80 F====
7 8 5
0 2 4 6 8 1 0 Time (min)
Fig 6. Stimulation of activated platelet adhesion to PMN by two activating anti-CD18 antibodies. Activated plateletlPMN mixed cell populations were incubated in standard conditions in the absence (control) or presence of20 pg/mLof KIM127, KIMl85, or LM2/1 Inega- tive control). The reaction was stopped at different tlmes by 1 volume of PFA 2% and samples analyzed by FACS. The data are from a single experiment representative of three dinerent experiments.
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom
4190 EVANGELISTA ET AL
PLT1100 PMN
EGTA b. l+ 63.3i4.1 8.3of1.3'
60.3 p* n=8 67.255.8 19.Oi3.7
n=3 58.4f9.4 11.7fl .g'
TS1118 m* H n=8 54.9s. 31.3f8.1
n=3 65.4f6.6 LPMlsC m * 13.3f1.5'
LMUl
OKMl ? n=2 48.8 46.0
n=4 62.3f4.7 4 50.9.112
Ts1122 4 n=4 50.9.110.9 63.6t10.3
AC11 m. - n=4 56.W7.1 24.053.2'
GA6 m. +-l n=4 60.135.8. 38.M.4
P20 " ,,=4 61.2i6.8 57.8f10.8
P1 8 n=3 58.4f9.4 71.M4.9
P k W 0 Z W V U) W
0 3 -l L
W Le
a
n
loo0
loo
10
TS 1/22 PLT/lOO PMN=259 PMN(+)% = 82.5
, . . ......, . . ......, . . ......, . . .... , ,
LPMlSc PLT/IOO PMN.36.6 PMN(+)X =13.4
10 loo loo0
PMN(+)% = 77.67
10 loo loo0 GREEN FLUORESCENCE (PLT)
Fig 7. Inhibition of activated platelet/PMN adhesion by anti-P-selectin and anti-CD18 antibodies. In the left panel, the bars show the number (mean and SEMI of platelets carried by 100 PMN (PLT/lOO PMN). The percentage of positive PMN [PMN (+) 961 is also shown. " P < .05 by paired Student's t-test. In the right panel, representative contour graphs are shown from FACS analysis of activated plateletlPMN adhesion in the presence of different antibodies, in particular: control (A), 184. an anti-CD18 (B), AC11, an anti-P-selectin (C), TS1/22, an anti-CDlla ID). LPMlSc, an anti-CDllb (E), and LM2/1, an anti-CDllb (F). Antibodies were preincubated for 10 minutes with the target cell at a concentration of 20 pg/mL, except for LPM19c. which was used as tissue culture supernatant (1:25 final dilution; the supernatant of LM2/1 hybridoma cell line was used as a control). EGTA (10 rnmol/L was added to the mixed cell population. Cells were incubated in standard conditions, the reaction stopped at 1 minute by 1 volume of PFA 2% and samples analyzed by FACS.
tion.' An essential step of platelet/PMN interaction is repre- sented by membrane to membrane adhesion that not only localizes both cell types at the same site, but also creates the optimal microenvironment for the exchange of soluble mediators and intermediate metabolites.'."
In addition, adhesive molecules engaged in the interaction with the counter-receptors transduce an activating signal to the cell^.^^^^^"^^' Clarification of the adhesion mechanism(s) is essential to interfere with functional interaction.
P-selectin, a granule integral protein expressed on platelets after stimulation, sustains a major adhesive mechanism rec- ognizing a recently cloned glycoprotein called PSGL-I on PMN In dynamic conditions, adhesion of P- selectin to its ligand is rapid and reversible probably because of the rapid on-off rates of the binding.".l5 The role of P- selectin and L-selectin in mediating PMN rolling on platelet monolayer in flow condition has been documented,'" while additional adhesive mechanisms have been suggested by Yeo et al.'' In the latter study. a previously unrecognized and not yet identified PMN activation-dependent phenomenon, necessary for complete adhesion to occur, was apparent. More recently, an important role of p2 integrins in mediating
attachment and spreading of flowing PMN on immobilized platelets was reported..'3.'4 Platelets were shown to interact with PMN when circulating in the blood of experimental animals."."
