
Phylogenetic Origin of Phyllolobium with a Further Implication for Diversification of Astragalus in...
25
1 Phylogenetic Origin of Phyllolobium with a Further Implication for Diversifiction of Astragalus in China Mingli Zhang 1* , Yun Kang 2* , Lihua Zhou 3 , Dietrich Podlech 4 ( 1 Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China; 2 Pharmacy School, Fudan University, Shanghai 200032, China; 3 Department of Botany, California Academy of Sciences, San Francisco, CA 94118-4503, USA; 4 Faculty of Biology, Münich University, Münich D-80336, Germany) *Email <[email protected], [email protected]> Abstract Astragalus is a species-rich genus occurring in western arid habitats in China. However, its diversification and infrageneric relationships in this region remain unclear. In this study, based on the molecualr data, we aim to (1) test whether Phyllolobium (previously treated as a subgenus Pogonophace in Astragalus) should be warranted and (2) date the origin of Phyllolobium and probable diversification of Astragalus sensu stricto (s.s.). We seqeunced five species from Phyllolobium firstly and collected all related sequences from this genus,Astragalus s.s and their close relatives (Oxytropis and Caragana etc.). Our phylogenetic analyses suggested that all species of Phyllolobium comprise a monophyletic group sister to genera of subtribe Coluteinae. Molecular dating suggested that Phyllolobium and Astragalus s.s. originated around 8 and 10 million years ago (Ma). These two estimations are highly consistent with the intense uplifts of the Qinghai- Tibetan Plateau (QTP) inferred from the geological evidence. In addition, one section of Pogonophace (Sect.Robusti) was estimated to originate 2.50 Ma and this section with a preferencing of dry habitats seems a trace of Asian intensified aridity resulting from the intense uplift of the QTP.
Transcript of Phylogenetic Origin of Phyllolobium with a Further Implication for Diversification of Astragalus in...

1
Phylogenetic Origin of Phyllolobium with a Further
Implication for Diversifiction of Astragalus in China
Mingli Zhang 1*, Yun Kang 2*, Lihua Zhou 3 , Dietrich Podlech 4
(1 Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China; 2 Pharmacy School, Fudan University, Shanghai 200032, China;
3 Department of Botany, California Academy of Sciences, San Francisco, CA 94118-4503, USA; 4 Faculty of Biology, Münich University, Münich D-80336, Germany)
*Email <[email protected], [email protected]>
Abstract
Astragalus is a species-rich genus occurring in western arid habitats in China. However,
its diversification and infrageneric relationships in this region remain unclear. In this
study, based on the molecualr data, we aim to (1) test whether Phyllolobium (previously
treated as a subgenus Pogonophace in Astragalus) should be warranted and (2) date the
origin of Phyllolobium and probable diversification of Astragalus sensu stricto (s.s.). We
seqeunced five species from Phyllolobium firstly and collected all related sequences from
this genus,Astragalus s.s and their close relatives (Oxytropis and Caragana etc.). Our
phylogenetic analyses suggested that all species of Phyllolobium comprise a
monophyletic group sister to genera of subtribe Coluteinae. Molecular dating suggested
that Phyllolobium and Astragalus s.s. originated around 8 and 10 million years ago (Ma).
These two estimations are highly consistent with the intense uplifts of the Qinghai-
Tibetan Plateau (QTP) inferred from the geological evidence. In addition, one section of
Pogonophace (Sect.Robusti) was estimated to originate 2.50 Ma and this section with a
preferencing of dry habitats seems a trace of Asian intensified aridity resulting from the
intense uplift of the QTP.

2
Key words: Astragalus, Phyllolobium, Sino-Himalayan flora, phylogeny, dating,
diversification, distribution pattern, Qinghai-Xizang (Tibet) Plateau (QTP).
As one of the largest genera of angiosperms, Astragalus comprises about 1800-2500
species worldwide (Polhill, 1981; Sanderson, 1991; Podlech, 1999; Lock, Schrire, 2005).
There are three diversification centers in the world, namely Iran-Turkey center,
Himalayan center and western North American center (Sanderson, 1991). As one of three
centers, Himalaya and adjacent regions, locating in southwestern China, is named the
Sino-Himalayan floristic region (Wu & Wu, 1999). Here has a rich biodiversity among
the Northern Temperate flora (Wu, 1988).
According to Fu et al. (1993), there are ca. 290 species belong to eight subgenera sensu
Bunge (1868, 1869, whole genus includes eleven subgenera) in China. Most species in
the genus have the characteristic of northern temperate herbaceous, appearing in the
southwestern cold and humid/drought area, and northwestern drought and semi-drought
area. For instance, subgenus Pogonophace, with a taxonomical diagnostic of the bearded
stigma, mainly occurs in southwestern cold and humid high mountains (Wenninger, 1991;
Zhang, 2000).
Astragalus molecular systematics in legume and the subfamily Faboideae
(Papilionoideae) (see Kajita et al., 2001; Dong et al. 2003; Wojciechowski, 2003, 2005;
Wojciechowski et al., 2004) shows that this genus should be regarded as one of
representative large groups in Galageae. Galageae, Vicieae and other temperate
herbaceous groups compose most of a young clade in legumes, to been named IRLC
(inverted-repeat-lacking clade) which is one of eleven subclades of the subfamily

3
Faboideae (Wojciechowski et al., 2004). Liston & Wheeler (1994), Sanderson & Liston
(1995), Sanderson & Wojciechowski (1996), and Wojciechowski et al. (1999) discussed
the monophyly of Astragalus based on the sequence data. As a larger world-cosmopolitan
group, non-monophyly at subgeneric rank has been presented based on the molecular
phylogeny from Eurasian taxa (Kang et al., 2003; Kazempour et al., 2003, 2005;
Wojciechowski 2005). Two new genera, Phyllolobium and Podlechiella have been
proposed to be segregated from Astragalus (Kang et al., 2003; Kazempour et al., 2003,
2005). For the historical biogeography, Sanderson & Wojciechowski (1996) used ITS
data indicated that the diversification rates in Astragalus were higher than its closest
relatives. Wojciechowski (2005) used matK and ITS data to estimate the ages of
Astragalus and relatives; the results shown that Astragalus and its sister genus Oxytropis
shared a crown clade time ca. 16.1 Ma. The estimated age of Astragalus was ca. 12.4 Ma,
but the so-named Neo-Astragalus (New World especially North America aneuploid,
chromosome number n=11-15) diverged ca. 4.4 Ma. The Old World groups are generally
euploid (n=8, 16) and probably older than that of New World. Lavin et al. (2005) recently
used fossils as the calibrations to date legumes.They inferred a rapid diversification of
lineages during the Tertiary. Astragalus was demonstrated to be a rapidly evolving group
in Tertiary, and the divergence time of Coluteinae and Astragalinae was estimated at ca.
14.8 Ma (see Fig. 2 and node 78 of Table 2, Lavin et al., 2005). The divergence time of
Astragalus should certainly be later of that time. However, for such a large genus, only
four species of Astragalus were sampled in their dataset (Lavin et al., 2005). Recently, a
South American Astragalus phylogenetic dating based on ITS, trnD-trnT and trnfM-trnS1
sequences, showed a recent and rapid diversification in this area (Scherson et al, 2008).

