Pharmacological management of appetite expression in obesity
15
NATURE REVIEWS | ENDOCRINOLOGY VOLUME 6 | MAY 2010 | 255 School of Psychology, University of Liverpool, Eleanor Rathbone Building, Bedford Street South, Liverpool L69 7ZA, UK (J. C. G. Halford, E. J. Boyland, T. C. Kirkham, J. A. Harrold). Institute of Psychological Sciences, Faculty of Medicine and Health, University of Leeds, Woodhouse Lane, Leeds LS2 9JT, UK (J. E. Blundell). Correspondence to: J. C. G. Halford j.c.g.halford@ liverpool.ac.uk Pharmacological management of appetite expression in obesity Jason C. G. Halford, Emma J. Boyland, John E. Blundell, Tim C. Kirkham and Joanne A. Harrold Abstract | For obese individuals, successful weight loss and maintenance are notoriously difficult. Traditional drug development fails to exploit knowledge of the psychological factors that crucially influence appetite, concentrating instead on restrictive criteria of intake and weight reduction, allied to a mechanistic view of energy regulation. Drugs are under development that may produce beneficial changes in appetite expression in the obese. These currently include glucagon-like peptide-1 analogs such as liraglutide, an amylin analog davalintide, the 5-HT 2C receptor agonist lorcaserin, the monoamine re-uptake inhibitor tesofensine, and a number of combination therapies such as pramlintide and metreleptin, bupropion and naltrexone, phentermine and topiramate, and bupropion and zonisamide. However, the effects of these treatments on eating behavior remain poorly characterized. Obesity is typically a consequence of overconsumption driven by an individual’s natural sensitivity to food stimuli and the pleasure derived from eating. Intuitively, these processes should be effective targets for pharmacotherapy, and behavioral analysis can identify drugs that selectively affect desire to eat, enjoyment of eating, satiation or postmeal satiety. Rational interventions designed specifically to modulate these processes could limit the normally aversive consequences of caloric restriction and maximize an individual’s capacity to successfully gain control over their appetite. Halford, J. C. G. et al. Nat. Rev. Endocrinol. 6, 255–269 (2010); published online 16 March 2010; doi:10.1038/nrendo.2010.19 Introduction Appropriate changes to energy balance (reducing energy intake and/or increasing expenditure) is fundamental to weight management. While it is estimated that over the long term even small adjustments in daily energy balance of the order of 50 kcal should reduce body weight, 1 a meta-analysis of UK clinical data suggests that meaningful weight loss (5 kg over 12 months) can only be achieved through adherence to a 600 kcal energy-deficit diet or by reducing dietary fat (thereby beneficially altering the satiating potency of meals and/or the general reduction of total calorie intake). 2 Unfortunately, individuals experience difficulty making the necessary behavioral adjustments needed to yield meaningful weight loss. These difficulties arise from conflicts with the strong biological forces that maintain weight status, and particularly the powerful motivational factors that promote consumption. For these reasons, substantial negative psychological consequences result from caloric restriction. Dieting produces clear changes in appetite, notably an increase in hunger. 3 The consequences of severe caloric restriction are increased preoccupation with food, persistent thoughts of eating and serious diffi- culty with dietary adherence in the presence of food. 4–7 Management of the natural inclination to eat, and speci- fically of diet-induced hunger, in order to help indivi- duals resist the strong environmental and situational cues to overconsume should, therefore, be an important factor in any weight-loss therapy. However, regulatory bodies and pharmaceutical companies focus almost exclusively on reduction of body mass and normaliza- tion of biomarkers of obesity-related disease as clinical end points in trials of obesity drugs. 8 Thus, the attention of researchers and regulators is directed at the conse- quences rather than the causes of obesity. Little regula- tory emphasis is currently placed on helping the obese manage their drive to consume. A vast array of factors has been targeted to counter obesity. The factors fall into four main groups. Firstly, factors involved in the preabsorptive processes that break down food and control the passage of nutrients from the gastrointestinal tract. Secondly, the post- ingestive hormones produced in the gastrointestinal tract that control intestinal function (and the pancre- atic hormones that control energy metabolism); both types of hormones may additionally influence appetite expression. Thirdly, the peripheral processes involved in fat deposition in adipose tissue and the utilization of energy. Lastly, the myriad of neurotransmitters and Competing interests J. C. G. Halford declares associations with the following companies/organizations: GlaxoSmithKline, Novo Nordisk, Prosidion OSI Pharmaceuticals. J. E. Blundell declares associations with the following companies/organizations: Amylin, Barilla, Biotechnology and Biological Sciences Research Council, British Nutrition Foundation, Coca-Cola, Covance, Danone, European Association for the Study of Obesity, European Food Safety Authority, Fonterra, Kellogg’s, Merck, National Starch, Sanofi-Aventis, Unilever. See the article online for full details of the relationships. The other authors declare no competing interests. REVIEWS © 20 Macmillan Publishers Limited. All rights reserved 10
Transcript of Pharmacological management of appetite expression in obesity

nature reviews | endocrinology volume 6 | may 2010 | 255
School of Psychology, University of Liverpool, Eleanor Rathbone Building, Bedford Street South, Liverpool L69 7ZA, UK (J. c. g. Halford, e. J. Boyland, T. c. Kirkham, J. A. Harrold). Institute of Psychological Sciences, Faculty of Medicine and Health, University of Leeds, Woodhouse Lane, Leeds LS2 9JT, UK (J. e. Blundell).
Correspondence to: J. C. G. Halford j.c.g.halford@ liverpool.ac.uk
Pharmacological management of appetite expression in obesityJason C. G. Halford, Emma J. Boyland, John E. Blundell, Tim C. Kirkham and Joanne A. Harrold
Abstract | For obese individuals, successful weight loss and maintenance are notoriously difficult. Traditional drug development fails to exploit knowledge of the psychological factors that crucially influence appetite, concentrating instead on restrictive criteria of intake and weight reduction, allied to a mechanistic view of energy regulation. Drugs are under development that may produce beneficial changes in appetite expression in the obese. These currently include glucagon-like peptide-1 analogs such as liraglutide, an amylin analog davalintide, the 5-HT2C receptor agonist lorcaserin, the monoamine re-uptake inhibitor tesofensine, and a number of combination therapies such as pramlintide and metreleptin, bupropion and naltrexone, phentermine and topiramate, and bupropion and zonisamide. However, the effects of these treatments on eating behavior remain poorly characterized. Obesity is typically a consequence of overconsumption driven by an individual’s natural sensitivity to food stimuli and the pleasure derived from eating. Intuitively, these processes should be effective targets for pharmacotherapy, and behavioral analysis can identify drugs that selectively affect desire to eat, enjoyment of eating, satiation or postmeal satiety. Rational interventions designed specifically to modulate these processes could limit the normally aversive consequences of caloric restriction and maximize an individual’s capacity to successfully gain control over their appetite.
Halford, J. C. G. et al. Nat. Rev. Endocrinol. 6, 255–269 (2010); published online 16 March 2010; doi:10.1038/nrendo.2010.19
Introductionappropriate changes to energy balance (reducing energy intake and/or increasing expenditure) is fundamental to weight management. while it is estimated that over the long term even small adjustments in daily energy balance of the order of 50 kcal should reduce body weight,1 a meta-analysis of uK clinical data suggests that meaningful weight loss (5 kg over 12 months) can only be achieved through adherence to a 600 kcal energy-deficit diet or by reducing dietary fat (thereby beneficially altering the satiating potency of meals and/or the general reduction of total calorie intake).2 unfortunately, individuals experience difficulty making the necessary behavioral adjustments needed to yield meaningful weight loss. these difficulties arise from conflicts with the strong biological forces that maintain weight status, and particularly the powerful motivational factors that promote consumption. For these reasons, sub stantial negative psychological consequences result from caloric restriction.
Dieting produces clear changes in appetite, notably an increase in hunger.3 the consequences of severe caloric restriction are increased preoccupation with food, persistent thoughts of eating and serious diffi-culty with dietary adherence in the presence of food.4–7 management of the natural inclination to eat, and speci-fically of diet-induced hunger, in order to help indivi-duals resist the strong environ mental and situational cues to overconsume should, therefore, be an important factor in any weight-loss therapy. However, regulatory bodies and pharmaceutical companies focus almost exclusively on reduction of body mass and normaliza-tion of biomarkers of obesity-related disease as clinical end points in trials of obesity drugs.8 thus, the attention of researchers and regulators is directed at the conse-quences rather than the causes of obesity. little regula-tory emphasis is currently placed on helping the obese manage their drive to consume.
a vast array of factors has been targeted to counter obesity. the factors fall into four main groups. Firstly, factors involved in the preabsorptive processes that break down food and control the passage of nutrients from the gastrointestinal tract. secondly, the post-ingestive hormones produced in the gastrointestinal tract that control intestinal function (and the pancre-atic hormones that control energy metabolism); both types of hormones may additionally influence appetite expression. thirdly, the peripheral processes involved in fat deposition in adipose tissue and the utilization of energy. lastly, the myriad of neurotransmitters and
competing interestsJ. C. G. Halford declares associations with the following companies/organizations: GlaxoSmithKline, Novo Nordisk, Prosidion OSI Pharmaceuticals. J. E. Blundell declares associations with the following companies/organizations: Amylin, Barilla, Biotechnology and Biological Sciences Research Council, British Nutrition Foundation, Coca-Cola, Covance, Danone, European Association for the Study of Obesity, European Food Safety Authority, Fonterra, Kellogg’s, Merck, National Starch, Sanofi-Aventis, Unilever. See the article online for full details of the relationships. The other authors declare no competing interests.
REvIEWS
nrendo_19_MAY10.indd 255 30/3/10 15:43:52
© 20 Macmillan Publishers Limited. All rights reserved10

256 | may 2010 | volume 6 www.nature.com/nrendo
neuromodulators within the central nervous system (Cns) that have been proposed to modulate energy balance and control eating behavior.
of all these factors, putative satiety hormones and neurotransmitters involved in energy regulation circuits centered on the hypothalamus have been key targets for appetite-suppressing agents. the systems underpinning energy regulation in the gut, the Cns and adipose tissues act in an integrated manner. For example, inhibition of digestion can affect nutrient-derived satiety signals,9,10 while changes in peripheral energy storage exert power-ful effects on appetite mediated via the hypothalamic energy regulatory systems.11,12 Consequently, weight- control interventions may lead to changes in eating behavior through a variety of indirect routes.
a complex interplay of integrative neural processes within the brain, which are sensitive to a plethora of signals that originate in the periphery, underpin the behavioral expression and conscious experience of appetite and represent the ultimate factors in the control of energy intake. episodic factors initiate, sustain, ter-minate and inhibit eating, whereas tonic factors derived from the body’s energy reserves modulate these episodic processes.13,14 the combination of episodic and tonic feedback integrates biological need with environmental influences through an individual’s subjective experience of appetite and its components; namely: hunger (the drive to consume that elicits and sustains eating), satiation (within-meal processes that terminate a meal) and satiety (postmeal satisfaction that suppresses further intake).
as the very existence of obesity demonstrates, energy balance is not perfectly regulated—in spite of the per-sistent theoretical dominance of homeostasis. more particularly, the psychological aspects of appetite are pre-eminent in the determination of nutrient intake and all too evidently override any inhibitory mecha-nisms that might operate in response to energetic suf-ficiency or surplus. sensory input (such as taste, flavor and mouth feel) and cognitive factors (including beliefs,
Key points
Traditional antiobesity drug development focuses on weight and ■obesity-related disease end points, without adequate consideration of behavioral and psychological changes that initiate weight gain or that impede weight management
Classical energy homeostasis models emphasize energy regulation systems, ■but fail to acknowledge key motivational, emotional and behavioral factors that contribute to energy intake and the propensity to gain weight
Drug efficacy has been limited, as key behavioral features of appetite have ■been ignored—namely, hedonic aspects of consumption and the powerful influence of the environment in triggering overeating
Pharmaceutical companies are beginning to focus on treatments that ■combine existing compounds to exploit additive, or supra-additive, actions of separate agents
This approach could be advanced by exploiting the specific actions of drugs ■on distinct components of eating motivation rather than merely emphasizing simple effects on energy intake and body weight
Behavioral analysis can isolate drugs that reduce the desire to eat by acting on ■the processes that initiate, sustain and terminate meals or inhibit consumption
expectations, learned associations and prior experience) are critical to the pleasurable and rewarding experiences of eating that are such important determinants of human appetite, particularly in an environment in which food choices are abundant and deprivation is rare.
increasingly, the roles of both wanting (the desire to consume food, often instigated by food stimuli) and liking (the positive hedonic response to food stimuli) in the control of human appetite are being addressed by those studying weight control.15,16 Food preferences and dietary habits result from lifelong learning and are determined both by the palatability of foods and the frequency and context of exposure. to alter established patterns of dietary behavior by changing the desire for specific foods and reinforcing the positive experience of other specific foods is, therefore, a challenge for any therapeutic intervention, but may provide the most effective strategy for weight management. Currently, despite the self-evident importance of these factors, they are not being appropriately addressed by pharmaceutical companies in their antiobesity programs. the current review examines behaviors associated with obesity that could provide strong indications of both efficacy and specificity in drug development and discusses the effects of various medications on appetite expression in obesity.