The present study was undertaken to better characterize the mechanism(s) regulating human platelet/PMN adhesion. as well as the adhesion molecule(s) involved, when cells interact in suspension. The experimental condition was cho- sen to highlight high-affinity adhesive cell-cell interaction not easily disrupted at moderate or high shear forces. Double color cytofluorimetry was used to evaluate the formation of mixed cell conjugates. This assay allows the detailed study of platelet/PMN mixed conjugates formation. as the evalua- tion of as many as 5.000 PMN per sample provides high sensitivity even for low levels of cell-cell interaction.
In the dynamic conditions used, the formation of mixed cell conjugates between activated platelets and nonexoge- nously activated PMN is very rapid. but transient. Electron microscopy showed morphological changes in PMN car- rying adherent platelets. Both Ca" and Mg" are essential, while the presence of Mn" strongly stimulated the formation of mixed cell conjugates. This is in agreement with the con-
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

CDllb/CD18 IN PLATELET/PMN LEUKOCYTE INTERACTION 4191
v! S
In
Z 200- n
c a
0 0
3 100-
0- cu 6
1 b B : +I
m (d +I m ci b
500 - 400- I
n 0 300- 0
5 200- 7
n 100 -
n- ., Control P1 RGDS peptide
Fig 8. Inefficacy of fibrinogen-binding inhibitor peptides. The pep- tides RGD 11,200 pmol/Ll and P1 1400 pmol/LI or control peptide (400 pmol/L) were preincubated with PMN for 10 minutes before addition of activated platelets in the absence of (AI or presence (B1 of fMLP. Combined cell suspensions were mixed in standard condi- tion for 1 minute. Reaction was stopped by 1 volume of PFA 2% and samples analyzed by FACS. The bars show the number of platelets carried by 100 PMN 1PLT/100 PMN). The percentage of positive PMN are also shown on top of the bars (mean 2 SEM, n = 41.
cept that selectins require Ca”, while Mg” is required for and Mn” stimulates integrin adhesion to their ligands.
PlateleVPMN optimal adhesion requires an intact tyrosine kinase activity. Recently Naccache et al” presented evidence that tyrosine phosphorylation regulates the expression and functional activity of the leukocyte integrin Mac-l.
The increased binding of MoAb 24 following PMN inter- action with activated platelets, without comparable increase in Mac-l expression, indicates that adherent platelets induce a qualitative change of PMN 02 integrins. rather than an increased surface expression. In addition, we found that ex- ogenous stimulation of PMN increased the formation of mixed conjugates: the observed kinetics of activated platelet adhesion to fMLP- or PMA-stimulated PMN correlated with the reported kinetics of 02 integrins activatiomJS
1000 1200 1 A z g n 8004
8 6004 F \ 5 400 n 3
-[3- Control + fMLP - pMA /
/
l o o 1 80 B
0’ I I I I 1
0 2 4 6 8 Time (min)
Fig 9. PMN stimulation induces adhesion with resting platelets: different effects of fMLP and PMA. Resting platelets were incubated with PMN for 2 minutes before addition of fMLP 1100 nmollL), PMA (100 nmol/L) or vehicle (control). Reaction was stopped at different times by 1 volume PFA 2% and samples analyzed by FACS In = 2 to 5).
