
Planning for the persistence of river biodiversity: exploring alternative futures using...
18
Planning for the persistence of river biodiversity: exploring alternative futures using process-based models EREN TURAK*, SIMON FERRIER* ,† , TOM BARRETT ‡ , EDWINA MESLEY*, MICHAEL DRIELSMA ‡ , GLENN MANION ‡ , GAVIN DOYLE § , JANET STEIN – AND GEOFF GORDON* *Department of Environment, Climate Change and Water, Sydney NSW, Australia † CSIRO Entomology, Canberra, ACT, Australia ‡ Department of Environment, Climate Change and Water, Armidale NSW, Australia § Hunter-Central Rivers CMA, Paterson NSW, Australia – The Fenner School of Environment and Society, Australian National University, Canberra, ACT, Australia SUMMARY 1. Planning for the conservation of river biodiversity must involve a wide range of management options and account for the complication that the effects of many actions are spatially removed from these actions. Reserve design algorithms widely used in conservation planning today are not well equipped to address such complexities. 2. We used process-based models to estimate the expected persistence of river biodiversity under alternative catchment-wide management scenarios and applied it in the Hunter Region (37 000 km 2 ) in southeastern Australia. 3. The biological condition of 12 197 subcatchments was estimated using a multiple linear regression model that related assessments of the integrity of macroinvertebrate assem- blages to human-induced disturbances at river sites. The best-fit model (R 2 = 0.76) used measures of both local and catchment-wide disturbances as well as elevation and distance from source as predictor variables. Based on the outputs of this model, we estimated that substantial loss of river biodiversity had occurred in some parts of the coastal fringes and the lower parts of the larger river systems. The most affected river type was small, low- gradient streams. 4. The predicted biodiversity condition together with river types based on macroinver- tebrate assemblages and abiotic attributes was used to estimate a biodiversity persistence index (BDI). 5. A priority value for each subcatchment was calculated for different actions by changing the disturbance values for that subcatchment and calculating the resulting marginal change in regional BDI. Maps were thereby created for three different types of priority: catchment protection priority, catchment restoration priority and river section conserva- tion priority. 6. The subcatchments of high catchment protection priority for river biodiversity were mostly in the uplands and within protected areas. The river sections of high conservation priority included many coastal lowland rivers in and around protected areas as well as many upland headwater streams. Subcatchments of high priority for catchment restoration were mostly in coastal areas or lowland floodplains. Correspondence: Eren Turak, Department of Environment, Climate Change and Water, 59-61 Goulburn St, Sydney, New South Wales 2000, Australia. E-mail: [email protected] Freshwater Biology (2011) 56, 39–56 doi:10.1111/j.1365-2427.2009.02394.x ȑ 2010 Blackwell Publishing Ltd 39
Transcript of Planning for the persistence of river biodiversity: exploring alternative futures using...

Planning for the persistence of river biodiversity:exploring alternative futures using process-based models
EREN TURAK*, SIMON FERRIER* , †, TOM BARRETT‡, EDWINA MESLEY*, MICHAEL
DRIELSMA ‡, GLENN MANION‡, GAVIN DOYLE § , JANET STEIN– AND GEOFF GORDON*
*Department of Environment, Climate Change and Water, Sydney NSW, Australia†CSIRO Entomology, Canberra, ACT, Australia‡Department of Environment, Climate Change and Water, Armidale NSW, Australia§Hunter-Central Rivers CMA, Paterson NSW, Australia–The Fenner School of Environment and Society, Australian National University, Canberra, ACT, Australia
SUMMARY
1. Planning for the conservation of river biodiversity must involve a wide range of
management options and account for the complication that the effects of many
actions are spatially removed from these actions. Reserve design algorithms widely
used in conservation planning today are not well equipped to address such
complexities.
2. We used process-based models to estimate the expected persistence of river biodiversity
under alternative catchment-wide management scenarios and applied it in the Hunter
Region (37 000 km2) in southeastern Australia.
3. The biological condition of 12 197 subcatchments was estimated using a multiple linear
regression model that related assessments of the integrity of macroinvertebrate assem-
blages to human-induced disturbances at river sites. The best-fit model (R2 = 0.76) used
measures of both local and catchment-wide disturbances as well as elevation and distance
from source as predictor variables. Based on the outputs of this model, we estimated that
substantial loss of river biodiversity had occurred in some parts of the coastal fringes and
the lower parts of the larger river systems. The most affected river type was small, low-
gradient streams.
4. The predicted biodiversity condition together with river types based on macroinver-
tebrate assemblages and abiotic attributes was used to estimate a biodiversity persistence
index (BDI).
5. A priority value for each subcatchment was calculated for different actions by changing
the disturbance values for that subcatchment and calculating the resulting marginal
change in regional BDI. Maps were thereby created for three different types of priority:
catchment protection priority, catchment restoration priority and river section conserva-
tion priority.
6. The subcatchments of high catchment protection priority for river biodiversity were
mostly in the uplands and within protected areas. The river sections of high conservation
priority included many coastal lowland rivers in and around protected areas as well as
many upland headwater streams. Subcatchments of high priority for catchment restoration
were mostly in coastal areas or lowland floodplains.
Correspondence: Eren Turak, Department of Environment, Climate Change and Water, 59-61 Goulburn St, Sydney, New South Wales
2000, Australia. E-mail: [email protected]
Freshwater Biology (2011) 56, 39–56 doi:10.1111/j.1365-2427.2009.02394.x
� 2010 Blackwell Publishing Ltd 39

7. This approach may be particularly well suited to guide the integrated implementation of
three place-based protection strategies proposed for freshwaters: focal areas, critical
management zones and catchment management zones.
Keywords: conservation planning, persistence, rivers, scenario modelling
Introduction
Freshwater biodiversity is imperilled across the world
(e.g. Malmqvist & Rundle, 2002; Dudgeon et al., 2006).
While interest in conserving river biodiversity is not
new, only recently, systematic approaches aimed at
selecting a complementary set of sites for conservation
at the scale of broad geographical regions (e.g.
Margules & Pressey, 2000) have been proposed for
rivers (e.g. Linke et al., 2007; Thieme et al., 2007;
Moilanen, Leathwick & Elith, 2008).
The central goal of systematic conservation plan-
ning is to allow representative examples of the
biodiversity of the planning region to persist (Mar-
gules & Pressey, 2000). To allow persistence, conser-
vation areas must be spatially configured to
accommodate ecological processes such as dispersal,
local extinctions and recolonisations and possible
adjustments of species distributions to climate change
(Sarkar et al., 2006). Systematic conservation planning
typically involves first identifying measureable fea-
tures of biodiversity (biodiversity surrogates; Mar-
gules & Pressey, 2000; Sarkar & Margules, 2002) and
then setting quantitative targets for including these
features in areas where protection measures will be
applied (Margules & Pressey, 2000). The setting of
these targets is based on assumptions about the
conservation return expected from protecting an area
of a given size. The world conservation union (IUCN)
has officially adopted a guideline that 10% of each
biome should be included in protected areas (McNe-
ely, 1993), leading to the widespread use of the target
of 10% of any biodiversity surrogate in conservation
plans at a wide range of spatial scales (Soule &
Sanjayan, 1998; Pressey, Cowling & Rouget, 2003;
Brooks et al., 2004). This target is loosely based on a
theoretical premise that a 10-fold increase in area will
double the number of species and hence 10% of an
area protects 50% of species (Diamond & May, 1976).
However, the unreserved and indiscriminate applica-
tion of this target has been questioned on the basis
that protecting 50% of species cannot universally be
considered as a satisfactory target (Soule & Sanjayan,
1998; Rodrigues & Gaston, 2001; Pressey et al., 2003)
and because the requirement for protection would
necessarily vary among species and ecosystem types
(Pressey et al., 2003; Desmet & Cowling, 2004). For
rivers, a specified percentage of the length of river
type has been used in several studies in this volume:
15% (P. C. Esselman & J. D. Allan, in review), 20%
(J. L. Nel, B. Reyers, D. J. Roux & R. M. Cowling, in
review) and different values depending on river size
(N. A. Rivers-Moore, P. S. Goodman & J. L. Nel, in
review). These targets were assigned on the basis of
expert opinion. While one of these was defined for
fish (P. C. Esselman & J. D. Allan, in review), the other
targets were not specifically assigned for any taxo-
nomic group but were considered to address the
protection of all river biodiversity (J. L. Nel, B. Reyers,
D. J. Roux & R. M. Cowling, in review; N. A. Rivers-
Moore, P. S. Goodman & J. L. Nel, in review).
Arbitrary targets without any ecological basis may
lead to inadequate protection (Soule & Sanjayan, 1998;
Pressey et al., 2003). Also, such targets do not explic-
itly recognise the additional contribution to biodiver-
sity of going beyond a target nor the contribution of
features that are below target (Arponen et al., 2005;
Ferrier et al., 2009). One solution when working with
community-level surrogates – e.g. vegetation types or
river types – is to explicitly model the proportion of
biodiversity expected to be retained in each surrogate
class as a continuously increasing function of the
proportion of that class conserved (e.g. Ferrier et al.,
2004; Allnutt et al., 2008; Faith, Ferrier & Williams,
2008). This is achieved using some form of species–
area relationship (SAR), most commonly formulated
as a power function (Arrhenius, 1921):
S = CAz
where S is number of species, A is habitat area and C
and z are constants. This formulation enables, for each
surrogate class, the proportion of species expected to
be retained in a given retained proportion of original
habitat to be estimated as:
Sretained ⁄Soriginal = (Aretained ⁄AOriginal)z
40 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

