
Differential tolerance of two Gammarus pulex populations transplanted from different metallogenic...
9
Aquatic Toxicology 102 (2011) 95–103 Contents lists available at ScienceDirect Aquatic Toxicology journal homepage: www.elsevier.com/locate/aquatox Differential tolerance of two Gammarus pulex populations transplanted from different metallogenic regions to a polymetal gradient Farhan R. Khan ∗ , Jennifer R. Irving, Nicolas R. Bury, Christer Hogstrand Nutritional Sciences Division, King’s College London, Franklin-Wilkins Building, 150 Stamford Street, London SE1 9NH, United Kingdom article info Article history: Received 28 September 2010 Received in revised form 23 December 2010 Accepted 4 January 2011 Keywords: Metal bioaccumulation Sub-cellular distribution Detoxification Population differences abstract The River Hayle, Cornwall, UK exhibits pronounced Cu and Zn concentration gradients which were used to compare the metal handling abilities of two populations of Gammarus pulex (Crustacea: Amphipoda). One population was native to the Hayle region (Drym) and presumably has been historically impacted by elevated Cu and Zn levels, whilst naïve gammarids were collected from the River Cray, Kent, UK. Both populations were subject to a 32 day in situ exposure at four R. Hayle sites (Drym, Godolphin, Relubbus and St. Erth). Mortality (LT50), Cu and Zn accumulation and sub-cellular distribution, and oxidative stress (malondialdehyde production) increased with the expected Cu and Zn bioavailabilities at the four sites (i.e. Godolphin > Relubbus > St. Erth > Drym). The naïve population experienced greater metal induced effects in terms of Cu and Zn accumulation, oxidative stress responses and lower LT50s. Analysis of Cu and Zn sub-cellular distribution, however, revealed no significant differences in metal handling. In both populations each metal was localised predominantly to the sub-cellular fraction containing metal bound to metallothionein-like proteins (MTLP) or that holding both metal-rich granules (MRG) and exoskeleton, MTLP and MRG binding being indicative of metal detoxification. However, a greater capacity for detoxi- fied metal storage is not a mechanism implicated in the perceived tolerance of the historically impacted gammarids. Instead our results suggest that the historically impacted population was adapted for lower uptake of Cu and Zn leading to lower bioaccumulation, stress response and ultimately mortality. These results demonstrate not only the usefulness of the in situ methodology, but also that differences in pop- ulation exposure history can cause significant differences in metal responses during exposure at higher concentrations. © 2011 Elsevier B.V. All rights reserved. 1. Introduction Typically, the impact of toxic metal enrichment has been assessed through the collection of field sampled endogenous pop- ulations in which metal accumulation and/or biomarker responses are studied (Cain and Luoma, 1998). However, this approach makes comparisons between naturally exposed and naïve populations dif- ficult, as metal impacted areas and non-impacted aquatic habitats rarely have exactly the same water physicochemical properties, which affect local trace metal bioavailabilities. Another approach is to subject field collected organisms to laboratory based experi- ments at concentrations within an environmentally relevant range (Muyssen et al., 2002). Although this often reveals differences in Abbreviations: MRG + exo, metal rich granule and exoskeleton fraction; MTLP, metallothionein-like proteins; MDA, malondialdehyde. ∗ Corresponding author. Present address: Department of Zoology, Natural History Museum, Cromwell Road, London SW7 5BD, United Kingdom. Tel.: +44 0207 942 6688. E-mail address: [email protected] (F.R. Khan). metal tolerances between populations, it does not generally mimic the metal bioavailabilities or other conditions in nature. An alter- native method that is gaining wider acceptance is the use of caged animals subjected to in situ studies (e.g. Gower and Darlington, 1990; Baudrimont et al., 1999; De Coen et al., 2006). This allows for the actual in-stream impact of metal discharges to be assessed on species with known responses. In this present paper we investi- gated the accumulation and sub-cellular distribution of copper and zinc, as well as the oxidative stress response, in two populations of the amphipod crustacean Gammarus pulex exposed within a stream that exhibits a severe metal gradient. Total metal accumulation in individual organisms is dependent on the balance of uptake, through waterborne and dietary routes, and excretion and growth (Luoma and Rainbow, 2005). Yet high levels of total bioaccumulation do not necessarily result in toxicity, as this is dependent on that fraction of bioaccumulated metal that is in metabolically available form directly or indirectly disrupting metabolic processes (Luoma and Rainbow, 2008). Any potentially toxic rise in this metabolically available metal component can be prevented if accumulated metal is detoxified for storage in inert forms (i.e. biologically detoxified metal; Wallace et al., 2003) that do 0166-445X/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.aquatox.2011.01.001
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Differential tolerance of two Gammarus pulex populations transplanted from different metallogenic...

Dd
FN
a
ARR2A
KMSDP
1
auacfirwim(
m
MT
0d
Aquatic Toxicology 102 (2011) 95–103
Contents lists available at ScienceDirect
Aquatic Toxicology
journa l homepage: www.e lsev ier .com/ locate /aquatox
ifferential tolerance of two Gammarus pulex populations transplanted fromifferent metallogenic regions to a polymetal gradient
arhan R. Khan ∗, Jennifer R. Irving, Nicolas R. Bury, Christer Hogstrandutritional Sciences Division, King’s College London, Franklin-Wilkins Building, 150 Stamford Street, London SE1 9NH, United Kingdom
r t i c l e i n f o
rticle history:eceived 28 September 2010eceived in revised form3 December 2010ccepted 4 January 2011
eywords:etal bioaccumulation
ub-cellular distributionetoxificationopulation differences
a b s t r a c t
The River Hayle, Cornwall, UK exhibits pronounced Cu and Zn concentration gradients which were usedto compare the metal handling abilities of two populations of Gammarus pulex (Crustacea: Amphipoda).One population was native to the Hayle region (Drym) and presumably has been historically impactedby elevated Cu and Zn levels, whilst naïve gammarids were collected from the River Cray, Kent, UK. Bothpopulations were subject to a 32 day in situ exposure at four R. Hayle sites (Drym, Godolphin, Relubbusand St. Erth). Mortality (LT50), Cu and Zn accumulation and sub-cellular distribution, and oxidative stress(malondialdehyde production) increased with the expected Cu and Zn bioavailabilities at the four sites(i.e. Godolphin > Relubbus > St. Erth > Drym). The naïve population experienced greater metal inducedeffects in terms of Cu and Zn accumulation, oxidative stress responses and lower LT50s. Analysis of Cuand Zn sub-cellular distribution, however, revealed no significant differences in metal handling. In bothpopulations each metal was localised predominantly to the sub-cellular fraction containing metal boundto metallothionein-like proteins (MTLP) or that holding both metal-rich granules (MRG) and exoskeleton,
MTLP and MRG binding being indicative of metal detoxification. However, a greater capacity for detoxi-fied metal storage is not a mechanism implicated in the perceived tolerance of the historically impactedgammarids. Instead our results suggest that the historically impacted population was adapted for loweruptake of Cu and Zn leading to lower bioaccumulation, stress response and ultimately mortality. Theseresults demonstrate not only the usefulness of the in situ methodology, but also that differences in pop-ulation exposure history can cause significant differences in metal responses during exposure at higher concentrations.. Introduction
Typically, the impact of toxic metal enrichment has beenssessed through the collection of field sampled endogenous pop-lations in which metal accumulation and/or biomarker responsesre studied (Cain and Luoma, 1998). However, this approach makesomparisons between naturally exposed and naïve populations dif-cult, as metal impacted areas and non-impacted aquatic habitatsarely have exactly the same water physicochemical properties,
hich affect local trace metal bioavailabilities. Another approachs to subject field collected organisms to laboratory based experi-ents at concentrations within an environmentally relevant range
Muyssen et al., 2002). Although this often reveals differences in
Abbreviations: MRG + exo, metal rich granule and exoskeleton fraction; MTLP,etallothionein-like proteins; MDA, malondialdehyde.∗ Corresponding author. Present address: Department of Zoology, Natural Historyuseum, Cromwell Road, London SW7 5BD, United Kingdom.
el.: +44 0207 942 6688.E-mail address: [email protected] (F.R. Khan).
