
A model of the cerebellum in adaptive control of saccadic gain
Upload
royalhollowayCategory
view
0download
0
~ Pergamon 0028 3932(95)00156 5
Neurops.vcholoclia, Voh 34, No. 6. pp. 493 508, 1996 Copyright ! 1996 Elsevier Science Ltd. All rights reserved
Printed in Great Britain 0028 3932/96 S15.00+0.00
Saccadic eye movement programming in unilateral neglect
R. WALKER* and J. M. FINDLAY
Department of Psychology, University of Durham, Durham DH1 3LE, U.K.
(Receired 19 April 1995; accepted 20 September 1995)
Abstract--The present study examined the eye movements made by patients with unilateral neglect under fixation gap and overlap conditions. The prior offset of fixation in a + 100 msec gap condition did not produce an increase in the numbers of contralesional saccades made by 3 out of 4 patients. This finding is incompatible with the view that the deficit in producing contralesional saccades reflects an inability to disengage visual attention from fixation. A significant reduction in saccade latency was, however, obtained in the gap condition ('gap effect'). The latency reduction in the gap condition is consistent with models which attribute the gap effect to warning signal effects and the release of an ocular rather than an attentional disengagement mechanism. Saccade latency was not increased when two targets were presented bilaterally and simultaneously in both hemifields (in contrast to the increase in latency shown by normal subjects). The lack of a normal 'bilateral target effect' in neglect is attributed to an imbalance in the level of activity in the saccadic system. Three patients showed visual 'extinction' and did not make saccades to the contralesional bilateral targets. In contrast R.R. who shows object-based neglect did not show extinction and made saccades to the contralesional bilateral targets. This suggests that visual extinction may be influenced by the form of neglect shown by the patient. The effects on saccade amplitude of presenting two targets in the same hemifield were also examined in a global effect task. One patient showed a much greater global effect than normal when pairs of targets were presented in his ipsilesional hemifield. In contrast R.R. showed a normal magnitude global effect. It appears that the form of neglect shown by a patient is a factor that influences their eye movement behaviour on simple saccade tasks and these eye movement abnormalities cannot be accounted for by a deficit of attentional disengagement. Copyright © 1996 Elsevier Science Ltd.
Key Words: unilateral neglect; gap effect; global effect; bilateral target effect; parietal lobe; human.
Introduction
Unilateral neglect is a condit ion in which a patient with brain damage, usually in the region of the right parietal lobe, fails to respond to, or be aware of, people and objects in the contralateral side of space. Patients show neglect on a range of tasks, such as: reading, drawing and mental imagery. Al though neglect does not result f rom a failure to scan the contralesional side o f space, neglect patients can show a range of deficits in making con- tralesional saccadic eye movements [9, 36]. The defective patterns o f saccades that result f rom neglect are dis- tinguishable fi'om those that result f rom a hemianopia [11, 18, 19]. Girotti et al. [9] showed that patients with unilateral neglect failed to make saccades to targets on a quarter o f all trials when a stimulus appeared in the contralesional visual field. His patients made multiple
* Address for correspondence: R. Walker, Academic Unit of Neuroscience, Charing Cross Hospital, Fulham Palace Road, London W6 8RF, U.K.; e-mail: ragt(a cxwms.ac.uk.
small saccades to locate targets in their left visual field. In contrast control patients with cortical lesions and hemianopias (without neglect) were able to make single saccades to locate targets in their blind field. The time to initiate a contralesional saccade (latency) was greater in neglect patients than in hemianopic patients. Girotti et al. found the same defective pattern o f eye movements in a neglect patient who had intact visual fields and con- cluded that the abnormal eye movement patterns cannot be attributed to a field defect. Furthermore, as patients with genuine field defects are known to be able to learn to compensate for their loss of vision [1, 18, 19], it appears that it is neglect and not a visual field defect that produces a long term impairment in saccade generation [ l l , 18, 4O].
Some models of neglect have proposed that the deficits in orienting attention in the direction contralateral to the lesion results f rom an inability to 'disengage' visual attention. The disengagement deficit is thought to be selective and to impair attentional shifts made in the contralesional, but not the ipsilesional, direction. Sup-
493

494 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
port for such models has come from manual reaction time studies with neglect patients, which have shown a selective deficit of covert attentional orienting in the contralesional direction following an ipsilesional cue [26]. The concept of attentional disengagement has also been used to explain the systematic reduction in saccade lat- ency observed with normal subjects when a central fix- ation point is extinguished before the onset of a saccade target ('gap paradigm') [34]. Fisher [5, 8] suggested that fixation offset in the gap situation automatically dis- engages visual attention. Disengagement of attention is held to be a process which has to be completed before an attentional shift (by an overt saccade) can be accomplished and fixation offset in the gap situation is thought to reduce saccade latency by automatically achieving the disengagement operation prior to the onset of the saccade target.
The gap paradigm has been shown to reduce saccade latency and to increase the numbers of contralateral sac- cades made by a patient with unilateral neglect [40]. Walker el al. [40] examined the effects of fixation offset in a patient, B.Q., who showed severe left-sided neglect following damage in the region of the right parietal lobe. In an overlap condition B.Q. did not make saccades to unilateral targets presented in the left visual field. However, in a gap condition where the fixation point was removed 100 msec before the onset of the target B.Q. made normal saccades on 50% of trials. B.Q.'s improve- ment in making contralesional saccades was interpreted as being consistent with the attentional disengagement deficit model of neglect [26] and the attentional dis- engagement hypothesis of the gap effect [6]. It was noted in this study however that the gap condition also increased the numbers of saccades made to targets pre- sented in the patient's ipsilesional visual field.
There is, however, an alternative explanation to that of attentional disengagement for the facilitation effect observed in gap situations [28, 29]. Ross and Ross [32, 33] showed that the onset, offset or change, of a visual event at fixation prior to the onset of the saccade target all produced a reduction in saccade latency although stimulus offset was found to be the most effective. They proposed that fixation onset, offset or change, act as warning signals that enable partial pre-programming of the saccade prior to the onset of the peripheral stimulus. Such a warning signal effect has been confirmed by Reuter-Lorenz et al. [29], who showed that the gap facili- tation effect was reduced when an auditory warning sig- nal occurred along with the offset of the fixation point. A second process is needed to account for the increased effectiveness of visual offsets. Tam and Stelmach [39] proposed that release from a process termed ocular fix- ation accounts for this additional part of the gap latency reduction. Munoz and Wurtz [21 23] showed that the superior colliculus contains a region which operates rather like this proposed ocular fixation mechanism. They found that monkeys produced faster contralateral sac- cades when cells in the rostral pole region of the superior
colliculus were inhibited (by the injection of the GABA agonist muscimol). It has been suggested that the rostral pole acts as a mechanism for suppressing gaze shifts dur- ing fixation via projections to the omnipause neurons in the brain stem that are involved in the generation of saccades [24]. Thus, it appears that the gap effect can be accounted for in terms of a combination of a generalized warning signal that enables partial saccade pre-pro- gramming together with the release of an ocular motor fixation mechanism.
Release of ocular fixation appears to be a similar con- cept to the disengagement of attention. However an important difference is that disengagement of attention implies that attention can be relocated away from fixation. Two recent experiments [12, 41] have tested whether the prior allocation of attention away from the fixation point influences the gap facilitation effect. In this situation attention should have been disengaged and moved away from fixation and so the latency reduction in gap conditions should not be as great as when the subject is attending to the fixation point. Both experi- ments found the latency reduction to be unaffected by the prior orienting of attention, weakening support for the attentional disengagement hypothesis and favouring the alternative of the release of an ocular fixation mech- anism.
If the gap effect is not produced by prior attentional disengagement then an alternative explanation must be found for the increased incidence of contralesional sac- cades produced in the gap condition by neglect patients. In this study, we aim to examine the eye movements made by patients with neglect to see if the gap situation produces a general improvement in the patient's ability to make saccades. The attentional disengagement hypothesis predicts a selective improvement in the pa- tient's ability to make contralesional saccades. The alter- native explanation of a warning signal effect predicts an improvement in contralesional saccades only in patients who are able to make use of such 'cues' to initiate a search for stimuli presented in their contralesional visual field [31].
A second point of interest is the effect of presenting targets bilaterally and simultaneously in both hemifields on saccade generation. In this situation the latency of saccades made by normal subjects is found to increase by some 20~40 msec [3, 16]. This latency increase has been found to be maximal for simultaneous onsets and is not thought to result from a simple conflict in saccade direc- tion [41]. In contrast it has been reported that the neglect patient B.Q. did not show the normal increase in latency with bilateral simultaneous target presentation [41]. B.Q.'s failure to show the normal latency increase cannot be accounted for in terms of a contralateral visual field defect as she was not hemianopic. It thus appears that bilateral target presentation produces an inhibitory effect for saccade generation in normal subjects and this mech- anism is defective in patients with unilateral neglect. The loss of such an inhibitory mechanism could explain the bias of saccades made by the patients in the ipsilesional

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 495
direct ion. The present exper iment aims to examine the effect o f b i la te ra l target p resen ta t ion on saccade la tency in o ther pat ients with neglect to see if the lack o f a bi la tera l la tency increase is a c o m m o n finding.