4
Therefore, we so far can investigate Astragalus diversification in China in the light of
previous molecular framework of phylogenetics and biogeography. Meanwhile, a
stimulating motivation for this paper also came from the Astragalus diversification in
Sino-Himalayan region/China. As the well-known, the intense uplifting of QTP during
the Late Tertiary tremendously changed the environment and ecology in Asia and China,
organic evolution must be influenced by this geological event. The Astragalus
distribution pattern of southwestern cold and dry alpine area and northwestern dry area in
China, should be rationally analyzed by joining the background of historical geological
processes in Sino-Himalayan region and China.
Now we turn our attention to the diversification of Astragalus in China, 1) to test
whether Phyllolobium should be warranted; 2) to date the origin of Phyllolobium and
probable diversification of Astragalus in China; 3) to understand the cause of Astragalus/
Phyllolobium diversification by linking it to the uplifting of QTP
Materials and Methods
Taxa sampling
All species of Astragalus in Sino-Himalaya/China, with nrDNA ITS 5.8S and
contributed to Genbank, and five species in Pogonophace to be sequenced firstly by us in
this paper, were sampled, see appendix 1. The outgroup genera of Astragalus, such
Oxytropis, Caragana, Hedysarum, Colutea, Swainsona, were also analyzed using data
from Genbank. Other related legume groups which have fossil records, such as Sophora,
Dalbergia, Pueraria, Cercis, Acacia, and so on, were also sampled so that use them as

5
the calibration constraint for the phylogenetic dating. thus, a total 101 species were
evaluated, see appendix 1.
DNA sequence data
Five species in Pogonophace were collected from wild populations in southwestern
China, see appendix 1. The leaves were dried by silica gel for DNA extraction. The
sequencing procedure of ITS 5.8S utilized was that of Kang et al. (2002). All other
sequence data for putatively related taxa were downloaded from the Genbank, see
appendix 1.
Phylogenetic analysis
ITS sequence data for all 101 species were transferred to BioEdit ver.5.0.9 (Hall, 1999);
the alignment was done by using Clustal X 1.83 (Jeanmougin et al., 1998). The alignment
data then was moved to BioEdit v.5.0.9 for manual improvement. An original nexus form
of the dataset was exported for phylogenetic analysis.
Maximum parsimony analysis
Parsimony analyses were implemented by employing PAUP ver. 4.0 (Swofford, 2002).
Multiple tree searches were conducted using heuristic search options, including random
additional sequences (1000 replicates) holding ten trees per replicate, and tree-bisection-
reconnection (TBR) branch swapping, with retention of multiple parsimonious trees.
These parsimonious trees then were used to calculate the consensus tree.
Bootstrap analyses were used to determine clade support. Non-parametric bootstrap
resampling proportions (Felsentein, 1985) were estimated from 1000 bootstrap replicates

6
incorporating heuristic parsimony searches using addition sequence and branch swapping
options, and TBR and maxtrees.
Modeltest
For the Maximum Likelihood (ML) and MrBayes analyses (MB), the best fit of DNA
substitution model should be found. The Akaike information criterion (AIC) and
hierarchical likelihood ratio test (hLRT) were calculated based on the log likelihood
scores of 56 models using Modeltest 3.5 (Posada & Crandall, 1998). In general, AIC is
chosen (Posada & Buckley, 2004).
For ITS spacer dataset of this paper, the GTR+I+G model by AIC was selected as the
most appreciated model in Modeltest 3.5. It was with the nucleotide frequencies A =
0.2039, C = 0.3075, G = 02741, T = 0.2145, a gamma shape parameter of 1.8037 and an
assumed proportion of invariable sites of 0.2904.
Maximum likelihood analysis
Maximum likelihood search was performed on the basis of the result of Modeltest in
PAUP. The parameters of best model, such as the base frequency, the mean relative
substitution rates, proportion of invariable sites, Gamma distribution shape, were all
employed. The heuristic search and bootstrap were implemented as in parsimony analysis
in PAUP above mentioned.
Bayesian inference
Bayesian inference of phylogenetic trees was analyzed by employing MrBayes version
3.0 (Huelsenbeck & Ronquist, 2001). Some parameters from the Modeltest, was also
included in the analysis. The option was set up using 1,000,000 generations of Markov

7
Chain Monte Carlo (MCMC) searches and a sample frequency of 1000. Saturation was
reached after a burn-in of 1000 generations.
The clade support was assessed using Bayesian posterior probabilities (Huelsenbeck et
al., 2002), these probabilities were estimated as the proportion of trees sampled after
‘burn-in’ that contained each of the observed bipartitions.
Dating
Reference fossils
Legumes are well represented in the fossil record (Heredeen et al., 1992). But there are
no fossils assignable to Astragalus and allied genera, such as Oxytropis, Caragana,
Hedysarum, Colutea, and Swainsona, etc. According to Tao (1992), Tao et al. (2000),
Institute of Botany & Nanjing Institute of Geology and Palaeontology (IB & NJGP),
Academia Sinica (1978), Li et al. (1995), and Guo & Zhou (1992) more than 20 legume
genera are recorded from fossil records from China. Most of these are dated to the
Miocene, several approach to Eocene and Paleocene, but only Bauhinia and Cercis
extend to the Late Cretaceous. Sophora occurring in Oligocene-Miocene, is credible,
since fossils are known from the Eocene of eastern Siberia and North America (IB &
NJGP, 1978; Tao et al., 2000). In China, Sophora fossils were found in Oligocene from
Heilongjiang, Miocene from Shandong and Yunnan, and Pliocene from Shanxi (Tao et al.,
2000). Acacia in Eocene is also well represented in museum collections. Dalbergia
fossils including leaves and fruit were recorded Eocene-Miocene boundary in North
America, and Oligocene-Miocene boundary in Europe, Miocene in Yunnan Province,
China. Fossil leaves of Pueraria were appeared in Miocene in Shandong and Yunnan

8
provinces, China (IB & NJGP, 1978; Tao et al., 2000). These genera were used as
outgroups and the constraint minimum ages, i.e., the maximum fossil times were selected
as the generic minimum age MRCA (most recent common ancestor) in dating. For details
of the fossil references refer to Table 1.
Table 1. Fossil references of five genera use for dating ____________________________________________________________________________________________ Taxa Time Ma Location Reference Cercis 60-11 Late Cretaceous-Miocene China Tao, 1992; Tao et al.,2000 Eocene American Lavin et al., 2005 Acacia 47-42 Eocene Liaoning, China; Tanzania Tao,et al.,2000; Heredeen et al.,2000 15 Miocene Dominican Lavin et al.,2005 Sophora 35-9 Oligocene-Miocene China, Siberia, N America Tao et al.,2000; IB & NJGP,1978 Pueraria 17-5 Miocene Yunnan, Shandong, China Tao,1992,2000 Dalbergia 19.5-5 Miocene China, N America, Europe Tao,1992,2000; IB & NJGP,1978 _____________________________________________________________________________________________
In terms of the ancient fossil record and phylogenetic tree, the most root should be
selected Cercis. In southern China Guangdong province, Cercis fossil found in Late
Cretaceous to Eocene, at Oligocene in Yunnan, and Miocene in Shandong and Qinghai.
So, both genera were used to root taxa for constraint ages, and root age was determined
as 60 Ma.
Dating implement
Phylogenetic dating was conducted by three approaches, r8s, PAML and BEAST. In r8s
version 1.7 (http://ginger.ucdavis.edu/r8s/) based on the Bayesian tree, Penalized
likelihood PL (semi-parametric rate smoothing, Sanderson, 2002) and NPRS (non-
parametric rate smoothing, Sanderson, 1997) methods were used. However, PL is
generally in favor with and many applications (e.g. Bremer et al., 2004; Renner & Zhang,
2004; Near & Sanderson, 2004; Renner & Givnish eds., 2004; Anderson et al., 2005; Bell
& Donoghue, 2005; Lavin et al., 2005). And the smoothing parameter of Cross
Validation option in r8s were estimated for using in PL and NPRS.

