
A Bacterium That Can Grow by Using Arsenic Instead of Phosphorus

Phenotypic Switching in Biofilm-Forming Marine BacteriumPaenibacillus lautus NE3B01
Neelam Mangwani • Supriya Kumari •
Sudhir K. Shukla • T. S. Rao • Surajit Das
Received: 18 November 2013 / Accepted: 28 November 2013 / Published online: 23 January 2014
� Springer Science+Business Media New York 2014
Abstract Biofilm-forming marine bacterium Paenibacillus
lautus NE3B01 was isolated from a mangrove ecosystem,
Odisha, India. This isolate formed a swarming type of colony
pattern on the solid culture medium with 0.5–2 % agar. Phase
contrast microscopy study of a growing colony of P. lautus on
solid media and swarming pattern revealed the existence of
two phenotypically distinct cells (i.e. cocci and rods) across
the colonies. However, in actively growing planktonic cul-
ture, only rod-shaped cells were observed. Biofilm growth
studies (crystal violet assay) with the isolate showed signifi-
cant biofilm formation by 6 h, and the detachment phase was
observed after 18 h. Biofilm parameters (such as total bio-
mass, roughness coefficient, biofilm thickness, etc.) of 24-h-
old P. lautus biofilm were studied by confocal scanning laser
microscopy (CSLM). The CSLM study showed that P. lautus
formed a biofilm with an average thickness of 14.8 ± 2.6 lm,
a high roughness coefficient (0.379 ± 0.103) and surface to
bio-volume ratio (4.59 ± 1.12 lm2/lm3), indicating a highly
uneven topography of the biofilm. This also indicates that the
24-h-old biofilm is in dispersal phase. Scanning electron
microphotographs of P. lautus also supported the existence of
two distinct phenotypes of P. lautus. The current findings
suggest that P. lautus has two vegetative phenotypes and to
decongest the overcrowded biofilm the bacterium can switch
over to motile rods from nonmotile cocci and vice versa.
Introduction
Bacteria tend to colonize when they encounter a solid
surface in aquatic environments. Colonization is a collec-
tive behaviour of bacterial cells that result in a swarming
pattern of growth and biofilm formation [32]. Multiple
factors together with intercellular and intracellular signal-
ling, extracellular polymers, cell surface appendages,
nutrition and environmental sensing are imperative aspects
that determine the spatial arrangement of cells in bacterial
colonies and biofilms [3, 4, 14]. Biofilms are ubiquitous in
environmental systems particularly in freshwater and
marine ecosystems [5]. Biofilm is a multicellular sessile
community of bacteria attached to a substratum and shel-
tered in a milieu of self-synthesised extracellular polymeric
substances [19, 20]. Apart from bacterial biofilms, the other
social aggregation that has received a great deal of interest
amongst researchers is the swarming motility. Many bac-
terial species undergo rapid and coordinated translocation
of their population across solid and semi-solid surface and
exhibit some intricate growth pattern known as swarming.
The swarming pattern formation represents multi-cellular
structure that provides protection and cooperation amongst
cells [9]. Swarming motility is a diffusion mechanism that
generates macroscopic patterns, which has been widely
studied in Pseudomonas, Bacillus, Vibrio, Escherichia,
Salmonella and Proteus [18]. In the recent time, some
Paenibacillus species (Paenibacillus vortex and P. den-
dritiformis) have been studied for their complex swarming
behaviour [1, 14].
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00284-014-0525-8) contains supplementarymaterial, which is available to authorized users.
N. Mangwani � S. Kumari � S. Das (&)
Laboratory of Environmental Microbiology and Ecology
(LEnME), Department of Life Science, National Institute of
Technology, Rourkela 769 008, Odisha, India
e-mail: [email protected]; [email protected]
S. K. Shukla � T. S. Rao
Biofouling and Biofilm Processes Section, Water & Steam
Chemistry Division, BARC Facilities,
Kalpakkam 603 102, Tamil Nadu, India
123
Curr Microbiol (2014) 68:648–656
DOI 10.1007/s00284-014-0525-8

The strategies for survival (e.g. endospores and canni-
balism) ensure bacterial population protection when
encountered with intense environmental conditions or
nutrient stress [8, 11]. In some cases, bacteria also adopt
phenotypic heterogeneity (or bistability) [7]. Bacteria
multiply by cell division that yields genetically and mor-
phologically identical siblings. Conversely, it is also well
established that within an isogenic population there may be
a regulatory diversification. As a result, bacterial cells can
display various phenotypes. The phenotypic variation can
benefit bacteria, as far as their growth and survival are
concerned, with changing environmental conditions [10]. A
recent study by Be’er et al. [1] reported similar phenom-
enon in P. dendrtifoirmis colonies.
