
Physiological and behavioural responses of Gammarus pulex (Crustacea: Amphipoda) exposed to cadmium
13
Aquatic Toxicology 86 (2008) 413–425 Physiological and behavioural responses of Gammarus pulex (Crustacea: Amphipoda) exposed to cadmium V. Felten a,∗ , G. Charmantier b , R. Mons a , A. Geffard c , P. Rousselle d , M. Coquery e , J. Garric a , O. Geffard a a Laboratoire d’Ecotoxicologie, CEMAGREF, 3 bis quai Chauveau, CP 220, 69336 Lyon Cedex 09, France b Equipe Adaptation Ecophysiologique et Ontogen` ese, UMR 5119 Ecolag, Universit´ e Montpellier II, Place E. Bataillon, 34095 Montpellier Cedex 05, France c Laboratoire d’Eco-toxicologie, Universit´ e de Reims Champagne Ardenne, Facult´ e des Sciences, Moulin de la Housse, BP 1039, 51687 Reims Cedex 2, France d Laboratoire Biodiversit´ e et Fonctionnement des Ecosyst` emes, Universit´ e de Metz, Campus Bridoux, Rue du G´ en´ eral Delestraint, 57 070 Metz, France e Laboratoire de Chimie Environnementale, CEMAGREF, 3 bis quai Chauveau, CP 220, 69336 Lyon Cedex 09, France Received 10 September 2007; received in revised form 8 December 2007; accepted 11 December 2007 Abstract The aim of this study was to investigate the effects of cadmium on physiological and behavioural responses in Gammarus pulex. In a first experiment, cadmium LC50s for different times were evaluated in 264h experiment under continuous mode of exposure (LC50 96h = 82.1 gL −1 , LC50 120h = 37.1 gL −1 , LC50 168h = 21.6 gL −1 , LC50 264h = 10.5 gL −1 ). In a second experiment, the physiological and behavioural responses of the amphipod exposed to cadmium (0, 7.5 and 15 gL −1 ) were investigated under laboratory conditions. The mortality and the whole body cadmium concentration of organisms exposed to cadmium were significantly higher than in controls. Con- cerning physiological responses, cadmium exposure exerted a significant decrease on osmolality and haemolymph Ca 2+ concentration, but not on haemolymph Na + and Cl − concentrations, whereas the Na + /K + -ATPase activity was significantly increased. Behavioural responses, such as feeding rate, locomotor and ventilatory activities, were significantly reduced in Cd exposed organisms. Mechanism of cadmium action and consequent energetic reallocation in favour of maintenance functions (i.e., osmoregulation) are discussed. The results of this study indicate that osmolality and locomotor activity in G. pulex could be effective ecophysiological/behavioural markers to monitor freshwater ecosystem and to assess the health of organisms. © 2007 Elsevier B.V. All rights reserved. Keywords: Cadmium; Gammarus pulex; Survival; Physiology; Behaviour 1. Introduction Cadmium is a nonessential heavy metal of high envi- ronmental concern (on some government priority-substance lists, e.g., Canadian Environmental Protection Act, 1994) due to its toxicity, general usage, industrial production and emissions from fossil fuel combustion. In natural fresh- waters, cadmium sometimes occurs at concentrations of less than 0.1 gL −1 (USEPA, 2001), but in environments impacted by man, concentrations may reach micrograms per liter or higher values (e.g., Bervoets et al., 2005: 67 gL −1 of dissolved cadmium in Flanders freshwa- ∗ Corresponding author at: Laboratoire d’Eco-toxicologie, Universit´ e de Reims Champagne Ardenne, Facult´ e des Sciences, Moulin de la Housse, BP 1039, 51687 Reims Cedex 2, France. Tel.: +33 3 26 91 33 43. E-mail address: [email protected] (V. Felten). ter; Canadian Environmental Protection Act, 1994: [Cd] ≥300 gL −1 ). Freshwater crustaceans are among the most Cd-sensitive macroinvertebrate species (Williams et al., 1985; Canadian Environmental Protection Act, 1994; and references therein). Although responses of crustacean to cadmium stress are rea- sonably well documented both in terms of (1) acute toxicity (McCahon and Pascoe, 1988a,b,c; Williams et al., 1985; Wright and Fain, 1981) and (2) sublethal effects (mainly reproduc- tion effect: Biesinger and Christensen, 1972), relatively little is known about its effect on the physiology of freshwater species, particularly in crustaceans. The freshwater gammarid Gammarus pulex was selected as the test organism in this study because this species is (1) widespread and common in West Paleartica (Barnard and Barnard, 1983), (2) known to be sensitive to a range of stresses, (3) often occurring at high density, (4) easy to identify down to 0166-445X/$ – see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.aquatox.2007.12.002
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Physiological and behavioural responses of Gammarus pulex (Crustacea: Amphipoda) exposed to cadmium

A
eLt
chr
Tm©
K
1
rldewlip6
R1
0d
Aquatic Toxicology 86 (2008) 413–425
Physiological and behavioural responses of Gammarus pulex(Crustacea: Amphipoda) exposed to cadmium
V. Felten a,∗, G. Charmantier b, R. Mons a, A. Geffard c, P. Rousselle d,M. Coquery e, J. Garric a, O. Geffard a
a Laboratoire d’Ecotoxicologie, CEMAGREF, 3 bis quai Chauveau, CP 220, 69336 Lyon Cedex 09, Franceb Equipe Adaptation Ecophysiologique et Ontogenese, UMR 5119 Ecolag, Universite Montpellier II, Place E. Bataillon, 34095 Montpellier Cedex 05, France
c Laboratoire d’Eco-toxicologie, Universite de Reims Champagne Ardenne, Faculte des Sciences, Moulin de la Housse, BP 1039, 51687 Reims Cedex 2, Franced Laboratoire Biodiversite et Fonctionnement des Ecosystemes, Universite de Metz, Campus Bridoux, Rue du General Delestraint, 57 070 Metz, France
e Laboratoire de Chimie Environnementale, CEMAGREF, 3 bis quai Chauveau, CP 220, 69336 Lyon Cedex 09, France
Received 10 September 2007; received in revised form 8 December 2007; accepted 11 December 2007
bstract
The aim of this study was to investigate the effects of cadmium on physiological and behavioural responses in Gammarus pulex. In a firstxperiment, cadmium LC50s for different times were evaluated in 264 h experiment under continuous mode of exposure (LC5096h = 82.1 �g L−1,C50120h = 37.1 �g L−1, LC50168h = 21.6 �g L−1, LC50264h = 10.5 �g L−1). In a second experiment, the physiological and behavioural responses of
he amphipod exposed to cadmium (0, 7.5 and 15 �g L−1) were investigated under laboratory conditions.The mortality and the whole body cadmium concentration of organisms exposed to cadmium were significantly higher than in controls. Con-
erning physiological responses, cadmium exposure exerted a significant decrease on osmolality and haemolymph Ca2+ concentration, but not onaemolymph Na+ and Cl− concentrations, whereas the Na+/K+-ATPase activity was significantly increased. Behavioural responses, such as feeding
ate, locomotor and ventilatory activities, were significantly reduced in Cd exposed organisms.Mechanism of cadmium action and consequent energetic reallocation in favour of maintenance functions (i.e., osmoregulation) are discussed.he results of this study indicate that osmolality and locomotor activity in G. pulex could be effective ecophysiological/behavioural markers toonitor freshwater ecosystem and to assess the health of organisms.2007 Elsevier B.V. All rights reserved.
t≥
mEAs(a
eywords: Cadmium; Gammarus pulex; Survival; Physiology; Behaviour
. Introduction
Cadmium is a nonessential heavy metal of high envi-onmental concern (on some government priority-substanceists, e.g., Canadian Environmental Protection Act, 1994)ue to its toxicity, general usage, industrial production andmissions from fossil fuel combustion. In natural fresh-aters, cadmium sometimes occurs at concentrations of
ess than 0.1 �g L−1 (USEPA, 2001), but in environments
mpacted by man, concentrations may reach microgramser liter or higher values (e.g., Bervoets et al., 2005:7 �g L−1 of dissolved cadmium in Flanders freshwa-∗ Corresponding author at: Laboratoire d’Eco-toxicologie, Universite deeims Champagne Ardenne, Faculte des Sciences, Moulin de la Housse, BP039, 51687 Reims Cedex 2, France. Tel.: +33 3 26 91 33 43.
E-mail address: [email protected] (V. Felten).
tkp
a(B(
166-445X/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.aquatox.2007.12.002
er; Canadian Environmental Protection Act, 1994: [Cd]300 �g L−1).Freshwater crustaceans are among the most Cd-sensitive
acroinvertebrate species (Williams et al., 1985; Canadiannvironmental Protection Act, 1994; and references therein).lthough responses of crustacean to cadmium stress are rea-
onably well documented both in terms of (1) acute toxicityMcCahon and Pascoe, 1988a,b,c; Williams et al., 1985; Wrightnd Fain, 1981) and (2) sublethal effects (mainly reproduc-ion effect: Biesinger and Christensen, 1972), relatively little isnown about its effect on the physiology of freshwater species,articularly in crustaceans.