9
PAML (Phylogenetic Analysis by Maximum Likelihood, Yang, 2007) was
implemented by PAML 4.2 (http://abacus.gene.ucl.ac.uk/software/paml.html). PAML
belongs to relaxed molecular clock method, Bayesian tree was used as BASEML, dating
was using MCMCTREE in PAML. The independent rate model (Rannala & Yang, 2007)
with HKY85+Γ5 model for nucleotide substitutions, and multiple fossil calibrations were
used. Two separate MCMC analyses were run for 1,000,000 generations (burnin 10%)
with parameters sample size every 1000 steps. The mean and 95% confidential internals
of MRCA (most recent common ancestor) nodes were yielded.
BEAST v1.46 (http://beast.bio.ed.ac.uk/) was also used to estimate divergence times
(Drummond & Rambaut, 2007). Best-fit models of nucleotide substitution parameters for
the priors in BEAST were yielded by Modeltest. A Yule process speciation prior and an
uncorrelated lognormal model of rate variation were accepted. And normal distribution
was chosen. Tracer v1.4 was used to measure the efficient sample size of each parameter
and mean and 95% confidential internals. Two separate MCMC analyses were run for
10,000,000 generations (burnin 10%) with parameters sample size every 1000 steps. The
mean and 95% confidential internals of MRCA nodes were produced.
Results
Phylogeny
In the maximum parsimony analysis, ITS sequence dataset consist of total 719 aligned
positions, with 385 potential parsimony-informative characters. Parsimonious trees was
with length=1730, CI=0.4965, RI=0.8756, a consensus tree of the most parsimonious
trees was yielded. ML resulted in one best tree. Bayesian analysis presented 10001 burn-

10
in trees, and a consensus tree was conducted with the support of posterior probabilities to
clades.
Three phylogenetic analyses resulted in almost identical topology of trees, especially at
the generic level, only exceptionally, Astragalus cysticalyx nested in the clade of
Sphaerophysa and Colutea, and Calophaca tianshanica nested in Caragana clade. The
clades were with high bootstrap and posterior probability values, most over 90%
bootstrap and 95% posterior probability. A Bayesian inference tree was shown as the
phylogenetic relationships in Fig. 1.
Dating
The dating provided estimated ages of Astragalus and relatives, see Table 2. In r8s, the
cross-validation value of all methods is identical, smoothing value 1. All fidelities of used
methods were shown “good”. So, the alternate estimated ages should be in favor of PL-
TN, likes most applications (e.g. Bremer et al., 2004; Anderson et al., 2005; Near &
Sanderson, 2004; Lavin et al., 2005; Wojciechowski, 2005; Near & Sanderson, 2004;
Bell & Donoghue, 2005). The dating results of Astragalus and relatives in r8s were
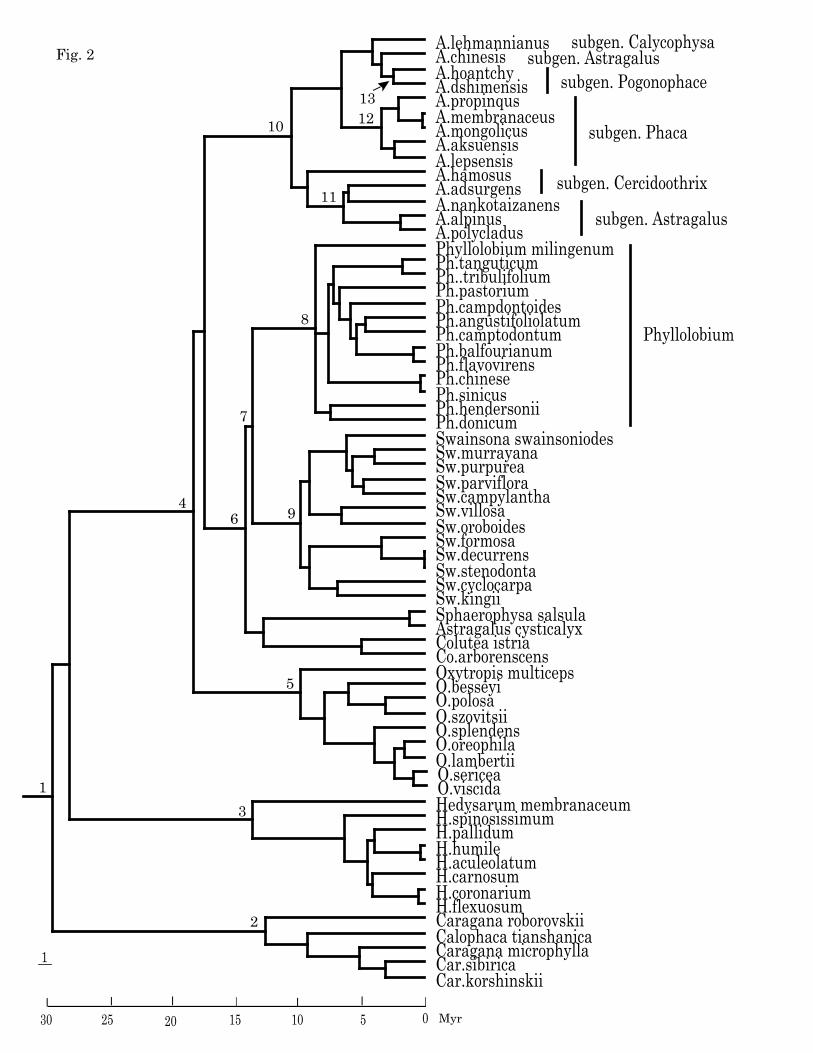
presented in Fig. 2 and Table 2.
The estimated ages of Astragalus and relatives in PAML and BEAST for related nodes
refer to Table 2, were offered in Table 2. Three approaches provide roughly congruent
estimated ages to the nodes. However, the estimated ages in PAML and BEAST are
higher than that of r8s especially those in BEAST, for instance, node 8 Phyllolobium 8.39
Ma in PL, 6.39 Ma in PAML, 12.43 in BEAST, and node 10 Astragalus 10.34 Ma in PL,
12.78 Ma in PAML, 18.90 in BEAST etc. The estimated ages from r8s and PAML are