Eating behavior and obesitynumerous gene polymorphisms have been linked to obe-sity.17 However, only in the last 5 years have specific genes that underpin appetite expression in general obesity been widely considered.18 Blundell and colleagues19 have iden-tified a constellation of behavioral traits which, in con-junction with a high-fat, energy-dense, palatable diet, promote adiposity and constitute an ‘at risk’ behavioral phenotype. recent research indicates that increased food intake and adiposity is associated with specific variants of the fat mass and obesity-associated (FTO) gene. thus, individuals who carry at-risk alleles of the FTO gene not only are heavier and more likely to be obese but also consume more calories relative to those individuals with the protective genotype. these genotypic differences contribute to a behavioral phenotype, which supports the notion that individual differences in eating behavior contribute importantly to general obesity risk.20–23
laboratory studies have demonstrated differences between lean and obese individuals in the way that they eat. thus, the obese tend to eat faster, take larger mouth-fuls, chew less before swallowing and are less likely to reduce their eating rate over the course of a meal than lean individuals,24–27 effects that can be associated with a deficit in the operation of normal satiation.28 Certainly, infusions of peripheral satiety peptides appear to slow the rate of consumption in the obese.29 although not all obese people fail to reduce eating rate over the course of a meal, the phenomenon of sustained high rates of intake is a marked characteristic of rare obesity-related conditions such as Prader–willi syndrome and leptin deficiency or insufficiency30,31 and may be particularly apparent in the severely obese.27 epidemiological data also confirm that
reviews
nrendo_19_MAY10.indd 256 30/3/10 15:43:53
© 20 Macmillan Publishers Limited. All rights reserved10

nature reviews | endocrinology volume 6 | may 2010 | 257
both obesity and the liability to gain weight are as sociated with self-reported high eating rates,32,33 and it seems that the relationship between eating rate and adiposity may well be heritable.34 unsurprisingly, fast eating rates gen-erally result in large meal sizes, which in turn are also associated with weight gain and obesity.35,36 interestingly, these large meals do not seem to result in proportionally strong postmeal satiety.35,36
with regard to mechanisms, a combination of dietary, physiological and neural factors seem to contribute to poor satiation and satiety responses. the consump-tion of energy-dense foods (those high in fat or refined sugar) promotes excessive energy intake and often pro-duces comparatively weak satiety on a calorie-for-calorie basis.19,35–38 reduced gastrointestinal peptide responses to ingested nutrients and larger stomach capacities may also underpin weaker satiation.39–45 Finally, diminished Cns responses to ingestion in brain areas associated with satiety are observed in the obese.46,47 thus food type, gastric capacity and deficiencies in the physio logical and neurological response to ingestion may combine to promote, or at least allow, overconsumption.
while such data are consistent with the notion that ingested calories have a less inhibitory effect on the appetite of obese individuals, clearly differences between the appetite of lean and obese individuals go beyond sati-ation and satiety. the idea that individuals with obesity have a heightened disposition to respond to stimuli to eat is a well-established concept.19,28 numerous studies have shown that the obese experience feelings of con-stant and/or excessive hunger and are overresponsive to the sensory and hedonic characteristics of food. in chil-dren, both epidemiological and experimental research suggest that obesity is related to overresponsiveness to food cues.48,49 moreover, in the obese, the effect of food cues on motivational systems seems to be resistant to the normal inhibitory consequences of ingestion.50,51 early studies have demonstrated that obese indivi duals chose a greater variety of foods than lean individuals when offered a range of palatable options.28 more over, food preferences in the obese and individuals who gain weight easily are characterized by a strong preference for fat,19,35,36,52 and pleasantness ratings for high-fat foods in these individuals have been shown to correlate with percentage body fat.53,54
overall, the data suggest that the hedonic experience of ingestion, particularly the enjoyment of energy-dense, high-fat foods, seems to be more influential in obese than lean individuals. Given the powerful influence of palatability, and contrary to prevailing, physiologically- centered views, differences in eating rate observed during meals may not indicate specific deficits in satiation so much as the overriding impact of the hedonic experience to promote consumption. in particular, responsiveness to a highly palatable food, even when an individual reports already being sated, remains a key risk factor for over-consumption.19 thus, obese individuals are more likely to be provoked to eat, irrespective of previous consump-tion, and are more likely to overconsume if presented with a palatable food. although this behavior reflects
evolutionarily advantageous adaptation, such traits are highly disadvantageous in an environment in which highly-palatable, energy-dense foods are both heavily promoted and widely available.
Measuring eating behavior and appetiteDetecting differences in eating behavior between lean and obese individuals in the laboratory or in more naturalis tic settings than the laboratory is challenging.55,56 How ever, laboratory-based studies that have employed ad libitum meals have successfully characterized the effects on eating behavior and appetite of clinically effective antiobesity agents, particularly serotonergic drugs.57 subjective experi ences of appetite, measured with visual analog scales, have been employed to confirm satiety-enhancing effects (an important indication of behavioral specificity), and changes in visual analog scales ratings can predict subsequent energy intake and indicate potential changes in body weight.58–61
measures of eating rate and within-meal appetite also appear particularly useful in discriminating between drugs that merely disrupt eating behavior by non selective actions from those that specifically enhance satiation or reduce the hedonic experience of ingestion.62–64 thus, a variety of well-validated designs (ad libitum intake to measure satiation, fixed caloric preloads to assess sa tiation), measures (indices of appetite, sensory evalua-tion, preference and choice, pleasure and other aspects of motivation to consume) and protocols (manipulations of foods, meals or diets, within-meal or between-meal measures of behavior) are available to researchers to charac terize the behavioral effects of drugs within the laboratory early in their clinical development.56 nevertheless, despite the potential of these methods to quickly indicate efficacy, specificity, mode of action and potential adverse effect issues, they remain surprisingly underutilized in drug development.
the experimental approach has its constraints, of course, and it remains difficult to model behavior exhibited in obesity-promoting environments within the laboratory. nevertheless, the utility of these assays has been sufficiently demonstrated for pharmaceutical companies to be advised to adopt them, in order to avoid the repeated licensing and marketing problems that have followed failures to properly test candidate drug effects on human eating behavior in early clinical trials.65 the argument could be made that such additional complexity in testing and data analysis may prove to be prohibitively costly in relation to the scale of clinical trials. However, attention to behavioral and motivational specificity of a candidate drug could easily be addressed in phase i and ii trials—guided, of course, by appropriately detailed assessment of drug specificity through observational analysis and other behavioral measures conducted in preclinical studies.
Certainly, to underuse or ignore the potential of human trials to improve understanding of the potential psycho-logical mechanisms of action of candidate antiobesity agents may well lead to greater costs at a later stage. the withdrawal of rimonabant and the discontinuation
reviews
nrendo_19_MAY10.indd 257 30/3/10 15:43:54
© 20 Macmillan Publishers Limited. All rights reserved10

258 | may 2010 | volume 6 www.nature.com/nrendo
of clinical research programs for type 1 cannabinoid re ceptor (CB1) antagonists and inverse agonists is a case in point (see below).
in the real world, food cues are more abundant, eating opportunities more frequent and food choices less restricted than in laboratory-based trials. Consequently, clinical trials often depend on retrospective accounts of free-living caloric intake, food choices and day-to-day experiences of appetite (satiety, cravings and food pr eferences). nonetheless, despite ecological validity, such measures remain subject to inaccuracy and report-ing bias. technology has enabled us to capture reliable real-time changes in appetite outside the lab,66,67 to sample behavior and obtain psychological evaluations and to generate detailed data on eating behavior suitable for large-scale clinical trials.68–71 these assays monitor appetite, food intake, food preferences and experience of other drug effects by use of a variety of response formats (validated questionnaires, motivational rating scales, image choices and dietary intake reports). moreover, the assays can include more detailed measures of enduring aberrations in eating behavior and the drive to consume that should provide useful data in long-term trials.
Behavioral traits assessed by tools such as the three Factor eating Questionnaire, Dutch eating Behavior Questionnaire, Binge eating scale and Power of Food scale often show only a weak overall association with body mass.72,73 importantly, however, these tools do assess issues in eating behavior control that are shared by many of those trying to manage their weight (irrespec tive of current weight status). Certainly, feelings of constant and provokable hunger74–77 and excessive, poorly-controlled eating behavior76,78–82 are associated with increased adi-posity. Heritability studies suggest that these factors may form a consistent, distinct behavioral phenotype linked to a susceptibility to weight gain.83,84 moreover, more extreme forms of eating behavior, such as binge eating, remain a critical issue for a subset of obese individuals.85–87 Consequently, despite an apparently weak relationship with Bmi, aberrant eating be havior provides valuable insight into the factors that may mi tigate against treatment compliance and success.88,89
Critically, some of these traits seem to predict which drug treatment patients are most likely to respond to; for example, those who exhibit high dietary restraint respond best to orlistat and those who exhibit low dietary restraint respond best to sibutramine.90,91 thus, self-reported reductions in unabated hunger, food cravings and preoccupations, responsiveness to food and corres-ponding uncontrolled eating behavior could be useful indicators of behavioral change in large-scale clinical trials. these, together with modifications in the desire for and appreciation of highly palatable, energy-dense, obesity-promoting foods, provide etiologically appro-priate behavioral outcomes for long-term pharmaco-logical trials. Frustratingly, measures of eating behavior are often employed as inclusion criteria for large-scale clinical studies, but rarely are the data analyzed to show which traits were associated with successful weight control (with or without pharmacotherapy).
Obesity pharmacotherapyGiven the critical role of behavior in the etiology of obesity, it is only logical that the effects of antiobesity drugs on appetite expression should be scrutinized. only limited behavioral data are available on many treatments that are undergoing regulatory approval. this situation is particularly true for many novel Cns-acting agents (see below). nonetheless, for peripherally acting agents at least, a strong experimental medicine literature details the effects of various hormones on food intake and appetite expression in the target population.
Peripherally acting agentsthe gastrointestinal tract and the pancreas secrete a rich array of hormones that control gut motility and glucose metabolism. many of these hormones, such as the gut peptides ghrelin, cholecystokinin, glucagon-like peptide-1 (GlP-1) and peptide yy (Pyy), act as compo-nents in episodic appetite control and mediate powerful postingestive influences of nutrients on hunger, sati ation and satiety.41,92–94 these hormones are proposed to influ-ence the expression of eating behavior by modifying intestinal transit, changing rates of nutrient absorption, stimulating afferent vagal signals and/or penetrating appetite control centers within the Cns.
endogenous levels of some of these hormones (GlP-1 and Pyy) are diminished in obese individuals. in addi-tion, despite the increased caloric loads of obese indivi-duals, these individuals’ hormonal responses to ingestion also seem to be attenuated.41,43,44,95 nonetheless, these systems still remain viable targets for obesity treatment. Currently, the development of weight- management treatments based on GlP-1 and the pancreatic hormone amylin appear to be the most advanced. although the stimulatory effects of ghrelin infusions on human eating behavior are well documented, clinical data on the effects of ghrelin antagonists on appetite remain unavailable, with most agents still in the early stages of clinical development. after previous failures,96 no cholecystokinin-based therapies appear to be under-going clinical trials because of the unsatisfactory effects of available formulations.
GlP-1, an incretin hormone synthesized by l cells of the distal small and large intestine, is released in response to various forms of carbohydrate, soluble fiber and free fatty acids within the gastrointestinal tract. the endo-genous GlP-1 response generally occurs some time after ingestion, and the hormone is thus associated with post-meal satiety.97 the hormone is also implicated in the ‘ileal brake’, in which nutrient signals from the small intestine feed back to inhibit gastric emptying.98,99 exogenous in fusions of GlP-1 modify appetite and reduce energy intake in lean individuals.100–103 similar changes in pre-meal hunger, prospective consumption and food intake have been observed in overweight patients with type 2 diabetes mellitus and obese individuals.104–108 subcu-taneous injections of GlP-1 induce weight loss in the obese;106 however, GlP-1 is rapidly inactivated by dipepti-dyl pepti dase 4 (DPP4), which limits its usefulne ss as a treatment option. a summary of the effects of GlP-1,
reviews
nrendo_19_MAY10.indd 258 30/3/10 15:43:55
© 20 Macmillan Publishers Limited. All rights reserved10
nature reviews | endocrinology volume 6 | may 2010 | 259
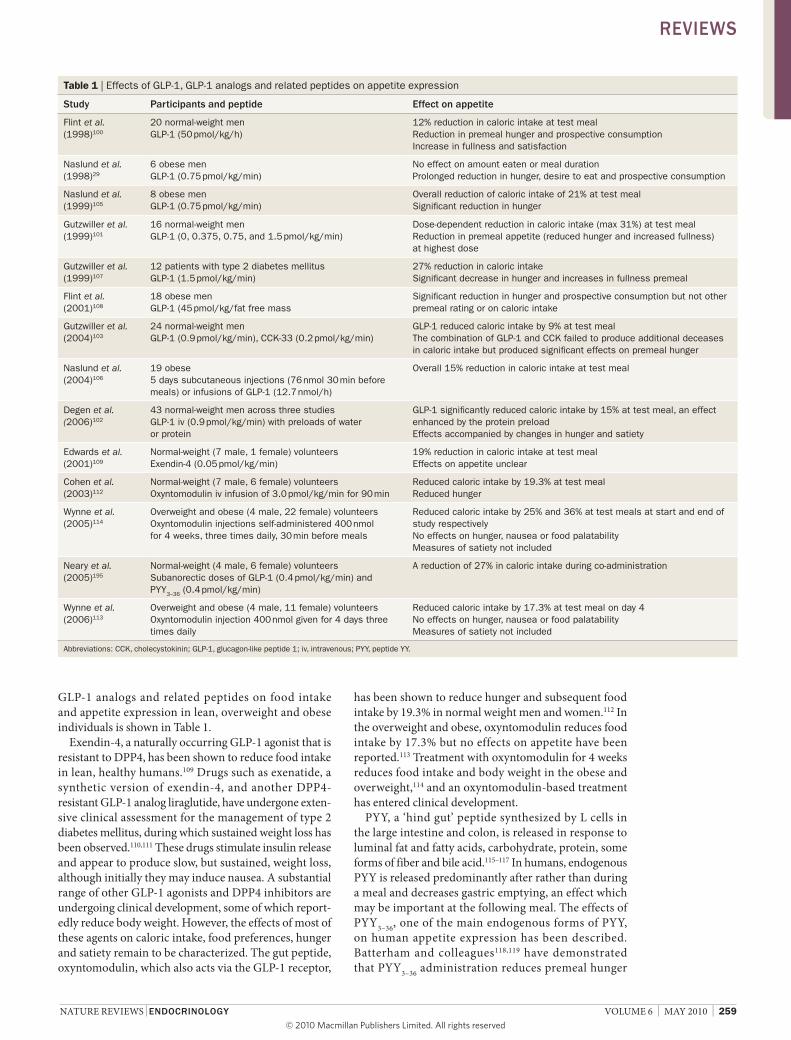
GlP-1 analogs and related peptides on food intake and appetite expression in lean, overweight and obese in dividuals is shown in table 1.
