
A regional pedal ischemia scoring system for decision analysis in patients with heel ulceration

Pedal Proportions of Poposaurusgracilis: Convergence and Divergence
in the Feet of ArchosaursJAMES O. FARLOW,1* EMMA R. SCHACHNER,2 JOHN CODY SARRAZIN,3
HENDRIK KLEIN,4 AND PHILIP J. CURRIE5
1Department of Geosciences, Indiana-Purdue University, Fort Wayne, Indiana2Department of Biology, University of Utah, Salt Lake City, Utah
3Department of Mechanical Engineering, University of Utah, Salt Lake City, Utah4Saurierwelt Pal€aontologisches Museum, D-92318, Neumarkt, Germany
5Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada
ABSTRACTThe crocodile-line basal suchian Poposaurus gracilis had body propor-
tions suggesting that it was an erect, bipedal form like many dinosaurs,prompting questions of whether its pedal proportions, and the shape ofits footprint, would likewise “mimic” those of bipedal dinosaurs. Weaddressed these questions through a comparison of phalangeal, digital,and metatarsal proportions of Poposaurus with those of extinct andextant crocodile-line archosaurs, obligate or facultatively bipedal non-avian dinosaurs, and ground birds of several clades, as well as a compari-son of the footprint reconstructed from the foot skeleton of Poposauruswith known early Mesozoic archosaurian ichnotaxa. Bivariate and multi-variate analyses of phalangeal and digital dimensions showed numerousinstances of convergence in pedal morphology among disparate archosau-rian clades. Overall, the foot of Poposaurus is indeed more like that ofbipedal dinosaurs than other archosaur groups, but is not exactly like thefoot of any particular bipedal dinosaur clade. Poposaurus likely had a dig-itigrade stance, and its footprint shape could have resembled grallatoridichnotaxa, unless digit I of the foot of Poposaurus commonly left animpression. Anat Rec, 00:000–000, 2014. VC 2014 Wiley Periodicals, Inc.
Key words: Poposaurus; archosaur pedal morphology;vertebrate ichnology
The early Mesozoic adaptive radiation of archosaurs pro-duced a diversity of reptilian life forms; these animalswere variously large or small; aquatic, terrestrial, or aerial;herbivorous or carnivorous; ponderous or cursorial (Ewer,1965; Sereno, 1991; Heckert and Lucas, 2002; Dzik, 2003;Ziegler et al., 2003; Nesbitt, 2003, 2011; Desojo and B�aez,2005; Parker et al., 2005; Sen, 2005; Fraser and Hender-son, 2006; Li et al., 2006; Jalil and Peyer, 2007; Kubo andBenton, 2007, 2009; Schoch, 2007; Peyer et al., 2008;Ezcurra, 2010; Heckert et al., 2010; Sues and Fraser, 2010;Brusatte et al., 2011a; Nesbitt et al., 2010, 2013a,b; Padianet al., 2010; Butler et al., 2011; Franca et al., 2011; Desojoet al., 2011; Lecuona and Desojo, 2011; Galton, 2012;
Grant sponsor: National Science Foundation; Grant number:9612880-EAR; Grant sponsor: Dinosaur Society; Grant sponsor:NSERC; Grant number: 203091-06; Grant sponsors: AmericanAssociation of Anatomists Scholar, American Association ofAnatomists.
*Correspondence to: James O. Farlow, Department of Geo-sciences, Indiana-Purdue University, 2101 East Coliseum Boule-vard, Fort Wayne, IN 46805. Fax: 260 481-6880. E-mail:[email protected]
Received 26 March 2013; Accepted 22 November 2013.
DOI 10.1002/ar.22863Published online in Wiley Online Library(wileyonlinelibrary.com).
THE ANATOMICAL RECORD 00:00–00 (2014)
VVC 2014 WILEY PERIODICALS, INC.

Kammerer et al. 2012; Li et al., 2012; Nied�zwiedzki et al.,2012; Parrish, 2012; Trotteyn et al., 2012, 2013; B�elen VonBaczko and Ezcurra, 2013; Ezcurra et al., 2013; Desojoet al., 2013; Irmis et al., 2013; Kubo and Kubo, 2013;Langer et al., 2013; Nied�ziedzki et al., 2013; Peecock et al.,2013; Sereno et al., 2013; Stocker and Butler, 2013; Turnerand Nesbitt, 2013). Some early Mesozoic archosaurs werebipeds, variously facultative or obligate (Kubo and Kubo,2012). Such animals were at or near the base of the adapt-ive radiation of dinosaurs and their close relatives (aveme-tatarsalians/ornithodirans/bird-line archosaurs: Sereno andArcucci, 1993, 1994; Benton, 1999, 2012; Langer, 2003;Butler et al., 2007; Marsicano et al., 2007; Nesbitt et al.,2007, 2009a,b; Padian, 2008; Martinez and Alcober, 2009;Rinehart et al., 2009; Butler, 2010; Irmis, 2011; Brusatteet al., 2011b; Nesbitt, 2011; Brusatte, 2012; Langer et al.,2013) but also evolved among non-ornithodiran/crurotar-san/crocodile-line archosaurs (Nesbitt and Norell, 2006;Nesbitt, 2007, 2011; Piechowski and Dzik, 2010; Sennikov,2012; Sookias and Butler, 2013; Weinbaum, 2013).
A particularly well-known crocodile-line form withbody proportions suggesting bipedalism is Poposaurus
gracilis from the Late Triassic of the western UnitedStates (Weinbaum and Hungerb€uhler, 2007; Gauthieret al., 2011; Parker and Nesbitt, 2013). Phylogenetic andfunctional morphological considerations suggest thatthis archosaur (Fig. 1) was an erect, striding biped(Gauthier et al., 2011; Schachner et al., 2011; Bates andSchachner, 2012; Kubo and Kubo, 2013). At least super-ficially, the foot of Poposaurus has a very dinosaur-likegestalt (Fig. 2E). This observation prompts a moredetailed, quantitative comparison of the pedal propor-tions of Poposaurus with those of other archosaurs. Wasthis crocodile-line archosaur indeed a dinosaur-mimic indetails of its foot structure? If so, to what dinosaurs isits foot most similar? And what is the likelihood thatfootprints made by such a “dinocroc” might be mistakenfor dinosaur footprints, and vice versa?
MATERIALS AND METHODS
This study builds upon previous comparisons of pedalproportions within and across groups of dinosaurs andground birds (Farlow and Lockley, 1993; Farlow, 2001;
Fig. 1. Skeleton of Poposaurus gracilis (YPM VP.057100) in field jackets, left lateral view. Courtesy ofthe Peabody Museum of Natural History, Yale University, New Haven, Connecticut, USA.
2 FARLOW ET AL.