60.3 v n=8 52.6f4.13 13.3k3.6‘
AC11 1 ,,=3 47.1f3.4
9.2-2
Fig 10. Inhibition of resting platelet adhesion to fMLP-activated PMN by anti-CD18, but not anti-P-selectin antibodies. Antibodies (20 pglmL1 were preincubated for 10 minutes with the target cell. Mixed cell populations were incubated for 2 minutes in standard conditions before addition of fMLP 1100 nmol/L). The reaction was stopped by 1 volume of PFA 2% 1 minute after stimulation and samples analyzed by FACS. The bars show the number (mean and SEMI of platelets carried by 100 PMN IPLT/lOO PMN). The percentage of positive PMN IPMN ( + l 461 is also shown. ‘ P < .05 by paired Student’s t-test.
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

4192 EVANGELISTA ET AL
Taken together, these data strongly suggest that 02 inte- grins were involved in activated platelet adhesion to PMN. Consistently, direct activation of /32 integrins induced by the activating anti-CD18 antibodies, KIM127 and KIM185, was able to stimulate the formation of activated pIatelet/PMN mixed conjugates stable for up to 10 minutes. Experiments with a panel of different antibodies showed an interesting pattern of inhibition. In fact, two different anti-CD18, as well as an anti-CD1 1 b inhibitory antibody abolished, as expected, activated platelet adhesion to nonexogenously activated PMN; but also two different anti-P-selectin antibodies re- duced by about 70% the participation of platelets to mixed cell conjugates formation. A similar inhibition was obtained with fMLP-stimulated PMN indicating that also in this case both adhesive mechanisms are required for optimal activated platelet/PMN adhesion.
Although alternative possibilities including negative sig- naling by the anti-CD18 antibodies46 cannot be completely excluded, the inhibition obtained with the anti-CD1 Ib anti- body, LPM19c, is suggestive of the direct engagement of this integrin in the adhesive event.
Because the direct binding between P-selectin and p 2 inte- grins can be excluded," it is reasonable to speculate that a two-step model takes place during activated platelet/PMN interaction: a first P-selectin-dependent (PMN activation- independent) step tethering platelets on PMN surface, fol- lowed by tyrosine kinase-dependent p 2 integrins activation, which subsequently results in stable cell-cell interaction.
Recently, it was reported that PMN activation reduces the P-selectin-dependent platelet binding.47 Subsequently, Lorant et a148 demonstrated that following activation, PSGL- 1 is redistributed on the surface of PMN with a consequent reduced binding capacity for P-selectin. This phenomenon probably also takes place in our system, but it does not explain the time course of activated platelet/PMN interac- tion. In fact, if disaggregation of mixed cell conjugates was only a consequence of the release of P-selectin following PSGL- 1 redistribution, one should expect that the metabolic inhibition of PMN would prevent it, allowing the formation of stable mixed cell conjugates. Because this was not the case, we can conclude that /32 integrins activation and inacti- vation is the major determinant of the time course of mixed- cell conjugate formation in our experimental condition.
This study extends to the PMN p2 integrin CD1 1 b/CD 18 the role previously ascribed mainly to P-selectin. The previ- ous studies on PMN-platelet adhesion were performed in static conditions in which the adhesive P-selectin-PSGLI interaction is probably more stable and at room tempera- ture"." or lower,IX which may dampen leukocyte activation and p 2 integrins function.49 Indeed, in the experimental con- ditions used in the present study, in particular the high shear rate and the physiological temperature, the role of p 2 inte- grins in mediating platelet/PMN adhesion, resistant to shear forces, was magnified.
We performed experiments aimed at characterizing the possible 02 integrin counter-receptor on platelets. Experi- ments using the inhibitory antibody TS1/22'' indicated that LFA-l/ICAM-2 interaction was not playing a major role in mediating the CD18-dependent adhesion in our system.
Fibrinogen is an important adhesion molecule able to bind both activated platelets and PMN, CD 1 1 b/CD 18 and CD 1 1 c/ CD18 being the main fibrinogen receptors on PMN.-5".5' It was already proposed to mediate, at least in part, platelet/ PMN, as well as PMN/endotheliai cell adhesion."-" Al- though fibrinogen may be a potential bridging molecule be- tween platelets and PMN, the inefficacy to reduce plateleu PMN adhesion of the anti-Mac- 1 antibodies, OKM I and LM2/1,53 and of the peptide P1, which mimicks the y chain region mediating the binding to Mac-l,?* indicated that in our experimental conditions, tibrinogedMac- 1 interaction does not seem to play a major role .