In the present study, we have used a combinational
microscopic approach to describe a cellular heterogeneity
and phenotypic switching in P. lautus micro-colony as well
as in biofilm. The bacterium showed to switch from non-
motile cocci to motile rods when overcrowding occurs in
the colony. The motile rods can switch back to cocci on
finding a nutrient-rich and open surface. To the best of our
knowledge, this is the first report on biofilm population
heterogeneity wherein P. lautus cells are coccoid in shape
in the centre of biofilm and at the periphery the cells are
transformed into motile rods and migrate to avoid
overcrowding.
Materials and Methods
Isolation and Characterization
Paenibacillus lautus NE3B01 was isolated from marine
water sample collected from Bhitarkanika mangrove eco-
system, Odisha, India. The strain was identified by 16S
rRNA gene sequencing, and phylogenetic tree was con-
structed following Dash et al. [5]. The culture was main-
tained in Luria–Bertani (LB) broth, and all the experiments
were performed in LB with 0.1 % of NaCl. The strain has
been submitted to Microbial Type Culture Collection and
Gene Bank, India (MTCC 11807).
Biofilm Screening and Growth Studies
Screening for biofilm formation was carried out by glass
tube assay following O’Toole et al. [24] and Jain et al. [16].
The tubes with attached cell mass visible after crystal
violet staining were considered as biofilm positive. Biofilm
growth of the isolate was monitored for 48 h using
microtiter plate assay. A colony was inoculated in LB broth
and incubated at 37 �C overnight. The strain was grown in
LB broth (37 �C/160 rpm); at log phase, cell mass was
harvested and washed with phosphate-buffered saline
(PBS) twice. It was resuspended in LB broth, and OD600nm
was adjusted to 0.5. 10 ll of this inoculum was transferred
to microtiter plate well with 190 ll of LB medium. At
regular time interval, the plate was washed with water
twice, air dried and stained with 0.2 % crystal violet for
5 min. Again, it was washed twice with water, air dried and
destained with 95 % ethanol for 30 min and OD595nm was
measured using 96 well plate reader (Multiplate reader,
PerkinElmer).
Swarming Motility
Swarming motility was evaluated by seeding 10 ll of
stationary phase culture mounted at the centre of 8-cm
diameter motility plates with 0.5, 1, 1.5 and 2 % LB agar
and incubated at 37 �C for 48–72 h. Swarm plates were
typically allowed to dry at room temperature under laminar
hood for few hours before being used. The swarming pat-
tern was observed under a transwhite illumination cham-
ber. 1.5 % LB agar swarming plate, for general cell
morphology and colony swarming pattern were observed
under inverted fluorescence microscope equipped with 40X
phase contrast objective lens (Olympus, 1 9 71)
Microscopy of Biofilm
A single colony was inoculated in LB broth and incubated at
37 �C, and the overnight grown culture was diluted 1:100 in
LB broth. 3 ml of the diluted suspension was transferred to
six-well cell culture plates. A sterile glass slide
(2 cm 9 2 cm) was placed in each well in order to grow the
biofilm. The plate was incubated at 37 �C for 24 h. After 24 h,
the slides were removed, gently washed with PBS and then
stained with 0.02 % acridine orange for 5 min and washed
again with PBS. Confocal scanning laser microscopy (CSLM)
studies were performed to monitor the biofilm architecture. A
thin cover slip was placed over the biofilms and mounted
upside down over the objective lens of CSLM (TCS-SP2-
AOBS) equipped with DM IRE 2-inverted microscope (Leica
Microsystems, Hessen, Wetzlar, Germany). About 10 image
stacks were collected randomly from different points in order
to get reliable data. Biofilm parameters were assayed as
described in Mangwani et al. [20] by analysing the image
stacks using COMSTAT [13]. Scanning electron microscopy
(SEM) of the P. lautus biofilm was carried by growing the
biofilms grown on glass slides (1 9 1 cm). The biofilm-
grown slides were fixed with 2.5 % glutaraldehyde (in PBS,
pH 7.2) at 4 �C for 12 h, washed with PBS and post fixed with
1 % aqueous solution of tannic acid. Later, the specimens
were dehydrated with 30, 70 and 100 % alcohol and coated
with platinum. The samples were analysed using scanning
electron microscope (Jeol T-330 Scanning Electron Micro-
scope, Germany). The planktonic cells were also stained with
N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus 649
123
0.2 % crystal violet, destained with 95 % ethanol for light
microscopy studies and observed under oil immersion lens
(1009).