Recent extensions to this approach also allow
information on overlap (similarity) in species compo-
sition between surrogate classes to be considered in
estimating the overall proportion of species retained
across all classes within a region (e.g. Ferrier et al.,
2004, 2009; Faith et al., 2008).
Another challenge confronting systematic conser-
vation planning is to develop effective ways to
account for the impacts (both positive and negative)
of multiple forms of off-reserve land use and man-
agement on conservation outcomes for biodiversity at
a regional scale (Sarkar et al., 2006). This problem
takes on an added level of complexity when dealing
with rivers because of the large influence that human
activities in the catchment can have on biodiversity in
downstream sections of river. Management actions,
therefore, must often be applied at locations spatially
removed (i.e. upstream) from the biodiversity features
intended to benefit from these actions.
The usefulness of conservation planning methods
and tools also depends on how they are linked to
implementation and hence to governance of conser-
vation. In most countries and regions, the governance
of freshwater conservation involves multiple agencies
and stakeholders. For example, in New South Wales,
Australia’s most populous state, three important areas
of natural resource management [(i) spatial configu-
ration and management of reserves, (ii) major water
allocation and sharing arrangements and (iii) land
management decisions at the scale of individual
properties] are the responsibility of different govern-
ment agencies. A tool that evaluates the effects of
management actions on river biodiversity in terms of
a single currency is likely to allow for greater
coordination and congruence among stakeholders in
taking action. It would also help to bring transparency
to trade-offs made between biodiversity and other
values.
In this article, we describe a new method for
prioritising actions to conserve river biodiversity,
based on process-based modelling of the level of
persistence of biodiversity expected under alternative
catchment-wide management scenarios. The method
is derived from an existing process-based modelling
approach to conservation planning employed widely
in terrestrial environments in New South Wales over
recent years (e.g. Ferrier, 2005; Drielsma & Ferrier,
2006; DECC 2007a). In adapting this approach to river
environments, we model the condition of river
sections as a function of upstream disturbance factors
(Stein, Stein & Nix, 1998, 2002). We then model the
consequences of predicted changes in river condition
for the persistence of biodiversity at a regional level,
using a SAR-based analysis with multi-attribute eco-
logical river types (Turak & Koop, 2008) serving as
broad surrogates for river biodiversity.
The method we describe here can be used to
generate three main types of output that may help
protect and improve river biodiversity in a planning
region:
1 maps that help identify spatial priorities for
different types of management actions under hypo-
thetical, region-wide management scenarios;
2 quantitative evaluation of specific solutions
involving a suite of management actions at multiple
locations; and
3 optimisation of the spatial configuration of spe-
cific management interventions, e.g. design of fresh-
water protected areas.
Here, we demonstrate the application of the method
to generate the first of these outputs (priority maps)
for the Hunter Region in southeastern Australia.
Methods
We constructed a model to predict river biodiversity
for alternative land management scenarios. We mod-
elled the condition (state) of local instream biodiver-
sity at a river section as a function of local and
upstream disturbance, allowing future change in this
condition to be predicted from anticipated future
change in disturbance resulting from proposed man-
agement actions. The conservation return of these
actions was then estimated in terms of the expected
change in persistence of biodiversity across the entire
region.
Study area
We applied this approach in the Hunter-Central
Rivers Region (37 000 km2) in New South Wales in
southeastern Australia (Fig. 1). The region covers a
variety of landscape types from the sub-alpine Bar-
rington Tops area (highest point, Brumlow Top,
1586 m) to the low-rainfall zones in the upper Hunter
Valley, and lowland swamps and coastal lakes in the
east of the region. Mean annual rainfall varies across
the region from below 700 mm in the west to more
Planning for persistence of river biodiversity 41
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

than 2100 mm in the mid- northern section. Large
geological differences in the region result in a range of
background levels of salinity in large lowland rivers,
from <100 lS cm)1 to more than 1000 lS cm)1 (Hose
& Turak, 2006).
Approximately, a million people live in the study
area with particularly large urban populations in the
coastal fringes to the south of the mouth of the Hunter
River, including the city of Newcastle.
There are numerous protected areas in the region
(Fig. 1). The biological condition of rivers in the region
varied greatly (Hose & Turak, 2006). Generally, the
poorest conditions occurred downstream of towns
and large mines and the best conditions within
protected areas, but there were some sites within
national parks that were in poor biological condition
and many sites outside of protected areas that were in
very good condition (Hose & Turak, 2006).
Delineating river sections and subcatchments
The fundamental spatial unit used in the model is a
river section, defined as the segment of river or stream
between nodes (confluences). The location of the
nodes will depend on the drainage network used.
The drainage network we used here was derived by
Stein (2005, 2007) from a 9-s resolution (c. 270 m)
Digital Elevation Model (DEM) (Hutchinson, Stein &
Stein, 2001) and associated definition of surface flow
pathways. The region was then divided into small
subcatchments delineating the area draining directly
to each stream section. The attributes of the contrib-
uting area to each stream section were determined by
adding the attributes of all subcatchments, weighted
by their relative contribution to catchment run-off
above that section. To determine upstream–down-
stream connectivity, we used a modified version
(Stein, 2005, 2007) of the Pfafstetter coding system
(Pfafstetter, 1989; Verdin & Verdin, 1999) that assigns
to each subcatchment a specific code based on its
location within the overall drainage system. These
codes allow ready identification of all the subcatch-
ments that are upstream from a stream section.
Quantifying disturbance
To quantify local pressure on river reaches, we used
an index of anthropogenic disturbance developed to
help identify wild rivers across Australia (Stein et al.,
1998, 2002). This index uses seven different indicators
to determine modification from a pre-European (pre-
1788) condition. The value of each of these indicators
(factor scores) was calculated for every subcatchment
or stream section, then weighted and combined to
produce two summary indicators of local disturbance:
the Subcatchment Disturbance Index (SCDI) and the
Section Flow Regime Disturbance Index (SFRDI). The
SCDI incorporates four factors calculated from sepa-
Wollemi NP
Myall Lakes NP
Barrington Tops NP
Nowendoc NP
Watagans NP
Jilliby SCA
Hunter River
Manning River
Goulburn RiverHunt
erRive
r
Newcastle
Myall R
iver
N
Protected areasWater bodies
0 50 100 Kilometres
Fig. 1 Major rivers and protected areas of
the Hunter Region. The inset shows the
location of the Hunter Region. The study
area is contained within New South Wales
(NSW), the borders of which define the
spatial extent of the ecological river types
used as biodiversity surrogates. The two
categories of protected areas shown in the
labels are National Parks (NP) and State
Conservation Areas (SCA).
42 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