exendin-4, a naturally occurring GlP-1 agonist that is resistant to DPP4, has been shown to reduce food intake in lean, healthy humans.109 Drugs such as exenatide, a synthetic version of exendin-4, and another DPP4-resistant GlP-1 analog liraglutide, have undergone exten-sive clinical assessment for the management of type 2 diabetes mellitus, during which sustained weight loss has been observed.110,111 these drugs stimulate insulin release and appear to produce slow, but sustained, weight loss, although initially they may induce nausea. a substantial range of other GlP-1 agonists and DPP4 inhibitors are undergoing clinical development, some of which report-edly reduce body weight. However, the effects of most of these agents on caloric intake, food preferences, hunger and satiety remain to be characterized. the gut peptide, oxyntomodulin, which also acts via the GlP-1 receptor,
has been shown to reduce hunger and subsequent food intake by 19.3% in normal weight men and women.112 in the overweight and obese, oxyntomodulin reduces food intake by 17.3% but no effects on appetite have been reported.113 treatment with oxyntomodulin for 4 weeks reduces food intake and body weight in the obese and overweight,114 and an oxyntomodulin-based treatment has entered clinical development.
Pyy, a ‘hind gut’ peptide synthesized by l cells in the large intestine and colon, is released in response to luminal fat and fatty acids, carbohydrate, protein, some forms of fiber and bile acid.115–117 in humans, endo genous Pyy is released predominantly after rather than during a meal and decreases gastric emptying, an effect which may be important at the following meal. the effects of Pyy3–36, one of the main endogenous forms of Pyy, on human appetite expression has been described. Batterham and colleagues118,119 have de monstrated that Pyy3–36 administration reduces premeal hunger
Table 1 | Effects of GLP-1, GLP-1 analogs and related peptides on appetite expression
study Participants and peptide effect on appetite
Flint et al. (1998)100
20 normal-weight men GLP-1 (50 pmol/kg/h)
12% reduction in caloric intake at test meal Reduction in premeal hunger and prospective consumption Increase in fullness and satisfaction
Naslund et al. (1998)29
6 obese men GLP-1 (0.75 pmol/kg/min)
No effect on amount eaten or meal duration Prolonged reduction in hunger, desire to eat and prospective consumption
Naslund et al. (1999)105
8 obese men GLP-1 (0.75 pmol/kg/min)
Overall reduction of caloric intake of 21% at test meal Significant reduction in hunger
Gutzwiller et al. (1999)101
16 normal-weight men GLP-1 (0, 0.375, 0.75, and 1.5 pmol/kg/min)
Dose-dependent reduction in caloric intake (max 31%) at test meal Reduction in premeal appetite (reduced hunger and increased fullness) at highest dose
Gutzwiller et al. (1999)107
12 patients with type 2 diabetes mellitus GLP-1 (1.5 pmol/kg/min)
27% reduction in caloric intake Significant decrease in hunger and increases in fullness premeal
Flint et al. (2001)108
18 obese men GLP-1 (45 pmol/kg/fat free mass
Significant reduction in hunger and prospective consumption but not other premeal rating or on caloric intake
Gutzwiller et al. (2004)103
24 normal-weight men GLP-1 (0.9 pmol/kg/min), CCK-33 (0.2 pmol/kg/min)
GLP-1 reduced caloric intake by 9% at test meal The combination of GLP-1 and CCK failed to produce additional deceases in caloric intake but produced significant effects on premeal hunger
Naslund et al. (2004)106
19 obese 5 days subcutaneous injections (76 nmol 30 min before meals) or infusions of GLP-1 (12.7 nmol/h)
Overall 15% reduction in caloric intake at test meal
Degen et al. (2006)102
43 normal-weight men across three studies GLP-1 iv (0.9 pmol/kg/min) with preloads of water or protein
GLP-1 significantly reduced caloric intake by 15% at test meal, an effect enhanced by the protein preload Effects accompanied by changes in hunger and satiety
Edwards et al. (2001)109
Normal-weight (7 male, 1 female) volunteers Exendin-4 (0.05 pmol/kg/min)
19% reduction in caloric intake at test meal Effects on appetite unclear
Cohen et al. (2003)112
Normal-weight (7 male, 6 female) volunteers Oxyntomodulin iv infusion of 3.0 pmol/kg/min for 90 min
Reduced caloric intake by 19.3% at test meal Reduced hunger
Wynne et al. (2005)114
Overweight and obese (4 male, 22 female) volunteers Oxyntomodulin injections self-administered 400 nmol for 4 weeks, three times daily, 30 min before meals
Reduced caloric intake by 25% and 36% at test meals at start and end of study respectively No effects on hunger, nausea or food palatability Measures of satiety not included
Neary et al. (2005)195
Normal-weight (4 male, 6 female) volunteers Subanorectic doses of GLP-1 (0.4 pmol/kg/min) and PYY3–36 (0.4 pmol/kg/min)
A reduction of 27% in caloric intake during co-administration
Wynne et al. (2006)113
Overweight and obese (4 male, 11 female) volunteers Oxyntomodulin injection 400 nmol given for 4 days three times daily
Reduced caloric intake by 17.3% at test meal on day 4 No effects on hunger, nausea or food palatability Measures of satiety not included
Abbreviations: CCK, cholecystokinin; GLP-1, glucagon-like peptide 1; iv, intravenous; PYY, peptide YY.
reviews
nrendo_19_MAY10.indd 259 30/3/10 15:43:57
© 20 Macmillan Publishers Limited. All rights reserved10
260 | may 2010 | volume 6 www.nature.com/nrendo
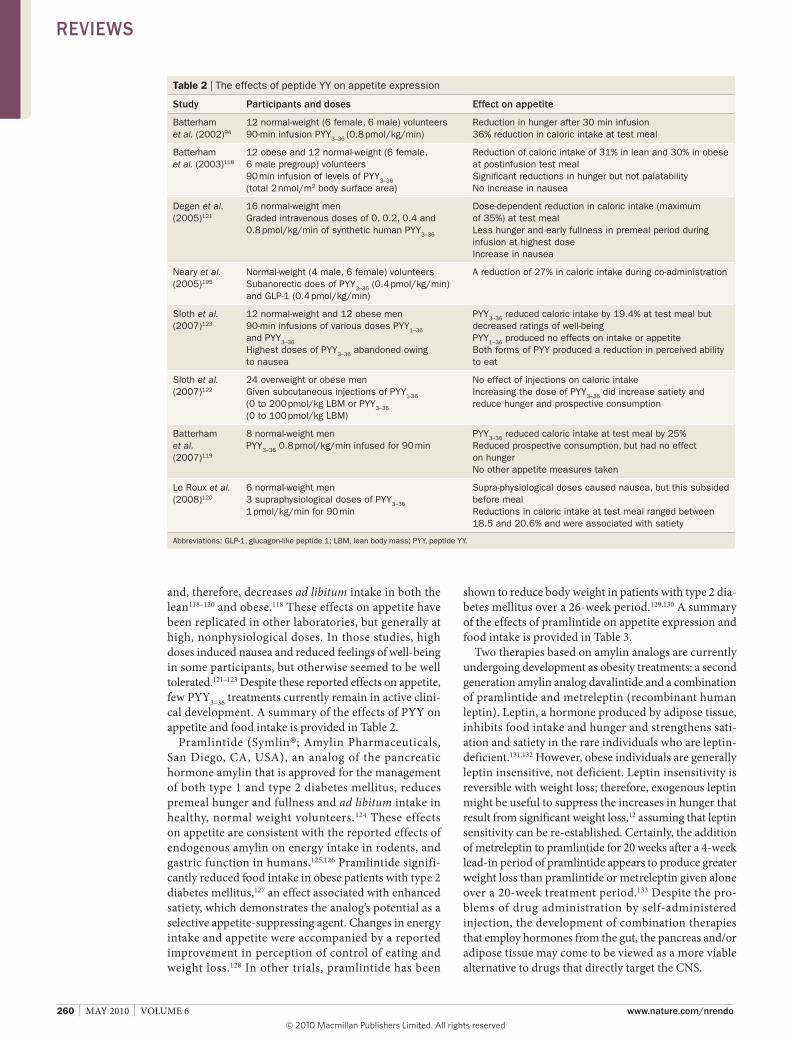
and, therefore, decreases ad libitum intake in both the lean118–120 and obese.118 these effects on appetite have been replicated in other laboratories, but generally at high, nonphysiological doses. in those studies, high doses induced nausea and reduced feelings of well-being in some participants, but otherwise seemed to be well tolerated.121–123 Despite these reported effects on appetite, few Pyy3–36 treatments currently remain in active clini-cal development. a summary of the effects of Pyy on appetite and food intake is provided in table 2.
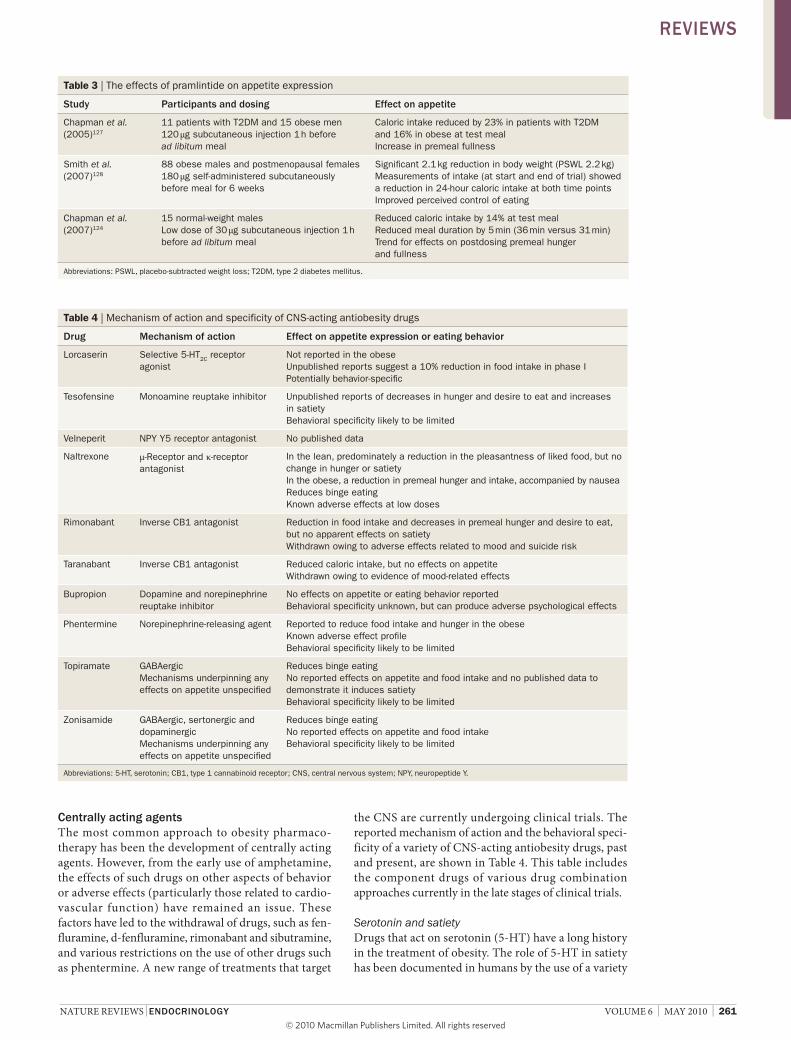
Pramlintide (symlin®; amylin Pharmaceuticals, san Diego, Ca, usa), an analog of the pancreatic hormone amylin that is approved for the management of both type 1 and type 2 diabetes mellitus, reduces premeal hunger and fullness and ad libitum intake in healthy, normal weight volunteers.124 these effects on appetite are consistent with the reported effects of endogenous amylin on energy intake in rodents, and gastric function in humans.125,126 Pramlintide signifi-cantly reduced food intake in obese patients with type 2 diabetes mellitus,127 an effect associated with enhanced satiety, which demon strates the analog’s potential as a selective appetite-suppressing agent. Changes in energy intake and appetite were accompanied by a reported improvement in perception of control of eating and weight loss.128 in other trials, pramlintide has been
shown to reduce body weight in patients with type 2 dia-betes mellitus over a 26-week period.129,130 a summary of the effects of pramlintide on appetite expression and food intake is provided in table 3.
two therapies based on amylin analogs are currently undergoing development as obesity treatments: a second generation amylin analog davalintide and a combination of pramlintide and metreleptin (recombinant human leptin). leptin, a hormone produced by adipose tissue, inhibits food intake and hunger and strengthens sati-ation and satiety in the rare individuals who are leptin-deficient.131,132 However, obese individuals are generally leptin insensitive, not deficient. leptin insensitivity is reversible with weight loss; therefore, exogenous leptin might be useful to suppress the increases in hunger that result from significant weight loss,12 assuming that leptin sensitivity can be re-established. Certainly, the addition of metreleptin to pramlintide for 20 weeks after a 4-week lead-in period of pramlintide appears to produce greater weight loss than pramlintide or metreleptin given alone over a 20-week treatment period.133 Despite the pro-blems of drug administration by self-administered injection, the development of combination therapies that employ hormones from the gut, the pancreas and/or adipose tissue may come to be viewed as a more viable al ternative to drugs that directly target the Cns.
