
Pitheciids in fragmented habitats: Land cover change and its implications for conservation
16
RESEARCH ARTICLE Pitheciids in Fragmented Habitats: Land Cover Change and Its Implications for Conservation SARAH A. BOYLE* Department of Biology, Rhodes College, Tennessee Pitheciids (Cacajao, Callicebus, Chiropotes, and Pithecia) have experienced habitat loss and fragmentation across their geographic range in South America. Some populations living in habitat fragments live in smaller groups, travel shorter distances, and consume items that are not regularly found in the diets of populations living in continuous habitat; however, these patterns are not consistent across species. I used the IUCN Red List of Threatened Species to delineate the geographic range and conservation status of 43 pitheciid species. I calculated the amount of modified land cover within the range of each species, as well as the extent to which the remaining habitat exists in small fragments and the amount of forest lost from 2000 to 2012. Mean forest fragment size ranged from 12 to 12,027 ha, and mean forest loss from 2000 to 2012 ranged from 10.7% for Chiropotes to 0.9% for Pithecia. Critically Endangered and Endangered species represented 20.9% of the pitheciid species, and 46.5% of these species had population trends documented as decreasing. Total modified land cover was greatest for Callicebus species (18.0% of geographic range), followed by Chiropotes (13.8%), Pithecia (4.4%), and Cacajao (1.1%). Species of greater conservation concern had smaller geographic ranges, and a greater percentage of their range consisting of modified land cover than species of lower conservation concern. Species of greater conservation concern also had a greater percentage of forest lost from 2000 to 2012 and a smaller percentage of the remaining forest being protected. Most studies of pitheciids in fragments have concentrated on census data; the behavior of pitheciids in fragments has been examined for only 9 of the 43 species. Increased data on the responses of pitheciid species to forest loss and fragmentation are necessary in order to address pitheciid conservation, especially in areas undergoing severe habitat loss. Am. J. Primatol. © 2014 Wiley Periodicals, Inc. Key words: Cacajao; Callicebus; Chiropotes; habitat fragmentation; Pithecia INTRODUCTION Globally most of the world’s habitats primarily exist as small fragments [Pimm and Brooks, 2013]. Habitat loss, degradation, and fragmentation, as well as hunting, are the main threats to primate conserva- tion [Oates, 2013]. Forest fragmentation can impact the distribution of primates [Hickey et al., 2013], and it has been correlated with increased parasite prevalence [Gillespie and Chapman, 2006], lowered genetic diversity [Chaves et al., 2011; Craul et al., 2009], and local extinction [Benchimol and Peres, 2014]. Studies [Anzures‐Dadda and Manson, 2007; Harcourt and Doherty, 2005; Johns and Skorupa, 1987; Sharma et al., 2014] have found one or a combination of habitat characteristics (e.g. fragment size, isolation) and biological characteristics of the species (e.g. home range, diet) to predict a primate’s presence in a fragment (or, alternatively, vulnerability for local extinction). However, there can be variations across taxa [Benchimol and Peres, 2014], and such correla- tions between primate traits and presence are not always evident [Gibbons and Harcourt, 2009]. The pitheciids (Cacajao, Callicebus, Chiropotes, and Pithecia) face pressures of habitat loss and fragmentation throughout their range in South America [Ferrari et al., 2013a,b; Lehman et al., 2013; Porter et al., 2013]. The factors driving habitat loss and fragmentation differ by geographic area, but include large‐scale agriculture [García et al., 2010; Lehman et al., 2006; Rowe and Martínez, 2003], mining [Lehman et al., 2013], oil Conflict of interest: None Special Issue: Progressing Pitheciids: Recent Advances in the Study of Cacajao, Callicebus, Chiropotes, and Pithecia—Papers by which to remember Liza Maria Veiga (1963–2012). Correspondence to: Sarah A. Boyle, Department of Biology Rhodes College 2000 North Parkway Memphis, TN 38112. E‐mail: [email protected] Received 1 May 2014; revised 26 June 2014; revision accepted 5 August 2014 DOI: 10.1002/ajp.22325 Published online XX Month Year in Wiley Online Library (wileyonlinelibrary.com). American Journal of Primatology © 2014 Wiley Periodicals, Inc.
Transcript of Pitheciids in fragmented habitats: Land cover change and its implications for conservation

RESEARCH ARTICLE
Pitheciids in Fragmented Habitats: Land Cover Change and Its Implications forConservation
SARAH A. BOYLE*Department of Biology, Rhodes College, Tennessee
Pitheciids (Cacajao, Callicebus, Chiropotes, and Pithecia) have experienced habitat loss andfragmentation across their geographic range in South America. Some populations living in habitatfragments live in smaller groups, travel shorter distances, and consume items that are not regularlyfound in the diets of populations living in continuous habitat; however, these patterns are not consistentacross species. I used the IUCN Red List of Threatened Species to delineate the geographic range andconservation status of 43 pitheciid species. I calculated the amount of modified land cover within therange of each species, as well as the extent to which the remaining habitat exists in small fragments andthe amount of forest lost from 2000 to 2012. Mean forest fragment size ranged from 12 to 12,027ha, andmean forest loss from 2000 to 2012 ranged from 10.7% for Chiropotes to 0.9% for Pithecia. CriticallyEndangered and Endangered species represented 20.9% of the pitheciid species, and 46.5% of thesespecies had population trends documented as decreasing. Total modified land cover was greatest forCallicebus species (18.0% of geographic range), followed by Chiropotes (13.8%), Pithecia (4.4%), andCacajao (1.1%). Species of greater conservation concern had smaller geographic ranges, and a greaterpercentage of their range consisting of modified land cover than species of lower conservation concern.Species of greater conservation concern also had a greater percentage of forest lost from2000 to 2012 anda smaller percentage of the remaining forest being protected. Most studies of pitheciids in fragmentshave concentrated on census data; the behavior of pitheciids in fragments has been examined for only 9 ofthe 43 species. Increased data on the responses of pitheciid species to forest loss and fragmentation arenecessary in order to address pitheciid conservation, especially in areas undergoing severe habitat loss.Am. J. Primatol. © 2014 Wiley Periodicals, Inc.
Key words: Cacajao; Callicebus; Chiropotes; habitat fragmentation; Pithecia
INTRODUCTION
Globally most of the world’s habitats primarilyexist as small fragments [Pimm and Brooks, 2013].Habitat loss, degradation, and fragmentation, as wellas hunting, are the main threats to primate conserva-tion [Oates, 2013]. Forest fragmentation can impactthe distribution of primates [Hickey et al., 2013], and ithas been correlatedwith increasedparasite prevalence[Gillespie and Chapman, 2006], lowered geneticdiversity [Chaves et al., 2011; Craul et al., 2009], andlocal extinction [Benchimol and Peres, 2014]. Studies[Anzures‐Dadda and Manson, 2007; Harcourt andDoherty, 2005; Johns and Skorupa, 1987; Sharmaet al., 2014] have found one or a combination of habitatcharacteristics (e.g. fragment size, isolation) andbiological characteristics of the species (e.g. homerange, diet) to predict a primate’s presence in afragment (or, alternatively, vulnerability for localextinction). However, there can be variations acrosstaxa [Benchimol and Peres, 2014], and such correla-tions between primate traits and presence are notalways evident [Gibbons and Harcourt, 2009].
The pitheciids (Cacajao, Callicebus, Chiropotes,and Pithecia) face pressures of habitat loss andfragmentation throughout their range in SouthAmerica [Ferrari et al., 2013a,b; Lehmanet al., 2013; Porter et al., 2013]. The factors drivinghabitat loss and fragmentation differ by geographicarea, but include large‐scale agriculture [Garcíaet al., 2010; Lehman et al., 2006; Rowe andMartínez, 2003], mining [Lehman et al., 2013], oil
Conflict of interest: NoneSpecial Issue: Progressing Pitheciids: Recent Advances in theStudy of Cacajao, Callicebus, Chiropotes, and Pithecia—Papersby which to remember Liza Maria Veiga (1963–2012).�Correspondence to: Sarah A. Boyle, Department of BiologyRhodes College 2000 North Parkway Memphis, TN 38112.E‐mail: [email protected]
Received 1 May 2014; revised 26 June 2014; revision accepted 5August 2014
DOI: 10.1002/ajp.22325Published online XX Month Year in Wiley Online Library(wileyonlinelibrary.com).
American Journal of Primatology
© 2014 Wiley Periodicals, Inc.

and gas development [Porter et al., 2013; Suárezet al., 2009], dams [Ferrari et al., 2013a], and logging[Martinez and Wallace, 2007]; some areas are undermultiple pressures [Urbani, 2006].
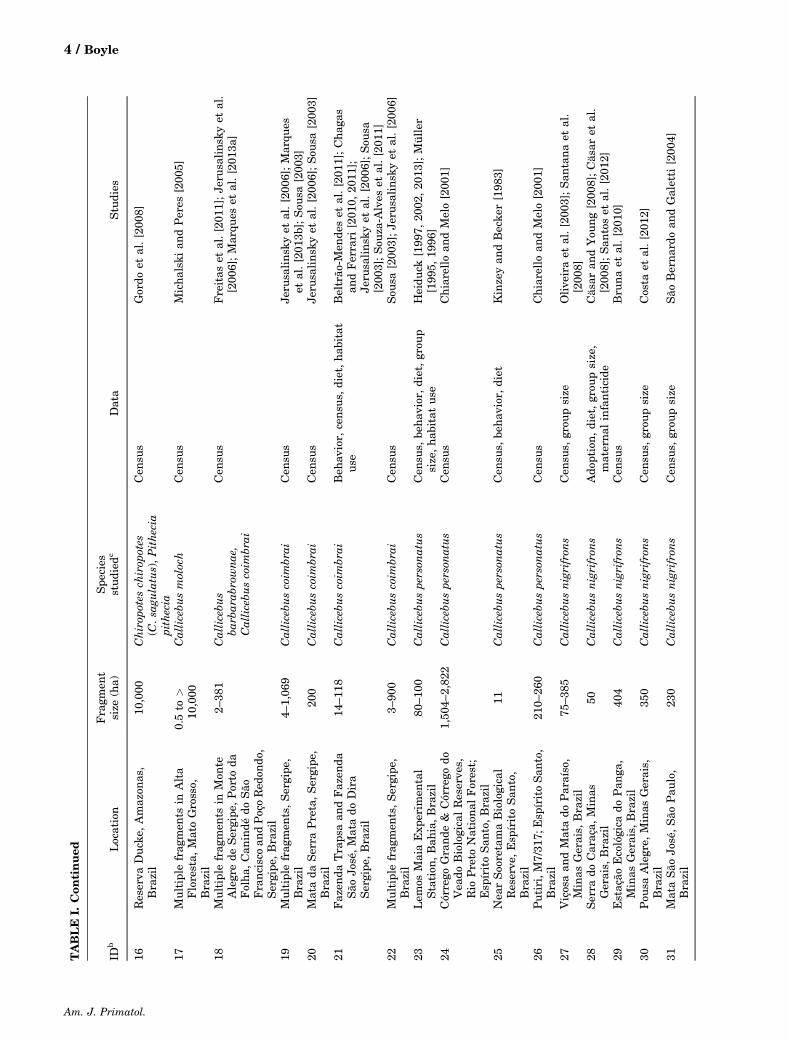
Three of the four‐pitheciid genera (Callicebus,Chiropotes, and Pithecia; Table I) have been studiedin fragmented habitats in six countries, with themajority of these studies occurring in Brazil (Fig. 1).To date there have been no published studies ofCacajao in forest fragments [Ferrari et al., 2013b;Peres, 2001]. Four study sites (Lago Gurí in Bolívar,Venezuela; Biological Dynamics of Forest FragmentsProject in Amazonas, Brazil; Tucuruí Dam in Pará,Brazil; and Lemos Maia Experimental Station inBahia, Brazil) have been the most‐studied areas todate, yet these sites represent only a handful ofpitheciid species (Table I). Of the 43 pitheciid specieslisted by the IUCN, only 14 species have been studiedin habitat fragments (with only 9 of these specieshaving their behavior documented in fragments).