The question whether the p 2 integrin counter-receptor on platelets requires platelet activation was addressed with three different experiments, which convincingly indicated that it is constitutively present on platelet surface. In fact, ( 1 ) resting platelets adhered to fMLP-activated PMN, although at an extent about four times lower with respect to activated plate- lets. In contrast, PMA-activated PMN bound resting platelets at a similar extent than activated platelets. This suggests that when p 2 integrin activation has a short half-life, as in the case of fMLP activation, tethering by P-selectin is essential for optimal adhesion. On the other hand, P-selectin would play a minor role when p 2 integrins are strongly activated by PMA. (2) The activating antibodies KIM 127 and KIM I85 stimulated fixed or unfixed resting platelet adhesion to PMN. (3) The major mechanism mediating resting platelet/fMLP- activated PMN adhesion is p 2 integrin-dependent, being that the anti-P-selectin antibodies are completely ineffective in this condition.
Although the counter-receptor of the p2 integrin CD I I b/ CD18 on platelets remains to be identified, our study demon- strates the existence of a previously unrecognized adhesive mechanism playing an essential role in mediating platelet/ PMN adhesion. Similar to PMN/endothelial cell interaction, platelet/PMN adhesion in dynamic conditions can be reason- ably modeled as a multistep adhesion cascade: a first P- selectin-dependent recognition step, followed by the PMN 0 2 integrin CD1 lb/CD18 activation, which in turn, through a subsequent adhesion step, stabilizes platelet/PMN interac- tion. The intermediate functional signal seems to proceed through tyrosine kinase activation and further studies are in progress to clarify this point.
P-selectin might directly induce a functional signal in leu- kocytes. In fact, purified and recombinant P-selectin soluble or expressed by Chinese Hamster Ovary (CH0)-cell was recently shown to activate CD 18-dependent phagocytosis on PMN4" and tissue factor expression in monocytes.x
On the other hand, more recently Weyrich et reported that P-selectin cooperates with another platelet-derived sig- naling factor, the chemokine RANTES, in inducing the pro- duction of monocyte chemotactic protein- 1 and interleukin- 8 by monocytes. Production of chemokines, although not directly induced by P-selectin, requires tethering of the monocytes by this adhesion molecule for their activation by RANTES. On the basis of this observation, it is reasonable to hypothesize that also in inducing PMN responses platelets use a mechanism in which tethering and signaling molecules act in concert.
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

CD1 Ib/CD18 IN PLATELET/PMN LEUKOCYTE INTERACTION 4193
In our study, we did not examine the possibility that other signaling molecules, such as ICAM-255 or fibrinogen ex- pressed by may induce p2 integrin activation. If this is the case, tethering by P-selectin, although not suffi- cient, would remain a necessary step for both functional and adhesive plateletPMN interaction.
Adherence of activated platelets to PMN may be a key event in the sequence of thrombus formation. Similarly, platelet recruitment by activated PMN has an important role in mounting an inflammatory reaction. Some in vivo studies demonstrated that 0 2 integrins blockade significantly ame- liorates PMN-dependent tissue damage in different models of inflammati~n~’.~’ and indicating that leukocyte integrins are potential targets for pharmacological interven- tion. Due to the possible role of platelet/PMN functional interaction in the pathogenesis of both thrombotic and in- flammatory events, the recognition of the essential role of the p2 integrin CD 1 1 b/CD18 also in plateletPMN adhesion provides additional evidence for the broad range of functions and mechanisms in which PMN integrins are involved.