rate indicators reflecting the spatial extent and
potential magnitude of impact of activities occurring
within the subcatchment: the extractive indus-
tries ⁄point sources factor (PSF), infrastructure factor
(IF), settlement factor (SF) and land use factor (LUF).
The SFRDI incorporates three factors that indicate
direct alterations to flow: impoundments factor (IMF),
levee bank factor (LBF) and flow diversion factor
(FDF). The SCDI and SFRDI values, weighted by their
relative contribution to catchment run-off, were accu-
mulated for all subcatchments upstream from any
river reach to account for upstream disturbances, to
give the Catchment Disturbance Index (CDI) and the
Flow Regime Disturbance Index (FRDI) for any river
reach. These values were then averaged to give the
River Disturbance Index (RDI), which we used as the
single measure that integrates both local and
upstream pressure on river ecosystems.
With the exception of the LUF, we used the values
generated by Stein (2007) to represent each factor in our
model. These values were derived from primary data
layers obtained from different state and national
databases (see Stein et al., 1998, 2002; Stein, 2007 for
details). We based the LUF values on the latest
available land use data from the Hunter Region (DECC
2007b). The 172 different land use classes were grouped
into to 20 new classes, each of which was given a
disturbance weighting indicating its potential impact
on biodiversity (Table 1). The grouping and weighting
was performed with the aim of meaningfully linking
management actions with likely improvements in river
biodiversity. It was based on local professional knowl-
edge of river ecosystems and the catchment action plan
for the region that defines specific management targets
(HCR CMA, 2007). We anticipated that actions taken to
meet these targets would change the land use class into
one that had a lower disturbance weighting (Table 2)
leading to lower values for the LUF, CDI and RDI
downstream from these actions.
Measuring biodiversity condition and predicting it from
disturbance
As a measure of biodiversity condition, we used
observed ⁄expected (O ⁄E) values generated from pre-
dictive models incorporated in the AUSRIVAS (Aus-
tralian River Assessment System, Davies, 2000;
Simpson & Norris, 2000). At any river location, the
AUSRIVAS values indicate the proportion of expected
macroinvertebrate taxa that were actually observed
there (Simpson & Norris, 2000). For this study, we
used the results of assessments made at 80 river sites
across the Hunter Region between 1994 and 1999
(Turak, Hose & Waddell, 2002; Hose & Turak, 2006).
These were the only available assessments suitable for
representing river condition across the whole region.
We used the medians of the AUSRIVAS O ⁄E values
from multiple samples collected at each site to
represent the biodiversity condition separately for
the edge and riffle habitat. Edge habitats are areas on
the edges of rivers with little or no flow, and riffle
habitats are zones with broken water over stony
substratum (Turak, Waddell & Johnstone, 2004). The
riffle habitat was not present at some sites in which
case the median O ⁄E value for the edge habitat was
used to represent the biodiversity condition at a site.
Where both habitats were sampled, we averaged the
results for the two habitats to obtain an overall
assessment for invertebrate assemblages for each site.
To predict biodiversity condition as a function of
disturbance, we fitted a linear regression model using
the R statistical program (R Development Core Team,
2004) to predict O ⁄E values as a function of distur-
bance indices. Both cumulative disturbances
upstream and local disturbances (including the
changes to the riparian zone and land use in the
immediate surrounding areas) will affect instream
biota at any location, so we included the measures of
local disturbance (SCDI, SFRDI and the seven factor
scores) as well as the accumulated disturbance mea-
sures (CDI, FRDI and RDI) as potential input variables
in the model. We assumed that the relationships
between upstream catchment disturbances and biodi-
versity measures may vary across the regions depend-
ing on the location and river type. To account for such
differences, we considered maximum distance from
source (DFS), elevation, slope, mean annual rainfall,
and ecological river types (Turak & Koop, 2008) as
potential input variables in the model. The regression
equation for this model was used to estimate the
biodiversity condition in all river sections in the
region and to predict how biodiversity condition
might change in the future in response to changes in
the degree of disturbance within the catchment.
The sampling sites in the study region were not
randomly selected. A large proportion of them had
been selected as reference sites for the development of
AUSRIVAS predictive models in NSW (Turak et al.,
Planning for persistence of river biodiversity 43
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56
1999, 2002) and were, therefore, presumed in very
good condition. This left very few sites in poor
condition. For details of condition assessments of
these sites, see Hose & Turak (2006). A graph of the
O ⁄E values predicted from the linear regression
against the actual O ⁄E values suggested that the
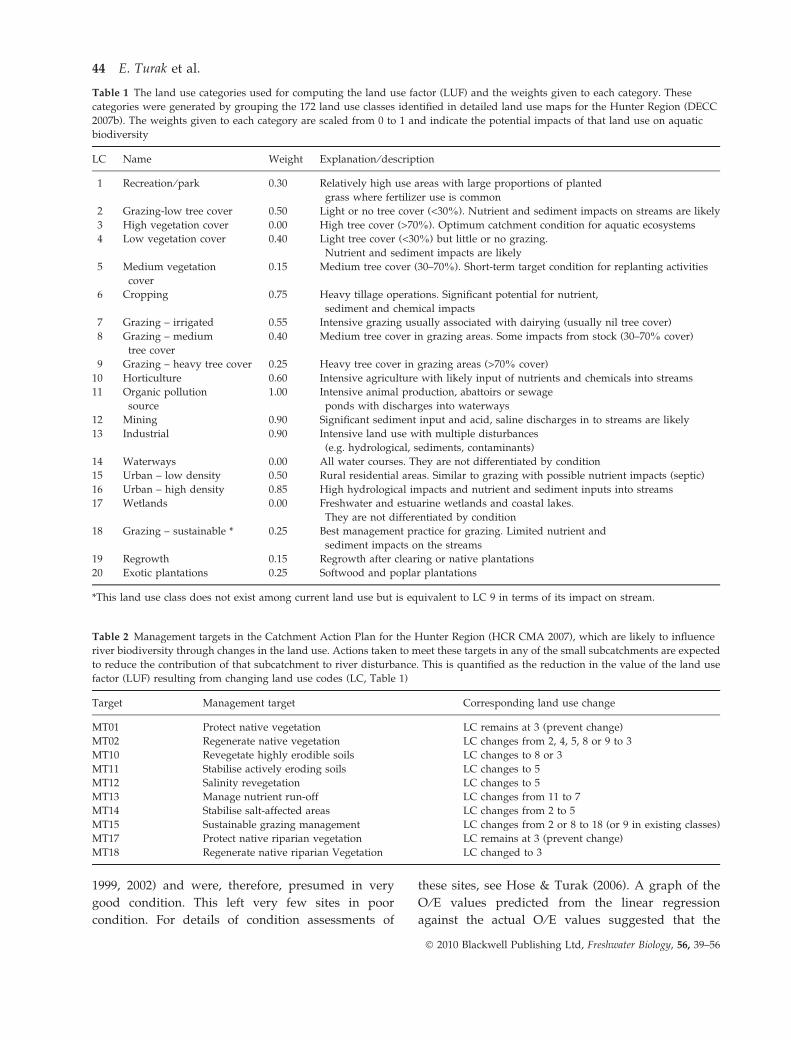
Table 1 The land use categories used for computing the land use factor (LUF) and the weights given to each category. These
categories were generated by grouping the 172 land use classes identified in detailed land use maps for the Hunter Region (DECC
2007b). The weights given to each category are scaled from 0 to 1 and indicate the potential impacts of that land use on aquatic
biodiversity
LC Name Weight Explanation ⁄ description
1 Recreation ⁄ park 0.30 Relatively high use areas with large proportions of planted
grass where fertilizer use is common
2 Grazing-low tree cover 0.50 Light or no tree cover (<30%). Nutrient and sediment impacts on streams are likely
3 High vegetation cover 0.00 High tree cover (>70%). Optimum catchment condition for aquatic ecosystems
4 Low vegetation cover 0.40 Light tree cover (<30%) but little or no grazing.
Nutrient and sediment impacts are likely
5 Medium vegetation
cover
0.15 Medium tree cover (30–70%). Short-term target condition for replanting activities
6 Cropping 0.75 Heavy tillage operations. Significant potential for nutrient,
sediment and chemical impacts
7 Grazing – irrigated 0.55 Intensive grazing usually associated with dairying (usually nil tree cover)
8 Grazing – medium
tree cover
0.40 Medium tree cover in grazing areas. Some impacts from stock (30–70% cover)
9 Grazing – heavy tree cover 0.25 Heavy tree cover in grazing areas (>70% cover)
10 Horticulture 0.60 Intensive agriculture with likely input of nutrients and chemicals into streams
11 Organic pollution
source
1.00 Intensive animal production, abattoirs or sewage
ponds with discharges into waterways
12 Mining 0.90 Significant sediment input and acid, saline discharges in to streams are likely
13 Industrial 0.90 Intensive land use with multiple disturbances
(e.g. hydrological, sediments, contaminants)
14 Waterways 0.00 All water courses. They are not differentiated by condition
15 Urban – low density 0.50 Rural residential areas. Similar to grazing with possible nutrient impacts (septic)
16 Urban – high density 0.85 High hydrological impacts and nutrient and sediment inputs into streams
17 Wetlands 0.00 Freshwater and estuarine wetlands and coastal lakes.
They are not differentiated by condition
18 Grazing – sustainable * 0.25 Best management practice for grazing. Limited nutrient and
sediment impacts on the streams
19 Regrowth 0.15 Regrowth after clearing or native plantations
20 Exotic plantations 0.25 Softwood and poplar plantations
*This land use class does not exist among current land use but is equivalent to LC 9 in terms of its impact on stream.
Table 2 Management targets in the Catchment Action Plan for the Hunter Region (HCR CMA 2007), which are likely to influence
river biodiversity through changes in the land use. Actions taken to meet these targets in any of the small subcatchments are expected
to reduce the contribution of that subcatchment to river disturbance. This is quantified as the reduction in the value of the land use
factor (LUF) resulting from changing land use codes (LC, Table 1)
Target Management target Corresponding land use change
MT01 Protect native vegetation LC remains at 3 (prevent change)
MT02 Regenerate native vegetation LC changes from 2, 4, 5, 8 or 9 to 3
MT10 Revegetate highly erodible soils LC changes to 8 or 3
MT11 Stabilise actively eroding soils LC changes to 5
MT12 Salinity revegetation LC changes to 5
MT13 Manage nutrient run-off LC changes from 11 to 7
MT14 Stabilise salt-affected areas LC changes from 2 to 5
MT15 Sustainable grazing management LC changes from 2 or 8 to 18 (or 9 in existing classes)
MT17 Protect native riparian vegetation LC remains at 3 (prevent change)
MT18 Regenerate native riparian Vegetation LC changed to 3
44 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