Callicebus,Chiropotes, andPithecia have all beenfound in habitat fragments that are smaller than theanimals’ typical home range size in continuous forest.For example, Callicebus spp. have home ranges upto 48ha [Bicca‐Marques and Heymann, 2013] buthave been found in fragments as small as 2–5ha[DeLuycker, 2012; Ferrari et al., 2013b; Mark, 2003];home ranges of 0.3–0.5ha were calculated for threegroups of Callicebus in Peru [Mason, 1968], but thesehome range estimates were based on four days of datacollection. Chiropotes spp. have home ranges up to1000ha in continuous forest [Gregory, 2011;Shaffer, 2013], but have lived for multiple yearsin fragments as small as 10–20ha [Boyle andSmith, 2010a; Silva and Ferrari, 2009]. Home rangesize reported forPithecia spp. varies from 10 to 204ha[Palminteri andPeres, 2012; Peres, 1993] butPitheciagroups have lived in fragments as small as 10ha[Setz, 1994].
Comparative studies of groups living in habitatfragments with groups in continuous forest (or largefragments >1000ha) have been limited, with resultsvarying by genus [Boyle and Smith, 2010b; Schwarz-kopf and Rylands, 1989]. For example, Chiropotes spp.have demonstrated significant modifications to theirbehavior and ecology in small (<20ha) forest frag-ments: they occurred in smaller groups and at greaterdensities, rested more and traveled less, consumedplant species that were not part of diet in thecontinuous forest, and revisited more feeding treesthroughout the day thanChiropotes spp. in continuousforest [Boyle et al., 2009, 2012; Boyle andSmith, 2010a; Silva and Ferrari, 2009; Veiga, 2006].However,Wagner etal. [2009] didnotfinda correlationbetween fragment size andCallicebus ornatus density;however, all fragments were small (�40ha). Incomparisons with other published studies, group sizeof Pithecia pithecia chrysocephala did not differbetween fragments and continuous forest [Setz, 1994].
The objectives of this study were to quantify landcover throughout the pitheciids’ ranges, test if landcover disturbance and extent of habitat fragmenta-tion are correlatedwith a species’ conservation status,and quantify the extent to which the remaining forestis protected in the species’ ranges. I predicted thatspecies of high conservation concern (i.e. CriticallyEndangered, Endangered) would have ranges thathave been heavily modified by humans, and a greaterpercentage of their remaining habitat would exist infragments when compared to species of lowerconservation concern. These findings are importantfor targeting geographic areas and species that arecurrently or likely to become critical for immediateattention (e.g. conservation plans, research) based onthe species’ conservation status, patterns of habitatloss, and current knowledge of the species’ distribu-tion, population status, and behavior.
METHODSI used the IUCN Red List of Threatened Species
[IUCN, 2013] to delineate the geographic range,conservation status, and population trend of the 43pitheciid species listed by IUCN. There are currenttaxonomic disagreementswith some pitheciid species[Auricchio, 2010; Bonvicino et al., 2003; Boubliet al., 2008; Silva Júnior et al., 2013], so I followedthe conservation status and delineation of thegeographic ranges noted by the IUCN [IUCN, 2013]in order to remain consistent with the analyses. As ofJune 2014, 41 of the 43 species were last assessed in2008 (Callicebus caquetensis and C. oenanthe wereassessed in 2011). I used GlobCover 2009 (300‐mresolution) land cover data to quantify the amount ofland cover that had been modified (defined using theGlobCover categories of artificial, mosaic cropland/vegetation, mosaic vegetation/cropland, rainfed crop-lands, bare areas, and irrigated croplands) in eachspecies’ geographic range [Bontemps et al., 2011].Pitheciids use a variety of forest types [Barnettet al., 2013; Bicca‐Marques and Heymann, 2013;Norconk and Sez, 2013; Veiga and Ferrari, 2013], andmultiple species can use forest fragments �10ha(Table I), so I quantified the amount of forest withinthe entire geographic distribution of the pitheciidspecies (total area: 6,247,459km2) using global forestcover data [Hansen et al., 2013]. These forest datarepresent forest extent and forest change from 2000to 2012 using Landsat 7 ETMþ imagery at a spatialresolution of 30m. Therefore, the forest data allowedfor more detailed high‐resolution analysis [Boyleet al., 2014; Hansen et al., 2013] of forest coverpatterns (i.e. amount of forest cover and size of forestfragments) within the pitheciids’ ranges. I created amosaic of the 19 tiles representing the entiregeographic range of all pitheciids. The modifiedland cover data, forest cover data, and IUCN data
Am. J. Primatol.
2 / Boyle

TABLE
I.Stu
diesof
Pithec
iidsin
Fra
gmen
tedHab
itat
sa
IDb
Location
Fragm
ent
size
(ha)
Spe
cies
stud
iedc
Data
Studies
1Lag
oGuri,Bolívar,
Ven
ezuelad
12–36
5Chirop
otes
chirop
otes,
Pithecia
pithecia
Beh
avior,
cens
us,d
iet,grou
psize,ha
bitatus
e,pr
edation,
repr
oduc
tive
physiology
Cun
ning
ham
andJa
nson
[200
6];
Cun
ning
ham
etal.[201
3];G
leas
onan
dNorconk
[200
2];Kinzeyan
dNorconk
[199
3];N
orconk
[199
6,20
06];
Norconk
andCon
klin‐B
rittain[200
4];
Norconk
andGrafton
[200
3];P
eetz
[200
1];Urban
i[200
2]2
Vered
aSan
Isidro
deChichim
ene,
Meta,
Colom
bia
2–40
Callicebu
sornatus
Cen
sus,
grou
psize,h
abitat
use,
pred
ation
Wag
neret
al.[20
09]
3Haciend
aBarba
scal,ea
stof
San
Martín,M
eta,
Colom
bia
7Callicebu
sornatus
Beh
avior,
cens
us,g
roup
size,
habitatus
eMas
on[196
8]
4Multiplefrag
men
tsin
Caq
uetá,
Colom
bia
Not
prov
ided
Callicebu
sca
queten
sis
Cen
sus,
grou
psize
Defleret
al.[20
10];Garcíaet
al.[20
10]
5Moy
obam
ba,Tarap
oto,
Sap
osoa
andJu
anjuiarea
,San
Martín,
Peru
2–64
0Callicebu
soenan
the
Beh
avior,
birthing
beha
vior,
cens
us,diet
Bóv
eda‐Pen
alba
etal.[20
09];
DeL
uyck
er[200
6,20
12,20
14];Mark
[200
3]6
Faz
enda
Mon
teVerde
,Pará,
Brazile
200
Chirop
otes
satanas
Cen
susd,grou
psize
Ferrari
etal.[19
99];Lop
esan
dFerrari
[200
0]7
Faz
enda
São
Marcos,
Pará,
Brazil
10,000
Chirop
otes
satanas
Cen
sus,
grou
psize
Ferrari
etal.[19
99];Lop
esan
dFerrari
[200
0]8
Faz
enda
Bad
ajós,P
ará,
Brazil
8,00
0Chirop
otes
satanas
Cen
sus,
grou
psize
Ferrari
etal.[19
99];Lop
esan
dFerrari
[200
0]9
Celmar
plan
tation
complex
,Maran
hão
,Brazil
8–20
00Chirop
otes
satanas
Beh
avior,
cens
us,g
roup
size,
diet
Port‐Carva
lhoan
dFerrari
[200
4]
10Tucu
ruíReservo
ir,P
ará,
Brazild
16–1,30
0Callicebu
smoloch,Chirop
otes
satanas
,Chirop
otes
utahicki
Beh
avior,
cens
us,d
iet,grou
psize,ha
bitatus
e,pr
edation
Ferrari
etal.[20
04];Sam
paio
and
Ferrari
[200
5];S
antoset
al.[20
13];
Silva
andFerrari
[200
9];Veiga
[200
6];Veiga
andFerrari
[200
6]11
Faz
enda
Arataú,P
ará,
Brazil
7,50
0Chirop
otes
utahicki
Cen
sus,grou
psize,h
abitat
use
Bob
adilla
andFerrari
[199
8,20
00];
Ferrari
etal.[19
99,20
07]
12Tab
ocal
andSão
Ben
edito,
Pará,
Brazil
30–90
Callicebu
smoloch
Cen
sus
Ferrari
etal.[20
03];Gon
çalves
etal.
[200
3]13
Multiple
frag
men
tsalon
gSan
tarém‐C
uiabá
Highw
ay,
Pará,
Brazil
70–3,00
0Callicebu
smoloch,
Chirop
otes
albinas
us
Cen
sus
Ferrari
etal.[20
03]
14Multiple
frag
men
tsalon
gSan
tarém‐C
uiabá
Highw
ay,
Pará,
Brazil
20–2,50
0Callicebu
smoloch,
Chirop
otes
albinas
us
Cen
sus
Ferrari
etal.[20
03]
15BiologicalDyn
amicsof
Forest
Fragm
ents,Amaz
onas
,Brazil
1–10
0Chirop
otes
chirop
otes
(C.sa
gulatus),Pithecia
pithecia
Beh
avior,
cens
us,d
iet,grou
psize,ha
bitatus
e,pr
edation
Ayres
[198
1];B
oyle
etal.