In con t ras t to the s i tua t ion with two targets presented in bo th hemifields, it has been shown that present ing two targets at different eccentricit ies in the same hemifield does not a l ter the la tency o f saccades made by norma l subjects, but does al ter the landing pos i t ion o f the saccade endpoint . W h e n two targets are presented at different eccentricit ies in the same hemifield saccades made to the near target show a systematic overshoot . This overshoo t has been te rmed the 'g loba l effect' [2]. Neglect pat ients are known to bias their a t ten t ion [13 15] and eye move- ments in the ipsi lesional d i rect ion [10]. It has been shown that neglect pat ients have an a t ten t iona l bias for the more per iphera l st imuli in their ipsi lesional visual field [14, 15]. I f this is the case then it could be predic ted tha t a greater g lobal effect will be shown by neglect pat ients when pairs of targets are presented in their ipsi lesional visual field than occurs in no rma l subjects. Similar ly a smaller g lobal effect would be expected when pairs of targets are pre- sented in their con t ra les iona l visual field as saccades m a y be di rec ted to the mos t ipsi lesional target which is the target nearest to the f ixat ion point . These predic t ions are examined in a g lobal effect task with two neglect pat ients .
We examined the effects o f f ixation offset (gap effect), b i la tera l target p resen ta t ion and the g lobal effect in pat- ients with uni la tera l neglect. The main quest ions o f inter- est are: (a) will the gap effect improve the pa t ien t ' s abi l i ty to p roduce cont ra les iona l saccades; (b) will pat ients with neglect show the la tency reduct ion character is t ic of a no rma l gap effect; (c) does saccade la tency increase when two targets are presented b i la tera l ly in oppos i te hem- ifields; and (d) do neglect pat ients show a no rma l g lobal effect when two targets are presented in the same hemi- field. Exper iment 1 examined the gap and bi la tera l target effect in four neglect pat ients . In Exper iment 2 the g lobal effect was examined in two neglect pat ients .
General method
Patients
Four male patients with right brain damage and a diagnosis of unilateral neglect acted as subjects. Their ages ranged from 59-64 years. None of the patients were known to have any other neurological disorders or general intellectual impairments. All of the patients were right handed. Table 1 gives the clinical details of the patients and their performance on tests sensitive to the presence of unilateral neglect. The visual fields of one patient, R.R., were assessed using a Henson automatic field plotter and the plot for one eye is shown in Fig. I. It is important to note that R.R. does not have a left hemianopia and that his upper left and upper and lower right visual fields have normal thresholds. An increase in threshold is apparent in his lower left visual field only for eccentricities greater than 10".
Apparatus
Stimuli were presented on a Phillips TP-200 monochrome monitor, controlled by a BBC microcomputer. Horizontal eye movements were recorded using the differential limbus reflec- tion technique [45] using the Skalar IRIS system [27]. The ana- logue eye movement signal was sampled and digitized every 5 msec by an Apple Macintosh computer using a National Instruments interface (LabNBoard). A signal from the BBC microcomputer, synchronized with the screen raster, initiated the start of the analogue-digital conversion of the eye movement signal following stimuli onset. A chin rest and head restraint were used to restrict movements of the head.
Experiment 1. The gap and bilateral target effects
Method
Subjects. Four patients with unilateral neglect following right brain damage (see Table 1) acted as subjects in Experiment 1.
Procedure. The patients viewed the VDU screen from a dis- tance of 28 cm. Each patient completed two blocks of fixation overlap trials and two blocks of + 100 msec gap trials, in an ABBA design (overlap, gap, gap, overlap). Single targets were presented unilaterally at eccentricities of 4 c', 8 or 12' to the left and right of fixation. Bilateral targets were presented sim- ultaneously, at equal and opposite eccentricity locations - 4 ' and + 4 , - 8 ° and +8" and - 1 2 ' and + 12. Ten targets were presented at each eccentricity location, producing a total of 30 left single targets, 30 right single targets and 30 bilateral simultaneous targets in each block. Patients completed two experimental blocks for each fixation condition, producing a total of 180 trials in each condition.
At the start of each trial a fixation cross (1.O appeared in the centre of the VDU screen. The target was a uniformly illuminated square (sides 1.0f), which was presented for 500 msec on a horizontal axis level with the fixation point. To encourage accurate saccades the central pixels of each stimulus square were not illuminated leaving a small black central point which the patient was asked to fixate. On single target trials a target appeared at one of the six eccentricity locations in a random order. On bilateral target trials, two targets appeared bilaterally and simultaneously, at equal and opposite eccen- tricity locations. The target was extinguished and an inter-trial interval of 2 sec occurred before the next trial.
Stimulus timing. Overlap condition: The fixation point remained on throughout each trial. Saccade targets appeared after an interval of 1000 msec following the onset of the fixation point.
Gap condition: The fixation point appeared for 900 rnsec and then went off. A delay of 100 msec then occurred during which time the screen remained black (+ 100 msec gap). Targets were presented immediately after the gap interval and appeared 1000 msec after the onset of fixation as was the case in the overlap condition.
A calibration routine was performed prior to each block of trials. The patient was asked to track a small dot surrounded by a circle (1.5 radius) which moved in a series of five equal steps from 12" left of fixation, to centre, to 12 right of fixation. As patients had neglect, care was taken to ensure that they moved their eyes to fixate on the far left sided calibration point. If necessary their eyes were oriented to the left position by asking them to track a pen which was moved slowly from right to left.
The instructions given were as follows. "Please look at the fixation cross when it appears in the centre of the screen. A small target square will then appear to either the left, or right,

496 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
Table 1. Patient details
Background details Assessment of neglect
Lesion Line Word Text Clock Patient location Age TPO Hemianopia Cancellation bisect reading reading drawing Chimaerics
R.R. Fronto-temporo-parietal 60 41 No 0 3 0 0 Yes 9 T.S. Undoc. 59 29 No 1 0 0 0 No 0 H.T. Parietal, and post. frontal 61 42 Left 2 10 0 1 Yes 10
small R. occip. D.R. MCA 64 6 ? 30 60 1 30 Yes 10
Notes: Background Details:
Lesion location: all lesions were to the right hemisphere in the region of the parietal lobe; MCA = middle cerebral artery infarct, Undoc. = scan unavailable.
Age: in years. TPO: time post onset, in months. Hemianopia: R.R. visual fields assessed using an automatic perimetric field plotter (Henson CFA) and shown not to be hemianopic.
T.S. not hemianopic and H.T. left homonymous hemianopia (both diagnosed using a field plotting technique described by Walker et al. 1991 [40]. ? = D.R.'s visual fields not assessed (patient unavailable for field plotting). Assessment of Neglect:
Cancellation: number of targets omitted from left side (max = 40). Line bisection: percentage deviation to right of true centre (line length 20 cm). Word reading: left-sided errors or omissions made when reading, 4, 5 and 6 letter words (max = 15). Text reading: number of whole words omitted from left side of a passage of text (max = 90). Clock drawing: patient made left-sided omissions when drawing from memory (yes or no). Chimaeric faces: number of left sides not reported (max = 10).
side of the cross and you should move your eyes to the target as quickly as possible. Try to look at the small black dot in the centre of each target square. On some trials two targets may appear both to the left, and right, of fixation. When two targets appear you are free to choose either one of them to move your eyes towards".
A short practice block of 30 trials was given to ensure that the patients could understand and comply with the instructions.
They were not informed of the difference between the overlap and gap blocks of trials.
As it has been argued that neglect is not a unitary syndrome and that neglect patients do not form a heterogeneous group [20], combining the results from different 'neglect' patients may disguise subtle differences in the performance of each patient. The following results section discusses each patient's results separately as a series of single case studies. As the latencies in
30
23 25
26 26 26
17 28 31 32
23 28 28 32
19 22 26 26
19 22 23 22
6 16 18
15 21
Thresholds (db)
23 29
22 30
32 31
32 30
32 28 + 2 7
28 29 26 29
28 28 26
27 27
24
28 25
30 27 30
0 cor responds to 300 c d / m 2
Fig. 1. R.R.'s visual fields expressed as raw threshold values where 0 = 300 cd/m 2, 20 = 3 cd/m 2 and 30 = 0.3 cd/m 2. The plot shows that R.R.'s upper left and upper and lower right visual fields are normal. His lower left visual field shows an increase in thresholds
for eccentricities greater than 10 "~.