The freshwater gammarid Gammarus pulex was selected
s the test organism in this study because this species is1) widespread and common in West Paleartica (Barnard andarnard, 1983), (2) known to be sensitive to a range of stresses,3) often occurring at high density, (4) easy to identify down to

4 oxico
tco(lw
xmaWbhewiibaemibMuf1u
bq2Pattba
TMw
CpTO
CNPSNKCM
tosu
pc(awmNba
2
2
uiwTw
Q
2
owcu
14 V. Felten et al. / Aquatic T
he species level and characterized by a sexual dimorphism, (5)urrently used in ecotoxicological tests, (6) an important reservef food for macroinvertebrates, fish, bird and amphibian speciesMacNeil et al., 2000), and (7) playing a major role in the leafitter breakdown process and consequently in the entire foodeb (Forrow and Maltby, 2000).Gills constitute a sensitive primary target for a variety of
enobiotics due to direct contact with the aquatic environ-ent. Gills of freshwater crustaceans are crucial for respiration,
cid–base balance, osmotic and ionic regulation (Henry andheatly, 1992). According to Soegianto et al. (1999), water-
orne cadmium accumulates primarily on crustacean’s gills andepatopancreas causing cellular damages (Xu, 1995; Soegiantot al., 1999; Silvestre et al., 2005). Crustaceans exposed toater-borne pollutants, environmental stressors and patholog-
cal agents usually exhibit disruption of iono/osmoregulation,ncluding alterations in the structure and ultrastructure of theranchial and excretory organs, and changes in Na+/K+-ATPasectivity, ion fluxes and surface permeability (review in Lignott al., 2000). As a result, Na+ and Cl− which are known toade up 90% or more of the osmotic pressure of haemolymph
n most crustaceans (Mantel and Farmer, 1983), can be impactedy stress (i.e., zinc: Johnson and Jones, 1990; copper: Brooks andills, 2003; acidification: Felten and Guerold, 2004). Osmoreg-
lation is an important and highly energy-consuming regulatoryunction in aquatic invertebrates. According to Sutcliffe (1984),1% of the total energy budget in G. pulex is devoted to osmoreg-lation.
In the past few years, a number of studies have usedehavioural responses as tools for ecotoxicity testing and wateruality monitoring (locomotor activity: Wallace and Estephan,004; Mills et al., 2006; ventilatory activity: Lawrence andoulter, 1998; Felten and Guerold, 2001; feeding rate: Maltby etl., 2002). The development of such tests is of interest for eco-oxicology because, in addition to being sensitive, fast, simple
o perform and cheap, they allow to link toxic effects obtained atiochemical/cellular levels to impacts observed on populationsnd communities (Wallace and Estephan, 2004). We hypothesizeable 1ean (±S.D.) values of various parameters and components of the dilutionater (well-water) used in the experiments (n = 4)
Mean ± S.D.
onductivity (�S cm−1) 589 ± 1H 7.19 ± 0.02emperature (◦C) 8.5 ± 2.1smolality (mOsm kg−1) 12.03 ± 0.07
mg L−1 �mol L−1
l− 66.7 ± 3.5 1881.4 ± 98.7O3
− 27 ± 1 435.5 ± 16.1O4
3− <0.03 <0.38O4
2− 68.3 ± 3.5 711.0 ± 36.4a+ 29.7 ± 1.5 1291.9 ± 65.2+ 2.43 ± 0.21 62.2 ± 5.4a2+ 99.3 ± 3.1 2477.5 ± 77.3g2+ 5.17 ± 0.29 212.7 ± 11.9
wwdto1t(“(112o1(aaeeai
logy 86 (2008) 413–425
hat adverse effects on osmoregulation should lead to alterationf behaviour (i.e., locomotor activity), since energetic allocationhould be in favour of maintenance functions (such as osmoreg-lation, detoxification).
The objectives of the present study were (1) to assess G.ulex’s LC50s (experiment 1), (2) to investigate the effects ofadmium on physiological and behavioural responses in G. pulexexperiment 2), and (3) to evaluate mechanisms of cadmiumction. To achieve these goals, different levels of cadmiumere used to assess its effect on survival, cadmium accu-ulation, physiological (haemolymph osmolality, haemolympha+, Cl− and Ca2+concentrations, Na+/K+-ATPase activity) andehavioural responses (feeding rate, ventilation and locomotorctivity) of G. pulex’s.
. Materials and methods
.1. Collection and maintenance of animals
Specimens of G. pulex were collected using a hand net fromnpolluted stream, La Bourbre (near Lyon, France). The organ-sms were quickly transported in plastic vessels to the laboratory,here they were kept at 12 ◦C in well-water (composition inable 1) for 10 days before being used in experiments. Animalsere fed on alder leaves (Alnus glutinosa).Chemicals used were provided by Sigma–Aldrich (St.
uentin Fallavier, France).
.2. Experiment 1: LC50 assessment
This first experiment was aimed at determining the valuef cadmium LC50 of G. pulex coming from La Bourbre. Itas also a prerequisite for defining the design and the tested
oncentrations of the second one. Test solutions were preparedsing well-water (Table 1). Continuous exposure to cadmiumas performed to avoid chemical variation (500 mL tanksere renewed 4 times a day: flow rate = 1.34 mL min−1). Eightifferent experimental Cd concentrations were prepared fromhe acclimation water using daily renewed CdCl2 stock solutionf 22 mg Cd L−1 (nominal concentration: 0, 15.6, 31.3, 62.5,25, 250, 500 and 1000 �g L−1). Peristaltic pumps were usedo dilute stock solution and to prepare all experimental mediaFig. 1): dilutions were established by controlling the flow ofacclimation water” and stock solution or intermediate dilutionreal Cd concentration, mean ± S.D.: control <50 ng L−1,5.6 = 15.8 ± 0.75 �g L−1, 31.3 = 30.4 ± 2.53 �g L−1,25 = 122.7 ± 3.51 �g L−1, 500 = 490.8 ± 23.8 �g L−1, 62.5,50 and 1000 media were not measured). Four replicatesf 20 gammarids with 7–9 mm body size were exposed at2 ◦C during 264 h to the 8 treatments in 500 mL glass tankphotoperiod of 14 h of light and 10 h of darkness). Deadnimals were removed and survivors were counted at 4, 8,nd 12 h and thereafter at 12 hourly intervals until 216 h of
xposure after which the count occurs every 24 h. During thentire experiment, animals were fed ad libitum on disks oflder leaves (A. glutinosa, see below – Section 2.3.7 – for morenformation).
V. Felten et al. / Aquatic Toxicology 86 (2008) 413–425 415
F 1. Si(
2r
w7cfifChdpcto2mgs
rrwAarhpoceb
Nt
2
l
2
wsucwdssH1amstIaHvi
ig. 1. Schematic description of the flow-through system used in experiment330 �g Cd L−1).
.3. Experiment 2: behavioural and physiologicalesponses
Test solutions were prepared using natural water (Table 1)ith addition of CdCl2 to obtain cadmium concentration of 0,.5 and 15 �g L−1 (Fig. 1, stock solution: 330 �g Cd L−1). Theseoncentrations were defined according to results obtained in therst experiment. In these conditions, water osmolality rangedrom 9 to 11 mOsm kg−1. Males G. pulex in similar intermoult
stage with 7–9 mm body size were selected for the study ofaemolymph concentration and osmolality. Moult stages wereetermined by microscopical observation of the tip of posteriorereopods according to Blanchet-Tournier (1980). Nine repli-ates of 30 individuals were exposed at 12 ◦C for 168 h to thehree different cadmium conditions in 500 mL glass tank. Inrder to avoid chemical variations, the water was renewed every4 h after the daily survival count. During the entire experi-ent, animals were fed ad libitum on disks of alder leaves (A.
lutinosa, see below: Section 2.3.7). The experimental design ishown in Fig. 2.
Briefly, except for survival/feeding rate assessment, eacheplicate was associated with one or more measurements toetain the same exposure and to reduce the number of animalshile having sufficient numbers to conduct the experiments.fter 120 and 168 h of exposure, locomotor and ventilatory
ctivities were assessed, and samples of haemolymph wereandomly chosen from 10 organisms in each condition foraemolymph concentration and osmolality measurements. Sam-les of haemolymph were collected on three groups of 10
rganisms permitting the measure of osmolality, Ca2+ con-entrations and Na+/Cl− concentrations. At the end of thexposure period (168 h), organisms were sorted and storedefore cadmium accumulation measurement, feeding rate and2
pc
milar exposure system was used in experiment 2, using lower stock solution
a+/K+-ATPase activity assessments (see corresponding sec-ions).
.3.1. SurvivalEvery 12 h, dead animals were removed in each tank and
iving ones were counted in 4 replicates of 30 animals.
.3.2. Cadmium accumulationAfter 168 h of exposure, three replicates of five organisms
ere sampled, frozen (at −80 ◦C), freeze-dried, weighed andtored before cadmium accumulation analysis. The samplessed for metal analysis were pretreated according to a pro-edure that best avoids secondary contamination. All labwareas soaked in 10% hydrochloric acid, rinsed three times witheionised water and dried in a desiccator sheltered from atmo-pheric dust. The five-pooled gammarids were digested withuprapur nitric acid at 80 ◦C (500 �L corresponding to 1 mLNO3 for 30 mg dry weight of gammarids) for a minimum of2 h until a limpid solution was obtained. The volume was thendjusted to 3 mL with deionized water. Cadmium was deter-ined in these acid solutions by flameless atomic absorption
pectrophotometry (AAS) using the Zeeman effect accordingo the analytical method described by Amiard et al. (1987).nternal quality controls were based on the analysis of the met-ls of interest in standard reference material (TORT-2, Lobsterepatopancreas NRC-CNRC; certified values: 26.7 ± 0.6, ouralues: 26.8 ± 2.0). The cadmium accumulation was expressedn �g Cd g dry weight−1.
.3.3. Haemolymph samplingThe animals were gently dried between two sheets of filter
aper. Samples of haemolymph for osmolality and haemolymphoncentration assessments were taken from the telson of each

416 V. Felten et al. / Aquatic Toxicology 86 (2008) 413–425
l desi
im
2
mwoum
2c
a1dfNabca1(ms
2
(1twwpi5waftD4
astuaKK
Fig. 2. Experimenta
ndividual (n = 10) using either a modified microcapillary or aicrosyringe, respectively.