[200
9,20
12,20
13];Boy
lean
dSmith
[201
0a,20
10b];Gilbe
rt[200
0];S
etz
[199
4];Setzan
dGas
par[199
7];S
etz
etal.[19
99]
Am. J. Primatol.
Pitheciids in Fragments / 3
TABLE
I.Con
tinued
IDb
Location
Fragm
ent
size
(ha)
Spe
cies
stud
iedc
Data
Studies
16Reserva
Duck
e,Amaz
onas
,Brazil
10,000
Chirop
otes
chirop
otes
(C.sa
gulatus),Pithecia
pithecia
Cen
sus
Gordo
etal.[20
08]
17Multiplefrag
men
tsin
Alta
Floresta,
MatoGrosso,
Brazil
0.5to
>10
,000
Callicebu
smoloch
Cen
sus
Micha
lski
andPeres
[200
5]
18Multiplefrag
men
tsin
Mon
teAlegrede
Sergipe
,Porto
daFolha
,Can
indé
doSão
Franciscoan
dPoçoRed
ondo
,Sergipe
,Brazil
2–38
1Callicebu
sba
rbar
abrownae
,Callicebu
scoim
brai
Cen
sus
Freitas
etal.[20
11];Je
rusa
lins
kyet
al.
[200
6];Marqu
eset
al.[20
13a]
19Multiple
frag
men
ts,Sergipe
,Brazil
4–1,06
9Callicebu
scoim
brai
Cen
sus
Jeru
salins
kyet
al.[20
06];Marqu
eset
al.[20
13b];Sou
sa[200
3]20
Matada
Serra
Preta,S
ergipe
,Brazil
200
Callicebu
scoim
brai
Cen
sus
Jeru
salins
kyet
al.[20
06];Sou
sa[200
3]
21Faz
enda
Traps
aan
dFaz
enda
São
José,M
atado
Dira
Sergipe
,Brazil
14–11
8Callicebu
scoim
brai
Beh
avior,
cens
us,d
iet,ha
bitat
use
Beltrão
‐Men
deset
al.[20
11];Cha
gas
andFerrari
[201
0,20
11];
Jeru
salins
kyet
al.[20
06];Sou
sa[200
3];Sou
za‐A
lves
etal.[20
11]
22Multiplefrag
men
ts,Sergipe
,Brazil
3–90
0Callicebu
scoim
brai
Cen
sus
Sou
sa[200
3];J
erus
alinsk
yet
al.[20
06]
23Lem
osMaiaExp
erim
ental
Station
,Bah
ia,B
razil
80–10
0Callicebu
spe
rson
atus
Cen
sus,
beha
vior,d
iet,grou
psize,ha
bitatus
eHeidu
ck[199
7,20
02,2
013];Müller
[199
5,19
96]
24Córrego
Grande
&Córrego
doVea
doBiologicalReserve
s,Rio
Preto
Nationa
lForest;
Esp
íritoSan
to,Brazil
1,50
4–2,82
2Callicebu
spe
rson
atus
Cen
sus
Chiarello
andMelo[200
1]
25Nea
rSooretamaBiological
Reserve
,Esp
íritoSan
to,
Brazil
11Callicebu
spe
rson
atus
Cen
sus,
beha
vior,d
iet
Kinzeyan
dBecke
r[198
3]
26Putiri,M
7/31
7;Esp
íritoSan
to,
Brazil
210–
260
Callicebu
spe
rson
atus
Cen
sus
Chiarello
andMelo[200
1]
27Viçosaan
dMatado
Paraíso,
Minas
Gerais,
Brazil
75–38
5Callicebu
snigrifron
sCen
sus,
grou
psize
Olive
iraet
al.[20
03];San
tana
etal.
[200
8]28
Serra
doCaraça,
Minas
Gerais,
Brazil
50Callicebu
snigrifron
sAdo
ption,
diet,grou
psize,
materna
linfanticide
Cäs
aran
dYou
ng[200
8];C
äsar
etal.
[200
8];San
toset
al.[20
12]
29Estação
Ecológica
doPan
ga,
Minas
Gerais,
Brazil
404
Callicebu
snigrifron
sCen
sus
Bru
naet
al.[20
10]
30Pou
saAlegre,
Minas
Gerais,
Brazil
350
Callicebu
snigrifron
sCen
sus,
grou
psize
Costa
etal.[20
12]
31MataSão
José,São
Pau
lo,
Brazil
230
Callicebu
snigrifron
sCen
sus,
grou
psize
São
Berna
rdoan
dGaletti
[200
4]
Am. J. Primatol.
4 / Boyle

(2008) were temporallymatched as closely as possible(years 2009, 2012, and 2008, respectively).
I determined the number of forest fragments �0.5 ha and mean size of these fragments in thegeographic range for each pitheciid species. Theminimum size of 0.5 ha was based on previouslypublished literature [Boyle and Smith, 2010b; Man-dujano et al., 2005]. I then calculated the degree offragmentation by determining the percentage ofthe remaining forest that was represented byforest fragments �10ha, �100ha, �1000ha, and�10,000ha. I downloaded the global distribution ofprotected areas from the World Database on Pro-tected Area [IUCN and UNEP, 2014], and thencalculated the percent of these remaining forestedareas that are within protected areas. I used ArcGIS10.1 for all spatial analyses.
I tested whether species of high conservationstatus (i.e. IUCN classifications of Critically Endan-gered, Endangered) were more likely to have agreater percentage of their ranges consisting ofmodified land cover and whether these species weremore likely to have a greater percentage of theirremaining forest consisting of forest fragments �10,100, 1000, and 10,000 ha using one‐tailed Pearson’scorrelations. These correlations did not include onespecies (Callicebus stephennashi) for which the IUCNstatus was Data Deficient. I tested if there was acorrelation between the percentage of forest lost from2000 to 2012 with the percentage of forest that wasprotected for each pitheciid species using a one‐tailedPearson’s correlation. I tested if there were differ-ences among the four‐pitheciid genera in the conser-vation status of their species using a one‐wayanalysis of variance. I used SPSS 20 for all statisticaltests. This research adhered to the American Societyof Primatologists principles for the ethical treatmentof primates.
Fig. 1. Geographic range of the pitheciids, extent ofmodified landcover, and locations of studies examining pitheciids in habitatfragments.
TABLE
I.Con
tinued
IDb
Location
Fragm
ent
size
(ha)
Spe
cies
stud
iedc
Data
Studies
32Parqu
eEstad
ual
daCan
tareira,
São
Pau
lo,
Brazil
7,91
7Callicebu
snigrifron
sCen
sus,
diet,ha
bitatus
eTreve
linet
al.[20
07]
33Multiplesitesin
San
taCru
zde
laSierra,
Bolivia
1–30
3Callicebu
sdon
acop
hilus
Cen
sus,
habitatus
ePyritzet
al.[20
10]
aDatainclud
edforfrag
men
ts�1
0,00
0ha
.bNum
bercorrespo
ndsto
site
numbe
rin
Figur
e1.
c Spe
cies
arelisted
basedon
theIU
CN
Red
List[IUCN,2
013]
forcompa
tibility
withtheIU
CN
geog
raph
icrang
es;c
ommon
syno
nymsarein
parentheses.
dIsland
form
edafterflooding
ofarea
forhy
droelectricda
mde
velopm
ent.
e Local
extinc
tion
ofChirop
otes
satanas
.
Am. J. Primatol.
Pitheciids in Fragments / 5

RESULTS
Land Cover in Pitheciids’ Geographic Ranges
Overall, 79.3% of the entire pitheciid rangeconsisted of forest and 14.3% modified land cover,with the remaining 6.4% cover consisting of grass-land, shrub, water, and sparse vegetation. Hotspots ofmodified land cover (Fig. 1) were concentrated alongthe Atlantic coast of Brazil (states: Sergipe, Bahia,Espírito Santo, Minas Gerais, Rio de Janeiro, and SãoPaulo), and also found in north‐east Brazil (states:Pará andMaranhão), central Brazil (states: Rondôniaand Mato Grosso), central Venezuela (state: Bolívar);central Colombia (departments: Meta and Caquetá),central Bolivia (department: Santa Cruz), and north‐central Paraguay (departments: Presidente Hayes,Boqueron, and Alto Paraguay). Within the pitheciidrange, there existed large variation in the amountof forest and modified land cover, with Callicebushaving the greatest percentage of modified landcover, followed by Chiropotes, Pithecia, and Cacajao(Table II; Fig. 2). Ten species had >10% of theirgeographic range consisting of modified cover; 6 ofthese 10 species had >60% (Table III).
Mean forest fragment size in2012varied from12ha(Callicebus coimbrai) to 12,027ha (Cacajao hosomi),with Cacajo and then Pithecia having the largest meanfragment sizes (4,058.8ha� 2683.5 and 1,641.6ha� 637.3, respectively), followed by Callicebus (991.9ha� 306.1) and then Chiropotes (240.3ha� 108.7).
Overall, pitheciid species lost 5.3%� 0.8 offorested area from 2000 to 2012. The areas of greatestforest loss were concentrated in the eastern andsouthern Brazilian Amazon, central Bolivia, Para-guayan Chaco, and southern Colombia (Fig. 3).Chiropotes had the greatest average forest lossfrom 2000 to 2012 (10.7%� 0.04), followed byCallicebus (5.9%� 0.9), Cacajao (1.0%� 0.6), andPithecia (0.9%� 0.2). Overall there was a correlationin that species of greater conservation concernexperienced greater forest loss from 2000 to 2010(N¼ 42, r¼ 0.60, P< 0.001); however, 3 species listedas Least Concern had had �10% of forest lost from2000 to 2010 (Table IV).
Due to large tracts of continuous forest, all of theCacajao species and all of the Pithecia species had95% or more of the forest remaining in areas>10,000ha. For the other two genera, all of theChiropotes species had �88% of the forest in largeforests >10,000, and 73.3% of Callicebus species(22 of 30) had �90% of the forest remaining in areas>10,000. However, two species hadmore than 50% ofthe remaining forest distributed in fragments�10,000ha (Callicebus coimbrai and C. nigrifrons;Table II). In fact, nearly 13% of the remaining forestin the geographic range of C. coimbrai consisted offragments �10ha (Table II).