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 497
each of the target conditions are not matched across subjects for single case studies, separate analysis of variance com- parisons were used to compare the different conditions for each patient.
Results
To enable a visual examination of each eye movement the saccade records were automatically plotted onto the VDU screen by the analysis program. Any record on which central fixation had not been maintained, or on which a blink or head movement occurred prior to target onset, was not analysed further.
The saccade records for each trial were analysed using a semi-automatic program which detected the start of each saccade using a velocity criterion of 30 ' /sec . A moveable cursor was placed at the point where the algo- rithm detected saccade onset and the cursor could be re- positioned by the examiner if required. The latency of a saccade was taken as the time from the onset of the target to the point at which the start of the saccade was detected. Saccades with a latency of less than 100 msec were excluded as being anticipations and saccades with a lat- ency of over 400 msec were excluded as not being stimulus driven. Saccade duration was taken as being approxi- mately 30 msec and the eye position 40 msec after saccade onset was used to calculate the amplitude of each saccade. Saccade amplitude was computed in relation to the mean values obtained from each of the calibration routines performed. The direction of each saccade was noted and when in opposition to the target direction was not included in the final analysis. The numbers of trials where more than one saccade was made to reach the target location were noted.
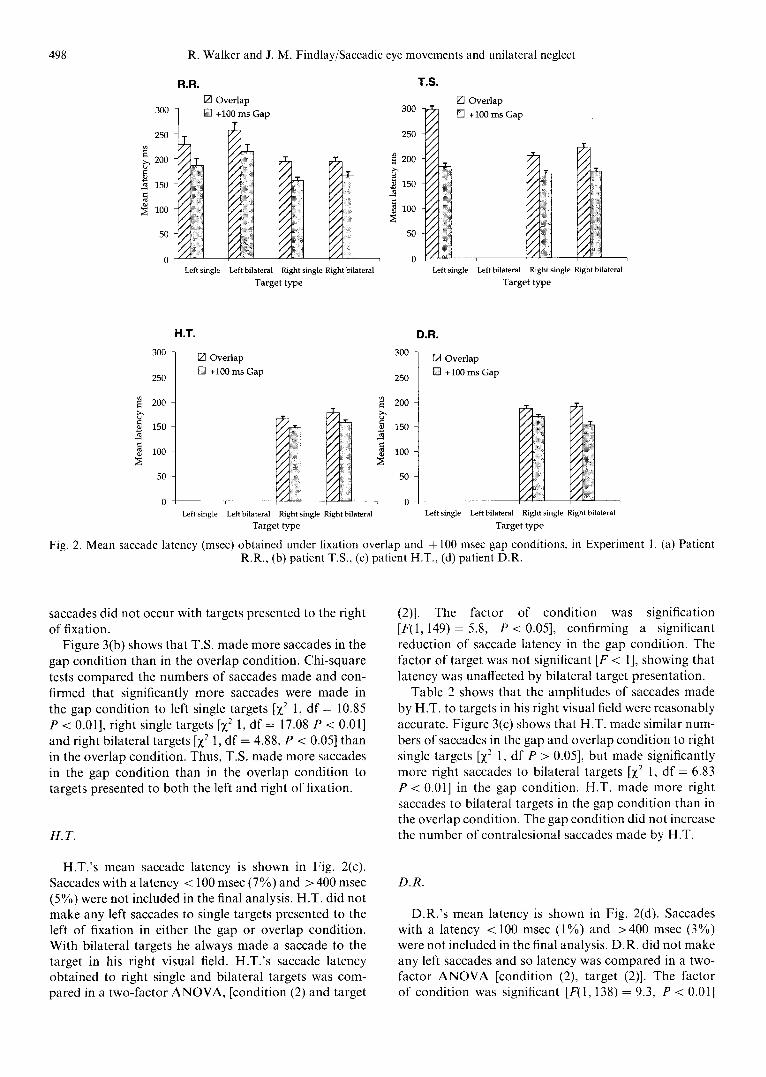
As there was no evidence of eccentricity effects on saccade latency and only small numbers of saccades made in some conditions the latencies obtained from the three target eccentricities were combined. There were no con- tralesional saccades made under some conditions for three of the patients. The latency of saccades made by the four patients in the overlap and + 100 msec gap condition are shown in Fig. 2(a-d) and the numbers of saccades made under each condition in Fig. 3(a d).
R.R
R.R. 's mean saccade latency is shown in Fig. 2(a). R.R. is of particular interest as he made left saccades with unilateral single and bilateral target presentation. Any saccades with a latency < 100 msec (2%) and > 400 msec (6%) were excluded from the final analysis. Saccade lat- ency was compared in a three-factor analysis of variance (ANOVA) [condition (gap/overlap), direction (left/right) and target (single/bilateral)]. The factor of condition was significant [F(1,194) = 17.6, P < 0.01] showing that sac- cade latency was significantly reduced in the gap
condition. The factor of direction was also significant [F(1,194) = 22.7, P < 0.01] showing that left saccades had longer latencies than those made to the right. The factor of target was not significant showing that latency was comparable for single and bilateral target pres- entation. There were no interaction effects.
Table 2 shows the mean amplitudes of saccades made by R.R. in the gap and overlap conditions. R.R. 's sac- cades made to targets presented to the right of fixation were accurate for targets at 4 and 8 , but tended to undershoot targets at 12. Saccade amplitude was unaffected by bilateral target presentation. In contrast R.R. 's left saccades had an amplitude of some 4~5 irres- pective of the eccentricity of the target. Saccades made to targets at 8 and 12 left of fixation tended to fall short of the target locations. On 49% of the trials with targets presented at 8 and 12: left of fixation more than one saccade was made to reach the target location. R.R. made multiple saccades to ipsilesional targets on only 2% of trials. The amplitude of left saccades was not improved in the gap condition.
Figure 3(a) shows that R.R. made similar numbers of contralesional saccades to single targets under gap and overlap conditions. However, there was an increase in the number of left saccades made with bilateral targets in the gap condition (producing a corresponding decrease in the numbers of right saccades made to bilateral targets). When questioned, R.R. stated that he was aware of the presence of both the left and right bilateral targets.
T.S.
The mean latency of T.S.'s saccades are shown in Fig. 2(b). Saccades with a latency < 100 msec (2%) and >400 msec (3%) were not included in the final analysis. T.S. did not make any left saccades with bilateral targets and so saccade latency to right single, right bilateral and left single targets were compared in a two-factor ANOVA, [condition (2) and target (3)]. The factor of condition was significant [F(I, 214) = 99.7, P < 0.1] confirming a significant gap facilitation effect. The factor of target was also found to be significant [F(2,214) = 23.8, P < 0.01]. Post hoc analysis (Newman-Keuls) showed that saccade latency was significantly greater to left single targets than when made to right single targets (P < 0.01). The latency of saccades made to single targets was comparable to that made to bilateral targets.
Table 2 shows that the amplitudes of saccades made by T.S. to targets in his right visual field were reasonably accurate. The amplitudes of saccades made to targets presented in his left visual field were small (approx. 2 ) and did not correspond to the eccentricity of the target. The amplitude ofcontralesional saccades did not improve in the gap condition. On 47% of trials with targets pre- sented left of fixation T.S. made more than one saccade to locate the target location. This pattern of multiple
498 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
300
250
200
150
o~ lOO
50
R,R,
[ ] Overlap
[ ] + 1 0 0 ms Gap
L
Z;') / /
I ~ ??:i / # ......... ,. #
" / ~ x / " < t " /
, , , " " ......, Z i-; ,,,'" , ,
Left single Left bilateral Right single Right bilateral Target type
T,S,
300 . x . f / / /
250 " j # / . . - j
200 f . ,
150
100
50
- - n 0
[ ] Overlap
[ ] + 1 0 0 ms Gap
, - j / # / / / / / / g / ¢- /
# /
Left single
/ / 1 i t- / # / 1 I
f pl;;~, .i 1.-141
Left bilateral Right single Right bilateral Target type
>,
H . T .
300 [ ] Overlap
[ ] + 1 0 0 ms Gap 250
200
150
100
5O
0
3 0 0
2 5 0
2 0 0
150
~ 1oo
50
0
D,R,
[ ] Overlap
[ ] + 1 0 0 ms Gap
Left single Left bilateral Right single Right bilateral Left single Left bilateral Right single Right bilateral Target type Target type
Fig. 2. Mean saccade latency (msec) obtained under fixation overlap and + 100 msec gap conditions, in Experiment I. (a) Patient R.R., (b) patient T.S., (c) patient H.T., (d) patient D.R.
saccades did not occur with targets presented to the right of fixation.
