
Phantom limb pain after unilateral arm amputation is ... - zora.uzh
21
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2022 Phantom limb pain after unilateral arm amputation is associated with decreased heat pain thresholds in the face Fuchs, Xaver ; Diers, Martin ; Trojan, Jörg ; Kirsch, Pinar ; Milde, Christopher ; Bekrater-Bodmann, Robin ; Rance, Mariela ; Foell, Jens ; Andoh, Jamila ; Becker, Susanne ; Flor, Herta Abstract: BACKGROUND The mechanisms underlying chronic phantom limb pain (PLP) are complex and insuffciently understood. Altered sensory thresholds are often associated with chronic pain but quantitative sensory testing (QST) in PLP has so far been inconclusive due to large methodological variation between studies and small sample sizes. METHODS In this study, we applied QST in 37 unilateral upper-limb amputees (23 with and 14 without PLP) and 19 healthy controls. We assessed heat pain (HPT), pressure pain, warmth detection and two-point discrimination thresholds at the residual limb, a homologous point and the thenar of the intact limb as well as both corners of the mouth. RESULTS We did not fnd signifcant diferences in any of the thresholds between the groups. However, PLP intensity was negatively associated with HPT at all measured body sites except for the residual limb, indicating lower pain thresholds with higher PLP levels. Correlations between HPT and PLP were strongest in the contralateral face (r = -0.65, p < 0.001). Facial HPT were specifcally associated with PLP, independent of residual limb pain (RLP) and various other covariates. HPT at the residual limb, however, were signifcantly associated with RLP, but not with PLP. CONCLUSION We conclude that the association between PLP and, especially facial, HPT could be related to central mechanisms. SIGNIFICANCE Phantom limb pain (PLP) is still poorly understood. We show that PLP intensity is associated with lower heat pain thresholds, especially in the face. This fnding could be related to central nervous changes in PLP. DOI: https://doi.org/10.1002/ejp.1842 Posted at the Zurich Open Repository and Archive, University of Zurich ZORA URL: https://doi.org/10.5167/uzh-212435 Journal Article Published Version The following work is licensed under a Creative Commons: Attribution-NonCommercial 4.0 International (CC BY-NC 4.0) License. Originally published at: Fuchs, Xaver; Diers, Martin; Trojan, Jörg; Kirsch, Pinar; Milde, Christopher; Bekrater-Bodmann, Robin; Rance, Mariela; Foell, Jens; Andoh, Jamila; Becker, Susanne; Flor, Herta (2022). Phantom limb pain after unilateral arm amputation is associated with decreased heat pain thresholds in the face. European Journal of Pain, 26(1):114-132.
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Phantom limb pain after unilateral arm amputation is ... - zora.uzh

Zurich Open Repository andArchiveUniversity of ZurichMain LibraryStrickhofstrasse 39CH-8057 Zurichwww.zora.uzh.ch
Year: 2022
Phantom limb pain after unilateral arm amputation is associated withdecreased heat pain thresholds in the face
Fuchs, Xaver ; Diers, Martin ; Trojan, Jörg ; Kirsch, Pinar ; Milde, Christopher ; Bekrater-Bodmann,Robin ; Rance, Mariela ; Foell, Jens ; Andoh, Jamila ; Becker, Susanne ; Flor, Herta
Abstract: BACKGROUND The mechanisms underlying chronic phantom limb pain (PLP) are complexand insufficiently understood. Altered sensory thresholds are often associated with chronic pain butquantitative sensory testing (QST) in PLP has so far been inconclusive due to large methodologicalvariation between studies and small sample sizes. METHODS In this study, we applied QST in 37unilateral upper-limb amputees (23 with and 14 without PLP) and 19 healthy controls. We assessed heatpain (HPT), pressure pain, warmth detection and two-point discrimination thresholds at the residuallimb, a homologous point and the thenar of the intact limb as well as both corners of the mouth.RESULTS We did not find significant differences in any of the thresholds between the groups. However,PLP intensity was negatively associated with HPT at all measured body sites except for the residuallimb, indicating lower pain thresholds with higher PLP levels. Correlations between HPT and PLPwere strongest in the contralateral face (r = -0.65, p < 0.001). Facial HPT were specifically associatedwith PLP, independent of residual limb pain (RLP) and various other covariates. HPT at the residuallimb, however, were significantly associated with RLP, but not with PLP. CONCLUSION We concludethat the association between PLP and, especially facial, HPT could be related to central mechanisms.SIGNIFICANCE Phantom limb pain (PLP) is still poorly understood. We show that PLP intensity isassociated with lower heat pain thresholds, especially in the face. This finding could be related to centralnervous changes in PLP.
DOI: https://doi.org/10.1002/ejp.1842
Posted at the Zurich Open Repository and Archive, University of ZurichZORA URL: https://doi.org/10.5167/uzh-212435Journal ArticlePublished Version
The following work is licensed under a Creative Commons: Attribution-NonCommercial 4.0 International(CC BY-NC 4.0) License.
Originally published at:Fuchs, Xaver; Diers, Martin; Trojan, Jörg; Kirsch, Pinar; Milde, Christopher; Bekrater-Bodmann, Robin;Rance, Mariela; Foell, Jens; Andoh, Jamila; Becker, Susanne; Flor, Herta (2022). Phantom limb painafter unilateral arm amputation is associated with decreased heat pain thresholds in the face. EuropeanJournal of Pain, 26(1):114-132.

DOI: https://doi.org/10.1002/ejp.1842
2

114 | Eur J Pain. 2022;26:114–132.wileyonlinelibrary.com/journal/ejp
DOI: 10.1002/ejp.1842
O R I G I N A L A R T I C L E
Phantom limb pain after unilateral arm amputation is associated
with decreased heat pain thresholds in the face
Xaver Fuchs1,2 | Martin Diers1,3 | Jörg Trojan1 | Pinar Kirsch1 | Christopher Milde1,4 | Robin Bekrater- Bodmann1 | Mariela Rance1,5 | Jens Foell1,6 | Jamila Andoh1,7 | Susanne Becker1,8 | Herta Flor1
1Institute of Cognitive and Clinical Neuroscience, Central Institute of Mental Health, Medical Faculty Mannheim, Heidelberg University, Mannheim,
Germany
2Biopsychology and Cognitive Neuroscience, Faculty of Psychology and Sports Science, Bielefeld University, Bielefeld, Germany
3Department of Psychosomatic Medicine and Psychotherapy, LWL University Hospital, Ruhr University Bochum, Bochum, Germany
4Department of Psychology, University of Koblenz- Landau, Landau, Germany
5Department of Radiology and Biomedical Imaging, Yale School of Medicine, New Haven, Connecticut, USA
6Department of Psychology, Florida State University, Tallahassee, Florida, USA
7Department of Psychiatry and Psychotherapy, Medical Faculty Mannheim, Central Institute of Mental Health, University of Heidelberg, Heidelberg,
Germany
8Integrative Spinal Research, Research Chiropractic, Balgrist University Hospital, University of Zurich, Zurich, Switzerland
This is an open access article under the terms of the Creat ive Commo ns Attri bution-NonCo mmercial License, which permits use, distribution and reproduction in any medium,
provided the original work is properly cited and is not used for commercial purposes.
© 2021 The Authors. European Journal of Pain published by John Wiley & Sons Ltd on behalf of European Pain Federation - EFIC ®
Correspondence
Xaver Fuchs, Institute of Cognitive and
Clinical Neuroscience, Central Institute of
Mental Health, Medical Faculty Mannheim,
J5, 68159 Mannheim, Germany.
Email: [email protected]
Funding information
Deutsche Forschungsgemeinschaft, Grant/
Award Number: SFB1158/B07; European
Research Council, Grant/Award Number:
FP7/2007– 2013/230249
Abstract
Background: The mechanisms underlying chronic phantom limb pain (PLP) are
complex and insufficiently understood. Altered sensory thresholds are often associ-
ated with chronic pain but quantitative sensory testing (QST) in PLP has so far been
inconclusive due to large methodological variation between studies and small sample
sizes.
Methods: In this study, we applied QST in 37 unilateral upper- limb amputees (23
with and 14 without PLP) and 19 healthy controls. We assessed heat pain (HPT),
pressure pain, warmth detection and two- point discrimination thresholds at the re-
sidual limb, a homologous point and the thenar of the intact limb as well as both
corners of the mouth.
Results: We did not find significant differences in any of the thresholds between the
groups. However, PLP intensity was negatively associated with HPT at all measured
body sites except for the residual limb, indicating lower pain thresholds with higher
PLP levels. Correlations between HPT and PLP were strongest in the contralateral
face (r = −0.65, p < 0.001). Facial HPT were specifically associated with PLP, inde-
pendent of residual limb pain (RLP) and various other covariates. HPT at the residual
limb, however, were significantly associated with RLP, but not with PLP.

| 115FUCHS ET AL.
1 | INTRODUCTION
Phantom limb pain (PLP)— pain perceived in an absent limb or
portions of an absent limb— is a common consequence of limb
amputation and occurs in about 50%– 80% (Nikolajsen, 2013)
of all amputees. Both peripheral and central mechanisms are
involved in chronic PLP (Flor & Fuchs, 2017; Flor et al., 2006;
Ramachandran & Hirstein, 1998). However, their exact roles
and impact are under debate. Peripheral factors relate to no-
ciceptive input stemming from the residual limb and can in-
volve peripheral sensitization, spontaneous discharges from
the residual limb due to involuntary movement or physiolog-
ical arousal (Sherman et al., 1989), neuromas (Devor, 1991;
Fried et al., 1991; Wiffen et al., 2006), or activity generated
in dorsal root ganglia (Vaso et al., 2014). Central sensitiza-
tion can develop as a consequence of long- term peripheral
nociceptive input and is mediated by spinal cord neurons
(Latremoliere & Woolf, 2009; Woolf & Salter, 2000) or the
brain (Woolf, 2014). In the brain, PLP has been associated
with maladaptive reorganization of body maps in the primary
somatosensory (S1) (Flor et al., 1995) and primary motor
cortex (Lotze et al., 2001) as well as preserved function of
the section of S1 that formerly represented the missing limb
(Kikkert et al., 2018; Makin, Scholz, et al., 2013).
Increased sensitivity to painful stimuli encompasses both
peripheral and central mechanisms (Woolf & Salter, 2000)
and is commonly assessed psychophysically by quanti-
tative sensory testing (QST; Magerl et al., 2010; Maier
et al., 2010; Rolke et al., 2006). Although QST has previ-
ously been applied in amputees, the results are mixed and
do not support an association with PLP. Some previous stud-
ies reported enhanced tactile (Haber, 1955; D. Katz, 1921;
Teuber et al., 1949; Wilson et al., 1962), thermal (Engkvist
et al., 1985; Wahren, 1990) or pressure pain sensitivity
(Cronholm, 1951; Korin et al., 1963; Vase et al., 2011) of
the residual limb compared to a homologous site on the in-
tact limb but did not relate these findings to PLP. The studies
that examined the relationship of tactile and pain sensitivity
to PLP mostly found no significant relationship with tac-
tile sensitivity (Grüsser et al., 2001; Hunter et al., 2005;
Katz, 1992; Li et al., 2015), heat pain (Flor et al., 1998;
Harden et al., 2010; Hunter et al., 2005; Li et al., 2015) or
cold perception (Harden et al., 2010). One study reported hy-
peralgesia for electrical painful stimulation on the intact limb
in amputees with but not without PLP, suggesting central
sensitization (Li et al., 2015).
Hence, findings of previous studies do not suggest that there
is an association of PLP and thresholds for non- painful stimuli
and are inconsistent with respect to the question whether PLP is
associated with pain thresholds. However, inconsistencies and
negative findings might be partly related to small sample sizes
and heterogeneity of the samples (e.g. lower- vs. upper- limb
amputations) and large heterogeneity in the applied assessment
methods and tested body sites. In particular, comparisons relat-
ing the residual to the intact limb might be problematic for vari-
ous reasons. First, it is difficult to standardize measures taken at
the residual limb due to variations in levels of amputation and
scar tissue. Second, QST of the residual limb may be related to
residual limb pain (RLP) rather than PLP. Third, testing only
the residual limb and a homologous contralateral site cannot
determine whether altered pain thresholds are also present at re-
mote sites, for instance at the thenar or the mouth. The face and
the missing upper limb have adjacent representations in sensory
and motor areas of the brain and the mouth might therefore have
altered sensitivity related to cortical reorganization.
As central sensitization might also play a role in PLP, the
present study tested whether PLP is accompanied by changes
in sensitivity to painful and non- painful stimuli applied at sev-
eral standardized body sites. The aims were (a) to investigate
whether there are changes in sensory thresholds, which are as-
sociated with the presence and/or the intensity of PLP, (b) to
test whether potential changes are restricted to the residual limb,
which could involve peripheral mechanisms, and/or whether
changes are also present in other areas of the body, which could
point to central mechanisms and finally, in order to test the
specificity to PLP, we sought to (c) take into account associa-
tions with other variables, which are potentially related to sen-
sitization and brain changes, especially RLP (Kern et al., 2009).
2 | METHODS
2.1 | Participants
Thirty- seven unilateral upper- limb amputees and 19 healthy
controls (HCs) took part in this study. We employed QST and
the assessment of phantom phenomena and chronic pain. The
sample was recruited from a nation- wide database on which
Conclusion: We conclude that the association between PLP and, especially facial,
HPT could be related to central mechanisms.
Significance: Phantom limb pain (PLP) is still poorly understood. We show that PLP
intensity is associated with lower heat pain thresholds, especially in the face. This
finding could be related to central nervous changes in PLP.