In 2012, 33.1%� 3.1 of the forest in the pitheciidspecies’ ranges was in a protected area. The percentprotected ranged from <1% protected (Cacajao
TABLE II. Pitheciid IUCN Conservation Status and Land Cover
Cacajao Callicebus Chiropotes Pithecia
Number species listeda 4 30 4 5Conservation status
% Critically endangered 0 10 25 0% Endangered 0 10 50 0% Vulnerable 75 13.3 0 20% Near threatened 0 3.3 0 0% Least concern 25 60 25 80% Data deficient 0 3.33 0 0
Population trend% Stable 25 6.7 25 20% Decreasing 75 40 75 40% Unknown 0 53.3 0 40
Land cover in geographic rangeb
% ForestEntire genus rangec 96.3 77.4 86.5 93.8Mean range for species (�SE) 96.5� 0.5 75.4�5.3 80.2� 6.4 91.8�3.2Range for species 95.7–97.8 8.3–97.6 63.8–91.5 79.3–96.2
% ModifiedEntire genus rangec 1.1 16.3 8.0 2.9Mean range for species (�SE) 1.1�0.2 18.0�4.8 13.8� 5.6 4.4�2.5Range for species 0.4–1.2 0.3–76.9 1.8–26.6 1.1–14.6
aNumber of species and the species’ geographic range follows the IUCN Red List [IUCN, 2013].bRemaining land cover categories consist of water, grassland, shrub, and sparse vegetation; values provided are based on each species’ values. Land coverclasses follow the GlobCover [Bontemps et al., 2011] classification.cGenus range includes species with overlapping ranges and some species’ ranges are larger than others, hence the division between the total range for thegenus and the mean range for the species within the genus.
Am. J. Primatol.
6 / Boyle
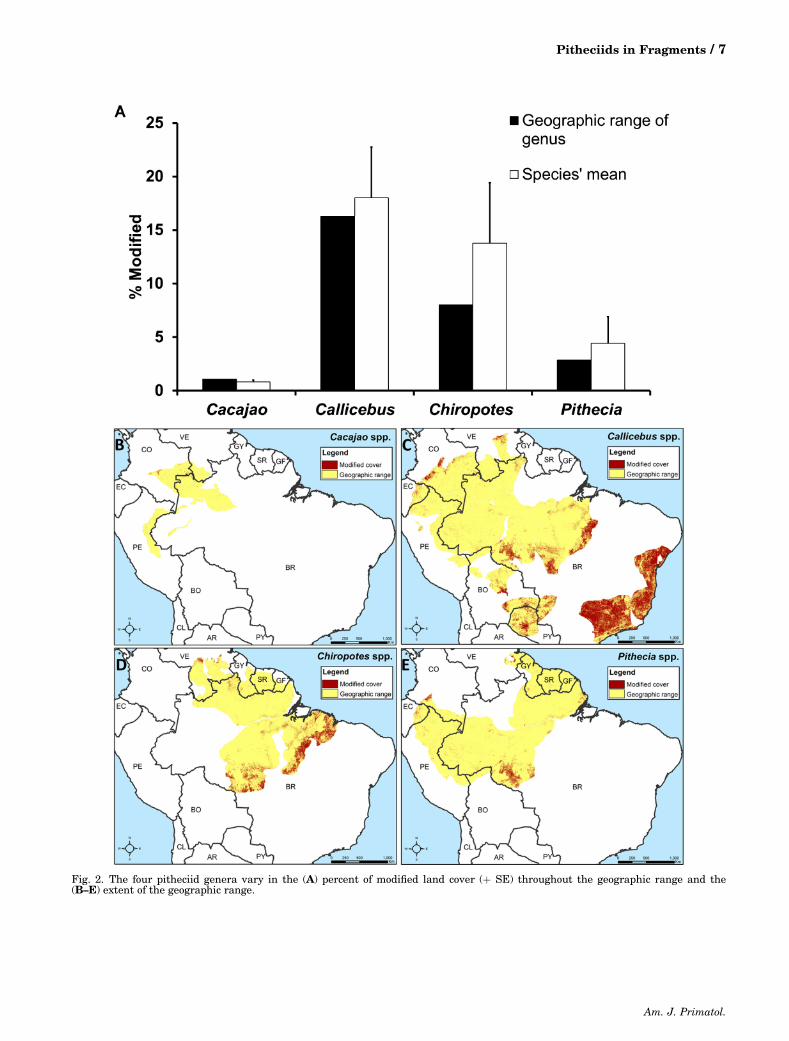
Fig. 2. The four pitheciid genera vary in the (A) percent of modified land cover (þ SE) throughout the geographic range and the(B–E) extent of the geographic range.
Am. J. Primatol.
Pitheciids in Fragments / 7
ayresi,Callicebus caquetensis,C. olallae,C. oenanthe)to 83.2% protected (Cacajao hosomi). Of the 43‐pitheciid species, 18 (41.9%) had �25% of its 2012forest in a protected area. Of these 18 species, 7 werelisted as Critically Endangered or Endangered.There were correlations in that species with lowerpercentage of the remaining forest protected hadgreater forest loss from 2000 to 2012 (N¼ 43,r¼�0.36, P¼ 0.01), smaller mean forest fragmentsize (N¼ 43, r¼ 0.43, P¼ 0.002), and the pitheciidspecies were classified as being in greater conserva-tion concern (N¼ 42, r¼�0.55, P< 0.001).
Conservation Status of Pitheciids
Critically Endangered and Endangered speciesrepresented 20.9% of the 43 pitheciid species, butCallicebus (C. barbarabrownae, C. caquetensis, C.oenanthe) and Chiropotes (C. satanas) were the onlygenera in these two categories (Table I). Populationtrends for the 43 species were listed as Decreasing(46.5%), Stable (11.6%), and Unknown (41.9%). Thefour pitheciid genera did not statistically differ intheir conservation status (F(38,3)¼ 1.8,P¼ 0.16). Only55.8% of pitheciid species were classified as LeastConcern by the IUCN.
The conservation status of a species and thepercentage of modified land cover in its range wascorrelated (N¼ 42, r¼�0.52, P< 0.001), with Criti-cally Endangered species having 44.9%� 15.3 oftheir range modified, and species classified as LeastConcern having 4.5%� 1.1 of their range modified.The conservation status was also correlated withgeographic range size (N¼ 42, r¼ 0.3, P¼ 0.03), withspecies of high conservation concern having smallerranges than species of lower conservation concern.Species of greater conservation concern had more ofthe remaining forest distributed in forest fragments�10ha (N¼ 42, r¼ 0.54, P< 0.001),�100ha (N¼ 42,r¼ 0.50, P< 0.001), �1,000ha (N¼ 42, r¼ 0.49,P< 0.001), and �10,000 (N¼ 42, r¼ 0.51,P< 0.001), but there was not a correlation betweenthe mean size of the forest fragments and conserva-tion status (N¼ 42, r¼�0.18, P¼ 0.12).
DISCUSSION
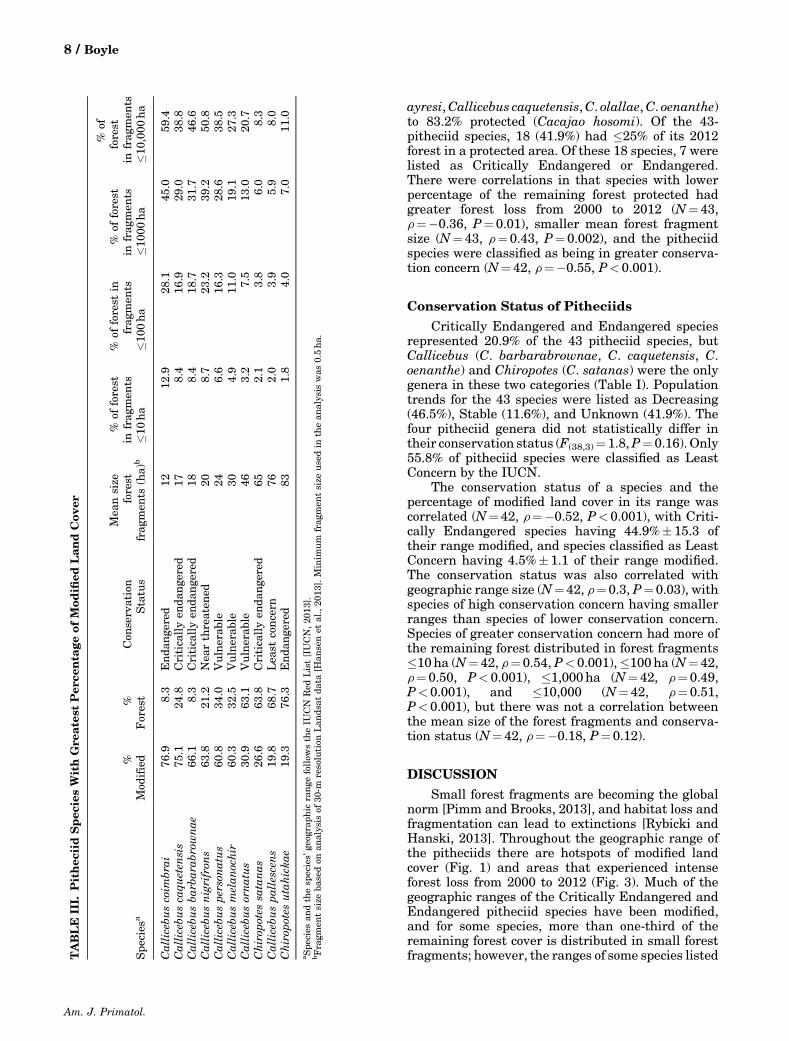
Small forest fragments are becoming the globalnorm [Pimm and Brooks, 2013], and habitat loss andfragmentation can lead to extinctions [Rybicki andHanski, 2013]. Throughout the geographic range ofthe pitheciids there are hotspots of modified landcover (Fig. 1) and areas that experienced intenseforest loss from 2000 to 2012 (Fig. 3). Much of thegeographic ranges of the Critically Endangered andEndangered pitheciid species have been modified,and for some species, more than one‐third of theremaining forest cover is distributed in small forestfragments; however, the ranges of some species listedT
ABLE
III.