.3.4. Measurement of osmolalityA drop of haemolymph was immediately transferred into
ineral oil to avoid evaporation. The haemolymph osmolalityas measured in 30 nL samples with a Kalber–Clifton nano-smometer (Clifton Technical Physics, Hartford, NY, USA)sing a 300 mOsm kg−1 standard. Water osmolality was alsoeasured on a Wescor 5100c microosmometer.
.3.5. Analysis of haemolymph Na+, Cl− and Ca2+
oncentrationsSamples of haemolymph (0.8–1.2 �L) were transferred to
gauged 5 �L microcapillary tube and were centrifuged for0 min at 6596 × g. After centrifugation the liquid phase wasiluted in 2 mL of Nanopur water. Prior to analysis, samplesor Na+/Cl− and Ca2+ were fixed by acidification with HNO3ormapur (0.5%). Lanthanum chloride was added to acidified
liquots in a final proportion of 5% (w/v), to avoid possi-le interferences in the determination of Ca2+. Sodium andalcium concentrations in haemolymph were evaluated usingtomic absorption spectrophotometry (PerkinElmer Analyst
00) whereas chloride was measured by ionic chromatographyDionex 4500i equipped with a Ion Pac AS4A column). Forore details concerning haemolymph sampling and analysis,ee Felten and Guerold (2004).
paac
gn of the exposure.
.3.6. Gill Na+/K+-ATPase activityThe third coxal gills was removed and rinsed in a solution
pH 7.4) containing 300 mM sucrose, 20 mM Na2EDTA and00 mM imidazole (Zaugg, 1982), placed in tubes containinghe same solution and stored at −20 ◦C until use. During thehole procedure, samples were kept on ice. Stored samplesere thawed at room temperature and briefly centrifuged. Thereservation medium was then removed and 500 �L of isotonicsolation medium (MIIM: 250 mM sucrose, 5 mM MgCl2 andmM Hepes/Tris; pH 7.4) were added in each tube. Samplesere then homogenized in a glass potter homogenisation device
nd the homogenate was subsequently centrifuged at 4000 rpmor 5 min at 4 ◦C to eliminate debris. The supernatant containinghe plasma membrane fragments was transferred to new tubes.uring the extraction procedure, samples were maintained at◦C on ice.
The protocol for the Na+/K+-ATPase activity titration wasdapted from Flik et al. (1983) using microplates. Enzymaticpecific activity was expressed per mg protein. To this end, pro-ein was determined by a colorimetric method (Bio-Rad, France)sing BSA as reference. Na+/K+-ATPase specific activity wasssessed as the difference of ATP hydrolysis in presence of Na+,+, Mg2+ and ATP (100 mM NaCl, 5 mM MgCl2, 13.4 mMCl, 3 mM ATP, 0.1mM Na2EDTA, 30mM Imisadole/Hepes,
H 7.4), and that in the same medium without KCl, but withn optimal concentration of ouabain (1 mg mL−1; see Flik etl., 1983). The amount of phosphate released was colorimetri-ally assessed against a certified standard (Sigma, France). Three
oxicology 86 (2008) 413–425 417
ma
2
MaAiardporcroawmd
F
ww
C
wo
2
(dm(r
2
iA(orp
2
p((Lc
Fig. 3. Toxicity of cadmium to Gammarus pulex. LC50 as defined time pertime using logistic curve-fitting (Vindimian et al., 1999; Isnard et al., 2001).Llt
deotTw
3
3
pc2sLLoc
3r
3
rtwsrefs1
V. Felten et al. / Aquatic T
easures were performed on each replicate. The enzyme specificctivity was expressed in �mol Pi mg protein−1 h−1.
.3.7. Feeding rateThis measurement was based on the method developed by
altby (1992). Alder leaves were collected from the field afterbscission but before fall. The leaves were then dried and stored.fter a 5-day leaching, 2 cm diameter leaf discs were cut (tak-
ng care not to include major veins), dried at 105 ◦C for 24 hnd weighed (Li, leaf initial dry weight in mg). Leaf discs wereehydrated in tanks containing natural uncontaminated wateruring 24 h and were then placed with the gammarids (n = 12er tank: 180.7 ± 12 mg dry weight). Eight tanks, containingnly leaf material in control water, were used as control to cor-ect for weight loss due to leaching (C). The experimental designonsisted of three treatments replicated three times. The fourtheplicate served as specimen stock to replace a dead animal withne coming from a similar treatment. After 168 h, leaf discs andnimals were separated, counted, dried at 105 ◦C for 24 h andeighed (leaf final dry weight in mg: Lf; organisms weight ing: W). The feeding rate, expressed in mg leaf mg gammarid−1
ay−1, was calculated using the formula (see Maltby, 1992):
eeding rate = (LfC) − Li
WT(1)
here T was the exposure time (in days: 168 h, 7 days). Leaveseight changes due to leaching were corrected by C, defined as:
=∑
(Lb/La)
8(2)
here La and Lb refer to the initial and final dry weights (mg)f leaves in control tanks.
.3.8. Ventilatory activityIt was recorded by observation of the frequency of pleopod
ventilatory appendages of malacostracean crustaceans) beatinguring 1 min according to Hervant et al. (1997). The measure-ents were performed rapidly at the same period of the day
fixed hours: 13–14 h) to avoid the effects of a possible circadianhythm of respiration (Rosas et al., 1992).
.3.9. Locomotor activityIt was monitored by counting the number of moving animals
n 800 mL tank containing 10 individuals (Issartel et al., 2005).piece of net was added in the tank to provide a resting surface
mesh size 200 �m, length × width: 6 cm × 5 cm). Each countf moving G. pulex was conduced on a period of 2 s and wasepeated 30 times. Measurements were performed at the sameeriod of the day, with similar light conditions and without noise.
.3.10. Statistical analysisLC50s were calculated, time per time, by logistic curve-fitting
rocedure according to the method described by Vindimian et al.
1999) and Isnard et al. (2001). A Microsoft Excel® spreadsheetRedmond, WA, USA) enabled automated macro-calculation ofC50, and resampling procedure for the bootstrap estimations ofonfidence intervals was used. All physiological and behavioural3
tc
C50 values (�g L−1) are surrounded with 95% confidence interval (dottedines). Correlation significance estimated using Pearson coefficient correlationest (n = 18; ***p < 0.001).
ata are reported as mean ± S.D. Statistical comparisons ofxperimental data were performed by nonparametric analysisf variance (Kruskal–Wallis ANOVA) and Mann–Whitney U-est (comparison between control mean and exposed means).he analyses were carried out using STATISTICA (Microsoft),ith a probability limit of p ≤ 0.05 considered as significant.
. Results
.1. Experiment 1: LC50 assessment
The relationship between cadmium exposure time and G.ulex’s LC50 is illustrated in Fig. 3. Mortality rates in theontrols were very low at all exposure times (≤8% during64 h). A dose–effect relationship of cadmium was mea-ured: LC5024h = 1130.1 �g L−1, LC5048h = 494 �g L−1,C5096h = 82.1 �g L−1, LC50120h = 37.1 �g L−1,C50168h = 21.6 �g L−1, and LC50264h = 10.5 �g L−1. More-ver, the TL50 for 15.625 �g L−1 was close to 205 h, and thisoncentration was selected for experiment 2.
.2. Experiment 2: physiological and behaviouralesponses
.2.1. SurvivalThe survival rate of G. pulex exposed to the control water
emained high and above 94% (Fig. 4a). After 120 h of exposure,he survival rates of organisms exposed to 7.5 and 15 �g Cd L−1
ere significantly different from mean control values for theame exposure time (p < 0.05), reaching 90 ± 3 and 88 ± 6%,espectively. After 168 h, the survival rates of the animalsxposed to 7.5 �g Cd L−1 (86 ± 6%) were not significantly dif-erent from the rate observed in controls (94 ± 6%) whereasignificant differences were reported for organisms exposed to5 �g Cd L−1 (79 ± 6%).
.2.2. Cadmium accumulationAfter 168 h of exposure, increasing cadmium concentra-
ion produced a significant increase in G. pulex whole bodyoncentration of cadmium (Table 2b). The whole body con-

418 V. Felten et al. / Aquatic Toxicology 86 (2008) 413–425
Table 2Summary of Kruskal–Wallis ANOVA
Exposure time (h) Kruskall–Wallis ANOVA
H-value d.f. N p-Value
(a) Survival 120 7.7 2 12 <0.05168 7 2 12 <0.05
(b) Cadmium accumulation 168 7.2 2 9 <0.05
(c) Heamolymph osmolality 120 13 2 30 <0.01168 10.3 2 30 <0.01
(d) Heamolymph Na+ 120 4.1 2 30 n.s.168 2.7 2 30 n.s.
(e) Heamolymph Cl− 120 0.4 2 30 n.s.168 1.2 2 30 n.s.
(f) Heamolymph Ca2+ 120 8.8 2 30 <0.05(g) Na+/K+-ATPase activity 168 11.8 2 27 <0.01(h) Feeding rate 168 3.5 2 9 n.s.
(i) Ventilatory activity 120 7.4 2 30 <0.05168 17.9 2 30 <0.001
(j) Locomotor activity 120 23.9 2 90 <0.00139
H ility.
cwfe2
F�
cM
c
168
-value, H-ratio; d.f., degrees of freedom; N, number of value; p-value, probab
entration of cadmium in G. pulex exposed to the control
ater was 0.28 ± 0.02 �g Cd g dry weight−1, which was 30-old and 76-fold lower than those measured for organismsxposed to 7.5 and 15 �g Cd L−1, respectively (8.65 ± 1.77 and1.51 ± 2.85 �g Cd g dry weight−1; Fig. 4b; p < 0.05). The con-
ig. 4. Mean (±S.D.) survival (a) and cadmium accumulation expressed ing Cd g dry weight−1 (b) of Gammarus pulex exposed to cadmium. Signifi-ant differences against the controls (0 �g L−1) are indicated by asterisks (n = 3;ann–Whitney U-test; *p < 0.05).