Results and Discussion
Isolation and Characterization
The partial 1335 bp 16S rRNA gene sequence of the isolate
NE3B01 was submitted to NCBI GenBank under accession
number JX273779. The 16S rRNA gene analysis indicated
that the strain is phylogenetically related to genus Paeniba-
cillus. The sequence alignment was carried out using
CLUSTAL W tool in MEGA version 5. A phylogenetic tree
was constructed by the Neighbour Joining method based on
16S rRNA partial gene sequence (Supplementary Fig. S1).
The selected isolate was designated as P. lautus NE3B01.
Biofilm Screening and Growth Studies
Screening for biofilm formation was done by visualization of
attached cell mass to the glass tube visible after crystal violet
staining (Supplementary Fig. S2a). Biofilm is a surface-
associated growth, composed of cells and EPS that gets
stained by crystal violet [12]. Biofilm could easily be visually
seen after destaining with 95 % ethanol. Biofilm growth
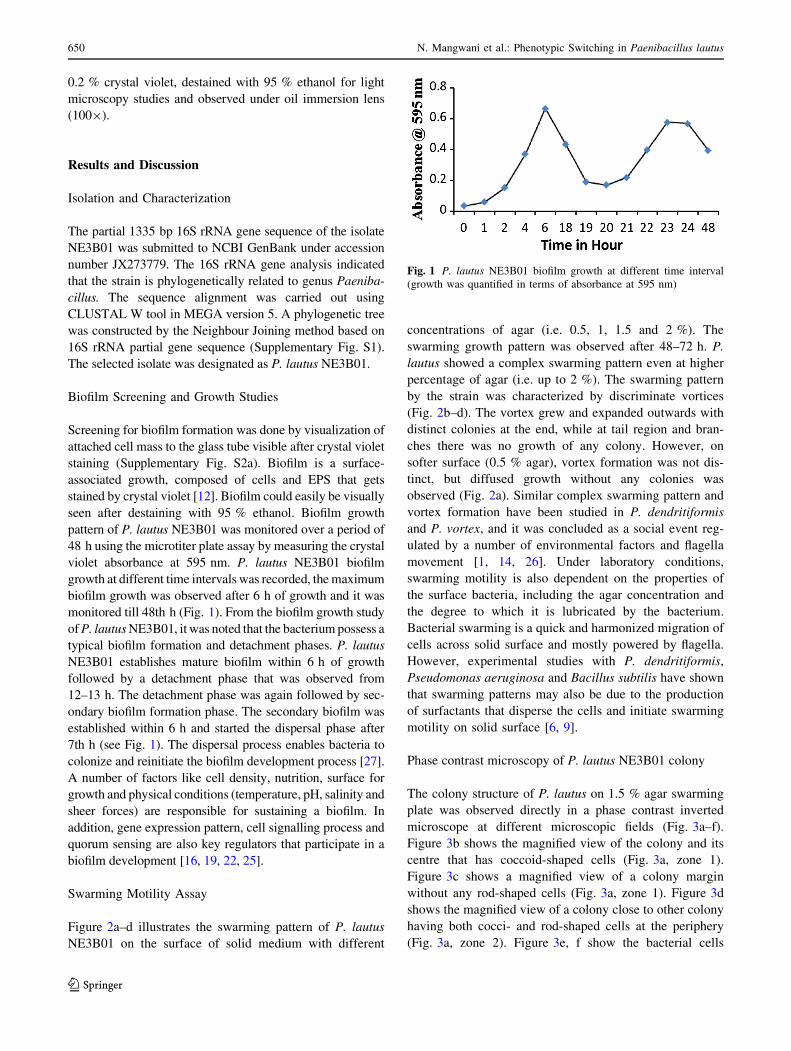
pattern of P. lautus NE3B01 was monitored over a period of
48 h using the microtiter plate assay by measuring the crystal
violet absorbance at 595 nm. P. lautus NE3B01 biofilm
growth at different time intervals was recorded, the maximum
biofilm growth was observed after 6 h of growth and it was
monitored till 48th h (Fig. 1). From the biofilm growth study
of P. lautus NE3B01, it was noted that the bacterium possess a
typical biofilm formation and detachment phases. P. lautus
NE3B01 establishes mature biofilm within 6 h of growth
followed by a detachment phase that was observed from
12–13 h. The detachment phase was again followed by sec-
ondary biofilm formation phase. The secondary biofilm was
established within 6 h and started the dispersal phase after
7th h (see Fig. 1). The dispersal process enables bacteria to
colonize and reinitiate the biofilm development process [27].