116 | FUCHS ET AL.
we had previously reported (Bekrater- Bodmann, Schredl,
et al., 2015). The participants were selected from the data-
base and contacted directly. The criteria for selection were
unilateral arm amputation and eligibility for magnetic reso-
nance imaging testing (used in another study). We recruited
amputees with and without PLP and attempted to achieve
comparable distributions of age, sex and side of amputation
between the groups. The study was approved by the Ethics
Commission II of the Medical Faculty Mannheim, Heidelberg
University. The amputees were divided into two groups— one
with PLP (PLP; N = 23) and one without PLP (non- PLP;
N = 14). Grouping was based on the Phantom Pain Severity
subscale of the German version of the West Haven- Yale
Multidimensional Pain Inventory (MPI; Flor et al., ,1990,
1995; Kerns et al., 1985) adapted for separate assessments of
PLP and RLP. The MPI is a 22- item questionnaire that uses a
7- point numeric rating scale for each item ranging from 0 to 6.
The Pain Severity subscale is calculated as the average value
of three items assessing (a) the intensity of momentary pain,
(b) the average pain during the last week and (c) the inten-
sity of pain- related suffering. Amputees with a Pain Severity
score of 0 were assigned to the non- PLP group and all other
amputees to the PLP group. The HC were matched for age,
sex and stimulation sites with amputees from both groups.
Sample and demographic details are provided in Table 1.
There were no statistically significant differences between
the three groups (PLP, non- PLP and HC) in the distribution of
sex [ratio male/female: PLP, 20/3; non- PLP, 12/2; HC, 16/3;
χ2(2, N = 56) = 0.06, p = 0.97] and age (mean ± standard devi-
ation [SD, years]: PLP, 51.3 ± 12.3; non- PLP, 53.1 ± 10.6;
HC, 51.6 ± 11.7; F(2, 53) = 0.10, p = 0.90). The PLP group and
the non- PLP group did not significantly differ in their level of
amputation (mean ± SD [% arm length]: PLP, 32.0 ± 18.8;
non- PLP, 40.2 ± 23.9; t(22.7) = −1.10, p = 0.28) or side of
amputation [χ2(1, N = 37) = 0.53, p = 0.47]. None of the ampu-
tees was amputated at an age below 5 years and the groups
did not differ significantly with respect to age of amputation
(mean ± SD, range [years]: PLP, 27.1 ± 12.5, 13– 55; non- PLP,
19.8 ± 11.5, 5– 45; t(29.5) = 1.80, p = 0.08). All amputees were
in a later stage after amputation with at minimum 3 years pass-
ing between amputation and sensory testing and the groups
did not differ significantly with respect to that (mean ± SD,
range [years]: PLP, 24.1 ± 11.4, 9– 58; non- PLP, 33.3 ± 15.6,
3– 55; t(29.5) = 1.84, p = 0.08). All amputees were right- handed
before amputation and so were all of the HC, according to the
Edinburgh Handedness Inventory (Oldfield, 1971).
2.2 | Body sites
Sensory testing was performed at five body sites. Amputees
were tested at the thenar of their intact hand, both corners of the
mouth, the residual limb and at a homologous site on the intact
arm. The sites at the corners of the mouth were symmetric and
located laterally of the lips on the hairy skin of the cheeks.
The test site at the residual limb was located 5 cm from the
distal end of the limb (Grüsser et al., 2001; Hunter et al., 2005)
and outside of scar tissue. Both sites on the arms were marked
using cosmetic colour and a photo was taken for documenta-
tion. The sites on the arms used in the HC were anatomically
homologous to the ones used in the matched amputees.
Body sites for sensory testing were selected based on the
following rationale: the residual limb was tested as the body
site directly affected by the amputation with a homologous site
on the intact arm as a control site. The corners of the mouth
were tested because they are remote from the amputation site
and clearly outside the area of potential nerve damage. In addi-
tion, the face is a relevant area with regards to central changes,
as the hand and the face are represented at adjacent sites
within the cortical body map in S1. Previous studies showed
that an expansion (in terms of functional S1 reorganization) of
the mouth area in the hemisphere contralateral to the amputa-
tion into the neighbouring former hand area is positively cor-
related with PLP (Flor et al., 1995; Lotze et al., 2001; MacIver
et al., 2008). It is, however, unclear whether cortical reorgani-
zation is reflected in altered sensitivity to stimuli presented at
the corners of the mouth. Additionally, we tested the thenar of
the existing hand, as sensory changes at the thenar might indi-
cate altered interhemispheric connectivity in the brain as the
cortical representation of the existing hand is functionally (van
den Heuvel & Hulshoff Pol, 2009) and structurally (Bonzano
et al., 2008) coupled to the corresponding area in the contra-
lateral hemisphere. Another reason for including the thenar is
that it is a standard site for QST (Rolke et al., 2006).
2.3 | Sensory testing
Measures were chosen to detect increased or decreased sen-
sitivity to noxious or non- noxious thermal or mechanical
stimuli. In total, we assessed four different measures: two
pain thresholds— heat pain threshold (HPT) and pressure pain
threshold (PPT)— and two detection thresholds— warmth de-
tection threshold (WDT) and tactile two- point discrimination
thresholds (2PDT). This allowed us to separate the effects
related to the nociceptive system from other somatosensory
modalities. The protocol used here adhered to the standards of
the German Research Network on Neuropathic Pain (DFNS)
but did not use the complete protocol. Thermal QST was se-
lected for comparability with several previous QST studies
with amputees (Flor et al., 1998; Harden et al., 2010; Hunter
et al., 2005; Li et al., 2015) and because it allows assessment of
both detection and pain thresholds using the same stimulus type
(temperature). Tactile 2PDT were selected because body sites
with larger S1 representation usually have smaller 2PDT and
the measure is sensitive to detect changes in S1 representation

| 117FUCHS ET AL.
TA
BL
E 1
D
escr
ipti
on o
f th
e st
udy s
ample
IDT
yp
eG
rou
pM
atc
hin
gS
exA
ge
EH
I
score
a
Lvl.
of
stim
. (%
)b
Age
at
am
p.
Tim
e
sin
ce a
mp
.
Sid
e of
am
p.c
Cau
se o
f
am
p.d
Ph
an
tom
lim
bT
eles
cop
eP
rost
hes
is
PL
P
inte
nsi
tye
RL
P
inte
nsi
tye
A01
Am
pute
eP
LP
None
M57
100.0
15
27
30
Rig
ht
Tra
um
aY
esY
esN
one
0.7
1.0
A02
Am
pute
eP
LP
HC
12
M48
75.0
70
18
30
Lef
tT
raum
aY
esY
esC
osm
etic
3.0
3.0
A03
Am
pute
eP
LP
HC
01
M69
57.1
45
45
24
Lef
tT
raum
aY
esY
esM
yo- e
l.3.3
2.7
A04
Am
pute
eP
LP
HC
06
M42
84.6
35
23
19
Rig
ht
Tra
um
aY
esY
esM
yo- e
l.0.7
1.7
A05
Am
pute
eP
LP
HC
02
M50
72.7
20
21
29
Rig
ht
Tra
um
aY
esY
esC
osm
etic
4.0
0.3
A06
Am
pute
eP
LP
HC
09
M52
84.6
042
10
Rig
ht
Tra
um
aY
esN
oC
osm
etic
2.3
0.0
A0
7A
mpute
eP
LP
HC
08
M64
66.7
055
9L
eft
Tum
or
Yes
Yes
None
2.0
0.0
A08
Am
pute
eP
LP
None
M62
100.0
15
43
19
Rig
ht
Tra
um
aY
esY
esN
one
2.3
2.0
A09
Am
pute
eP
LP
None
M39
80.0
35
15
24
Lef
tT
raum
aY
esN
oN
one
0.3
0.0
A1
0A
mpute
eP
LP
HC
15
M37
50.0
50
13
24
Lef
tT
raum
aY
esY
esC
osm
etic
0.3
0.0
A11
Am
pute
eP
LP
None
M30
75.0
25
19
11
Rig
ht
Tra
um
aY
esY
esN
one
2.0
2.0
A12
Am
pute
eP
LP
None
M67
83.3
45
38
29
Lef
tT
raum
aY
esN
oC
osm
etic
1.7
4.7
A13
Am
pute
eP
LP
HC
14
M54
84.6
21
42
12
Rig
ht
Tra
um
aY
esY
esN
one
1.7
1.3
A14
Am
pute
eP
LP
None
M73
84.6
15
15
58
Rig
ht
Tra
um
aY
esN
oN
one
0.7
0.0
A1
5A
mpute
eP
LP
HC
13
M26
57.1
38
17
9R
ight
Tra
um
aY
esY
esN
one
2.3
2.0
A16
Am
pute
eP
LP
HC
04
M42
100.0
43
19
23
Lef
tT
raum
aY
esN
oN
one
1.0
1.7
A17
Am
pute
eP
LP
None
M54
46.7
15
14
40
Rig
ht
Tra
um
aY
esY
esN
one
0.7
1.0
A1
8A
mpute
eP
LP
None
M63
75.0
30
40
23
Rig
ht
Tra
um
aY
esY
esM
yo- e
l.1.7
1.0
A19
Am
pute
eP
LP
None
M52
69.2
50
16
36
Rig
ht
Tra
um
aY
esN
oM
anual
1.3
2.0
A20
Am
pute
eP
LP
None
M48
76.5
30
18
30
Rig
ht
Tra
um
aY
esY
esN
one
4.3
3.0
A2
1A
mpute
eP
LP
HC
19
F40
100.0
45
25
15
Lef
tO
ther
Yes
Yes
None
2.7
0.0
A22
Am
pute
eP
LP
HC
17
F58
100.0
25
37
21
Lef
tT
raum
aY
esY
esN
one
2.3
0.7
A23
Am
pute
eP
LP
None
F53
66.7
70
23
30
Rig
ht
Tra
um
aY
esN
oN
one
3.7
2.7
A24
Am
pute
enon- P
LP
None
M59
84.6
20
10
49
Rig
ht
Tra
um
aN
oN
oM
yo- e
l.0.0
0.0
A25
Am
pute
enon- P
LP
HC
05
M67
90.0
62
19
48
Rig
ht
Tra
um
aY
esY
esM
yo- e
l.0.0
0.0
A26
Am
pute
enon- P
LP
HC
07
M54
100.0
72
23
31
Rig
ht
Tra
um
aY
esN
oN
one
0.0
0.0
A27
Am
pute
enon- P
LP
None
M24
83.3
45
618
Lef
tT
raum
aN
oN
oM
yo- e
l.0.0
A28
Am
pute
enon- P
LP
None
M60
57.1
60
555
Lef
tT
raum
aN
ono
None
0.0
0.0
A29
Am
pute
enon- P
LP
HC
11
M62
71.4
10
10
52
Lef
tT
raum
aN
oN
oN
one
0.0
0.0
A30
Am
pute
enon- P
LP
HC
10
M47
60.0
022
25
Lef
tT
raum
aY
esN
oN
one
0.0
0.0
A31
Am
pute
enon- P
LP
None
M48
70.0
62
45
3R
ight
Tra
um
aN
oN
oN
one
0.0
0.0
(Conti
nues
)