166-445X/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.aquatox.2011.01.001
© 2011 Elsevier B.V. All rights reserved.
metal tolerances between populations, it does not generally mimicthe metal bioavailabilities or other conditions in nature. An alter-native method that is gaining wider acceptance is the use of cagedanimals subjected to in situ studies (e.g. Gower and Darlington,1990; Baudrimont et al., 1999; De Coen et al., 2006). This allowsfor the actual in-stream impact of metal discharges to be assessedon species with known responses. In this present paper we investi-gated the accumulation and sub-cellular distribution of copper andzinc, as well as the oxidative stress response, in two populations ofthe amphipod crustacean Gammarus pulex exposed within a streamthat exhibits a severe metal gradient.
Total metal accumulation in individual organisms is dependenton the balance of uptake, through waterborne and dietary routes,and excretion and growth (Luoma and Rainbow, 2005). Yet highlevels of total bioaccumulation do not necessarily result in toxicity,as this is dependent on that fraction of bioaccumulated metal that
is in metabolically available form directly or indirectly disruptingmetabolic processes (Luoma and Rainbow, 2008). Any potentiallytoxic rise in this metabolically available metal component can beprevented if accumulated metal is detoxified for storage in inertforms (i.e. biologically detoxified metal; Wallace et al., 2003) that do
9 Toxico
nmimemnott
ogcutp2tEgtectpims
wcebMaaMHostanhiFtaswadTdtw
2
2
c(
6 F.R. Khan et al. / Aquatic
ot allow more labile metal to cause metabolic damage. Commonacro-invertebrate methods for detoxifying accumulated metal
nclude sequestration by glutathione and intracellular proteins (e.g.etallothioneins), or binding within metal rich granules (Vijver
t al., 2004; Luoma and Rainbow, 2008). Cu and Zn, as essentialicronutrients, are subject to tight homeostatic control and inter-
al concentrations can be actively regulated by increased turnoverr decreased uptake (Bossuyt and Janssen, 2005). Thus, the needo detoxify these elements may be negated if organisms can limitheir initial accumulation.
The development of strategies to avoid metal induced toxicityften occurs out of necessity through exposure to elevated back-round concentrations. Consequently, prior natural exposure mayonfer a degree of tolerance that is absent in populations fromncontaminated areas. Examples of naturally impacted popula-ions being more tolerant than naive populations include particularopulations of the polychaete Nereis diversicolor (Rainbow et al.,006), the midge larva Chironomus riparius (Postma et al., 1996) andhe collembolan springtail Orchesella cincta (Roelofs et al., 2007).ach tolerant population used a different method of surviving atreater concentrations; N. diversicular distributed greater propor-ion of Ag to insoluble metal rich granules, C. riparius increasedxcretion of cadmium, and previously exposed populations of O.incta had increased metallothionein gene expression in responseo cadmium exposure. Tolerance occurs via genetic adaptation orhysiological acclimation. Acclimation is a response to a metal dur-
ng the lifetime of an organism, whereas adaptation evolves overany generations being exposed to the same stressor leading to
election for tolerant genotypes (Morgan et al., 2007).In the present study G. pulex were collected from two regions
ith differing histories of natural Cu and Zn loads. G. pulex werehosen for this study as they are, as amphipods are in gen-ral, net accumulators of trace metals which makes them reliableiomonitors of environmental metal bioavailability (Rainbow andoore, 1986; Rainbow, 2002; Fialkowski et al., 2003). Addition-
lly, G. pulex are found in high population densities in streamsnd rivers throughout the UK and Europe (Graca et al., 1994;acNeil et al., 1997), including the upper reaches of the Riverayle at Drym in Cornwall, South West England from whereur historically impacted population was collected. Caged sub-ets from our two collected populations were then transplantedo four sites along River Hayle, which exhibits pronounced coppernd zinc concentration gradients owing to the legacy of the Cor-ish mining industry. The gradient in toxic metal concentrationsas been previously shown as the cause of spatial disturbances
n both macro-invertebrate and piscine populations (River Hayleisheries Survey, 1970; Gower et al., 1994). Following transplan-ation, gammarids were collected during the 32 day exposure andnalysis of mortality rate, copper and zinc bioaccumulation andub-cellular distribution was undertaken. Additionally, cytotoxityas measured in the form of malondialdehyde (MDA) production,by-product of lipid peroxidation and one of the widely used
eterminants of metal-induced oxidative stress (Liu et al., 1997).hrough this range of measures, our aim is to consider whetherifferences in the long-term exposure history of the two popula-ions have an effect during subsequent exposure in metal impactedaters.
. Methods
.1. Study area and sampling procedures
The catchment of the River Hayle was an important source ofopper and tin during the seventeenth and eighteenth centuriesBrown, 1977). Mining ceased in the early part of the twentieth
logy 102 (2011) 95–103
century, but the river continues to receive drainage water rich inCu and Zn, principally from four sources at Binner Down, GreatWork, Wheal Godolphin and Penberthy Cross (Fig. 1; Brown, 1977).Based on the influx of these mine discharges, the freshwater RiverHayle can be broadly divided into three distinctive sections; (1)upstream of the Binner Down Adit includes Drym (50◦09′23.07′′N05◦19′54.90′′W) where our historically impacted G. pulex popu-lation is found. G. pulex are the dominant macro-invertebratesat Drym and the site also supports local populations of browntrout (Salmo trutta L.) and eel (Anguilla anguilla L.); (2) Down-stream of the Great Work and Binner Down Adits is Godolphin(50◦08′42.70′′N 05◦21′42.97′′W), the most Cu and Zn enriched areaalong the River Hayle. The river is fishless for approximately 2miles and macro-invertebrate densities are vanishingly low, butplant life remains in the form of submerged macrophytes (RiverHayle Fisheries Survey, 1970; Chemical Survey of R. Hayle, 1983);(3) From the edge of Tregembo Marsh to where the River Haylemeets estuarine water preceding St. Ives Bay is the third river sec-tion. Tregembo Marsh has been shown to remediate water qualitywith respect to trace metal contamination which results in thesites of Relubbus (50◦08′23.17′′N 05◦24′32.93′′W), following themarsh, and St. Erth (50◦09′57.60′′N 05◦26′05.37′′W) having lowerCu and Zn concentrations than Godolphin. Macro-invertebrateand fish densities increase following remediation and Relubbussustains populations of caseless caddisfly larvae, Hydropsyche pel-licidula (Trichoptera), and dragonfly larvae, Cordulegaster boltoni(Odonata). The trichopterans, in particular, exhibit a high degree ofmetal tolerance (Gower et al., 1994). Table 1 describes the dramaticincrease in dissolved Cu and Zn concentrations caused by the minedrainage waters between Drym and Godolphin, and the subsequentdecreases between Godolphin and Relubbus, and St. Erth.
Hyrdochemical parameters (total hardness, pH and conductivity(as a representation of ionic strength)) that are known to effect tracemetal bioavailability were measured throughout the 32 d study ateach transplantation site and did not vary significantly. Therefore,we assume that exposure conditions at each site were reasonablyconsistent as were the differences between sites. Total hardness,pH, conductivity and metal concentrations all increase with theinflux of mine water between Drym and Godolphin, but thereafterare subject only to slight further increases at Relubbus and St. Erth(Table 1). Dissolved organic carbon concentrations throughout thecatchment area of the River Hayle are <5 mg C L−1 and decreaseas mine adit discharges enter the river (Chemical, 1983; Fig. 1).The site with lowest metal concentrations (Drym) has the high-est DOC concentration, after which metal and DOC concentrationsfollow the same pattern i.e. Godolphin > Relubbus > St Erth. Accord-ingly, we expect bioavailable dissolved Cu and Zn concentrationsat each site to be relative to the total dissolved metal concentra-tions. Our measurements were taken during November 2006 whenthe in situ study was conducted and although metal concentrationsand hydrochemical parameters are subject to seasonal variation therelative differences between the sites can be expected to be largelyconstant.