Pithec
iidSpec
iesWithGre
ates
tPer
centa
geof
Mod
ified
Lan
dCov
er
Spe
cies
a%
Mod
ified
%Forest
Con
servation
Status
Mea
nsize
forest
frag
men
ts(ha)
b
%of
forest
infrag
men
ts�1
0ha
%of
forest
infrag
men
ts�1
00ha
%of
forest
infrag
men
ts�1
000ha
%of
forest
infrag
men
ts�1
0,00
0ha
Callicebu
scoim
brai
76.9
8.3
End
ange
red
1212
.928
.145
.059
.4Callicebu
sca
queten
sis
75.1
24.8
Criticallyen
dang
ered
178.4
16.9
29.0
38.8
Callicebu
sba
rbar
abrownae
66.1
8.3
Criticallyen
dang
ered
188.4
18.7
31.7
46.6
Callicebu
snigrifron
s63
.821
.2Nea
rthreaten
ed20
8.7
23.2
39.2
50.8
Callicebu
spe
rson
atus
60.8
34.0
Vulne
rable
246.6
16.3
28.6
38.5
Callicebu
smelan
ochir
60.3
32.5
Vulne
rable
304.9
11.0
19.1
27.3
Callicebu
sornatus
30.9
63.1
Vulne
rable
463.2
7.5
13.0
20.7
Chirop
otes
satanas
26.6
63.8
Criticallyen
dang
ered
652.1
3.8
6.0
8.3
Callicebu
spa
llescens
19.8
68.7
Lea
stconc
ern
762.0
3.9
5.9
8.0
Chirop
otes
utahicka
e19
.376
.3End
ange
red
831.8
4.0
7.0
11.0
aSpe
cies
andthesp
ecies’ge
ograph
icrang
efollow
stheIU
CN
Red
List[IUCN,2
013].
bFragm
entsize
basedon
analysis
of30
‐mresolution
Lan
dsat
data
[Han
senet
al.,20
13].Minim
umfrag
men
tsize
used
inthean
alysis
was
0.5ha.
Am. J. Primatol.
8 / Boyle

as Least Concern have also undergone heavymodification since 2000. Therefore forest loss andfragmentation appear to have had recent impacts onthe geographic range ofmany pitheciid species, whichis worrisome when considering that the small frag-
mentsmay not be able to sustain these populations inthe long term [Gibbons andHarcourt, 2009; Harcourtand Doherty, 2005].
Although 23% of the pitheciid species have �50%of the remaining forest in protected areas, 40% of the
Fig. 3. The geographic range of the pitheciids experienced forest loss from 2000 to 2012, primarily in southern Colombia (1: 45 kmsoutheast of San Vicente del Caguán, Caquetá, Colombia), southern and eastern Brazilian Amazon (2: 135km southwest of Tucurí, Pará,Brazil), and the Paraguyan Chaco (3: north of Filadelfia, Boquerón, Paraguay). Data for forest extent and loss derive from Hansen et al.[2013].
Am. J. Primatol.
Pitheciids in Fragments / 9

pitheciid species have �25% of the remaining forestprotected. In general, pitheciid species that hadsmallerpercentagesof their remaining forest protectedhad greater forest loss from 2000 to 2012, suggestingthat unprotected forests are under heavy deforestationpressure. However, protected areas alone cannotconserve primates in the long‐term [Estrada, 2013].In addition to direct human pressures (e.g. hunting,deforestation), changes in forest structure, climate,disease, and predators can also impact primatepopulations [Chapman et al., 2005]. In the pitheciids,populations in someprotectedareasappear to beunderhuman pressures (e.g. Brownsberg Nature Park,Suriname [Gregory, 2011]; Saracá‐Taquera NationalForest [Melo et al., 2013b]; Yasuní [Kolowski andAlonso, 2012]; Fig. 4). Given the fragmented nature ofmuch of the landscape in parts of the world, it maybe beneficial to protect clusters of fragments [Rybickiand Hanski, 2013], which could improve connectivitybetween habitat fragments and can allow for gene flow[Bergl and Vigilant, 2007].
Forest fragmentation can decrease gene flowbetween forest fragments, decrease genetic diversity,and increase inbreeding [Keller and Waller, 2002;Knapp, 2013], all of which may negatively impactreproductive success [Amos et al., 2001] and conser-vation [Allendorf et al., 2013]. In primates, popula-tions living in small forest fragments can have lowergenetic diversity than populations living in more‐intact forest [Chaves et al., 2011; Craul et al., 2009],but to date there have been minimal studies ofgenetic diversity of pitheciids living in forest frag-ments [Ferrari et al., 2013b].
Most pitheciid studies addressing habitat lossand fragmentation have concentrated on populationestimates, with limited data on behavioral andecological changes (Table I). Therefore, although itis known that some species are capable of residing inhabitat fragments for the short‐term, there is stilllittle known about the geographic distribution,ecology, and behavior of many pitheciid species[Martinez andWallace, 2007; Pinto et al., 2013; SilvaJúnior et al., 2013]. Callicebus pallescens occurs in
the Paraguayan Chaco, but to date there are nobehavioral studies of these primates in Paraguay[Porter et al., 2013]. Furthermore, some areas areprimarily represented by checklists of mammals[Bruna et al., 2010] instead of behavioral andecological studies. The outlook does not look toopromising for many species. For example, Callicebuspallescens was listed as in 2008 with 19.8% of itsrange consisting of modified land cover, but there hasbeen a huge increase in deforestation of this region,particularly in Paraguay where 294,566ha were lostin 2012 alone [Palacios et al., 2013].
Callicebus, Chiropotes, and Pithecia can reside inforest fragments that are fractions of the size of thespecies’ typical home range in continuous forest[Boyle et al., 2009, 2012; Boyle and Smith, 2010a;DeLuycker, 2006, 2012, 2014; Kinzey andBecker, 1983; Setz, 1994; Setz and Gaspar, 1997;Setz et al., 1999; Silva and Ferrari, 2009; Veiga, 2006;Wagner et al., 2009]. Ferrari et al. [1999] provided arelatively positive assessment of the ability ofChiropotes to live in forest fragments; however, themonkeys were locally extinct from a 200‐ha fragment,as well as the surrounding area (given the small sizeof the other fragments), and the “successful” frag-ments were relatively large (�7500ha). AlthoughCallicebus, Chiropotes, and Pithecia have beendocumented in small (�20ha) habitat fragments,and some species have been noted to be tolerant ofhabitat fragmentation (e.g.Callicebus coimbrai [Cha-gas and Ferrari, 2010; Jerusalinsky et al., 2006];Chiropotes utahicki [Bobadilla and Ferrari, 2000]),the long‐term implications of such habitat fragmen-tation is not currently known. It may be thatindividuals in small, isolated fragments experiencelower reproductive success [Boyle and Smith, 2010a],or that the ability to use disturbed habitat is criticalfor the survival of the population [Heiduck, 2002]. Inmultiple studies the habitat fragments have beenvery isolated from other habitat areas by largedistances [Ferrari et al., 2007; García et al., 2010;Jerusalinsky et al., 2006] or by physical barriers suchas barbedwire [Defler et al., 2010]. As there appear to
TABLE IV. Pitheciid Species With Greatest Percentage of Forest Loss From 2000 to 2012
Speciesa % Forest loss Conservation status Population trend % of forest in protected areas
Chiropotes utahickae 21.3 Endangered Decreasing 40.9Callicebus coimbrai 17.7 Endangered Decreasing 10.8Callicebus oenanthe 14.5 Critically Endangered Decreasing 0.06Callicebus caquetensis 13.8 Critically Endangered Decreasing 0.0Chiropotes satanas 13.5 Critically Endangered Decreasing 25.0Callicebus moloch 11.6 Least Concern Unknown 55.2Callicebus brunneus 10.7 Least Concern Unknown 39.2Callicebus pallescens 9.8 Least Concern Stable 25.3Callicebus melanochir 9.7 Vulnerable Decreasing 16.6
aSpecies and the species’ geographic range follows the IUCN Red List [IUCN, 2013].
Am. J. Primatol.
10 / Boyle

Fig. 4. Recent land cover change (2000–2013) is visible in three areas that had been relatively intact in the year 2000: BrownsbergNaturePark, Suriname (A: 2000; B: 2013), Saracá‐Taquera National Forest, Brazil (C: 2000; D: 2013), and outside of Defensores del ChacoNational Park, Paraguay (E: 2000; F: 2013; black line indicates the park’s boundaries). Year 2000 imagery is from Landsat 5; year 2013imagery is from Landsat 8.
Am. J. Primatol.
Pitheciids in Fragments / 11

be differences in the frequency that pitheciid speciesengage in terrestrial movement [Barnett et al., 2012],a landscape of small, isolated forest fragments mayhave serious negative implications for species’ move-ment between habitat fragments.
Although there have not been any publishedstudies of Cacajao spp. in human‐modified habitatfragments, some species have likely experienced localextinction or severe population declines as a resultof hunting and logging [Barnett, 2005; Bowleret al., 2009], but the extent to which hunting hasimpacted populations is not fully known [Barnett,2005; Barnett et al., 2013]. Furthermore, there is nota full understanding of how human‐modified land-scapes have impacted Cacajao, as Peres [2001]reported that Cacajao went locally extinct in theextension of the highway BR‐364 in Acre, Brazil.Ferrari et al. [2013b] suggested that Cacajao may becapable of adjusting to fragmented habitat, given thesimilarities between its behavior and ecology withthose of Chiropotes; however, additional field work isnecessary in order to test this hypothesis, assimilarities to Chiropotes in home range and dietecology do not necessarily indicate that Cacajao farewell in fragmented habitat. Overall the extent ofhabitat fragmentation in the range of Cacajao hasbeenminimal comparedwith other pitheciids (Fig. 1).Unfortunately, current patterns of deforestationmayresult in additional opportunities to study Cacajao inhabitat fragments in the future. As hunting ofCacajao occurs [Bowler et al., 2009; Peres, 2001], itmay be that multiple factors (e.g. habitat loss,hunting, restricted ranges) drive increased pressureon Cacajao in the future [Ferrari et al., 2013c,d].
Increased data on the responses of pitheciidspecies to habitat loss and fragmentation are neces-sary in order to address pitheciid conservation in thefuture, especially in areas where there has beensevere habitat loss. Future studies should examineland cover change in the specific areas wherepitheciids have been found, as range maps canoverestimate distribution and omit areas that arepart of the actual distribution [Palminteri et al.,2009], and some species are patchily distributed andtherefore not found throughout their entire geo-graphic range [Boubli and de Lima, 2009; Ferrariet al., 2007]. Through this current study, two sites(Fig. 1, sites 2 and 27) where pitheciids have beennoted in habitat fragments (i.e., Callicebus ornatus[Wagner et al., 2009]; Callicebus nigrifrons [Santanaet al., 2008]) were not included within the IUCNgeographic ranges. Geographic range extensionshave also been reported for Cacajao in continuousforest [Bowler et al., 2009; Cardoso et al., 2014;Vermeer et al., 2013]. As additional studies areconducted, the geographic ranges of each species willneed to be verified.