Figure 3(b) shows that T.S. made more saccades in the gap condition than in the overlap condition. Chi-square tests compared the numbers of saccades made and con- firmed that significantly more saccades were made in the gap condition to left single targets [;(2 1, df = 10.85 P < 0.01], right single targets [Z 2 1, df = 17.08 P < 0.01] and right bilateral targets [9( 2 1, df = 4.88, P < 0.05] than in the overlap condition. Thus, T.S. made more saccades in the gap condition than in the overlap condition to targets presented to both the left and right of fixation.
H.T.
H.T.'s mean saccade latency is shown in Fig. 2(c). Saccades with a latency < 100 msec (7%) and > 400 msec (5%) were not included in the final analysis. H.T. did not make any left saccades to single targets presented to the left o f fixation in either the gap or overlap condition. With bilateral targets he always made a saccade to the target in his right visual field. H.T.'s saccade latency obtained to right single and bilateral targets was com- pared in a two-factor A N O V A , [condition (2) and target
(2)]. The factor of condition was signification [F(1,149) = 5.8, P < 0.05], confirming a significant reduction of saccade latency in the gap condition. The factor of target was not significant IF < 1], showing that latency was unaffected by bilateral target presentation.
Table 2 shows that the amplitudes of saccades made by H.T. to targets in his right visual field were reasonably accurate. Figure 3(c) shows that H.T. made similar num- bers of saccades in the gap and overlap condition to right single targets [;(2 1, df P > 0.05], but made significantly more right saccades to bilateral targets [Z 2 1, df = 6.83 P < 0.01] in the gap condition. H.T. made more right saccades to bilateral targets in the gap condition than in the overlap condition. The gap condition did not increase the number of contralesional saccades made by H.T.
D.R.
D.R.'s mean latency is shown in Fig. 2(d). Saccades with a latency < 100 msec (1%) and >400 msec (3%) were not included in the final analysis. D.R. did not make any left saccades and so latency was compared in a two- factor A N O V A [condition (2), target (2)]. The factor of condition was significant [F(I, 138) = 9.3, P < 0.01]

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 499
R.Ro
60 [] Over lap
50 [] +100 ms Gap
"O 40
"83o
o
0 , Left single Left bilateral
60
50
40
"8 30
20
10
, , D
Right single Right bilateral
Target type
T . S .
[] Overlap [] +100 ms Gap
Left single Left bilateral Right single Right bilateral Target type
Z
60
50
40
30
20
10
0
H.T.
[] Overlap [] +100 ms Gap
60
50
40
30
zo Z
10
, 0
D.R.
[] Overlap [] +100 ms Gap
q i
Leftsingle Left bilateral Right single Right bilateral Leftsingle LeRbilateral Right single Right bilateral Target type Target type
Fig. 3. Number of saccades made left and right of fixation with single and bilateral target presentation in the overlap and + 100 msec gap condition.
Table 2. Mean saccade amplitude (and S.D.) in degrees of visual angle obtained in Experiment 1
Target type and eccentricity (degrees):
Left single Right single Left bilateral Right bilateral
- 1 2 - 8 - 4 +12 +9 + 4 - 1 2 - 8 - 4 +12 +8 + 4
(a) Overlap condition
R.R. - 5 . 3 - 4 . 2 (2.9) (I .9)
T.S. - 4 . 2 - 2 . 7 (2.4) (1.4)
H.T. none none
D.R. none none
- 3 . 8 (l.S)
- 2 . 7 (1.7)
n o n e
n o n e
(b) + 100 msec gap condition
R.R. - 5 . 4 - 5 . 5 (1.6) (2.7)
T.S. - 3 . 0 --2.0 (1.0) (0.8)
H.T. none none
D.R. none none
- 3 . 0 (1.0)
- 1 . 6 (0.5)
none
none
+9.7 (4.0)
+8.2 (3.9)
+8.8 (5.3)
+11.2 (2.2)
+8.5 (2.8)
+7.6 (2.3)
+8.6 (1.8)
+8.1 (4.1)
+7.7 (2.4)
+6.2 (2.3)
+8.6 (4.1)
+8.3 (2.1)
+5.2 (3.3)
+4.3 (1.2)
+6.6 (0.9)
+7.4 (3.0)
+4.3 (1.5)
+3.8 (1.8)
+4.6 (2.2)
+5.1 (1.4)
+3.6 (2.1)
+2.5 (1.5)
+4.3 (1.1)
+5.3 (1.8)
- 4 . 6 (3.6)
none
none
none
- 7 . 0 (2.2)
none
none
none
none
none
none
none
--4.8 (1.0)
none
none
none
--3.0 (1.0)
none
none
none
- 2 . 3 (1.2)
n o n e
none
none
+9.3 (3.0)
+8.7 (4)
+8.2 (4.6)
+10.8 (3.5)
+8.2 (2.9)
+6.8 (1.8)
+8.1 (2.2)
+6.9 (3.9)
+6.4 (2.7)
+6.7 (1.8)
+7.3 (3.1)
+6.8 (3.5)
+3.5 (1.3)
+4.3 (1.4)
+7.0 (1.4)
+7.0 (4.1)
+4.6 (1.9)
+2.8 (0.8)
+5.0 (2.3)
+4.7 (1.3)
+3.1 (1.1)
+2.7 (0.8)
+4.4 (0.9)
+5.2 (1.9)

500 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
showing a significant reduction in saccade latency in the gap condition. The factor of target was not significant [F < 1] showing that latency was not increased with bilat- eral target presentation.
Table 2 shows that the amplitudes of saccades made by D.R. to targets in his right visual field were reasonably accurate. Figure 3(d) shows that D.R. made similar num- bers of right saccades under gap and overlap conditions to right single [X 2 1, df, P > 0.05] and bilateral targets [Z 2 1, dr, P > 0.05]. The gap condition has not increased the numbers of saccades made to targets presented in the left or right visual fields. Thus, there was no improvement in D.R. 's ability to make left saccades in the gap condition.
Discussion
In Experiment 1 the saccadic eye movements made by four patients with unilateral neglect were recorded under fixation gap and overlap conditions. Saccades were made to a single target presented in the left or right visual field or to one target when two were presented bilaterally and simultaneously in both visual fields. The prior offset of fixation in the gap condition did not improve the patients' ability to make contralesional saccades. Two of the pat- ients (H.T. and D.R.) failed to make any contralesional saccades in either the gap or overlap condition and R.R. made similar numbers of contralesional saccades under both conditions. Only T.S. made significantly more con- tralesional saccades in the gap condition than in the over- lap condition, replicating the result found by Walker et al. [40]. However, T.S. also made more ipsilesional saccades in the gap condition than in the overlap condition. A similar result emerged for patient H.T., who also made more ipsilesional saccades in the gap condition than in the overlap condition.
An examination of saccade amplitude showed that the contralesional saccades made by R.R. and T.S. were hypometric and landed short of the exact target location. Both R.R. and T.S. showed a tendency to make multiple saccades to locate stimuli presented in their con- tralesional visual fields. The amplitude of contralesional saccades did not improve in the gap condition. None of the patients made multiple saccades to locate stimuli presented in their ipsilesional visual fields.
Although the prior offset of fixation in the gap con- dition did not increase the numbers of contralesional saccades made it did produce a consistent gap effect. The mean latency of saccades made in the ipsilesional direction was significantly reduced in the gap condition compared to that in the overlap condition for all patients. Thus it appears that the latency reduction characteristic of the gap effect occurs in neglect patients. The magnitude of the latency reduction is comparable to that found with normal subjects [8, 28] with the sole exception of the contralesional saccades made by patient T.S., where a substantially larger gap decrease was found.
The effect of presenting two targets simultaneously in
both visual fields was also examined in Experiment 1. With bilateral targets three patients always made sac- cades to the target presented in their ipsilesional visual field. These patients were also unaware that a target had also appeared simultaneously in their contralesional vis- ual field (visual extinction). In contrast, R.R. 's first sac- cades were made to the bilateral target presented in his contralesional hemifield on a number of trials. The num- ber of saccades made by R.R. to the contralesional bilat- eral target increased in the gap condition. R.R. did not show extinction of the contralesional bilateral target and was aware of the presence of both bilateral targets when questioned.
A second important finding was that saccade latency was not increased with bilateral target presentation. This absence of a latency increase contrasts with the reliable increase in latency shown by normal subjects under simi- lar conditions [3, 16, 41]. Two of the neglect patients (R.R. and T.S.) are not hemianopic and so the failure to show the normal increase in latency observed with bilat- eral targets cannot just be attributed to the lack of a contralesional sensory input. An absence of a latency increase with bilateral target presentation was also noted in an earlier study of eye movements in a non-hemianopic neglect patient [40].