At each transplantation site on the River Hayle (and at the RiverCray) Cu and Zn concentrations were determined in water and sed-iments. Dissolved metal concentrations in filtered (0.45 �m filter,Scleicher & Schuell Bioscience Inc., New Hampshire, USA) acidified(0.1% ultrapure HNO3, Merck KGaA, Darmstadt, Germany) watersamples were measured by graphite furnace atomic absorptionspectrometry (Perkin Elmer 4100ZL with Zeeman background cor-rection, Perkin Elmer, UK). One gram of dried (70 ◦C overnight)
sieved (125 �m mesh) sediment was refluxed in 69% HNO3 for 6 h.Cu and Zn sediment concentrations were measured by acetyleneflame atomic absorption spectrometry (Unicam 929 AAS, Unicam,UK). For all metal analysis, including G. pulex tissue and fractiondigests, TORT-2 lobster hepatopancreas (National Research Council
F.R. Khan et al. / Aquatic Toxicology 102 (2011) 95–103 97
Table 1Hyrdochemical parameters at each River Hayle transplantation site and at the River Cray G. pulex collection site.
Site Dissolved water Sediment Total hardness pH Conductivity
Cu Zn Cu Zn
R. Cray 0.8 ± 0.1 3.6 ± 0.8 83.6 ± 5.7 300.6 ± 13.6Drym 1.3 ± 0.2 7.2 ± 1.3 133.4 ± 38.6 390.4 ± 95.0 72.1 ± 3.4 6.58 ± 0.02 47.4 ± 0.1Godolphin 42.7 ± 5.1 881.5 ± 72.0 1722.8 ± 325.7 883.5 ± 216.8 95.6 ± 5.2 6.63 ± 0.01 66.2 ± 0.1Relubbus 10.7 ± 0.9 664.5 ± 43.9 1279.6 ± 17.6 479.2 ± 68.0 96.6 ± 2.1 6.68 ± 0.03 69.6 ± 0.1St. Erth 7.9 ± 1.3 500.5 ± 32.7 1561.9 ± 169.8 709.8 ± 271.3 109.9 ± 4.9 6.71 ± 0.02 77.3 ± 0.1
M ed waa L−1 Cs
CdsrcpspaM
fEawGaeCaaCtgethmmlpwo
FHaa1
ean values ± standard deviation are shown. Cu and Zn concentrations in dissolvdits between Drym and Godolphin. Hydrochemical parameters total hardness (mgtrength)) are known to influence metal bioavailabilities.
anada) and SRM 1566b oyster tissue (National Institute of Stan-ards & Technology, U.S. Department of Commerce) were used astandard reference materials. Recoveries of both Cu and Zn for botheference materials were consistently within ±10% of the certifiedoncentrations. Additionally, total hardness (mg L−1 CaCO3) andH were measured in filtered (0.45 �m filter) unacidified wateramples by EDTA titration and pH meter (Mettler Toleda MP220H meter), respectively. Ionic strength of the water was measureds conductivity with an in situ probe (WTW LF320 Conductivityeter).Individuals from the naïve population of G. pulex were collected
rom the River Cray (51◦23′09.47′′N 00◦05′59.02′′W), in Kent, Southast England. The River Cray is an urban watercourse with abundantquatic vegetation (mainly Cladaphora spp. and Sparganium spp.)hich supports a wide range of macro-invertebrates. In addition to. pulex, Asellus aquaticus (isopoda), Lymnaea stagnalis (gastropoda)nd Baetis vernus (ephemeroptera) are all present. G. pulex, how-ver, is the most common species (Dawkins and Donoghue, 1992).oncentrations of dissolved Cu and Zn at the R. Cray were 0.81nd 3.62 �g L−1, respectively which are approximately half thoset Drym (Table 1). The Cu and Zn concentrations at Drym and R.ray are both in the range of background concentrations withinhe EU (Comber et al., 2008) and therefore during the lifetime ofammarids used in this study, none would have experienced metalxposure of any significant magnitude. However, with the popula-ion at Drym being located within a highly metallogenic region it isypothesised that at least some of their ancestors were selected foretal tolerance over the last few centuries. However, as the majorining work in the area ceased more than 100 years ago and no eco-
ogical or water chemistry data were recorded during the miningeriod this is ultimately difficult to verify. Our aim was to determinehether this possible scenario could influence the metal responses
f the current population in comparison to a metal naïve popula-
ig. 1. Location of the four transplantation sites and mine adits along the Riverayle in Cornwall. Transplantation sites (Drym, Godolphin, Relubbus and St. Erth)re marked with black circles and the influx of metal rich waters from major minedits (large arrow) and minor adits (small arrow) are shown (re-drawn from Brown,977 and Chemical Survey of R. Hayle, 1983).
ter (�g L−1, n = 5) and in sediment (�g g−1, n = 8) show the influence of the mineaCO3, n = 6), pH (n = 6) and conductivity (�S cm−1, n = 6 (as a representation of ionic
tion during the subsequent in situ exposure at sites with higher Cuand Zn concentrations.
2.2. In situ transplantation
G. pulex were collected from the R. Cray and R. Hayle at Drym bykick net sampling.
R. Cray G. pulex were acclimated to US E.P.A moderately hardsynthetic water (80–100 mg L CaCO3
−1) for one week prior to trans-plantation and were transported to Cornwall in this moderatelyhard synthetic water within a cooler. After collection of Drym G.pulex, a sub-sample of both populations were rinsed in deionisedwater, blotted dry and frozen on dry ice to provide Day 0 samples.
For the in situ study, approximately 2 g of adult G. pulex (80–120individual gammarids) from either Drym or R. Cray were placed in5 exposure chambers at each transplantation site (Drym, Godol-phin, Relubbus and St. Erth). Exposure chambers were constructedfrom plastic tubes (20 cm long, inner diameter 2.8 cm) and cov-ered at both ends with 1 mm nylon mesh. All tubes and mesh weresoaked in 1% HNO3 and rinsed thoroughly with deionised water.Leaf litter from R. Cray was placed in each chamber as a source offood, micro-habitat and to discourage cannibalism (Maltby et al.,2002). Leaf litter from the River Cray was used in preference tothat of R. Hayle, as the latter may have introduced dietary metal.Other sources of food received would be from the natural environ-ment at each R. Hayle site. Thus gammarids were primarily exposedto waterborne Cu and Zn. For transplantation, exposure chamberswere attached to bricks as ballast. One chamber per population, persite was collected on Days 1, 2, 4, 16 and 32 post-transplantation.On collection gammarids were rinsed out of the exposure cham-ber with deionised water and blotted dry. Live and dead animalswere separated and weighed. From the known weight of the G.pulex in each exposure chamber at t = 0, an estimated percentagemortality was calculated. Though factors such as decomposition,growth and reproduction can influence this estimate of mortalitybased on biomass, we attempted to control for this by omittingcopulating pairs at t = 0 and by determining the recovery of tissuefrom each exposure which was <90% (range 92.6–98.3%, n = 32).Live gammarids were frozen on dry ice for transport back to thelaboratory where they were promptly stored at −80 ◦C.
2.3. Metal accumulation
Total Cu and Zn accumulated concentrations were measured insurviving gammarids by acetylene flame atomic absorption spec-trometry (Unicam 929 AAS). G. pulex samples of 100 mg fresh
weight (fw) were dried to constant weight overnight at 70 ◦C,digested in 1:5 (w/v) ratio with 60% ultrapure HNO3 and made upto 5 mL in MilliQ water. Where necessary dilutions were made withMilliQ water. Metal accumulation is expressed as �g metal g−1 G.pulex dry weight (dw).
9 Toxico
2
aWpUHwaptppthspN1etfi
pwubGttaad
2
aaaots(
uappcawfc
2
tsETst
8 F.R. Khan et al. / Aquatic
.4. Metal distribution
To determine the sub-cellular distribution of accumulated Cund Zn within G. pulex the sub-cellular fractionation protocol ofallace et al. (2003) was followed. Approximately 200 mg (fw) G.
ulex (∼10 individuals) were homogenised (TissueLyser, Qiagen,K) in a 1:4 (w/v) ratio in ice cold 10 mM HCL–TRIS buffer (pH 7.6).omogenates were centrifuged at 800 × g for 15 min at 4 ◦C fromhich 150 �L of the supernatant was removed for oxidative stress
nalysis. This aliquot was stored at −80 ◦C until analysis could takelace. The remaining supernatant was subjected to the two fur-her ultra-centrifugation steps. Firstly, 100,000 × g centrifugationroduced a combined mitochondrial, lysosomal and microsomalellet termed the organelle fraction, and secondly after heat dena-uring (80 ◦C for 10 min) 50,000 × g centrifugation resulted in aeat denatured protein (HDP) pellet and heat stable protein (HSP)upernatant containing metallothionein-like proteins (MTLP). Theellet obtained from the initial homogenate centrifugation wasaOH digested and heated (70 ◦C for 1 h) before being centrifuged at0,000 × g for 10 min producing a supernatant containing digestedxoskeleton, nuclei and cell debris (debris fraction) and pellet con-aining metal rich granules and undigested exoskeleton (MRG + exoraction). The centrifuges used and the specific details of each stepn this protocol can be found in Khan et al. (2010).