11
resemble and available. So, in following discussion, we will prefer the estimated ages
from r8s and PAML.
Table 2. The estimated ages using different methods and algorithms, in PAMLand BEAST mean and 95% confidential intervals of the nodes are shown. The classification system following Lock & Schrire (2005), the subtribes following Polhill (1981), the number in front of taxa refer to the nodes in Fig. 2. ______________________________________________________________________________________________ r8s PAML4.2 BEAST
PL-TN PL-Powell NPRS-Powell MCMCTREE 1 Tribe Hedysareae 28.65 29.27 32.01 29.35 (20.29-39.75) 25.68 (24.76-40.56) 2 Caragana 12.22 12.77 20.57 9.28 (4.01-17.20) 14.02 (5.16-32.49) 3 Hedysarum 13.29 13.85 16.80 12.22 (6.20-20.84) 16.13 (6.32-27.46) 4 Tribe Galageae 17.87 18.68 21.04 18.88 (13.01-26.29) 25.56 (16.07-34.18) 4 Subtribe Astragaleae s.str. 17.87 18.68 21.04 18.88 (13.01-26.29) 25.56 (16.07-34.18) 5 Oxytropis 9.63 11.65 15.97 3.66 (1.41-7.86) 9.27 (2.2-20.2)
6 Subtribe Coluteoid s.str. 13.80 14.51 16.34 12.31 (7.85-17.25) 20.96 (12.12-27.93) 7 Phyllolobium+Swainsona 13.30 14.04 15.69 11.00 (7.14-16.10) 19.21 (10.85-25.61) 8 Phyllolobium 8.39 8.82 9.68 6.39 (3.73-10.50) 12.43 (6.74-19.88),
9 Swainsona 9.62 10.21 12.07 7.64 (5.04-11.22) 15.02 (7.99-21.12) 10 Astragalus 10.34 10.92 13.83 12.78 (8.59-18.66) 18.90 (10.4-27.05)
11 Subgen. Astragalus 6.25 6.60 8.63 5.79 (2.72-9.75) 9.44 (3.46-16.79) 12 Subgen. Phaca 3.29 3.51 5.80 3.80 (1.60-7.11) 8.28 (1.91-14.82) 13 Subgen. Pogonophace 2.50 2.68 4.41 1.53 (0.45-3.43) 3.28 (2.07-14.4) ______________________________________________________________________________________________
Discussions
Phyllolobium, a credible monophylenetic genus, and related remarkable to Coluteae
changes the monophyly of Astragalus
From Fig. 1, we determined that Phyllolobium was phylogenetically far away from
Astragalus in contrast to previous presentations (Kang et al., 2003; Wojciechowski &
Sanderson, 2004; Wojciechowski, 2005), which did not sample sufficient taxa from
Phyllolobium. For instance, Wojciechowski & Sanderson (2004) and Wojciechowski
(2005) only sampled one species, A. complanatus (=Phyllolobium chinense. Kang et al.
(2002) selected 7 species from Phyllolobium. Thus it is not surprising that their result
based on a few species is not affirmed or somewhat suspect. In this paper Fig. 1 and Fig.
2, we smapled a total of 15 species including 13 from Phyllolobium (Ph. sinicus in

12
Genbank is perhaps Ph. chinense) and two A. hoatchy and A. dshimensis from Astragalus,
were sampled from previous Pogonophace sensu Zhang (2002). In Phyllolobium, Ph.
chinense (=A. complanatus) belongs to sect. Phyllolobium, Ph. hendensornii to sect.
Trichostylus, the rest species is included in sect. Bibracteola. These three sections and
their members in fact present a taxonomical framework within Phyllolobium (Zhang &
Podlech, 2006). Therefore, Phyllolobium can be confirmed confidently to be a genus and
monophyletic. This is very different from the traditional circumscriptions of taxonomical
treatment to put Phyllolobium to Astragalus (Fu et al., 1993; Polhill, 1981). In addition,
Phyllolobium is allied to the subtribe Coluteinae rather than Astragaleae. Consequently,
the diversification of Phyllolobium extracting from Astragalus demonstrates that the
customary circumscription of Astragalus is not monophyletic. The Old World Astragalus
molecular systematics recently revealed that eight traditionally recognized Astragalus
subgenera Epiglottis, Trimeniaeus, Phaca, Hypoglottis, Calycophysa, Tragacantha,
Cercidothrix and Calycocystis have no monophyletic support (Kazempour et al., 2003,
2005). Clearly, Phyllolobium provides another non-monophyletic evidence to Astragalus,
and it jumped over the circumscription of Astragalus.
At the same time, in the light of Phyllolobium appearing concentrately in Hengduan
Mts. and eastern Himalaya (Zhang, 2000), the variation of Phyllolobium could be
considered as one of most notable diversification in Astragalus and in Sino-Himalayan
center of this genus.
Phylogenetic relationships of Astragalus/Phyllolobium and ralatives
The topology of phylogenetic tree concerning tribe Galegeae and tribe Hydesareae in the
present paper (see Fig. 1, Fig. 2), is in agreement with Wojciechowski et al (1999, 2000).

13
The molecular phylogeny (Wojciechowski et al., 1999, 2000) obviously affected previous
tribe Galageae classification (Polhill, 1981), see recent delimitation of taxa (Lock &
Schrire, 2005). A remarkable change in recent system (Lock & Schrire, 2005) is that
Caragana and allied groups, Calophaca and Halimodendron were extracted from tribe
Galegeae and transferred into tribe Hedysareae. This change was also shown clearly in
Fig. 1, Fig. 2. Astragalus and Oxytropis are generally considered as a sister group and the
latter is considered to be dereived from the former (Pohill, 1981), but Oxytropis was in
front of Astragalus in Fig. 2.
Within Astragalus, according to Fu et al. (1993), five subgenera were involved in this
paper. In Fig. 2, three phylogenetic clades were clearly present. Subgenus Phaca was
monophyletic. A. hoatchy and A. dshimensis in subgenus Pogonophace also constituted a
monophyletic group. Subgenus Cercidothrix (with A. hamosus and A. adsurgens) and
subgenus Astragalus shared an ancestral node. Subgenus Astragalus, a large group
comprising more than 60 species in China, does not appear to be monophyletic. So, like
as previous studies (see Kazempour et al., 2005), most subgenera in Astragalus have less
monophyletic support.
Infra-Phyllolobium, there is no a clear taxonomic status to be shown, but both sect.
Phyllolobium with Ph. chinese (=A. complanatus) and sect. Trichostylus with A.
hendensornii respectively are distantly related to other species of sect. Bibracteola in
phylogenetic tree, it implies three sections are resolved (Fig. 2).
Diversification ages of Astragalus/Phyllolobium and relatives
As the outgroup of Astragalus, Hedysaroid clade had an estimated age ca. 28.65 Ma in
PL 29.35 Ma in PAML 25.68 Ma in BEAST, see Table 2, it approaches Lavin et al.