Table 2 | The effects of peptide YY on appetite expression
study Participants and doses effect on appetite
Batterham et al. (2002)94
12 normal-weight (6 female, 6 male) volunteers 90-min infusion PYY3–36 (0.8 pmol/kg/min)
Reduction in hunger after 30 min infusion 36% reduction in caloric intake at test meal
Batterham et al. (2003)118
12 obese and 12 normal-weight (6 female, 6 male pregroup) volunteers 90 min infusion of levels of PYY3–36 (total 2 nmol/m2 body surface area)
Reduction of caloric intake of 31% in lean and 30% in obese at postinfusion test meal Significant reductions in hunger but not palatability No increase in nausea
Degen et al. (2005)121
16 normal-weight men Graded intravenous doses of 0, 0.2, 0.4 and 0.8 pmol/kg/min of synthetic human PYY3–36
Dose-dependent reduction in caloric intake (maximum of 35%) at test meal Less hunger and early fullness in premeal period during infusion at highest dose Increase in nausea
Neary et al. (2005)195
Normal-weight (4 male, 6 female) volunteers Subanorectic does of PYY3–36 (0.4 pmol/kg/min) and GLP-1 (0.4 pmol/kg/min)
A reduction of 27% in caloric intake during co-administration
Sloth et al. (2007)123
12 normal-weight and 12 obese men 90-min infusions of various doses PYY1–36
and PYY3–36 Highest doses of PYY3–36 abandoned owing to nausea
PYY3–36 reduced caloric intake by 19.4% at test meal but decreased ratings of well-being PYY1–36 produced no effects on intake or appetite Both forms of PYY produced a reduction in perceived ability to eat
Sloth et al. (2007)122
24 overweight or obese men Given subcutaneous injections of PYY1-36 (0 to 200 pmol/kg LBM or PYY3–36 (0 to 100 pmol/kg LBM)
No effect of injections on caloric intake Increasing the dose of PYY3–36 did increase satiety and reduce hunger and prospective consumption
Batterham et al. (2007)119
8 normal-weight men PYY3–36 0.8 pmol/kg/min infused for 90 min
PYY3–36 reduced caloric intake at test meal by 25% Reduced prospective consumption, but had no effect on hunger No other appetite measures taken
Le Roux et al. (2008)120
6 normal-weight men 3 supraphysiological doses of PYY3–36 1 pmol/kg/min for 90 min
Supra-physiological doses caused nausea, but this subsided before meal Reductions in caloric intake at test meal ranged between 18.5 and 20.6% and were associated with satiety
Abbreviations: GLP-1, glucagon-like peptide 1; LBM, lean body mass; PYY, peptide YY.
reviews
nrendo_19_MAY10.indd 260 30/3/10 15:43:58
© 20 Macmillan Publishers Limited. All rights reserved10
nature reviews | endocrinology volume 6 | may 2010 | 261
centrally acting agentsthe most common approach to obesity pharmaco-therapy has been the development of centrally acting agents. However, from the early use of amphetamine, the effects of such drugs on other aspects of behavior or adverse effects (particularly those related to cardio-vascular function) have remained an issue. these factors have led to the withdrawal of drugs, such as fen-fluramine, d- fenfluramine, rimonabant and sibutramine, and various restrictions on the use of other drugs such as phentermine. a new range of treatments that target
the Cns are currently undergoing clinical trials. the reported mechanism of action and the behavioral speci-ficity of a variety of Cns-acting antiobesity drugs, past and present, are shown in table 4. this table includes the component drugs of various drug combination approaches currently in the late stages of clinical trials.
Serotonin and satietyDrugs that act on serotonin (5-Ht) have a long history in the treatment of obesity. the role of 5-Ht in satiety has been documented in humans by the use of a variety
Table 3 | The effects of pramlintide on appetite expression
study Participants and dosing effect on appetite
Chapman et al. (2005)127
11 patients with T2DM and 15 obese men 120 μg subcutaneous injection 1 h before ad libitum meal
Caloric intake reduced by 23% in patients with T2DM and 16% in obese at test meal Increase in premeal fullness
Smith et al. (2007)128
88 obese males and postmenopausal females 180 μg self-administered subcutaneously before meal for 6 weeks
Significant 2.1 kg reduction in body weight (PSWL 2.2 kg) Measurements of intake (at start and end of trial) showed a reduction in 24-hour caloric intake at both time points Improved perceived control of eating
Chapman et al. (2007)124
15 normal-weight males Low dose of 30 μg subcutaneous injection 1 h before ad libitum meal
Reduced caloric intake by 14% at test meal Reduced meal duration by 5 min (36 min versus 31 min) Trend for effects on postdosing premeal hunger and fullness
Abbreviations: PSWL, placebo-subtracted weight loss; T2DM, type 2 diabetes mellitus.
Table 4 | Mechanism of action and specificity of CNS-acting antiobesity drugs
drug Mechanism of action effect on appetite expression or eating behavior
Lorcaserin Selective 5-HT2C receptor agonist
Not reported in the obese Unpublished reports suggest a 10% reduction in food intake in phase I Potentially behavior-specific
Tesofensine Monoamine reuptake inhibitor Unpublished reports of decreases in hunger and desire to eat and increases in satiety Behavioral specificity likely to be limited
velneperit NPY Y5 receptor antagonist No published data
Naltrexone μ-Receptor and κ-receptor antagonist
In the lean, predominately a reduction in the pleasantness of liked food, but no change in hunger or satiety In the obese, a reduction in premeal hunger and intake, accompanied by nausea Reduces binge eating Known adverse effects at low doses
Rimonabant Inverse CB1 antagonist Reduction in food intake and decreases in premeal hunger and desire to eat, but no apparent effects on satiety Withdrawn owing to adverse effects related to mood and suicide risk
Taranabant Inverse CB1 antagonist Reduced caloric intake, but no effects on appetite Withdrawn owing to evidence of mood-related effects
Bupropion Dopamine and norepinephrine reuptake inhibitor
No effects on appetite or eating behavior reported Behavioral specificity unknown, but can produce adverse psychological effects
Phentermine Norepinephrine-releasing agent Reported to reduce food intake and hunger in the obese Known adverse effect profile Behavioral specificity likely to be limited
Topiramate GABAergic Mechanisms underpinning any effects on appetite unspecified
Reduces binge eating No reported effects on appetite and food intake and no published data to demonstrate it induces satiety Behavioral specificity likely to be limited
Zonisamide GABAergic, sertonergic and dopaminergic Mechanisms underpinning any effects on appetite unspecified
Reduces binge eating No reported effects on appetite and food intake Behavioral specificity likely to be limited
Abbreviations: 5-HT, serotonin; CB1, type 1 cannabinoid receptor; CNS, central nervous system; NPY, neuropeptide Y.
reviews
nrendo_19_MAY10.indd 261 30/3/10 15:43:59
© 20 Macmillan Publishers Limited. All rights reserved10

262 | may 2010 | volume 6 www.nature.com/nrendo
of drugs such as the 5-Ht releaser d-fenfluramine, the 5-Ht reuptake inhibitor fluoxetine, and 5-Ht recep-tor agonists meta-chlorophenylpiperazine (mCPP) and sumatriptan.57 in the 1980s, fenfluramine was widely used to treat obesity, and during the 1990s, the more 5-Ht-selective d-fenfluramine also became available. However, in 1997, all forms of fenfluramine were with-drawn owing to the increased risk of cardiac valvulo-pathy and pulmonary hypertension associated with the use of peripheral 5-Ht2B receptors.134,135 sibutramine, a selective serotonergic and noradrenergic reuptake inhibi-tor, was until recently globally licensed to treat obesity. sibutramine had been shown to reduce food intake136,137 and enhance within-meal satiation in the obese.64 the drug also reduces the number of binges or the amount of food ingested during binges in patients with binge-eating disorder.138–140 interestingly, sibutramine seems to be more effective in obese individuals with poor regula-tion of eating behavior, particularly those who have low levels of cognitive dietary restraint.88 nonetheless, similar to many other monoaminergic treatments, sibutramine commonly produces transient increases in blood pres-sure and heart rate and has very occasionally been associ-ated with adverse psychological events.141,142 the effects on blood pressure and heart rate preclude the drug’s use in patients with cardio vascular problems, although the long-term outcome of sibutramine treatment in this at-risk group is currently under assessment.143
Despite the adverse effect issues that surround the use of fenfluramine and sibutramine, certain 5-Ht receptors appear to perform key roles in facilitating anorexigenic or inhibiting orexigenic inputs to key hypothalamic nuclei. Data from Heisler and colleagues demonstrate the critical role of 5-Ht2C and 5-Ht1B receptors in the inhibition of orexigenic and stimulation of anorexigenic neurons that mediate the effects of leptin on energy balance.144,145 in humans, the drug mCPP (a preferen-tial agonist of the 5-Ht1B/2C receptor) has been shown in a number of studies to reduce hunger and enhance satiety, an effect associated with a significant reduction in food intake.146,147 the drug also induced weight loss in obese individuals over 2 weeks of treatment.148 However, mCPP also produces transient increases in blood pres-sure and heart rate149 and increases lightheadedness, anxiety and nausea.146,147
the development of sufficiently specific 5-Ht2C ago-nists with little activity at 5-Ht2B or 5-Ht2a receptors has proven difficult, with nearly all compounds failing in early clinical trials for reasons of efficacy or issues of safety. Despite this fact, one selective 5-Ht2C agonist, lorcaserin has completed a number of phase iii clinical trials.150,151 this drug has little affinity for 5-Ht2B recep-tors and, equally importantly, little affinity for 5-Ht2a receptors, which reduces the likelihood of adverse psycho logical effects. Phase ii clinical data demonstrated that 12 weeks of treatment with lorcaserin, at various doses (10 mg daily, 15 mg daily and 10 mg twice a day), significantly reduced body weight.151 no unwarranted psychological or cardiovascular effects were reported; the most frequent adverse events being headache, nausea and
dizziness. these adverse effects were reported mainly at the start of the trial.
no published effects of lorcaserin on appetite are currently available, and it is assumed that the drug is con sidered to act on satiety. while placebo-subtracted weight loss appears modest, the selectivity of the drug should limit unwarranted behavioral or psychological adverse effects, making it a safer and more tolerable therapeutic alternative than existing monoaminergic therapies and possibly other Cns-acting agents currently under development. whether a potentially modest effect of lorcaserin on satiety is sufficient to have an impact on hedonically driven or poorly controlled eating behavior remains to be seen.
other drugs that act on 5-Ht that are under develop-ment include the monoamine reuptake inhibitor teso-fensine, which increases synaptic levels of dopamine, norepinephrine and 5-Ht.152,153 tesofensine, initially developed to treat neurodegenerative disease, pro-duced weight loss in early clinical studies.154 results of a 24-week, phase ii obesity study of tesofensine have recently been published.155 significant weight loss was reported and at the highest doses the placebo-subtracted weight loss appeared far greater than that produced by currently approved drugs. Dose-dependent decreases in hunger and increases in satiety are mentioned but not detailed in the original paper, while reductions in satiety and desire to eat are reported in a separate conference abstract.156 trends in anger and hostility and significant effects on confusion were reported during the trial along with increases in blood pressure and heart rate. the researchers declared, however, that no serious psychi atric adverse reactions (agitation, panic attacks and mood disorders) were observed during the trial, even though their sample included patients with a history of anxiety or depression.157,158
the therapeutic potential implied by the appetite data is interesting, as the developers believe that the drug not only affects serotonergic mechanisms tradition-ally believed to underpin satiation and satiety, but also modu lates dopaminergic mechanisms believed to be involved in ‘wanting’ (possibly reflected in the reported reduction in desire to eat). Given this potential dual action on appetite, tesofensine could produce more profound effects on eating behavior than lorcaserin, perhaps reflecting apparent differences in the magni-tude of weight loss reported in clinical trials. However, the mechanisms underlying the effects of tesofensine on either satiety or wanting remain to be charac terized. Certainly, it would be premature to attribute any effects on appetite to specific neurotransmitter systems or motivational processes, given the lack of relevant pre-clinical research. indeed, how reduction of dopamine reuptake might be translated into a reduction in wanting is unclear, since dopamine activation is regarded as being critical to this aspect of appetite.
For a drug entering the later stages of clinical develop-ment the lack of published behavioral pharmaco logy on tesofensine is surprising, but does exemplify the limitations of current approaches to the development
reviews
nrendo_19_MAY10.indd 262 30/3/10 15:44:00
© 20 Macmillan Publishers Limited. All rights reserved10

nature reviews | endocrinology volume 6 | may 2010 | 263
of antiobesity agents that we have outlined. overall, few appetite data are available on tesofensine and, despite denials,158 the reported adverse effect profile by astrup et al.155 seems to indicate some nonspecific psychological and behavioral effects.