Smith and Farlow, 2003; Farlow et al. 2006, 2012, 2013)by adding new data (especially for crocodile-line archo-saurs), correcting errors in earlier versions of the database, and by analyzing the data in different ways thanpreviously. We analyzed metatarsal (or tarsometatarsal),digital, and pedal phalangeal lengths, and in someinstances phalangeal distal widths, for a diverse set ofliving and extinct species (Figs. 2, 3; institutional abbre-viations in Table 1) of basal archosauriforms, basalsuchians (including Poposaurus), basal crocodyliforms,crocodylians, basal dinosauromorphs, non-avian thero-pods, basal birds, living and extinct ground birds (palae-ognaths as well as neognaths), and bipedal or potentiallybipedal basal sauropodomorphs (Eoraptor, “prosauropods”[cf. Mallison, 2011]) and ornithischians (Lesothosaurus,ornithopods, basal ceratopsians [cf. Senter, 2007; Leeet al., 2011]). Several issues of specimen preparation or
preservation, or of data reporting, complicated ouranalyses.
For fossil forms the number of specimens with com-plete phalangeal counts is often limited, and due to thevagaries of preservation it is not always possible tomake all the desired measurements on all pedal pha-langes. The smaller and/or more distal non-ungual pha-langes are frequently missing, and the ungualsthemselves often have significant portions of their distaltips missing. For osteological specimens of modern taxain museum collections, dried tissues frequently envelopmetatarsals and phalanges, making it difficult or impos-sible to measure phalangeal dimensions. In osteologicalspecimens of extant crocodylians, the horny claw sheathsoften are so strongly attached to their underlyingunguals that they cannot easily be removed for meas-uring the unguals. Finally, published descriptions of
Fig. 2. Gallery of diagrammatic images of pedal skeletons of selectedarchosauriform reptiles. Scale bars 5 50 mm. Left/right symmetry ofspecimens indicated by labeling digit II. (A–I) Crocodile-line archosaurs.(A) Proterosuchus vanhoepeni (redrawn from Cruickshank [1972]). (B)Revueltosaurus callenderi (PEFO 34561). (C) Stagonolepis robertsoni(redrawn from Walker [1961]). (D) Ticinosuchus ferox (redrawn fromKrebs [1965]). (E) Poposaurus gracilis (YPM VP.057100). (F) Incompletefoot of Shuvosaurus inexpectatus (TTU-P09001). (G) Postosuchus aliso-nae (UNC 15575). (H) Terrestrisuchus gracilis (redrawn from Crush
[1984]). (I) Protosuchus richardsoni (redrawn from Colbert and Mook[1951]). (J–S) Bird-line archosaurs. (J) Silesaurus opolensis (redrawnfrom Dzik [2003]). (K) Coelophysis bauri (MNA V3320). (L) Tyrannosaurusrex (BHI-6230). (M) Dinornis robustus (NMNZ S.28225). (N) Plateosauruslongiceps (Skelett 45, MB.R.4404.67–77). (O) Leptoceratops gracilis(CMN 8889). (P) Thescelosaurus garbanii (Bugenasaura infernalis [cf.Boyd et al., 2009]; LACM 33542). (Q) Parksosaurus warreni (ROM 804).(R) Tenontosaurus tilletti (OU 10132). (S) Brachylophosaurus canadensis(MOR 794).
POPOSAURUS GRACILIS PEDAL PROPORTIONS 3

fossil feet that report measurements of pedal phalangesgenerally are not explicit about how the measurementswere made; in our experience, differences in phalangealproportions due to different measurement protocols aremost striking for ungual rather than non-ungualphalanges.
We therefore did parallel analyses using two differentdata sets. Data set 1 consisted of lengths of the non-ungual phalanges of digits I, II, and III, and the firstthree phalanges of digit IV, and the length of metatarsalIII. The measurements in data set 1 were made by our-selves, or taken from the literature, the latter withoutworrying about the details of how measurements weremade by other authors. In some cases, phalanx lengthswere estimated from scaled drawings of foot skeletons.Data set 1 therefore provided an extensive coverage oftaxa, and a large number of specimens, but at theexpense of measuring fewer bones, and risking “operatorerror” in measurements due to different procedures pos-sibly being followed by different authors. (The cost ofmeasuring fewer bones may not be excessive, however;Farlow et al. [2013] found that a canonical variate anal-ysis [CVA] based on the dimensions of just the first twophalanges of digits II–IV performed almost as well as aCVA based on the dimensions of all the phalanges ofthose digits in discriminating among the foot skeletonsof tridactyl ratites.)
Data set 2 consisted of measurements made by our-selves, or by others following our explicit measurementprotocols (Fig. 4), of the lengths of metatarsal III and allpedal phalanges, and of the distal widths of phalangesII2, III2, and IV2. As in previous studies (Farlow and
Fig. 3. CT scan of the foot of a large (total animal length from snoutto tail tip 3 m), male Alligator mississippiensis, Rockefeller Wildlife Ref-uge, Louisiana. Note that soft tissues surrounding the digits result ingreater splaying of the digits than would be expected from the skele-ton alone. Scale bar 5 20 mm.
TABLE 1. Institutional abbreviations
Abbreviation Institution Location
AM Auckland War Memorial Museum Auckland, New ZealandAMNH American Museum of Natural History New YorkBHI Black Hills Institute of Geological Research Hill City, South DakotaCEUM Utah State University Eastern Prehistoric Museum Price, UtahCM Canterbury Museum Christchurch, New ZealandCMN and GSC Canadian Museum of Nature Ottawa, Ontario, CanadaDMNH Denver Museum of Nature and Science Denver, ColoradoFMNH Field Museum of Natural History Chicago, IllinoisIRSNB Institut Royal des Sciences Naturelles de Belgique Brussels, BelgiumLACM Natural History Museum of Los Angeles County Los Angeles, CaliforniaMB Museum f€ur Naturkunde Berlin, GermanyMCF Museo Municipal Carmen Funes Neuqu�en, ArgentinaMNA Museum of Northern Arizona Flagstaff, ArizonaMOR Museum of the Rockies, Montana State University Bozeman, MontanaNHM Natural History Museum London, EnglandNMNZ Museum of New Zealand Te Papa Tongarewa Wellington, New ZealandOU & OMNH Sam Noble Oklahoma Museum of Natural History Norman, OklahomaPEFO Petrified Forest National Park ArizonaPU Princeton University (Yale Peabody Museum) New Haven, ConnecticutPVSJ Museo de Ciencias Naturales, Universidad de San Juan San Juan, ArgentinaQM Queensland Museum Brisbane, AustraliaROM Royal Ontario Museum Toronto, Ontario, CanadaRTMP Royal Tyrrell Museum of Palaeontology Drumheller, Alberta, CanadaSMA Sauriermuseum Aathal Aathal, SwitzerlandTTU Texas Tech University Lubbock, TexasUCMP University of California Museum of Paleontology Berkeley, CaliforniaU Illinois University of Illinois Museum of Natural History Urbana, IllinoisUNC University of North Carolina Chapel Hill, North CarolinaUSNM National Museum of Natural History, Smithsonian Institution Washington, D.C.YPM Peabody Museum of Natural History, Yale University New Haven, Connecticut
4 FARLOW ET AL.