14
(2005) estimated age ca. 29.3 (node 77). Caragana, Hedysarum, Oxytropis, and
Swainsona etc. had the estimated ages at Late Miocene ca. 13.85-7 Ma.
In the previous dating for Astragalus and relatives, Wojciechowski (2003) estimated
Astragalean crown age was 8-10 Ma (8.8-10 Ma use PL, 10.1 Ma use NPRS), or 16.4 Ma
(Wojciechowski, 2005), or ca. 10 Ma (Lavin et al., 2004), or ca. 14.8 Ma (Lavin et al.,
2005). All of these differences are perhaps mainly resulted from different taxa or
different sequence datasets, such as matK, rbcL and ITS sequence datasets. Our dating to
Astragalean clade was ca 17.87 Ma in PL (18.88 Ma in PAML), which is approach 16.4
Ma (Wojciechowski, 2005).
For Astragalus age, Wojciechowski (2005) estimated it as 12.4 Ma; our estimated age
based on Chinese species in Table 2, was 10.34 Ma in PL (12.78 Ma in PAML), which is
behind the world-wide Astragalus age and older than that of 4.4 Ma of the New World
“Neo-Astragalus” aneuploid group (Wojciechowski, 2005), it should be rational.
Meanwhile, this older age 10.34 (12.78) Ma near 12.4 Ma, implies that the Chinese
Astragalus groups should be ancient in the genus.
Within Astragalus, the estimated ages of three subgenera Phaca, Astragalus, and
Pogonophace, are shown in Table 2 and Fig. 2. Their divergence time was approximately
at 6.5-3.3 Ma in PL. Subgenus Phaca formed a natural group; its estimated age was ca.
3.29 Ma in PL (3.80 Ma in PAML). The clade age of subgenus Astragalus was older, ca.
6.25 Ma in PL (5.79 Ma in PAML). While subgenus Pogonophace (Fu et al., 1993;
Zhang, 2002), comprising section Robusti with two species A. hoantchy and A.
dshimensis and distributed in the arid area of northwestern China (Zhang, 2000), had an
young estimated age 2.50 Ma in PL (1.53 Ma in PAML)

15
Phyllolobium has an estimated age 8.39 Ma in PL (6.39 Ma in PAML), indicating its
diversification is older than that of the subgenera within Astragalus mentioned just above,
not only this, considering the born age 13.30 Ma in PL (11 Ma in PAML) of
Phyllolobium and Swainosa, probably means these two groups respectively from Tibet
and Australia have an early diversification since this born age approaches the diversified
age 13.80 Ma in PL (12.31 Ma in PAML) of three groups respectively from South
Europe, Mediterranean, West and Central Asia (groups of Coluteae, such as Colutea,
Sphaerophysa), Tibet (Phyllolobium) and Australia (Swainosa). Obviously, these legume
groups present a case of three systematic and phytogeographical disjunctive
diversifications from Europe-Mediterranean-W & C Asia, Tibet and Australia.
Analyses of the distribution pattern from QTP uplift and the dating
According to the distribution pattern of Astragalus in Sino-Himalayan region/China, a
rational explanation of diversification should be linked to the geological process of the
QTP uplifting in Miocene. The ecological environment and organic evolution in
Himalayas and adjacent region are tremendously influenced by QTP uplifting, which has
been illustrated by many organisms, e.g. Nannoglottis (Asteraceae) (Liu et al., 2002),
Ligularia-Cremanthodium-Parasenecio complex (Asteraceae) (Liu et al., 2006), the
schiozothoracine fishes (He et al., 2003). Astragalus and allied genera, as mentioned
above, with the Miocene estimated ages, should be certainly explained under the
background of QTP intense uplifting in Miocene.
Based on the investigations, the QTP intense uplifting appears at about 8 Ma (Harrison
et al, 1992; Molnar et al., 1993) or early 14-15 Ma (Coleman & Hodges, 1995; Spicer et
al., 2003), or late 3.6 Ma (Li et al., 1979, 1998; Zhong et al., 1996; Zheng et al., 2000).

16
Anyway, the period 8-3.6 Ma is very important yet since the evidence of 3.6 Ma of Asian
monsoon origin (An et al., 2001). The Asian monsoon was speculated to be resulted from
the QTP intense uplifting. Consequent climate change heavily influenced biological
evolution.
The estimated age ca. 10.34 Ma in PL (12.78 Ma in PAML)of Astragalus in the Sino-
Himalaya/China, was approximately at the beginning stage of the plateau intense uplift.
The divergence times 6.5-3.3 Ma in PL at subgeneric level in Astragalus, fall well into
time 8-3.6 Ma of the plateau intense uplift. It implies that Astragalus diversification in
Sino-Himalayan region was triggered by the uplifting, in other words, Astragalus and the
geology in this region are co-evolutionary.
The Phyllolobium estimated age was ca. 8.39 Ma in PL (6.39 Ma in PAML) just
approaching 8 Ma of the QTP intense uplifting, clearly it implies that Phyllolobium is a
product of the plateau intense uplifting. In particular, most Phyllolobium species densely
endemic to Hengduan Mts. and eastern Himalaya (Zhang, 2000), an observation that
tends to support our proposal of the relationship of geological uplift and biological
diversification.
Like the Asian monsoon formation, another striking result of QTP intense uplifting, is
the enhanced aridity in Asian interior land since Pliocene 3.6 Ma (Li et al., 1979, 1998;
Zhong & Jing, 1996; Cheng et al., 2000). The most strongly influenced area is
northwestern China, located just north of the plateau. To annotate this environment and
ecological process, section Robusti with A. hoantchy and A. dshimensis of subgenus
Pogonophace, and its estimated age, should be a well performer. These two species are
located at north of the plateau, Xinjiang and western Inner Mongolia provinces in China

17
(Zhang, 2000), and had an estimated age 2.50 in PL (1.53 Ma in PAML)Ma, later than
the intense time of plateau uplift of 3.6 Ma. So, it could be regarded as one of arid-
adapted lineages in Astragalus resulting from the plateau intense uplift.
Acknowledgments
Thanks to Peter W. Fritsch (San Francisco, CAS, USA), Joachim W. Kadereit (Mainz,
Mainz University, Germany), and Gordon C. Tucker (Eastern Illinois University, USA)
for their valuable comments to manuscript, especially G.C. Tucker’ English improvement.
Thanks also to Q.B. Wang and B.J. Zhong (Fudan Unversity, Shanghai) for their
calculating helps. Funding from the Chinese Academy of Sciences (KSCX2-YW-2-
069),National Natural Science Foundation of China (NSFC 30500035), and China
Scholarship Council (CSC) supported M.L. Zhang’s visit to the CAS for this study.
References Anderson C L, Bremer K, Friis E M (2005). Dating phyogenetically basal eudicots using rbcL
sequences and multiple fossil reference points. Amer J Bot, 92(10):1737-1748
Bell C D, Donoghue M J (2005). Dating the Dipsacales: comparing models, genes, and evolutionary
implications. Amer J Bot, 92(2):284-296
Bremer K, Friis E M, Bremer B (2004). Molecular phylogenetic dating of Asterid flowering plants
shows Early Cretaceous diversification. Syst Biol, 53(3):496-505
Bunge A (1868). Astragali. Memorial Academy Science Petersberg. 11(16):1-140
Bunge A (1869). Astragali, specierum enumeratio. Memorial Academy Science Petersberg. 15(1):1-
140
Cheng H B, Powell C, An Z S, Zhou J, Dong G R (2000). Pliocene uplift of the northern Tibetan
Plateau. Geology, 28:715-718
Coleman M, Hodges K (1995). Evidence for Tibetan Plateau uplift before 14 Myr age from new
minimum estimate for east-west extension. Nature, 374:49-52