NOTE ADDED IN PROOF
We acknowledge the work of Diacovo et a1 who, using a different experimental model, recently showed that PMN adhesion and transmigration across surface adherent platelets occur via P-selectin and CD1 lb/CD18 (Blood 88: 146, 1996).
ACKNOWLEDGMENT
We thank Dr Licia Totani for fruitful discussion, Antonio Piccoli for help in obtaining TS1/18 and TS1122 from hybridoma cell lines; Lorenzo Naccarella and Rita Sideri for technical assistance; Filo- mena Cinalli, the Gustavus A. Pfeiffer Memorial Library staff, Raf- faella Bertazzi, and Sabrina Menna for their contribution in editing the manuscript and the figures.
REFERENCES
1. Marcus A J : Thrombosis and inflammation as multicellular pro- cesses: Pathophysiologic significance of transcellular metabolism. Blood 76:1903, 1990
2. Cerletti C, Evangelista V, de Gaetano G: Polymorphonuclear leucocyte-dependent modulation of platelet function: Relevance to the pathogenesis of thrombosis. Pharmacol Res 26:261, 1992
3. Bazzoni G, Dejana E, Del Maschio A: Platelet-dependent mod- ulation of neutrophil function. Pharmacol Res 26:269, 1992
4. Jungi TW, Spycher MO, Nydegger UE, Barandun S: Platelet- leukocyte interaction: Selective binding of thrombin-stimulated platelets to human monocytes, polymorphonuclear leukocytes, and related cell lines. Blood 67:629, 1986
5. Evangelista V, Piccardoni P, White JG, de Gaetano G , Cerletti C: Cathepsin G-dependent platelet stimulation by activated polymor- phonuclear leukocytes and its inhibition by antiproteinases: Role of P-selectin-mediated cell-cell adhesion. Blood 81:2947, 1993
6. Maugeri N. Evangelista V, Celardo A, Dell’Elba G, Martelli N, Piccardoni P, de Gaetano G, Cerletti C: Polymorphonuclear leu- kocyte-platelet interaction: Role of P-selectin in thromboxane B2 and leukotriene C, cooperative synthesis. Thromb Haemost 72:450, 1994
7. Nagata K, Tsuji T, Todoroki N, Katagiri Y, Tanoue K, Yama- zaki H, Hanai N, Irimura T: Activated platelets induce superoxide anion release by monocytes and neutrophils through P-selectin (CD62). J lmmunol 151:3267, 1993
8. Celi A, Pellegrini G , Lorenzet R, De Blasi A, Ready N, Furie BC, Furie B: P-selectin induces the expression of tissue factor on monocytes. Proc Natl Acad Sci USA 91:8767, 1994
9. Larsen E, Celi A, Gilbert GE, Furie BC, Erban JK, Bonfanti R, Wagner DD, Furie B: PADGEM protein: A receptor that mediates the interaction of activated platelets with neutrophils and monocytes. Cell 59:305, 1989
IO. Hamburger SA, McEver RP: GMP-I40 mediates adhesion of stimulated platelets to neutrophils. Blood 75:550, 1990
11. Varki A: Selectin ligands. Proc Natl Acad Sci USA 91:7390, 1994
12. Moore KL, Varki A, McEver RP: GMP- 140 binds to a glyco- protein receptor on human neutrophils: Evidence for a lectin-like interaction. J Cell Biol 112:491, 1991
13. Moore K L , Stults NL, Diaz S, Smith DF, Cummings RD, Varki A, McEver R P Identification of a specific glycoprotein ligand for P-selectin (CD62) on myeloid cells. J Cell Biol 1 18:445, 1992
14. Lawrence MB, Springer TA: Leukocytes roll on a selectin at physiologic flow rates: Distinction from and prerequisite for adhe- sion through integrins. Cell 65:859, 1991