Predicting a species’ threat to local extinction isnot straightforward, as it is important to take into
consideration of the multiple variables associatedwith the type of extinction threat and the biology ofthe species in question [Isaac and Cowlishaw, 2004].However, when one examines the extent of habitatremaining for the pitheciid species, several species(i.e. Callicebus combrai, Callicebus caquetensis;Table III) have very little native habitat remaining,and what is left is distributed among many smallfragments. Therefore it is critical that efforts aremade to reduce the rate of habitat loss andfragmentation, and provide possible solutions toimprove the conservation status of these species inthe near future. It may be beneficial, particularly inareas that are already heavily modified, to examinethe landscapes from multiple perspectives [Arroyo‐Rodríguez and Fahrig, 2014; Melo et al., 2013a],given that such a great extent of some pitheciidspecies’ ranges have undergone intense change.
ACKNOWLEDGEMENTS
I thank Paul Garber for his support of this specialissue, Adrian Barnett and Cynthia Thompson forcoediting this special issue, and Marilyn Norconk fororganizing the symposium where this paper was firstpresented. I also thank two anonymous reviewers forfeedback that helped improve the paper. Theresearch presented complieswith all legal and ethicalrequirements. I dedicate this paper to the memory ofLiza Veiga.
REFERENCES
Allendorf FW, Luikart G, Aitken SN. 2013. Conservation andthe genetics of populations. Second edition. New York: JohnWiley & Sons. 624 p.
Amos W, Wilmer JW, Fullard K., et al. 2001. The influence ofparental relatedness on reproductive success. Proceedings ofthe Royal Society of London B 268:2021–2027.
Anzures‐Dadda A, Manson RH. 2007. Patch‐ and landscape‐scale effects on howler monkey distribution and abundancein rainforest fragments. Animal Conservation 10:69–76.
Arroyo‐Rodríguez V, Fahrig L. 2014. Why is a landscapeperspective important in studies of primates? AmericanJournal of Primatology doi: 10.1002/ajp.22282.
Auricchio P. 2010. A morphological analysis of some species ofCallicebus, Thomas, 1903 (Pitheciidae – Callicebinae).Neotropical Primates 17:47–58.
Ayres JM. 1981. Observações sobre a ecologia e o comporta-mento dos cuxiús (Chiropotes albinasus e Chiropotessatanas, Cebidae: Primates) [thesis]. Manaus (Amazonas):Instituto Nacional de Pesquisas da Amazônia and FundaçãoUniversidade do Amazonas, Manaus. 142 p.
Barnett AA. 2005. Cacajao melanocephalus. MammalianSpecies 776:1–6.
Barnett AA, Bowler M, Bezerra BM, Defler TR. 2013. Ecologyand behavior of uacaris (genus Cacajao). In: Veiga LM,Barnett AA, Ferrari SF, Norconk MA, editors. Evolutionarybiology and conservation of titis, sakis and uacaris. Cam-bridge: Cambridge University Press. p 151–172.
Barnett AA, Boyle SA, Norconk MA., et al. 2012. Terrestrialactivity in pitheciins (Cacajao, Chiropotes, and Pithecia).American Journal of Primatology 74:1106–1127.
Am. J. Primatol.
12 / Boyle

Beltrão‐Mendes R, Cunha AA, Ferrari SF. 2011. New localitiesand perspectives on the sympatry between two endangeredprimates (Callicebus coimbrai and Cebus xanthosternos) innortheastern Brazil. Mammalia 75:103–105.
Benchimol M, Peres CA. 2014. Predicting primate localextinctions within “real‐world” forest fragments: a Pan‐Neotropical analysis. American Journal of Primatology76:289–302.
Bergl RA, Vigilant L. 2007. Genetic analysis reveals populationstructure and recentmigrationwithin the highly fragmentedrange of the Cross River gorilla (Gorilla gorilla diehli).Molecular Ecology 16:501–516.
Bicca‐Marques JC, Heymann EW. 2013. Ecology and behaviorof titimonkeys (genusCallicebus). In: VeigaLM,Barnett AA,Ferrari SF, Norconk MA, editors. Evolutionary biologyand conservation of titis, sakis and uacaris. Cambridge:Cambridge University Press. p 196–207.
Bobadilla UL, Ferrari SF. 1998. First detailed field data onChiropotes satanas utahicki Hershkovitz, 1985. NeotropicalPrimates 6:17–18.
Bobadilla U, Ferrari SF. 2000. Habitat use by Chiropotessatanas utahicki and syntopic platyrrhines in easternAmazonia. American Journal of Primatology 50:215–224.
Bontemps S, Defourny P, Van Bogaert E., et al. 2011.GlobCover 2009: Product description and validation report.53. pp. http://due.esrin.esa.int/globcover/LandCover2009/GLOBCOVER2009_Validation_Report_2.2.pdf.
Bonvicino CR, Boubli JP, Otazú IB., et al. 2003. Morphologic,karyotypic, and molecular evidence of a new form ofChiropotes (Primates, Pitheciinae). American Journal ofPrimatology 61:123–133.
Boubli JP, de Lima MG. 2009. Modeling the geographicaldistribution and fundamental niches of Cacajao spp. andChiropotes israelita in northwestern Amazonia via a maxi-mum entropy algorithm. International Journal of Primatol-ogy 30:217–228.
Boubli JP, Silva MNF, Amado MV., et al. 2008. A taxonomicreassessment of Cacajao melanocephalus Humboldt (1811)with the description of two new species. InternationalJournal of Primatology 29:723–741.
Bóveda‐Penalba AJ, Vermeer J, Rodrigo F, Guerra‐Vásquez F.2009. Preliminary report on the distribution of Callicebusoenanthe on the eastern feet of the Andes. InternationalJournal of Primatology 30:467–480.
Bowler M, Murrieta JN, Recharte M, Puertas P, Bodmer R.2009. Peruvian red uakari monkeys (Cacajao calvusucayalii) in the Pacaya‐Samiria National Reserve—A rangeextension across a major river barrier. Neotropical Primates16:34–37.
Boyle SA, Lourenço WC, da Silva LR, Smith AT. 2009. Traveland spatial patterns change when northern bearded sakimonkeys (Chiropotes satanas chiropotes) live in forestfragments. International Journal of Primatology 30:515–531.
Boyle SA, Smith AT. 2010a. Behavioral modifications innorthern bearded saki monkeys (Chiropotes satanas chiro-potes) in forest fragments of central Amazonia. Primates51:43–51.
Boyle SA, Smith AT. 2010b. Can landscape and speciescharacteristics predict primate presence in forest fragmentsin the BrazilianAmazon? Biological Conservation 143:1134–1143.
Boyle SA, Zartman CE, Spironello WR, Smith AT. 2012.Implications of habitat fragmentation on the diet of beardedsaki monkeys in central Amazonian forest. Journal ofMammalogy 93:957–976.
Boyle SA, Lenz BB, Gilbert KA, et al. 2013. Primates of theBiological Dynamics of Forest Fragments Project: A history.In: Marsh LK, Chapman C, editors. Primates in fragments:Ecology and conservation. New York: Springer. p 57–74.
Boyle SA, Kennedy CM, Torres J., et al. 2014. High‐resolutionsatellite imagery is an important yet underutilized resourcein conservation biology. PLoS ONE 9:e86908.
Bruna EM, Guimarães FJ, Lopes CT., et al. 2010. Mammalia,Estação Ecológica do Panga, a cerrado protected area inMinais Gerais state, Brazil. Check List 6:668–675.
Cardoso NA, Valsecchi J, Vieira T, Queiroz HL. 2014. Newrecords and range expansion of white bald uakari (Cacajaocalvus calvus, I. Geoffroy, 1847) in Central BrazilianAmazonia. Primates 55:199–206.
Cäsar C, Young RJ. 2008. A case of adoption in a wild group ofblack‐fronted titi monkeys (Callicebus nigrifrons). Primates49:146–148.
Cäsar C, Franco ES, Soares GCN, Young RJ. 2008. Observedcase of maternal infanticide in a wild group of black‐frontedtiti monkeys (Callicebus nigrifrons). Primates 49:143–145.
Chagas RRD, Ferrari SF. 2010. Habitat use by Callicebuscoimbrai (Primates: Pitheciidae) and sympatric species inthe fragmented landscape of the Atlantic Forest of southernSergipe, Brazil. Zoologia 27:853–860.
Chagas RRD, Ferrari SF. 2011. Population parameters of theendangered titi monkey, Callicebus coimbraiKobayashi andLangguth, 1999, in the fragmented landscape of southernSergipe, Brazil. Brazilian Journal of Biology 71:569–575.
Chapman CA, Struhsaker TT, Lambert JE. 2005. Thirty yearsof research in Kibale National Park, Uganda, reveals acomplex picture for conservation. International Journal ofPrimatology 26:539–555.
Chaves PB, Alvarenga CS, Possamai CB., et al. 2011. Geneticdiversity and population history of a critically endangeredprimate, the northern muriqui (Brachyteles hypoxanthus).PLoS ONE 6:e20722.
Chiarello AG,Melo FR. 2001. Primate population densities andsizes in Atlantic Forest remnants of Northern EspíritoSanto, Brazil. International Journal of Primatology 22:379–396.
Costa MD, Fernandes FAB, Hilário RR, Gonçalves AV, deSouza JM. 2012. Densidade, tamanho populacional econservação de primatas em fragment de Mata Atlânticano sul do Estado de Minas Gerais, Brasil. Iheringia, SérieZoologia 102:5–10.
Craul M, Chikhi L, Sousa V., et al. 2009. Influence of forestfragmentation on an endangered large‐bodied lemur innorthwestern Madagascar. Biological Conservation 142:2862–2871.
Cunningham EP, Janson CH. 2006. Pithecia pithecia’s behav-ioral response to decreasing fruit abundance. AmericanJournal of Primatology 68:491–497.
Cunningham EP, Harrison‐Levine AL, Norman RG. 2013.Finding the balance: optimizing predator avoidance and foodencounters through individual positioning in Pitheciapithecia during travel. In: Veiga LM, Barnett AA, FerrariSF, Norconk MA, editors. Evolutionary biology and conser-vation of titis, sakis and uacaris. Cambridge: CambridgeUniversity Press. p 272–276.
Defler TR, Bueno ML, García J. 2010. Callicebus caquetensis: anew and critically endangered titi monkey from southernCaquetá, Colombia. Primate Conservation 25:1–9.
DeLuycker AM. 2006. Preliminary report and conservationstatus of the Río Mayo titi monkey, Callicebus oenantheThomas, 1924, in the Alto Mayo Valley, Northeastern Peru.Primate Conservation 21:33–39.
DeLuycker AM. 2012. Insect prey foraging strategies inCallicebus oenanthe in Northern Peru. American Journalof Primatology 74:450–461.