The five fractions were digested in a 1:5 w/v ratio with 60% Ultra-ure HNO3 at 70 ◦C overnight and then made up to 3 mL with MilliQater. For the digestion of the liquid fractions 100 �L of sample wassed. The Cu and Zn concentration in each fraction was determinedy acetylene flame AAS. After determining the moisture content of. pulex (80 ± 2%, n = 20), Cu and Zn concentrations were related
o the dry weight of each initial 200 mg pool. Thus metal concen-rations are expressed as �g g−1 G. pulex (dw) which takes intoccount the size of fraction in relation to the size of the organismnd allows the relative distribution of whole body burdens to beetermined.
.5. Oxidative stress analysis
Oxidative stress was measured as the production of malondi-ldehyde (MDA) by thiobarbituric acid reactive substances (TBARS)ssay (ZeptoMetric Corporation, New York, USA) in the 150 �Lliquot of the first centrifuge supernatant. MDA is a by-productf lipid peroxidation which reacts with thiobarbituric acid (TBA)o form a 1 MDA: 2 TBA complex which, in our study, was mea-ured fluorometrically owing to the greater sensitivity of detection0–4 nmol/mL).
From each 150 �L aliquot three replicate 40 �L of samples weresed for the TBARs reaction. The TBARs reaction was performeds described in Khan et al. (2010) according to the manufacturer’srotocol. A volume of 200 �L of each assay product was addeder well to a black 96-well plate. Plates were read fluorometri-ally on a Biotek Synergy HT plate reader with excitation at 528 nmnd emission at 550 nm. The concentration of MDA in each sampleas normalised to the protein concentration, determined by Brad-
ord’s Assay (Bradford, 1976; protein assay kit, Biorad) so that MDAoncentration is expressed as nmol MDA mg−1 protein.
.6. Statistical analysis
The estimated percentage mortality in each chamber was usedo calculate an LT50 (median lethal time for 50% mortality) for each
ite and both populations using Probit regression (Comprehensivenvironment Toxicity Information Software (CETIS) version 1.6.1,idepool Scientific Software). The LT50s of both populations at eachite were compared using an LC50 ratio test in which the ratio ofhe two log LT50s (�) is divided by the ratio of their respective loglogy 102 (2011) 95–103
standard errors (se) to produce a z value, thus z = log(�)/se[log(�)](Wheeler et al., 2006).
In the statistical analysis of metal accumulation, oxidative stressand sub-cellular fractionation, copper and zinc were treated sep-arately. Our reasoning for this is that they have been shownto enter organisms via different trans-membrane uptake routes(transporter proteins and major ion channels) and have differingmechanisms of toxicity. Cu+ can be taken up by Na+ channels andis a redox active metal that participates in Fenton-like reactionscausing direct oxidative stress (Grosell and Wood, 2002; Kendricket al., 1992), whereas Zn2+ can be transported via Ca2+ channelsand disruption of calcium homeostasis is considered to be a likelymechanism of toxicity (Hogstrand et al., 1996).
In the present study, the following definitions were used: Basalmetal concentration is the concentration of Cu or Zn in G. pulex(whole body or sub-cellular fraction) collected from Drym or R.Cray at Day 0. Total metal concentration is the concentration of Cuor Zn within the organism (whole body or sub-cellular fraction).This incorporates the basal metal concentration. Metal accumula-tion (net accumulation) is the concentration of Cu or Zn that wasincorporated by the organism (whole body or fraction) over thelength of exposure. It is the total metal concentration minus thebasal metal concentration.
Overall analysis of metal accumulation over the exposure periodwas determined by three-way ANOVA with post-hoc Tukey test.The three variables were population origin, Drym or R. Cray, whichdetermined whether Cu and Zn exposure history influenced accu-mulation in subsequent exposure; site (the influence of the Hayle’smetal gradient) and the exposure duration (time). The high concen-trations of accumulated metal at Day 16 caused a skew in the dataset and all other time and sites were found not to be significantlydifferent from each other. A two-way ANOVA with post-hoc Tukeytest was then performed with population and site as variables. Inthis, each time point was treated individually which allowed sig-nificant differences between sites and populations to be identifiedon Days 1, 2 and 4. The changes in accumulation within a singlepopulation were determined by a two-way ANOVA with time andsite as variables. Oxidative stress levels were analysed to determinethe influences of time, exposure site and population origin by thesame methods used for metal accumulation.
Enrichment of metal in each sub-cellular fraction was analysedusing the methods of Bonneris et al. (2005). Both total metal con-centration in each fraction and total metal body burden were Log10transformed to allow for analysis of linear relationships by Pear-son’s correlation. For correlations the total metal concentration wasused in preference to accumulated concentrations. Furthermore,sub-cellular fractions that were enriched during the in situ expo-sure were analysed by two-way ANOVA with post hoc Tukey testusing accumulated metal concentrations in the fraction.
3. Results
3.1. Mortality
Mortality in both populations followed the expected pattern ofincreasing with Cu and Zn dissolved concentration along the R.Hayle gradient. At the most metal impacted site (Godolphin) 100%mortality occurred within 4 days and there was no difference in theestimated LT50s between populations (z = 0.06, p = 0.95, Fig. 2). AtRelubbus, Cray and Drym gammarids suffered 50% mortality at 4.9 d
(95% CI: 4.3–5.5 d) and 6.7 d (95% CI: 5.9–7.7 d) respectively, whichwas significantly different (z = 6.06, p < 0.01). Again at St. Erth thenaïve R. Cray gammarids had a significantly lower LT50 (7.8 d, 955CI: 6.9–8.8 d) compared to the historically impacted Drym popula-tion (12.1 d, 95% CI: 10.4–13.5 d; z = 11.11, p < 0.01). No mortalities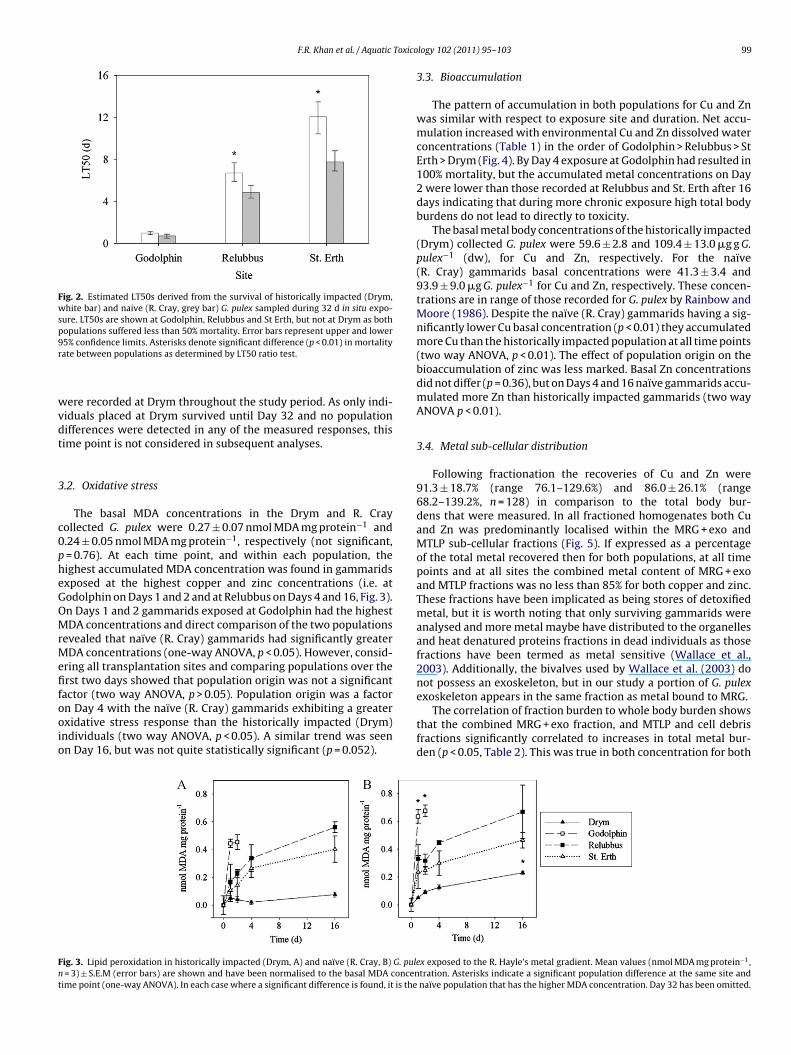
F.R. Khan et al. / Aquatic Toxico
Fig. 2. Estimated LT50s derived from the survival of historically impacted (Drym,white bar) and naive (R. Cray, grey bar) G. pulex sampled during 32 d in situ expo-sure. LT50s are shown at Godolphin, Relubbus and St Erth, but not at Drym as bothpopulations suffered less than 50% mortality. Error bars represent upper and lower9r
wvdt
3
c0pheGOMrMefifooio
exoskeleton appears in the same fraction as metal bound to MRG.