Experiment 2. The global effect
Experiment 2 examines eye movements when two tar- gets are presented in the same hemifield at different eccen- tricities along the horizontal axis. With normal subjects, saccades to target pairs tend to be directed towards the 'centre of gravity' of the configuration. The landing pos- ition of saccades made by normal subjects to a single target tends to undershoot the target location, but when two targets are presented in the same hemifield, the sac- cade usually overshoots the nearer target. Findlay [2] has reported an increase in saccade amplitude of 20M0% with pairs of targets (global effect) compared to the amplitude obtained to single targets. It is of interest to study the global effect in patients with unilateral neglect as certain models have suggested that one factor in neg- lect is an attentional bias in the ipsilesional direction. Kinsbourne's model of neglect [13] is based on the idea that each hemisphere of the brain contains an opponent processor which is involved in orienting attention in the contralateral direction. Neglect is thought to arise due to damage to one hemisphere that leaves the opponent processor in the other hemisphere disinhibited and results in a bias of attentional orienting in the ipsilesional direc- tion. Indeed covert attentional orienting studies have shown that neglect patients have a bias of attentional orienting to stimuli located further ipsilesionally from fixation [15]. If a hyper-attentional orienting bias is a component of neglect then the amplitude of saccades made by patients to global effect targets might be expected to be different from those of normal subjects. A

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 501
generalized rightwards attentional bias could result in an alteration of the centre of gravity weightings so that tar- gets presented in the patient's ipsilesional hemifield would be expected to show a greater overshoot than is found in normal subjects. Conversely targets presented in the contralesional hemifield would be expected to show a reduced overshoot. Attentional biases have been shown to influence the global effect [4].
As only R.R. and T.S. made contralesional saccades in Experiment 1 they acted as subjects in the following Experiment. The amplitudes of saccades made with pairs of targets presented simultaneously in one hemifield were compared to the amplitude of saccades made to single targets presented in the same hemifield. The effects of bilateral target presentation on saccade amplitude and latency was also examined.
Method
Subjects. R.R. and T.S. acted as the subjects in this experiment. A 52-year-old male, C.R., with no known neurological impairments acted as a normal control.
Procedure. The subjects viewed the V D U screen from a distance of 57 cm. The stimuli used were the same square targets as used in Experiment 1, but in this experi- ment a similar square was also presented as a fixation stimulus. At the viewing distance of 57 cm the sides of the stimuli subtended a visual angle of 0.6'. At the start of each trial a fixation square appeared in the centre of the screen. Targets were presented on a horizontal axis level with fixation at eccentricities of 2 ' and 4 '~. A block of trials included single targets presented at one of the eccentricities used, pairs of targets presented sim- ultaneously at 2 '~ and 4 in the same hemifield and bilat- eral targets presented at equal and opposite locations (as in Experiment 1). In order to maximize the numbers of saccades the patients would make to left targets there were two blocks of trials, one in which single and global effect targets were all presented to the left of fixation (contralesional block) and one in which they were pre- sented to the right of fixation (ipsilesional block). Each block of trials contained 20 single targets, 20 global effect targets and 20 bilateral targets for each eccentricity location, giving a total of 100 trials in each. The order of presentation of single and global effect targets was randomized within each block of trials.
The d(/Jbrent stimulus types used in each block of trials" were:
Single targets: a single target appeared unilaterally in one hemifield at eccentricities of either 2 or 4 ' from fixation.
Global effect targets: two targets appeared sim- ultaneously 2 ~ and 4 from fixation in the same hemifield.
Bilateral targets: two targets appeared simultaneously to the left and right of fixation at equal and opposite
eccentricity locations (at - -2: and + 2 , or - 4 '~ and + 4 ' locations).
The timin 9 sequence used was as Jollows: The fixation square appeared at the centre of the VDU for 1000 msec and then went off. The onset of the saccade target was simultaneous with the offset of fixation (0 gap). Targets were presented for 1000 msec. An inter-trial delay of 1000 msec occurred between each trial. The timing sequence used gave the appearance of a rapid tracking experiment and subjects were instructed to move their eyes from fixation to locate the centre of the target square on each trial. To ensure that the subject maintained fix- ation prior to target onset the analogue-digital con- version started with the onset of the fixation point and continued until target offset. The instructions given to subjects were the same as those used in Experiment 1.
As Experiment 2 required accurate saccade amplitude measures care was taken to restrict the patients' head movements and reduce any inaccuracies during the rec- ording procedure. The calibration routine (as for Experi- ment 1) was performed twice at the start and end of each block of trials. To reduce any inaccuracies that may result from any small head movements during the experiment the average of these calibrations was used to calculate saccade amplitude. The analysis of each record was per- formed using the same method as that described for Experiment 1.
Results
Trials on which central fixation was not maintained prior to target onset were not included in the analysis. The amplitude of saccades made to single targets, global effect targets and bilateral targets is shown in Fig. 4. The global effect parameter was calculated as a percentage in the following way [1]: (global amp near single amp/near a m p - - f a r amplitude). The ipsilesional and contralesional global effect results from the two patients will be dis- cussed separately below.
Control subject C.R.
When single targets were presented at 2 ' and 4 ' eccen- tricities in either the left or right visual field C.R. 's saccade amplitude showed a small undershoot of the target location. However, when target pairs appeared at 2 ' and 4 locations C.R. 's saccades showed an overshoot of the near target position. The global effect parameter with pairs of targets in the left visual field was 25% and with pairs of targets in the right visual field was 26%. Mean saccade latency was unaffected by the presentation of two targets in the same hemifield, but was increased when a target appeared in both hemifields. C.R. used a deliberate strategy of always making saccades to the bilateral target

502 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
(a) CO~TRALESIONAL TARGETS . 'S lL~S~ONA~ TARGETS
Normal C.R. + ~ 2 ° Single
+ ~ Z ] ,o~ing,e + + + ~ U] 2O,OGlo~.lE.~,
D 4-] +
r~ + ~ 4o Bila feral
(b) CONTRALESIONAL TARGETS IPSILESIONAL TARGETS
Patient R.R. + ~ 2 ° Single
+ [ ~ 4° Single
+
[ - - 1~ + + D ~ - ~ D 2o ,OG,o~lE.eo,
[3 ~ + [ 3 + ~ 2o ...,eral
(c) CONTRALESIONAL TARGETS 1PSILESIONAL TARGETS
Patient T.S. + [ ~ 2 ° Single
+ ~ 40 Sing le
I3. + 7"1 ~ - ~ +
+ D ~fi] ~O,OGlo~,~,oo,
E] fi~ + F-1 + ~ ~O.lla,e,o,
[Z] + qE] 4o ~i~o~l
Fig. 4. Mean saccades amplitude (with S.D.) in degrees obtained in Experiment 2. Saccades were made to: single targets presented 2 ° and 4 ° left and right of fixation, to targets presented bilaterally at 2 ° or 4 ° in opposite hemifields and with pairs of targets presented
at 2 ° and 4 ° locations in the same hemifield. (a) Normal subject C.R. (b) neglect patient R.R. (c) neglect patient T.S.
in the r ight visual field and la tency was increased by 20 27 msec c o m p a r e d to tha t ob ta ined to single targets.
R.R.
Ipsilesional global effect. R.R. ' s saccades to single tar- gets presented 2 ° f rom fixation had a mean ampl i tude o f 2.28 ° indica t ing a small overshoo t and saccades made to
a target at 4 ° had a mean ampl i tude o f 4.05 °. R .R . ' s saccades with target pairs presented at 2 ° and 4 ~' showed an increase in ampl i tude c o m p a r e d to that ob ta ined to single targets with a mean o f 2.9 °. R.R. showed an ips- i lesional g lobal effect o f 35%, which is slightly greater than that shown by the cont ro l subject C.R. , but com- pa rab le with previous figures [2].
R .R . ' s saccade ampl i tude with bi la tera l targets was s imilar to tha t ob ta ined to uni la tera l single targets. On

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 503
two trials with bilateral targets R.R. 's first saccade was directed to the left bilateral target and on a further four trials he made saccades to the left bilateral target after making an initial saccade to the right target. A one-way ANOVA with five levels of targets showed that R.R. 's saccade latency was unaffected by the type of target pre- sented [ F < 1]. Saccade latency was not significantly increased with bilateral target presentation.
Contralesionalglobal efJ~'ct. R.R. 's left saccades tended to undershoot the position of single targets presented in his contralesional hemifield. R.R. 's mean saccade ampli- tude to single targets presented at 2 ° and 4 ' in his left hemifield was of 1.79 ~ and 2.T', respectively. With global effect targets R.R. 's mean saccade amplitude was increased to a mean of 2.05 ~, showing a contralesional global effect of 34%.
With bilateral targets R.R. made 12 saccades to the target in his left hemifield and 10 to the target in his right hemifield. There were a further seven trials where he made a saccade to the right target and then to the left targets on bilateral trials. A one-way ANOVA compared the latency of saccades made to the five target types and showed that R.R. 's saccade latency was unaffected by target type [F < 1]. Thus R.R. 's saccade latency was not increased with bilateral target presentation.