A number of factors like cell density, nutrition, surface for
growth and physical conditions (temperature, pH, salinity and
sheer forces) are responsible for sustaining a biofilm. In
addition, gene expression pattern, cell signalling process and
quorum sensing are also key regulators that participate in a
biofilm development [16, 19, 22, 25].
Swarming Motility Assay
Figure 2a–d illustrates the swarming pattern of P. lautus
NE3B01 on the surface of solid medium with different
concentrations of agar (i.e. 0.5, 1, 1.5 and 2 %). The
swarming growth pattern was observed after 48–72 h. P.
lautus showed a complex swarming pattern even at higher
percentage of agar (i.e. up to 2 %). The swarming pattern
by the strain was characterized by discriminate vortices
(Fig. 2b–d). The vortex grew and expanded outwards with
distinct colonies at the end, while at tail region and bran-
ches there was no growth of any colony. However, on
softer surface (0.5 % agar), vortex formation was not dis-
tinct, but diffused growth without any colonies was
observed (Fig. 2a). Similar complex swarming pattern and
vortex formation have been studied in P. dendritiformis
and P. vortex, and it was concluded as a social event reg-
ulated by a number of environmental factors and flagella
movement [1, 14, 26]. Under laboratory conditions,
swarming motility is also dependent on the properties of
the surface bacteria, including the agar concentration and
the degree to which it is lubricated by the bacterium.
Bacterial swarming is a quick and harmonized migration of
cells across solid surface and mostly powered by flagella.
However, experimental studies with P. dendritiformis,
Pseudomonas aeruginosa and Bacillus subtilis have shown
that swarming patterns may also be due to the production
of surfactants that disperse the cells and initiate swarming
motility on solid surface [6, 9].
Phase contrast microscopy of P. lautus NE3B01 colony
The colony structure of P. lautus on 1.5 % agar swarming
plate was observed directly in a phase contrast inverted
microscope at different microscopic fields (Fig. 3a–f).
Figure 3b shows the magnified view of the colony and its
centre that has coccoid-shaped cells (Fig. 3a, zone 1).
Figure 3c shows a magnified view of a colony margin
without any rod-shaped cells (Fig. 3a, zone 1). Figure 3d
shows the magnified view of a colony close to other colony
having both cocci- and rod-shaped cells at the periphery
(Fig. 3a, zone 2). Figure 3e, f show the bacterial cells
Fig. 1 P. lautus NE3B01 biofilm growth at different time interval
(growth was quantified in terms of absorbance at 595 nm)
650 N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus
123

between colonies. Only rod-shaped cell is visible in the
region where fewer cocci can be seen in Fig. 3f (Fig 3a,
zone 3, 4). From the phase contrast microscopic study, it
was observed that the fraction of rod-shaped population
increased with increasing distance from the colony,
whereas the proportion of cocci was higher within colonies.