Neuropeptides and energy regulationtherapeutically targeting the key systems that integrate episodic and tonic signals critical to energy regulation has the potential to produce profound effects on eating behavior.159,160 since 1995, various neuropeptides that mediate the effects of leptin on energy balance have been reported. the effect of leptin on neurons that project from the arcuate nucleus to a number of hypo-thalamic nuclei, including the paraventricular nucleus has been of particular interest. the anorexigenic PomC–Cart containing neurons and the orexigenic nPy–agrP contain ing neurons appear to provide a key con vergence point for peripherally generated epi-sodic signals (such as 5-Ht) and tonic signals such as leptin.14,145 in turn, these neurons have reciprocal con-nections with other appetite-regulating systems, such as the hypo thalamic melanin-concentrating hormone (mCH) system and orexin-expressing populations of the lateral hy pothalamic area.160
on the basis of studies of hypothalamic circuits, a variety of potentially satiety-enhancing drugs have been developed, including nPy and mCH1 receptor antago-nists and melanocortin receptor 4 agonists. a number of these drugs have reached clinical trials.161,162 How-ever, it seems many of these programs have yet to come to fruition or have been abandoned, either because the promising results from rodent trials could not be repli-cated in humans (for example, several candidate mCH1 antagonists), or because the efficacy in phase i trials was not maintained in phase ii trials (for example, a nPy y5 receptor antagonist candidate56). the reasons for the differences in efficacy between species or study duration are unclear. nonetheless, a number of drugs still appear to be in active clinical development including the nPy y5 receptor antagonist velneperit. unpublished data indicate that, in a recent phase iib trial of velneperit of 1-year duration, participants responded with a ≥5% weight loss.163,164
unfortunately, behavioral data on these drugs, irrespec tive of their success or failure, remain elusive. Given the many Cns systems implicated in the effects of endogenous leptin on energy balance, the benefit gained from targeting any system in isolation may be limited. nevertheless, given the potentially powerful effects of leptin on appetite expression (hunger, sati ation and satiety), directly targeting elements within this circuitry could form an important component of future thera-peutic interventions. ultimately, however, we must be aware of the all too frequent interpretation of intake reduction in animal models as representing specific actions on appetite control mechanisms. the transla-tion from animal paradigms to clinical development fre-quently lacks any detailed analysis of behavioral and/or motivational changes—in either animals or people.
Nonsatiety approachesGiven the role of hedonic experience in obesity, par-ticularly the enjoyment of energy-dense, high-fat foods, approaches that focus solely on satiation and satiety have potential limitations. in other words, not all drugs that reduce the motivation to eat do so through actions on satiety. therapeutically targeting Cns systems associated with the pleasure or rewarding aspects of food intake have also been assessed. one candidate system is the endogenous opioid system, as evidence mounts that μ, δ and κ opioid receptors located in reward-related brain areas are involved in the short-term control of feeding.
the distinct effects produced by opioid antagonists on the liking of pleasant and preferred foods are well docu-mented in human laboratory studies. naloxone, a general opioid receptor antagonist, reduces food palatability in the obese.165 in lean individuals, nonselective antago-nists, naltrexone and nalmefene reduce palatability, food pleasant ness and fat intake.166–169 these effects on hedonic evaluation of foods seem to be selective, since they gen-erally occur without any effect on ratings of hunger or satiety. Currently, naltrexone is a component of a com-bination therapy (Contrave®; orexigen therapeutics, la Jolla, Ca, usa) that is in clinical development.
early studies of naltrexone in obese individuals sug-gested that the drug reduced premeal hunger and intake but did not promote satiation or satiety, although treat-ment was also accompanied by nausea.170 naltrexone has also been shown to reduce binge eating.171 the precise effects of long-term naltrexone treatment on the moti-vation to eat in individuals with obesity remain to be characterized. Data from early clinical trials suggested that naltrexone produced minimal weight loss as mono-therapy172 and, therefore, may produce little effect on food intake. However, most studies in this area preceded the development of models of opioid involvement in reward processes and failed to specifically assess opioid antagonist effects in situations where they would most probably exert their effects, such as on the consumption of palatable foods. in light of our current knowledge of the role of opioid peptide systems in food liking, more detailed analysis of the effects of antagonists on appetite measures would be timely.
research in the past decade suggests that the endo-cannabinoids, such as anandamide, may be critical to the anticipation and wanting of food and, through inter-actions with the opioid peptides, the liking of food.173 in laboratory studies, the cannabinoid receptor agonist Δ9-tetrahydrocannabinol (tHC), derived from Cannabis sativa, has been shown to amplify premeal hunger and increase the ingestion of preferred foods.174,175 the influence of tHC on feeding behavior is mediated by the canna binoid CB1 receptor and reflects the speci-fic actions of the endocannabinoids on eating motiva-tion that have been demonstrated in animal models. thus, the blockade of cannabinoid receptors should sup-press the desire to eat and possibly reduce the enjoy ment experienced during the consumption of palatable food with a consequent reduction of energy intake. indeed, a number of endocannabinoid CB1 receptor inverse
reviews
nrendo_19_MAY10.indd 263 30/3/10 15:44:02
© 20 Macmillan Publishers Limited. All rights reserved10

264 | may 2010 | volume 6 www.nature.com/nrendo
agonists and antagonists, including rimonabant and taranabant, were developed as antiobesity agents. many of these drugs produced significant placebo-controlled weight loss and improvements in obesity disease-related risk factors that were superior or at least equivalent to the effects of currently licensed treatments.176–179
Despite the well-known hyperphagic effects of cannabi-noid receptor agonists, the effects of CB1 antagonists on actual eating behavior and appetite have only been reported in two trials. in line with the proposed role of the endocannabinoids and the actions of CB1 agonists, rimonabant reduced food intake, premeal hunger and desire to eat with little effect on satiety, which suggests that the blockade of endocannabinoid receptors specifically suppresses the motivation to initiate eating.180 taranabant also reduced caloric intake, but the effect could not be attributed to any change in appetite expression—at least as assessed by limited retr ospective measures.181 rimonabant was eventually licensed in a number of coun-tries for the treatment of obesity. However, in october 2008, it was recommended that rimonabant should no longer be prescribed in europe owing to an increased risk of anxiety and depressive disorders, including suicidality. advanced CB1 antagonist and inverse agonist programs were discontinued by all the major pharmaceutical com-panies. Given the psychological adverse effects associated with these drugs, the inadequate characterization of their behavioral effects in early clinical research appears to have contributed markedly to their eventual failure.
CNS combination drug approachesselectively targeting newly-discovered systems within the Cns has produced drugs that, in the case of some centrally-acting neuropeptide agonists or antagonists, lack the expected efficacy, or in the case of the cannabinoid receptor antagonists, promote unwanted psycho logical adverse effects. to a large extent, these failures reflect the premature targeting of those systems, before sufficient basic research could produce the theoretical models neces sary for the development of efficacious drug thera-pies. By contrast, exploitation of established, more com-pletely characterized neurochemical or endocrine systems could reduce the occurrence of un anticipated adverse effects. moreover, use of drugs with a broad spectrum of pharmacological effects or combining drugs with distinct pharmacological actions has the potential to increase treatment efficacy. monoamine reuptake inhibi tors such as tesofensine should influence a variety of systems that underpin the motivation to eat, such as the dopaminergic system implicated in the wanting of pleasur able food, and not just enhance satiety through the serotonergic system. By use of this rationale, a number of combinations using drugs already licensed for other indications have reached phase iii trials. these include a bupropion–naltrexone combination (Contrave®), a phentermine–topiramate combination (Qnexa®; vivus, mountain view, Ca, usa) and a bupropion–zonisamide sr combination (empatic®; orexigen therapeutics, la Jolla, Ca, usa).
Considering Contrave®, the combination of naltrexo ne and bupropion is designed to combine the separate
anorectic properties of the two drugs, while avoiding the high-doses of each individual compound that may reduce food intake or body weight, but can also be associ ated with adverse effects.182 Clinical trials indicate that the combination of drugs produces supra-additive effects on weight loss. the specific mechanisms through which this combination suppresses appetite and reduces body weight are unknown, but the development of Contrave® rests on the findings of animal and in vitro studies which suggest that naltrexone increases the release of the melanocortin-4 receptor agonist α-msH—an anorexi-genic peptide. the dopamine and norepinephrine reuptake inhibitor bupropion is also thought to stimulate the release of α-msH. via this mecha nism, bupropion, also used as an anti depressant, could reduce energy intake and increase energy expenditure.183 in turn, this may explain the weight loss produced in obese patients.184,185 meta-analysis of these effects suggests that bupropion placebo-subtracted weight loss would be 2.8 kg over 6 to 12 months.186 although the combined therapy appears to be well tolerated, adverse effects previously associ ated with bupropion include tachycardia arrhythmias and hypertension. whatever the mechanism, it is clear once again that the specific actions of each drug on psycho-logical processes in appetite have largely been ignored in the develop ment of this antiobesity application.
the combination therapy Qnexa®, currently the most efficacious weight-loss drug on trial, contains phen-termine, which also has well-documented adverse effects. Phentermine is a norepinephrine-releasing agent that has been used in weight control since the late 1950s. For a brief period it was given in combination with fen-fluramine. Compared to amphetamine, phentermine has low abuse potential, but at higher doses issues of cardio-vascular stimulation, insomnia, anxiety and irritability remain, and the drug is generally not recommended for routine use. Few behavioral studies of phentermine have been conducted, but an early study suggested that the drug reduced food intake in the obese by reduc-ing hunger.187 one potential mechanism of action is via phentermine-stimulated release of catecholamines (including dopamine and norepinephrine), which are proposed to indirectly raise leptin levels. Catecholamines may also reduce levels of orexigenic nPy independently of their effects on leptin. Given the promising phase ii trial results of combinational therapies, the effects of both phentermine and bupropion on human appetite warrant further study.
anticonvulsants such as topiramate and zonisamide induce weight loss.188–192 in a 2-year trial of topira-mate, patients with obesity who received a daily dose of 256 mg lost 9.7% of their body weight over a 60-week period compared with a loss of 1.7% in participants who received placebo.193 topiramate and zonisamide also reduce binge eating.194 However, both drugs produce adverse Cns-related adverse effects that limit toler-ability (including psychomotor difficulties); therefore, the development of topiramate as a monotherapy for obesity was dis continued. to date, the effects of topiramate and zo nisamide on food intake and eating behavior remain
reviews
nrendo_19_MAY10.indd 264 30/3/10 15:44:03
© 20 Macmillan Publishers Limited. All rights reserved10

nature reviews | endocrinology volume 6 | may 2010 | 265
unclear, as do the underlying pharmacological mecha-nisms. to attribute the action of these drugs to changes in the motivation to eat, the hedonic experience of ingestion, sati ation or satiety is entirely premature. nonetheless, these drugs remain components of two combination therapies in the final stages of clinical development.
Conclusionsa wide variety of treatments for type 2 diabetes mel-litus that target GlP-1 are undergoing clinical trials, and many of these may strengthen satiety and reduce intake. Conceivably, these treatments will form a new class of antiobesity agents. nausea is associated, at least in the short term, with the administration of many peri-pherally acting agents. Combinations of low-dose GlP-1 analogs with other gut peptides might strengthen the effect of treatment on appetite,195 while reducing the adverse effects profile. similarly, amylin-based treat-ments that target episodic satiety signals in combination with leptin could provide a novel approach that may effectively deal with the deleterious effects of chronic caloric restriction on appetite. one study suggests that leptin therapy reverses weight-loss-induced brain ac tivity in response to food cues.196 with the clinical failure of endo cannabinoid antagonists, current Cns drug develop-ment now re- emphasizes mono aminergic mechanisms. However, the only mono aminergic drug recently licensed to treat obesity, sibutramine, was withdrawn from euro-pean markets following a safety review by the european medicines agency, although the drug currently remains available in the us.
the precise effects of any of the novel mono aminergic drugs such as lorcaserin and tesofensine on appetite expression in the obese remain to be published.
irrespective of mechanism, current drug develop-ment fails to fully address the contribution of appetite and eating behavior in either the etiology or the treat-ment of obesity, despite behavioral and psychological factors having been shown to be key to successful weight loss and long-term weight management.88 some of the factors that contribute specifically to weight regain relate to appetite control; these include disinhibited eating, poorly controlled hunger, binge eating and eating in response to negative emotions and stress. although no pharma ceutical treatment approach can substitute for the behavioral change that individuals must commit to in their own lives to lose weight, interventions that allevi-ate the detri mental effects of dieting on appetite may be particularly effective in treating obesity.
Given that appetite is a dynamic system that responds to caloric restriction and that food preferences are habits that might be resistant to change, what evidence— beyond modest weight loss—is there that current anti-obesity drugs effectively help the obese manage their eating behavior?
Pharmaceutical approaches to the management of body weight tend to focus on simple end points, such as the absolute reduction in body weight over the course of treatment. weight loss can be the consequence of a reduction of energy intake, but we are often left guessing
as to the precise changes in behavior or motivation that might bring about reduced consumption. even now, many drug treatments progress to clinical trials simply on the basis of a demonstration of reduced food intake with consequent body-weight reduction.
a simple reduction of intake is all too often interpreted as an indication of the amplification of one or other of the numerous satiety signals that have been proposed to regu-late energy balance. such interpretations, in turn, derived from very simplistic homeostatic conceptualizations, ignore the complexity of human behavior and our psycho-logical relationship with food. However, these changes may actually occur through nonspecific actions of drugs, the lack of specificity often only receiving attention when nonspecific effects become reported as adverse effects that culminate in withdrawal of a treatment. admittedly, the pharmaceutical industry would face additional barriers to drug development if adequate behavioral protocols were incorporated into clinical testing. However, this would be a small cost to pay compared with the benefits of develop-ing an effective and selective antiobesity drug. Preclinical models of feeding behavior may at least provide an early indication of behavioral specificity.
a substantial body of science exists that relates the description and analysis of appetite and the changes in emotions, experiences, and motivations of people in rela-tion to food and its consumption. Differences between lean and obese individuals in their style of eating, sen-sitivity to food stimuli and enjoyment of food have been charac terized. these factors, which largely focus on our innate attraction to food and susceptibility to over-consumption, could provide important and effective targets for specific antiobesity drug treatments. as we have described, drugs exist that do act specifically on these psychological factors—and affect wanting and/or liking of food, for example. Closer attention to the subtleties of the eating process and the precise psychological effects of different drugs could conceivably lead to very direct manipulations that might be targeted towards the avoid-ance and reduced consumption of fattening foods. such increased scrutiny could also lead to the development of credible drug therapies that can directly aid benefi cial behavioral change in the overweight and obese.