T.S.
Ipsilesional global efJect. T.S.'s saccade amplitude to single targets showed a small overshoot. His mean sac- cade amplitude to targets presented at 2 ~' and 4 c was 2.4 '-~ and 4.04", respectively. His mean saccade amplitude with pairs of targets presented at 2 "~ and 4 ° was 3.6 ~' indicating a large overshoot of the near target location. T.S. showed an ipsilesional global effect of 73%, which is much greater than that observed with normal subjects.
When presented with bilateral saccade targets T.S. made a saccade to the target in his right hemifield on every trial. Mean saccade amplitude to targets at 2 ~ and 4 c with bilateral presentation were 2.07 ° and 3.7 '~, respec- tively. Saccade latency made to bilateral targets was not found to be increased in comparison to that observed with single targets. A one-way ANOVA confirmed that T.S.'s saccade latency was comparable for the five target types [F(1,54) = 2.34, P > 0.05].
Contralesional global effect. T.S.'s saccades made to single targets presented in his left hemifield tended to undershoot the target location. Mean saccade amplitude to targets presented at 2:' and 4 ~ eccentricities was 1.6 '~ and 2.5 "~, respectively. Saccade amplitude made with tar- get pairs at 2'- and 4 c~ eccentricities was some 1.7 ° indi- cating a small contralesional global effect of only 11%.
With bilateral targets T.S. always made saccades to the target in his right ipsilesional hemifield. As three-quarters of saccade targets appeared in the contralesional hemi- field T.S.'s tendency to make an ipsilesional saccade with bilateral targets appeared to be strong. A one-way
ANOVA showed that T.S.'s saccade latency was com- parable for single, bilateral and global effect targets [ F < 1].
Discussion
The presentation of two targets in one hemifield of normal subjects is known to influence the landing pos- ition of saccades with amplitudes increasing in com- parison to that shown to a single target at the same location [2]. In this experiment an increase in amplitude of some 25% was shown in the saccades made by a normal control subject when pairs of targets appeared in one hemifield. The amplitudes of saccades made by the two neglect patients were differentially affected in the global effect situation. R.R. showed a similar magnitude global effect overshoot of 35% when pairs of targets were presented in either his ipsilesional or contralesional visual fields. This is slightly greater than the global effect shown by the control subject C.R., but is within the range shown by normal subjects [1, 2]. In contrast T.S. showed a large global effect overshoot of 73% with pairs of targets pre- sented in the ipsilesional visual field and a much smaller global effect of 11% with targets pairs in his con- tralesional visual field. T.S.'s saccades landed close to the far target with target pairs in the ipsilesional visual field, but fell short of the near target location with pairs of targets in his contralesional visual field.
Experiment 2 also confirmed the finding of Experiment 1 that the latency of saccades made by patients with neglect is not increased with bilateral target presentation. Mean latency of saccades made to a single target pre- sented in the patients' ipsilesional visual field was com- parable to the mean latency obtained when two targets were presented simultaneously in both visual fields. The amplitude of saccades made in the ipsilesional direction was unaffected by the presentation of bilateral targets. When targets were presented bilaterally R.R. did not show visual extinction and his first saccades were often made to the target in his left (contralesional) visual field in preference to that in his right visual field. In contrast, T.S. showed extinction of the contralesional target and always made saccades to the target in his ipsilesional visual field.
General discussion
The present study has revealed a complex but coherent picture of fractionation of eye movement programming in patients with unilateral neglect. In Experiment 1 the number, latency and amplitude of saccades made were examined under fixation gap and overlap conditions. The main findings can be summarized as follows:
(1) For three patients the prior offset of fixation in the + 100 msec gap condition did not increase the num-

504 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
bers of saccades made into their contralesional visual field;
(2) Prior fixation offset did produce a consistent reduction in saccade latency (gap effect);
(3) Two patients made some contralesional saccades which were of small amplitude and long latency;
(4) Saccade latency was not significantly increased when two targets were presented bilaterally and sim- ultaneously in both hemifields. This performance is in contrast to the latency increase shown by normal subjects in such situations [41]. In Experiment 2 the amplitude of saccades were examined in a global effect task where two targets were presented in the same hemifield;
(5) One patient, T.S., showed a greater than normal glo- bal effect in his ipsilesional field and a smaller global effect in his contralesional field. In contrast, R.R., a patient with apparent object-based neglect [42], showed a normal global effect in both his ipsilesional and contralesional visual fields. The implication of these results will now be discussed
Neglect patients are known to have deficits in pro- ducing saccades to locate stimuli presented in their con- tralesional hemifields [9, 11, 12, 18, 36, 40]. The present study examined the possibility that the failure to initiate contralesional saccades may reflect a deficit of an atten- tional disengagement mechanism [26]. According to this model the removal of the central fixation point prior to the onset of the saccade target should automatically disengage visual attention [5, 8] and improve the patients' ability to make contralesional saccades. The present study revealed no selective improvement in the numbers of contralesional saccades made in a + 100 msec gap condition. Two patients, H.T. and D.R., failed to make contralesional saccades in either the gap or overlap con- dition. R.R. made equal numbers of contralesional sac- cades in the gap and overlap conditions. Only T.S. made more contralesional saccades in the gap condition than in the overlap condition and he also made more ipsilesional saccades. Furthermore, the amplitude of the con- tralesional saccades made by R.R. and T.S. were hypo- metric and landed short of the target location. Both patients showed a tendency to make more than one sac- cade to locate targets in their contralesional visual field. Contralesional saccade amplitude was not improved in the + 100 msec gap condition.
The failure to increase the numbers of contralesional saccades made in the gap condition is incompatible with the view that the deficit is produced by an inability to disengage attention from fixation. This finding contrasts with a previous report where a dramatic increase in the numbers of contralesional saccades was observed under a similar gap condition by a neglect patient, B.Q. [40]. We have discussed in the Introduction alternative expla- nations of the gap effect to that of attentional dis- engagement. The alternatives were warning signal events [32, 33] and ocular disengagement [22, 39]. Can these
alternative models account for the results from neglect patients? A notable feature of the results from the patient T.S. is that the gap condition resulted in an increase in the numbers of both contralesional and also ipsilesional saccades. Similarly, in the earlier study B.Q. was also shown to make more contralesional and ipsilesional sac- cades under the gap than overlap condition. We believe that the increase in the number of saccades made in the gap condition by patients such as T.S. and B.Q. can be explained in part in terms of an increase in non-spatial attention caused by warning signal effects of fixation offset in the gap condition.
The finding that some patients with neglect can make use of a warning signal event is consistent with models of neglect that have stressed that an important factor in poor recovery from the condition is a deficit of a non- spatial attentional arousal system [30]. Patients with dam- age to their arousal/vigilance system may fail to learn to compensate for their deficits and do not use cues to orient their attention to the contralesional side of space. Patients who show some degree of recovery from neglect have most likely learned some strategies to enable a degree of compensation for their deficits [17]. It appears that pa- tients such as T.S. with mild neglect may be able to use fixation offset as a warning signal which alerts them to make a contralesional saccade. Although fixation offset can increase the numbers of saccades made by some pat- ients, it was not found to have improved the hypometric amplitude of these saccades. The contralesional saccades were hypometric and typically landed short of the target locations. T.S. and R.R. made multiple left saccades to locate contralesional target locations supporting the view that a search strategy may have been used.
The removal of the fixation point in the gap condition produced a significant reduction in saccade latency com- pared to that obtained in the overlap condition. This 'gap effect' was consistently shown by all four neglect patients. The reduction in saccade latency in the gap condition is consistent with both the warning signal and ocular disengagement models of the gap effect. Reuter-Lorenz et al. [29] demonstrated that part of the latency reduction in gap conditions can be attributed to a warning signal effect. Our results have shown that neglect patients may also be able to make use of such warning signal events which can enable partial pre-programming of saccade parameters to take place prior to the onset of the saccade target. Reuter-Lorenz et al. also showed that the gap effect was not abolished when such warning signals were controlled. The remaining gap effect latency reduction in this situation was attributed to an ocular fixation mech- anism [22, 23]. The latency reduction shown by neglect patients in gap conditions is consistent with both the warning signal and ocular disengagement models of the gap effect.
It should be noted that the latency of ipsilesional sac- cades made by the patients in the present study was within the expected range for normal human subjects [7]. This is in contrast to some reports of a bilateral saccade deficit

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 505
following unilateral posterior parietal cortex lesions. Pierrot-Deseilligny et al. [25] examined the latency of saccades made by patients with right posterior parietal lobe damage and found that the latency of saccades made in both the ipsilesional and contralesional direction was increased compared to normal subjects. This bilateral increase in saccade latency was interpreted as showing that the right posterior parietal cortex is involved in trig- gering visually guided saccades made to the left and right of fixation. In the present study the latency of ipsilesional saccades made by the parietal patients was not increased and was comparable to those obtained from younger normal subjects on a similar task [41] One factor that may explain the difference between the results of these two studies could be the exact location of the lesions involved. However, animal lesion studies [17] have also failed to demonstrate an ipsilesional increase in saccade latency following right inferior parietal lesions (including area 7) in monkeys. A degree of caution is therefore required in attributing the role of the parietal cortex to triggering saccades in both directions.