Lockley, 1993; Farlow, 2001; Smith and Farlow, 2003;Farlow et al., 2006, 2012, 2013), lengths of non-ungualphalanges were measured from the dorso-ventral mid-point (or close to it) of the concave proximal edge of thebone to the dorsoventral midpoint of the convex distalend of the bone; where possible, this measurement wasmade on both the medial and lateral sides of the bone,and the measurements averaged. The maximum trans-verse width of non-ungual phalanges was measuredacross the distal articular end of the bone. Ungual lengthswere measured in a straight-line fashion from the dorso-ventral midpoint of the concave proximal edge of the boneto its tip, again averaging measurements of the medialand lateral sides, when possible. Frequently “composite”measurements of the left and right feet of a single indi-vidual were made, when a complete set of measurementscould not be made on the left or right foot alone.
Data were analyzed in several ways. Multivariateanalyses were employed to analyze phalangeal lengths(and sometimes widths). Principal components analyses(PCAs) were done using log-transformed data and acovariate matrix. Two versions of PCA were performed.The first used data set 1 and the log-transformedlengths of phalanges I1, II1-2, III1-3, and IV1-3. To ena-ble inclusion of taxa in which digit I has been lost, 5 mmwas added to lengths of phalanx I1 of all specimens priorto log transformation. The second PCA employed data
set 2 and the lengths of all phalanges of digits I-IV, andalso the distal widths of phalanges II2, III2, and IV2(which are positioned about halfway out along thelengths of their respective toes). We added 5 mm to thelengths of phalanges I1 and I2 to allow inclusion of taxain which digit I has been lost.
A problem arises with some birds (particularly somemoa species) in which there are only four phalanges(including the ungual) in digit IV. Typically in groundbirds and dinosaurs the smallest bone in digit IV is thefinal non-ungual phalanx, IV4, which can be very short(e.g. in Rhea and Pterocnemia). As in a previous study(Farlow et al., 2013), in forms with only four phalangeson digit IV we treated the ungual as phalanx IV5, andassigned phalanx IV4 a length of zero. In our PCA withdata set 2, instead of using separate log-transformedlengths of both IV3 and IV4 (which would have necessi-tated discarding specimens in which IV4 had zerolength), we used the log-transformed summed lengths ofIV3 and IV4.
Cluster analyses of phalangeal lengths and widthsusing data set 2 were performed after first log-transforming all variables used in the analysis, comput-ing the mean of all the log-transformed variables, andfinally scaling each log-transformed variable by subtract-ing the mean of all the log-transformed variables in theanalysis from each log-transformed variable. Clusteranalyses used between-groups linkage employing thesquared Euclidean distance.
Finally, simple bivariate comparisons were madewhenever possible, to increase the sample size involvedin the comparison. Because the number of specimensrepresenting any given taxon in any particular bivariatecomparison was highly variable (some species, such asAlligator mississippiensis, were represented by manyspecimens, but most species were represented by one ora few specimens), no attempt was made to test statisti-cally significant differences among groups in bivariatecomparisons. Instead we looked for consistent visual pat-terns in which data cases for particular groups plottedin different regions of graphs. For some bivariategraphs, we log-transformed data, to better show pointsfor both very large and very small forms. In other bivari-ate graphs, we did not log-transform data before plot-ting, but rather restricted the plotted data to formscomparable to or smaller than Poposaurus in size. Thiswas a strictly ad hoc decision, based on which form ofgraph was judged better to show how Poposaurus com-pared with other taxa.
All data analyses were performed in IBM SPSS version20. The data upon which analyses were based are availablefrom Farlow. In the graphs that follow (Figs. 5-8, 10, 11),limitations in the number of different symbols the plottingroutines permitted us to use forced some uncomfortablechoices in labeling points. We tried to use consistent sym-bols and color schemes (Table 2). Where possible to do sowithout creating excessive clutter, we also labeled points(or groups of points) for individual genera (Table 2).
Some of the groups used in the graphs are paraphy-letic or otherwise idiosyncratic. Thus Archaeopteryx wasgiven its own symbol, and not included with other basalbirds, partly because of the intrinsic interest of thisform, and partly because of controversy over exactly howit relates to non-avian theropods and avialians (Xuet al., 2011b; Lee and Worthy, 2012).
Fig. 4. Single phalanges of Poposaurus illustrating protocols formeasuring pedal phalanges. (A) Lengths of non-ungual phalanges. (B)Distal widths of non-ungual phalanges. (C) Lengths of ungualphalanges.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 5
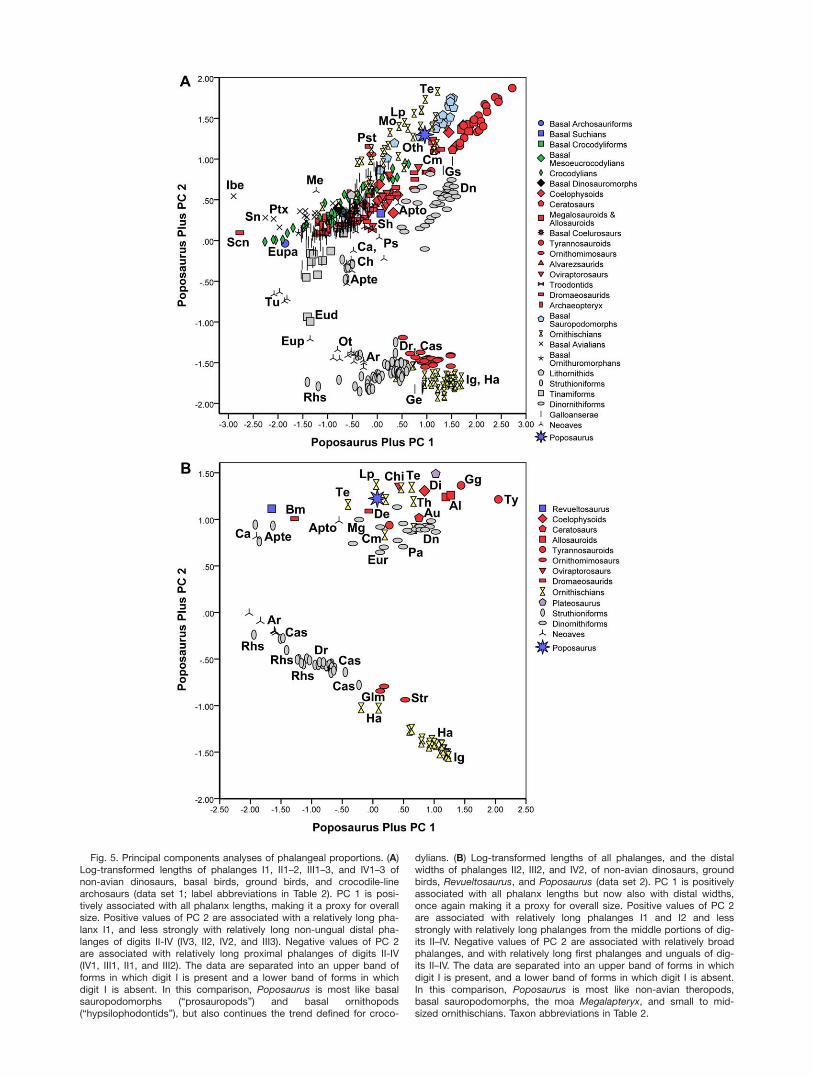
Fig. 5. Principal components analyses of phalangeal proportions. (A)Log-transformed lengths of phalanges I1, II1–2, III1–3, and IV1–3 ofnon-avian dinosaurs, basal birds, ground birds, and crocodile-linearchosaurs (data set 1; label abbreviations in Table 2). PC 1 is posi-tively associated with all phalanx lengths, making it a proxy for overallsize. Positive values of PC 2 are associated with a relatively long pha-lanx I1, and less strongly with relatively long non-ungual distal pha-langes of digits II-IV (IV3, II2, IV2, and III3). Negative values of PC 2are associated with relatively long proximal phalanges of digits II-IV(IV1, III1, II1, and III2). The data are separated into an upper band offorms in which digit I is present and a lower band of forms in whichdigit I is absent. In this comparison, Poposaurus is most like basalsauropodomorphs (“prosauropods”) and basal ornithopods(“hypsilophodontids”), but also continues the trend defined for croco-
dylians. (B) Log-transformed lengths of all phalanges, and the distalwidths of phalanges II2, III2, and IV2, of non-avian dinosaurs, groundbirds, Revueltosaurus, and Poposaurus (data set 2). PC 1 is positivelyassociated with all phalanx lengths but now also with distal widths,once again making it a proxy for overall size. Positive values of PC 2are associated with relatively long phalanges I1 and I2 and lessstrongly with relatively long phalanges from the middle portions of dig-its II–IV. Negative values of PC 2 are associated with relatively broadphalanges, and with relatively long first phalanges and unguals of dig-its II–IV. The data are separated into an upper band of forms in whichdigit I is present, and a lower band of forms in which digit I is absent.In this comparison, Poposaurus is most like non-avian theropods,basal sauropodomorphs, the moa Megalapteryx, and small to mid-sized ornithischians. Taxon abbreviations in Table 2.