(e
3
t2TE(d1Ss12
3
i1rh7mCthw1Fc
.4 2 90 <0.001
entration of cadmium in G. pulex was significantly higher2.5-fold) in organisms exposed to 15 �g Cd L−1 tan in animalsxposed to 7.5 �g Cd L−1.
.2.3. Haemolymph osmolalityThe baseline level of haemolymph osmolality in con-
rol organisms remained constant and was 285 ± 5 and88 ± 8 mOsm kg−1 after 120 and 168 h, respectively (Fig. 5a).he distribution range of control organism osmolality was small.xposure to cadmium decreased the haemolymph osmolality
Table 2c): after 120 h of exposure, haemolymph osmolalityecreased significantly by 8% at 7.5 �g Cd L−1 and by 9% at5 �g Cd L−1 (261 ± 17 and 260 ± 28 mOsm kg−1; Fig. 5a).lightly higher decreases were noted following a 168 h expo-ure to cadmium: lower values compared to controls, 9 and1% at 7.5 and 15 �g Cd L−1, respectively (262 ± 23 and56 ± 25 mOsm kg−1).
.2.4. Haemolymph ion concentrationsThe baseline level of haemolymph Na+ concentrations
n control organisms remained constant with values of04.7 ± 8.8 and 108.2 ± 12.6 mmol L−1 after 120 and 168 h,espectively (Fig. 5b). The same trend was reported foraemolymph Cl− concentrations in control organisms (120 h:3.5 ± 10 mmol L−1; 168 h: 79 ± 19 mmol L−1; Fig. 5c). Cad-ium had no significant effect on haemolymph Na+ andl− concentrations (Table 2d and e). However, compared
o controls (31 ± 7.4 mmol L−1), a significant reduction ofaemolymph Ca2+ concentration in G. pulex by 24 and 37%
as observed in organisms exposed for 120 h to 7.5 and5 �g Cd L−1, respectively (24.6 ± 5.4 and 20.4 ± 4 mmol L−1;ig. 5d and Table 2f). Results in haemolymph Ca2+ con-entration after 168 h of exposure to cadmium are not
V. Felten et al. / Aquatic Toxicology 86 (2008) 413–425 419
Fig. 5. Mean (±S.D.) osmolality (a), haemolymph [Na+] (b), [Cl−] (c) and[aU
pt
3
2tA
Fcb
t(7s
3
lco(cg
3
rbo1td3o1t(2
3
n(s5tity decreased significantly by 20% at 7.5 �g Cd L and by 31%
Ca2+] (d) of Gammarus pulex exposed to cadmium. Significant differencesgainst the controls (0 �g L−1) are indicated by asterisks (n = 10; Mann–Whitney-test; *p < 0.05; **p < 0.01).
resented here due to the loss of these samples (brokenubes).
.2.5. Gill Na+/K+-ATPase activity
The Na+/K+-ATPase activity of control organisms was.59 ± 0.22 �mol Pi mg protein−1 h−1 (Fig. 6). Exposureo cadmium concentration tended to increase gill Na+/K+-TPase activity of G. pulex (Table 2g): after 168 h of exposure,
arma
ig. 6. Mean (±S.D.) Na+/K+-ATPase activity of Gammarus pulex exposed toadmium. Significant differences against the controls (0 �g L−1) are indicatedy asterisks (n = 9; Mann–Whitney U-test; ***p < 0.001).
he enzyme activity increased by 19% at 7.5 �g Cd L−1
3.08 ± 0.94 �mol Pi mg protein−1 h−1; not significant) and by1% at 15 �g Cd L−1 (4.42 ± 0.49 �mol Pi mg protein−1 h−1;ignificant) (Fig. 6).
.2.6. Feeding rateThe feeding rate of control organisms was 0.058 ± 0.014 mg
eaf mg gammarid−1 day−1 (Fig. 7a). Exposure to cadmium con-entration tended to decrease G. pulex feeding rate: after 168 hf exposure, feeding rate decreased by 30% at 7.5 �g Cd L−1
0.041 ± 0.012 mg leaf mg gammarid−1 day−1; not signifi-ant) and by 36% at 15 �g Cd L−1 (0.037 ± 0.003 mg leaf mgammarid−1 day−1; significant) (Fig. 7a).
.2.7. Ventilatory activityIncreasing cadmium concentration produced a significant
eduction of ventilation in G. pulex (Table 2i). The pleopodeat (PB) number of control organisms remained constantver a 168 h exposure period (120 h: 227.8 ± 7.7 PB min−1;68 h: 230.4 ± 7.0 PB min−1; Fig. 7b). After 120 h, the ventila-ion of animals exposed to 7.5 �g Cd L−1 was not significantlyifferent from ventilation in controls (220.8 ± 10.1 PB min−1;.1%), whereas significant hypoventilation was observed inrganisms exposed to 15 �g Cd L−1 (199.2 ± 51.9 PB min−1;2.6%). After a 168 h exposure to cadmium, animals exposedo both cadmium concentrations hypoventilaed significantly7.5 �g Cd L−1: 217.2 ± 11.2 PB min−1, −5.7%; 15 �g Cd L−1:02.2 ± 15.2 PB min−1, −12.2%).
.2.8. Locomotor activityThe exposure to cadmium contaminated waters led to a sig-
ificant reduction in the percentage of G. pulex in movementTable 2j). Locomotor activity of control G. pulex remained con-tant over a 168 h exposure period (120 h: 56.3 ± 12.9%; 168 h:6.3 ± 11.6%), but decreased significantly in organisms exposedo cadmium (Fig. 7c). After 120 h of exposure, locomotor activ-
−1
t 15 �g Cd L−1 (45.3 ± 11.4 and 38.7 ± 11.1%; Fig. 7c). Theseeductions were more pronounced after a 168 h exposure to cad-ium: reduced by 38% (34.7 ± 12.5%) and 39% (34.3 ± 11.6%)
t 7.5 and 15 �g Cd L−1, respectively.
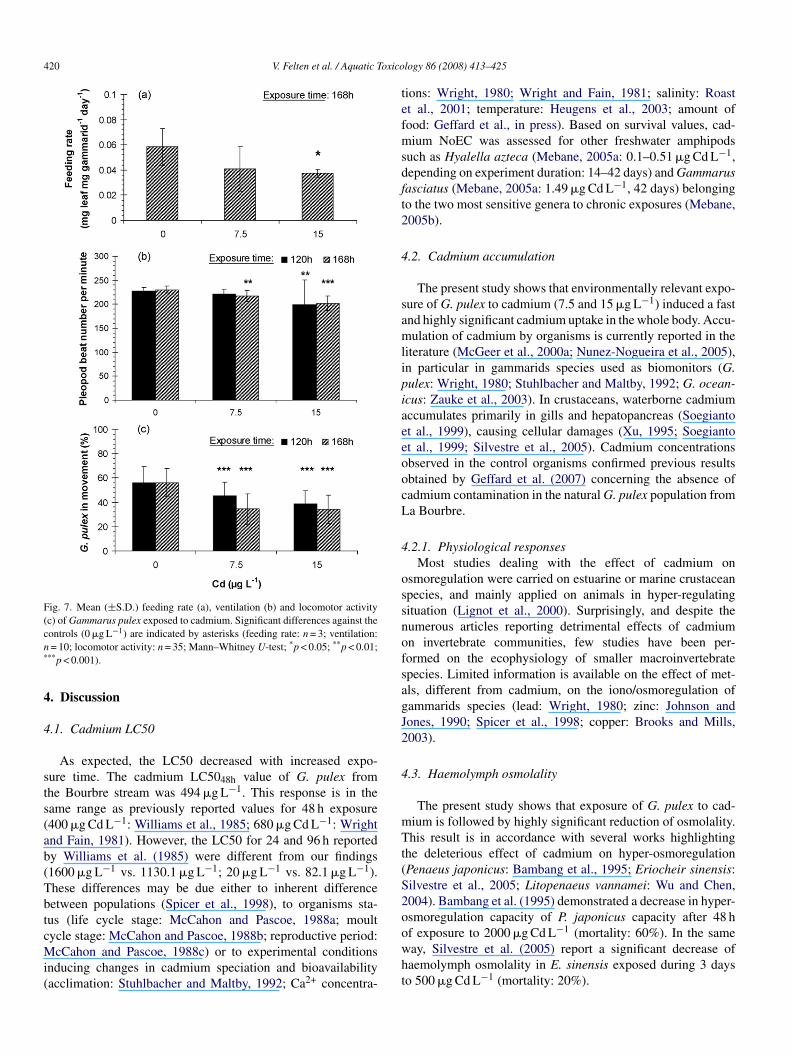
420 V. Felten et al. / Aquatic Toxico
Fig. 7. Mean (±S.D.) feeding rate (a), ventilation (b) and locomotor activity(c) of Gammarus pulex exposed to cadmium. Significant differences against thecn*
4
4
sts(ab(TbtcMi(
tefmsdft2
4
samlipiaeeoocL
4
ossnofsagJ2
4
mTt(S2o
−1
ontrols (0 �g L−1) are indicated by asterisks (feeding rate: n = 3; ventilation:= 10; locomotor activity: n = 35; Mann–Whitney U-test; *p < 0.05; **p < 0.01;
**p < 0.001).
. Discussion
.1. Cadmium LC50
As expected, the LC50 decreased with increased expo-ure time. The cadmium LC5048h value of G. pulex fromhe Bourbre stream was 494 �g L−1. This response is in theame range as previously reported values for 48 h exposure400 �g Cd L−1: Williams et al., 1985; 680 �g Cd L−1: Wrightnd Fain, 1981). However, the LC50 for 24 and 96 h reportedy Williams et al. (1985) were different from our findings1600 �g L−1 vs. 1130.1 �g L−1; 20 �g L−1 vs. 82.1 �g L−1).hese differences may be due either to inherent differenceetween populations (Spicer et al., 1998), to organisms sta-us (life cycle stage: McCahon and Pascoe, 1988a; moult
ycle stage: McCahon and Pascoe, 1988b; reproductive period:cCahon and Pascoe, 1988c) or to experimental conditionsnducing changes in cadmium speciation and bioavailabilityacclimation: Stuhlbacher and Maltby, 1992; Ca2+ concentra-
owht
logy 86 (2008) 413–425
ions: Wright, 1980; Wright and Fain, 1981; salinity: Roastt al., 2001; temperature: Heugens et al., 2003; amount ofood: Geffard et al., in press). Based on survival values, cad-ium NoEC was assessed for other freshwater amphipods
uch as Hyalella azteca (Mebane, 2005a: 0.1–0.51 �g Cd L−1,epending on experiment duration: 14–42 days) and Gammarusasciatus (Mebane, 2005a: 1.49 �g Cd L−1, 42 days) belongingo the two most sensitive genera to chronic exposures (Mebane,005b).