regression relationship was dominated by the large
number of sites having good condition. Given the
current lack of sufficient data from sites with poor
condition, as a temporary ‘work-around’, we
weighted the regression relationship between biolog-
ical condition and disturbance to give more weight to
the relatively small number of poor condition sites.
An ad hoc weighting was used – all points were
weighted inversely according to their observed O ⁄Evalues. This meant that the lower the observed O ⁄Evalue of a site, the greater was its influence on the
regression.
Because insufficient data were available to test the
predictions of the regressions on an independent
data set, we used two cross-validation approaches
to ensure that there was no over-fitting and that
model predictions were well grounded. In the first
approach, the model was used repeatedly with a
single point at a time dropped from the data and
the prediction for the data point dropped compared
with the actual value. In the second approach, a
folded cross-validation was used, in which one-
tenth of the data was removed at a time, and
predictions for the group dropped compared with
their actual values. The cross-validations were per-
formed using the cv.glm function of the R library
‘boot’ (Canty & Ripley, 2009), with a mean square
prediction error. The cross-validations were used to
compare the prediction efficiencies of alternative
models to check that steps taken to improve the
goodness of fit (weighting and the removal of high
leverage points) did not result in over-fitting.
We used a stepwise variable selection procedure,
using both forward and backward selection, to select
the most appropriate models for both unweighted and
weighted regressions. This was performed using the
function stepAIC of the R library MASS (Venables &
Ripley, 2002), which uses the Akaike Information
Criterion as a selection criterion to select the most
parsimonious fits. We also used diagnostic plots to
identify high leverage points. In some cases, a small
number of these points were removed from the data.
This was only performed in exceptional circum-
stances, namely, when assessments were based on
just one or two samples, when there was evidence that
samples had been poorly collected, or when the site in
question had been subject to an impact of a type that
would not be well accounted for by the disturbance
indicators of Stein et al. (2002).
This regression was then used to make predictions
for the remaining of the 12 197 subcatchments, for
which no biological assessment data were available.
The resulting predicted O ⁄E values were then stan-
dardised, so that their values all lay between 0 and 1,
by dividing them by the maximum predicted value in
the catchment. The standardised values were mapped
using different colour bands to represent seven
biodiversity condition categories: reference (0.95–1),
very good (0.85–0.95), good (0.65–0.85), moderate
(0.35–0.65), poor (0.15–0.35), very poor (0.05–0.15)
and extremely poor (0–0.05). We refer to scores above
0.95 as ‘reference’ because the biodiversity condition
scores were predicted from site assessments based on
comparisons with best available reference sites (Turak
et al., 1999; Simpson & Norris, 2000). The number
of condition categories and the score intervals repre-
senting these categories were chosen to be consistent
with the rank priority categories for subcatchments
explained later in this section. These biodiversity
condition categories are used only for visually repre-
senting the spatial patterns in predicted biodiversity
condition.
Mapping ecological river types
Ecological river types (Turak, 2007; Turak & Koop,
2008) were used as surrogate features for river
biodiversity composition. It was necessary to map
them in the study area, so that each river section could
be assigned to a river type. This was performed using
identification keys based on slope, elevation, maxi-
mum DFS, mean annual rainfall and latitude (Turak,
2007). To do this, first a comprehensive drainage
network in the Hunter Region was determined using
ESRI’s ArcHydro extension in ArcGIS (ESRI, 2005).
This hydrological analysis used existing high-resolu-
tion drainage data and created a network of river
reaches with contributing catchment area ‡1.6 km2.
Maximum DFS was calculated for the catchment
using Arcview 3.3¢s Hydrotools extension (ESRI,
1999). Values along the network for DFS, elevation
and rainfall were extracted from a drainage data layer
of the Department of Water and Energy, of NSW, a
25 m DEM and some NSW-wide annual rainfall data,
respectively. River reach slope was calculated using
the elevation network grid and a method involving
neighbourhood analysis undertaken in ArcGIS (ESRI,
2005).
Planning for persistence of river biodiversity 45
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

For this study, we used two of the four river
typologies defined by Turak & Koop (2008). These are
the macroinvertebrate edge and abiotic typologies. We
will refer to the macroinvertebrate edge river types as
‘macroinvertebrate types’ throughout this article.
The biodiversity persistence index (BDI)
The common currency by which the regional status of
biodiversity condition in rivers was estimated is the
BDI. Calculations of BDI values were based on
the concept of the original habitat area (OHA) and
the effective habitat area (EHA) for each subcatch-
ment, where the former represents a condition in
which all river types are in an undisturbed state (i.e.
has an RDI value of 0) and the latter represents the
current state. Given the linearity of rivers, we have
used river length as a surrogate for habitat area.
The original and EHAs were calculated for each
river type i within a river typology (abiotic and
macroinvertebrate) as follows.
oi ¼Xn
j¼1
length of river type i in river section j ð1Þ
ei ¼Xn
j¼1
condition in river section j
� length of river type i in section j ð2Þ
Given that we are using river length as a surrogate
of habitat area, here
oi and ei represent OHA for river type i and EHA for
river type i, respectively, and n is the number of river
sections in the region.
The values for the original and EHAs for all river
types in a given typology were then used to derive the
BDI representing the persistence of biodiversity for
the region as a whole under specified land manage-
ment scenarios. This was achieved using the approach
described by Ferrier et al. (2004) reformulated slightly
by Allnutt et al. (2008) to work with discrete types of
vegetation communities rather than a continuum of
compositional turnover. We estimated the proportion
of biodiversity historically occurring in a river type
that is more likely to have persisted as follows.
pi ¼Xn
j¼1
sijej
,Xn
j¼1
sijoj
24
35
z
ð3Þ
where pi is the proportion of biodiversity predicted to
persist in river type i, sij is a surrogate for similarity
among river types i and j, and was computed as 1 – dij, dij
being the dissimilarity in species composition between
river types i and j. These were estimated as Bray-Curtis
dissimilarities between classes from the numerical
classification of macroinvertebrate assemblages, and
generalised squared distances between classes from
numerical classification based on abiotic variables
(Turak, 2007). This is based on the assumption that that
each river type shares species with other river types.
The species–area exponent z (Arrhenius, 1921)
accounts for diminishing biodiversity benefit (conser-
vation return) for increases in EHA. Here, the z value
represents river biodiversity as a whole rather than a
specific taxonomic group. Specific z values have not
been determined for estimating loss of river biodiver-
sity in a fragmented landscape for given habitat loss in
rivers. Hence, we chose a z value of 0.25 which has also
been used in similar applications in other ecosystem
types (e.g. Zurlini, Grossi & Rossi, 2002; Ferrier et al.,
2004 and A.-G. E. Ausseil, W. L. Chadderton, R. T.
Stephens, P. Gerbeux & J. Leathwick, in review
Following from eqn 3, the persistence of river
biodiversity may be computed as the proportion of
persisting species for all river types together. How-
ever, given that the contribution of a river type to
regional river biodiversity will decrease with the
proportion of species that are shared with other river
types, it is necessary to weight river types using the
proportion of shared species. The weight factor for
each river type was computed as follows:
wi ¼oiPn
j¼1
sijoj
� � ð4Þ
The BDI can then be computed as the weighted sum of
p forallrivertypes,scaledtorangefrom0to1bydividing
it by the sum of the weighting factors as follows:
BDI ¼
Pni¼1
wipi
Pni¼1
wi
ð5Þ
Alternative futures
Alternative futures may be explored through sce-
nario modelling which allows biodiversity outcomes
46 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