18
Dong T T X, Ma X Q, Clarke C, Song Z H, Ji Z N, Lo C K, Karl W. K. Tsim K W K (2003).
Phylogeny of Astragalus in China: molecular evidence from the DNA sequences of 5S rRNA spacer,
ITS, and 18S rRNA. J Agric Food Chem, 51:6709-6714
Drummond AJ, Rambaut A (2007). ‘BEAST: Bayesian evolutionary analysis by sampling trees’.
BMC Evolutionary Biology, 7:214
Felsenstein J (1985). Confidence limits on phylogenies: an approach using the bootstrap. Evolution,
39:783–791
Fu K T, He S B, He Y C (1993). Astragalus. In Flora Reipublicae Popularis Sinicae. Vol. 42, No.1.
Beijing: Science Press, 80-114
Guo S X, Zhou Z K (1992). The mega fossil legumes from China. In: Advances in legume
systematics, part 4. Herendeen P S & Dilcher D L, eds. Kew: Royal Botanic Gardens, 303-316
Hall, T (1999). BioEdit: a user-grendly biological sequence alinment editor and analysis program for
Windows 95/98/NT. Nucl Acids Symp, Ser. 41:95-98
Harrison T M, Coplend P, Kidd W S F, Yin A (1992). Raising Tibet. Science, 255:1663-1670
He D K, Chen Y F, Chen Y Y, Chen Z M (2004). Molecular phylogeny of the specialized
schizothoracine fishes (Teleostei: Cyprinidae), with their implications for the uplift of the Qinghai-
Tibetan Plateau. Chinese Science Bulletin, 48(22):2354-2362
Heredeen P S, Jocobs B F (2000). Fossil legumes from the Middle Eocene (46.0 Ma) Maheng flora
of Singda, Tanzania. Amer J Bot, 87:1358-1366
Herendeen P S, Crepet W L, Dilcher D L (1992). The fossil history of the Leguminosae:
phylogenetic and biogeographic implications. In: Advances in legume systematics, part 4.
Herendeen P S & Dilcher D L, eds. Kew: Royal Botanic Gardens, 303-316
Huelsenbeck J P, Ronquist F (2001). MrBayes: Bayesian inference of phylogeny: Bioinformatics,
17:754–755
Institute of Botany, Nanjing Institute of Geology and Palaeontology, Academia Sinica (1978).
Chinese plant fossils, Vol.3, Neogene Floras. Beijing: Academic Press
Jeanmougin F, Thompson J D, Gibson T J, Gouy M, Higgins D G (1998). Multiple sequence
alignment with Clustal X. Trends in Biochem Sci, 23: 403-405
Kajita T, Ohashi H, Tateishi Y, Bailey C D, Doyle J J (2001). rbcL and legume phylogeny, with
particular reference to Phaseoleae, Millettieae, and allies. Syst Bot, 26:515–536
Kang Y, Zhang M L (2003). Study of pollen brush in selected species of Astragalus L. subgenus
Pogonophace Bunge (Leguminosae). Plant Syst Evol, 249(1-2):1-8
Kang Y, Zhang M L, Chen Z D (2002). A preliminary phylogenetic study of the Subgenus
Pogonophace (Astragalus) in China based on ITS sequence data. Acta Botanica Sinica, 45(2):140-
145

19
Kazempour O S, Maassoumi A A, Murakami N (2003). Molecular systematics of the genus
Astragalus L. (Fabaceae): phylogenetic analyses of nuclear ribosomal DNA internal transcribed
spacers and chloroplast gene ndhF sequences. Plant Syst Evol, 242:1-32
Kazempour O S, Maassoumi A A, Murakami N (2005). Molecular systematics of the Old World
Astragalus L. (Fabaceae) as inferred from nrDNA ITS sequence data: Brittonia, 57:367-381
Langley C H, Fitch W (1974). An estimation of the constancy of the rate molecular evolutiojn. J Mol
Evol, 3:161-177
Lavin M, Shrire B P, Lewis G, Pennington RT, Delgado-Salinas A, Thulin M, Hughes C E,
Matos A B, Wojciechowski M F (2004). Metacommunity process rather than continental tectonic
history better explains geographically structured phylogenies in legumes. Phil Trans R Soc Lond B,
359:1509-1522
Lavin M, Herendeen PS, Wojciechowski M F (2005). Evolutionary rates analysis of Leguminosae
implicates a rapid diversification of lineages during the Tertiary: Syst Biol, 54:575–594
Lewis GP, Schrire BD, Mackinder BA, Lock M. (2005). Legumes of the world. Kew: Royal
Botanic Gardens
Li J J, Fang X M (1998). Research on the uplift of the Qinghai-Xizang Plateau and environmental
changes. Chinese Science Bulletin, 43(15):1569-1574
Li J J, Wen S X, Zhang Q S, Wang F B, Zheng B X, Li B Y (1979). A discussion on the period,
amplitude and type of the uplift of the Qinghai-Xizang Plateau. Science in China, Ser B, 6:608-616
Li X X (eds.) 1995. Fossil floras of China through the geological ages. Guangzhou: Guangdong
Scientific and Technology Press
Liston A, Wheeler J A (1994). The phylogenetic position of the genus Astragalus (Fabaceae):
evidence from the chloroplast genus rpoC1 and rpoC2. Biochem Syst Ecol, 22(4): 377-388.
Liu J Q, Gao T G, Chen Z D, Lu A M (2002). Molecular phylogeny and biogeographyof the
Qinghai-Tibet Plateau endemic Nannoglottis (Asteraceae). Mol Phylogen Evol, 23:307-325
Liu J Q, Wang Y J, Wang A L, Hedeaki O, Abbott R (2006). Radiation and diversification within
the Ligularia-Cremanthodium-Parasenecio complex (Asteraceae) triggered by uplift of the Qinghai-
Tibetan Plateau. Mol Phylogen Evol, 38:31-49
Lock JM, Schrire BD (2005). Tribe Galegeae. In: Legumes of the world. Lewis G, Shrive B,
Mackinder B, Lock M, eds. Kew: Royal Botanical Garden, 475-488
Molnar P, England P, Martiod J (1993). Mantle dynamics, uplift of the Tibetan Plateau and the
Indian monsoon development. Rev Geophys, 34:357-396
Near TJ, Sanderson M J (2004). Assessing the quality of molecular divergence time estimating by
fossil calibration and fossil-based model selection. Phil Trans R Soc Lond B, 359:1477-1483