The relationships of neornithine paleognaths are par-ticularly controversial (cf. Livezey and Zusi, 2007;Harshman et al., 2008; Bourdon et al., 2009; Phillipset al., 2010; Smith et al., 2013). Different analyses agreein finding the two extant rhea species (Rhea americanaplus Pterocnemia pennata), the emu (Dromaius novae-hollandiae) plus the cassowaries (Casuarius spp.), andthe various moa species (dinornithiforms), respectively,to constitute clades. In contrast, the relationships amongtinamous, moa, and the extant ratites vary markedlyamong studies. In our graphs we will use separate sym-bols for lithornithids, tinamiforms, and dinornithiforms.We will label emu, cassowaries, rheas, ostrich, and kiwicollectively as struthioniforms (Clements, 2007) but forconvenience more than anything else. Indeed, a recur-rent theme in our analyses (as in Farlow et al., 2013)will be how kiwi (Apteryx) consistently differ from otherextant ratites in pedal proportions.
To create a three-dimensional digital model of the dis-tal hindlimb and pes of Poposaurus, the left femur, tibia,fibula, metatarsals, and pes were scanned with a Polhe-mus FastSCAN cobra laser scanner (www.polhemus.
TABLE 2. Abbreviations of genus or group names(alphabetical order) used in Figures 5-8, 10, and 11,and the group names associated with each taxon in
the figure symbol keys
Abbreviation Genus or group
Al Allosaurus (Allosauroids)Alb Albertosaurus (Tyrannosauroids)Ali Alligator (Crocodylians)An Anomalopteryx (Dinornithiforms)Apte Apteryx (Struthioniforms)Apto Aptornis (Neoaves)Ar Ardeotis (Neoaves)Arc Archaeoceratops (Ornithischians)Ari Araripesuchus (Basal
Mesoeucrocodylians)Au Aucasaurus (Ceratosaurs)Bm Bambiraptor (Dromaeosaurids)Ca Cariama (Neoaves)Cai Caiman (Crocodylians)Cas Casuarius (Struthioniforms)Cer Cerasinops (Ornithischians)Ch Chunga (Neoaves)Chi Chirostenotes (Oviraptorosaurs)Chr Chanaresuchus (Basal Archosauriforms)Chry Chrysolophus (Galloanserae)Cm Camptosaurus (Ornithischians)Co Confuciusornis (Basal Avialians)Cro Crocodylus (Crocodylians)Cyr Cyrtonyx (Galloanserae)Das Daspletosaurus (Tyrannosauroids)De Deinonychus (Dromaeosaurids)Di Dilophosaurus (Coelophysoids)Dn Dinornis (Dinornithiforms)Dr Dromaius (Struthioniforms)Dry Dryosaurus (Ornithischians)Eud Eudromia (Tinamiforms)Eup Eupodotis (Neoaves)Eupa Euparkeria (Basal Archosauriforms)Eur Euryapteryx (Dinornithiforms)Gav Gavialis (Crocodylians)Ge Genyornis (Galloanserae)Gg Gorgosaurus (Tyrannosauroids)Glm Gallimimus (Ornithomimosaurs)Gs Gastornis (Galloanserae)Ha Hadrosaurids (several genera;
Ornithischians)Het Heterodontosaurus (Ornithischians)Hng Hongshanornis (Basal
Ornithuromorphans)Hy Haya (Ornithischians)Ibe Iberomesornis (Basal Avialians)Ig Iguanodon and Mantellisaurus
(Ornithischians)Lg Lagerpeton (Basal Dinosauromorphs)Lp Leptoceratops (Ornithischians)Lph Lophura (Galloanserae)Lu Lufengosaurus (Basal Sauropodomorphs)Me Mesitornis (Neoaves)Mg Megalapteryx (Dinornithiforms)Mi Microceratops (Ornithischians)Mo Montanoceratops (Ornithischians)Ms Massospondylus (Basal
Sauropodomorphs)Or Orodromeus (Ornithischians)Ost Osteolaemus (Crocodylians)Ot Otis (Neoaves)Oth Othnielosaurus (Ornithischians)Pa Pachyornis (Dinornithiforms)Pal Palaeociconia (Neoaves)Ph Phasianus (Galloanserae)
TABLE 2. (continued)
Abbreviation Genus or group
Pk Parksosaurus (Ornithischians)Pl Plateosaurus (Basal Sauropodomorphs)PL Proctor Lake ornithopod (Ornithischians)Plsh Paleosuchus (Crocodylians)Post Postosuchus (Basal Suchians)Pr Protosuchus (Basal Crocodyliforms)Prt Proterosuchus (Basal Archosauriforms)Ps Psilopterus (Neoaves)Pso Psophia (Neoaves)Pst Psittacosaurus (Ornithischians)Ptg Patagopteryx (Basal Ornithuromorphans)Ptx Protopteryx (Basal Avialians)Re Revueltosaurus (Basal Suchians)Rhn Rhynchotus (Tinamiforms)Rhs Rhea and Pterocnemia (Struthioniforms)Sa Saurosuchus (Basal Suchians)Sat Saturnalia (Basal Sauropodomorphs)Scn Scansoriopteryx (Dromaeosaurids)Sgt Sagittarius (Neoaves)Sgx Saurophaganax (Allosauroids)Sh Shuvosaurus (Basal Suchians)Sn Sinornis (Basal Avialians)St Struthio (Struthioniforms)Ste Steneosaurus (Basal Mesoeucrocodylians)Str Struthiomimus (Ornithomimosaurs)Te Tenontosaurus (Ornithischians)Th Thescelosaurus (including Bugenasaura;
Ornithischians)To Tomistoma (Crocodylians)Tu Turnix (Neoaves)Tx Typothorax (Basal Suchians)Ty Tyrannosaurus (Tyrannosauroids)
The genera listed here are not the only ones used in theanalyses, but merely the ones that could be labeled on thegraphs. Color conventions for points belonging to groups ingraphs employing data set 1: green 5 crocodyliforms; darkblue 5 crocodile-line archosaurs other than crocodyliforms;black 5 basal dinosauromorphs; red 5 non-avian theropodsand Archaeopteryx; light blue 5 basal sauropodomorphs;light gray 5 paleognaths; yellow 5 ornithischians. There area few departures from these color conventions in graphsemploying data set 2.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 7