These observations suggest the presence of two distinct
phenotypes i.e. motile rods and non-motile cocci when
grown on a solid medium. Figure 3g shows an isolated
colony of P. lautus NE3B01 from which some motile rods
get detached from the colony and form a new colony
nearby (Fig. 3a, zone 5). It can be speculated that P. lautus
possesses a phenotypic switching i.e. transition from cocci
to rod under the condition of overcrowding. The rod gets
switched to cocci again on finding a new settlement surface
nearby where nutrients are sufficient and no competition is
present in close proximity. This speculation corroborates
with the study of Be’er et al. [1] where an inducible and
reversible phenotypic switching phenomenon was descri-
bed as a mechanism of survival by P. dendritiformis due to
overcrowding and sibling colony competition. P. dendrit-
iformis switch between rods and cocci by toxin and sig-
nalling proteins known as Slf and Ris (sibling lethal factor
and rod-inducing signals, respectively), which induce the
cocci to rod shape in response to local environmental
conditions. It might be possible that both P. dendritiformis
and P. lautus have some common regulator network
conserved in Paenibacillus genus responsible for the two
distinct phenotypes. However, from the current study on P.
lautus, the mechanism of transformation appears to be
different from what is reported in P. dendritiformis as no
spores were observed in the present study in the culture of
P. lautus NE3B01.
SEM and CSLM analysis of P. lautus NE3B01 biofilm
The CSLM images (see Fig. 4a, b) show the biofilm growth
of P. lautus NE3B01 on the glass surface after 24 h of
growth at 37 �C. Figure 4a shows a 24-h-old biofilm con-
sisting of many rod-shaped bacteria spaced interstitially
along with the mature micro-colonies. In contrast, Fig. 4b
illustrates a mature 24-h-old biofilm of P. lautus NE3B01
with a maximum biofilm thickness of *21 lm. Figure 3
shows that P. lautus NE3B01 forms a dense biofilm in a
distinct manner, having branched growth at the terminals
with vortices. A number of rod-shape single cells are
clearly visible in Fig. 4a. Table 1 illustrates the COM-
STAT analysis of P. lautus NE3B01 biofilm, which had an
average thickness of 14.82 ± 2.61 lm and maximum
thickness of 21.02 ± 2.10. P. lautus NE3B01 biofilm
showed high roughness coefficient (0.379 ± 0.103) and
surface to biovolume ratio (4.59 ± 1.12 lm2/lm3), indi-
cating either highly uneven topography of biofilm or dis-
persal event [30, 31]. In general, bacterial cells are
Fig. 2 Swarming pattern of P.
lautus NE3B01 at different
concentrations of agar a 0.5 %
agar showing bacterial mat,
b 1 % agar, c 1.5 % agar, d 2 %
agar. At higher concentration of
agar, vortices can be seen.
Diameter of vortices decreases
with increase in agar
percentage, whereas swarming
pattern becomes more distinct at
high concentration of agar
N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus 651
123

compactly entrapped in a biofilm, resulting in loss of
motility [16]. However, CSLM studies with P. lautus
NE3B01 biofilm manifest that a number of individual cells
were present in nearby biofilm (see Fig. 4b), which might
have dispersed from the parent biofilm. This suggests that
there was a detachment of cells resulting from dissociation
of biofilm. Thus, it can be hypothesized that P. lautus
NE3B01 forms a dynamic biofilm community with higher
cell detachment rate as compared to biofilms formed by
other bacteria.
Paenibacillus lautus NE3B01 biofilm and cell mor-
phology were also studied by high resolution SEM at dif-
ferent magnifications. Figure 5a, b show that P. lautus
NE3B01 formed a dense biofilm showing two distinct
phenotypes of cells. Both rod and coccoid cells were
observed in the biofilm. However, the shape of both rod-
and cocci-shaped cells was quite different from their typ-
ical morphology as reported in P. dendritiformis [1]. Some
cocci-shaped cells were more or less like a ring or bicon-
cave disc. The cocci cells were more in number compactly
Fig. 3 a–g Colony and cell morphology of P. lautus NE3B01
observed under phase contrast microscope. a Swarming pattern over
1.5 % agar. b Magnified view of colony and at its centre (a, zone 1).
c Magnified view of a colony margin (a, zone 1). d Two adjacent
colonies having both cocci and rod at the periphery (a, zone 2).
e Bacterial cell between colonies and only rod-shaped cell are visible
(a, zone 3). f At the place fewer cocci can be seen (a, zone 4). g An
isolated colony of P. lautus NE3B01 from which some motile rods get
detached from the colony and forming a new colony nearby (a, zone
5)
652 N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus
123

arranged and attached to the glass surface, while the rod-
shaped cell were present at the periphery of the biofilm.