Review criteria
The Review examines the effects of various medications on appetite expression in the obese, despite limited published data. The content of this Review is based on the collective knowledge of the authors and literature collected over the course of their careers. Thus, a formal literature search was not required to find the majority of the papers cited. However, to find the most recent papers and to confirm no relevant papers had been excluded, searches on Web of Knowledge and Scopus were conducted using search terms such as “hunger”, “palatability”, “wanting”, “liking”, “satiation”, “satiety”, “eating behavior” (US and UK spelling), “caloric intake”, “energy intake”, “food intake”, “food preferences” and “food choice” along with peptide, neurotransmitter or drug names. All papers quoted were written in English. No date limits were imposed in the searches.
reviews
nrendo_19_MAY10.indd 265 30/3/10 15:44:04
© 20 Macmillan Publishers Limited. All rights reserved10

266 | may 2010 | volume 6 www.nature.com/nrendo
1. Brown, W. J., Williams, L., Ford, J. H., Ball, K. & Dobson, A. J. Identifying the energy gap: magnitude and determinants of 5-year weight gain in mid-age women. Obes. Res. 13, 1431–1441 (2005).
2. National Institute for Clinical Excellence. Obesity: the prevention, identification, assessment and management of overweight and obesity in adults and children [online], http://guidance.nice.org.uk/CG43 (2006).
3. Pasman, W. J., Saris, W. H. & Westerterp-Plantenga, M. S. Predictors of weight maintenance. Obes. Res. 7, 43–50 (2006).
4. Keys, A., Brozek, J., Henschel, A., Mickelsen, O. & Taylor, H. L. The Biology of Human Starvation (2 volumes) (University of Minnesota Press, Minneapolis, 1950).
5. Warren, C. & Cooper, P. J. Psychological effects of dieting. Br. J. Clin. Psychol. 27, 269–270 (1988).
6. Ogden, J. & Evans, C. The problem with weighing: Effects of mood, self-esteem and body image. Int. J. Obes. Relat. Metab. Disord. 20, 272–277 (1996).
7. Laessle, R. G., Platte, P., Schweiger, U. & Pirke, K. M. Biological and psychological correlates of intermittent dieting behavior in young women. A model for bulimia nervosa. Physiol. Behav. 60, 1–5 (1996).
8. Halford, J. C. Clinical pharmacotherapy for obesity: current drugs and those in advanced development. Curr. Drug Targets 5, 637–646 (2004).
9. O’Donovan, D., Feinle-Bisset, C., Wishart, J. & Horowitz, M. Lipase inhibition attenuates the acute inhibitory effects of oral fat on food intake in healthy subjects. Br. J. Nutr. 90, 849–852 (2003).
10. Swhwizer, A. et al. Role of lipase in the regulation of upper gastrointestinal function in humans. Am. J. Physiol. 273, G612–G620 (1997).
11. Rosenbaum, M. et al. Low-dose leptin reverses sketetal muscle, autonomic, and neuroendocrine adaptation to maintenance of reduced body weight. J. Clin. Invest. 115, 3579–3586 (2005).
12. Mars, M., de Graaf, C., de Groot, C. P., van Rossum, C. T. & Kok, F. J. Fasting leptin and appetite responses induced by a 4-day 65%-energy-restricted diet. Int. J. Obes. 30, 122–128 (2006).
13. Blundell, J. E. & Halford, J. C. Regulation of nutrient supply: the brain and appetite control. Proc. Nutr. Soc. 53, 407–418 (1994).
14. Halford, J. C. & Blundell, J. E. Separate systems for serotonin and leptin in appetite control. Ann. Med. 32, 222–232 (2000).
15. Mela, D. J. Eating for pleasure or just wanting to eat? Reconsidering sensory hedonic responses as a driver of obesity. Appetite 47, 10–17 (2006).
16. Finlayson, G., King, N. & Blundell, J. E. Liking vs wanting food: importance for human appetite control and weight regulation. Neurosci. Biobehav. Rev. 31, 987–1002 (2007).
17. Rankinen, T. et al. The human obesity gene map: The 2005 update. Obesity 14, 529–644 (2006).
18. O’Rahilly, S. & Farooqi, I. S. Genetics of obesity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 361, 1095–1105 (2006).
19. Blundell, J. E. et al. Resistance and susceptibility to weight gain: individual variability in response to a high fat diet. Physiol. Behav. 86, 614–622 (2005).
20. Wardle, J. et al. Obesity associated genetic variation in FTO is associated with diminished satiety. J. Clin. Endocrinol. Metab. 93, 3640–3643 (2008).
21. Cecil, J. E., Tavendale, R., Watt, P., Hetherington, M. M. & Palmer, C. N. An obesity-
associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 359, 2558–2566 (2008).
22. Wardle, J., Llewellyn, C., Sanderson, S. & Plomin, R. The FTO gene and measured food intake in children. Int. J. Obes. 33, 42–45 (2009).
23. Haupt, A. et al. variation in the FTO gene influences food intake but not energy expenditure. Exp. Clin. Endocrinol. Diabetes 117, 194–197 (2009).
24. Spiegel, T. A., Wadden, T. A. & Foster, G. D. Objective measurement of eating rate during behavioral treatment of obesity. Behav. Ther. 22, 61–67 (1991).
25. Barkeling, B., Ekman, S. & Rössner, S. Eating behaviour in obese and normal weight 11-year-old children. Int. J. Obes. Relat. Metab. Disord. 16, 335–360 (1992).
26. Laessle, R. G., Lehrke, S. & Dückers, S. Laboratory eating behavior in obesity. Appetite 49, 399–404 (2007).
27. Meyer, J. E. & Pudel, v. Experimental studies on food-intake in obese and normal weight subjects. J. Psychosom. Res. 16, 305–308 (1972).
28. Stunkard, A. & Kaplan, D. Eating in public places: a review of reports of the direct observation of eating behavior. Int. J. Obes. 1, 89–101 (1977).
29. Näslund, E., Gutniak, M., Skogar, S., Rössner, S. & Hellström, P. M. Glucagon-like peptide 1 increases the period of postprandial satiety and slows gastric emptying in obese men. Am. J. Clin. Nutr. 68, 525–530 (1998).
30. Lindgren, A. C. et al. Eating behaviour in Prader–Willi syndrome, normal weight and obese control groups. J. Pediatr. 137, 50–55 (2000).
31. Williamson, D. A. et al. Microanalysis of eating behavior of three leptin deficient adults treated with leptin therapy. Appetite 45, 75–80 (2005).
32. Otsuka, R. et al. Eating fast leads to obesity: Findings based on self-administered questionnaires among middle-aged Japanese men and women. J. Epidemiol. 16, 117–124 (2006).
33. Maruyama, K. et al. The joint impact on being overweight of self reported behaviours of eating quickly and eating until full: cross sectional survey. BMJ 337, a2002 (2008).
34. Llewellyn, C. H., van Jaarsveld, C. H., Boniface, D., Carnell, S. & Wardle, J. Eating rate is a heritable phenotype related to weight in children. Am. J. Clin. Nutr. 88, 1560–1566 (2008).
35. Pearcey, S. M. & de Castro, J. M. Food intake and meal patterns of weight-stable and weight-gaining persons. Am. J. Clin. Nutr. 76, 107–112 (2002).
36. Rissanen, A. et al. Acquired preference especially for dietary fat and obesity: a study of weight-discordant monozygotic twin pairs. Int. J. Obes. Relat. Metab. Disord. 26, 973–977 (2002).
37. Bray, G. A., Paeratakul, S. & Popkin, B. M. Dietary fat and obesity: a review of animal, clinical and epidemiological studies. Physiol. Behav. 83, 549–555 (2004).
38. Kral, T. v., Roe, L. S. & Rolls, B. J. Combined effects of energy density and portion size on energy intake in women. Am. J. Clin. Nutr. 79, 962–968 (2004).
39. Geliebter, A. & Hashim, S. A. Gastric capacity in normal obese, and bulimic women. Physiol. Behav. 74, 743–746 (2001).
40. Geliebter, A., Yahav, E. K., Gluck, M. E. & Hashim, S. A. Gastric capacity, test meal intake, and appetitive hormones in binge eating disorder. Physiol. Behav. 81, 735–740 (2004).
41. Ranganath, L. R. et al. Attenuated GLP-1 secretion in obesity: cause or consequence? Gut 38, 916–919 (1996).
42. De Graaf, C., Blom, W. A., Smeets, P. A., Staflue, A. & Hendricks, H. F. Biomarkers of satiation and satiety. Am. J. Clin. Nutr. 17, 949–961 (2004).
43. Adam, T. C. & Westerterp-Plantenga, M. S. Glucagon-like peptide-1 release and satiety after a nutrient challenge in normal-weight and obese. Br. J. Nutr. 93, 845–851 (2005).
44. le Roux, C. W. et al. Attenuated peptide YY release in obese subjects is associated with reduced satiety. Endocrinology 147, 3–8 (2006).
45. Blundell, J. E. et al. Over-consumption and obesity: peptides and susceptibility to weight gain. Regul. Pept. 149, 32–38 (2008).
46. Matsuda, M. et al. Altered hypothalamic function in response to glucose ingestion in obese humans. Diabetes 48, 1801–1806 (1999).
47. Gautier, J. F. et al. Differential brain responses to satiation in obese and lean men. Diabetes 49, 838–846 (2000).
48. Carnell, S. & Wardle, J. Appetite and adiposity in children: evidence for a behavioral susceptibility theory of obesity. Am. J. Clin. Nutr. 88, 22–29 (2008).
49. Halford, J. C. et al. Beyond-brand effect of television food advertisements on food choice in children: the effects of weight status. Public Health Nutr. 11, 897–904 (2008).
50. Rothemund, Y. et al. Differential activation of the dorsal striatum by high-calorie visual food stimuli in obese individuals. Neuroimage 37, 410–421 (2007).
51. Stoeckel, L. E. et al. Widespread reward-system activation in obese women in response to pictures of high-calorie foods. Neuroimage 41, 636–647 (2008).
52. Drewnowski, A., Brunzell, J. D., Sande, K., Iverise, K. & Greenwood, M. R. Sweet tooth reconsidered: taste responsiveness in human obesity. Physiol. Behav. 35, 617–622 (1985).
53. Mela, D. J. & Sacchetti, D. A. Sensory preference for fats: relationships with diet and body composition. Am. J. Clin. Nutr. 53, 908–915 (1991).
54. Blundell, J. E. & Finlayson, G. Is susceptibility to weight gain characterized by homeostatic or hedonic risk factors for over-consumption? Physiol. Behav. 82, 21–25 (2004).
55. Hill, A. J., Rogers, P. J. & Blundell, J. E. Techniques for the experimental measurement of human eating behaviour and food intake: a practical guide. Int. J. Obes. Relat. Metab. Disord. 19, 361–375 (1995).
56. Blundell, J. E. et al. in Handbook of Assessment Methods for Eating Behaviors and Weight-Related Problems: Measures, Theory, and Research (eds Allison, D. B. & Baskin, M. L.) 283–326 (Sage Publications Inc., Thousand Oaks, 2009).
57. Halford, J. C., Harrold, J. A., Boyland, E. J., Lawton, C. L. & Blundell, J. E. Serotonergic drugs: effects on appetite expression and use for the treatment of obesity. Drugs 67, 27–55 (2007).
58. Flint, A., Raben, A., Blundell, J. E. & Astrup, A. Reproducibility, power and validity of visual analogue scales in assessment of appetite sensations in a single test meal studies. Int. J. Obes. Relat. Metab. Disord. 24, 38–48 (2000).
59. Stubbs, R. J. et al. The use of visual analogue scales to assess motivation to eat in human subjects: A review of their reliability and validity with an evaluation of new hand-held computerized systems for temporal tracking of appetite ratings. Br. J. Nutr. 84, 405–415 (2000).
reviews
nrendo_19_MAY10.indd 266 30/3/10 15:44:06
© 20 Macmillan Publishers Limited. All rights reserved10

nature reviews | endocrinology volume 6 | may 2010 | 267
60. Drapeau, v. et al. Appetite sensations as a marker of overall intake. Br. J. Nutr. 93, 273–280 (2005).
61. Drapeau, v. et al. Appetite sensations and satiety quotient: predictors of energy intake and weight loss. Appetite 48, 159–166 (2007).
62. Rogers, P. J. & Blundell, J. E. Effect of anorexic drugs on food intake and the micro-structure of eating in human subjects. Psychopharmacology 66, 159–165 (1979).
63. Yeomans, M. R. & Gray, R. W. Effects of naltrexone on food intake and changes in subjective appetite during eating: Evidence for opioid involvement in the appetizer effect. Physiol. Behav. 62, 15–21 (1997).