Fnt
5% confidence limits. Asterisks denote significant difference (p < 0.01) in mortalityate between populations as determined by LT50 ratio test.
ere recorded at Drym throughout the study period. As only indi-iduals placed at Drym survived until Day 32 and no populationifferences were detected in any of the measured responses, thisime point is not considered in subsequent analyses.
.2. Oxidative stress
The basal MDA concentrations in the Drym and R. Crayollected G. pulex were 0.27 ± 0.07 nmol MDA mg protein−1 and.24 ± 0.05 nmol MDA mg protein−1, respectively (not significant,= 0.76). At each time point, and within each population, theighest accumulated MDA concentration was found in gammaridsxposed at the highest copper and zinc concentrations (i.e. atodolphin on Days 1 and 2 and at Relubbus on Days 4 and 16, Fig. 3).n Days 1 and 2 gammarids exposed at Godolphin had the highestDA concentrations and direct comparison of the two populations
evealed that naïve (R. Cray) gammarids had significantly greaterDA concentrations (one-way ANOVA, p < 0.05). However, consid-
ring all transplantation sites and comparing populations over therst two days showed that population origin was not a significant
actor (two way ANOVA, p > 0.05). Population origin was a factorn Day 4 with the naïve (R. Cray) gammarids exhibiting a greater
xidative stress response than the historically impacted (Drym)ndividuals (two way ANOVA, p < 0.05). A similar trend was seenn Day 16, but was not quite statistically significant (p = 0.052).ig. 3. Lipid peroxidation in historically impacted (Drym, A) and naïve (R. Cray, B) G. pule= 3) ± S.E.M (error bars) are shown and have been normalised to the basal MDA concen
ime point (one-way ANOVA). In each case where a significant difference is found, it is the
logy 102 (2011) 95–103 99
3.3. Bioaccumulation
The pattern of accumulation in both populations for Cu and Znwas similar with respect to exposure site and duration. Net accu-mulation increased with environmental Cu and Zn dissolved waterconcentrations (Table 1) in the order of Godolphin > Relubbus > StErth > Drym (Fig. 4). By Day 4 exposure at Godolphin had resulted in100% mortality, but the accumulated metal concentrations on Day2 were lower than those recorded at Relubbus and St. Erth after 16days indicating that during more chronic exposure high total bodyburdens do not lead to directly to toxicity.
The basal metal body concentrations of the historically impacted(Drym) collected G. pulex were 59.6 ± 2.8 and 109.4 ± 13.0 �g g G.pulex−1 (dw), for Cu and Zn, respectively. For the naïve(R. Cray) gammarids basal concentrations were 41.3 ± 3.4 and93.9 ± 9.0 �g G. pulex−1 for Cu and Zn, respectively. These concen-trations are in range of those recorded for G. pulex by Rainbow andMoore (1986). Despite the naïve (R. Cray) gammarids having a sig-nificantly lower Cu basal concentration (p < 0.01) they accumulatedmore Cu than the historically impacted population at all time points(two way ANOVA, p < 0.01). The effect of population origin on thebioaccumulation of zinc was less marked. Basal Zn concentrationsdid not differ (p = 0.36), but on Days 4 and 16 naïve gammarids accu-mulated more Zn than historically impacted gammarids (two wayANOVA p < 0.01).
3.4. Metal sub-cellular distribution
Following fractionation the recoveries of Cu and Zn were91.3 ± 18.7% (range 76.1–129.6%) and 86.0 ± 26.1% (range68.2–139.2%, n = 128) in comparison to the total body bur-dens that were measured. In all fractioned homogenates both Cuand Zn was predominantly localised within the MRG + exo andMTLP sub-cellular fractions (Fig. 5). If expressed as a percentageof the total metal recovered then for both populations, at all timepoints and at all sites the combined metal content of MRG + exoand MTLP fractions was no less than 85% for both copper and zinc.These fractions have been implicated as being stores of detoxifiedmetal, but it is worth noting that only surviving gammarids wereanalysed and more metal maybe have distributed to the organellesand heat denatured proteins fractions in dead individuals as thosefractions have been termed as metal sensitive (Wallace et al.,2003). Additionally, the bivalves used by Wallace et al. (2003) donot possess an exoskeleton, but in our study a portion of G. pulex
The correlation of fraction burden to whole body burden showsthat the combined MRG + exo fraction, and MTLP and cell debrisfractions significantly correlated to increases in total metal bur-den (p < 0.05, Table 2). This was true in both concentration for both
x exposed to the R. Hayle’s metal gradient. Mean values (nmol MDA mg protein−1,tration. Asterisks indicate a significant population difference at the same site andnaïve population that has the higher MDA concentration. Day 32 has been omitted.

100 F.R. Khan et al. / Aquatic Toxicology 102 (2011) 95–103
Fig. 4. Accumulation of copper (A and B) and zinc (C and D) in historically impacted (Drym collected, A and C) and naïve (R. Cray collected, B and D) G. pulex exposed to fours valuen thereft cally sd
Cois
t
FeIl
ites along the R. Hayle. Data points show mean accumulated metal concentrationsormalised to the body burdens of copper and zinc on Day 0 (‘basal metal burden’),he metal burden at the site and time point. Day 32 has been omitted. The statistiuration were determined by three-way ANOVA and are discussed in Section 3.3.