118 | FUCHS ET AL.
IDT
yp
eG
rou
pM
atc
hin
gS
exA
ge
EH
I
score
a
Lvl.
of
stim
. (%
)b
Age
at
am
p.
Tim
e
sin
ce a
mp
.
Sid
e of
am
p.c
Cau
se o
f
am
p.d
Ph
an
tom
lim
bT
eles
cop
eP
rost
hes
is
PL
P
inte
nsi
tye
RL
P
inte
nsi
tye
A32
Am
pute
enon- P
LP
HC
03
M65
69.2
52
15
50
Rig
ht
Tra
um
aY
esY
esC
osm
etic
0.0
0.0
A33
Am
pute
enon- P
LP
None
M51
83.3
10
23
28
Lef
tT
raum
aN
oN
oC
osm
etic
0.0
0.0
A34
Am
pute
enon- P
LP
HC
16
M56
100.0
65
38
18
Rig
ht
Tra
um
aN
oN
oC
osm
etic
0.0
0.0
A35
Am
pute
enon- P
LP
None
M47
100.0
25
15
32
Lef
tT
um
or
Yes
Yes
Myo- e
l.0.0
0.0
A36
Am
pute
enon- P
LP
HC
18
F52
100.0
50
18
34
Lef
tT
raum
aN
oN
oM
yo- e
l.0.0
3.0
A37
Am
pute
enon- P
LP
None
F51
86.7
30
28
23
Lef
tT
raum
aY
esY
esN
one
0.0
0.7
HC
01
Contr
ol
HC
A03
M71
100.0
45
Lef
t
HC
02
Contr
ol
HC
A05
M50
61.3
20
Rig
ht
HC
03
Contr
ol
HC
A32
M64
100.0
52
Rig
ht
HC
04
Contr
ol
HC
A16
M41
70.4
43
Lef
t
HC
05
Contr
ol
HC
A25
M67
88.6
62
Rig
ht
HC
06
Contr
ol
HC
A04
M44
75.0
35
Rig
ht
HC
07
Contr
ol
HC
A26
M54
73.9
72
Rig
ht
HC
08
Contr
ol
HC
A07
M64
93.3
0L
eft
HC
09
Contr
ol
HC
A06
M53
88.9
0R
ight
HC
10
Contr
ol
HC
A30
M47
51.7
0L
eft
HC
11
Contr
ol
HC
A29
M63
89.5
10
Lef
t
HC
12
Contr
ol
HC
A02
M47
63.6
70
Lef
t
HC
13
Contr
ol
HC
A15
M25
53.8
38
Rig
ht
HC
14
Contr
ol
HC
A13
M51
65.2
21
Rig
ht
HC
15
Contr
ol
HC
A10
M35
89.5
50
Lef
t
HC
16
Contr
ol
HC
A34
M55
71.4
65
Rig
ht
HC
17
Contr
ol
HC
A22
F58
100.0
25
Lef
t
HC
18
Contr
ol
HC
A36
F53
31.2
50
Lef
t
HC
19
Contr
ol
HC
A21
F39
82.9
45
Lef
t
Abbre
via
tions:
EH
I, E
din
burg
h h
anded
nes
s in
ven
tory
; H
C, hea
lthy c
ontr
ol;
myo- e
l., m
yoel
ectr
ic p
rost
hes
is;
non- P
LP
, w
ithout
phan
tom
lim
b p
ain;
PL
P, phan
tom
lim
b p
ain;
RL
P, re
sidual
lim
b p
ain.
a A v
alue
of
100 i
n t
he
EH
I in
dic
ates
the
stro
nges
t deg
ree
of
right-
han
ded
nes
s, a
val
ue
of
−100 i
ndic
ates
the
stro
nges
t deg
ree
of
left
- han
ded
nes
s.bL
evel
of
stim
ula
tion w
as d
efin
ed a
s th
e posi
tion o
f m
easu
rem
ents
at
the
arm
rel
ativ
e to
the
length
of
the
(inta
ct)
arm
, m
easu
red f
rom
the
caput
hum
eri
to t
he
tip o
f th
e th
ird d
igit
. In
the
ampute
es, th
is p
osi
tion w
as d
efin
ed b
y t
he
length
of
the
resi
dual
lim
b. H
om
olo
gous
site
s w
ere
mea
sure
d i
n t
he
mat
ched
HC
.c S
ide
of
amputa
tion i
n t
he
HC
(in
bra
cket
s) r
efer
s to
the
side
of
amputa
tion o
f th
e m
atch
ed a
mpute
e.dT
he
cate
gory
‘oth
er’
cause
s of
amputa
tion s
ubsu
mes
unknow
n c
ause
s not
fall
ing i
nto
the
cate
gori
es t
raum
atic
, in
jury
, dysp
lasi
a, i
nfe
ctio
n, tu
mour
or
vas
cula
r dis
ease
.e P
ain s
ever
ity s
ubsc
ale
of
the
MP
I ad
apte
d f
or
PL
P a
nd R
LP
, re
spec
tivel
y. V
alues
ran
ge
from
0 (
low
est
sever
ity)
to 6
(hig
hes
t se
ver
ity).
TA
BL
E 1
(C
onti
nued
)

| 119FUCHS ET AL.
(Dinse, 2003; Muret et al., 2016). PPTs were selected to assess
an additional pain threshold referring to mechanical stimuli.
The procedures for HPT, WDT and PPT were based on the
study protocol developed by the DFNS (Rolke et al., 2006)
and were carried out by experimenters trained in QST.
2.3.1 | Thermal thresholds
Warmth detection thresholds and HPT were measured using
a Medoc ATS thermode (3 x 3 cm surface) included in
the Medoc Pathway system (PATHWAY Pain & Sensory
Evaluation System, Medoc Ltd. Advanced Medical System).
Stimuli were applied with a baseline temperature of 32 ℃
and a rising rate of 1.5 ℃/s (ascending method of limits)
(Bekrater- Bodmann, Chung, et al., 2015). The maximum
temperature of the system was set to 52 ℃ to avoid skin
injuries.
Because amputees, when tested at the thenar, could not
respond with button presses, all responses for all body sites
and groups, including the ones in the HC, were given verbally
to ensure comparability between sites and groups. The as-
sessment of WDT and HPT only differed with respect to the
instructions (‘say “stop” as soon as you feel that the tempera-
ture is getting warmer’ for WDT and ‘say “stop” when the
temperature just becomes painful’ for HPT). When partici-
pants said ‘stop’, the experimenter immediately terminated
the ascending stimulus and the thermode returned to the
baseline temperature at a rate of 8 ℃/s. The participants were
asked to keep their eyes closed throughout the test.
When assessing WDT and HPT in the face, the thermode
was attached to the skin with adhesive tape and the participant
was additionally asked to gently press the index finger against
the thermode to ensure good contact with the skin. In each
series, we conducted five consecutive trials. The first trial of
each series was presented to familiarize participants with the
test and the remaining four stimuli were averaged to calculate
the threshold (Bekrater- Bodmann, Chung, et al., 2015).
2.3.2 | Pressure pain thresholds
Pressure pain threshold were measured using a Medoc
Algomed (Medoc Ltd. Advanced Medical System) comput-
erized pressure algometer (surface of 1 cm2). Four consecu-
tive stimuli were applied using a rate of 50 kPa/s (ascending
method of limits) (Rolke et al., 2006). The participants were
instructed to say ‘stop’ when the pressure started to feel pain-
ful. Four consecutive stimuli were applied; the first stimulus
was excluded and the remaining three were averaged for cal-
culation of the pain threshold. When testing at the corners of
the mouth, the participants clenched the teeth to ensure that
the tested tissue was pushed against the teeth when applying
the pressure algometer. The experimenter gently stabilized
the participant's head using the hand not applying the algom-
eter. This prevented the participant from compensating the
pressure using the neck muscles, avoiding discomfort.
2.3.3 | Two- point discrimination thresholds
Two- point discrimination threshold were measured using a
set of 28 calibrated compasses (Rotring “Centro”; Rotring)
ranging in size from 1 to 82 mm in steps of 3 mm. The tips
of the compasses were exchanged with blunt rods in order to
induce a light touch. Stimuli were applied by placing the com-
pass perpendicularly onto the skin from above. The skin was
touched briefly (1– 2 s) while the participants kept their eyes
closed. We used a simple adaptive staircase (‘up- down’) pro-
cedure (Dixon & Mood, 1948; Levitt, 1971), stopping after
seven reversals. The first reversal point was excluded and the
following six were averaged to calculate the absolute (50%)
threshold. To avoid biases in the estimation of the thresholds
that can be introduced when the starting point is far away
from the threshold, we used different compass sizes as start-
ing points, depending on the body site: 25 mm for the corners
of the mouth where the threshold was expected to be lower
than on the arms (Mancini et al., 2014; Weinstein, 1968) and
46 mm for the arms. 2PDT at the arms were always meas-
ured in proximal- to- distal direction while the participant sat
in a chair. 2PDT at the corners of the mouth were measured
parallel to the inferior- to- superior axis while the participant
was lying on a bench to facilitate applying the stimuli per-
pendicularly from above. Beforehand, the protocol had been
evaluated in a pilot study, testing the dorsal forearms of 10
participants. The procedure showed good within- session reli-
ability (Pearson's r = 0.81). The data of this pilot are provided
in Supplementary Material (Figure S5). 2PDT were measured
in a subsample of 27 amputees (19 in the PLP and 8 in the
non- PLP group) and all HC and at all body sites except for
the thenar.
2.4 | Specificity for PLP
To test whether associations between PLP and sensory
thresholds were specific to PLP and not mediated by other
correlates of PLP or related brain plasticity, we performed
additional analyses to control their potential influence.
Especially, RLP is correlated with PLP (Kern et al., 2009)
and can therefore confound interpretations if not statistically
controlled. Apart from RLP, we also ruled out confound-
ers and the results of these analyses are reported in detail
in Supplementary Material: (a) use of prostheses (Dietrich
et al., 2012; Lotze et al., 1999), (b) telescoping (Flor
et al., 2006; Grüsser et al., 2001), (c) compensation in daily

120 | FUCHS ET AL.
activities by the existing hand (Makin, Cramer, et al., 2013),
(d) level of the amputation and (e) time since amputation.
2.5 | Statistical analyses
2.5.1 | Data inspection and outliers
Because large variation was observed in the sensory data (see
the boxplots in Figure 2 in the Section 3), we performed a sys-
tematic analysis for the presence and the influence of outliers
which is included in Supplementary Material. Despite the pres-
ence of some outliers, the results were not strongly influenced,
and the conclusions remain identical for statistical analyses
with and without outliers. Therefore, no data were removed for
the analyses presented in the article. Supplementary Material
includes a reanalysis of the data after removal of outliers.
2.5.2 | Transformation of sensory data
In accordance with the recommendations given by Rolke
et al. (2006), we transformed WDT and PPT logarithmically
prior to the statistical analyses using a log- transformation.
WDT were coded as deviation from the baseline, with the
baseline temperature of 32℃ subtracted from the thresholds,
prior to the log- transformation.
2.5.3 | Statistical models for comparisons between groups and body sites
To compare thresholds between the groups and body sites, we
fitted a (repeated measures) linear mixed model (LMM) for
each sensory measure using group (PLP, non- PLP and HC) as
a between- subject fixed factor and body site (arm ipsilateral,
arm contralateral, corner of the mouth ipsilateral, corner of the
mouth contralateral and thenar contralateral to the amputation)
as a within- subject fixed factor. All models included partici-
pants as a random intercept effect. LMMs are a flexible and ro-
bust alternative to repeated measures analyses of variance and
permit simultaneous analysis of discrete grouping variables,
repeated factors and continuous covariates, providing a frame-
work for both factorial and regression- type analyses (Pinheiro
& Bates, 2000). To test whether differences between groups
depended on the body site (or vice versa), we also included the
interaction between group and body site in the models.
The coding of body site (‘ipsilateral’ and ‘contralat-
eral’) in the HC was defined by the side of amputation of
the matched (yoked) amputee. For example, if an HC was
matched to a right- sided amputee, his or her right corner of
the mouth was coded as ipsilateral. Note that in the amputees,
‘ipsilateral arm’ refers to the residual limb. This allowed a
balanced mixture of right- and left- side body parts of the HCs
within the categories.
Significant effects were followed up by pairwise post-
hoc comparisons to test which mean values differed signifi-
cantly. To avoid alpha error inflation, post- hoc test p- values
were corrected using the false discovery rate (Benjamini &
Hochberg, 1995).
2.5.4 | Associations between sensory thresholds and PLP
To test the association between the sensory thresholds and
PLP intensity, we correlated each sensory threshold in the
PLP group with the Phantom Pain Severity score from the
MPI using Pearson correlations.
Next, to test whether PLP intensity was associated with a
combination of sensory thresholds and to account for the inter-
correlation of sensory variables, we performed a stepwise mul-
tiple regression analysis. We predicted PLP intensity in the PLP
group by a combination of HPT, PPT and WDT at all body sites
and determined the best- fitting model using stepwise elimina-
tion of regression terms (based on Akaike's information criterion
[AIC], as implemented in R’s stepwise function). As missing
values are a problem for multiple regression approaches, we im-
puted missing data with the mean values prior to the analyses
(1 value [0.8%] for HPT and PPT, respectively). As 2PDT were
only measured in a subset of the participants, this would have
resulted in an imputation of 18% of the values. Therefore, we
excluded 2PDT from the multiple regression analyses.
2.5.5 | Specificity of associations for PLP versus RLP intensity
To test whether the types of pain (PLP and RLP intensity)
are predicted differentially by the sensory thresholds, we
performed an LMM that resembled the multiple regression
model described in the previous paragraph but included both
PLP and RLP intensity as predicted values, coded by a factor
‘pain type’ that was included as a within- subject fixed fac-
tor. The model comprised all sensory thresholds as predic-
tors and all two- fold interactions between the pain type and
the sensory thresholds. A significant interaction between the
pain type and a sensory threshold variable indicates that the
prediction is significantly different (and therefore specific)
for PLP versus RLP intensity. As for the regression model,
we used a stepwise approach to determine the most appropri-
ate model based on AIC. The analysis included all amputees
showing either PLP or RLP values above zero. This was the
case for the 23 amputees from the PLP group and three cases
from the non- PLP group who reported RLP but no PLP; the
analysis therefore included 26 participants.