com), which generated a high-resolution, digital, three-dimensional point cloud. All data post processing andimage creation was performed using Geomagic Studio 12(www.geomagic.com). As each element was scanned inmultiple parts, it was necessary to re-align individualscans to produce a single 3D mesh of each bone. Individ-ual scans were first manually aligned using the pickpoints function, after which their alignment was refinedusing the surface-matching algorithm on overlappingareas. Once all scans of an element were maximally reg-istered, mesh repair and ‘wrapping’ were applied to sealthe two halves, to fix any non-uniformities in the pointcloud data, and to ultimately produce congruent ‘watertight’ meshes for further utilization (for more informa-tion on 3D models and mesh production see the Supple-mental Information of Bates and Schachner [2012]).Special attention was paid to the calcaneal/astrageal andphalangeal scans, where multiple bones were fossilizedas one unit and could not be separated during prepara-tion without risking extensive damage. After the appro-priate scans were mated, the available exposed surfaceof each constitutive bone was systematically removedfrom the overall scan computationally, and the holes leftby the unexposed surface area in the fossil clump wererepaired in Geomagic. Some liberty was taken in recon-structing surfaces where the scans were incomplete, butanatomical insight and comparison with the oppositelimb allowed for increased accuracy. Once a completesurface mesh was created for each fossilized bone, theentire lower limb was rearticulated into the various con-figurations for further analysis, based upon wear facetsand the original pedal articulation of the fossil specimen.Anatomical abbreviations for features of our digitalmodel of the Poposaurus foot are in Table 3.
We used the proportions of the foot skeleton of Popo-saurus to reconstruct what a pedal footprint of this rep-tile would look like, using standard procedures (Baird,1957). The reconstructed footprint was then comparedwith several Triassic reptilian ichnotaxa to see if anyknown forms are similar.
RESULTS
Phalangeal and Digital Proportions inPoposaurus and Other Archosaurs
Phalangeal proportions. PCA (data set 1) of log-transformed lengths of phalanges I1, II1-II2, III1-III3,and IV1-IV3 (Table 4; Fig. 5A) identifies a first principalcomponent strongly associated with overall foot size thataccounts for the bulk of data variance, as found in previ-ous analyses of this kind (Farlow, 2001; Farlow et al.,2012, 2013). A much smaller, but still important, secondprincipal component distinguishes forms that have adigit I, and also relatively long more distal non-ungualphalanges (positive values of PC 2), from forms that lackdigit I and also have relatively long more proximal non-ungual phalanges (negative values of PC 2). Plotting PC2 from PC 1 separates specimens into two bands thatdiverge with increasing overall foot size.
The lower band (negative values of PC 2) includesmany extant struthioniforms (Casuarius, Dromaius, Pter-ocnemia, Rhea; Struthio was excluded from this analysis(and most other analyses in this study) because it lacksdigit II as well as digit I), some tinamous (Eudromia), thedromornithid Genyornis, bustards (Ardeotis, Eupodotis,Neotis, Otis) and buttonquail (Turnix), most ornithomimo-saurs (Dromiceiomimus, Gallimimus, Ornithomimus,Struthiomimus), and very large ornithopods (Iguanodon,Mantellisaurus and several genera of hadrosaurs). Thedivergence of this lower band from the upper band withincreasing animal size is largely a result of the way pha-lanx 1 length was log-transformed in order to includetaxa lacking digit I; with very large ornithopods in whichdigit I has been lost (cf. Moreno et al., 2007), the artificial5-mm length of phalanx I1 becomes trivial compared withthe lengths of phalanges in digits II–IV.
The upper band (positive values of PC 2) includesmany genera of archosaurs and their close relatives from
TABLE 3. Anatomical abbreviations
Abbreviation Meaning of Abbreviation
a Astragalusc Calcaneumf Fibulamt Metatarsalp Phalangest Tibia
TABLE 4. Principal components analysis (using a covariance matrix) of log-transformed lengths of phalangesI1 (5 mm were added to all lengths of this bone prior to log-transformation to enable inclusion of specimens
in which digit I has been lost), II1-II2, III1-III3, and IV1-IV3 of basal archosaurs and their close relatives,basal suchians, basal mesoeucrocodylians, crocodylians, basal dinosauromorphs, non-avian dinosaurs, and
ground birds (number of specimens 5 593)
ParameterComponent 1 Loading
(Raw [Rescaled])Component 2 Loading
(Raw [Rescaled])
I1 Length 0.109 (0.308) 0.332 (0.940)II1 Length 0.377 (0.971) 20.072 (20.186)II2 Length 0.307 (0.956) 0.051 (0.158)III1 Length 0.367 (0.974) 20.072 (20.192)III2 Length 0.313 (0.972) 20.023 (20.070)III3 Length 0.289 (0.968) 0.023 (0.077)IV1 Length 0.377 (0.966) 20.076 (20.195)IV2 Length 0.345 (0.978) 0.032 (0.092)IV3 Length 0.327 (0.961) 0.061 (0.180)Eigen values (% of variance) 0.932 (84.291) 0.135 (12.209)Cumulative variance explained (%) 84.291 96.500
Kaiser-Meyer-Olkin measure of sampling adequacy 5 0.883; Bartlett’s test of sphericity: chi-square 5 13462.273, p<0.001.
8 FARLOW ET AL.

several major groups: basal archosauriforms (Euparke-ria), basal suchians (Poposaurus, Revueltosaurus, Shuvo-saurus, Stagonolepis, Ticinosuchus), basal crocodyliforms(Protosuchus), basal mesoeucrocodylians (Araripesuchus,Steneosaurus), crocodylians (Alligator, Caiman, Crocody-lus, Osteolaemus, Paleosuchus, Tomistoma), basal dino-sauromorphs (Lagerpeton), coelophysoids (Coelophysis,Dilophosaurus, Megapnosaurus, Segisaurus), ceratosaurs(Aucasaurus), allosauroids (Allosaurus), basal coeluro-saurs (Compsognathus, Juravenator, Sinosauropteryx,Tanycolagreus), tyrannosauroids (Albertosaurus, Dasple-tosaurus, Gorgosaurus, Tarbosaurus, Tyrannosaurus), theornithomimosaur Harpymimus, alvarezsaurids (Albiny-kus, Mononykus), oviraptorosaurs (Caudipteryx, Chiroste-notes, Conchoraptor, Ingenia, Khaan), troodontids(Sinornithoides, Troodon), dromaeosaurids (Bambiraptor,Deinonychus, Neuquenraptor, Scansoriopteryx, Sinorni-thosaurus, Velociraptor), Archaeopteryx and other basalavialians (Bohaiornis, Confuciusornis, Iberomesornis,Jeholornis, Protopteryx, Sinornis, Zhouronis), basal orni-thuromorphans (Changmaornis, Gansus, Hongshanornis,Yanornis, Yixianornis), kiwi (Apteryx), moa (Anomalopteryx,Dinornis, Euryapteryx, Megalapteryx, Pachyornis), severalmembers of Galloanserae (Gastornis and many species ofcurrasows, pheasants, grouse, and quail), several taxa ofNeoaves (Aptornis, Cariama, Chunga, Foro, Gallirallus,Idiornis, Juncitarsus, Mesitornis, Rhynochetos, Psilopterus,Psophia, Sagittarius, Salmila), basal sauropodomorphs(Gyposaurus, Lufengosaurus, Massospondylus, Plateosau-rus), and several taxa of small to medium-sized ornithi-schians (Lesothosaurus, Heterodontosaurus, Agilisaurus,Haya, Orodromeus, Othnielosaurus, Parksosaurus, Thesce-losaurus, Yandusaurus, Tenontosaurus, Camptosaurus,Psittacosaurus, Archaeoceratops, Cerasinops, Leptoceratops,Microceratops, Montanoceratops).
Within the upper cluster, data for some groups repre-sented by multiple specimens fall in a series of roughlyparallel bands, all of which show more positive values of
PC 2 with increasing animal size. The lowest band is formoa, with non-avian theropods, crocodylians, basal sau-ropodomorphs, and small to medium-sized ornithischians(“hypsilophodontids,” Tenontosaurus, basal ceratopsians)plotting progressively higher. Poposaurus plots amongthe basal sauropodomorphs and ornithischians but alsois arguably along the trend defined by crocodylians.
PCA using data set 2 yields analogous results (Table5). PC 1 again is associated with animal size, and againaccounts for more than half the data variance, but notas much as in PCA using data set 1. The cumulativepercentage of variance explained by principal compo-nents 1 and 2 is about the same for data sets 1 and 2,and so PC 2 accounts for substantially more of the datavariance in data set 2 than in data set 1. As in PC 2 ofdata set 1, positive values are associated with having adigit I, and also relatively long phalanges from the mid-dle portions of toes (that is, phalanges between the mostproximal phalanx and the ungual). Negative values ofPC 2 are associated with relatively long proximal pha-langes of digits II-IV, as in PC 2 of data set 1, but alsowith relatively long unguals, and with relatively broadphalanges (particularly in large ornithopods [Morenoet al., 2007]).
Plotting PC 2 against PC 1 of data set 2 (Fig. 5B)again separates the data into two bands that divergewith increasing values of PC 1 (animal size), againlargely on the basis of whether digit I is absent (bus-tards, most tridactyl struthioniforms, most ornithomimo-saurs, Iguanodon, Mantellisaurus, and hadrosaurids) orpresent (most non-avian theropods, kiwi [Apteryx], serie-mas [Cariama, Chunga], adzebill [Aptornis], moa, basalsauropodomorphs, smaller ornithopods, basal ceratop-sians, Revueltosaurus, and Poposaurus). Pedal propor-tions of Poposaurus are most like those of non-aviantheropods, the moa Megalapteryx, the ornithischiansTenontosaurus, Thescelosaurus, and Leptoceratops, andthe crurotarsan Revueltosaurus.
TABLE 5. Principal components analysis (using a covariance matrix) of log-transformed phalanx lengths ofall phalanges of digits I–IV (5 mm were added to all lengths of phalanges I1 and of I2 prior to log-
transformation to enable inclusion of specimens in which digit I has been lost), and distal widths ofphalanges II2, III2, and IV2, in Revueltosaurus, Poposaurus, non-avian dinosaurs, and ground birds (number
of specimens 5 108)
ParameterComponent 1 Loading
(Raw [Rescaled])Component 2 Loading
(Raw [Rescaled])
I1 Length 0.198 (0.431) 0.410 (0.894)I2 Length 0.196 (0.404) 0.439 (0.906)II1 Length 0.249 (0.924) 20.069 (20.257)II2 Length 0.302 (0.952) 0.031 (0.097)II2 Distal Width 0.357 (0.947) 20.094 (20.249)II3 Length 0.267 (0.933) 20.041 (20.143)III1 Length 0.223 (0.909) 20.061 (20.250)III2 Length 0.166 (0.818) 0.005 (0.025)III2 Distal Width 0.343 (0.909) 20.141 (20.373)III3 Length 0.210 (0.904) 0.023 (0.101)III4 Length 0.268 (0.958) 20.048 (20.178)IV1 Length 0.224 (0.910) 20.073 (20.299)IV2 Length 0.269 (0.942) 0.036 (0.125)IV2 Distal Width 0.358 (0.943) 20.107 (20.283)Combined IV3 Length 1 IV4 Length 0.290 (0.956) 0.046 (0.152)IV5 length 0.284 (0.966) 20.050 (20.170)Eigen values (% of variance) 1.149 (68.577) 0.426 (25.415)Cumulative variance explained (%) 68.577 93.993
Kaiser-Meyer-Olkin measure of sampling adequacy 5 0.902; Bartlett’s test of sphericity: chi-square 5 4612.581, p< 0.001.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 9