.2. Cadmium accumulation
The present study shows that environmentally relevant expo-ure of G. pulex to cadmium (7.5 and 15 �g L−1) induced a fastnd highly significant cadmium uptake in the whole body. Accu-ulation of cadmium by organisms is currently reported in the
iterature (McGeer et al., 2000a; Nunez-Nogueira et al., 2005),n particular in gammarids species used as biomonitors (G.ulex: Wright, 1980; Stuhlbacher and Maltby, 1992; G. ocean-cus: Zauke et al., 2003). In crustaceans, waterborne cadmiumccumulates primarily in gills and hepatopancreas (Soegiantot al., 1999), causing cellular damages (Xu, 1995; Soegiantot al., 1999; Silvestre et al., 2005). Cadmium concentrationsbserved in the control organisms confirmed previous resultsbtained by Geffard et al. (2007) concerning the absence ofadmium contamination in the natural G. pulex population froma Bourbre.
.2.1. Physiological responsesMost studies dealing with the effect of cadmium on
smoregulation were carried on estuarine or marine crustaceanpecies, and mainly applied on animals in hyper-regulatingituation (Lignot et al., 2000). Surprisingly, and despite theumerous articles reporting detrimental effects of cadmiumn invertebrate communities, few studies have been per-ormed on the ecophysiology of smaller macroinvertebratepecies. Limited information is available on the effect of met-ls, different from cadmium, on the iono/osmoregulation ofammarids species (lead: Wright, 1980; zinc: Johnson andones, 1990; Spicer et al., 1998; copper: Brooks and Mills,003).
.3. Haemolymph osmolality
The present study shows that exposure of G. pulex to cad-ium is followed by highly significant reduction of osmolality.his result is in accordance with several works highlighting
he deleterious effect of cadmium on hyper-osmoregulationPenaeus japonicus: Bambang et al., 1995; Eriocheir sinensis:ilvestre et al., 2005; Litopenaeus vannamei: Wu and Chen,004). Bambang et al. (1995) demonstrated a decrease in hyper-smoregulation capacity of P. japonicus capacity after 48 h
f exposure to 2000 �g Cd L (mortality: 60%). In the sameay, Silvestre et al. (2005) report a significant decrease ofaemolymph osmolality in E. sinensis exposed during 3 dayso 500 �g Cd L−1 (mortality: 20%).
oxico
4
oats(nma
hCMsaP(cNSittoca(
ctnR(tMthmco
4
AmharaemeAT
oaNgrec
4
thlptfl
i1(
2tnfawHaa
iwit−aeNhiwsamc
4
aio
V. Felten et al. / Aquatic T
.4. Haemolymph ion concentrations
In G. pulex, the decrease in osmolality is not linked to lossesf the major haemolymph ions (Na+ and Cl−) which are notffected by cadmium, but is partly due to the significant reduc-ion in haemolymph Ca2+ concentration. Although calcium lossubsequent to cadmium exposure is commonly reported in fishOreochromis mossambicus: Pratap et al., 1989; Salmo gaird-eri: Verbost et al., 1989; McGeer et al., 2000b; Oncorhynchusykiss: Chowdhury et al., 2004), the results concerning sodium
nd chloride concentrations are more variable.According to Silvestre et al. (2005), the decrease in
aemolymph osmolality in E. sinensis results from Na+ andl− losses (3-day exposure to 500 �g Cd L−1, mortality: 20%).cGeer et al. (2000b) reports that a chronic cadmium expo-
ure (3 �g L−1 during 65 days) leads to severe losses of Na+
nd Ca2+ in rainbow trout (as soon as 2 days of exposure). Butratap et al. (1989) showed that waterborne cadmium exposure10 �g L−1 during 35 days) exerts no effect on plasma Na+ con-entration of O. mossambicus whereas hypocalcemia is reported.a+ loss following cadmium exposure (McGeer et al., 2000b;ilvestre et al., 2005) seems to be mainly due to differences
n experimental design (cadmium concentration and exposureime): in a preliminary experiment with high cadmium concen-ration exposure (15–1000 �g L−1, see experiment 1), after 96 hf exposure, we observed significant Na+ loss at the higher con-entrations (≥125 �g Cd L−1) which was accompanied by highnd significant G. pulex mortalities as LC5096h = 82.1 �g L−1
unpublished results).It has been suggested that cadmium enters mainly through
hloride cells in the fish gill (Wicklund-Glynn et al., 1994) viahe same uptake mechanisms as calcium, such as Ca2+ chan-els, Na+/Ca2+ exchangers or Ca2+-ATPase (Verbost et al., 1989;ainbow, 1997). Thus, a Cd-induced inhibition of Ca2+ influx
Verbost et al., 1987, 1989) has been linked to competition athe apical uptake channels (Playle et al., 1993a,b). According to
cGeer et al. (2000a), cadmium exposure conducts to a deple-ion of Ca2+ which can ultimately lead to death. On the otherand, Xu (1995) demonstrated that G. pulex exposed to cad-ium (14.5 �g Cd L−1 for 2 days) have lower free amino acids
oncentrations, which can also partly explain the decrease ofsmolality in exposed organisms.
.5. Na+/K+-ATPase activity
Cadmium exposure significantly increased the Na+/K+-TPase activity in G. pulex gills, which could be a compensatoryechanism explaining the absence of cadmium effect on
aemolymph Na+ concentration (induction of Na+/K+-ATPasectivity and/or increase of ionocytes number/surface). Thisesult is in disagreement with numerous papers reporting
reduction in Na+/K+-ATPase activity following cadmiumxposure (Anguilla anguilla, in vitro: Lionetto et al., 2000; Chas-
agnathus granulata, in vitro and in vivo: Rodriguez Morenot al., 1998). In this last crab species, an inhibition of Na+/K+-TPase was induced by cadmium (250 �g Cd L−1 for 18 days).hus, according to Silvestre et al. (2005), a possible mode
bloe
logy 86 (2008) 413–425 421
f action concerns the direct effect on enzymes involved inctive ionic transport and located in gill epithelial cells (e.g.,a+/K+-ATPase and carbonic anhydrase) or an impairment ofill permeability. As for the haemolymph Na+ concentrationesults, response differences can be explained by the differ-nce in experimental design, since in these studies the testedoncentration were higher than in the present one.
.6. Synthesis
The body fluid of all freshwater organisms including crus-aceans is hyperosmotic to the aqueous environment. Thus, theseyperosmotic regulators are faced by water invasion and ionosses. To maintain osmolality level, most but not all organismsroduce dilute urine to eliminate excess water and perform activeransport of salts from the water across the gills into the bodyuids (reviews in Mantel and Farmer, 1983; Pequeux, 1995).
In this study, the sum of the considered haemolymphonic concentration in control G. pulex for a20 h exposure was 241 mOsm kg−1 approximatelyNa+ + Cl− + 2Ca2+ = 105 + 74 + 2 × 31).
The measured haemolymph osmolality mean value was85 mOsm kg−1 which was near to the total of these three ions:he difference can be explained by other ions (potassium, mag-esium, sulfates, . . .) and by organic osmoeffectors (such asree amino acids and glucids). In G. pulex, haemolymph Na+
nd Cl− concentrations made up 66% of the osmotic pressurehich is less than in most crustacean (Mantel and Farmer, 1983).owever, osmotic coefficient (undissociation of the ions), usu-
lly neglected in physiological studies, has not been taken intoccount.
Cadmium exposure led to a decrease in G. pulex’s osmolal-ty (168 h exposure to 15 �g Cd L−1: −25 mOsm kg−1, −11%)hich is not explained by the losses of the major haemolymph
ons (Na+ and Cl−), but probably partially due to the reduc-ion in haemolymph Ca2+ concentrations (−11 meq L−1, i.e.,
22 mOsm kg−1, −33%). Simultaneouslyy, Na+/K+-ATPasectivity is increased by cadmium exposure (+71%). How-ver, this is not associated with an increase in haemolympha+ concentration, suggesting a compensatory mechanism. Ourypothesis is that cadmium exposure also induces an increasen ion losses and water invasion, perhaps through altered gills,hich tends to dilute haemolymph Na+, which is compen-
ated by the increase in Na+/K+-ATPase activity (number and/orctivity of Na+/K+-ATPase). In the same way, compensatoryechanisms could explain the maintenance of haemolymph Cl−
oncentration.