| 121FUCHS ET AL.
For another regression model aiming at ruling out
other confounding variables (described in Section 2.5), see
Supplementary Material.
2.5.6 | Software and data resources
All data and code to reproduce all statistical analyses and
figures reported in this article are publicly available online
in a repository hosted on the website of the Open Science
Framework and can be accessed via https://osf.io/v6hjy/
We used the R statistical software (R Core Team, 2014).
LMMs were performed using the “lme4” (Bates
et al., 2014) and the “lmerTest” (Kuznetsova et al., 2014)
packages for R. Degrees of freedom in the LMMs were
estimated using Satterthwaite's approximation as imple-
mented in the lmerTest (Kuznetsova et al., 2014) package.
Figures were created using the “ggplot2” package for R
(Wickham, 2009).
3 | RESULTS
3.1 | Effects related to the presence of PLP: differences between the groups and body sites
Results from the LMMs for all measures are shown in
Figure 1. Table 2 summarizes the coefficients for the full
models (including the factor group and body site, and the
group × body site interaction). Table 3 summarizes the re-
sults from post- hoc comparisons between the body sites.
3.1.1 | Heat pain thresholds
There was no significant main effect of group [F(2, 53) = 0.1,
p = 0.901] and no significant interaction between group and
body site [F(8, 211) = 0.65, p = 0.732]. The main effect of body
site was significant [F(4, 211) = 50.5, p < 0.001]. Post- hoc
comparisons (Table 3) showed that HPT were significantly
T A B L E 2 Results for main effects and interactions from linear
mixed models for heat pain thresholds, pressure pain thresholds,
warmth detection thresholds and two- point discrimination thresholds
including the factors group, body site and their interaction
Measure Term F
df (num./
denom.) p
HPT Group 0.10 2/52.9 0.906
Body site 51.77 4/211 <0 .001
Group × Body site 0.69 8/211 0.701
PPT Group 0.86 2/53 0.43
Body site 130.60 4/211 <0 .001
Group × Body site 1.91 8/211 0.061
WDT Group 2.88 2/53 0.065
Body site 88.91 4/212 <0 .001
Group × Body site 0.94 8/212 0.485
2PDT Group 1.43 2/44.2 0.25
Body site 304.21 3/128.5 <0 .001
Group × Body site 0.54 6/128 0.775
Abbreviations: 2PDT, two- point discrimination thresholds; HPT, heat pain
thresholds; PPT, pressure pain thresholds; WDT, warmth detection thresholds.
F I G U R E 1 Results of the linear mixed models comparing the thresholds for heat pain (HPT), pressure pain (PPT), warmth detection (WDT)
and two- point discrimination (2PDT) between the groups (amputees with phantom limb pain [PLP]: orange; amputees without phantom limb
pain [non- PLP]: blue; healthy controls [HC]: grey) and the body sites. The values represent estimated marginal means derived from the statistical
models with 95% confidence intervals shown as error bars. There were no significant differences between the groups in any of the pairwise
comparisons. The body sites differed significantly (see also section 3.1, Tables 2 and 3 for details)

122 | FUCHS ET AL.
lower at the ipsilateral arm compared to the contralateral arm
and also at both corners of the mouth compared to both arms
and the thenar. The corners of the mouth did not differ sig-
nificantly from each other, and the thenar did not differ sig-
nificantly from both arms.
3.1.2 | Pressure pain thresholds
For PPT, there was no significant effect of group [F(2, 53) =
0.86, p = 0.430] or group × body site [F(8, 211) = 1.91,
p = 0.061]. The factor body site was significant [F(4,
T A B L E 3 Results from post- hoc comparisons for the significant factor body site
Measure Contrast Estimate SE t ratio df p
HPT Arm contra– arm ipsi 1.19 0.448 2.65 211 0.012
Arm contra– mouth contra 4.55 0.447 10.20 211 <0 .001
Arm contra– mouth ipsi 4.57 0.447 10.23 211 <0 .001
Arm contra– thenar contra 0.28 0.447 0.63 211 0.587
Arm ipsi– mouth contra 3.37 0.448 7.51 211 <0 .001
Arm ipsi– mouth ipsi 3.38 0.448 7.54 211 <0.001
Arm ipsi– thenar contra −0.91 0.448 −2.02 211 0.055
Mouth contra– mouth ipsi 0.02 0.447 0.03 211 0.973
Mouth contra– thenar contra −4.27 0.447 −9.57 211 <0.001
Mouth ipsi– thenar contra −4.29 0.447 −9.60 211 <0.001
PPT Arm contra– arm ipsi 0.07 0.0250 2.68 211 0.009
Arm contra– mouth contra 0.31 0.0250 12.46 211 <0.001
Arm contra– mouth ipsi 0.33 0.0250 13.30 211 <0.001
Arm contra– thenar contra −0.14 0.0251 −5.40 211 <0.001
Arm ipsi– mouth contra 0.24 0.0250 9.77 211 <0.001
Arm ipsi– mouth ipsi 0.27 0.0250 10.62 211 <0.001
Arm ipsi– thenar contra −0.20 0.0251 −8.07 211 <0.001
Mouth contra– mouth ipsi 0.02 0.0250 0.85 211 0.398
Mouth contra– thenar contra −0.45 0.0251 −17.81 211 <0.001
Mouth ipsi– thenar contra −0.47 0.0251 −18.65 211 <0.001
WDT Arm contra– arm ipsi 0.04 0.0288 1.36 212 0.196
Arm contra– mouth contra 0.40 0.0288 13.70 212 <0.001
Arm contra– mouth ipsi 0.41 0.0288 14.14 212 <0.001
Arm contra– thenar contra 0.17 0.0288 5.94 212 <0.001
Arm ipsi– mouth contra 0.36 0.0288 12.34 212 <0.001
Arm ipsi– mouth ipsi 0.37 0.0288 12.78 212 <0.001
Arm ipsi– thenar contra 0.13 0.0288 4.58 212 <0.001
Mouth contra– mouth ipsi 0.01 0.0288 0.44 212 0.66
Mouth contra– thenar contra −0.22 0.0288 −7.76 212 <0.001
Mouth ipsi– thenar contra −0.24 0.0288 −8.20 212 <0.001
2PDT Arm contra– arm ipsi −0.08 0.213 −0.38 131 0.705
Arm contra– mouth contra 4.35 0.202 21.47 126 <0.001
Arm contra– mouth ipsi 4.43 0.202 21.88 126 <0.001
Arm ipsi– mouth contra 4.43 0.213 20.79 131 <0.001
Arm ipsi– mouth ipsi 4.51 0.213 21.19 131 <0.001
Mouth contra– mouth ipsi 0.08 0.202 0.41 126 0.705
Note: Post- hoc tests followed linear mixed models for each of the sensory thresholds (HPT, PPT, WDT and 2PDT). Within each model, p values were adjusted for
multiple comparisons using false discovery rate.
Abbreviations: 2PDT, two- point discrimination thresholds; HPT, heat pain thresholds; PPT, pressure pain thresholds; SE, standard error; WDT, warmth detection
thresholds.
| 123FUCHS ET AL.
207) = 87.1, p < 0.001]. Post- hoc comparisons between body
sites revealed that all body sites differed significantly from
each other except for the homologous body parts at the corners
of the mouth, which did not differ significantly. At the arms,
PPT were significantly lower ipsilateral than contralateral.
3.1.3 | Warmth detection thresholds
No significant effect of group [F(2, 53) = 2.88, p = 0.065] and
group × body site was found [F(8, 212) = 0.94, p = 0.485]. The
factor body site was significant [F(4, 204) = 88.91, p < 0.001]. Post-
hoc comparisons revealed significant differences between all body
sites, except for the comparison between the corners of the mouth
and between both arms where the differences were not significant.
3.1.4 | Two- point discrimination thresholds
For 2PDT, there was no significant effect of group [F(2, 44) =
1.43, p = 0.25] or group × body site [F(6, 128) = 0.54,
p = 0.775]. Body site was significant [F(3, 129) = 304.21,
p < 0.001]. Post- hoc tests showed that thresholds were lower
at both corners of the mouth than at both arms. The homolo-
gous parts at the corners of the mouth and at the arms did not
differ significantly from each other.
3.2 | Effects related to the intensity of PLP: correlational analyses
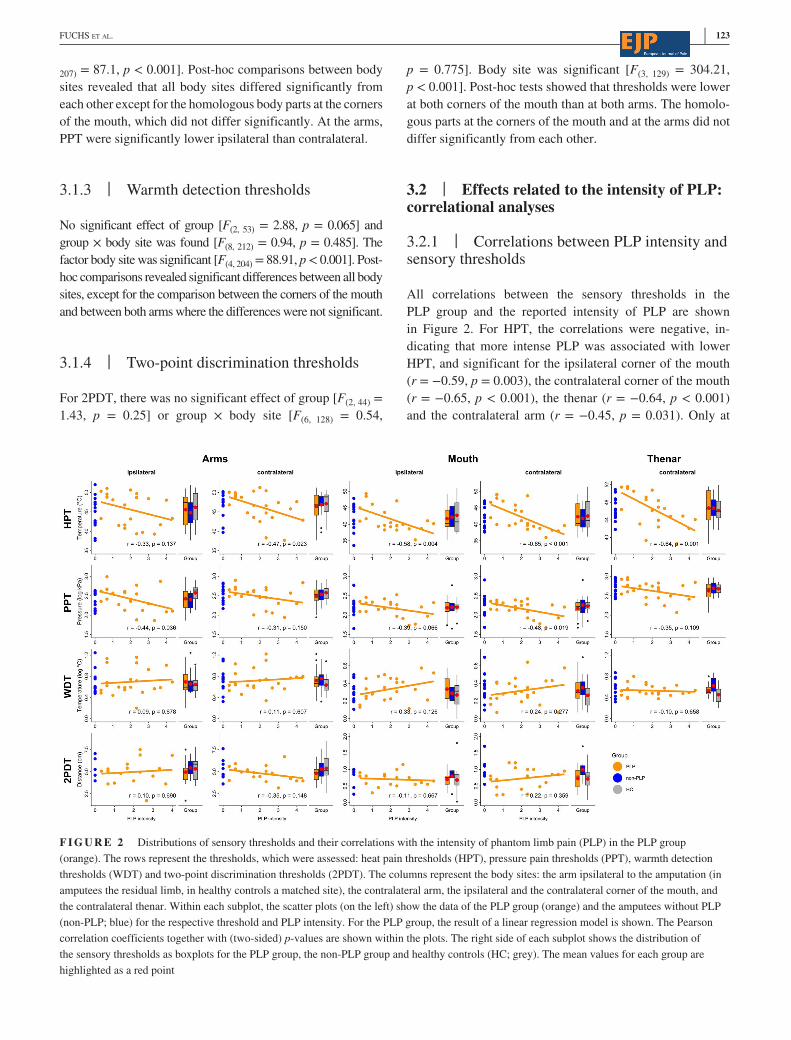
3.2.1 | Correlations between PLP intensity and sensory thresholds
All correlations between the sensory thresholds in the
PLP group and the reported intensity of PLP are shown
in Figure 2. For HPT, the correlations were negative, in-
dicating that more intense PLP was associated with lower
HPT, and significant for the ipsilateral corner of the mouth
(r = −0.59, p = 0.003), the contralateral corner of the mouth
(r = −0.65, p < 0.001), the thenar (r = −0.64, p < 0.001)
and the contralateral arm (r = −0.45, p = 0.031). Only at
F I G U R E 2 Distributions of sensory thresholds and their correlations with the intensity of phantom limb pain (PLP) in the PLP group
(orange). The rows represent the thresholds, which were assessed: heat pain thresholds (HPT), pressure pain thresholds (PPT), warmth detection
thresholds (WDT) and two- point discrimination thresholds (2PDT). The columns represent the body sites: the arm ipsilateral to the amputation (in
amputees the residual limb, in healthy controls a matched site), the contralateral arm, the ipsilateral and the contralateral corner of the mouth, and
the contralateral thenar. Within each subplot, the scatter plots (on the left) show the data of the PLP group (orange) and the amputees without PLP
(non- PLP; blue) for the respective threshold and PLP intensity. For the PLP group, the result of a linear regression model is shown. The Pearson
correlation coefficients together with (two- sided) p- values are shown within the plots. The right side of each subplot shows the distribution of
the sensory thresholds as boxplots for the PLP group, the non- PLP group and healthy controls (HC; grey). The mean values for each group are
highlighted as a red point