to be forecast for a range of alternative management
regimes (Peterson, Cumming & Carpenter, 2003;
Drielsma & Ferrier, 2006). Scenario modelling
involves hypothetically manipulating actions with
known or expected links to outcomes of interest. In
the context of conservation planning for rivers, the
outcome of greatest interest is river biodiversity. One
application of scenario evaluation is the generation of
priority maps for specific types of management
actions. This involves hypothetically making a par-
ticular type of change to the land use across the
entire landscape, ranking every river section in terms
of the effect the changes in them have on regional
river biodiversity and converting these rankings into
a priority map. To generate such maps for the
Hunter Region, we made changes in the LUF by
changing the land use types on the land use data
layer. Some changes in the other disturbance factors
were made by altering the tabulated values for each
subcatchment directly. The steps taken to generate
three priority values for the Hunter Region are given
below.
1. Protection priority maps for subcatchments:
These are intended to help identify subcatchments to
target for protection. They provide an estimate of the
relative contribution that protecting each subcatch-
ment makes to the maintenance of current river
biodiversity in the region based on the consequences
of this action on the biodiversity of river sections
downstream. The priority value for each subcatch-
ment was determined as follows:
• The ‘current BDI’ was calculated using current
LUF, SF and IF factor values.
• Degraded condition was simulated by systemat-
ically changing the LUF, SF and IF factors of each
subcatchment to 1 (irrespective of current value).
• BDI value under this degraded condition was
calculated for the whole region.
• Priority value is the difference between current
and ‘degraded BDI’.
This priority value is a measure of the impact of
degrading the current condition of each subcatch-
ment, in terms of expected change in regional biodi-
versity status. It, therefore, indicates the priority for
preventing further loss of condition within each
subcatchment (but only for changes that affect the
LUF, SF and IF factors).
2. Restoration priority maps for subcatchments:
These are intended to help identify subcatchments to
target for restorative, remedial action. They provide
an estimate of the relative contributions of such
actions based on their predicted effect on river
biodiversity downstream. Calculations were made as
follows.
• Current BDI was calculated using current LUF, SF
and IF factor values.
• Improvement in condition was simulated in
accordance with the following rules for each of
the LUF, SF and IF factors:
(i) If factor value £0.2, then it was adjusted to 0.
(ii) If factor value >0.2, then 0.1 was subtracted from
factor value.
• BDI was then re-calculated for the whole scenario.
• Priority value is the difference between current
and ‘restored’ BDI.
This priority value is a measure of the impact of
improving the condition of each subcatchment, in
terms of expected change in regional biodiversity
status. It, therefore, indicates the priority for restora-
tion or improving the condition within each subcatch-
ment (but only for changes that affect the LUF, SF and
IF factors).
3. Conservation priority maps for river sections:
These are intended to help identify river sections that
have high conservation value because of the signifi-
cance of their biodiversity for the region. Because it is
particularly important to protect the biodiversity in
these river sections, they may be suitable for inclusion
into freshwater protected areas and be the focus of
intensive and costly protection and restoration activ-
ities both within that river section and across its entire
catchment. To estimate the importance of an individ-
ual river section, regional biodiversity persistence
(BDI) with and without that river section was calculated.
The difference between these two scenarios divided
by the subcatchment area can be taken as the relative
importance of that river section within the region.
Priority = (BDIwith river section ) BDIwithout river sec-
tion) ⁄area of subcatchment.
The priority index values were converted into rank
percentiles and these were then mapped across the
region under seven priority categories (1 being the
highest and 7 the lowest) as follows. 1:0.95–1, 2: 0.85–
095, 3: 0.65–0.85, 4:0.35–0.65, 5:0.15–0.35, 6:0.05–0.15,
7:0–0.05. In choosing these categories, we used an
approach developed for producing similar priority
maps to conserve or repair terrestrial biodiversity in
the Northern Rivers Region (DECC 2009) based on the
Planning for persistence of river biodiversity 47
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56
normal distribution of ranked priority values of all
grid cells in the planning regions (A. Steed, Pers.
Comm.).
Results
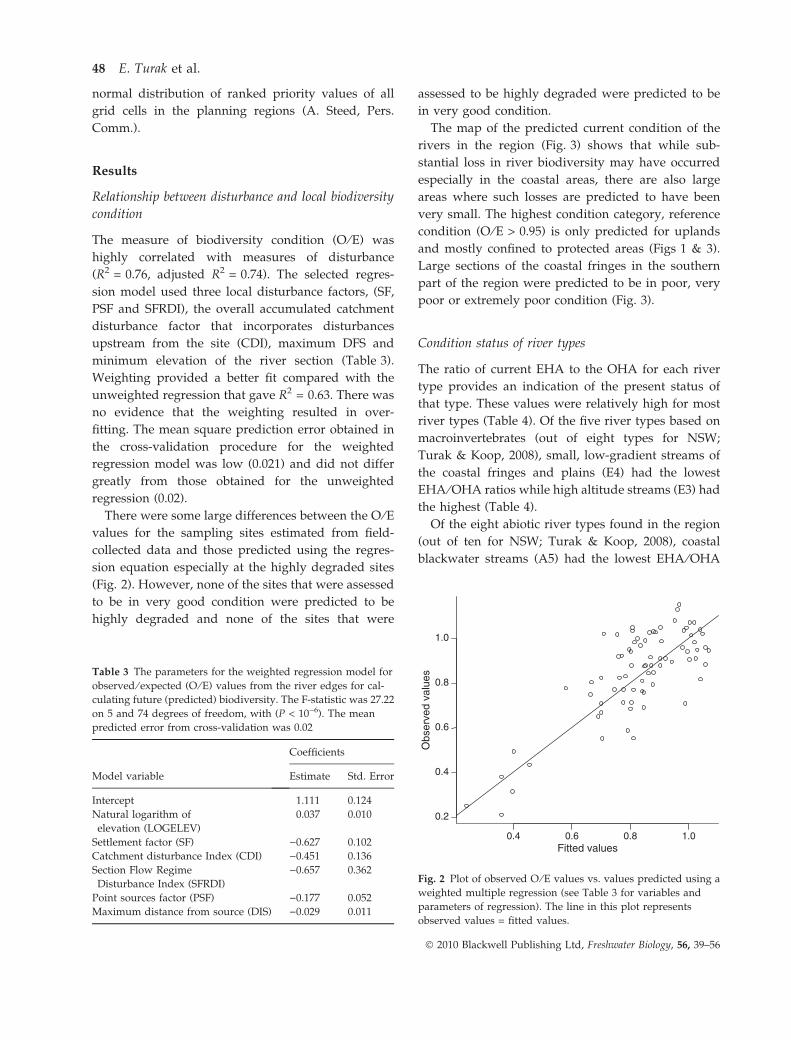
Relationship between disturbance and local biodiversity
condition
The measure of biodiversity condition (O ⁄E) was
highly correlated with measures of disturbance
(R2 = 0.76, adjusted R2 = 0.74). The selected regres-
sion model used three local disturbance factors, (SF,
PSF and SFRDI), the overall accumulated catchment
disturbance factor that incorporates disturbances
upstream from the site (CDI), maximum DFS and
minimum elevation of the river section (Table 3).
Weighting provided a better fit compared with the
unweighted regression that gave R2 = 0.63. There was
no evidence that the weighting resulted in over-
fitting. The mean square prediction error obtained in
the cross-validation procedure for the weighted
regression model was low (0.021) and did not differ
greatly from those obtained for the unweighted
regression (0.02).
There were some large differences between the O ⁄Evalues for the sampling sites estimated from field-
collected data and those predicted using the regres-
sion equation especially at the highly degraded sites
(Fig. 2). However, none of the sites that were assessed
to be in very good condition were predicted to be
highly degraded and none of the sites that were
assessed to be highly degraded were predicted to be
in very good condition.
The map of the predicted current condition of the
rivers in the region (Fig. 3) shows that while sub-
stantial loss in river biodiversity may have occurred
especially in the coastal areas, there are also large
areas where such losses are predicted to have been
very small. The highest condition category, reference
condition (O ⁄E > 0.95) is only predicted for uplands
and mostly confined to protected areas (Figs 1 & 3).
Large sections of the coastal fringes in the southern
part of the region were predicted to be in poor, very
poor or extremely poor condition (Fig. 3).
Condition status of river types
The ratio of current EHA to the OHA for each river
type provides an indication of the present status of
that type. These values were relatively high for most
river types (Table 4). Of the five river types based on
macroinvertebrates (out of eight types for NSW;
Turak & Koop, 2008), small, low-gradient streams of
the coastal fringes and plains (E4) had the lowest
EHA ⁄OHA ratios while high altitude streams (E3) had
the highest (Table 4).
Of the eight abiotic river types found in the region
(out of ten for NSW; Turak & Koop, 2008), coastal
blackwater streams (A5) had the lowest EHA ⁄OHA
Table 3 The parameters for the weighted regression model for
observed ⁄ expected (O ⁄ E) values from the river edges for cal-
culating future (predicted) biodiversity. The F-statistic was 27.22
on 5 and 74 degrees of freedom, with (P < 10)6). The mean
predicted error from cross-validation was 0.02
Model variable
Coefficients
Estimate Std. Error
Intercept 1.111 0.124
Natural logarithm of
elevation (LOGELEV)
0.037 0.010
Settlement factor (SF) )0.627 0.102
Catchment disturbance Index (CDI) )0.451 0.136
Section Flow Regime
Disturbance Index (SFRDI)
)0.657 0.362
Point sources factor (PSF) )0.177 0.052
Maximum distance from source (DIS) )0.029 0.011
0.4 0.6 0.8 1.0
0.2
0.4
0.6
0.8
1.0
Fitted values
Obs
erve
d va
lues
Fig. 2 Plot of observed O ⁄ E values vs. values predicted using a
weighted multiple regression (see Table 3 for variables and
parameters of regression). The line in this plot represents
observed values = fitted values.
48 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

value and high altitude streams (A3) had the highest
(Table 4).
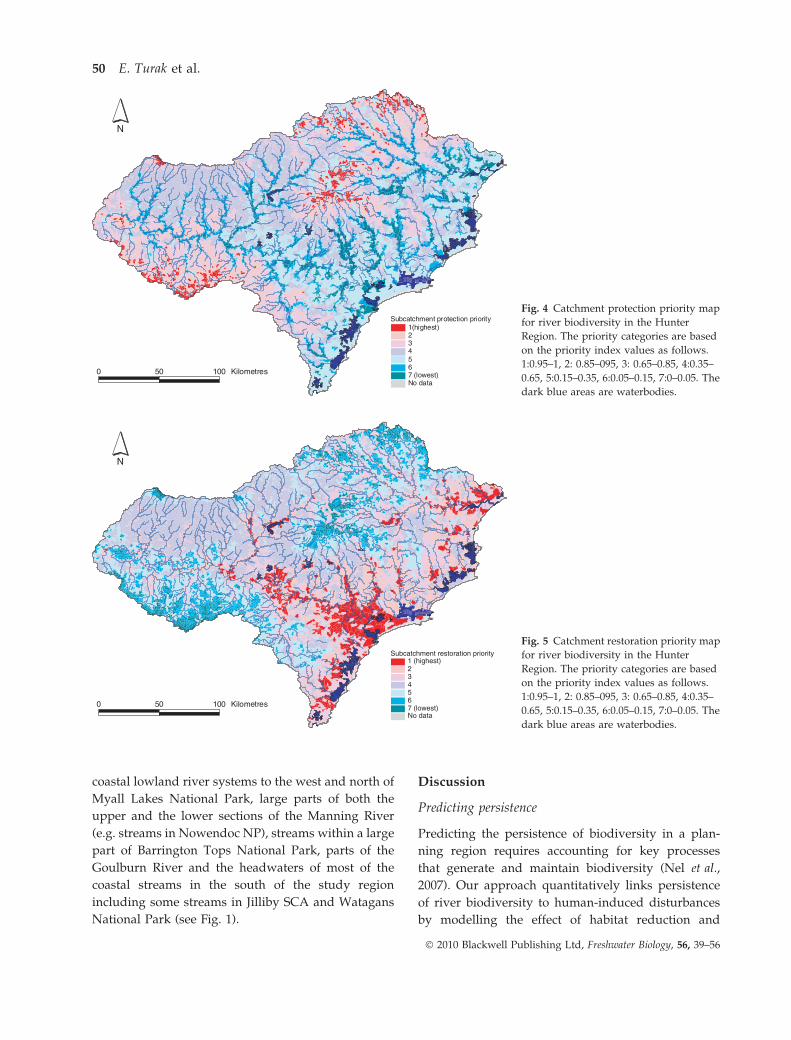
Priority maps
The areas of high priority for the protection of river
biodiversity (Fig. 4) were mostly either within
protected areas or adjacent to these and were largely
confined to the uplands. The greatest density of such
areas was in and around Barrington Tops National
Park, the upper reaches of Wollemi National Park and
areas to its west and the upper parts of some
tributaries of the Manning River including some
streams in Nowendoc National Park (see Fig. 1).
The areas of high catchment restoration priority for
river biodiversity (Fig. 5) were mostly at lower
altitudes on the coastal fringes and the lower parts
of the valleys of the Hunter and Manning Rivers (See
Fig. 1). Some of the upper sections of coastal streams
in the southern section of the regions and headwater
streams of the upper Manning catchments also had
high restoration priority.
River sections of high priority for the conservation
of river biodiversity (Fig. 6) included parts of many
N
No data
Biodiversity conditionReferenceVery goodGoodModeratePoorVery PoorExtremely poor
0 50 100 Kilometres
Fig. 3 Predicted current condition of river
biodiversity relative to a pre-European
reference condition. The condition cate-
gories were based on predicted O ⁄ Evalues as follows. Reference: 0.95–1, very
good: 0.85–095, good: 0.65–0.85, moderate:
0.35–0.65; poor: 0.15–0.35, very poor: 0.05–
0.15, extremely poor: 0–0.05. The dark
blue areas are waterbodies.
Table 4 The status of river types (Turak,
2007; Turak & Koop 2008) in the Hunter
Region. EHA ⁄ OHA (effective habitat
area ⁄ original habitat area) represents the
predicted overall biodiversity condition of
each river type relative to a pre-European
condition. Percentage of total is the
percentage of the length of rivers in the
region that were assigned to each river
type. Percentage in PAs is the percentage
of the length of each river type that fall
within the boundaries of protected areas
River types and their key attributes
EHA ⁄OHA
Percentage
of total
Percentage
in PAs
Macroinvertebrate typology (edge fauna)
E1: Small streams with a rich fauna 0.83 65.7 17.8
E2: Large lowland rivers with a rich fauna 0.72 12.3 8.1
E3: High altitude streams of intermediate richness 0.91 1.2 40
E4: Small coastal streams of intermediate richness 0.64 16.0 8
E7: Small, rocky streams of intermediate richness 0.71 4.9 5.5
Abiotic typology
A1: Large lowland rivers 0.67 9.6 6.3
A2: Small steep lowland streams 0.72 10.3 6.4
A3: Highland streams 0.88 2.8 29.8
A4: Streams and rivers of the western slopes 0.90 0.1 14.9
A5: Low gradient, acidic,
blackwater coastal streams
0.55 7.3 10.3
A7: Lowland streams of intermediate gradient 0.77 22.3 10.6
A9: Small to medium upland streams
with low bedrock
0.83 3.2 9.3
A10 Small, steep, rocky upland streams 0.86 44.5 20.7
Planning for persistence of river biodiversity 49
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56
coastal lowland river systems to the west and north of
Myall Lakes National Park, large parts of both the
upper and the lower sections of the Manning River
(e.g. streams in Nowendoc NP), streams within a large
part of Barrington Tops National Park, parts of the
Goulburn River and the headwaters of most of the
coastal streams in the south of the study region
including some streams in Jilliby SCA and Watagans
National Park (see Fig. 1).
Discussion
Predicting persistence
Predicting the persistence of biodiversity in a plan-
ning region requires accounting for key processes
that generate and maintain biodiversity (Nel et al.,
2007). Our approach quantitatively links persistence
of river biodiversity to human-induced disturbances
by modelling the effect of habitat reduction and
0 50 100 Kilometres
N
No data
Subcatchment protection priority1(highest)234567 (lowest)
Fig. 4 Catchment protection priority map
for river biodiversity in the Hunter
Region. The priority categories are based
on the priority index values as follows.
1:0.95–1, 2: 0.85–095, 3: 0.65–0.85, 4:0.35–
0.65, 5:0.15–0.35, 6:0.05–0.15, 7:0–0.05. The
dark blue areas are waterbodies.
0 50 100 Kilometres
N
No data
Subcatchment restoration priority1 (highest)234567 (lowest)
Fig. 5 Catchment restoration priority map
for river biodiversity in the Hunter
Region. The priority categories are based
on the priority index values as follows.
1:0.95–1, 2: 0.85–095, 3: 0.65–0.85, 4:0.35–
0.65, 5:0.15–0.35, 6:0.05–0.15, 7:0–0.05. The
dark blue areas are waterbodies.
50 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