.6.1. Behavioural responsesAnimal behaviour is being used increasingly as a sensitive
nd integrated measure of sub-lethal exposure to toxic contam-nants. Cadmium exposure is known to exert significant effectsn behaviour, such as filtration activity in the bivalve Macoma
althica (Duquesne et al., 2004), time spent fighting by intruderarvae of Hydropsyche contubernalis (Vuori, 1994), disruptionf swimming (ability and orientation) in Neomysis integer (Roastt al., 2000), capture efficiency of prey in Salvelinus fontinalis
4 oxico
((
4
iwitcdstaTmti
4
c7ftenh(BtrcSsitcgCc
4
aIpaes≥vei
sMcN
4i
soatI(tttcaewbaualviaef(fah(zaFoClbfsrd
4
d
22 V. Felten et al. / Aquatic T
Riddell et al., 2005) and precopulatory behaviour in G. pulexPoulton and Pascoe, 1990).
.7. Feeding rate
The exposure to cadmium induced significant lowered feed-ng rate in G. pulex. Although a 30% decrease in feeding activityas observed for a 168 h exposure to 7.5 �g Cd L−1, this effect
s not significant due to the number of replicates (only 3). Inhe literature, nutrition is commonly reported to be impacted byadmium exposure (Chironomidae: Heinis et al., 1990; Baeti-ae: Riddell et al., 2005). Thus, Heinis et al. (1990) showedlight deviations in the feeding pattern of the filter feeder Glyp-otendipes pallens exposed during 96 h to 100 �g Cd L−1, withmuch greater variability in the length of the pumping period.he grazing rate of Baetis tricaudatus was also affected by cad-ium exposure (0.5 and 5 �g Cd L−1). In a series of field studies,
he feeding rate of G. pulex was demonstrated to be a sensitivendicator of water quality (Crane and Maltby, 1991).
.8. Ventilatory activity
The ventilatory activity of G. pulex decreased signifi-antly within 120 h of exposure to 15 �g Cd L−1, and also at.5 �g Cd L−1 following a 168 h exposure. To our knowledge,ew studies have reported the effect of cadmium on ventila-ion: no effect was detected in rainbow trout by Chowdhuryt al. (2004), whereas Bishop and McIntosh (1981) found sig-ificant increases of bluegill ventilation frequency. However,ypoventilation was observed in gammarids exposed to copperGammarus duebeni: Lawrence and Poulter, 1998; G. pulex:rooks et al., 2002). According to Lawrence and Poulter (1998),
he pleopod beat number can be used as a measure of respi-ation rates. Oxygen consumption generally decreases whenrustaceans are acutely exposed to heavy metals (reviewed inpicer and Weber, 1991). For instance, Wu and Chen (2004)howed deleterious effect of cadmium on oxygen consumptionn the white shrimp L. vannamei. Chinni et al. (2000) showedhat the reduction in oxygen consumption might be related toytological damage (i.e., necrosis, Soegianto et al., 1999), sinceills are most likely the first target of waterborne heavy metals.admium also interfered with the respiratory system, includingellular respiration (Spicer and Weber, 1991; Sokolova, 2004).
.9. Locomotor activity
A significant reduction in locomotor activity was observedfter 120 and 168 h of exposure to 7.5 and 15 �g Cd L−1.n earlier studies, chemicals such as lead, copper, cadmium,harmaceuticals have been shown to decrease the locomotorctivity of gammarids (Wallace and Estephan, 2004; De Langet al., 2006; Mills et al., 2006). Wallace and Estephan (2004)howed that a 72 h exposure of Gammarus lawrencianus to [Cd]
500 �g L−1 lead to a significant decrease of horizontal andertical swimming whereas 72 h exposure to [Cd] ≥125 �g L−1
xerts only a significant reduction of vertical swimming. Stud-es carried on other heavy metals, such as copper, report the
trCt
logy 86 (2008) 413–425
ame trend (i.e., locomotor activity reduction, Mills et al., 2006).oreover, several works highlighted the deleterious effect of
admium on locomotor activity (G. pallens: Heinis et al., 1990;. integer: Roast et al., 2001).
.10. Linking physiological and behavioural endpoints:mplication for populations
The physiological and behavioural responses of G. pulex wereeverely impacted by cadmium exposure. The analysis of physi-logical responses shows that compensatory mechanisms (suchs the induction of Na+/K+-ATPase activity) take place to main-ain homeostasis. These mechanisms were energy-consuming.n response, organisms can have three nonexclusives choices:1) the increase of the entry of energy (e.g., nutrition), (2)he draw on its energetic reserves and/or (3) the limitation ofhe loss of energy in nonvital functions. However, we showedhat the feeding rate was significantly reduced subsequent toadmium exposure whereas significant decreases in locomotornd ventilatory activities were simultaneously reported, whichxcludes the first possibility and suggests that the third oneas selected. Thus, although a direct effect of cadmium cannote excluded, decreases observed in locomotor and ventilatoryctivities might be induced by redirection of energy for osmoreg-lation compensation mechanisms, such as Na+/K+-ATPasectivity (activity, number/surface of ionocytes), which is directlyinked to energetic pathways. Lawrence and Poulter (1998) pre-iously hypothesised that the hypoventilation and the decreasen swimming endurance as a result of metal exposure indicatereduction or redirection of available energy from these mark-
rs to detoxification (metallothioneins, stress proteins, mixedunctional enzyme systems). According to Maltby and Naylor1990), zinc exposure induced a significant reduction in scopeor growth of brooding females over a 7-day period (main energybsorbed: leaf energy consumed minus energy lost in faeces), butad also more long-term effects (2 months) on their reproductionoffspring size and abortion rate). Thus, the authors suggest thatinc acts on the reproductive output of G. pulex thought energyllocation rather than by killing broodlings or eggs. Moore andarrar (1996) also showed that growth rates and reproductionf H. azteca decrease significantly with reduced food rations.onsequently, such stress may profoundly influence the popu-
ation as a whole. Movement is a highly ecologically relevantehavioural marker as well, since locomotion is required to findood, escape predation and obtain mates. Thus, any pollutant,uch as cadmium, that interferes with this activity is likely toeduce the fitness of organisms and could involve ‘ecologicaleath’ (Scott and Sloman, 2004).
.11. Implications for biotic ligand model (BLM)
The present study shows that cadmium exposure induceseleterious effects on iono/osmoregulation (particularly a reduc-
ion of haemolymph Ca2+ concentrations) and that biologicalesponses (physiological and behavioural) should be related toa2+ reduction. Results of our study are particularly relevanto mechanisms of toxicity postulated by biotic ligand model.

oxico
Bcb1mtisluEtiol
5
AwCpmNmhitsan
pelcishmtpftl(on
A
dlrF
R
A
B
B
B
B
B
B
B
B
C
C
C
C
D
D
F
F
F
F
V. Felten et al. / Aquatic T
LM is a conceptual framework which has been developed toharacterize the modifying effects of water chemistry on metalioavailability (bio-accumulation) and toxicity (Playle et al.,993c; Paquin et al., 2002; Niyogi and Wood, 2004). In thisodel, gill membrane (and particularly ionocytes) is defined as
he primary site of toxic action and is considered as complex-ng ligands. Based on physical and chemical characteristics oftudied water, BLM has been used to predict metal bioaccumu-ation and toxicity. However, this model has been developedsing biological responses more or less ecological relevant.ven if more work is needed to clarify direct mechanisms of
oxicity, our results support the interest on continually develop-ng BLM in this species, allowing to predict cadmium impactsn ecological relevant responses from exposure contaminationevels.
. Conclusion
Osmolality, haemolymph Ca2+ concentration, Na+/K+-TPase activity, ventilation, feeding rate and locomotor activityere all impacted by cadmium, but not haemolymph Na+ andl− concentration. Each response provides information on G.ulex health. Although the main mechanisms of action of cad-ium on osmoregulation is the alteration of Ca2+ regulation,a+ regulation seems to be also affected since compensatoryechanisms (Na+/K+-ATPase activity) is induced to maintain
aemolymph Na+ concentration. We showed that a disturbancen an essential regulatory function, such as osmoregulation, ledo an energetic reallocation to the detriment of other functions,uch as locomotor activity, which reduce the organism fitnessnd may lead to deleterious impact on population and commu-ities.
Locomotor activity is one of the most pronouncedly affectedarameter. This behavioural response is highly relevant as ancological marker reflecting potential effects at the populationevel. The haemolymph osmolality response (1) indicates globalhanges in haemolymph composition, (2) shows low variabil-ty in control organisms, (3) is linked to locomotor activity,howing its relevance as a physiological health marker. Thus,aemolymph osmolality appears as an effective and relevantarker of G. pulex physiological health. The application of this
ool may grant a significant contribution to the assessment, inter-retation and understanding of the impact of contaminants inreshwater ecosystem. Further studies are needed to elucidatehe mechanisms involved at the cellular (structure of osmoregu-ators organ: gill epithelium, green gland) and molecular levelsNa+/K+-ATPase activity in green gland), to assess the relevancef these markers with other pollutants and also to determine theiratural variability in different gammarid populations (in situ).
cknowledgements
We would like to thank Francois Guerold (Laboratoire Bio-
iversite et Fonctionnement des Ecosystemes, Metz, France) forending different instruments, Fabien Aujoulat et Lysiane Dher-et for technical help. We received financial support from therench program ECODYN (ANR “Ecosphere Continentale”).G
logy 86 (2008) 413–425 423
eferences
miard, J.-C., Pineau, A., Boiteau, H.L., Metayer, C., Amiard-Triquet, C., 1987.Application de la spectrometrie d’absorption atomique Zeeman aux dosagesde huit elements traces (Ag, Cd, Cr, Cu, Mn, Ni, Pb et Se) dans les matricesbiologiques solides. Water. Res. 21, 696–697.
ambang, Y., Charmantier, G., Thuet, P., Trilles, J.-P., 1995. Effect of cad-mium on survival and osmoregulation of various developmental stages ofthe shrimp Penaeus japonicus (Crustacea: Decapoda). Mar. Biol. 123, 443–450.