124 | FUCHS ET AL.
the ipsilateral arm (residual limb), the correlation was not
significant (r = −0.33, p = 0.137). PPT were significantly
negatively correlated with PLP intensity at the contralateral
corner of the mouth (r = −0.48, p = 0.019) and at the ipsi-
lateral arm (r = −0.44, p = 0.036). However, the result for
the ipsilateral arm was no longer significant when outliers
were removed from the data (see Supplementary Material,
Table S3). At all other body sites, the correlations were nega-
tive but not significant (Figure 2). WDT and 2PDT were not
significantly correlated with PLP intensity at any of the body
sites (Figure 2).
Intra- individual differences between the homologous
areas at the arms and the corners of the mouth were not sig-
nificantly correlated with PLP intensity for any of the sen-
sory thresholds (all r > −0.32 and <0.37, all p > 0.07). All
correlations are shown Supplementary Material (Figure S1).
3.2.2 | Association of PLP intensity with a linear combination of all sensory thresholds
The stepwise multiple regression procedure removed 12 out
of the 15 sensory thresholds and resulted in a model in which
PLP intensity was predicted by a combination of three sen-
sory thresholds, which are listed in Table 4 (Model A). The
resulting model was statistically significant [F(9, 13) = 8.67,
p < 0.001; R2 = 0.57; R2adj = 0.51]. Two thresholds in this
model significantly predicted higher degrees of PLP in-
tensity: lower HPT [b = −0.21, t(19) = −4.74, p < 0.001]
and higher WDT at the contralateral corner of the mouth
[b = 2.96, t(19) = 2.96, p = 0.03] Note that HPT received
negative and WDT positive weights, indicating that higher
PLP intensity was associated with lower HPT and higher
WDT, which is shown in Figure 3.
Further multiple regression analyses (reported in
Supplementary Material, Table S5) confirmed that the asso-
ciations of PLP intensity with HPT and WDT at the corner
of the mouth were still present when a number of other po-
tentially relevant correlates of PLP were controlled for. This
shows that the relationship is robust and not mediated by any
of these variables.
3.2.3 | Specificity of associations of PLP and RLP intensities with sensory thresholds
Only two sensory thresholds remained in the resulting model
(Model B): HPT at the contralateral corner of the mouth and
HPT at the ipsilateral arm. Importantly, there were signifi-
cant interactions with type of pain (PLP vs. RLP intensity)
for both HPT at the contralateral corner of the mouth [F(1,
23) = 8.69, p < 0.001] and at the ipsilateral arm [F(1, 23) = 4.41,
p = 0.046]. The interactions are visualized in Figure 4. On
the ipsilateral arm, lower HPT was associated with higher
levels of RLP but not with PLP [b = −0.16, t(23) = −2.10,
p = 0.047]. On the contralateral corner of the mouth, how-
ever, lower HPT was specifically associated with more PLP
and not with RLP [b = 0.23, t(23) = 2.95, p = 0.007]. This
indicates that the two concurrent types of clinical pain, PLP
and RLP were differentially associated with sensory thresh-
olds: PLP was mainly associated with HPT in the face and
RLP with HPT at the residual limb.
Model Term b t df p
A (Intercept) 8.99 4.84 19 <0.001
HPT mouth contra −0.21 −4.75 19 <0.001
WDT arm ipsi 1.07 1.49 19 0.152
WDT mouth contra 2.96 2.38 19 0.028
B (Intercept) 9.54 3.33 42.6 0.002
Pain typea −2.48 −0.72 23.0 0.478
HPT arm ipsi 0.01 0.15 42.6 0.882
HPT mouth contra −0.19 −3.01 42.6 0.004
Pain type × HPT arm ipsi −0.16 −2.10 23.0 0.047
Pain type × HPT mouth contra 0.23 2.95 23.0 0.007
Note: The table lists regression terms that remained in the final model after iterative removal of terms not
significantly contributing to the model fit. (A) Statistical prediction of PLP by HPT, PPT and WDT measured
at all body sites. (B) Statistical prediction of both PLP and RLP by the same combination of thresholds like in
model A including the factor ‘pain type’ coding for PLP and RLP, respectively.
Abbreviations: HPT, heat pain thresholds; PLP, phantom limb pain intensity; PPT, pressure pain thresholds;
RLP, residual limb pain intensity; WDT, warmth detection thresholds.aThe baseline level was PLP, hence the regression coefficient codes the influence of RLP.
T A B L E 4 Results from stepwise
multiple regression and linear mixed models

| 125FUCHS ET AL.
4 | DISCUSSION
Based on research on other types of chronic pain and the find-
ings in cortical reorganization in PLP, we assumed that PLP
might be associated with lowered pain thresholds also at body
sites that are remote from the residual limb. Previous QST
studies in amputees have provided an inconclusive picture due
to inconsistencies in measures, small and heterogenous sam-
ples and a focus restricted to the residual limb. In this study,
we tested a relatively large and homogeneous sample of upper-
limb amputees. We investigated whether changes in pain and
detection thresholds assessed at different body sites correlate
with the presence and/or the perceived intensity of PLP.
Our most important finding is that the presence of PLP
(whether amputees reported PLP or not) was not related to
any sensory measure, whereas the perceived intensity of PLP
in amputees who reported PLP was clearly associated with
lower pain thresholds, especially in the face.
4.1 | No association between sensory thresholds and the presence of PLP
We tested for associations with the presence of PLP by com-
paring pain and perception thresholds measured in the PLP
group to those measured in the non- PLP group and in HC.
Surprisingly, there were no significant differences between
HPT and PPT at any of the measured body sites as a function
of the absence/presence of PLP. We also did not find any
evidence for an association between detection thresholds and
the presence of PLP. This negative finding was against our
primary hypothesis that the presence of PLP might be associ-
ated with pain perception.
4.2 | PLP intensity correlates with sensory thresholds
Contrary to the non- significant differences in sensory thresh-
olds between the groups, we found that higher PLP intensity
was associated with lower HPT at all body sites except for the
residual limb. Similar correlations were also found for PPT
although the relationships were numerically weaker and a ro-
bust significant correlation was only found at the contralateral
corner of the mouth. In contrast to the pain thresholds, detec-
tion thresholds for non- painful stimuli were not significantly
correlated with PLP intensity. However, when combining
pain and detection thresholds of all body sites to statistically
predict PLP intensity in multiple regression, we found a dif-
ferential relationship of PLP intensity with lowered HPT but
heightened WDT in the face. At the contralateral corner of
the mouth, the combination of HPT and WDT explained a
high proportion (of up to 51%) of the variance in PLP inten-
sity. This suggests that when PLP is present, its intensity is
strongly associated with decreased pain but increased warmth
detection thresholds in the face. Because the face is remote
from the residual limb, the observed patterns of sensory
thresholds are therefore not likely to have a peripheral origin.
It might strike as a surprise that PLP was differentially
associated with, on the one side, lower pain but, on the other
side, higher perception thresholds. However, it is well doc-
umented that chronic pain is associated with reduced preci-
sion in perception of non- painful stimuli (Catley et al., 2014;
Moseley, 2008; Moseley et al., 2012). Our results fit well
with the ‘imprecision hypothesis’ of chronic pain (Moseley
& Vlaeyen, 2015). This hypothesis posits that chronic pain
patients learn associations between multisensory events,
or bodily states, and painful events (e.g. during an injury).
Similar bodily states can later increase the likelihood to per-
ceive pain again due to classical conditioning. This learning
mechanism is enhanced when bodily states are less precisely
perceived.
F I G U R E 3 Results of a multiple regression model in which
the intensity of phantom limb pain (PLP intensity) in the group of
amputees with phantom limb pain was statistically predicted by
a linear combination of heat pain thresholds (HPT) and warmth
detection thresholds (WDT) at the corner of the mouth contralateral
to the amputation. The three- dimensional scatter plots show the data
together with a regression plane, representing the values predicted by
the multiple regression model. Higher PLP intensity was associated
with lower HPT and higher WDT. The proportion of variance in PLP
intensity explained by the model and the regression coefficients for
HPT and WDT (with statistical significance: *p < .05; ***p < .001)
are shown within the three- dimensional (3D) scatter plot

126 | FUCHS ET AL.
4.3 | Differences between body sites and their possible relationship to peripheral versus central processes
For all sensory measures, we found significant differences
between body sites. Many of the results were expected based
on prior research. For example, the corners of the mouth
showed lower thresholds than all other body sites in all sen-
sory measures. With respect to processes of PLP, compari-
sons between the homologous body sites at the corners of
the mouth and the arms are of particular interest. The com-
parison between the arms (i.e. between the residual limb and
its contralateral counterpart) can be indicative of peripheral
changes. Differences between the corners of the mouth could
be indicative of cortical changes in early sensory processing
areas such as S1. We found that HPT and PPT were signifi-
cantly lower at the residual limb compared to the intact limb,
which could be viewed as a sign of peripheral hyperalgesia.
However, this local hyperalgesia was observed in both the
PLP and in the non- PLP group. Moreover, the difference
between limbs was not significantly correlated with PLP
intensity. These findings do not suggest a close relation-
ship between PLP and pain thresholds at the residual limb.
We also revisited the hypothesis of increased tactile acuity
of the residual limb. Contrary to early studies (Haber, 1955;
D. Katz, 1921; Teuber et al., 1949; Wilson et al., 1962),
but in line with more recent ones (Flor et al., 1998; Grüsser
et al., 2001; Hunter et al., 2005), we did not find significantly
lower 2PDT at the residual limb. There were also no sig-
nificant differences between the limbs in WDT. Taken to-
gether, these results support the conclusion drawn by Hunter
et al. (2005) that sensory measures taken at the residual limb
do not in any simple way reflect phantom phenomena. Our
data further suggest that they are instead more strongly re-
lated to RLP.
Because cortical reorganization in S1 is both associated
with PLP (Flor et al., 1995) and with sensory changes in the
face of healthy subjects (Muret et al., 2016), sensory thresh-
olds might be altered in the mouth region of arm amputees.
We did not find significant differences between the groups at
the corners of the mouth and there were no significant intra-
individual differences between the corners of the mouth for
any of the thresholds. As discussed above, HPT and WDT
in the face correlated with PLP intensity in a multivariate
analysis. Because the face is remote from the residual limb,
this likely represents a centrally mediated effect. The fact that
facial pain thresholds were specifically associated with PLP
is in line with the notion that preamputation pain leads to the
formation of cortical pain memories via growth or broader
recruitment of nociceptive neurons in S1, resulting in an in-
creased range of stimuli eliciting pain (Flor, 2002; Vierck
et al., 2013; Yi & Zhang, 2011). It is possible that this process
also spreads to the neighbouring face area due to PLP- related
S1 reorganization. However, this association was present at
both sides of the face, whereas PLP- related S1 reorganization
of the face was found to be lateralized (Flor et al., 1995), that
is mainly affecting S1 contralateral to the amputation. If fa-
cial sensitivity closely paralleled cortical reorganization, the
sensory changes would be expected to be lateralized as well,
with changes primarily in the half of the face that is ipsilat-
eral to the amputation. From this view, it is surprising that
we found the strongest associations for the contralateral face.
However, associations were also present for the ipsilateral
F I G U R E 4 Results from a linear mixed model (LMM) testing statistical prediction of both phantom limb pain intensity (PLP) and residual
limb pain intensity (RLP) by a linear combination of heat pain thresholds (HPT) and warmth detection thresholds (WDT) at the corner of the
mouth contralateral to the amputation as well as by HPT at the ipsilateral arm (residual limb). The plots highlight the two statistically significant
interactions between the type of pain (PLP vs. RLP) and the sensory threshold predictor. The left panel shows that PLP— but not RLP— was
associated with lower HPT at the corner of the mouth; the right panel shows that RLP— but not PLP— was associated with lower HPT at the
residual limb. The lines represent regression parameters derived from the LMM. The points represent partial residuals (i.e., data corrected for the
influence of the not shown predictors)