The present study has confirmed that the latency of saccades made by neglect patients was not influenced by bilateral target presentation [40]. This is in marked contrast to the situation observed with normal subjects who show an increase in latency of some 20 40 msec with bilateral target presentation [3, 16, 41]. Furthermore, the lack of a bilateral target effect and the increased latency shown for contralesional saccades cannot simply be attri- buted to the presence of a hemianopia, or to reduced sensitivity of the contralesional visual field. The visual field plot for R.R. (Fig. 1) shows that his upper left visual field is intact and increased thresholds are apparent only for the lower left quadrant at eccentricities greater than 10'. R.R. did not show a bilateral target effect and this cannot be attributed to a reduced level of sensitivity in the contralesional visual field.
To explain our findings we propose that unilateral parietal damage may result in an alteration of the balance in the orienting system, possibly at the collicular level. Saccades produced quasi-reflexively following an abrupt stimulus onset are thought to involve the superior col- liculus [37]. The parietal lobe does not have a direct projection to the brainstem saccade generators, but it does have both direct and indirect projects (via the sub- stantia nigra) to the colliculus. The suggestion that the left and right superior colliculus are normally in a state of dynamic equilibrium resulting partly from mutual crossed inhibitory inputs has been made previously by Sprague [38]. Sprague showed that cats failed to respond to stimuli presented in their contralesional visual fields after a unilateral occipital lesion, However, the sub- sequent removal of the contralateral superior colliculus enabled the cats to respond to stimuli presented in the previously hemianopic neglected field. The explanation of the Sprague effect is that the initial cortical lesion results in a loss of a tonic excitatory input to the ipsilateral colliculus, which in consequence is subject to deep inhi-
bition from the contralateral colliculus. Subsequent work by Sprague's group [43, 44] has established that the crossed inhibition does not arise from the contralateral colliculus but is mediated by the substantia nigra.
We propose a speculative account of our findings based on a similar model to that used to account for the Sprague effect. It is plausible to suggest that unilateral parietal lobe damage could result in a similar corticotectal imbal- ance to that suggested by Sprague. Damage to the right parietal lobe may result in under-activity of the ipsi- lesional (right) superior colliculus which is involved in programming saccades in the contralesional direction. One consequence of the ipsilesional colliculus being over- inhibited would, therefore, be a deficit in producing con- tralesional saccades. Furthermore, if the ipsilesional col- liculus remains permanently over-inhibited following the parietal lesion then the contralesional colliculus may not be subject to normal crossed inhibitory inputs. Thus, when targets are presented bilaterally and simultaneously in both visual fields the system would be biased to pro- ducing a saccade in the ipsilesional direction. The absence of the normal latency increase following bilateral target presentation could be accounted for by the lack of crossed inhibition resulting from the constant depression of the ipsilesional colliculus. In this imbalanced situation the latency of saccades made in the ipsilesional direction would be comparable following either unilateral or bilat- eral target presentation.
A further finding which emerged from the presentation of bilateral targets appears to be relevant to the issue of the relationship between visual extinction and visual neglect. One neglect patient, R.R., did not show visual extinction with bilateral targets and could report both bilateral targets in these situations. This performance is in contrast to that of T.S., who was unaware of the contralesional bilateral targets. We believe that this find- ing is consistent with the view that neglect is not a unitary syndrome [20] and that it can be fractionated into differ- ent forms. R.R. shows neglect for the contralesional side of his own drawings and with chimaeric stimuli [42], but not on spatial tasks such as crossing out. In contrast, T.S, shows mild neglect on spatial tasks such as crossing out but not for drawing or with chimaeric stimuli. It is plaus- ible that visual extinction may be fractionated into spatial and object-based forms as has been suggested for visual neglect. So, a patient like T.S. with spatial neglect shows extinction with the spatially separated bilateral targets, while R.R. who has object-based neglect does not show extinction with the spatially separate bilateral targets.
Experiment 2 examined the effects of presenting two targets simultaneously in the s a m e hemifield on the ampli- tude of saccades made by two neglect patients. For nor- mal subjects, these conditions result in the global effect [2], whereby saccades overshoot the nearer target. We anticipated that in neglect patients a rightward atten- tional bias might modify the global effect, resulting in a larger overshoot in the right visual field and a smaller one in the left. The results from T.S. were consistent

506 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
with this prediction. However the patient R.R. showed a normal global effect overshoot with pairs of targets in both his ipsilesional and contralesional visual fields. T.S.'s performance on the global effect task is consistent with the view that some neglect patients have a spatial bias to stimuli located further in the ipsilesional direction [15, 16]. The difference between the performance of the two patients on this task could be due to the different nature of the patients' deficits. R.R. shows evidence of object-based neglect, while T.S.'s deficits appear to be spatially based.
The performance of T.S. is consistent with what might be expected in patients with mild neglect. T.S. made small numbers of hypometric contralesional saccades with uni- lateral targets and extinguished the contralesional bilat- eral targets. T.S. also showed a large ipsilesional global effect indicating an ipsilesional attentional bias. The per- formance of R.R. was, however, different and cannot be explained in terms of a mild form of neglect. R.R. made contralesional saccades to both single and bilateral tar- gets and showed a normal size ipsilesional and con- tralesional global effect. We have suggested that R.R. remains unaffected by bilateral target presentation because his deficits are object-based [42] while T.S. shows extinction as his deficits are spatial in nature. However, other variables that could account for the differences in performance should also be considered. It is worth noting that R.R. and T.S. were of similar age and both had made some degree of recovery in the years following their stroke. Neither patient had been included in any formal rehabilitation programmes and both had taken part in a similar number of eye movement recording sessions. Both patients were right handed and neither one had a con- tralesional hemianopia. The main difference between these two patients appears to be the form of neglect they exhibit. Although T.S. made almost no errors on any of these tests used to assess neglect he did appear to use a strategy when performing letter cancellation. T.S. was observed to work slowly from the right side of the page to the left side until all of the targets had been crossed. This suggests that he has residual spatial neglect but is able to compensate for it by using a compensatory strat- egy. T.S. did not show evidence of neglect when drawing or with chimaeric-faces. R.R. 's neglect has been the sub- ject of a separate detailed study [42] and he shows impair- ment when drawing and with chimaeric-faces indicating object-based neglect, but he shows only mild spatial neglect. Unfortunately, it is not possible to make com- parisons about underlying pathology differences between R.R. and T.S. as the exact location of T.S.'s lesion is undocumented.
In summary, the present study has demonstrated a normal latency reduction (gap effect) in saccades made by patients with visual neglect. However, the numbers of contralesional saccades made was not increased in the gap condition. This result argues against the view that a deficit of an attentional disengagement mechanism is the cause of the patient 's failure to initiate normal con-
tralesional saccades. Although some patients with neglect appear to be able to use fixation offset as a warning signal event that enables them to make contralesional saccades, these saccades are of small amplitude which implies the use of a search strategy. In contrast to the situation with normal subjects the latency of saccades was not increased with bilateral target presentation. We propose that a consequence of the unilateral parietal damage is an imbal- ance in activity within the saccadic system. This imbal- ance results in the ipsilesional colliculus being over- inhibited, which results in deficits in making con- tralesional saccades and a bias in making saccades in the ipsilesional direction. This imbalance could also account for the lack of a bilateral target effect on saccade latency. One patient with object-based neglect did not show evi- dence of visual extinction with bilateral targets that were spatially separated. This patient also showed a normal ipsilesional and contralesional global effect. We conclude that an important factor in the patients' performance on simple saccade tasks appears to be the form of neglect that they show.
Acknowledgements--This work was funded by an MRC Grant awarded to J. M. Findlay and A. W. Young. We would like to thank two anonymous referees for their helpful suggestions and criticisms made to an earlier version of this paper. We are also grateful to Denise Taylor and Nadina Lincoln for allowing us access to patients.
References
1. Barbur, J. L., Forsyth, P. M. and Findlay, J. M. Human saccadic eye movements in the absence of the geniculocalcarine projection. Brain 111, 63-82, 1988.
2. Findlay, J. M. Global visual processing for saccadic eye movements. Vis. Res. 22, 1033 1045, 1982.
3. Findlay, J. M. Visual information processing for sac- cadic eye movements. In Spatially Oriented Behavior, A. Hein and M. Jeannerod (Editors), pp. 281 303. Springer-Verlag, New York, 1983.