Aspects of these relationships can be further elabo-rated, with larger sample sizes, by selected bivariatecomparisons. Principal component 2 in PCA of both datasets is closely associated with the relative length (orabsence) of digit I. For those forms that retain digit I,plotting phalanx I1 length against the length of digit IIIexcluding the ungual (Fig. 6A; data set 1) separates thevarious taxa into a series of distinct bands. Moa, kiwi,the lithornithid Pseudocrypturus, some tinamous, manygalloanseriforms and Neoaves, Patagopteryx, Rahonavis,Archaeopteryx, Protarchaeopteryx, and most non-aviantheropods plot together in a band of points with a pha-lanx I1 that is present, but relatively short comparedwith the length of digit III. Basal archosauriforms (Cha-naresuchus, Euparkeria, Proterosuchus), basal suchians(Postosuchus, Revueltosaurus, Shuvosaurus, Stagonole-pis, Ticinosuchus, Typothorax), basal mesoeucrocodylians(Araripesuchus, Steneosaurus), crocodylians (Alligator,Caiman, Crocodylus, Gavialis, Osteolaemus, Paleosu-chus, Tomistoma, Wannaganosuchus), basal sauropodo-morphs, and small to medium-sized ornithischians(Lesothosaurus, Heterodontosaurus, basal ornithopods,basal ceratopsians) comprise a heterogeneous group witha particularly long phalanx I1 compared to the length ofdigit III. Poposaurus (itself a basal suchian) falls withinthis last group, plotting among basal sauropodomorphsand “hypsilophodontids”, but is aligned with the trenddefined by crocodylians and other non-ornithodirans.
After the presence or absence of digit I, two of thevariables with the greatest loadings on PC 2 (but withopposite signs) in data set 1 are the lengths of pha-langes II2 (positive association) and IV1 (negative asso-ciation) (Fig. 6B). Large ornithopods (hadrosaurs,Iguanodon, and Mantellisaurus) and several kinds ofground birds (bustards, Gastornis, Genyornis, manymoa, and many struthioniforms) have a relatively longphalanx IV1 compared with the length of phalanx II2(exaggeratedly so in the ostrich Struthio, in which digitII is absent, and so II2 length is assigned a value ofzero). A variety of crocodile-line archosaurs, non-aviantheropods, basal sauropodomorphs, ground birds, andornithischians form a dense “main sequence” of pointsin which the length of IV1 relative to that of II2 is moremodest. Poposaurus plots among this latter group ofpoints but plots noticeably lower than the trend definedby crocodylians.
In data set 2, one of the most important contrasts inPC 2 after the presence or absence of digit I involves therelative widths of phalanges. Plotting the distal width ofphalanx III2 against the length of the same phalanx(Fig. 7A) separates the very stout-toed large ornithopodsfrom other non-avian dinosaurs, ground birds, Poposau-rus, and Revueltosaurus. Non-avian theropods and manyground birds are also slimmer-toed than most small tomid-sized ornithischians, and Poposaurus is more likenon-avian theropods than nearly all ornithischians inthis comparison.
Another aspect of PC 2 in data set 2 is the length ofunguals relative to the lengths of phalanges from themiddle portions of digits (Fig. 7B). Most ornithischianshave relatively longer unguals than do theropods (includ-ing ground birds) and Revueltosaurus, although small tomid-sized ornithopods overlap the theropod points. Popo-saurus plots among the larger birds, close to points forboth non-avian theropods and “hypsilophodontids.”
One feature in which Poposaurus is distinct frommany non-avian dinosaurs (but like galliform birds) is inhaving a rather short ungual on digit I compared withthe length of phalanx I1 (Fig. 8A). In contrast, Revuelto-saurus, Plateosaurus, basal avialians, basal ornithuro-morphans, kiwi, moa, and some other ground birds havea relatively long ungual I2 compared with the length ofphalanx I1.
A cluster analysis of the scaled lengths of the pha-langes of digits I-IV, along with the scaled distal widthsof phalanges II2, III2, and IV2, in birds, dinosaurs,Revueltosaurus, and Poposaurus yields a very large den-drogram (Fig. 9). At low levels in the dendrogram, thereare several phylogenetically reasonable groupings: large,narrow-toed moa (Dinornis plus Pachyornis); cassowaries(Casuarius); most rhea (Pterocnemia and Rhea) speci-mens; emus (Dromaius); bustards (Ardeotis); dromaeo-saurids (Bambiraptor, Deinonychus); ornithomimids(Gallimimus, Struthiomimus); specimens of Iguanodonbernissartensis; most hadrosaur specimens.
There are also some phylogenetically satisfying clus-ters at higher levels in the dendrogram. Most specimensof extant large struthioniforms (cassowaries, emu, rheas)cluster together (cf. Farlow et al., 2013), as do Iguano-don, Mantellisaurus, and hadrosaurids.
Other clusters are phylogenetically more problematic.Some specimens of large theropods (Aucasaurus, Allo-saurus, Gorgosaurus, Tyrannosaurus) of all sampledclades cluster together (cf. Farlow et al., 2013). Kiwi(Apteryx) cluster with the small moa Megalapteryx, theadzebill Aptornis, and the seriema Cariama. Tenontosau-rus plausibly clusters with the “hypsilophodontid” The-scelosaurus (?Bugenasaura), as well as Camptosaurusand Leptoceratops, but also with the basal sauropodo-morph Plateosaurus and the basal suchian Revueltosau-rus. Bustards (a group within Neoaves) cluster withextant large ratites, as do ornithomimosaurs (therebyliving up to their nickname of “ostrich mimics”). Largeornithopods form a cluster with all other groups com-bined. Although phalangeal proportions work reasonablywell at grouping congeneric (and often even conspecific)forms together, they seem to be phylogenetically unreli-able at higher taxonomic levels. Of greatest interest forthe present study, Poposaurus groups with the non-avian theropods Dilophosaurus, Chirostenotes, and onespecimen of Gorgosaurus.
Digital proportions. Comparisons of the overalllengths of digits II-IV (Fig. 10) show ornithischians gen-erally to have relatively long digits II and IV comparedwith the length of digit III, while most non-avian thero-pods and ground birds have proportionally shorter digitsII and IV compared with digit III. Dromaeosaurids, troo-dontids (cf. Zanno et al., 2011), basal sauropodomorphsShuvosaurus, Revueltosaurus, and to a lesser extent croc-odylians, have a relatively high digit IV/digit III lengthratio, but without a correspondingly high digit II/digit IIIlength ratio (Fig. 10A,B). Poposaurus plots among pile-ups of points for large theropods, oviraptorosaurs, thebasal bird Confuciusornis, moa (Dinornis, Megalapteryx),the quail Cyrtonyx, and “hypsilophodontids.”
In those ornithischians that have a digit I, this digitis generally fairly long, relative to the length of digit III(Fig. 8B). The same is true of Revueltosaurus and some
10 FARLOW ET AL.