| 127FUCHS ET AL.
face, as evident from the correlations. Due to the high degree
of intercorrelations between the thresholds, they are redun-
dant, and the ipsilateral face dropped in multiple regression
models. Second, it is possible that different generalization
mechanisms overlap and that changes might have affected
the ipsilateral face at an earlier and the entire face at a later
stage. Further studies are needed to investigate whether there
is a relationship between facial cortical reorganization in PLP
and changes in the facial thermal thresholds.
Most studies on cortical reorganization processes in am-
putees used tactile (e.g. Flor et al., 1995) or motor methods
(e.g. Lotze et al., 2001) to map the cortical reorganization
of the hand and the lips. As a tactile measure, this study as-
sessed 2PDT. There was no significant association between
PLP presence or intensity and facial 2PDT. Studies in healthy
participants have shown that peripheral input to the hand al-
ters its S1 reorganization and that this effect spreads to the
face, leading to altered facial 2PDT (Muret et al., 2016).
From this perspective, 2PDT might have been altered in the
amputation group. Other studies, however, suggest that 2PDT
is not closely related to S1 organization and rather reflects
processes outside of S1, for example in the inferior parietal
cortex (Akatsuka et al., 2008).
4.4 | Specificity of associations for PLP versus RLP and other chronic pain
Residual limb pain intensity should be carefully compared
with PLP, because as a frequent concomitant in chronic PLP,
it might be a driver of plastic changes in the nociceptive sys-
tem. We therefore compared the statistical association be-
tween PLP and RLP intensity with sensory thresholds. This
analysis showed that HPT at the contralateral corner of the
mouth specifically correlates with PLP whereas HPT at the
residual limb is specifically associated with RLP. In fact, the
residual limb was the only measured body site where HPT
was not significantly correlated with PLP. With respect to
the finding that it was mainly PLP, and not RLP, that was
associated with pain thresholds at areas of the body that were
remote from the residual limb, it can be assumed that it might
primarily be PLP that is associated with central factors of
pain enhancement, whereas RLP might rather be associated
with peripheral factors related to sensitivity of the residual
limb.
There are similarities with other types of chronic pain. In
fibromyalgia, pain thresholds are lowered in a widespread
manner, both at so- called tender points and at other areas,
and PPT correlates negatively with clinical pain assess-
ments (Lautenbacher et al., 1994). Other studies reported
contralateral transfer of lowered PPT in complex regional
pain syndrome (CRPS), but not in arthritis patients (Palmer
et al., 2019), or in CRPS but not in a control group with other
chronic pain syndromes (Vatine et al., 1998). One study found
that transfer of lowered PPT to the forehead in CRPS was
more pronounced ipsilateral than contralateral to the affected
limb (Drummond & Finch, 2006). This finding is interesting
because it is in line with the idea that generalization might
be related to S1 reorganization, which is seen both in PLP
and CRPS (for a review see Di Pietro et al., 2013). Future
research could directly compare sensory processes in PLP,
CRPS and other chronic pain to advance the understanding of
specific processes in these chronic pain syndromes.
4.5 | Discrepancy between presence and intensity of PLP
Our results showed that, contrary to our hypothesis, lower
pain thresholds were not associated with having PLP. Lower
pain thresholds were observed in the PLP as well as in the
non- PLP group and were, as such, unspecific. However, for
those amputees who reported PLP, pain thresholds correlated
with PLP intensity. These two findings seem, at first glance,
almost incompatible. However, they might indicate that
lowered pain thresholds play a different role depending on
whether PLP is present or not. Such differential processing
of sensory information depending on the presence or absence
of pain is not unusual. In line with this, Davis et al. (2015)
suggested that the presence or absence of chronic pain is not
merely a quantitatively, but rather a qualitatively different
state. When, in their words, the ‘pain switch is turned on’,
pain- modulating factors might interact with each other in
a different way. As the presence of chronic pain is associ-
ated with changes in neural processing of sensory informa-
tion (Davis & Moayedi, 2013; Latremoliere & Woolf, 2009;
Woolf & Salter, 2000), memory, affective learning and emo-
tional processes (Flor & Turk, 2011), we propose that pain
thresholds might play a different role when PLP is present
compared to absent and influence its perceived intensity.
As another explanation, pain thresholds correlating
with PLP intensity, but not its presence per se, could also
be the end product of an etiological process. Central sen-
sitization resulting from long- term nociceptive input is a
potential explanation for the correlation between PLP in-
tensity and enhanced sensitivity to painful stimuli (Woolf
& Salter, 2000) and its generalization to remote areas of the
body (Le Bars, 2002) in participants with strong PLP. The
strong nociceptive input might decrease both pain thresh-
olds and lead to the formation of pain memories related to
PLP, resulting in an association between them. Yet, it is also
possible that participants who developed strong PLP already
had low pain thresholds before the amputation (Nikolajsen
et al., 2000). Low pain thresholds might have been present as
a preamputation trait, potentially promoting the emergence of
PLP. Preoperation pain sensitivity has been shown to predict

128 | FUCHS ET AL.
postoperative pain (Granot, 2009) and PLP early after ampu-
tation correlated positively with sensitivity to pain stimuli as-
sessed right before the amputation (Nikolajsen et al., 2000).
However, whether later- stage chronic pain is also associated
with pain sensitivity before injury is not known yet.
In one way or the other, sensory processes as such do not
seem to be able to explain why amputees develop chronic PLP
but it is possible that they interact with other factors. Factors
determining whether or not amputees develop chronic pain
are manifold (Flor, 2002) and altered pain processing might
be one among them (Flor et al., 2006; Nikolajsen et al., ,1997,
2000). Functional (Apkarian et al., 2013) and structural
(Mansour et al., 2013) brain connectivities have been shown
to predict the transition from acute to chronic back pain in
longitudinal studies. Although PLP patients might differ from
chronic back pain patients especially with respect to the role of
cognitive and emotional variables (Fuchs et al., 2015, 2018),
it should be clarified whether brain connectivity also predicts
chronic PLP and how this relates to sensory processing and
cortical reorganization. As these relevant questions need to be
addressed using a longitudinal design, the present study can,
by itself, not contribute to clarify these etiological questions.
Future studies should also investigate relationships be-
tween the decreased pain thresholds observed in this study
and other alterations of sensory processing that have been ob-
served in chronic pain. For example, diffuse noxious inhibi-
tory controls (DNIC) or conditioned pain modulation refer to
the fact that the response to a painful stimulus is reduced or
inhibited due to another concurrently applied painful stim-
ulus (Le Bars et al., 1979) and this process has been found
to be deficient in chronic pain (Lewis et al., 2012). In this
study, we only tested single painful stimuli but did not as-
sess pain inhibition as assessed in dual stimulation paradigms
testing DNIC. Sensitization and habituation to painful stim-
uli have been found to be altered in chronic pain (Kleinböhl
et al., 1999). In this study, habituation across HPT trials oc-
curred and reduced habituation correlated with PLP intensity,
which did, however, not explain the reduced pain thresholds
we have reported (see Supplementary Material, Figure S6).
Future research should use sensitization and habituation par-
adigms (e.g. Kleinböhl et al., 1999) to assess the role of these
variables in PLP.
4.6 | Limitations
There are some limitations for the present study. First, the
study is only based on psychophysical thresholds. While the
study design invites conclusions about underlying processes,
further studies using neuroimaging methods are needed
to further investigate whether the mechanisms at play are
peripheral or central. Second, the study relied on a cross-
sectional design and its findings are only correlational and
allow no direct conclusions about the etiology of PLP. Third,
the study was restricted to sensory mechanisms related to
the amputation and PLP and, in correlational analyses, RLP.
This leaves open how specific these findings are to PLP. This
could have been investigated by including another clinical
group with a pain condition other than PLP (e.g. chronic back
pain, CRPS, or fibromyalgia).
Another limitation arises from the choice of tested body
sites and used measures. In this study, even the remote sites,
such as the thenar of the contralateral limb, are connected
to the missing limb via interhemispheric connections. If
lowered pain thresholds more strongly generalize to body
parts with S1 representations adjacent to the hand than to
remote body parts, this suggests that generalization paral-
lels brain reorganization processes. For clarification of the
mechanisms of generalization, assessment of another, more
remote, site such as the contralateral foot would have been
informative. Furthermore, most studies on brain changes in
amputees are based on tactile or motor processes and the
present study used warmth detection as the primary detection
measure. The inclusion of mechanical detection thresholds
would have been advantageous but would have prolonged
the study protocol, which was not feasible in the context of
this study. Unfortunately, 2PDT, which have a close relation-
ship with S1 reorganization, were only available in a sub-
sample of the amputees, resulting in reduced testing power
for 2PDT and their exclusion from the multivariate analyses.
Hence, this study cannot answer some questions concerning
potential changes in tactile spatial acuity and its relationship
with PLP. The choice of body sites and the testing protocol
also diverged in some respects from the protocol proposed
by the DFNS (Rolke et al., 2006), which limits the compa-
rability of the results to other studies and to normative data
provided by the network (Magerl et al., 2010). For exam-
ple, the normative data for the face were assessed on the
cheeks for thermal thresholds and over the masseter muscle
for PPT, but we assessed them at the corners of the mouth
for theoretical reasons related to previous data on reorgani-
zation. Further, the DFNS protocol assessed thermal thresh-
olds on the hand dorsum while we used the thenar. While we
had theoretical reasons for using the corners of the mouth
instead of the cheeks, the thermal thresholds on the hand
could instead have been assessed on the dorsum of the hand
with the benefit of better comparability with the normative
data. Another deviation is that, for the thermal thresholds,
we used verbal responses and the experimenter pressed the
button as fast as possible. This procedure is not ideal be-
cause it creates a bias (overestimation) of the threshold due
to the reaction time of the experimenter. Second, the experi-
menter was not blinded concerning the group assignment of
the participants and could have been implicitly biased and
reacted slower (or faster) depending on the group assign-
ment of the participants, which could confound the data. The