degradation on the net outcome of the processes of
emigration, re-colonisation and local extinctions.
Undoubtedly, these links are much more complex
than what our models account for. For example, we
have ignored the spatial configuration of the remain-
ing habitat and assumed that recovery of biodiver-
sity from disturbance will occur rapidly and
predictably once the disturbance is removed (i.e.
rubber band model of recovery; Sarr, 2002). Process-
based models that allow accurate predictions of the
persistence of river biodiversity would require,
among other things, accounting for the effect of
habitat fragmentation on recolonisation (e.g. see
Fagan, 2002) and, where possible, making distinc-
tions between the pathways and rates of recovery
and those of degradation (e.g. see Lake, Bond &
Reich, 2007).
Biodiversity condition and catchment disturbance
Estimating the biodiversity condition of any section of
river as a function of anthropogenic disturbances in its
catchment is a critical step in predicting the persis-
tence of biodiversity at a broad spatial scale. The
method we have used here represents just one of
many possible approaches. Our regression model
predicted that both local and catchment-scale distur-
bances affect local biodiversity condition (Table 3).
However, the estimated contributions of local distur-
bances to downstream impacts currently do not
depend on distance. Consequently, processes other
than dilution that may reduce impacts with distance
have been ignored. To make more reliable predictions
of the effects of disturbance on biodiversity, it will be
necessary to incorporate a decay function that
accounts for such processes.
The uneven spread of the sampling sites along
disturbance gradients (see Fig. 2) limited the useful-
ness of the biological datasets available for construct-
ing the regression models in our study area. By using
an ad hoc weighting, we have emphasised a part of the
data where we have poor coverage and have
attempted to compensate for the bias in the data
towards undisturbed sites. This, however, may have
addressed only part of the problem. Because of the
lack of data from highly disturbed environments such
as large urban centres, we may have extrapolated too
far beyond the range of the data to be able to make
reliable predictions of the condition of river biodiver-
sity in such environments. Even though the study
region was relatively rich in terms of biological data,
many of the sampling sites contribute little to the
predictions because they are concentrated at the lower
end of the disturbance gradient. Monitoring programs
targeting sites that are more evenly distributed along
this gradient than those used here should produce
0 50 100 Kilometres
N
No data
River section conservation priority1 (highest)234567
Fig. 6 River section conservation priority
map for aquatic biodiversity in the Hunter
Region. The priority categories are based
on the priority index values as follows.
1:0.95–1, 2: 0.85–095, 3: 0.65–0.85, 4:0.35–
0.65, 5:0.15–0.35, 6:0.05–0.15, 7:0–0.05. The
dark blue areas are waterbodies.
Planning for persistence of river biodiversity 51
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

much better input data for predicting biodiversity
condition across the region with considerably less
sampling effort.
Our ability to predict biodiversity condition from
measures of disturbance may be improved by refining
the methods. Steps that can be taken in this direction
include: lowering the taxa probability threshold in
calculating the AUSRIVAS O ⁄E values (Turak &
Koop, 2003); constructing new predictive models that
use only rare taxa (Linke & Norris, 2003); replacing
the disturbance factors determined at a continental
scale (Stein et al., 1998, 2002) with those that better
account for the effects of local land management
practices on river ecosystems; and incorporating
additional disturbance indicators for riparian condi-
tion and instream barriers. The predictions may also
be improved by replacing the linear regressions used
here with more complex relationships, such as gener-
alised additive models (Wood, 2006), which allow for
nonlinearities in response to predictor variables, or
geostatistical models generated for stream networks
(e.g. Peterson, Theobald & Ver Hoef, 2007), which
allow for spatial correlations among sites, or a
combination of such approaches.
Our predictive model was developed in a relatively
data-rich context especially in terms of biological data.
However, the input variables in the model are either
known topographical variables or indices that depend
on land management data which should in principle
be available from remote sensing data independently
of field observations. If our predictive model is
sufficiently robust to be used in other nearby catch-
ments, then the approach we have used may be
extended to catchments elsewhere. The potential to
extend our approach using only remotely obtained
data needs to be explored further.
Estimating conservation return
In our approach, the conservation benefit, or return,
from any management action is estimated in the
currency of the BDI. We used river types based on
abiotic attributes or family-level assemblage structure
of macroinvertebrates (Turak, 2007; Turak & Koop,
2008) to represent the likely influence of climate,
geology, topography and geography on river biodi-
versity at a broad spatial scale. Given that large
sections of the study area such as the southeastern
corner had poor values for biodiversity condition
(Fig. 3), the relatively small distinctions among river
types in terms of their overall biodiversity condition
(EHA ⁄OHA, Table 4) indicates that individually, the
two single attribute typologies used here may be too
coarse as surrogates of river biodiversity. The inclu-
sion of at least 5.5% of the length of each of the river
types in existing protected areas (Table 4) may,
therefore, simply reflect the excessively coarse spatial
resolution of the surrogates rather than the represen-
tativeness of river biodiversity of the protected
areas. Using multi-attribute river types based on the
intersection of different river typologies (Turak &
Koop, 2008), greater distinction may be attained
among surrogate classes in terms of both representa-
tion in protected areas and overall biodiversity
condition.
The relative contribution of each river type to the
regional BDI values depends on the choice of the
species–area relationship coefficient (z value) as well
as the estimated dissimilarities between river types.
Consequently, the relative priority of different sub-
catchments for any given conservation action will
depend on the z value as will the difference in the BDI
values from scenarios that affect different river types.
This means that choosing a different z value may
change priority maps (Figs 4–6) and the BDI values
obtained from any given scenario. We chose a z value
of 0.25 because we judged that this was appropriate to
represent an archipelagic species–area relationship
(Rosenzweig, 1995, 2003) averaged across all taxo-
nomic groups in a fragmented landscape based on
previous use of this value for similar purposes
(Zurlini et al., 2002; Ferrier et al., 2004; Thomas et al.,
2004). Currently, not enough is known to confidently
assign z values to broad groups of biota in rivers.
Marchant, Ryan & Metzeling (2006) and Heino &
Korsu (2008) found no evidence of a predictive
relationship between habitat area and species richness
for macroinvertebrate assemblages. While Sepkoski &
Rex (1974) found that area was the single best
predictor of the number of freshwater mussel species
in 44 coastal rivers in eastern U.S.A., they reported
that the estimated z values for five different subsets of
these rivers ranged from 0.193 to 0.342. Given our
limited knowledge and the potential for large varia-
tion in species–area relationships among freshwater
taxa, it is appropriate to use a z value of 0.25 in
conservation plans aimed at protecting all river
biodiversity in a region.
52 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