Some appendages could easily be seen on rod cells
(Fig. 5b); the rod-shaped cell may represent the motile cell
getting detached from the biofilm as was also observed
under phase contrast microscope (See supplementary file
video 1). From the SEM images, it is inferred that phe-
notypic switching occurred in biofilm under overcrowding
condition to facilitate the cells to escape in planktonic form
and allow it to settle at a new place. Phase contrast
microscopy and SEM examination of actively growing
planktonic culture showed abundance of rod-shaped cells
(Fig. 5c, d). These findings support the occurrence of
phenotypic variation and bistable switching between two
phenotypes in P. lautus biofilm. In general, being a clone
population bacterial culture is believed to be homogenous.
However, differential expression of genes is possible
amongst bacterial subpopulations because of some random
fluctuations in transcription and translation [2, 28]. SEM
and phase contrast microscopy results indicate that P.
lautus cells have surface structures that can help in folding
and unfolding of cells that give perception of cocci- and
rod-shaped populations, which are not a truly different cell
population. These structures also help in twitching motility
over hard agar surface (supplementary file, video 1).
Bacteria may exhibit a wide variety of morphologies in
its growth phase; these changes have to be made to
maintain the consistency of a bacterial cell as per its sur-
rounding milieu. However, the consistency of a bacterial
shape could be affected due to environmental stress. To
cope with changing environment, adaptation via pheno-
typic heterogeneity is an imperative biological function.
The pleomorphism phenomenon is well established in
bacteria such as Helicobacter pylori, Deinococcus radio-
durans, Mycobacterium tuberculosis, Bacillus bifidus and
Staphylococcus aureus [21, 23, 33]. For instance, in D.
radiodurans, nutritional stress can alter its phenotype and
radiotolerance [17, 29]. Phenotypic heterogeneity helps to
optimize interactions amongst cells and with the surfaces to
which they attach. Cells in such communities possess
alterations in phenotype with respect to growth and gene
expression [15, 22]. In the present study, the biofilm
formed by the P. lautus showed coccoid cells in the middle
of the micro-colony whereas at the periphery most of the P.
lautus cells were rod shaped and were motile. It is specu-
lated that disc-like cells resembling a doughnut are an
intermediate stage between cocci- and rod-shaped mor-
photypes and transformation from cocci to rod may be the
stage that helps the detachment of cells from the biofilm. A
schematic diagram Fig. 6 explains hypothetically the dif-
ferent stages of P. lautus transformation. The capability of
P. lautus NE3B01 to survive as rod and coccoid morpho-
types comes into sight because of static growth condition
and increased population density. The bacterium acquires
the rod shape to disperse itself from the competitive
overcrowded colony and re-establishes itself on to a virgin
surface. The current findings on coexistence of two sub-
populations of P. lautus provide a distinctive feature of
Fig. 4 a Confocal scanning
laser microscopic image of P.
lautus NE3B01 after acridine
orange staining. Arrow pointing
the dispersing planktonic rod
cells around the biofilm. bMagnified view of P. lautus
NE3B01 biofilm
Table 1 Biofilm parameters of 24-h-old P. lautus NE3B01 biofilm
measured by CSLM
S.
no.
Biofilm parameters (Mean ± SE)
1. Total biomass (lm3/lm2) 10.81 ± 2.67
2. Roughness coefficient
(dimensionless)
0.379 ± 0.103
3. Surface area of biomass (lm2) 132401.6 ± 11083.39
4. Surface to biovolume ratio (lm2/
lm3)
4.59 ± 1.12
5. Maximum thickness (lm) 21.02 ± 2.10
6. Average thickness (lm) 14.82 ± 2.61
N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus 653
123

Fig. 5 a Scanning electron
microscopic image of P. lautus
NE3B01 biofilm, showing mat-
like compact structure.
b Differential cell morphology
of P. lautus NE3B01. Arrow
and circle pointing rod and
cocci cells, respectively. c Rod-
shaped planktonic cells after
crystal violet staining observed
under light microscope.
d Typical doughnut-shaped
cells of P. lautus NE3B01
observed under scanning
electron microscope
Fig. 6 A schematic diagram illustrating the different stages in phenotypic switching of P. lautus NE3B01 during biofilm growth
654 N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus
123

differential phenotype and bacterial adaptation to conges-
tion or overcrowding in a biofilm.