15. Alon R, Hammer DA, Springer TA: Lifetime of the P-selec- tin-carbohydrate bond and its response to tensile force in hydrody- namic flow. Nature 374:539, 1995
16. Buttrum SM, Hatton R, Nash GB: Selectin-mediated rolling of neutrophils on immobilized platelets. Blood 82: 1165, 1993
17. Yeo EL, Sheppard J-AI, Feuerstein IA: Role of P-selectin and leukocyte activation in polymorphonuclear cell adhesion to surface adherent activated platelets under physiologic shear conditions (an injury vessel wall model). Blood 83:2498, 1994
18. Spangenberg P, Redlich H, Bergmann I, Losche W, Gotzrath M, Kehrel B: The platelet glycoprotein IIb/IIIa complex is involved in the adhesion of activated platelets to leukocytes. Thromb Haemost 70:5 14, 1993
19. Diacovo TG, deFougerolles AR, Bainton DF, Springer TA: A functional integrin ligand on the surface of platelets: Intercellular adhesion molecule-2. J Clin Invest 94:1243, 1994
20. Carlos TM, Harlan JM: Leukocyte-endothelial adhesion mol- ecules. Blood 84:2068, 1994
21. Lehr H-A, Olofsson AM, Carew TE, Vajkoczy P, von An- drian UH, Hiibner C, Berndt MC, Steinberg D, Messmer K, Arfors KE: P-selectin mediates the interaction of circulating leukocytes with platelets and microvascular endothelium in response to oxidized lipoprotein in vivo. Lab Invest 71:380, 1994
22. Lehr H-A, Frei B, Arfors K-E: Vitamin C prevents cigarette smoke-induced leukocyte aggregation and adhesion to endothelium in vivo. Proc Natl Acad Sci USA 91:7688, 1994
23. Evangelista V, Rajtar G, de Gaetano G, White JG, Cerletti C: Platelet activation by fMLP-stimulated polymorphonuclear leuko- cytes: The activity of cathepsin G is not prevented by antiproteinases. Blood 77:2379, 1991
24. Parmentier S, McGregor L, Catimel B, Leung LLK, McGregor JL: Inhibition of platelet functions by a monoclonal anti- body (LYP20) directed against a granule membrane glycoprotein
25. Palabrica T, Lobb R, Furie BC, Aronovitz M, Benjamin C, Hsu Y-M, Sajer SA, Furie B: Leukocyte accumulation promoting fibrin deposition is mediated in vivo by P-selectin on adherent plate- lets. Nature 359:848, 1992
26. Murphy JF, Bordet J-C, Wyler B, Rissoan M-C, Chomarat P, Defrance T, Miossec P, McGregor JL: The vitronectin receptor (a,&) is implicated, in cooperation with P-selectin and platelet- activating factor, in the adhesion of monocytes to activated endothe- lial cells. Biochem J 304:537, 1994
27. Sanchez-Madrid F, Krensky AM, Ware CF. Robbins E,
(GMP-140PADGEM). Blood 77~1734, 1991
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom

4194 EVANGELISTA ET AL
Strominger JL, Burakoff SJ, Springer TA: Three distinct antigenes associated with human T-lymphocyte-mediated cytolysis: LFA- I , LFA-2, and LFA-3. Proc Natl Acad Sci USA 79:7489, 1982
28. Wright SD, Rao PE, Van Voorhis WC, Craigmyle LS, Iida K, Talle MA, Westberg EF, Goldstein G, Silverstein SC: Identifica- tion of the C3bi receptor on human monocytes and macrophages by using monoclonal antibodies. Proc Natl Acad Sci USA 80:5699, I983
29. Harlan JM, Killen PD. Seneca1 FM, Schwartz BR, Yee EK. Taylor RF, Beatty PG, Price TH, Ochs HD: The role of neutrophil membrane glycoprotein GP-150 in neutrophil adherence to endothe- lium in vitro. Blood 66:167, 1985