20
Podlech D (1999). Thesaurus Astralorum 1. Index of all taxa within the genus Astragalus and other
genera belonging to the Astragalus. Taxa of the Old World and related taxa of the New World. URL:
http://www.botanik.biologie.uni-muenchen.de/botsyst /thesaur1. html
Polhill R M (1981). Galegeae. In: Advances in legume systematics, part 1. Polhill R M, Raven P H,
eds. Kew: Royal Botanical Gardens, 357-363
Posada D, Buckley T (2004). Model selection and model averaging in phylogenetics: advantages of
Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst Biol, 53:793-
808
Posada D, Crandall K A (1998). Modeltest: testing the model of DNA substitution: Bioinformatics,
14 817–818
Rannala B, Yang Z (2007). Inferring speciation times under an episodic molecular clock. Syst Biol,
56:453-466
Renner S S, Zhang L B (2004). Biogeography of the Pistia clade (Araceaae):based on chloroplast
and mitochondrial DNA sequences and Bayesian divergence time inference. Syst Biol, 53:422-432
Renner S S, Givnish T J (eds.) (2004). Tropical intercontinental disjuctions. Int Jour Plant Sci, 165(4
Suppl.):S1-S138
Sanderson M J (1991). Phylogenetic relationships within North American Astragalus (Fabaceae).
Syst Bot, 16: 414-430
Sanderson M J (1997). A nonparametric approach to estimating divergence times in the absence of
rate constancy. Mol Biol Evol, 14:1218-1231
Sanderson M J (2002). Estimating absolute rates of molecular evolution and divergence times: a
penalized likelihood approach. Mol Biol Evol, 19:101-109
Sanderson M J (2003). r8s: inferring absolute rates of molecular evolution and divergence times in
the absence of a molecular clock. Bioinformatics, 19:301-302
Sanderson M J, Liston A (1995). Molecular phylogenetic systematics of Galegeae, with special
reference to Astragalus. In: Advances in legume systematics, part 7, phylogeny. Crisp M D, Doyle J
J, eds. Kew: Royal Botanical Gardens, 331-350
Sanderson M J, Wojciechowski M F (1996). Diversification rates in a temperate legume clade: are
there “so many species” of Astragalus (Fabaceae)? Amer J Bot, 83(11):1488-1502
Scherson R A, Vidal R, Sanderson M J (2008). Phylogeny, biogeography, and rates of
diversification of New World Astragalus (Leguminosae) with an emphasis on South American
radiations. Amer J Bot, 95:1030-1039
Spicer R A, Harris N B W, Widdowson M, Herman A B, Guo S X, Valdes P J, Wolfe J A, Kelly
S P (2003). Constant elevation of southern Tibet over the past 15 million years. Nature, 421:622-
624

21
Swofford D L (2002). PAUP, phylogenetic analysis using parsimony, version 4. Sinauer Associates,
Sunderland, Massachusetts
Tao J R, Zhou Z K, Liu Y S (2000). The evolution of the Late Cretaceous-Cenozonic floras in China.
Beijing: Science Press
Tao J R (1992). The Tertiary vegetation and flora and floristic regions in China. Acta Phytotaxon
Sinica, 31(1):25-43
Wenninger J (1991). Revision von Astragalus L. sect. Chlorostachys Bunge, sect. Phyllolobium
Bunge und sect. Skythropos Bunge (Leguminosae). Mitteilungen der botanischen Staatssammlung
Munich, 30:1-196
Wojciechowski M F (2003). Reconstructing the phylogeny of legumes (Fabaceae): an early 21st
century perspective. In: Advances in Legume Systematics, part 10. Klitgaard, BB, Bruneau A, eds.
Kew: Royal Botanical Garden, 5-35
Wojciechowski M F (2005). Astragalus (Fabaceae): a molecular phylogenetic perspective. Brittonia,
57(4):382–396
Wojciechowski M F, Lavin M, Sanderson M J (2004). A phylogeny of legumes (Leguminosae)
based on the analysis of the plastid matK gene resolves many well-supported subclades within the
family. Amer J Bot, 91:1846-1862
Wojciechowski M F, Sanderson M J, Hu J M (1999). Evidence on the monophyly of Astragalus
(Fabaceae) and its major subgroups based on nuclear ribosomal DNA ITS and chloroplast DNA trnL
intron data. Syst Bot, 24(3): 409-437
Wu Z Y, Wu S G (1996). A proposal for new floristic kingdom (realm). In: Floristic characteristics
and diversity of Eastern Asian Plnats. Zhang A L, Wu S G, eds. Beijing:China Higher Education
Press, Hongkong:Springer-Verlag
Wu Z Y (1988). Hengduan Mountain flora and her significance. J Japan Bot, 63:297-311
Yang Z (2007) PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol Biol Evol,
2007 24(8):1586-1591
Zhang M L (2000). Studies on geographical distribution pattern of the Subgenus Pogonophace
(Fabaceae: Astragalus) in China using GIS technique. Acta Botanica Sinica, 42(8): 849-854
Zhang M L (2002). Systematics of Astragalus Subgenus Pogonophace (Leguminosae). Acta Botanica
Yunnanica, 24(5):543-553
Zhang M L (2003). Biogeography of Astragalus Subgenus Pogonophace (Leguminosae). Acta
Botanica Yunnanica, 25(1):25-32
Zhang M L, Podlech D (2006). Revision of the Genus Phyllolobium Fisch. (Leguminosae-
Papilionoideae). Feddes Repertorium, 117(1-2):41-64

22
Zhong D L, Jing L (1996). A discussion on the uplift process and mechanism of Tibetan Plateau.
Science in China, Ser D, 26(4):289-295
Appendix. Taxa samples, first five marking “*” are new reports of the ITS 5.8S from this paper,
the rest are downloaded from the GenBank. Total 101 species are listed.
*A.angustifoliolatus K.T. Fu Lijiang, Yunnna, China, Zhu S.X., Kang Y., No.43-2 (PE) FJ236887
*A.campdontoides Simps. Eryuan, Yunnan, Zhu S.X., Kang Y., No.39 (PE) FJ236888
*A.camptodontus Franch. Zhongdian, Yunnan, China, Zhu S.X., Kang Y., No.64 (PE) FJ236899
*A.flavovirens K.T. Fu Zhongdian, Yunnan, China, Zhu S.X., Kang Y., No.61 (PE) FJ236898
*A.pastorius Tsai et Yu Xiangchen, Sichuan,China, Zhu S.X., Kang Y., No.47 (PE) FJ236892
Astragalus adsurgens Pall. AF121674; A.aksuensis Bunge AF359753; A.alpinus L. ITS1 L10760+
ITS2 L10761; A.balfourianus Simps. AF521951; A.chinesis L. f. AF121681; A.complanatus R. Br. ex
Bunge AF521950; A.cysticalyx Ledeb. AF121682; A.dshimensis Gontsch. AF359755; A.hamosus L.
ITS1 L10778+ ITS2 L10779; A.hendersonii Baker AF521957; A.hoantchy Franch. AF521952;
A.lehmannianus Bunge AF359756; A.lepsensis Bunge AF359752; A.membranaceus Bunge
AF359749; A.milingensis Ni et P.C. Li AF521954; A.mongolicus Bunge AF359750;
A.nankotaizanensis Sasaki AF121680; A.polycladus Bur. et Franch. AF121676; A.propinquus B.
Schischk. AF359751; A.sinicus L. ITS1 U50502+ ITS2 U50503; A.tanguticus Batalin AF521956;
A.tribulifolius Benth. ex Bunge AF521953; A.yatungensis Ni et P.C. Li AF521955;
Acacia farnesiana Wall. AF360728; Ac.mearnsi De Will.AF360750; Ac.senegal AF360727;
Calophaca tianshanica (B. Fedtsch.) Boris. ITS1 U51220+ ITS2 U51221;
Cararagana korshinskii Kom. AY626914; Car.microphylla Lam. AY626915; Car.roboroviskyi Kom.
AF521958; Car.sibirica Fabr. (=Car. arborescens Lam.) AY626912;
Cercis canadensis L. AF390188; Ce.chinensis Bunge AF286351; Ce.chingii Chun AF286350;
Ce.chuniana Metcalf AF286349; Ce.gigantea AF390194; Ce.occidentalis Torr. AF286352;
Ce.racemosa Oliver AF288782; Ce.siliquastrum L. AF286353;
Colutea arborescens L. ITS1 U56009+ ITS2 U56010; Co.istria Mill. ITS1 U69544+ ITS2 U69545
Dalbergia congestiflora Pittier AF068140; D.foliolosa Benth. AF189002; D.sissoo Roxb. AF189023;
Hedysarum aculeolatum Michx AY772222; H.carnosulum Greene AY772224; H.coronarium L.
AY772225
H.flexuosum L. AY775312; H.humile L. AY772227; H.membranaceum Coss. et Bal. AY772228;
H.pallidum Desf. AY772229; H.spinosissimum L. subsp. capitatum AY772223;
Oxytropis besseyi (Rydb.) Blank. var. ventosa (Greene) Barneby AF121756; O.lamberti Pursh
AF121753; O.multiceps Nutt. AF121760; O.oreophila A. Gray AF121755; O.pilosa DC. AF121759;