arnard, J.L., Barnard, C.M., 1983. Freshwater Amphipoda of the World (PartsI & II). Hayfield Associates, Mt. Vernon.
ervoets, L., Voets, J., Covaci, A., Chu, S., Qadah, D., Smolders, R., Schepens,P., Blust, R., 2005. Use of transplanted zebra mussels (Dreissena polymor-pha) to assess the bioavailability of microcontaminants in Flemish surfacewaters. Environ. Sci. Technol. 39, 1492–1505.
iesinger, K.E., Christensen, G.M., 1972. Effect of various metals on survival,growth, reproduction and metabolism of Daphnia magna. J. Fish. Res. BoardCan. 29, 1691–1700.
ishop, W.E., McIntosh, A.W., 1981. Acute lethality and effects of sub-lethal cadmium exposure on ventilation frequency and cough rate ofbluegill (Lepomis macrochirus). Arch. Environ. Contam. Toxicol. 10, 519–530.
lanchet-Tournier, M.F., 1980. Mue et vitellogenese chez le crustace amphipodeOrchestia gammarellus (Pallas): controles endocrines et interactions. Thesis.Paris 6 University, 198 pp.
rooks, S.J., Mills, C.L., 2003. The effect of copper on osmoregulation in thefreshwater amphipod Gammarus pulex. Comp. Biochem. Physiol. 135A,527–537.
rooks, S., Foster, J.L., Mills, C.L., 2002. The effect of copper on heart rate,pleopod beat and osmoregulation in the freshwater amphipod Gammaruspulex. Abstracts/Comp. Biochem. Physiol. 132A, S1–S12.
anadian Environmental Protection Act, 1994. Priority Substances List, Assess-ment Report, Cadmium and its Compounds. Government of Canada.
hinni, S., Khan, R.N., Yallapragada, P.R., 2000. Oxygen consumption,ammonia-N excretion, and metal accumulation in Penaeus indicus postlarvaeexposed to lead. Bull. Environ. Contam. Toxicol. 64, 144–151.
howdhury, M.J., Pane, E.F., Wood, C.M., 2004. Physiological effects of dietarycadmium acclimation and waterborne cadmium challenge in rainbow trout:respiratory, ionoregulatory, and stress parameters. Comp. Biochem. Physiol.139C, 163–173.
rane, M., Maltby, L., 1991. The lethal and sub-lethal response of Gammaruspulex to stress: sensitivity and variation in an in situ bioassay. Environ.Toxicol. Chem. 10, 1331–1339.
e Lange, H.J., Noordoven, W., Murk, A.J., Lurling, M., Peeters, E.T.,2006. Behavioural responses of Gammarus pulex (Crustacea, Amphipoda)to low concentrations of pharmaceuticals. Aquat. Toxicol. 78, 209–216.
uquesne, S., Liess, M., Bird, D.J., 2004. Sub-lethal effects of metal exposure:physiological and behavioral responses of the estuarine bivalve Macomabalthica. Mar. Environ. Res. 58, 245–250.
elten, V., Guerold, F., 2001. Hyperventilation and loss of hemolymph Na+ andCl− in the freshwater amphipod Gammarus fossarum exposed to acid stress:a preliminary study. Dis. Aquat. Org. 45, 77–80.
elten, V., Guerold, F., 2004. Haemolymph [Na+] and [Cl−] loss in Gammarusfossarum exposed in situ to a wide range of acidic streams. Dis. Aquat. Org.61, 113–121.
lik, G., Wendelaar Bonga, S.E., Fenwich, J.C., 1983. Ca2+-dependent phos-phatase and ATPase activities in eel gill plasma membranes. I. Identificationof Ca2+-activated activities with non-specific phosphatase activities. Comp.Biochem. Physiol. 76B, 745–754.
orrow, D.M., Maltby, L., 2000. Toward a mechanistic understanding ofcontaminant-induced changes in detritus processing in streams: direct
and indirect effects on detritivore feeding. Environ. Toxicol. Chem. 19,2100–2106.effard, A., Queau, H., Dedourge, O., Biagiant-Risbourg, S., Geffard, O., 2007.Influence of biotic and abiotic factors on metallothionein level in Gammaruspulex. Comp. Biochem. Physiol. 145C, 631–640.

4 oxico
G
H
H
H
H
I
I
J
L
L
L
M
M
M
M
M
M
M
M
M
M
M
M
M
M
N
N
P
P
P
P
P
P
P
R
R
R
R
R
R
24 V. Felten et al. / Aquatic T
effard, O., Geffard, A., Chaumot, A., Alvarez, C., Vollat, B., Tusseau-Vuillemin, M.-H., Garric, J. Effects of chronic dietary and waterborne Cdexposures on the contamination level and reproduction of Daphnia magna.Environ. Toxicol. Chem, in press.
einis, F., Timmermans, K.R., Swain, W., 1990. Short-term sublethal effectsof cadmium on the filter feeding chironomid larva Glyptotendipes pallens(Meigen) (Diptera). Aquat. Toxicol. 16, 73–86.
enry, R.P., Wheatly, M.G., 1992. Interaction of respiration, ion regulation, andacid–base balance in the everyday life of aquatic crustaceans. Am. Zool. 32,407–416.
ervant, F., Mathieu, J., Barre, H., Simon, K., Pinon, C., 1997. Comparativestudy on the behavioral, ventilatory and respiratory responses of hypogeanand epigean crustacean to long-term starvation and subsequent feeding.Comp. Biochem. Physiol. 118A, 1277–1283.
eugens, E.H.W., Jager, T., Creyghton, R., Kraak, M.H.S., Hendriks, A.J., VanStraalen, N.M., Admiraal, W., 2003. Temperature-dependent effects of cad-mium on Daphnia magna: accumulation versus sensitivity. Environ. Sci.Technol. 37, 2145–2151.
snard, P., Flammarion, P., Roman, G., Babut, M., Bastien, P., Bintein, S.,Essermeant, L., Ferard, J.F., Gallotti-schmitt, S., Saouter, E., Saroli, M.,Thiebaud, H., Tomassone, R., Vindimian, E., 2001. Statistical analysis ofregulatory ecotoxicity tests. Chemosphere 45, 659–669.
ssartel, J., Hervant, F., Voituron, Y., Renault, D., Vernon, P., 2005. Behavioural,ventilatory and respiratory responses of epigean and hypogean crustaceansto different temperatures. Comp. Biochem. Physiol. 141A, 1–7.
ohnson, I.T., Jones, M.B., 1990. Effect of zinc on osmoregulation of Gammarusduebeni (Crustacea: Amphipoda) from the estuary and the sewage treatmentworks at Looe, Corwall. Ophelia 31, 187–196.
awrence, A.J., Poulter, C., 1998. Development of a sub-lethal pollution biossayusing the estuarine amphipod Gammarus duebeni. Water. Res. 32, 569–578.
ignot, J.H., Spanings-Pierrot, C., Charmantier, G., 2000. Osmoregulatorycapacity as a tool in monitoring the physiological condition and the effectof stress in crustaceans. Aquaculture 191, 209–245.
ionetto, M.O., Giordano, M.E., Vilella, S., Schettino, T., 2000. Inhibition ofeel enzymatic activities by cadmium. Aquat. Toxicol. 48, 561–571.
acNeil, C., Elwood, R.W., Dick, J.T.A., 2000. Factors influencing the impor-tance of Gammarus spp. (Crustacea: Amphipoda) in riverine salmonid diets.Archiv. fur Hydrobiologie 149, 87–107.
altby, L., 1992. The use of the physiological energetics of Gammarus pulexto assess toxicity: a study using artificial streams. Environ. Toxicol. Chem.11, 79–85.
altby, C., Naylor, C., 1990. Preliminary observations on the ecological rel-evance of the Gammarus ‘scope for growth’ assay: effect of zinc onreproduction. Funct. Ecol. 4, 393–397.
altby, L., Clayton, S.A., Wood, R.M., McLoughlin, N., 2002. Evaluation ofthe Gammarus pulex in situ feeding assay as a biomonitor of water qual-ity: robustness, responsiveness and relevance. Environ. Toxicol. Chem. 21,361–368.
antel, L.H., Farmer, L.L., 1983. Osmotic and ionic regulation. The biologyof Crustacea. In: Bliss, D.E., Mantel, L.H. (Eds.), Internal Anatomy andPhysiological Regulation, vol. 5. Academic Press, New York, pp. 53–161.
cCahon, C.P., Pascoe, D., 1988a. Use of Gammarus pulex (L.) in safety evalua-tion tests: culture and selection of a sensitive life stage. Ecotoxicol. Environ.Saf. 15, 245–252.
cCahon, C.P., Pascoe, D., 1988b. Cadmium toxicity to the freshwater amphi-pod Gammarus pulex (L.) during the moult cycle. Freshwater Biol. 19,197–203.
cCahon, C.P., Pascoe, D., 1988c. Increased sensitivity to cadmium of thefreshwater amphipod Gammarus pulex (L.) during the reproductive period.Aquat. Toxicol. 13, 183–194.
cGeer, J.C., Szebedinszky, C., McDonald, D.G., Wood, C.M., 2000a. Effectsof chronic sublethal exposure to waterborne Cu, Cd or Zn in rainbowtrout. 2. Tissue specific metal accumulation. Aquat. Toxicol. 50, 245–
256.cGeer, J.C., Szebedinszky, C., McDonald, D.G., Wood, C.M., 2000b. Effectsof chronic sublethal exposure to waterborne Cu, Cd or Zn in rainbow trout.1. Iono-regulatory disturbance and metabolic costs. Aquat. Toxicol. 50,231–243.