30. Robinson MK, Andrew D, Rosen H, Brown D, Ortlepp S, Stephens P, Butcher EC: Antibody against the Leu-CAM P-chain (CD1 8) promotes both LFA-I- and CR3-dependent adhesion events. J Immunol 148: 1080, 1992
3 1 . Cabanas C, Hogg N: Ligand intercellular adhesion molecule 1 has a necessary role in activation of integrin lymphocyte function- associated molecule 1 [published erratum appears in Proc Natl Acad Sci USA 90:8302, 19931. Proc Natl Acad Sci USA 90:5838, 1993
32. Altieri DC, Plescia J, Plow EF: The structural motif glycine 190-valine 202 of the fibrinogen yg chain interacts with CDlIb/ CD18 integrin (aM P2, Mac-l) and promotes leukocyte adhesion. J Biol Chem 268:1847, 1993
33. Simon SI, Chambers JD, Sklar LA: Flow cytometric analysis and modeling of cell-cell adhesive interactions: The neutrophil as a model. J Cell Biol 1 1 1 :2747, 1990
34. Kuijpers TW, Hakkert BC, van Mourik JA, Roos D: Distinct adhesive properties of granulocytes and monocytes to endothelial cells under static and stirred conditions. J Immunol 145:2588, 1990
35. Rinder HM, Bonan JL, Rinder CS, Ault KA, Smith BR: Acti- vated and unactivated platelet adhesion to monocytes and neutro- phils. Blood 78:1760, 1991
36. Remuzzi A, Languino LR, Costantini V, Guardabasso V, de Gaetano G, Dejana E: Platelet adhesion to subendothelium-Effect of shear rate, hematocrit and platelet count on the dynamic equilib- rium between platelets adhering to and detaching from the surface. Thromb Haemost 54:857, 1985
37. Naccache PH, Jean N, Liao NW, Bator JM, McColl SR, Kubes P: Regulation of stimulated integrin surface expression in human neutrophils by tyrosine phosphorylation. Blood 84:616, 1994
38. Cerletti C, Evangelista V, Molino M, de Gaetano G: Platelet activation by polymorphonuclear leukocyte: Role of cathepsin G and P-selectin. Thromb Haemost 74:213, 1995
39. Weyrich AS, McIntyre TM, McEver RP, Prescott SM, Zim- merman GA: Monocyte tethering by P-selectin regulates monocyte chemotactic protein-l and tumor necrosis factor-a secretion. Signal integration and NF-K B translocation [see comments]. J Clin Invest 95:2297, 1995
40. Cooper D, Butcher CM, Berndt MC, Vadas MA: P-selectin interacts with a P2-integrin to enhance phagocytosis. J Immunol 153:3199, 1994
4 I . Berman ME, Muller WA: Ligation of platelet/endothelial cell adhesion molecule 1 (PECAM-IKD31) on monocytes and neutro- phils increases binding capacity of leukocyte CR3 (CD1 Ib/CD18). J Immunol 154:299, 1995
42. Sako D, Chang X-J, Barone KM, Vachino G, White HM, Shaw G, Veldman GM, Bean KM, Ahern TJ, Furie B, Cumming DA, Larsen GR: Expression cloning of a functional glycoprotein ligand for P-selectin. Cell 75:1179, 1993
43. Kuijper PHM, Gallardo Torres HI, van der Linden JAM,
Lammers J-WJ, Sixma JJ, Koenderman L, Zwaginga JJ: Platelet- dependent primary hemostasis promotes selectin- and integrin-medi- ated neutrophil adhesion to damaged endothelium under flow condi- tions. Blood 87:3271, 1996
44. Sheikh S, Nash GB: Continuous activation and deactivation of integrin CD1 Ib/CD18 during de novo expression enables rolling neutrophils to immobilize on platelets. Blood 87:5040, 1996