Translating predictions of persistence into conservation
actions
The priority maps we generated here may help guide
the implementation of the three different place-based
protection strategies proposed by Abell, Allan &
Lehner (2007): freshwater focal areas, critical management
zones and catchment management zones. The river
section conservation priority maps identify river
sections that are predicted to have the high conserva-
tion values based on macroinvertebrate assemblages.
These river sections may include good candidates for
freshwater focal areas while the subcatchment resto-
ration and protection priority maps may help delin-
eate critical management zones and catchment
protection zones and guide the choice of management
actions within these zones.
We illustrate a hypothetical application for streams
on the coastal fringe in and around Myall Lakes
National Park (Fig. 1). Some of these streams had very
high river section conservation priorities, contrasting
sharply with streams on the coastal fringes in the
southern part of the region which mostly had very low
priorities (Fig. 6). This result may be explained by the
relatively good condition of streams in the Myall Lakes
area (Fig. 3) together with the overall poor condition of
the river types they belong to (E4 and A5) in the
Hunter Region (Table 4). If some of these streams were
designated as freshwater focal areas, critical manage-
ment zones may then be delineated and priorities for
actions within these zones identified using the priority
maps for restoration (Fig. 5) and protection (Fig. 4).
Our model can then be used to compare the biodiver-
sity outcomes of alternative futures formulated as
spatially explicit management scenarios involving
multiple actions at multiple locations. Importantly,
the improvement to BDI predicted under the preferred
management scenario for streams in the Myall Lakes
region can be quantitatively compared with those from
management scenarios requiring similar resources in
other parts of the Hunter Region.
Priority maps for different conservation actions for
different biodiversity surrogates, and the scenario
evaluations for different surrogates, collectively
amount to a large number of outputs. This is useful
in a complex planning process such as the develop-
ment and implementation of the Catchment Action
Plans undertaken by the Catchment Management
Authorities across Australia. However, it could also
lead to confusion. There is a need to put in place rules
about how different outputs may be used together in
the planning process. For example, methods might
need to be developed for combining the terrestrial
biodiversity priorities for catchment protection with
the river biodiversity priority for catchment protection
(Fig. 4) and for combining both these biodiversity
priorities with economic and social priorities. One
way to deal with this would be to use the various
priority maps to formulate a number of alternative
futures using multi-criteria analysis (e.g. Faith, 1995;
Moffett & Sarkar, 2006) or related methods.
Finding the optimal solution to the protection of
biodiversity under specified resource constraints
underpins many recent approaches in conservation
planning (e.g. Possingham et al., 2001). So far, we have
not considered the costs of conservation, nor have we
attempted to optimise conservation solutions. How-
ever, there is considerable potential to link our
process-based modelling of biodiversity persistence
to an optimisation algorithm that automatically
searches through possible spatial configurations to
identify the solution that maximises overall conserva-
tion benefit. Other important factors such as the
material cost per hectare of revegetation could also be
factored into such an analysis, thereby optimising the
ratio of benefit to cost.
Conclusions
Our new approach to freshwater conservation plan-
ning differs from most other complementarity-based
approaches by simultaneously evaluating the effects
of all management actions at all locations in a
planning region and by not relying on protection
targets. It is explicitly underpinned by current eco-
logical theory and readily allows the incorporation of
new data and improved knowledge of ecological
processes in river ecosystems. It enables practitioners
of conservation planning to explore alternative
futures for regional river biodiversity, resulting from
various spatially explicit management scenarios.
The reliability of the quantitative estimates of the
persistence of river biodiversity can be improved
substantially from this first application. There is a
need to refine estimates of biodiversity condition and
develop methods of accounting for the spatial config-
uration of remaining habitats. The influence of the
direction and rates of change in disturbance on
Planning for persistence of river biodiversity 53
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

biodiversity condition and the likely end points of
restoration activities must also be taken into consid-
eration. These – together with refinements of bio-
diversity surrogates – are likely to greatly improve the
utility of our methods in freshwater conservation
planning.
Acknowledgments
This work was partly funded by the Australian
Commonwealth under the Natural Heritage Trust
Funding. We thank Klaus Koop for comments on the
manuscript.
References
Abell R., Allan J.D. & Lehner B. (2007) Unlocking the
potential of protected areas for freshwaters. Biological
Conservation, 134, 48–63.
Allnutt T.F., Ferrier S., Manion G. et al. (2008) A method
for quantifying biodiversity loss and its application to
a 50-year record of deforestation across Madagascar.
Conservation Letters, 1, 173–181.
Arponen A., Heikkinen R.K., Thomas C.D. & Moilanen
A. (2005) The value of biodiversity in reserve selection:
representation, species weighting, and benefit func-
tions. Conservation Biology, 19, 2009–2014.
Arrhenius O. (1921) Species and area. Journal of Ecology,
9, 95–99.
Brooks T.M., Bakarr M.I., Boucher T. et al. (2004) Cover-
age provided by the Global Protected-Area System: is
it enough? BioScience, 54, 1081–1091.
Canty A. & Ripley B.D. (2009) boot: Bootstrap R (S-PLUS)
Functions. R package version 1.2–35. Available at:
http://CRAN.R-project.org/package=boot.(lastaccessed
on 30 December 2009).
Davies P.E. (2000) Development of the National River
Bioassessment System (AUSRIVAS) in Australia. In:
Assessing the Biological Quality of Freshwaters: RIVPACS
and Other Techniques (Eds J.F. Wright, D.W. Sutcliffe, &
M.T. Furse), pp. 113–124. Freshwater Biological Asso-
ciation, Cumbria, UK.
DECC (2007a) Lord Howe Island Biodiversity Management
Plan. NSW Department of Environment and Climate Change.
Available at: http://www.environment.nsw.gov.au/
parkmanagement/LordHoweBiodiversityMgmtplan
Draft.htm. (last accessed on 30 December 2009).
DECC (2007b) 100m Land use Grid for Hunter-Central
Rivers CMA (ANZLIC v1.2 Metadata Standard). NSW
Department of Environment and Climate Change.
Sydney.
DECC (2009) Working Draft Northern Rivers Region Biodi-
versity Management Plan, National Recovery Plan for the
Northern Rivers Region. Department of Environment
and Climate Change NSW, Sydney.
Desmet P. & Cowling R. (2004) Using the species-area
relationship to set baseline targets for conservation.
Ecology and Society 9: 11. Available at: http://www.
ecologyandsociety.org/vol9/iss2/art11/ (last accessed
on 30 December 2009).
Diamond J. & May R. (1976) Island biogeography and the
design of natural reserves. In: Theoretical Ecology:
Principles and Applications (Ed. R. May), pp. 163–186.
Blackwell Scientific Publications, Oxford, UK.
Drielsma M. & Ferrier S. (2006) Landscape scenario
modelling of vegetation condition. Ecological Manage-
ment and Restoration, 7, S45–S52.
Dudgeon D., Arthington A.H., Gessner M.O. et al. (2006)
Freshwater biodiversity: importance, threats, status and
conservation challenges. Biological Reviews, 81, 163–182.
ESRI (1999) ArcView Version 3.2. Environmental Systems
Research Institute, Inc., Redlands, CA.
ESRI (2005) ArcGIS Version 9.1. Environmental Systems
Research Institute, Inc, Redlands, CA.
Fagan W.F. (2002) Connectivity, fragmentation and
extinction risks in dendritic populations. Ecology, 83,
3243–3249.
Faith D.P. (1995) Biodiversity and Regional Sustainability
Analysis. CSIRO Division of Wildlife and Ecology,
Lyneham, Australia.
Faith D.P., Ferrier S. & Williams K.J. (2008) Getting
biodiversity intactness indices right: ensuring that
‘biodiversity’ reflects ‘diversity’. Global Change Biology
14, 207–217.
Ferrier S. (2005) An Integrated Approach to Addressing
Terrestrial Biodiversity in NRM Investment Planning and
Evaluation Across State, Catchment and Property Scales.
NSW Department of Environment and Conservation,
February 2005. Available at: http://www.nrc.nsw.gov.
au/content/documents/Submission%20-%20ST%20-%
20DEC%20Biodiversity.pdf (last accessed on 30 Dec-
ember 2009).
Ferrier S., Powell G.V.N., Richardson K.S. et al. (2004)
Mapping more of terrestrial biodiversity for Global
Conservation Assessment. BioScience, 54, 1101–1109.
Ferrier S., Faith D.P., Arponen A. & Drielsma M. (2009)
Community-level approaches to spatial conservation
prioritization. In: Spatial Conservation Prioritization:
Quantitative Methods and Computational Tools (Eds. A.
Moilanen, H. Possingham, K. Wilson), pp 94–109.
Oxford University Press, Oxford.
HCR CMA (2007) Hunter-Central Rivers Catchment Action
Plan. Hunter-Central Rivers Catchment Management
54 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