u and Zn and in both populations. Metal concentrations in therganelles and heat denatured proteins fractions did not vary with
ncreasing total metal body burden, suggesting that these were notub-cellular compartments in which metals were accumulated.The change in distribution of Cu and Zn with particular referenceo the two most enriched fractions (MRG + exo and MTLP fractions)
ig. 5. Accumulated copper (A) and zinc (B) sub-cellular distribution in historically impaach site along the R. Hayle gradient at each time point. Each bar represents the accumulaten each bar the accumulated metal concentrations in each sub-cellular fraction has beenocalised within the MRG + exo and MTLP fractions. Please note the different scales on the
s on logarithmic scale (n = 5–6 pooled samples) ± S.E.M (error bars). Each value wasore negative values arise when the basal metal burden measured on Day 0 exceedsignificant differences relating to population origin, site of exposure and exposure
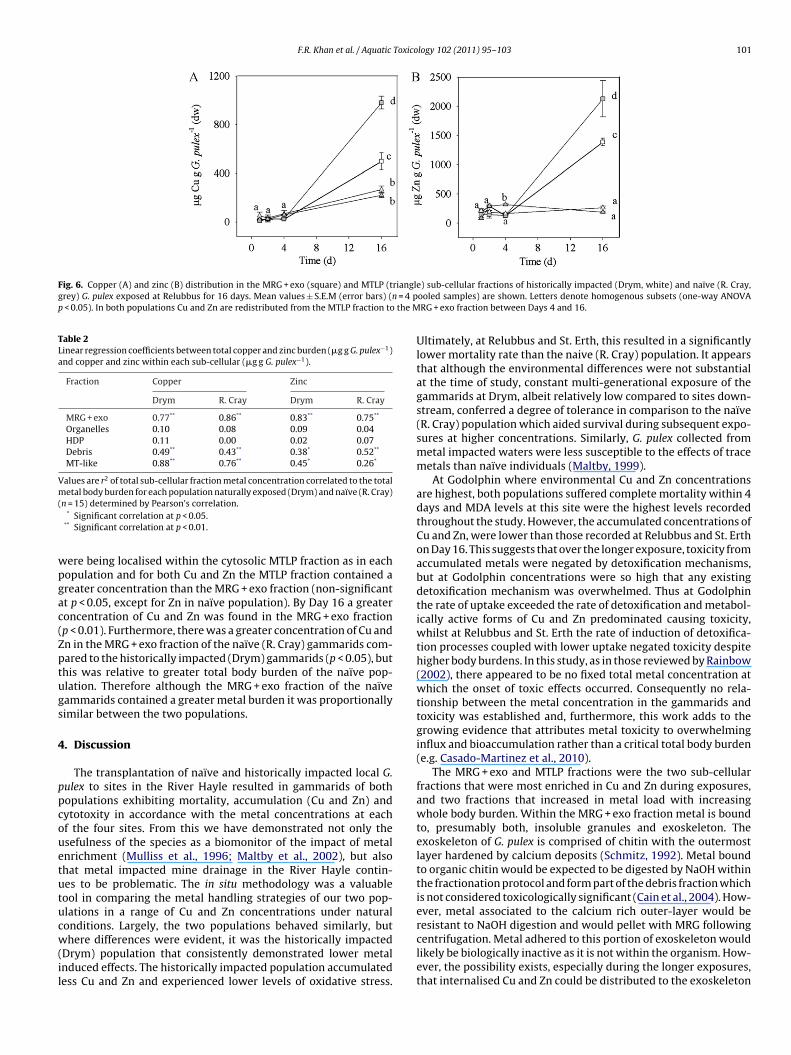
and the differences between the two populations is best illustratedby those gammarids that were exposed at Relubbus (i.e. 16 d expo-
sure at relatively high concentrations, Fig. 6). On Days 1 and 2 nosignificant differences in the concentrations of accumulated Cu orZn were found between MRG + exo and MTLP fractions or betweenthe two populations. On Day 4 a trend emerged to show that metalscted (Drym, non-striped bars) and naïve (R. Cray, striped bars) G. pulex exposed atd metal from G. pulex exposed at that site from that duration (n = 4 pooled samples).normalised to the basal (Day 0) metal burden. Copper and zinc are predominantlyy-axis. Day 32 has been omitted.
F.R. Khan et al. / Aquatic Toxicology 102 (2011) 95–103 101
Fig. 6. Copper (A) and zinc (B) distribution in the MRG + exo (square) and MTLP (trianglgrey) G. pulex exposed at Relubbus for 16 days. Mean values ± S.E.M (error bars) (n = 4 pp < 0.05). In both populations Cu and Zn are redistributed from the MTLP fraction to the M
Table 2Linear regression coefficients between total copper and zinc burden (�g g G. pulex−1)and copper and zinc within each sub-cellular (�g g G. pulex−1).
Fraction Copper Zinc
Drym R. Cray Drym R. Cray
MRG + exo 0.77** 0.86** 0.83** 0.75**
Organelles 0.10 0.08 0.09 0.04HDP 0.11 0.00 0.02 0.07Debris 0.49** 0.43** 0.38* 0.52**
MT-like 0.88** 0.76** 0.45* 0.26*
Values are r2 of total sub-cellular fraction metal concentration correlated to the totalm(
wpgac(Zptugs
4
ppcouetutucw(il
etal body burden for each population naturally exposed (Drym) and naïve (R. Cray)n = 15) determined by Pearson’s correlation.
* Significant correlation at p < 0.05.** Significant correlation at p < 0.01.
ere being localised within the cytosolic MTLP fraction as in eachopulation and for both Cu and Zn the MTLP fraction contained areater concentration than the MRG + exo fraction (non-significantt p < 0.05, except for Zn in naïve population). By Day 16 a greateroncentration of Cu and Zn was found in the MRG + exo fractionp < 0.01). Furthermore, there was a greater concentration of Cu andn in the MRG + exo fraction of the naïve (R. Cray) gammarids com-ared to the historically impacted (Drym) gammarids (p < 0.05), buthis was relative to greater total body burden of the naïve pop-lation. Therefore although the MRG + exo fraction of the naïveammarids contained a greater metal burden it was proportionallyimilar between the two populations.
. Discussion
The transplantation of naïve and historically impacted local G.ulex to sites in the River Hayle resulted in gammarids of bothopulations exhibiting mortality, accumulation (Cu and Zn) andytotoxity in accordance with the metal concentrations at eachf the four sites. From this we have demonstrated not only thesefulness of the species as a biomonitor of the impact of metalnrichment (Mulliss et al., 1996; Maltby et al., 2002), but alsohat metal impacted mine drainage in the River Hayle contin-es to be problematic. The in situ methodology was a valuableool in comparing the metal handling strategies of our two pop-lations in a range of Cu and Zn concentrations under natural
onditions. Largely, the two populations behaved similarly, buthere differences were evident, it was the historically impactedDrym) population that consistently demonstrated lower metalnduced effects. The historically impacted population accumulatedess Cu and Zn and experienced lower levels of oxidative stress.
e) sub-cellular fractions of historically impacted (Drym, white) and naïve (R. Cray,ooled samples) are shown. Letters denote homogenous subsets (one-way ANOVARG + exo fraction between Days 4 and 16.
Ultimately, at Relubbus and St. Erth, this resulted in a significantlylower mortality rate than the naive (R. Cray) population. It appearsthat although the environmental differences were not substantialat the time of study, constant multi-generational exposure of thegammarids at Drym, albeit relatively low compared to sites down-stream, conferred a degree of tolerance in comparison to the naïve(R. Cray) population which aided survival during subsequent expo-sures at higher concentrations. Similarly, G. pulex collected frommetal impacted waters were less susceptible to the effects of tracemetals than naïve individuals (Maltby, 1999).
At Godolphin where environmental Cu and Zn concentrationsare highest, both populations suffered complete mortality within 4days and MDA levels at this site were the highest levels recordedthroughout the study. However, the accumulated concentrations ofCu and Zn, were lower than those recorded at Relubbus and St. Erthon Day 16. This suggests that over the longer exposure, toxicity fromaccumulated metals were negated by detoxification mechanisms,but at Godolphin concentrations were so high that any existingdetoxification mechanism was overwhelmed. Thus at Godolphinthe rate of uptake exceeded the rate of detoxification and metabol-ically active forms of Cu and Zn predominated causing toxicity,whilst at Relubbus and St. Erth the rate of induction of detoxifica-tion processes coupled with lower uptake negated toxicity despitehigher body burdens. In this study, as in those reviewed by Rainbow(2002), there appeared to be no fixed total metal concentration atwhich the onset of toxic effects occurred. Consequently no rela-tionship between the metal concentration in the gammarids andtoxicity was established and, furthermore, this work adds to thegrowing evidence that attributes metal toxicity to overwhelminginflux and bioaccumulation rather than a critical total body burden(e.g. Casado-Martinez et al., 2010).