| 129FUCHS ET AL.
experimenter tried best to always react as fast as possible
and to stay neutral. The finding of some results that were
contrary to our hypothesis suggests that this was successful.
In future studies, participants could use their feet to press the
response buttons, for example. In Supplementary Material,
we compared our thresholds with the DFNS normative data,
and we found that our WDT were inflated. This difference
could be related to the additional delay due to the experi-
menter's reaction time but should be interpreted with caution
due to the different body sites as mentioned above. Note that
these deviations are not problematic for the interpretation of
the present results because the protocol was identical for all
tested groups, but they limit the possibilities of comparison
with the normative data. This could be addressed in future
studies. For PPT at the thenar, where the protocols and body
site were identical, the thresholds match with the normative
data.
4.7 | Conclusions
In summary, our assessments of pain thresholds at different
body sites, remote from the residual limb, suggest that global
enhanced sensitivity to painful stimuli is related to the in-
tensity of PLP. The global pattern showed that amputees in
the PLP group with low levels of PLP were hypoalgesic and
those with strong PLP were hyperalgesic. For facial HPT, for
which the association with PLP was strongest, associations
were even stronger when WDT was also taken into account.
PLP was associated with decreased HPT but increased WDT
in the face. This pattern was specific to PLP. Pain thresholds
at the residual limb, however, were unrelated to PLP but spe-
cifically related to RLP.
Surprisingly, any differences in sensory thresholds be-
tween the groups were absent and so the mere presence of PLP
did not coincide with sensory thresholds at any body site. We
suggest that these global sensitivity changes reflect a central
aspect of sensory processing rather than peripheral mecha-
nisms, emphasizing the presence of ‘widespread hypersensi-
tivity’ which has already been suggested by Cronholm in 1951
(p. 119). Whether this phenomenon is related to plasticity pro-
cesses occurring after or to factors already present before am-
putation cannot be clarified based on the present data.
AUTHOR CONTRIBUTION
HF, MD and XF conceptualized and designed the study. XF
and PK collected the data. XF analyzed the data and drafted
the manuscript. All authors wrote and critically revised the
article and approved the final version.
ACKNOWLEDGEMENTS
This study was supported by a grant from the Deutsche
Forschungsgemeinschaft (SFB1158/B07) to HF and
JA and European Research Council Advanced grant
‘Phantom phenomena: A window to the mind and the brain'
(PHANTOMMIND, FP7/2007– 2013/230249) to HF. We
would also like to thank Astrid Wolf for help with recruit-
ment, Kristina Staudt and student assistants for their help
with data acquisition.
DATA AVAILABILITY STATEMENT
Data and code of this study are publicly available and can
be accessed via the Open Science Framework (https://osf.io/
v6hjy/).
REFERENCES
Akatsuka, K., Noguchi, Y., Harada, T., Sadato, N., & Kakigi, R. (2008).
Neural codes for somatosensory two- point discrimination in infe-
rior parietal lobule: An fMRI study. NeuroImage, 40(2), 852– 858.
https://doi.org/10.1016/j.neuro image.2007.12.013
Apkarian, A. V., Baliki, M. N., & Farmer, M. A. (2013). Predicting tran-
sition to chronic pain. Current Opinion in Neurology, 26(4), 360–
367. https://doi.org/10.1097/WCO.0b013 e3283 6336ad
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2014). Fitting linear
mixed- effects models using lme4. ArXiv Preprint arXiv:1406.5823.
Bekrater- Bodmann, R., Chung, B. Y., Richter, I., Wicking, M., Foell,
J., Mancke, F., Schmahl, C., & Flor, H. (2015). Deficits in pain per-
ception in borderline personality disorder: Results from the thermal
grill illusion. Pain, 156(10), 2084– 2092. https://doi.org/10.1097/j.
pain.00000 00000 000275
Bekrater- Bodmann, R., Schredl, M., Diers, M., Reinhard, I., Foell, J.,
Trojan, J., Fuchs, X., & Flor, H. (2015). Post- amputation pain is asso-
ciated with the recall of an impaired body representation in dreams—
results from a nation- wide survey on limb amputees. PLoS One,
10(3), e0119552. https://doi.org/10.1371/journ al.pone.0119552
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery
rate: A practical and powerful approach to multiple testing. Journal
of the Royal Statistical Society. Series B (Methodological), 57(1),
289– 300. https://doi.org/10.2307/2346101
Bonzano, L., Tacchino, A., Roccatagliata, L., Abbruzzese, G., Mancardi,
G. L., & Bove, M. (2008). Callosal contributions to simultaneous
bimanual finger movements. The Journal of Neuroscience, 28(12),
3227– 3233. https://doi.org/10.1523/JNEUR OSCI.4076- 07.2008
Catley, M. J., O’Connell, N. E., Berryman, C., Ayhan, F. F., & Moseley,
G. L. (2014). Is tactile acuity altered in people with chronic pain? A
systematic review and meta- analysis. The Journal of Pain, 15(10),
985– 1000. https://doi.org/10.1016/j.jpain.2014.06.009
Cronholm, B. (1951). Phantom limbs in amputees; a study of changes
in the integration of centripetal impulses with special reference to
referred sensations. Acta Psychiatrica Et Neurologica Scandinavica
Supplementum, 72, 1– 310.
Davis, K. D., Kucyi, A., & Moayedi, M. (2015). The pain switch:
An “ouch” detector. Pain, 156(11), 2164– 2166. https://doi.
org/10.1097/j.pain.00000 00000 000303
Davis, K. D., & Moayedi, M. (2013). Central mechanisms of pain revealed
through functional and structural MRI. Journal of Neuroimmune
Pharmacology, 8(3), 518– 534. https://doi.org/10.1007/s1148
1- 012- 9386- 8
Devor, M. (1991). Neuropathic pain and injured nerve: Peripheral
mechanisms. British Medical Bulletin, 47(3), 619– 630. https://doi.
org/10.1093/oxfor djour nals.bmb.a072496

130 | FUCHS ET AL.
Di Pietro, F., McAuley, J. H., Parkitny, L., Lotze, M., Wand, B. M.,
Moseley, G. L., & Stanton, T. R. (2013). Primary somatosensory
cortex function in complex regional pain syndrome: A systematic
review and meta- analysis. The Journal of Pain, 14(10), 1001– 1018.
https://doi.org/10.1016/j.jpain.2013.04.001
Dietrich, C., Walter- Walsh, K., Preißler, S., Hofmann, G. O., Witte,
O. W., Miltner, W. H. R., & Weiss, T. (2012). Sensory feed-
back prosthesis reduces phantom limb pain: Proof of a principle.
Neuroscience Letters, 507(2), 97– 100. https://doi.org/10.1016/j.
neulet.2011.10.068
Dinse, H. R. (2003). Pharmacological modulation of perceptual learning
and associated cortical reorganization. Science, 301(5629), 91– 94.
https://doi.org/10.1126/scien ce.1085423
Dixon, W. J., & Mood, A. M. (1948). A method for obtaining and
analyzing sensitivity data. Journal of the American Statistical
Association, 43(241), 109– 126. https://doi.org/10.1080/01621
459.1948.10483254
Drummond, P. D., & Finch, P. M. (2006). Sensory changes in the fore-
head of patients with complex regional pain syndrome. Pain, 123(1–
2), 83– 89. https://doi.org/10.1016/j.pain.2006.02.013
Engkvist, O., Wahren, L. K., Wallin, G., Torebjrk, E., & Nystrm, B.
(1985). Effects of regional intravenous guanethidine block in post-
traumatic cold intolerance in hand amputees. Journal of Hand
Surgery (British and European Volume), 10(2), 145– 150. https://
doi.org/10.1016/0266- 7681(85)90003 - 8
Flor, H. (2002). Phantom- limb pain: Characteristics, causes, and treat-
ment. The Lancet Neurology, 1(3), 182– 189. https://doi.org/10.1016/
S1474 - 4422(02)00074 - 1
Flor, H., Elbert, T., Knecht, S., Wienbruch, C., Pantev, C., Birbaumers,
N., Larbig, W., & Taub, E. (1995). Phantom- limb pain as a percep-
tual correlate of cortical reorganization following arm amputation.
Nature, 375(6531), 482– 484. https://doi.org/10.1038/375482a0
Flor, H., Elbert, T., Mühlnickel, W., Pantev, C., Wienbruch, C., & Taub,
E. (1998). Cortical reorganization and phantom phenomena in
congenital and traumatic upper- extremity amputees. Experimental
Brain Research, 119(2), 205– 212. https://doi.org/10.1007/s0022
10050334
Flor, H., & Fuchs, X. (2017). Phantom limb pain. In Reference module in
neuroscience and biobehavioral psychology. Elsevier. https://www.
scien cedir ect.com/scien ce/artic le/pii/B9780 12809 32450 31436
Flor, H., Nikolajsen, L., & Jensen, T. S. (2006). Phantom limb pain: A
case of maladaptive CNS plasticity? Nature Reviews Neuroscience,
7(11), 873– 881. https://doi.org/10.1038/nrn1991
Flor, H., Rudy, T. E., Birbaumer, N., Streit, B., & Schugens, M. M.
(1990). Zur Anwendbarkeit des West Haven- Yale Multidimensional
Pain Inventory im deutschen Sprachraum [The applicability of the
West Haven- Yale multidimensional pain inventory in German-
speaking countries. Data on the reliability and validity of the MPI-
D.]. Der Schmerz, 4(2), 82– 87. https://doi.org/10.1007/BF025 27839
Flor, H., & Turk, D. C. (2011). Chronic pain: An integrated biobehav-
ioral approach (1 Pap/Cdr edition). IASP Press.
Fried, K., Govrin- Lippmann, R., Rosenthal, F., Ellisman, M. H., &
Devor, M. (1991). Ultrastructure of afferent axon endings in a
neuroma. Journal of Neurocytology, 20(8), 682– 701. https://doi.
org/10.1007/BF011 87069
Fuchs, X., Bekrater- Bodmann, R., & Flor, H. (2015). Phantom pain:
The role of maladaptive plasticity and emotional and cognitive vari-
ables. In G. Pickering & S. Gibson (Eds.), Pain, emotion and cog-
nition (pp. 189– 207). Springer International Publishing. https://doi.
org/10.1007/978- 3- 319- 12033 - 1_12
Fuchs, X., Flor, H., & Bekrater- Bodmann, R. (2018). Psychological
factors associated with phantom limb pain: A review of recent find-
ings. Pain Research and Management, 2018, 5080123. https://doi.
org/10.1155/2018/5080123
Granot, M. (2009). Can we predict persistent postoperative pain
by testing preoperative experimental pain? Current Opinion
in Anaesthesiology, 22(3), 425– 430. https://doi.org/10.1097/
ACO.0b013 e3283 2a40e1
Grüsser, S. M., Winter, C., Mühlnickel, W., Denke, C., Karl, A.,
Villringer, K., & Flor, H. (2001). The relationship of perceptual
phenomena and cortical reorganization in upper extremity ampu-
tees. Neuroscience, 102(2), 263– 272. https://doi.org/10.1016/S0306
- 4522(00)00491 - 7
Haber, W. B. (1955). Effects of loss of limb on sensory func-
tions. The Journal of Psychology, 40(1), 115– 123. https://doi.
org/10.1080/00223 980.1955.9712969
Harden, R. N., Gagnon, C. M., Khan, A., Wallach, G., & Zereshki,
A. (2010). Hypoesthesia in the distal residual limb of am-
putees. PM&R, 2(7), 607– 611. https://doi.org/10.1016/j.
pmrj.2010.03.033
Hunter, J. P., Katz, J., & Davis, K. D. (2005). Dissociation of phan-
tom limb phenomena from stump tactile spatial acuity and sensory
thresholds. Brain, 128(2), 308– 320. https://doi.org/10.1093/brain/
awh350
Katz, D. (1921). Zur Psychologie des Amputierten und seiner Prothese.
Barth.
Katz, J. (1992). Psychophysical correlates of phantom limb experience.
Journal of Neurology, Neurosurgery & Psychiatry, 55(9), 811– 821.
https://doi.org/10.1136/jnnp.55.9.811
Kern, U., Busch, V., Rockland, M., Kohl, M., & Birklein, F. (2009).
Prävalenz und Risikofaktoren von Phantomschmerzen und
Phantomwahrnehmungen in Deutschland [Prevalence and risk fac-
tors of phantom limb pain and phantom limb sensations in Germany.
A nationwide field survey]. Schmerz (Berlin, Germany), 23(5), 479–
488. https://doi.org/10.1007/s0048 2- 009- 0786- 5
Kerns, R. D., Turk, D. C., & Rudy, T. E. (1985). The West Haven- Yale
Multidimensional Pain Inventory (WHYMPI). Pain, 23(4), 345–
356. https://doi.org/10.1016/0304- 3959(85)90004 - 1
Kikkert, S., Johansen- Berg, H., Tracey, I., & Makin, T. R. (2018).
Reaffirming the link between chronic phantom limb pain and main-
tained missing hand representation. Cortex, 106, 174– 184. https://
doi.org/10.1016/j.cortex.2018.05.013
Kleinböhl, D., Hölzl, R., Möltner, A., Rommel, C., Weber, C., &
Osswald, P. M. (1999). Psychophysical measures of sensitization to
tonic heat discriminate chronic pain patients. Pain, 81(1), 35– 43.
https://doi.org/10.1016/S0304 - 3959(98)00266 - 8
Korin, H., Weiss, S. A., & Fishman, S. (1963). Pain sensitivity of am-
putation extremities. The Journal of Psychology, 55(2), 345– 355.
https://doi.org/10.1080/00223 980.1963.9916627
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. B. (2014).
lmerTest: Tests for random and fixed effects for linear mixed effect
models (lmer objects of lme4 package). http://CRAN.R- proje ct.org/
packa ge=lmerTest
Latremoliere, A., & Woolf, C. J. (2009). Central sensitization: A gener-
ator of pain hypersensitivity by central neural plasticity. The Journal
of Pain, 10(9), 895– 926. https://doi.org/10.1016/j.jpain.2009.06.012
Lautenbacher, S., Rollman, G. B., & McCain, G. A. (1994).
Multi- method assessment of experimental and clinical pain
in patients with fibromyalgia. Pain, 59(1), 45– 53. https://doi.
org/10.1016/0304- 3959(94)90046 - 9