Authority, New South Wales, Australia. Available at:
http://www.hcr.cma.nsw.gov.au/catchmentactionplan.
php3 (last accessed on 30 December 2009).
Heino J. & Korsu K. (2008) Testing species-stone area and
species-bryophyte cover relationships in riverine macr-
oinvertebrates at small scales. Freshwater Biology, 53,
558–568.
HoseG.&TurakE.(2006)RiverHealth intheNewSouthWales
Lower North Coast, Hunter and Central Coast Catchments.
River Health Bioassessment Report No. 41 Department
of the Environment and Heritage, and NSW Department
of Environment and Conservation. Available at: http://
www.environment.gov.au/water/publications/envi
ronmental/rivers/nrhp/catchments-nsw/index.html
(last accessed on 30 December 2009).
Hutchinson M.F., Stein J. & Stein J. (2001) Upgrade of the 9
Second Australian Digital Elevation Model. A Joint Project
of CRES and AUSLIG. Retrieved 2nd, August, 2004.
Available at: http://cres.anu.edu.au/dem/index.
php#4 (last accessed on 30 December 2009).
Lake P.S., Bond N. & Reich P. (2007) Linking ecological
theory with stream restoration. Freshwater Biology, 52,
597–615.
Linke S. & Norris R.H. (2003) Biodiversity: bridging the
gap between condition and conservation. Hydrobiolo-
gia, 500, 203–211.
Linke S., Pressey R.L., Bailey R.C. & Norris R.H. (2007)
Management options for river conservation planning.
Freshwater Biology, 5, 918–938.
Malmqvist B. & Rundle S. (2002) Threats to running
water ecosystems of the world. Environmental Conser-
vation, 29, 134–153.
Marchant R., Ryan D. & Metzeling L. (2006) Regional and
local species diversity patterns for lotic invertebrates
across multiple drainage basins in Victoria. Marine and
Freshwater Research, 47, 675–684.
Margules C.R. & Pressey R.L. (2000) Systematic conser-
vation planning. Nature, 405, 243–253.
McNeely J.A.Ed. (1993) Parks for Life: Report of the IVth
World Congress on National Parks and Protected Areas.
IUCN Communications Division, Gland (Switzerland).
Moffett A. & Sarkar S. (2006) Incorporating multiple
criteria into the design of conservation area networks:
a mini review with recommendations. Diversity and
Distributions, 12, 125–137.
Moilanen A., Leathwick J. & Elith J. (2008) A method for
spatial freshwater conservation prioritization. Freshwa-
ter Biology, 53, 577–592.
Nel J.L., Roux D.J., Maree G., Kleynhans C.J., Moolman J.,
Reyers B., Rouget M. & Cowling R.M. (2007) Rivers in
peril inside and outside protected areas: a systematic
approach to conservation assessment of river ecosys-
tems. Diversity and Distributions, 13, 341–352.
Peterson G.D., Cumming G.S. & Carpenter S.R. (2003)
Scenario planning: a tool for conservation in an
uncertain world. Conservation Biology, 17, 358–366.
Peterson E.E., Theobald D.M. & Ver Hoef J.M. (2007)
Geostatistical modelling on stream networks: develop-
ing valid covariance matrices based on hydrologic
distance and stream flow. Freshwater Biology, 52, 267–
269.
Pfafstetter O. (1989) Classification of hydrographic
basins: coding methodology, unpublished manuscript,
Departamento Nacional de Obras de Saneamento,
August 18, 1989, Rio de Janeiro; available from J.P.
Verdin, U.S. Geological Survey, EROS Data Center,
Sioux Falls, South Dakota 57198 USA.
Possingham H.P., Andelman S.J., Noon B.R., Trombulak
S. & Pulliam H.R. (2001) Making smart conservation
decisions. In: Conservation Biology: Research Priorities for
the Next Decade (Eds M.E. Soule & G.H. Orians), pp.
225–244. Island Press, Washington, DC.
Pressey R.L., Cowling R.M. & Rouget M. (2003) Formu-
lating conservation targets for biodiversity pattern and
process in the Cape Floristic Region, South Africa.
Biological Conservation, 112, 99–127.
R Development Core Team (2004) R: A Language and
Environment for Statistical Computing. R Foundation for
Statistical Computing, Vienna. ISBN 3-900051-00-3.
Available at: http://www.R-project.org (last accessed
30 December 2009).
Rodrigues A.S.L. & Gaston K.J. (2001) How large do
reserve networks need to be? Ecology Letters, 4, 602–
609.
Rosenzweig M.L. (1995) Species Diversity in Space and
Time. Cambridge University Press, Cambridge, UK.
Rosenzweig M.L. (2003) Reconciliation ecology and the
future of species diversity. Oryx, 37, 194–205.
Sarkar S. & Margules C. (2002) Operationalizing biodi-
versity for conservation planning. Journal of Biosciences,
27, 299–308.
Sarkar S., Pressey R.L., Faith D.P. et al. (2006) Biodiver-
sity conservation planning tools: present status and
challenges for the future. Annual Review of Environment
and Resources, 31, 123–159.
Sarr D.A. (2002) Riparian livestock exclosure research in
the Western United States: a critique and some
recommendations. Environmental Management, 30,
516–526.
Sepkoski J.J. Jr & Rex M.A. (1974) Distribution of
freshwater mussels: coastal rivers as biogeographic
islands. Systematic Zoology, 23, 165–188.
Simpson J.C. & Norris R.H. (2000) Biological assessment
of river quality: development of AUSRIVAS models
and outputs. In: Assessing the Biological Quality
of Freshwaters: RIVPACS and Other Techniques (Eds
Planning for persistence of river biodiversity 55
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56

J.F. Wright, D.W. Sutcliffe, & M.T. Furse), pp. 143–
163. Freshwater Biological Association, Cumbria,
UK.
Soule M.E. & Sanjayan M.A. (1998) Conservation targets:
do they help? Science 279, 2060–2061.
Stein J.L. (2005) A national landscape framework for river
and stream conservation: developing a new stream
and catchment reference system. In: Fourth Australian
Stream Management Conference: linking rivers to land-
scapes. 19–22 October 2004 (Eds I.D. Rutherford, I.
Wiszniewski, M. Askey-Doran & R. Glazik), pp. 548–
552. Department of Primary Industries, Water and the
Environment, Launceston Tasmania.
Stein J.L. (2007) A Continental Landscape Framework for
Systematic Conservation Planning for Australian Rivers
and Streams. PhD Thesis, Centre for Resource and
Environmental Studies, Australian National Univer-
sity, Canberra.
Stein J.L., Stein J.A. & Nix H.A. (1998) The Identification of
Wild Rivers: Methodology and Database Development.
Environment Australia, Canberra. Available at:
http://www.environment.gov.au/heritage/publications/
anlr/wild-river-identification.html (last accessed on 30
December 2009).
Stein J.L., Stein J.A. & Nix H.A. (2002) Spatial analysis of
anthropogenic river disturbance at regional and con-
tinental scales: identifying the wild rivers of Australia.
Landscape and Urban Planning, 60, 1–25.
Thieme M., Lehner B., Abell R.A., Hamilton S.K.,
Kellndorfer J., Powell G. & Riveros J.C. (2007) Fresh-
water conservation planning in data poor areas: an
example from a remote Amazonian basin (Madre de
Dios River, Peru and Bolivia). Biological Conservation
135, 500–517.
Thomas C.D., Cameron A., Green R.E. et al. (2004) Extinc-
tion risk from climate change. Nature, 427, 145–148.
Turak E. (2007) An Ecological Typology of the Rivers of New
South Wales, Australia. PhD thesis. University of Tech-
nology Sydney, NSW Australia.
Turak E. & Koop K. (2003) Use of rare macroinvertebrate
taxa and multiple-year data to detect low-level impacts
in rivers. Aquatic Ecosystem Health and Management, 6,
167–175.
Turak E. & Koop K. (2008) A multi-attribute ecological
river typology for assessing river condition and con-
servation planning. Hydrobiologia, 603, 83–104.
Turak E., Flack L.K., Norris R.H., Simpson J. & Waddell
N. (1999) Assessment of river condition at a large
spatial scale using predictive models. Freshwater Biol-
ogy, 41, 283–298.
Turak E., Hose G. & Waddell N. (2002) Australia-wide
assessment of River Health: New South Wales Bioassessment
Report (NSW Final Report), Monitoring River Health Initia-
tive Technical Report no 2a. Commonwealth of Australia
and NSW Environment Protection Authority, Canberra
and Sydney. Available at: http://www.environment.
gov.au/water/publications/environmental/rivers/nrhp/
nsw/index.html (last accessed on 30 December 2009).
Turak E., Waddell N. & Johnstone G. (2004). New South
Wales Australian River Assessment System (AUSRIVAS)
Sampling and Processing Manual. Commonwealth of
Australia and NSW Department of Environment
and Conservation, Canberra and Sydney. Available
at: http://ausrivas.canberra.edu.au/Bioassessment/
Macroinvertebrates/Man/Sampling/NSW/NSW_Aus
rivas_protocol_Version2_2004.pdf (last accessed on 30
December 2009).
Venables W.N. & Ripley B.D. (2002) Modern Applied
Statistics with S, 4th edn. Springer, New York.
Verdin K. & Verdin J.P. (1999) A topological system for
delineation and codification of the Earth’s river basins.
Journal of Hydrology, 218, 1–12.
Wood S.N. (2006) Generalized Additive Models, An Intro-
duction with R. Chapman and Hall ⁄CRC Taylor and
Francis Group, New York.
Zurlini G., Grossi L. & Rossi O. (2002) Spatial-accumu-
lation pattern and extinction rates of Mediterranean
flora as related to species confinement to habitats in
preserves and larger areas. Conservation Biology, 16,
948–963.
(Manuscript accepted 8 December 2009)
56 E. Turak et al.
� 2010 Blackwell Publishing Ltd, Freshwater Biology, 56, 39–56




