23
O.sericea Schur AF121757; O.splendens Douql. AF121761; O.szovitsii Boiss. et BuhseAF121754;
O.viscida Nutt. ex Torr. et GrayAF121758
Pueraria lobata (Willd.) Ohwi (=P. montana var. lobata) AF338215; P.montana (Lour.) Merr.
AF338216; P.thomsonii Benth. (=P. montana var. thomsonii) AF338217;
Sophora affinis Torr. et Gary U59886; S.cassioides (Phil.) Sparre AY056081; S.chathamica Cockayne
AY046513; S.chrysophylla Seem. AY056070; S.davidii Kom. ex Pavol. AF467496; S.denudata Bory
AY056071; S.flavescens Ait. AF123452; S.fulvida (Allan) Heenan & de Lange AY056072; S.godleyi
Heenan & de Lange AY056073; S.howinsula (W. R. B. Oliver) P.S. Green AY046514; S.microphylla
Ait. var. longicarinata AY056074; S.molloyi Heenan & de Lange AY056076; S.prostrata Buchanan
AY056077; S.raivavaeensis H.St. John AY056080; S.secundiflora Laq. ex DC. AF174638;
S.tetraphylla (ex Ives) A. Gary AJ310734; S.tetraptera J.S. Mill. AY056078; S.tomentosa L.
AY725482; S.toromira (R.A. Phil.) Skottsb. ST409921;
Sphaerophysa salsula DC. ITS1 U560011+ITS2 U560012;
Swainsona campylantha AF113855; Sw.cyclocarpa AF113856; Sw.decurrens AF113857
Sw.formosa F. Muell. AF113859; Sw.kingii F. Muell. AF113860; Sw.murrayana Wawra AF113861;
Sw.oroboides F. Muell. ex Benth. AF113862; Sw.parviflora Benth. AF113863; Sw.purpurea (A.T.
Lee) Joy Thomps. AF113864; Sw.stenodonta F. Muell. AF113865; Sw.swainsonioides AF113866;
Sw.villosa AF113867.
Fig. 1 Phylogenetic trees reconstruction were used three approaches, maximum
parsimony, maximum likelihood and Bayesian inference. Three phylogenetic analyses
resulted in almost identical topology of trees, especially at the generic level. The clades
were with high bootstrap to parsimony and posterior probability values to Bayesian,
most over 90% bootstrap and 95% posterior probability respectively. A Bayesian
inference tree Fig. 1 is used to show the phylogenetic relationships of legumes taxa and
circles at the nodes shown 95% posterior probability.
Fig. 2 Chronogram yielded by dating PL in r8s, focusing on Astragalus and relatives, the
estimated ages of the related taxa in r8s, PAML and BEAST refer to Table 2.

0.1
Acacia mearnsiiAc.farnesianaAc.senegal
Cercis giganteaCe.racemosa Ce.chinensis Ce.chingii Ce.chuniana Ce.canadensis Ce.occidentalis Ce.siliquastrum
Astragalus lehmannianusA.chinesis A.hoantchy A.dshimensis A.propinqus A.membranaceus A.mongolicusA.aksuensis A.lepsensis A.hamosus A.adsurgens
A.nankotaizanens A.alpinus A.polycladus
A.milingensis A.tanguticus A.tribulifoliusA.pastorius A.camptodontoides A.angustifoliolatusA.camptodontus A.balfourianusA.flavovirens
A.complanatusA.sinicus
A.hendersoniiA.yatungensisSwainsona swainsoniodesSw.murrayanaSw.purpurea Sw.parviflora Sw.campylantha
Sw.villosa Sw.oroboides Sw.formosa Sw.decurrens Sw.stenodonta Sw.cyclocarpa Swainsona kingii Sphaerophysa salsula Astragalus cysticalyxColutea istriaCo.arborescens
Oxytropis multiceps O.besseyi O.pilosa O.szovitsii O.splendens O.oreophila O.lambertii O.sericea O.viscida Hedysarum membranceum
H.spinosissimumH.pallidum H.humile H.aculeolatum H.carnosum H.coronarium Hedysarum flexuosum
Caragana roborovskii Calophaca tianshanicaCaragana microphylla Car.sibirica Car.korshinskii
Pueraria montana P.thomsonii P.lobata
Sophora affinis S.secundiflora S.flavescens
S.davidii S.raivavaeensis S.fulvida S.godleyi S.prostrata S.tetraptera
S.tomentosa S.chathamica S.howinsu S.tetraphylla S.toromira S.cassioides S.denudata S.molloyi S.chrysophylla S.microphylla
Dalbergia foliolosa D.congestiflora D.sissoo
o
o
o
o
o
o o
o
o
o
o
o
o
o
o
o
o
o
o
o o
o
o
o
o
o
o
o
o
1
A.lehmannianus A.chinesis A.hoantchy A.dshimensis A.propinqus A.membranaceus A.mongolicus A.aksuensis A.lepsensis A.hamosus A.adsurgens A.nankotaizanens A.alpinus A.polycladus Phyllolobium milingenum Ph.tanguticum Ph..tribulifolium Ph.pastorium Ph.campdontoides Ph.angustifoliolatum Ph.camptodontum Ph.balfourianum Ph.flavovirens Ph.chinese Ph.sinicus Ph.hendersonii Ph.donicumSwainsona swainsoniodes Sw.murrayana Sw.purpurea Sw.parviflora Sw.campylantha Sw.villosa Sw.oroboides Sw.formosa Sw.decurrens Sw.stenodonta Sw.cyclocarpa Sw.kingii Sphaerophysa salsula Astragalus cysticalyx Colutea istria Co.arborenscens Oxytropis multiceps O.besseyi O.polosa O.szovitsii O.splendens O.oreophila O.lambertii O.sericea O.viscida Hedysarum membranaceum H.spinosissimum H.pallidum H.humile H.aculeolatum H.carnosum H.coronarium H.flexuosum Caragana roborovskii Calophaca tianshanica Caragana microphylla Car.sibirica Car.korshinskii
25 10 15 0 5 20 30
subgen. Calycophysa
subgen. Astragalus
subgen. Pogonophace
subgen. Cercidoothrix
Phyllolobium
subgen. Phaca
subgen. Astragalus
1
2
11
6 9
5
7
8
13 12
4
3
10
Myr
Fig. 2




