The MRG + exo and MTLP fractions were the two sub-cellularfractions that were most enriched in Cu and Zn during exposures,and two fractions that increased in metal load with increasingwhole body burden. Within the MRG + exo fraction metal is boundto, presumably both, insoluble granules and exoskeleton. Theexoskeleton of G. pulex is comprised of chitin with the outermostlayer hardened by calcium deposits (Schmitz, 1992). Metal boundto organic chitin would be expected to be digested by NaOH withinthe fractionation protocol and form part of the debris fraction whichis not considered toxicologically significant (Cain et al., 2004). How-ever, metal associated to the calcium rich outer-layer would be
resistant to NaOH digestion and would pellet with MRG followingcentrifugation. Metal adhered to this portion of exoskeleton wouldlikely be biologically inactive as it is not within the organism. How-ever, the possibility exists, especially during the longer exposures,that internalised Cu and Zn could be distributed to the exoskeleton
1 Toxico
a<eaeMtM
ri2mRMariittifdtt2wl
tbfdrTirstatt(itsiialim2isiiingcfirl
02 F.R. Khan et al. / Aquatic
s was the case with the marine copepod, Acartia spp., in which97% of total Cd, Co and Zn body burdens were stored within thexoskeleton (Reinfelder and Fisher, 1994). Importantly, Acartia spp.re without other forms of insoluble detoxification. Although, thexact nature of metal binding to the exoskeleton in the G. pulexRG + exo fraction is debatable, we can hypothesise that it adds
o the overall fraction burden, but is distinct from metal bound toRG.Within amphipods MTLP and MRG fractions have typically been
elated to the detoxification of trace metals through metalloth-onein (Correia et al., 2002a) and insoluble granules (Nassiri et al.,000; Correia et al., 2002b). Our results indicate that a change inetal distribution occurred within those gammarids exposed at
elubbus and St. Erth with both Cu and Zn being distributed intoRG + exo fraction between Days 4 and 16. Granules are usually
ssociated with the digestive system and are excreted in order toelease accumulated toxicants (Icely and Nott, 1980). The increasen the concentration of Cu and Zn in these fractions, particularlyn gammarids with high metal body burdens, might explain howhese organisms were able to cope with internal metal concentra-ions that had caused mortality at Godolphin. Although the changen the metal distribution from the MTLP fraction to the MRG + exoraction was not statistically evident, it would support a previouslyescribed amphipod detoxification pathway in which metal boundo metallothionein is degraded within the acidic environment ofhe lysosome and gives rise to insoluble granules (Nassiri et al.,000). Within these granules metal is bound to sulfhydral groupshich originate from the sulfur rich cysteine amino acid in metal-
othionein (Luoma and Rainbow, 2008).However, there was no difference in the sub-cellular distribu-
ion of Cu or Zn between the populations as the proportion fractionurdens remained constant to overall body burden. Thus the con-erred tolerance of the historically impacted population was notue to a greater capacity for metal detoxification that has beenecorded in other invertebrate species (e.g. Postma et al., 1996).he question remains: by what mechanism did the historicallympacted population achieve lower metal induced responses? Theeview by Maltby (1999) elegantly proposes that changes in theensitivity to metals at the gill surface can account for popula-ion differences. Waterborne metal uptake in crustaceans occurst the gill which has been shown to have the dual role of respira-ion and active ion uptake (Péqueax, 1995). High concentrations ofrace metals can cause ultrastructural damage to gill epithelial cellsLawson et al., 1995) resulting in osmoregulatory and respiratorympairment (Nonnette et al., 1993; Maltby, 1999 and referencesherein). The respiratory function in individuals from contaminatedites is more resistant to damage caused by trace metals than naïvendividuals and furthermore resistance has been linked to a phys-ological adaptation in respiration (Maltby, 1999). However, metalccumulation on the gill surface also disrupts iono- and osmoregu-atory functions through the impaired function of proteins involvedn those functions and these effects can occur at relatively low water
etal concentrations. An exposure of 10 �g L−1 Cu at a hardness of31 mg L CaCO3
−1 significantly inhibited gill Na+/K+ ATPase activityn G. pulex (Brooks and Lloyd Mills, 2003) and 1.25 mg L−1 Zn expo-ure in ASTM hard water (160–180 mg L CaCO3
−1) resulted in annability to maintain osmolality with the haemolymph increasingn osmotic pressure and both Na+ and K+ concentrations caus-ng haemoconcentration (Spicer et al., 1998). In the latter studyaïve individuals were again more sensitive to adverse affects thanammarids collected from contaminated sites. Through this we
ould hypothesise that the gammarids from Drym, a populationrom a metallogenic region, were adapted at the gill for greaternsensitivity to metal damage. This adaptation may have causededuced transport of Cu and Zn across the gill which resulted in theower levels of accumulation. The differences in oxidative stresslogy 102 (2011) 95–103
and mortality between the Drym and naïve (R. Cray) populationscan be attributed to the accumulated metal concentration. Thoughthe partitioning of metals into MRG + exo and MTLP fractions didappear to allow greater body burdens to be non-toxic, our resultssuggest that between populations the advantageous adaptation ofthe Drym population was a reduction in uptake, most likely via thegill, which lead to greater survivability in particular at Relubbusand St. Erth.
In conclusion, the method of exposing two populations withinthe natural environment was successful in revealing small dif-ferences in responses that were caused by their respective metalexposure history. The difference between the two populations wasmanifested in bioaccumulation, oxidative stress responses and ulti-mately mortality with the historically impacted population beingless susceptible to Cu and Zn uptake and toxicity. As previouslyfound with comparisons of metal tolerant and naïve G. pulex pop-ulations, metal handling strategies (MTLP and MRG) were notdifferent. However, our results appear to suggest the Drym pop-ulation was adapted for lower Cu and Zn uptake resulting in lowerstress and mortality. This study shows that the difference in expo-sure histories of the two populations can cause great differences, interms of mortality rates, bioaccumulation and toxic stress responseduring subsequent exposure to higher concentrations.
Acknowledgements
The authors acknowledge to P.S. Rainbow (Natural HistoryMuseum) for the critical reading of a draft version of thismanuscript, to C. Moncrief (Natural History Museum) for hisstatistical guidance and the comments provided by two anony-mous reviewers. This research was supported by PhD researchgrant from the Natural Environment Research Council (NERC)NER/S/A2004/1215.
References
Baudrimont, M., Andrès, S., Metivaud, J., Lapaquellerie, Y., Ribeyre, F., Maillet,N., Latouche, C., Boudou, A., 1999. Field transplantation of the freshwaterbivalve Corbicula fluminea along a polymetallic contamination gradient (RiverLot France). II. Metallothionein response to metal exposures. Environ. Toxicol.Chem. 18, 2472–2477.
Bonneris, E., Giguere, A., Perceval, O., Buronfosse, T., Masson, S., Hare, L., Camp-bell, P.G.C., 2005. Sub-cellular partitioning of metals (Cd, Cu, Zn) in the gills ofa freshwater bivalve, Pyganodon grandis: role of calcium concretions in metalsequestration. Aquat. Toxicol. 71, 319–334.
Bossuyt, B.T.A., Janssen, C.R., 2005. Copper regulation and homeostasis of Daphniamagna and Pseudokirchneriella subcapitata: influence of acclimation. Environ.Pollut. 136, 135–144.
Bradford, M.M., 1976. Rapid and sensitive method for quantitation of microgramquantities of protein utilizing principle of protein-dye binding. Anal. Biochem.72, 248–254.
Brooks, S.J., Lloyd Mills, C., 2003. The effect of copper on osmoregulation in thefreshwater amphipod Gammarus pulex. Comp. Biochem. Physiol. A 135, 527–537.
Brown, B.E., 1977. Effects of mine drainage on River Hayle, Cornwall. A. Factors affect-ing concentrations of copper, zinc and iron in water, Sediments and DominantInvertebrate Fauna. Hydrobiologia 52, 221–233.
Cain, D.J., Luoma, S.N., 1998. Metal exposures to native populations of the caddis-fly Hydropsyche (Trichoptera: Hydropsychidae) determined from cytosolic andwhole body metal concentrations. Hydrobiologia 386, 103–117.
Cain, D.J., Luoma, S.N., Wallace, W.G., 2004. Linking metal bioaccumulation of aquaticinsects to their distribution patterns in a mining impacted river. Environ. Toxicol.Chem. 23, 1463–1473.
Casado-Martinez, M.C., Smith, B.D., Luoma, S.N., Rainbow, P.S., 2010. Metal toxicity ina sediment-dwelling polychaete: threshold body concentrations or overwhelm-ing accumulation rates? Environ. Pollut. 158, 3071–3076.
Chemical Survey of R. Hayle, 1983. Rectorate of Engineering and EnvironmentalServices. South West Water.
Comber, S.D.W., Merrington, G., Sturdy, L., Delbeke, K., van Assched, F., 2008. Copper
and zinc water quality standards under the EU Water Framework Directive: theuse of a tiered approach to estimate the levels of failure. Sci. Total Environ. 403,12–22.Correia, A.D., Livingstone, D.R., Costa, M.H., 2002a. Effects of water-borne copperon metallothionein and lipid peroxidation in the marine amphipod Gammaruslocusta. Mar. Environ. Res. 54, 357–360.