Conclusion
The present study reveals pleomorphism in P. lautus
NE3B01 micro-colonies and biofilm when grown on a solid
surface. Cells of the biofilm community switched from
non-motile cocci to motile rods due to competition for
space and survival. The strain showed typical phases of
biofilm formation and dispersal as well as swarming
motility on agar surface. This suggests that phenotypic
variation is reversible and is interconnected with cocci and
rods. Coccoid cells preferred to live in association, whereas
their transformation into rods results in acquisition of fla-
gellar motility and dispersal. The findings are suggestive of
mechanism to deal with space and nutrient scarcity for
long-term survival. The ability to replicate and maintain
coccoid forms and promote transformation to motile rods
requires further molecular investigations. However, in a
holistic view, the findings can be useful in understanding
bacterial adaptation and social behaviour while existing as
a colony or a biofilm.
Acknowledgments The authors would like to acknowledge the
authorities of NIT, Rourkela and BARC Facilities, Kalpakkam for
providing facilities. N.M. gratefully acknowledges the receipt of
fellowship from Ministry of Human Resource Development, Gov-
ernment of India for doctoral research. S.D. thanks the Department of
Biotechnology, Government of India for research grants on biofilm-
based enhanced bioremediation.
Conflicts of interest Authors of the manuscript declare no conflict
of interest.
References
1. Be’er A, Florin EL, Fisher CR, Swinney HL, Payne SM (2011)
Surviving bacterial sibling rivalry: inducible and reversible phe-
notypic switching in Paenibacillus dendritiformis. MBio. doi:10.
1128/mBio.00069-11
2. Becker P, Hufnagle W, Peters G, Herrmann M (2001) Detection
of differential gene expression in biofilm-forming versus plank-
tonic populations of Staphylococcus aureus using micro-repre-
sentational-difference analysis. Appl Environ Microbiol
67(7):2958–2965
3. Ben-Jacob E, Cohen I, Golding I, Gutnick DL, Tcherpakov M,
Helbing D, Ron IG (2000) Bacterial cooperative organization
under antibiotic stress. Phys Stat Mech Appl 282(1):247–282
4. Ben-Jacob E, Cohen I, Levine H (2000) Cooperative self-orga-
nization of microorganisms. Adv Phys 49(4):395–554
5. Dash H, Mangwani N, Das S (2013) Characterization and
potential application in mercury bioremediation of highly mer-
cury-resistant marine bacterium Bacillus thuringiensis PW-05.
Environ Sci Pollut Res. doi:10.1007/s11356-013-2206-8
6. Debois D, Hamze K, Guerineau V, LeCaer JP, Holland IB, Lopes
P et al (2008) In situ localisation and quantification of surfactins
in a Bacillus subtilis swarming community by imaging mass
spectrometry. Proteomics 8(18):3682–3691
7. Dubnau D, Losick R (2006) Bistability in bacteria. Mol Microbiol
61(3):564–572
8. Errington J (2003) Regulation of endospore formation in Bacillus
subtilis. Nat Rev Microbiol 1(2):117–126
9. Fauvart M, Phillips P, Bachaspatimayum D, Verstraeten N,
Fransaer J, Michiels J, Vermant J (2012) Surface tension gradient
control of bacterial swarming in colonies of Pseudomonas
aeruginosa. Soft Matter 8(1):70–76
10. Finkel SE (2006) Long-term survival during stationary phase:
evolution and the GASP phenotype. Nat Rev Microbiol
4(2):113–120
11. Gonzalez-Pastor JE, Hobbs EC, Losick R (2003) Cannibalism by
sporulating bacteria. Sci Signal 301(5632):510
12. Hall-Stoodley L, Nistico L, Sambanthamoorthy K, Dice B,
Nguyen D, Mershon WJ, Post JC (2008) Characterization of
biofilm matrix, degradation by DNase treatment and evidence of
capsule downregulation in Streptococcus pneumoniae clinical
isolates. BMC Microbiol 8(1):173
13. Heydorn A, Nielsen AT, Hentzer M, Sternberg C, Givskov M,
Ersbøll BK, Molin S (2000) Quantification of biofilm structures
by the novel computer program COMSTAT. Microbiology
146(10):2395–2407
14. Ingham CJ, Jacob EB (2008) Swarming and complex pattern
formation in Paenibacillus vortex studied by imaging and
tracking cells. BMC Microbiol 8(1):36
15. Irie Y, Parsek MR (2008) Quorum sensing and microbial bio-
films. In: Tony R (ed) Bacterial biofilms. Springer, Berlin,
pp 67–84
16. Jain K, Parida S, Mangwani N, Dash HR, Das S (2013) Isolation
and characterization of biofilm-forming bacteria and associated
extracellular polymeric substances from oral cavity. Ann
Microbiol. doi:10.1007/s13213-013-0618-9
17. Joshi HM, Rao TS (2009) Nutrition induced pleomorphism and
budding mode of reproduction in Deinococcus radiodurans.