45. Chatila TA, Geha RS, Arnaout MA: Constitutive and stimu- lus-induced phosphorylation of CD1 I/CD18 leukocyte adhesion molecules. J Cell Biol 109:3435, 1989
46. van Noesel C, Miedema F, Brouwer M, de Rie MA, Aarden LA, van Lier RAW: Regulatory properties of LFA-I a and P chains in human T-lymphocyte activation. Nature 333:850, 1988
47. Rinder HM, Tracey JL, Rinder CS, Leitenberg D, Smith BR: Neutrophil but not monocyte activation inhibits P-selectin-mediated platelet adhesion. Thromb Haemost 71:750, 1994
48. Lorant DE, McEver RP, Mclntyre TM, Moore KL, Prescott SM, Zimmerman GA: Activation of polymorphonuclear leukocytes reduces their adhesion to P-selectin and causes redistribution of ligands for P-selectin on their surfaces. J Clin Invest 96: 171. 1995
49. Cai T-Q, Wright SD: Energetics of leukocyte integrin activa- tion. J Biol Chem 270:14358, 1995
SO. Altieri DC, Agbanyo FR, Plescia J, Ginsberg MH, Edgington TS, Plow EF: A unique recognition site mediates the interaction of fibrinogen with the leukocyte integrin Mac-] (CD1 Ib/CDI8). J Biol Chem 265: I2 1 19, 1990
51. Loike JD, Sodeik B, Cao L, Leucona S, Weitz JI, Detmers PA, Wright SD, Silverstein SC: CD1 Ic/CD18 on neutrophils recog- nizes a domain at the N terminus of the Aa chain of fibrinogen. Proc Natl Acad Sci USA 88:1044, 1991
52. Languino LR, Plescia J, Duperray A, Brian AA, Plow EF. Geltosky JE, Altieri DC: Fibrinogen mediates leukocyte adhesion to vascular endothelium through an ICAM- I -dependent pathway. Cell 73:1423, 1993
53. Zhou L, Lee DHS, Plescia J, Lau CY, Altieri DC: Differential ligand binding specificities of recombinant CD I 1 b/CD18 integrin I- domain. J Biol Chem 269:17075, 1994
54. Weyrich AS, Elstad MR, McEver RP, McIntyre TM, Moore LK, Morrissey JH. Prescott SM, Zimmerman GA: Activated platelets signal chemokine synthesis by human monocytes. J Clin Invest 97:1525, 1996
S S . Li R. Xie J, Kantor C, Koistinen V, Altieri DC, Nortamo P, Gahmberg CG: A peptide derived from the intercellular adhesion molecule-2 regulates the avidity of the leukocyte integrins CD1 1 b/ CD18 and CDIIc/CD18. J Cell Biol 129:1143, 1995
56. Ruf A, Patscheke H: Platelet-induced neutrophil activation: Platelet-expressed fibrinogen induces the oxidative burst in neutro- phils by an interaction with CDllc/CD18. Br J Haematol 90:791. 1995
57. Arfors K-E, Lundberg C, Lindbom L, Lundberg K, Beatty PG, Harlan JM: A monoclonal antibody to the membrane glycoprotein complex CD18 inhibits polymorphonuclear leukocyte accumulation and plasma leakage in vivo. Blood 69:338, 1987
58. Vedder NB, Winn RK, Rice CL, Chi EY, Arfors K-E. Harlan JM: Inhibition of leukocyte adherence by anti-CD18 monoclonal antibody attenuates reperfusion injury in the rabbit ear. Proc Natl Acad Sci USA 87:2643, 1990
59. Simpson PJ, Todd 111 RF, Fantone JC, Mickelson JK, Griffin JD, Lucchesi BR: Reduction of experimental canine myocardial re- perfusion injury by a monoclonal antibody (anti-Mol, anti-CD1 Ib) that inhibits leukocyte adhesion. J Clin Invest 81:624, 1988
For personal use only. by guest on January 9, 2014. bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN




