Toxico
C
D
D
F
G
G
G
G
H
I
K
K
L
L
L
L
M
M
F.R. Khan et al. / Aquatic
orreia, A.D., Costa, M.H., Ryan, K.P., Nott, J.A., 2002b. Studies on biomarkers ofcopper exposure and toxicity in the marine amphipod Gammarus locusta (Crus-tacea). I. Copper-containing granules within the midgut gland. J. Mar. Biol. Assoc.U.K. 82, 827–834.
awkins, J., Donoghue, S., 1992. Invertebrates associated with aquatic vegetation inthe River Cray. London Naturalist 71, 71–74.
e Coen, W., Robbens, J., Janssen, C., 2006. Ecological impact assessment of metal-lurgic effluents using in situ biomarker assays. Environ. Pollut. 141, 283–294.
ialkowski, W., Fialkowska, E., Smith, B.D., Rainbow, P.S., 2003. Biomonitoring sur-vey of trace metal pollution in streams of a catchment draining a zinc and leadmining area of Upper Silesia Poland using the amphipod Gammarus fossarum.Int. Rev. Hydrobiol. 88, 187–200.
ower, A.M., Darlington, S.T., 1990. Relationships between copper concentrationsin larvae of Plectrocnemia conspersa (Curtis) (Trichoptera) and in mine drainagestreams. Environ. Pollut. 65, 155–168.
ower, A.M., Myers, G., Kent, M., Foulkes, M.E., 1994. Relationships betweenmacroinvertebrate communities and environmental variables in metal-contaminated streams in South-West England. Freshw. Biol. 32, 199–221.
raca, M.A.S., Maltby, L., Calow, P., 1994. Comparative ecology of Gammarus pulex (L)and Asellus Aquaticus (L). 1. Population-dynamics and microdistribution. Hydro-biologia 281, 155–162.
rosell, M., Wood, C.M., 2002. Copper uptake across rainbow trout gills: mechanismsof apical entry. J. Exp. Biol. 205, 1179–1188.
ogstrand, C., Verbost, P.M., Wendelaar Bonga, S.E., Wood, C.M., 1996. Mechanismof zinc uptake in the gills of freshwater rainbow trout: interplay with calciumtransport. Am. J. Physiol. 270, R1141–R1147.
cely, J.D., Nott, J.A., 1980. Accumulation of copper within the hepatopancreatic cecaof Corophium volutator (Crustacea, Amphipoda). Mar. Biol. 57, 193–199.
endrick, M.J., May, M.T., Plishka, M.J., Robinson, K.D., 1992. Metals in BiologicalSystems. Ellis Horwood Limited.
han, F.R., Bury, N.R., Hogstrand, C., 2010. Cadmium bound to metal rich granulesand exoskeleton from Gammarus pulex causes increased gut lipid peroxidationin zebrafish following single dietary exposure. Aquat. Toxicol. 96, 124–129.
awson, S.L., Jones, M.B., Moate, R.M., 1995. Effect of copper on the ultrastructure ofthe gill epithelium of Carcinus maenus (Decapoda: Brachyura). Mar. Pollut. Bull.31, 63–72.
iu, J., Yeo, H.C., Daniger, S.J., Ames, B.N., 1997. Assay of aldehydes from lipid peroxi-dation: gas chromatography–mass spectrometry compared with thiobarbituricacid. Anal. Chem. 245, 161–166.
uoma, S.N., Rainbow, P.S., 2005. Why is metal bioaccumulation so variable? Biody-namics as a unifying concept. Environ. Sci. Technol. 37, 1921–1931.
uoma, S.N., Rainbow, P.S., 2008. Metal Contamination in Aquatic Environments.Science and Lateral Management. Cambridge University Press, Cambridge, UK.
acNeil, C., Dick, J.T., Elwood, R.W., 1997. The trophic ecology of freshwaterGammarus spp. (Crustacea: Amphipoda): problems and perspectives concern-ing the functional feeding group concept. Biol. Rev. Cambr. Philos. Soc. 72,349–364.
altby, L., 1999. Studying stress: the importance of organism-level responses. Ecol.Appl. 9, 431–440.
logy 102 (2011) 95–103 103
Maltby, L., Clayton, S.A., Wood, R.M., McLoughlin, N., 2002. Evaluation of the Gam-marus pulex in situ feeding assay as a biomonitor of water quality: robustness,responsiveness, and relevance. Environ. Toxicol. Chem. 21, 361–368.
Morgan, A.J., Kille, P., Sturzenbaum, S.R., 2007. Microevolution and ecotoxicology ofmetals in invertebrates. Environ. Sci. Technol. 41, 1085–1096.
Mulliss, R.M., Revitt, D.M., Shutes, R.B.E., 1996. The determination of the toxicinfluences to Gammarus pulex (Amphipoda) caged in urban receiving waters.Ecotoxicology 5, 209–215.
Muyssen, B.T.A., Janssen, C.R., Bossuyt, B.T.A., 2002. Tolerance and acclimation tozinc of field-collected Daphnia magna populations. Aquat. Toxicol. 56, 68–79.
Nassiri, Y., Rainbow, P.S., Amiard-Triquet, C., Rainglet, F., Smith, B.D., 2000. Trace-metal detoxification in the ventral caeca of Orchestia gammarellus (Crustacea:Amphipoda). Mar. Biol. 136, 477–484.
Nonnette, L., Biotel, F., Truchot, J.P., 1993. Waterborne copper causes gill damageand haemolymph hypoxia in the shore crab Carcinus maenas. Can. J. Zool. 71,1569–1576.
Postma, J.F., van Nugteren, P., BuckertdeJong, M.B., 1996. Increased cadmium excre-tion in metal-adapted populations of the midge Chironomus riparius (Diptera).Environ. Toxicol. Chem. 15, 332–339.
Péqueax, A., 1995. Osmotic regulation in Crustacea. J. Crust. Biol. 15, 1–60.Rainbow, P.S., 2002. Trace metal concentrations in aquatic invertebrates: why and
so what? Environ. Pollut. 120, 497–507.Rainbow, P.S., Poirier, L., Smith, B.D., Brix, K.V., Luoma, S.N., 2006. Trophic transfer of
trace metals: subcellular compartmentalization in a polychaete and assimilationby a decapod crustacean. Mar. Ecol. Prog. Ser. 308, 91–100.
Rainbow, P.S., Moore, P.G., 1986. Comparative metal analyses in amphipod crus-taceans. Hydrobiologia 141, 273–289.
Reinfelder, J.R., Fisher, N.S., 1994. Retention of elements absorbed by juvenile fish(Menidia-menidia, Menidia-beryllina) from zooplankton prey. Limnol. Oceanogr.39, 1783–1789.
River Hayle Fisheries Survey, 1970. Cornwall River Authority.Roelofs, D., Mariën, J., van Straalen, N.M., 2007. Differential gene expression profiles
associated with heavy metal tolerance in the soil insect Orchesella cincta. InsectBiochem. Mol. Biol. 37, 287–295.
Schmitz, E.H., 1992. Amphipoda. In: Harrison, F.W., Humes, A.G. (Eds.), MicroscopicAnatomy of Invertebrates. Vol. 9, Crustacea. Wiley-Liss Inc., New York, pp.443–528.
Spicer, J.I., Morritt, D., Maltby, L., 1998. Effect of water-borne zinc on osmoregulationin the freshwater amphipod Gammarus pulex (L) from populations that differ intheir sensitivity to metal stress. Funct. Ecol. 12, 242–247.
Vijver, M.G., Van Gestel, C.M., Lanno, R.P., Van Straalen, N.M., Peijnenburg, W.J.G.M.,2004. Internal metal sequestration and its ecotoxicological relevance: a review.Environ. Sci. Technol. 38, 4705–4712.
Wallace, W.G., Lee, B.G., Luoma, S.N., 2003. Subcellular compartmentalization of Cdand Zn in two bivalves. I. Significance of metal-sensitive fractions (MSF) andbiologically detoxified metal (BDM). Mar. Ecol. Prog. Ser. 249, 183–197.
Wheeler, M.W., Park, R.M., Bailer, A.J., 2006. Comparing median lethal concentrationvalues using confidence interval overlap or ratio tests. Environ. Toxicol. Chem.25, 1441–1444.


