Fig. 6. Bivariate phalangeal proportions across all groups (data set 1).(A) Phalanx I1 length vs. digit III length (excluding the ungual) in archo-saurs that retain digit I. Data are log-transformed to spread out cases ofsmaller forms; no particular structural relationship between the two vari-ables is assumed. Poposaurus plots among or along the trends defined
by basal sauropodomorphs (“prosauropods”), small to medium-sizedornithischians, and crocodylians. (B) Phalanx II2 length vs. phalanx IV1length in archosaurs of about the same size as Poposaurus or smaller;Poposaurus has a relatively short phalanx IV1 in this comparison.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 11

Fig. 7. Bivariate phalangeal proportions in Poposaurus, Revueltosau-rus, and dinosaurs similar to or smaller than Poposaurus in size (dataset 2). (A) Phalanx III2 distal width vs. length of the same phalanx.Poposaurus is most like non-avian theropods, ground birds, and
“hypsilophodontids.” (B) Ungual III4 length vs. phalanx III2 length.Ornithischians tend to have relatively longer unguals than do thero-pods (including ground birds). Poposaurus is theropod-like in this fea-ture, but plots close to points for “hypsilophodontids.”
12 FARLOW ET AL.
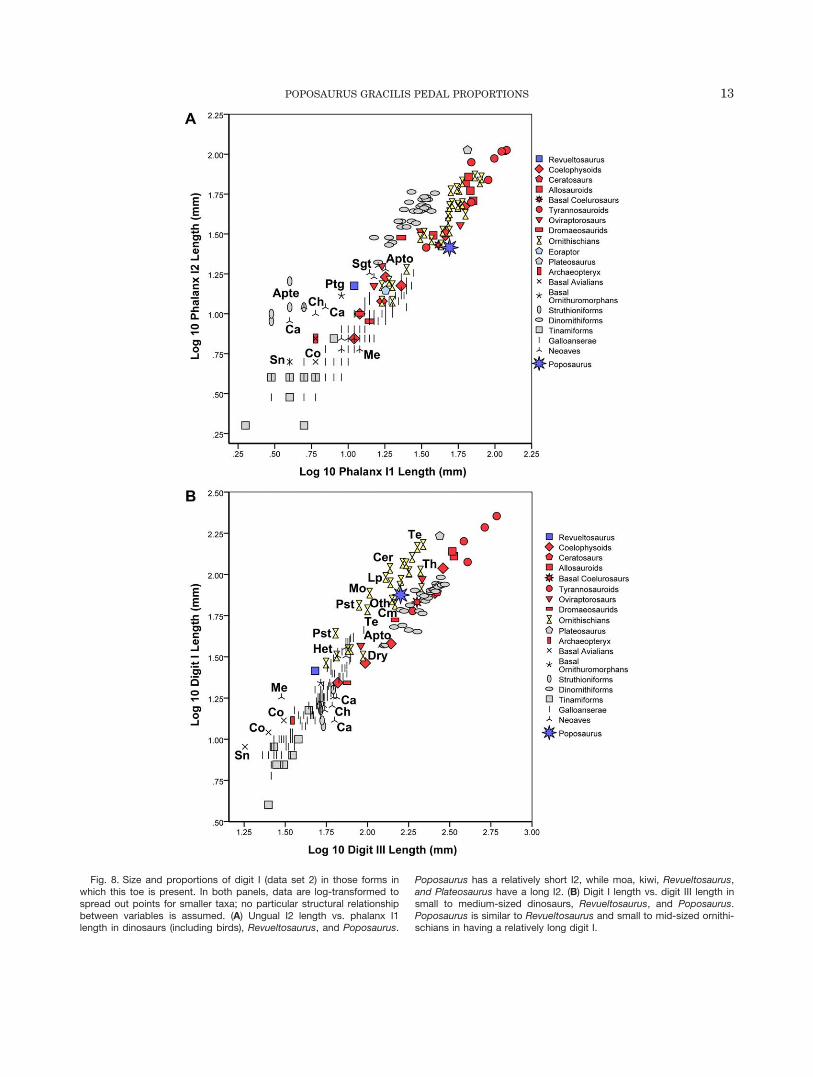
Fig. 8. Size and proportions of digit I (data set 2) in those forms inwhich this toe is present. In both panels, data are log-transformed tospread out points for smaller taxa; no particular structural relationshipbetween variables is assumed. (A) Ungual I2 length vs. phalanx I1length in dinosaurs (including birds), Revueltosaurus, and Poposaurus.
Poposaurus has a relatively short I2, while moa, kiwi, Revueltosaurus,and Plateosaurus have a long I2. (B) Digit I length vs. digit III length insmall to medium-sized dinosaurs, Revueltosaurus, and Poposaurus.Poposaurus is similar to Revueltosaurus and small to mid-sized ornithi-schians in having a relatively long digit I.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 13