DeLuycker AM. 2014. Observations of a daytime birthing eventin wild titi monkeys (Callicebus oenanthe): implications ofthe male parental role. Primates 55:59–67.
Estrada A. 2013. Socioeconomic contexts of primate conserva-tion: population, poverty, global economic demands, and
Am. J. Primatol.
Pitheciids in Fragments / 13

sustainable land use. American Journal of Primatology75:30–45.
Ferrari SF, Emidio‐Silva C, Lopes MA, Bobadilla UL. 1999.Bearded sakis in south‐eastern Amazonia—back from thebrink? Oryx 33:346–351.
Ferrari SF, Iwanaga S, Ravetta AL., et al. 2003. Dynamics ofprimate communities along the Santarém‐Cuiabá Highwayin south‐central Brazilian Amazonia. In: Marsh LM, editor.Primates in fragments: Ecology and conservation. NewYork:Kluwer Academic/Plenum Publishers. p 123–144.
Ferrari SF, Pereira WLA, Santos RR, Veiga LM. 2004. Fatalattack of a Boa constrictor on a bearded saki (Chiropotessatanas utahicki). Folia Primatologica 803:1–3.
Ferrari SF, Bobadilla UL, Emidio‐Silva C. 2007. Where haveall the titis gone? The heterogeneous distribution ofCallicebus moloch in eastern Amazonia, and its implicationsfor the conservation of Amazonian primates. PrimateConservation 22:49–54.
Ferrari SF, Boyle SA, Marsh LK, et al. 2013a. The challenge ofliving in fragments. In: Veiga LM, Barnett AA, Ferrari SF,NorconkMA, editors. Evolutionary biology and conservationof titis, sakis and uacaris. Cambridge: CambridgeUniversityPress. p 350–358.
Ferrari SF, Santos Junior EM, Freitas EB., et al. 2013b. Livingon the edge: habitat fragmentation at the interface of thesemiarid zone in the Brazilian northeast. In: Marsh LK,Chapman CA, editors. Primates in fragments: complexityand resilience. New York: Springer. p 121–135.
Ferrari SF, Silva Júnior JS, de Souza MA. 2013c. Brazil. In:Veiga LM, Barnett AA, Ferrari SF, Norconk MA, editors.Evolutionary biology and conservation of titis, sakis anduacaris. Cambridge: Cambridge University Press. p 334–343.
Ferrari SF, Silva Júnior JS, de Souza MA, Albernaz ALK.2013d. Conservation fact sheet: Brazil. In: Veiga LM,Barnett AA, Ferrari SF, Norconk MA, editors. Evolutionarybiology and conservation of titis, sakis and uacaris. Cam-bridge: Cambridge University Press. p 373–377.
Freitas EB, de Carvalho CB, Ferrari SF. 2011. Abundance ofCallicebus barbarabrownae (Hershkovitz 1990), (Primates:Pitheciidae) and other nonvolant mammals in a fragmentof arboreal Caatinga in northeastern Brazil. Mammalia75:339–343.
García J, Defler TR, Bueno ML. 2010. The conservation statusof Callicebus caquetensis (Pitheciidae): a new speciesin southern Caquetá Department, Colombia. NeotropicalPrimates 17:37–46.
Gibbons MA, Harcourt AH. 2009. Biological correlates ofextinction and persistence of primates in small forestfragments: a global analysis. Tropical Conservation Science2:388–403.
Gilbert KA. 2000. Attempted predation on a white‐facedsaki in the central Amazon. Neotropical Primates 8:103–104.
Gillespie TR, Chapman CA. 2006. Prediction of parasiteinfection dynamics in primate metapopulations based onattributes of forest fragmentation. Conservation Biology20:441–448.
Gleason TM, Norconk MA. 2002. Predation risk and antipred-ator adaptations in white‐faced sakis, Pithecia pithecia. In:Miller LE, editor. Eat or be eaten: Predation sensitiveforaging among primates. New York: Cambridge UniversityPress. p 169–184.
Gonçalves EC, Ferrari SF, Silva A., et al. 2003. Effects ofhabitat fragmentation on the genetic variability of silverymarmosets,Mico argentatus. In:MarshLM, editor. Primatesin fragments: Ecology and conservation. New York: KluwerAcademic/Plenum Publishers. p 17–28.
Gordo M, Rodrigues LF, Vidal MD, Spironello WR. 2008.Primatas. In: Oliveira ML, Baccaro FB, Braga‐Neto R,Magnusson WE, editors. Reserva Ducke: A biodiverisdade
Amazônica através de uma grade. Manaus: Áttema DesignEditorial Ltda. p 39–49.
Gregory TL. 2011. Socioecology of the Guianan bearded saki,Chiropotes sagulatus [dissertation]. Kent (Ohio): Kent StateUniversity. 214 p.
HansenMC, Potapov PV,Moore R., et al. 2013. High‐resolutionglobal maps of 21st‐century forest cover change. Science342:850–853. Data available on‐line from: http://earthengi-nepartners.appspot.com/science‐2013‐global‐forest.
Harcourt AH, Doherty DA. 2005. Species‐area relationships ofprimates in tropical forest fragments: a global analysis.Journal of Applied Ecology 42:630–637.
Heiduck S. 1997. Food choice in masked titi monkeys(Callicebus personatus melanochir): selectivity or opportun-ism? International Journal of Primatology 18:487–503.
Heiduck S. 2002. The use of disturbed and undisturbed forestby masked titi monkeys Callicebus personatus melanochir isproportional to food availability. Oryx 36:133–139.
Heiduck S. 2013. Costs of foraging in the southern Bahianmasked titi monkey (Callicebus melanochir). In: Veiga LM,Barnett AA, Ferrari SF, Norconk MA, editors. Evolutionarybiology and conservation of titis, sakis and uacaris. Cam-bridge: Cambridge University Press. p 208–214.
Hickey JR, Nackoney J, Nibbelink NP., et al. 2013. Humanproximity and habitat fragmentation are key drivers of therangewide bonobo distribution. Biodiversity and Conserva-tion 22:3085–3104.
Isaac NJB, Cowlishaw G. 2004. How species respond tomultiple extinction threats. Proceedings of the Royal SocietyB 271:1135–1141.
IUCN. 2013. IUCN Red List of Threatened Species. Version2013.1 www.iucnredlist.org.
IUCN. UNEP. 2014. The World Database on Protected Areas(WDPA). Cambridge, UK: UNEP‐WCMC. www.protected-planet.net.
Jerusalinsky L, Oliveira MM, Pereira RF., et al. 2006.Preliminary evaluation of the conservation status ofCallicebus coimbrai Kobayashi & Langguth, 1999 in theBrazilian state of Sergipe. Primate Conservation 21:25–32.
Johns AD, Skorupa JP. 1987. Responses of rain‐forest primatesto habitat disturbance – a review. International Journal ofPrimatology 8:157–191.
Keller LF, Waller DM. 2002. Inbreeding effects in wildpopulations. Trends in Ecology and Evolution 17:230–241.
KinzeyWG, BeckerM. 1983. Activity pattern of themasked titimonkey, Callicebus personatus. Primates 24:337–343.
Kinzey WG, Norconk MA. 1993. Physical and chemicalproperties of fruit and seeds eaten by Pithecia andChiropotes in Surinam and Venezuela. International Jour-nal of Primatology 14:207–227.
Knapp LA. 2013. Molecular genetic tools for evaluating theconsequences of habitat fragmentation. In: Marsh LK,Chapman CA, editors. Primates in fragments: complexityand resilience. New York: Springer. p 389–398.
Kolowski JM, Alonso A. 2012. Primate abundance in anunhunted region of the northern Peruvian Amazon and theinfluence of seismic oil exploration. International Journal ofPrimatology 33:958–971.
Lehman SM, Sussman RW, Phillips‐Conroy J, Prince W. 2006.Ecological biogeography of primates in Guyana. In: LehmanSM, Fleagle JG, editors. Primate biogeography. New York:Springer. p. 105–130.
LehmanSM,Vié J‐C,NorconkMA,Portillo‐QuinteroC,UrbaniB. 2013. TheGuyana Shield: Venezuela and theGuyanas. In:Veiga LM, Barnett AA, Ferrari SF, Norconk MA, editors.Evolutionary biology and conservation of titis, sakis anduacaris. Cambridge: Cambridge University Press. p 311–319.
Lopes MA, Ferrari SF. 2000. Effects of human colonization onthe abundance and diversity of mammals in eastern Brazil-ian Amazonia. Conservation Biology 14:1658–1665.
Am. J. Primatol.
14 / Boyle

Mandujano S, Escobedo‐Morales LA, Palacios‐Silva R, Arroyo‐Rodríguez V, Rodríguez‐Toledo EM. 2005. Ametapopulationapproach to conserving the howler monkey in a highlyfragmented landscape in Los Tuxtlas, Mexico. In: Estrada A,Garber PA, Pavelka MSM, Luecke L, editors. New perspec-tives in the study of Mesoamerican primates: Distribution,ecology, behavior, and conservation. New York: Springer. p513–538.
Mark MM. 2003. Some observations on Callicebus oenanthe inthe upper Río Mayo Valley, Peru. Neotropical Primates11:183–187.
Marques EN, Beltrão‐Mendes R, Ferrari SF. 2013a. Primates,Pitheciidae, Callicebus barbarabrownae Hershkovitz, 1990:new localities for the critically endangered titi monkey in theSão Francisco basin, state of Sergipe, Brazil. Check List9:113–115.
Marques EN, Jerusalinsky L, Rocha JCAG., et al. 2013b.Primates, Pitheciidae, Callicebus coimbrai Kobayashi andLangguth, 1999: new localities for an endangered titimonkey in eastern Sergipe, Brazil. Check List 9:696–699.
Martinez J, Wallace RB. 2007. Further notes on the distribu-tion of endemic Bolivian titi monkeys, Callicebus modestusand Callicebus olallae. Neotropical Primates 14:47–54.
MasonWA. 1968.Use of space byCallicebus groups. In: Jay PC,editor. Primates studies in adaption and variability. NewYork: Holt, Rinehart & Winston. p 200–216.
Melo FPL, Arroyo‐Rodríguez V, Fahrig L, Martínez‐Ramos M,Tabarelli M. 2013a. On the hope for biodiversity‐friendlytropical landscapes. Trends in Ecology and Evolution28:462–468.
Melo FR, Moreira LS, Moura VS., et al. 2013b. Ecology andbehaviour of two groups of Chiropotes sagulatus inhabitingareas under different pressures of mining in the Saraca‐Taquera National Forest, Pará, Brazil. American Journal ofPrimatology 75:94.