64. Halford, J. C. et al. The effects of sibutramine on the microstructure of feeding behaviour as measured by the Universal Eating Monitor (UEM). J. Psychopharmacol. 24, 99–109 (2010).
65. Erondu, N. et al. Neuropeptide Y5 receptor antagonism does not induce clinically meaningful weight loss in overweight and obese adults. Cell Metab. 4, 275–282 (2006).
66. King, N. A., Lawton, C. L., Delargy, H. J., Smith, F. C. & Blundell, J. E. in Ingestive Behavior Protocols (eds Wellmann, P. J. & Hoebel, B. G.) 71–76 (Society for the Study of Ingestive Behavior, New York, 1997).
67. Stubbs, R. J. et al. Description and evaluation of a Newton-based electronic appetite rating system for temporal tracking of appetite in human subjects. Physiol. Behav. 72, 615–619 (2001).
68. Bolger, N., Davis, A. & Rafaeli, E. Diary methods: capturing life as it is lived. Annu. Rev. Psychol. 54, 579–616 (2003).
69. Burton, C., Weller, D. & Sharpe, M. Are electronic diaries useful for symptoms research? A systematic review. J. Psychosom. Res. 62, 553–561 (2007).
70. Shiffman, S., Stone, A. A. & Hufford, M. R. Ecological momentary assessment. Annu. Rev. Clin. Psychol. 4, 1–32 (2008).
71. Ebner-Priemer, U. W. & Trull, T. J. Ambulatory assessment: An innovative and promising approach for clinical psychology. European Psychol. 14, 109–119 (2009).
72. Cappelleri, J. C. et al. Evaluating the Power of Food Scale in obese subjects and a general sample of individuals: development and measurement properties. Int. J. Obes. 33, 913–922 (2009).
73. Cappelleri, J. C. et al. Psychometric analysis of the Three-Factor Eating Questionnaire-R21: results from a large diverse sample of obese and non-obese participants. Int. J. Obes. 33, 611–620 (2009).
74. Karlsson, J., Persson, L. O., Sjöström, L. & Sullivan, M. Psychometric properties and factor structure of the three-factor eating questionnaire (TFEQ) in obese men and women, results from the Swedish obese subjects (SOS) study. Int. J. Obes. 24, 1715–1725 (2000).
75. Barkeling, B., King, N. A., Näslund, E. & Blundell, J. E. Characterization of obese individuals who claim to detect no relationship between their eating pattern and sensations of hunger or fullness. Int. J. Obes. 31, 435–439 (2007).
76. Provencher, v., Drapeau, v., Tremblay, A., Després, J. P. & Lemieux, S. Eating behaviors and indexes of body composition in men and women from the Quebéc family study. Obes. Res. 11, 783–792 (2003).
77. Dykes, J., Brunner, E. J., Martikeainene, P. T. & Wardle, J. Socioeconomic gradient in body size and oboist among women: The role of dietary
restraint, disinhibition and hunger in the Whitehall II study. Int. J. Obes. Relat. Metab. Disord. 28, 262–268 (2004).
78. Lawson, O. J. et al. The association of body weight, dietary intake, and energy expenditure with dietary restraint and disinhibition. Obes. Res. 3, 153–161 (1995).
79. Williamson, D. A. et al. Association of body mass with dietary restraint and disinhibition. Appetite 25, 31–41 (1995).
80. Hays, N. P. et al. Eating behaviour correlates of adult weight gain and obesity in healthy women aged 55–65 y. Am. J. Clin. Nutr. 75, 476–483 (2002).
81. Bellisle, F. et al. The eating inventory and body adiposity from leanness to massive obesity: A study of 2509 adults. Obes. Res. 12, 2023–2030 (2004).
82. Borg, P., Fogelholm, M. & Kukkonen-Harjula, K. Food selection and eating behaviour during weight maintenance intervention and 2-y follow-up in obese men. Int. J. Obes. Relat. Metab. Disord. 28, 1548–1554 (2004).
83. Provencher, v. et al. Familial resemblance in eating behaviors in men and women from the Quebéc family study. Obes. Res. 13, 1624–1629 (2005).
84. Bouchard, L. et al. Neuromedin β: a strong candidate gene linking eating behaviors and susceptibility to obesity. Am. J. Clin. Nutr. 80, 1478–1486 (2004).
85. Stunkard, A. J. Eating patterns in obesity. Psychiatr. Q. 33, 284–292 (1959).
86. Hsu, L. K. et al. Binge eating disorder in extreme obesity. Int. J. Obes. Relat. Metab. Disord. 26, 1398–1403 (2002).
87. Grucza, R. A., Przybeck, T. R. & Cloninger, C. R. Prevalence and correlates of binge eating disorder in a community sample. Compr. Psychiatr. 48, 124–131 (2007).
88. Elfhag, K. & Rössner, S. Who succeeds in maintaining weight loss? A conceptual review of factors associated with weight loss maintenance and weight regain. Obes. Rev. 6, 67–85 (2005).
89. Bryant, E. J., King, N. A. & Blundell, J. E. Disinhibition: its effects on appetite and weight regulation. Obes. Rev. 9, 409–419 (2008).
90. Elfhag, K., Rössner, S., Barkeling, B. & Rooth, P. Sibutramine treatment in obesity: initial eating behaviour in relation to weight loss results and changes in mood. Pharmacol. Res. 51, 159–163 (2005).
91. Elfhag, K., Finer, N. & Rössner, S. Who will lose weight on sibutramine and orlistat? Psychological correlates for treatment success. Diabetes Obes. Metab. 10, 498–505 (2008).
92. Wren, A. M. et al. Ghrelin enhances appetite and increases food intake in humans. J. Clin. Endocrinol. Metab. 86, 5992–5995 (2001).
93. Ballinger, A., McLoughlin, L., Medbak, S. & Clark, M. Cholecystokinin is a satiety hormone in humans at physiological postprandial plasma concentrations. Clin. Sci. 89, 375–381 (1995).
94. Batterham, R. L. et al. Gut hormone PYY(3–36) physiologically inhibits food intake. Nature 418, 650–654 (2002).
95. English, P. J., Ghatei, M. A., Malik, I. A., Bloom, S. R. & Wilding, J. P. Food fails to suppress ghrelin levels in obese humans. J. Clin. Endocrinol. Metab. 87, 2984–2987 (2002).
96. Jordan, J. et al. Stimulation of cholecystokinin-A receptors with GI181771X does not cause weight loss in overweight or obese patients. Clin. Pharmacol. Ther. 83, 281–287 (2008).
97. Bojanowska, E. Physiology and pathophysiology of glucagon-like peptide 1 (GLP-1): the role of GLP-1 in the pathogenesis of diabetes mellitus,
obesity, and stress. Med. Sci. Monit. 11, RA271–RA278 (2005).
98. Nauck, M. A. et al. Glucagon-like peptide-1 inhibition of gastric emptying outweighs its insulinotropic effects in healthy humans. Am. J. Physiol. Endocrinol. Metab. 273, E981–E988 (1997).
99. Little, T. J. et al. Effects of intravenous glucagon-like peptide-1 on gastric emptying and intragastric distribution in healthy subjects: relationships with postprandial glycemic and insulinemic responses. J. Clin. Endocrinol. Metab. 91, 1916–1923 (2006).
100. Flint, A., Raben, A., Astrup, A. & Holst, J. J. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J. Clin. Invest. 101, 515–520 (1998).
101. Gutzwiller, J. P. et al. Glucagon-like peptide-1: a potent regulator of food intake in humans. Gut 44, 81–86 (1999).
102. Degen, L. et al. Effects of a preload on reduction of food intake by GLP-1 in healthy subjects. Digestion 74, 78–84 (2006).
103. Gutzwiller, J. P., Degen, L., Matzinger, D., Prestin, S. & Beglinger, C. Interaction between GLP-1 and CCK-33 in inhibiting food intake and appetite in men. Am. J. Physiol. Regul. Comp. Physiol. 287, R562–R567 (2004).
104. Näslund, E., Gutniak, M., Skogar, S., Rössner, S. & Hellström, P. M. Glucagon-like peptide 1 increase the period of postprandial satiety and slows gastric emptying in obese men. Am. J. Clin. Nutr. 68, 525–530 (1998).
105. Näslund, E. et al. Energy intake and appetite are suppressed by glucagon-like peptide-1 (GLP-1) in obese men. Int. J. Obes. Relat. Metab. Disord. 23, 304–311 (1999).
106. Näslund, E. et al. Prandial subcutaneous injections of glucagon like peptide 1 cause weight loss in obese human subjects. Br. J. Nutr. 91, 439–446 (2004).
107. Gutzwiller, J. P. et al. Glucagon-like peptide-1 promotes satiety and reduced food intake in patients with diabetes mellitus. Am. J. Physiol. 276, R1541–R1545 (1999).
108. Flint, A., Raben, A., Ersbøll, A. K., Holst, J. J. & Astrup, A. The effect of physiological levels of glucagon-like peptide-1 on appetite, gastric emptying, energy and substrate metabolism in obesity. Int. J. Obes. Relat. Metab. Disord. 25, 781–792 (2001).
109. Edwards, C. M. et al. Exendin-4 reduces fasting and postprandial glucose and decreases energy intake in healthy volunteers. Am. J. Physiol. Endocrinol. Metab. 291, E155–E161 (2001).
110. Madsbad, S. Exenatide and liraglutide: different approaches to develop GLP-1 receptor agonists (incretin mimetics)—preclinical and clinical results. Best Pract. Res. Clin. Endocrinol. Metab. 23, 463–477 (2009).
111. Hanssen, K. B., Knop, F. K., Holst, J. J. & vilsbøll, T. Treatment of type 2 diabetes with glucagon-like peptide-1 receptor agonists. Int. J. Clin. Pract. 63, 1154–1160 (2009).
112. Cohen, M. A. et al. Oxyntomodulin suppresses appetite and reduced food intake in humans. J. Clin. Endocrinol. Metab. 88, 4696–4701 (2003).
113. Wynne, K. et al. Oxyntomodulin increases energy expenditure in addition to decreasing energy intake in overweight and obese humans: a randomised controlled trial. Int. J. Obes. 30, 1729–1736 (2006).
114. Wynne, K. et al. Subcutaneous oxyntomodulin reduces body weight in overweight and obese subjects: a double-blind, randomized, controlled trial. Diabetes 54, 2390–2395 (2005).
reviews
nrendo_19_MAY10.indd 267 30/3/10 15:44:13
© 20 Macmillan Publishers Limited. All rights reserved10

268 | may 2010 | volume 6 www.nature.com/nrendo
115. Onga, T., Zabielski, R. & Kato, S. Multiple regulation of peptide YY secretion in the digestive tract. Peptides 23, 279–290 (2002).
116. Renshaw, D. & Batterham, R. L. Peptide YY: a potential therapy for obesity. Curr. Drug Targets 6, 171–179 (2005).
117. Baggiano, M. M. et al. PYY3–36 as an anti-obesity drug target. Obes. Rev. 6, 307–322 (2005).
118. Batterham, R. L. et al. Inhibition of food intake in obese subjects by peptide YY3–36. N. Engl. J. Med. 349, 914–918 (2003).
119. Batterham, R. L. et al. PYY modulation of cortical and hypothalamic brain areas predicts feeding behaviour in humans. Nature 450, 106–109 (2007).
120. le Roux, C. W. et al. Supraphysiological doses of intravenous PYY3–36 cause nausea, but no additional reduction in food intake. Ann. Clin. Biochem. 45, 93–95 (2008).
121. Degen, L. et al. Effect of peptide PYY3–36 on food intake in humans. Gastroenterology 129, 1430–1436 (2005).
122. Sloth, B., Davidson, L., Holst, J. J., Flint, A. & Astrup, A. Effect of subcutaneous injections of PYY1–36 and PYY3–36 on appetite, ad libitum energy intake, and plasma free fatty acid concentration in obese males. Am. J. Physiol. Endocrinol. Metab. 293, E604–E609 (2007).
123. Sloth, B., Holst, J. J., Flint, A., Gregersen, N. T. & Astrup, A. Effects of PYY1–36 and PYY3–36 on appetite, energy intake, energy expenditure, glucose and fat metabolism in obese and lean subjects. Am. J. Physiol. Endocrinol. Metab. 292, E1062–E1068 (2007).
124. Chapman, I. et al. Low-dose pramlintide reduced food intake and meal duration in healthy, normal-weight subjects. Obesity 15, 1179–1186 (2007).
125. Reda, T. K., Geliebter, A. & Pi-Sunyer, F. X. Amylin, food intake and obesity. Obes. Res. 10, 1087–1091 (2002).
126. Lutz, T. A. Pancreatic amylin as a centrally acting satiating hormone. Curr. Drug Targets 6, 181–189 (2005).
127. Chapman, I. et al. Effect of pramlintide on satiety and food intake in obese subjects and subjects with type 2 diabetes. Diabetologia 48, 838–848 (2005).
128. Smith, S. R. et al. Pramlintide treatment reduces 24-h caloric intake and meal sizes and improves control of eating in obese subjects: a 6-wk translational research study. Am. J. Physiol. Endocrinol. Metab. 293, E620–E627 (2007).
129. Hollander, P. et al. Addition of pramlintide to insulin therapy lowers HbA1c in conjunction with weight loss in patients with type 2 diabetes approaching glycaemic targets. Diabetes Obes. Metab. 5, 408–414 (2003).
130. Hollander P. et al. Effect of pramlintide on weight in overweight and obese insulin treated type 2 diabetes patients. Obes. Res. 12, 661–668 (2004).
131. Farooqi, I. S. et al. Effects of recombinant leptin therapy in a child with congenital leptin deficiency. N. Engl. J. Med. 341, 879–884 (1999).