4. Findlay, J. M. and Kapoula, Z. Scrutinization, spa- tial attention, and the spatial programming of sac- cadic eye movements. Q. J. Exp. Psychol. 45A, 633- 647, 1992.
5. Fischer, B. The preparation of visually guided sacca- des. Rev. Physiol. Biochem. Pharmacol. 106, 1 35, 1987.
6. Fischer, B. and Breitmeyer, B. Mechanisms of visual attention revealed by saccadic eye movements. Neu- ropsychologia 25, 73-83, 1987.
7. Fischer, B. and Ramsperger, E. Human express sac- cades: Extremely short reaction times of goal directed eye movements. Exp. Brain Res. 57, 191-195, 1984.
8. Fischer, B. and Weber, H. Express saccades and vis- ual attention. Behav. Brain Sci. 16, 553 610, 1993.
9. Girotti, F., Casazza, M., Musicco, M. and Avanzini, G. Oculomotor disorders in cortical lesions in man:

R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect 507
The role of unilateral neglect. Neuropsychologia 5, 543-553, 1983.
10. Hornak, J. Ocular exploration in the dark by patients with visual neglect. Neuropsychologia 30, 547-552, 1992.
11. Ishiai, S., Furukawa, T. and Tsukagoshi, H. Eye- fixation patterns in homonymous hemianopia and unilateral spatial neglect. Neuropsvchologia 25, 675 670, 1987.
12. Kingstone, A. and Klein, R.M. Visual offsets facili- tate saccadic latency: Does predisengagement of vis- uospatial attention mediate this gap effect? J. Exp. Psychol. Human Perception and Perjormance, 19, 1251 1265, 1993.
13. Kinsbourne, M. Orientational bias model of uni- lateral neglect: Evidence from attentional gradients within hemispace. In Unilateral Neglect: Clinical and Experimental Studies, I. H. Robertson and J. C. Mar- shall (Editors), pp. 63-86. Lawrence Erlbaum Associates, Hillsdale, NJ, 1993.
14. L~.davas, E. Is the hemispatial deficit produced by right parietal lobe damage associated with retinal or gravitational coordinates? Brain 110, 167 180, 1987.
15. Lfidavas, E., Petronio, A. and Umiltfi, C. The deploy- ment of visual attention in the intact field of hemi- neglect patients. Cortex 26, 307 317, 1990
16. L6vy-Schoen, A. Determination et latence de la reponse oculomotrice a deux stimulus. L'Anee Psy- chologique 74, 43 66, 1969.
17. Lynch, J. C. and McLaren, J. W. Deficits of visual attention and saccadic eye movements after lesions of parieto-occipital cortex in monkeys. J. Neuro- physiol. 61, 74-90, 1989.
18. Meienberg, O., Harrer, M. and Wehren, C. Ocu- lographic diagnosis of hemineglect in patients with homonymous hemianopia. J. Neurol. 233, 97 101, 1986.
19. Meienberg, O., Zangemeister, W. H., Rosenberg, M., Hoyt, W. F. and Stark, L. Saccadic eye movement strategies in patients with homonymous hemianopia. Ann. Neurol. 9, 537 544, 1981.
20. Milner, A. D. and Harvey, M. Towards a taxonomy of spatial neglect. In Spatial Neglect: Positional Pap- ers on Theory and Practice, P. W. Halligan and J. C. Marshall (Editors), pp. 177 183. Lawrence Erlbaum Associates, Hove, 1994.
21. Munoz, D. P. Pelisson, D. and Guitton, D. Move- ment of neural activity on the superior colliculus motor map during gaze shifts. Science 251, 1358 1360, 1991.
22. Munoz, D. P. and Wurtz, R. H. Role of the rostral superior colliculus in active visual fixation and execution of express saccades. J. Neurophysiol. 67, 1000-1002, 1992.
23. Munoz, D. P. and Wurtz, R. H. Fixation cells in monkey superior colliculus 1. Characteristics of cell discharge. J. Neurophysiol. 70, 559-575, 1993.
24. Par6, M. and Guitton, D. The fixation area of the cat superior colliculus: Effects of electrical stimu- lation and direct connection with brainstem omni- pause neurons. Exp. Brain Res. 101, 109-122, 1994.
25. Pierrot-Deseilligny, C., Rivaud, S., Gaymard, B. and Agid, Y. Cortical control of reflexive visually-guided saccades. Brain 114, 1473 1485, 1991.
26. Posner, M. L., Walker, J. A., Friedrich, F. J. and Rafal, R. D. Effects of parietal injury on covert ori- enting of attention. J. Neurosci. 4, 1863 1874, 1984.
27. Reulen, J. P. H., Marcus, J. T., Koops, D., de Fries, F. R., Tiesinger, G., Boshuizen, K. and Bos, J. E. Precise recording of eye movements: The IRIS tech- nique. Med. Biol. Eng. Comput. 26, 20-26, 1988.
28. Reuter-Lorenz, P. A., Hughes, H. C. and Fendrich, R. The reduction of saccadic latency by prior offset of the fixation point: An analysis of the gap effect. Percept. Psychophys. 49, 167-175, 1991.
29. Reuter-Lorenz, P. A., Oonk, H. M. Barnes, L. L. and Hughes, H. C. Effects of warning signals and fixation point offsets on the latencies of pro- vs anti- saccades: Implications for an interpretation of the gap effect. Exp. Brain Res. 103, 287-293, 1995.
30. Robertson, 1. H. Anomalies in the lateralisation omissions in unilateral left neglect: Implications for an attentional theory of neglect. Neuropsychologia 27, 157-165, 1989.
31. Robertson, I. H., Halligan, P. W. and Marshall, J. C. Prospects for the rehabilitation of unilateral neglect. In Unilateral Neglect: Clinical and Exper- imental Studies, I. H. Robertson and J. C. Marshall (Editors), pp. 279-292. Lawrence Erlbaum Associ- ates, Hillsdale, N J, 1993.
32. Ross, L. E. and Ross, S. M. Saccade latency and warning signals: Stimulus onset, offset, and change as warning events. Percept. Psychophys. 27, 251 257, 1980.
33. Ross, S. M. and Ross, L. E. Saccade latency and warning signals: Effects of auditory and visual stimu- lus onset and offset. Percept. Psychophys. 29, 429 437, 1981.
34. Saslow, M. G. Effects of components of dis- placement-step stimuli upon latency for saccadic eye movement. J. Opt. Soc. Am. 57, 1024-1029, 1967.
35. Schiller, P. S., True, S. D. and Conway, J. L. Deficits in eye movements following frontal eye-field and superior colliculus ablations. J. Neurophysiol. 44, 1175 1189, 1980.
36. Silberpfennig, J. Contributions to the problem of eye movements. III. Disturbances of ocular movements with pseudohemianopsia in frontal lobe tumours. Confinia Neurologica IV, 1-13, 1941.
37. Sparks, D. L. Translation of sensory signals into commands for control of saccadic eye movements: Role of primate superior colliculus. Physiol. Rev. 66, 118 171, 1986.
38. Sprague, J. M. Interaction of cortex and superior colliculus in mediation of visually guided behavior in the cat. Science 153, 1544-1547, 1966.
39. Tam, W. J. and Stelmach, L. B. Viewing behaviour: Ocular and attentional disengagement. Percept. Psy- chophys. 54, 211-222, 1993.
40. Walker, R. Findlay, J. M., Young, A. W. and Welch, J. Disentangling neglect and hemianopia. Neu- ropsychologia 29, 1019 1027, 1991.
41. Walker, R., Kentridge, R. W. and Findlay, J. M. Independent contributions of the orienting of atten- tion, fixation offset and bilateral stimulation on human saccadic latency. Exp. Brain Res. 103, 294 310, 1995.

508 R. Walker and J. M. Findlay/Saccadic eye movements and unilateral neglect
42. Walker, R., Findlay, J. M., Young, A. W. and Lincoln, N. B. Saccadic eye movements in object- based neglect. Cognit. Neuropsychol. (in press).
43. Wallace, S. F., Rosenquist, A. C. and Sprague, J. M. Recovery from cortical blindness mediated by destruction of nontectotectal fibres in the commisure of the superior colliculus in the cat. J. Comp. Neurol. 284, 429M50, 1989.
44. Wallace, S. F., Rosenquist, A. C. and Sprague, J. M. Ibotenic acid lesions of the lateral substantia nigra restores visual orientation behaviour in the hemianopic cat. J. Comp. Neurol. 296, 222 252, 1990.
45. Young, R. L. and Sheena, D. Eye-movement measurement techniques. Am. Psvchol. 30, 315 330, 1975.
Copyright © 2022 FDOKUMEN




