BMC Res Notes 2(1):123
18. Kearns DB (2010) A field guide to bacterial swarming motility.
Nat Rev Microbiol 8(9):634–644
19. Mangwani N, Dash HR, Chauhan A, Das S (2012) Bacterial
quorum sensing: functional features and potential applications in
biotechnology. J Mol Microbiol Biotechnol 22(4):215–227
20. Mangwani N, Shukla SK, Rao TS, Das S (2013) Calcium-med-
iated modulation of Pseudomonas mendocina NR802 biofilm
influences the phenanthrene degradation. Colloids Surf B. doi:10.
1016/j.colsurfb.2013.10.003
21. Markova N, Slavchev G, Michailova L (2012) Unique biological
properties of Mycobacterium tuberculosis L-form variants:
impact for survival under stress. Int Microbiol 15(2):61–68
22. Nadell CD, Xavier JB, Levin SA, Foster KR (2008) The evolu-
tion of quorum sensing in bacterial biofilms. PLoS Biol 6(1):e14
23. Noguchi H (1910) Pleomorphism and pleobiosis of Bacillus
bifiduscommunis. J Exp Med 12(2):182–195
24. O’Toole G, Kaplan HB, Kolter R (2000) Biofilm formation as
microbial development. Ann Rev Microbiol 54(1):49–79
25. Parsek MR, Greenberg EP (2005) Sociomicrobiology: the con-
nections between quorum sensing and biofilms. Trends Microbiol
13(1):27–33
26. Roth D, Finkelshtein A, Ingham C, Helman Y, Sirota-Madi A,
Brodsky L, Ben-Jacob E (2013) Identification and characteriza-
tion of a highly motile and antibiotic refractory subpopulation
involved in the expansion of swarming colonies of Paenibacillus
vortex. Environ Microbiol 15(9):2532–2544
N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus 655
123

27. Sandal I, Hong W, Swords WE, Inzana TJ (2007) Characterization
and comparison of biofilm development by pathogenic and com-
mensal isolates of Histophilussomni. J Bacteriol 189(22):8179–8185
28. Shemesh M, Tam A, Steinberg D (2007) Differential gene
expression profiling of Streptococcus mutans cultured under
biofilm and planktonic conditions. Microbiology 153(5):1307–
1317
29. Shukla SK, Sankar GG, Paraneeiswaran A, Rao TS (2013) Dif-
ferential radio-tolerance of nutrition-induced morphotypes of
Deinococcus radiodurans R1. Curr Microbiol. doi:10.1007/
s00284-013-0472-9
30. Shukla SK, Rao TS (2013) Dispersal of Bap-mediated Staphy-
lococcus aureus biofilm by proteinase K. J Antibiot 66(2):55–60
31. Shukla SK, Rao TS (2013) Effect of calcium on Staphylococcus
aureus biofilm architecture: a confocal laser scanning micro-
scopic study. Colloids Surf B 103:448–454
32. Verstraeten N, Braeken K, Debkumari B, Fauvart M, Fransaer J,
Vermant J, Michiels J (2008) Living on a surface: swarming and
biofilm formation. Trends Microbiol 16(10):496–506
33. Wainwright M (2000) Highly pleomorphic Staphylococci as a
cause of cancer. Med Hypotheses 54(1):91–94
656 N. Mangwani et al.: Phenotypic Switching in Paenibacillus lautus
123
Copyright © 2022 FDOKUMEN




