Michalski F, Peres CA. 2005. Anthropogenic determinants ofprimate and carnivore local extinctions in a fragmentedforest landscape of southern Amazonia. Biological Conser-vation 124:383–396.
Müller K‐H. 1995. Ranging in masked titi monkeys (Callicebuspersonatus) in Brazil. Folia Primatologica 65:224–228.
Müller K‐H. 1996. Diet and feeding ecology of masked titis(Callicebus personatus). In: Norconk MA, Rosenberger AL,Garber PA, editors. Adaptive radiations of Neotropicalprimates. New York: Plenum Press. p 383–401.
Norconk MA. 1996. Seasonal variation in the diets of white‐faced and bearded sakis (Pithecia pithecia and Chiropotessatanas) in Guri Lake, Venezuela. In: Norconk MA, Rose-nberger AL, Garber PA, editors. Adaptive radiations ofNeotropical primates. New York: Plenum Press. p 403–423.
Norconk MA. 2006. Long‐term study of group dynamics andfemale reproduction in Venezuela Pithecia pithecia. Inter-national Journal of Primatology 27:653–674.
Norconk MA, Conklin‐Brittain NL. 2004. Variation in frugi-vory: the diet of Venezuelan white‐faced sakis. InternationalJournal of Primatology 25:1–26.
NorconkMA, Grafton BW. 2003. Changes in forest compositionand potential feeding tree availability on a small land‐bridgeisland in Lago Guri, Venezuela. In: Marsh LK, editor.Primates in fragments. New York: Kluwer Academic. p 211–227.
Norconk MA, Sez EZ. 2013. Ecology and behavior of sakimonkeys (genusPithecia). In: Veiga LM,Barnett AA, FerrariSF, Norconk MA, editors. Evolutionary biology and conser-vation of titis, sakis and uacaris. Cambridge: CambridgeUniversity Press. p 262–271.
Oates J. 2013. Primate conservation: unmet challenges and therole of the International Primatological Society. Interna-tional Journal of Primatology 34:235–245.
Oliveira RCR, Coelho AS, de Melo FR. 2003. Estimativa dedensidade e tamanho populacional de sauá (Callicebus
nigrifrons) em um fragment de mata em regeneração,Viçosa, Minas Gerais, Brasil. Neotropical Primates 11:91–94.
Palacios F, Cardozo R, Caballero J, Paradeda C, Yanosky A.2013. Monitoreo de los cambios de uso de la tierra, incendiose inundaciones del Gran Chaco: Informe final – ãno 2012.Guyra Paraguay, Asunción, Paraguay, 22 p.
Palminteri S, Powell G, Endo W., et al. 2009. Usefulness ofspecies range polygons for predicting local primate occur-rences in southeastern Peru. American Journal of Primatol-ogy 71:1–9.
Palminteri S, Peres CA. 2012. Habitat selection and use ofspace by bald‐faced sakis (Pithecia irrorata) in southwesternAmazonia: lessons from a multiyear, multigroup study.International Journal of Primatology 33:401–417.
Peetz A. 2001. Ecology and social organization of the beardedsakiChiropotes satanas chiropotes (Primates: Pitheciinae) inVenezuela. Ecotropical Monographs 1:1–170.
Peres CA. 1993. Notes on the ecology of buffy saki monkeys(Pithecia albicans, Gray 1860): a canopy seed‐predator.American Journal of Primatology 31:129–140.
Peres CA. 2001. Synergistic effects of subsistence hunting andhabitat fragmentation on Amazonian forest vertebrates.Conservation Biology 15:1490–1504.
Pimm SL, Brooks T. 2013. Conservation: forest fragments,facts, and fallacies. Current Biology 23:R1098–1101.
Pinto LP, Barnett AA, Bezerra BM., et al. 2013. Why we knowso little: the challenges of fieldwork on the pitheciids. In:Veiga LM, Barnett AA, Ferrari SF, Norconk MA, editors.Evolutionary biology and conservation of titis, sakis anduacaris. Cambridge: Cambridge University Press. p 145–150.
Port‐Carvalho M, Ferrari SF. 2004. Occurrence and diet of theblack bearded saki (Chiropotes satanas satanas) in thefragmented landscape of western Maranhão, Brazil. Neo-tropical Primates 12:17–21.
Porter L, Chism J, Defler TR., et al. 2013. Pitheciid conserva-tion in Ecuador, Colombia, Peru, Bolivia and Paraguay. In:Veiga LM, Barnett AA, Ferrari SF, Norconk MA, editors.Evolutionary biology and conservation of titis, sakis anduacaris. Cambridge: Cambridge University Press. p 320–333.
Pyritz LW, Büntge ABS, Herzog SK, Kessler M. 2010. Effectsof habitat structure and fragmentation on diversityand abundance of primates in tropical deciduous forestsin Bolivia. International Journal of Primatology 31:796–812.
Rowe N, Martínez W. 2003. Callicebus sightings in Bolivia,Peru and Ecuador. Neotropical Primates 11:32–35.
Rybicki J, Hanski I. 2013. Species‐area relationships andextinctions caused by habitat loss and fragmentation.Ecology Letters 16:27–38.
Sampaio DT, Ferrari SF. 2005. Predation of an infant titimonkey (Callicebus moloch) by a tufted capuchin (Cebusapella). Folia Primatologica 76:113–115.
Santana BEMM, do Prado MR, Lessa G, Rocha EC, Melo FR.2008. Densidade, tamanho populacional e abundância dosprimatas em um fragmento de floresta Atlântica em MinasGerais, Brasil. Revista Árvore 32:1109–1117.
Santos GP, Galvão C, Young RJ. 2012. The diet of wild black‐fronted titi monkeys Callicebus nigrifrons during a bamboomasting year. Primates 53:265–272.
Santos RR, Vieira TM, Ferrari SF. 2013. Feeding ecology of UtaHick’s bearded saki (Chiropotes utahickae) on a man‐madeisland in southeastern Brazilian Amazonia: seasonal andlongitudinal variation. In: Veiga LM, Barnett AA, FerrariSF, Norconk MA, editors. Evolutionary biology and conser-vation of titis, sakis and uacaris. Cambridge: CambridgeUniversity Press. p 250–254.
São Bernardo CS, Galetti M. 2004. Densidade e tamanhopopulacional de primatas em um fragmento florestal no
Am. J. Primatol.
Pitheciids in Fragments / 15

sudeste do Brasil. Revista Brasileira de Zoologia 21:827–832.
Schwarzkopf L, Rylands AB. 1989. Primate species richness inrelation to habitat structure in Amazonian rainforest frag-ments. Biological Conservation 48:1–12.
Setz EFZ. 1994. Feeding ecology of golden‐faced sakis.Neotropical Primates 2:13–14.
Setz EFZ, Gaspar DA. 1997. Scent‐marking behavior in free‐ranging golden‐faced saki monkeys, Pithecia pithecia chrys-ocephala: sex differences and context. Journal of Zoology241:603–611.
Setz EFZ, Enzweiler J, Solferini VN, AmêndolaMP, Berton RS.1999. Geophagy in the golden‐faced saki monkey (Pitheciapithecia chrysocephala) in the Central Amazon. Journal ofZoology 247:91–103.
Shaffer CA. 2013. Ecological correlates of ranging behavior inbearded sakis (Chiropotes sagulatus) in a continuous forestin Guyana. International Journal of Primatology 34:515–532.
Sharma N, Madhusudan MD, Sinha A. 2014. Distribution andpersistence in the remnant lowland rainforests of the UpperBrahmaputra Valley, northeastern India. ConservationBiology 28:95–106.
Silva SSB, Ferrari SF. 2009. Behavior patterns of southernbearded sakis (Chiropotes satanas) in the fragmentedlandscape of eastern Brazilian Amazonia. American Journalof Primatology 70:1–7.
Silva Júnior JS, Figueiredo‐Ready WMB, Ferrari SF. 2013.Taxonomy and geographic distribution of the Pitheciidae. In:Veiga LM, Barnett AA, Ferrari SF, Norconk MA, editors.Evolutionary biology and conservation of titis, sakis anduacaris. Cambridge: Cambridge University Press. p 31–42.
SousaMC. 2003. Distribuição do guigó (Callicebus coimbrai) noestado de Sergipe. Neotropical Primates 11:89–91.
Souza‐Alves JP, Fontes IP, Chagas RRD, Ferrari SF. 2011.Seasonal versatility in the feeding ecology of a group of titis
(Callicebus coimbrai) in the northern Brazilian AtlanticForest. American Journal of Primatology 73:1199–1209.
Suárez E, Morales M, Cueva R., et al. 2009. Oil industry, wildmeat trade and roads: indirect effects of oil extractionactivities in a protected area in north‐eastern Ecuador.Animal Conservation 12:364–373.
Trevelin LC, Port‐Carvalho M, Silveira M, Morell E. 2007.Abundance, habitat use and diet of Callicebus nigrifronsSpix (Primates, Pitheciidae) in Cantareira State Park, SãoPaulo, Brazil. Revista Brasileira de Zoologia 24:1071–1077.
Urbani B. 2002. A field observation on color selection by NewWorld sympatric primates, Pithecia pithecia and Alouattaseniculus. Primates 43:95–101.
Urbani B. 2006. A survey of primate populations in northeast-ern Venezuelan Guayana. Primate Conservation 20:47–52.
Veiga LM. 2006. Ecologia e organização social de cuxiús,Chiropotes satanas satanas, emTucuruí, Pará [dissertation].Belém (Brazil): Universidade Federal do Pará, 207 p.
Veiga LM, Ferrari SF. 2006. Predation of arthropods bysouthern bearded sakis (Chiropotes satanas) in easternBrazilian Amazonian. American Journal of Primatology 68:209–215.
Veiga LM, Ferrari SF. 2013. Ecology and behavior of beardedsakis (genus Chiropotes). In: Veiga LM, Barnett AA, FerrariSF, Norconk MA, editors. Evolutionary biology and conser-vation of titis, sakis and uacaris. Cambridge: CambridgeUniversity Press. p 240–249.
Vermeer J, Tello‐Alvarado JC, Castillo JTV, Bóveda‐PenalbaAJ. 2013. A new population of red uakaris (Cacajao calvusssp.) in the mountains of north‐eastern Peru. NeotropicalPrimates 20:12–17.
Wagner M, Castro F, Stevenson PR. 2009. Habitat characteri-zation and population status of the dusky titi (Callicebusornatus) in fragmented forests, Meta, Colombia. NeotropicalPrimates 16:18–24.
Am. J. Primatol.
16 / Boyle




