132. Williamson, D. A. et al. Microanalysis of eating behavior of three leptin deficient adults treated with leptin therapy. Appetite 45, 75–80 (2005).
133. Ravussin, E. et al. Enhanced weight loss with pramlintide/metreleptin: an integrated neurohormonal approach to obesity pharmacotherapy. Obesity 17, 1736–1743 (2009).
134. Abenhaim, L. et al. Appetite suppressant drugs and the risk of primary pulmonary hypertension. International Primary Pulmonary Hypertension Study Group. N. Engl. J. Med. 335, 609–616 (1996).
135. Rothman, R. B. & Baumann, M. H. Serotonergic drugs and valvular heart disease. Expert Opin. Drug Saf. 8, 317–329 (2009).
136. Rolls, B. J., Shide, D. J., Thorwart, M. L. & Ulbrecht, J. S. Sibutramine reduces food intake in non-dieting women with obesity. Obes. Res. 6, 1–11 (1998).
137. Barkeling, B., Elfhag, K., Rooth, P. & Rössner, S. Short-term effects of sibutramine (ReductilTM) on appetite and eating behaviour and the long-term therapeutic outcomes. Int. J. Obes. Relat. Metab. Disord. 27, 693–700 (2003).
138. Appolinario, J. C. et al. A randomized, double-blind, placebo-controlled study of sibutramine in the treatment of binge-eating disorder. Arch. Gen. Psychiatry 60, 1109–1116 (2003).
139. Appolinario, J. C. et al. An open-label trial of sibutramine in obese patients with binge-eating disorder. J. Clin. Psychiatry 63, 28–30 (2002).
140. Milano, W. et al. Use of sibutramine, an inhibitor of the reuptake of serotonin and noradrenaline, in the treatment of binge eating disorder: A placebo-controlled study. Adv. Ther. 22, 25–31 (2005).
141. Taflinski, T. & Chojnacka, J. Sibutramine-associated psychotic episode. Am. J. Psychiatry 157, 2057–2058 (2000).
142. Binkley, K. & Knowles, S. R. Sibutramine and panic attacks. Am. J. Psychiatry 159, 1793–1794 (2002).
143. Sharma, S. M. et al. Blood pressure changes associated with sibutramine and weight management – an analysis from the 6-week lead-in period of the sibutramine cardiovascular outcomes trial (SCOUT). Diabetes Obes. Metab. 11, 239–250 (2009).
144. Heisler, L. K. et al. Activation of central melanocortin pathways by fenfluramine. Science 297, 609–611 (2002).
145. Heisler, L. K. et al. Serotonin reciprocally regulates melanocortin neurons to modulate food intake. Neuron 51, 239–249 (2006).
146. Walsh, A. E. et al. m-Chlorophenylpiperazine decreases food intake in a test meal. Psychopharmacology 116, 120–122 (1994).
147. Cowen, P. J., Sargent, P. A., Williams, C., Goodall, E. M. & Orlikov, A. B. Hypophagic, endocrine and subjective responses to m-chlorophenylpiperazine in healthy-men and women. Hum. Psychopharmacol. 10, 385–391 (1995).
148. Sargent, P. A., Sharpley, A. L., Williams, C., Goodall, E. M. & Cowen, P. J. 5-HT2C activation decreases appetite and body weight in obese subjects. Psychopharmacology 133, 309–312 (1997).
149. Ghaziuddin, N., Welch, K. & Greden, J. Central serotonergic effects of m-chlorophenylpiperazine (mCPP) among normal control adolescents. Neuropsychopharmacology 28, 133–139 (2003).
150. Smith, B. M. et al. Discovery and SAR of new benzazepines as potent and selective 5-HT(2C) receptor agonists for the treatment of obesity. Bioorg. Med. Chem. Lett. 15, 1467–1470 (2005).
151. Smith, S. R. et al. Lorcaserin (APD356), a selective 5-HT(2C) agonist, reduces body weight in obese men and women. Obesity 17, 494–503 (2009).
152. Doggrell, S. A. Tesofensine—a novel potent weight loss medication. Expert Opinion Invest. Drugs 18, 1043–1046 (2009).
153. Bello, N. T. & Zahner, M. R. Tesofensine, a monoanime reuptake inhibitor for the treatment of obesity. Curr. Opinion Investig. Drugs 10, 1105–1116 (2009).
154. Hauser, R. A., Saline, L., Judel, N., Konyago, v. L. & The NS 2330 Monotherepy PC Study Group.
Randomized trial of the triple monoamine reuptake inhibitor NS 2330 (tesofensine) in early Parkinson’s disease. Mov. Disord. 22, 359–365 (2007).
155. Astrup, A. et al. Effect of tesofensine on bodyweight loss, body composition, and quality of life in obese patients: a randomised, double-blind, placebo-controlled trial. Lancet 372, 1906–1913 (2008).
156. Gasteyger, C., Sjödin, A., Meier, D. H. & Astrup, A. Effects of tesofensine on appetite in obese subjects. Obesity 16 (Suppl. 1), S138–S139 (2008).
157. Sommet, A., Pathak, A. & Montastruc, J. L. Tesofensine and weight loss. Lancet 373, 719 (2009).
158. Astrup, A., Madsbad, S. & Larsen, T. M. Tesofensine and weight loss—Authors’ reply. Lancet 373, 720 (2009).
159. Kalra, S. P. et al. Interacting appetite-regulating pathways in the hypothalamic regulation of body weight. Endocr. Rev. 20, 68–100 (1999).
160. Berthoud, H. R. & Morrison, C. The brain, appetite and obesity. Annu. Rev. Psychol. 59, 55–92 (2008).
161. Kamiji, M. M. & Inui, A. Neuropeptide Y receptor selective ligands in the treatment of obesity. Endocr. Rev. 28, 664–684 (2007).
162. Jeon, M. K. & Cheon, H. G. Promising strategies for obesity pharmacotherapy: melanocortin-4 (MC-4) receptor agonists and melanin concentrating hormone (MCH) receptor-1 antagonists. Curr. Top. Med. Chem. 9, 504–538 (2009).
163. ClinicalTrials.gov. Phase IIb one year efficacy and safety study of S-2367 (Velneperit) in obese subjects with initial 6-week low calorie diet (LDC) [online], http://clinicaltrials.gov/ct2/show/NCT00748605 (2009).
164. Shionogi & Co., Ltd. Shionogi announces positive top-line efficacy results from year-long studies of velneperit, a novel NPY Y5 receptor antagonist being investigated for the treatment of obesity [online], http://www.shionogi.co.jp/ir_en/news/detail/e_090217.pdf (2009).
165. Drewnowski, A., Krahn, D. D., Demitrack, M. A., Nairn, K. & Gosnell, B. A. Naloxone, an opiate blocker, reduces the consumption of sweet high-fat foods in obese and lean female binge eaters. Am. J. Clin. Nutr. 61, 1206–1212 (1995).
166. Yeomans, M. R. & Gray, R. W. Selective effects of naltrexone on food pleasantness and intake. Physiol. Behav. 60, 439–446 (1996).
167. Yeomans, M. R. & Gray, R. W. Effects of naltrexone on food intake and changes in subjective appetite during eating: Evidence for opioid involvement in the appetizer effect. Physiol. Behav. 62, 15–21 (1997).
168. Yeomans, M. R., Wright, P., Macleod, H. A. & Critchley, J. A. Effects of nalmefene on feeding in humans—dissociation of hunger and palatability. Psychopharmacology 100, 426–432 (1990).
169. Yeomans, M. R. & Wright, P. Lower pleasantness of palatable foods in nalmefene-treated human volunteers. Appetite 16, 249–259 (1991).
170. Speigal, T. A. et al. Effect of naltrexone on food intake, hunger, and satiety in obese men. Physiol. Behav. 40, 135–141 (1987).
171. Alger, S. A., Schwalberg, M. D., Bigaouette, J. M., Michalek, A. v. & Howard, L. J. Effect of a tricyclic antidepressant and opiate antagonist on binge-eating behavior in normoweight bulimic and obese, binge-eating subjects. Am. J. Clin. Nutr. 53, 865–871 (1991).
172. Lee, M. W. & Fujioka, K. Naltrexone for the treatment of obesity: review and update. Expert Opin. Pharmacother. 10, 1841–1845 (2009).
reviews
nrendo_19_MAY10.indd 268 30/3/10 15:44:16
© 20 Macmillan Publishers Limited. All rights reserved10

nature reviews | endocrinology volume 6 | may 2010 | 269
173. Kirkham, T. C. Endocannabinoids in the regulation of appetite and body weight. Behav. Pharmacol. 16, 297–313 (2005).
174. Haney, M., Ward, A. S., Comer, S. D., Foltin, R. W. & Fischman, M. W. Abstinence symptoms following oral THC administration to humans. Psychopharmacol. 141, 385–394 (1999).
175. Kirkham, T. C., Townson, A. L. & Halford, J. C. Oromucosal application of a cannabis extract accentuates pre-prandial hunger and increases caloric intake in healthy volunteers. Int. J. Obes. 31 (Suppl. 1), T2:PO.86, S99 (2007).
176. van Gaal, L. F., Rissanen, A. M., Scheen, A. J., Ziegler, O. & Rossner, S. Effects of the cannabinoid-1 receptor blocker rimonabant on weight reduction and cardiovascular risk factors in overweight patients: 1-year experience from the RIO-Europe study. Lancet 365, 1389–1397 (2005).
177. Pi-Sunyer, X., Despres, J. P., Scheen, A. & van Gaal, L. Improvement of metabolic parameters with rimonabant beyond the effect attributable to weight loss alone: Pooled 1-year data from the RIO (rimonabant in obesity and related metabolic disorders) program. J. Am. Coll. Cardiol. 47 (Suppl. A), 362A (2006).
178. Pi-Sunyer, F. X. et al. Effect of rimonabant, a cannabinoid-1 receptor blocker, on weight and cardiometabolic risk factors in overweight or obese patients: RIO-North America: A randomized controlled trial. JAMA. 295, 761–775 (2006).
179. Scheen, A. J., Finer, N., Hollander, P., Jensen, M. D. & van Gaal, L. F. Efficacy and
tolerability of rimonabant in overweight or obese patients with type 2 diabetes: a randomised controlled study. Lancet 368, 1660–1672 (2006).
180. Blundell, J. E. et al. Effect of rimonabant on energy intake, motivation to eat and body weight with or without hypocaloric diet: the REBA study. Obes. Rev. 7 (Suppl. 2), PP0021, 104 (2006).
181. Addy, C. et al. Multiple-dose pharmacokinetics, pharmacodynamics, and safety of taranabant, a novel selective cannabinoid-1 receptor inverse agonist, in healthy male volunteers. J. Clin. Pharmacol. 48, 734–744 (2008).
182. Plodowski, R. A. et al. Bupropion and naltrexone: a review of their use individually and in combination for the treatment of obesity. Expert Opin. Pharmacother. 10, 1069–1081 (2009).
183. Hausenloy, D. J. ContraveTM: a novel treatment for obesity. Clin. Lipidol. 4, 279–285 (2009).
184. Jain, A. K. et al. Buproprion SR vs placebo for weight loss in obese patients with depressive symptoms. Obes. Res. 10, 1049–1056 (2002).
185. Anderson, J. W. et al. Buproprion SR enhances weight loss: A 48-week double blind, placebo-controlled trial. Obes. Res. 10, 633–641 (2002).
186. Li, Z. et al. Meta-analysis: pharmacologic treatments of obesity. Ann. Intern. Med. 142, 532–546 (2005).
187. Silverstone, T. Anorectic effect of a long-acting preparation of phentermine (duromine). Psychopharmacologica 25, 315–320 (1972).
188. van Ameringen, M., Mancini, C., Pipe, B., Campbell, M. & Oakman, J. Topiramate treatment
for SSRI-incuded weight gain in anxiety disorders. J. Clin. Psychiatry 63, 981–984 (2002).
189. Ben-Menachem, A., Axelsen, M., Johanson, E. H., Stagge, A. & Smith, U. Predictors of weight loss in adults with topiramate-treated epilepsy. Obes. Res. 11, 556–562 (2003).
190. Gadde, K. M., Francisey, D. M., Wagner, H. R. & Krishnan, K. R. Zonisamide for weight loss in obese adults: a randomized controlled trial. JAMA 289, 1820–1825 (2003).
191. Appolinario, J. C. & McElroy, S. L. Pharmacological approaches in the treatment of binge eating disorder. Curr. Drug Targets 5, 301–307 (2004).
192. Bray, G. A. et al. A 6-month randomized, placebo-controlled, dose-ranging trial of topiramate for weight loss in obesity. Obes. Res. 11, 722–733 (2003).
193. Wilding, J. et al. A randomised double-blind placebo controlled study of the long-term efficacy and safety of topiramate in the treatment of obese subjects. Int. J. Obes. Relat. Metab. Disord. 28, 1399–1410 (2004).
194. McElroy, S. L. et al. Role of antiepileptic drugs in the management of eating disorders. CNS Drugs 23, 139–156 (2009).
195. Neary, N. M. et al. Peptide YY3–36 and glucagon-like peptide-1(7–36) inhibit food intake additively. Endocrinology 146, 5120–5127 (2005).
196. Rosenbaum, M., Sy, M., Pavlovich, K., Leibel, R. L. & Hirsch, J. Leptin reverses weight loss-induced changes in regional neural activity responses to visual food stimuli. J. Clin. Invest. 118, 2583–2591 (2008).Ut nos adiatinit,
reviews
nrendo_19_MAY10.indd 269 30/3/10 15:44:17
© 20 Macmillan Publishers Limited. All rights reserved10