| 131FUCHS ET AL.
Le Bars, D. (2002). The whole body receptive field of dorsal horn multi-
receptive neurones. Brain Research. Brain Research Reviews, 40(1–
3), 29– 44. https://doi.org/10.1016/S0165 - 0173(02)00186 - 8
Le Bars, D., Dickenson, A. H., & Besson, J.- M. (1979). Diffuse nox-
ious inhibitory controls (DNIC). I. Effects on dorsal horn con-
vergent neurones in the rat. Pain, 6(3), 283– 304. https://doi.
org/10.1016/0304- 3959(79)90049 - 6
Levitt, H. (1971). Transformed up- down methods in psychoacous-
tics. The Journal of the Acoustical Society of America, 49(2),
Suppl2:467+.
Lewis, G. N., Rice, D. A., & McNair, P. J. (2012). Conditioned pain
modulation in populations with chronic pain: A systematic review
and meta- analysis. The Journal of Pain, 13(10), 936– 944. https://
doi.org/10.1016/j.jpain.2012.07.005
Li, S., Melton, D. H., & Li, S. (2015). Tactile, thermal, and electrical
thresholds in patients with and without phantom limb pain after
traumatic lower limb amputation. Journal of Pain Research, 8, 169–
174. https://doi.org/10.2147/JPR.S77412
Lotze, M., Flor, H., Grodd, W., Larbig, W., & Birbaumer, N. (2001).
Phantom movements and pain An fMRI study in upper limb am-
putees. Brain, 124(11), 2268– 2277. https://doi.org/10.1093/brain/
124.11.2268
Lotze, M., Grodd, W., Birbaumer, N., Erb, M., Huse, E., & Flor, H.
(1999). Does use of a myoelectric prosthesis prevent cortical reorga-
nization and phantom limb pain? Nature Neuroscience, 2(6), 501–
502. https://doi.org/10.1038/9145
MacIver, K., Lloyd, D. M., Kelly, S., Roberts, N., & Nurmikko, T.
(2008). Phantom limb pain, cortical reorganization and the thera-
peutic effect of mental imagery. Brain, 131(8), 2181– 2191. https://
doi.org/10.1093/brain/ awn124
Magerl, W., Krumova, E. K., Baron, R., Tölle, T., Treede, R.- D., &
Maier, C. (2010). Reference data for quantitative sensory testing
(QST): Refined stratification for age and a novel method for statis-
tical comparison of group data. Pain, 151(3), 598– 605. https://doi.
org/10.1016/j.pain.2010.07.026
Maier, C., Baron, R., Tölle, T. R., Binder, A., Birbaumer, N., Birklein,
F., Gierthmühlen, J., Flor, H., Geber, C., Huge, V., Krumova, E.
K., Landwehrmeyer, G. B., Magerl, W., Maihöfner, C., Richter,
H., Rolke, R., Scherens, A., Schwarz, A., Sommer, C., …
Treede, D.- R. (2010). Quantitative sensory testing in the German
Research Network on Neuropathic Pain (DFNS): Somatosensory
abnormalities in 1236 patients with different neuropathic pain
syndromes. Pain, 150(3), 439– 450. https://doi.org/10.1016/j.
pain.2010.05.002
Makin, T. R., Cramer, A. O., Scholz, J., Hahamy, A., Slater, D. H.,
Tracey, I., & Johansen- Berg, H. (2013). Deprivation- related and
use- dependent plasticity go hand in hand. Elife, 2, e01273. https://
doi.org/10.7554/eLife.01273
Makin, T. R., Scholz, J., Filippini, N., Henderson Slater, D., Tracey,
I., & Johansen- Berg, H. (2013). Phantom pain is associated with
preserved structure and function in the former hand area. Nature
Communications, 4, 1570. https://doi.org/10.1038/ncomm s2571
Mancini, F., Bauleo, A., Cole, J., Lui, F., Porro, C. A., Haggard, P.,
& Iannetti, G. D. (2014). Whole- body mapping of spatial acu-
ity for pain and touch: Mapping spatial acuity for pain. Annals of
Neurology, 75(6), 917– 924, https://doi.org/10.1002/ana.24179
Mansour, A. R., Baliki, M. N., Huang, L., Torbey, S., Herrmann, K. M.,
Schnitzer, T. J., & Apkarian, A. V. (2013). Brain white matter struc-
tural properties predict transition to chronic pain. Pain, 154(10),
2160– 2168. https://doi.org/10.1016/j.pain.2013.06.044
Moseley, G. L. (2008). I can’t find it! Distorted body image and tactile
dysfunction in patients with chronic back pain. Pain, 140(1), 239–
243. https://doi.org/10.1016/j.pain.2008.08.001
Moseley, G. L., Gallagher, L., & Gallace, A. (2012). Neglect- like tac-
tile dysfunction in chronic back pain. Neurology, 79(4), 327– 332.
https://doi.org/10.1212/WNL.0b013 e3182 60cba2
Moseley, G. L., & Vlaeyen, J. W. S. (2015). Beyond nociception: The
imprecision hypothesis of chronic pain. Pain, 156(1), 35– 38. https://
doi.org/10.1016/j.pain.00000 00000 000014
Muret, D., Daligault, S., Dinse, H. R., Delpuech, C., Mattout, J., Reilly,
K. T., & Farnè, A. (2016). Neuromagnetic correlates of adaptive
plasticity across the hand- face border in human primary somato-
sensory cortex. Journal of Neurophysiology, 115(4), 2095– 2104.
https://doi.org/10.1152/jn.00628.2015
Nikolajsen, L. (2013). Phantom limb. In S. B. McMahon, P. D. Wall, &
R. Melzack (Eds.), Wall and Melzack’s textbook of pain (6th ed., pp.
915– 925). Elsevier, Saunders.
Nikolajsen, L., Ilkjær, S., & Jensen, T. S. (2000). Relationship be-
tween mechanical sensitivity and postamputation pain: A prospec-
tive study. European Journal of Pain, 4(4), 327– 334. https://doi.
org/10.1053/eujp.2000.0194
Nikolajsen, L., Ilkjær, S., Krøner, K., Christensen, J. H., & Jensen, T.
S. (1997). The influence of preamputation pain on postamputa-
tion stump and phantom pain. Pain, 72(3), 393– 405. https://doi.
org/10.1016/S0304 - 3959(97)00061 - 4
Oldfield, R. C. (1971). The assessment and analysis of handedness: The
Edinburgh inventory. Neuropsychologia, 9(1), 97– 113. https://doi.
org/10.1016/0028- 3932(71)90067 - 4
Palmer, S., Bailey, J., Brown, C., Jones, A., & McCabe, C. S. (2019).
Sensory function and pain experience in arthritis, complex regional
pain syndrome, fibromyalgia syndrome, and pain- free volunteers:
A cross- sectional study. The Clinical Journal of Pain, 35(11), 894–
900. https://doi.org/10.1097/AJP.00000 00000 000751
Pinheiro, J. C., & Bates, D. M. (2000). Mixed- effects models in S and
S- PLUS. Springer Science & Business Media.
R Core Team (2014). R: A language and environment for statistical
computing. R Foundation for Statistical Computing. http://www.R-
proje ct.org/
Ramachandran, V. S., & Hirstein, W. (1998). The perception of phantom
limbs. The D. O. Hebb lecture. Brain, 121(9), 1603– 1630. https://
doi.org/10.1093/brain/ 121.9.1603
Rolke, R., Baron, R., Maier, C., Tölle, T. R., Treede, D. R., Beyer, A.,
Binder, A., Birbaumer, N., Birklein, F., Bötefür, I. C., Braune, S.,
Flor, H., Huge, V., Klug, R., Landwehrmeyer, G. B., Magerl, W.,
Maihöfner, C., Rolko, C., Schaub, C., … Wasserka, B. (2006).
Quantitative sensory testing in the German Research Network
on Neuropathic Pain (DFNS): Standardized protocol and refer-
ence values. Pain, 123(3), 231– 243. https://doi.org/10.1016/j.
pain.2006.01.041
Sherman, R. A., Arena, J. G., Sherman, C. J., & Ernst, J. L. (1989). The
mystery of phantom pain: Growing evidence for psychophysiologi-
cal mechanisms. Biofeedback and Self- Regulation, 14(4), 267– 280.
https://doi.org/10.1007/BF009 99118
Teuber, H.- L., Krieger, H., & Bender, M. (1949). Reorganization of
sensory function in amputation stumps: Two- point discrimination.
Federal Proceedings, 8:156.
van den Heuvel, M. P., & Hulshoff Pol, H. E. (2009). Specific somato-
topic organization of functional connections of the primary motor
network during resting state. Human Brain Mapping, 4(31), 631–
644. https://doi.org/10.1002/hbm.20893

132 | FUCHS ET AL.
Vase, L., Nikolajsen, L., Christensen, B., Egsgaard, L. L., Arendt-
Nielsen, L., Svensson, P., & Staehelin Jensen, T. (2011). Cognitive-
emotional sensitization contributes to wind- up- like pain in phantom
limb pain patients. Pain, 152(1), 157– 162. https://doi.org/10.1016/j.
pain.2010.10.013
Vaso, A., Adahan, H.- M., Gjika, A., Zahaj, S., Zhurda, T., Vyshka, G.,
& Devor, M. (2014). Peripheral nervous system origin of phan-
tom limb pain. Pain, 155(7), 1384– 1391. https://doi.org/10.1016/j.
pain.2014.04.018
Vatine, J.- J., Tsenter, J., & Nirel, R. (1998). Experimental pressure pain
in patients with complex regional pain syndrome, type I (reflex
sympathetic dystrophy). American Journal of Physical Medicine &
Rehabilitation, 77(5), 382– 387.
Vierck, C. J., Whitsel, B. L., Favorov, O. V., Brown, A. W., &
Tommerdahl, M. (2013). Role of primary somatosensory cortex in
the coding of pain. Pain, 154(3), 334– 344. https://doi.org/10.1016/j.
pain.2012.10.021
Wahren, L. K. (1990). Changes in thermal and mechanical pain thresholds
in hand amputees. A clinical and physiological long- term follow- up.
Pain, 42(3), 269– 277. https://doi.org/10.1016/0304- 3959(90)91139 - A
Weinstein, S. (1968). Intensive and extensive aspects of tactile sensitiv-
ity as a function of body part, sex, and laterality. In D. R. Kenshalo
(Ed.), The skin senses: Proceedings of the first International
Symposium on the Skin Senses, held at the Florida State University in
Tallahassee, Florida (pp. 195– 218). Charles C. Thomas Publishing.
Wickham, H. (2009). ggplot2: Elegant graphics for data analysis.
Springer Science & Business Media.
Wiffen, P., Meynadier, J., Dubois, M., Thurel, C., DeSmet, J., & Harden,
R. N. (2006). Chapter 4: Diagnostic and treatment issues in postam-
putation pain after landmine injury. Pain Medicine, 7(Suppl 2),
S209– S212. https://doi.org/10.1111/j.1526- 4637.2006.00234_6.x
Wilson, J. J., Wilson, B. C., & Swinyard, C. A. (1962). Two- point dis-
crimination in congenital amputees. Journal of Comparative and
Physiological Psychology, 55(4), 482– 485. https://doi.org/10.1037/
h0042122
Woolf, C. J. (2014). What to call the amplification of nociceptive
signals in the central nervous system that contribute to wide-
spread pain? Pain, 155(10), 1911– 1912. https://doi.org/10.1016/
j.pain.2014.07.021
Woolf, C. J., & Salter, M. W. (2000). Neuronal Plasticity: Increasing
the Gain in Pain. Science, 288(5472), 1765– 1768. https://doi.
org/10.1126/scien ce.288.5472.1765
Yi, M., & Zhang, H. (2011). Nociceptive memory in the brain: Cortical
mechanisms of chronic pain. The Journal of Neuroscience, 31(38),
13343– 13345. https://doi.org/10.1523/JNEUR OSCI.3279- 11.2011
SUPPORTING INFORMATION
Additional Supporting Information may be found online in
the Supporting Information section.
How to cite this article: Fuchs, X., Diers, M., Trojan,
J., Kirsch, P., Milde, C., Bekrater- Bodmann, R.,
Rance, M., Foell, J., Andoh, J., Becker, S., & Flor, H.
(2022). Phantom limb pain after unilateral arm
amputation is associated with decreased heat pain
thresholds in the face. European Journal of Pain, 26,
114– 132. https://doi.org/10.1002/ejp.1842




