S
logy 86 (2008) 413–425
ebane, C., 2005a. Cadmium risks to freshwater life: predicted low-effect val-ues in ambient waters based upon laboratory and field studies. Metals inthe Environment: Aquatic Biological Perspectives. Scientific InvestigationReport. USGS (Reviewed draft, see on http://www.deq.state.id.us/water/datareports/surface water/monitoring/cadmium risks freshwater life.pdf).
ebane, C., 2005b. Cadmium risks to freshwater life: predicted low-effect val-ues in ambient waters based upon laboratory and field studies. In: Metalsin the Environment: Aquatic Biological Perspectives. SETAC 26th AnnualMeeting in North America, Baltimore, MD, USA.
ills, C.L., Shukla, D.H., Compton, G.J., 2006. Development of a new low costhigh sensitivity system for behavioural ecotoxicity testing. Aquat. Toxicol.77, 197–201.
oore, D.W., Farrar, J.D., 1996. Effect of growth on reproduction in thefreshwater amphipod, Hyalella azteca (Saussure). Hydrobiologia 328, 127–134.
iyogi, S., Wood, C.M., 2004. Kinetic analyses of waterborne Ca and Cd trans-port and their interactions in the gills of rainbowtrout (Oncorhynchus mykiss)and yellow perch (Perca.avescens), two species differing greatly in acutewaterborne Cd sensitivity. J. Comp. Physiol. 174B, 243–253.
unez-Nogueira, G., Rainbow, P.S., Smith, B.D., 2005. Assimilation efficiencyof zinc and cadmium in the decapod crustacean Penaeus indicus. J. Exp.Biol. 332, 75–83.
aquin, P.R., Gorsuch, J.W., Apte, S., Batley, G.E., Bowles, K.C., Campbell,P.G.C., Delos, C.G., Di Toro, D.M., Dwyer, R.L., Galvez, F., Gensemer,R.W., Goss, G.G., Hogstrand, C., Janssen, C.R., McGeer, J.C., Naddy, R.B.,Playle, R.C., Santore, R.C., Schneider, U., Stubblefield, W.A., Wood, C.M.,Wu, K.B., 2002. The biotic ligand model: a historical overview. Comp.Biochem. Physiol. 133C, 3–35.
equeux, A., 1995. Osmotic regulation in crustaceans. J. Crust. Biol. 15 (1),1–60.
layle, R.C., Dixon, D.G., Burnison, K., 1993a. Copper and cadmium binding tofish gills: modifications by dissolved organic carbon and synthetic ligands.Can. J. Fish. Aquat. Sci. 50, 2667–2677.
layle, R.C., Dixon, D.G., Burnison, K., 1993b. Copper and cadmium binding tofish gills: estimates of metal-gill stability constants and modelling of metalaccumulation. Can. J. Fish. Aquat. Sci. 50, 2678–2687.
layle, R.C., Dixon, D.G., Burnison, K., 1993c. Copper and cadmium bindingto fish gills: modification by dissolved organic carbon and synthetic ligands.Can. J. Fish. Aquat. Sci. 50, 2667–2677.
oulton, M., Pascoe, D., 1990. Disruption of precopula in Gammarus pulex(L.)—development of a behavioural bioassay for evaluating pollutant andparasite induced stress. Chemosphere 20, 403–415.
ratap, H.B., Fu, H., Lock, R.A.C., Wendelaar Bonga, S.E., 1989. Effect ofwaterborne and dietary cadmium on plasma ions on the teleost Oreochromismossambicus in relation to calcium levels. Arch. Environ. Contam. Toxicol.18, 568–575.
ainbow, P.S., 1997. Ecophysiology of trace metal uptake in crustaceans. Estuar.Coast. Shelf Sci. 44, 169–176.
iddell, D.J., Culp, J.M., Baird, D.J., 2005. Behavioral responses to sublethalcadmium exposure within an experimental aquatic food web. Environ. Tox-icol. Chem. 24, 431–441.
oast, S.D., Widdows, J., Jones, M.B., 2000. Mysids and trace metals: disruptionof swimming as a behavioural indicator of environmental contamination.Mar. Environ. Res. 50, 107–112.
oast, S.D., Widdows, J., Jones, M.B., 2001. Effects of salinity and chemicalspeciation on cadmium accumulation and toxicity to two mysid species.Environ. Toxicol. Chem. 20, 1078–1084.
odriguez Moreno, P.A., Schwarzbaum, P.J., Rodriguez, E.M., 1998. Effects ofcadmium on gill Na,K-ATPase of the estuarine crab Chasmagnathus gran-ulata (Decapoda, Grapsidae) during postmolt: in vivo and in vitro studies.Bull. Environ. Contam. Toxicol. 61, 629–636.
osas, C., Sanchez, A., Escohar, E., Soto, L., Bolongaro-Crevenna, A., 1992.Daily variations of oxygen consumption and glucose hemolymph level
related to morphophysiological and ecological adaptations of crustacea.Comp. Biochem. Physiol. 101A, 323–328.cott, G.R., Sloman, K.A., 2004. The effects of environmental pollutants oncomplex fish behaviour: interating behavioural and physiological indicatorsof toxicity. Aquat. Toxicol. 68, 369–392.

oxico
S
S
S
S
S
S
S
U
V
V
V
V
W
W
W
W
W
W
X
V. Felten et al. / Aquatic T
ilvestre, F., Trausch, G., Devos, P., 2005. Hyper-osmoregulation capacity of theChinese mitten crab (Eriocheir sinensis) exposed to cadmium; acclimationduring chronic exposure. Comp. Biochem. Physiol. 140C, 29–37.
oegianto, A., Charmantier-Daures, M., Trilles, J.-P., Charmantier, G., 1999.Impact of cadmium on the structure of gills and epipodites of the shrimpPenaeus japonicus (Crustacea: Decapoda). Aquat. Living Resour. 12, 57–70.
okolova, I.M., 2004. Cadmium effect on mitochondrial function are enhancedby elevated temperature in marine poikilotherm, Crassostrea virginicaGmelin (Bivalvia: Ostreidae). J. Exp. Biol. 207, 2639–2648.
picer, J.I., Weber, R.E., 1991. Respiratory impairment in crustacean and mol-luscs due to exposure to heavy metals. Comp. Biochem. Physiol. 100C,339–342.
picer, J.I., Morritt, D., Maltby, L., 1998. Effect of water-borne zinc on osmoreg-ulation in the freshwater amphipod Gammarus pulex (L.) from populationthat differ in their sensitivity to metal stress. Funct. Ecol. 12, 242–247.
tuhlbacher, A., Maltby, L., 1992. Cadmium resistance in Gammarus pulex (L.).Arch. Environ. Contam. Toxicol. 22, 319–324.
utcliffe, D.W., 1984. Quantitative aspects of oxygen uptake by Gammarus(Crustacea, Amphipoda): a critical review. Freshwater Biol. 14, 443–489.
SEPA, 2001. 2001 Update of Ambient Water Quality Criteria for Cadmium.United States Environmental Protection Agency. EPA-822-R-01-001.
erbost, P.M., Flik, G., Lock, R.A.C., Wendelaar Bonga, S.E., 1987. Cad-mium inhibition of Ca2+ uptake in rainbow trout gills. Am. J. Physiol. 253,R216–R221.
erbost, P.M., Van Rooiij, J., Flik, G., Lock, R.A.C., Wendelaar Bonga, S.E.,
1989. The movement of cadmium through fresh water trout branchial epithe-lium and its interference with calcium transport. J. Exp. Biol. 145, 185–197.indimian, E., Garric, J., Flammarion, P., Thybaud, E., Babut, M., 1999. Anindex of effluent aquatic toxicity designed by partial least squares regres-
Z
Z
logy 86 (2008) 413–425 425
sion using acute and chronic tests and expert judgements. Environ. Toxicol.Chem. 18, 2386–2391.
uori, K.-M., 1994. Rapid behavioural and morphological responses ofhydropsychid larvae (Trichoptera, Hydropsychidae) to sublethal cadmiumexposure. Environ. Pollut. 84, 291–299.
allace, W.G., Estephan, A., 2004. Differential susceptibility of horizontaland vertical swimming to cadmium exposure in gammaridean amphipod(Gammarus lawrencianus). Aquat. Toxicol. 69, 289–297.
icklund-Glynn, A., Norrgren, L., Mussener, A., 1994. Differences in uptakeof inorganic mercury and cadmium in the gills of the zebrafish, Brachydaniorerio. Aquat. Toxicol. 30, 13–26.
illiams, K.A., Green, W.J., Pascoe, D., 1985. Studies on the acute toxicity ofpollutants to freshwater macroinvertebrates. 1. Cadmium. Arch. Hydrobiol.102, 461–471.
right, D.A., 1980. Cadmium and calcium interactions in the freshwater amphi-pod Gammarus pulex. Freshwater Biol. 10, 123–133.
right, D.A., Fain, J.W., 1981. The effect of calcium on cadmium toxicity inthe freshwater amphipod, Gammarus pulex (L.). Arch. Environ. Contam.Toxicol. 10, 321–328.
u, J.M., Chen, H.-C., 2004. Effects of cadmium and zinc on oxygenconsumption, ammonium excretion, and osmoregulation of white shrimp(Liptopenaeus vannamei). Chemosphere 57, 1591–1598.
u, Q., 1995. The effects of exposure to zinc and cadmium separately and jointlyof the free amino acid pool of Gammarus pulex. Toxicol. Environ. Chem.50, 183–196.
augg, W.S., 1982. A simplified preparation for adenosine triphosphatase deter-mination in gill tissue. Can. J. Fish. Aquat. Sci. 39, 215–217.
auke, G.-P., Clason, B., Savinov, V.M., Savinova, T., 2003. Heavy metals ofinshore Benthic invertebrates from the Barents Sea. Sci. Total Environ. 306,99–110.




