Meosozic birds. In contrast, non-avian theropods andmoa have a relatively short digit I. Poposaurus plotsnear the lower edge of the ornithischian points.
Metatarsal/digital proportions. Crocodyliansand most other crocodile-line archosaurs in our sample,along with Dilophosaurus, allosauroids, dromaeosaurids,Chirostenotes, moa other than Dinornis, basal sauropo-domorphs most small ornithischians, Tenontosaurus,Camptosaurus, Iguanodon, and Mantellisaurus have arelatively long digit III compared with the length ofmetatarsal III (or the tarsometatarsus) (Fig. 11A,B). Incontrast, most tyrannosaurs and ornithomimosaurs, aswell as Dinornis and most other ground birds, have aproportionally shorter digit III (or proportionally longermetatarsus). (There is, however, a complication in suchcomparisons, created by the fact that in birds the lengthof metatarsal III will be inflated, compared to that ofnon-avian archosaurs, due to inclusion of ankle bones inthe length of the tarsometatarsus). Poposaurus contin-ues a trend defined by crocodylians (Fig. 11A) and is oth-erwise most similar to Deinonychus, “prosauropods,” andsmall ornithischians.
Other pedal features. Metatarsal I length variesgreatly relative to the lengths of metatarsals II–IV inarchosaurs and their close relatives (Figs. 2, 3). It is sub-stantially shorter than the other metatarsals in basalarchosauriforms like Chanaresuchus (Romer, 1972), Pro-terosuchus, and Euparkeria (Ewer, 1965) but is also rela-tively quite short or even absent in theropods (includingground birds) and derived ornithopods. In contrast, meta-tarsal I is only a little shorter than the other metatarsalsin crocodyliforms (e.g. Protosuchus, Terrestrisuchus, andAlligator). Poposaurus is one of several phylogeneticallyheterogeneous taxa (along with forms like Stagonolepis,Ticinosuchus, Shuvosaurus, Plateosaurus, Tenontosau-rus, basal ornithopods [“hypsilophodontids”], and Lepto-ceratops) in which the relative length of metatarsal I isneither particularly long nor short. However, the aggre-gate length of the phalanges of digit I (especially theshortness of ungual I2 [Fig. 8A]), added to the modestlength of metatarsal I, causes the first toe of Poposaurusto terminate well proximally to the distal termination ofdigits II–IV (Fig. 2E). This configuration differs from thatof crocodylians and their close relatives, basal sauropodo-morphs, basal ceratopsians, Tenontosaurus, and some
Fig. 9. Cluster analysis of scaled lengths of all phalanges of digits I-IV, and the distal widths of phalanges II2, III2, and IV2 in Revueltosau-rus, Poposaurus, non-avian dinosaurs, and ground birds (data set 2:N 5 108; see Table 1 for institutional abbreviations). As in the PCA(Fig. 5B), 5 mm were added to the lengths of phalanges I1 and I2before scaling so that taxa in which digit I is lost could be included.Phalangeal measurements were scaled by subtracting the mean of alllog-transformed phalanx lengths and widths from each log-transformed phalanx length or width. Feet of conspecific or congene-ric specimens tend to cluster together. At the highest levels are threeclusters, the first two of which are phylogenetically problematic: acluster composed of moa, most non-avian theropods, Poposaurus,Revueltosaurus, Plateosaurus, kiwi, seriema, adzebill (Aptornis), andsmaller ornithischians; a cluster composed of ornithomimosaurs,rheas, emu, cassowaries, and bustards; a cluster composed of Igua-nodon, Mantellisaurus, and hadrosaurids.
14 FARLOW ET AL.

Fig. 10. Relative digit lengths. (A) Length of digit II (excluding theungual), and the aggregate lengths of phalanges 1–3 of digit IV, aspercentages of the length of digit III (excluding the ungual), for allgroups (data set 1). Compared with the length of digit III, most ornithi-schian dinosaurs tend to have relatively longer digits II and IV than domost other forms. Troodontids, dromaeosaurids, and basal sauropo-domorphs, and to a lesser extent some struthioniforms (particularlyCasuarius), have a relatively long digit IV length compared with thelength of digit II. Poposaurus plots near the lower edge of points for
crocodylians, surrounded by points for non-avian theropods, kiwi, tin-amous, moa, and “hypsilophodontids.” (B) Digit II and digit IV lengths,both as percentages of the length of digit III (all including unguallengths), in non-avian dinosaurs, birds, Revueltosaurus, and Poposau-rus (data set 2). Most ornithischians are separated from other groupsby having relatively long digits IV and especially II. Poposaurus plotsamong points for large non-avian theropods, oviraptorosaurs, moa,the basal bird Confuciusornis, the quail Cyrtonyx, and basalornithopods.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 15

Fig. 11. Length of digit III as a function of metatarsal III (or tarsome-tatarsus) length. (A) Digit III length excluding the ungual; all groups(data set 1), forms comparable to, or smaller than, Poposaurus in size.Poposaurus continues a trend defined by crocodylians, in which digitIII is relatively long compared with the length of metatarsal III; basalsauropodomorphs (“prosauropods”), dromaeosaurids, and“hypsilophodontids” also fall along this trend. (B) Digit III length includ-ing the ungual; forms across the entire size range of dinosaurs, birds,Revueltosaurus, and Poposaurus (data set 2). Data are log-
transformed to spread out cases of smaller forms; no particular struc-tural relationship between the variable is assumed. Poposaurus is sim-ilar to the more basal theropods (Herrerasaurus, Dilophosaurus,Allosaurus, Saurophaganx), some non-avian coelurosaurs (Deinony-chus, Chirostenotes), “hypsilophodontids”, Tenontosaurus, Iguanodon(and Mantellisaurus), basal ceratopsians, kiwi, and moa other thanDinornis in having a relatively long digit III compared to the length ofmetatarsal III.
16 FARLOW ET AL.

basal ornithopods, in which the very large ungual on digitI (Figs. 2, 3, 8A) contributes to making the first digit pro-ject farther forward with respect to digits II and IV thanin Poposaurus.
Unlike many bipedal dinosaurs, in which the distalends of metatarsals II and IV bend away from the longaxis of digit III (e.g. Fig. 2L,M,P), in Poposaurus andother crocodile-line archosaurs metatarsals II and IV arestraight. Consequently any divarication between digitsII and IV in Poposaurus related to the metatarsalswould involve increasing distalward separation of themetatarsals along their lengths (e.g. Figs. 2I, 3), ratherthan the shapes of the metatarsals.
In crocodylians and some other crocodile-line archo-saurs (e.g. Fig. 2I, 3), digit IV is markedly slimmer thandigits I-III. Digit IV is also relatively more slender thanthe other toes in many dinosaurs, but not as much as in,for example, Alligator (Fig. 3). In this feature, Poposau-rus is distinctly more dinosaur-like than crocodylian-like.
A curious feature seen on the unguals of Poposaurus,and also on the unguals of some ornithopods (Fig.2P,R,S; also see Gilmore, 1915; Galton, 1974; Weisham-pel et al., 2003), basal ceratopsians (Fig. 2O) and thero-pods (Osborn, 1916; Shapiro et al., 2003; Longrich, 2008;Makovicky et al., 2004, 2010; Xu et al., 2011a; Cullenet al., 2013; White et al., 2013), is the presence of dis-tinct flanges or “wings” running longitudinally alongboth sides of the bottom of the ungual. Such wingsbecome exaggerated in large ornithopods (Fig. 2S), giv-ing unguals a rather arrowhead-like shape.
DISCUSSION
Limitations of data set 1 notwithstanding, some gen-eralizations about variability in pedal morphology acrossthe archosaurian groups in our sample can be made. InPCA, size alone accounts for more than three-quarters ofthe data variance (Table 4), leaving only modest shapedisparity after the effects of size are considered. Whatshape variability remains mostly involves the presenceand relative size of digit I, and the relative lengths ofmore proximal vs. more distal non-ungual phalanges(Figs. 5A, 6A). Although members of some clades tend togroup together, there is no obvious overall phylogeneticsignature. Thus large ornithopods, most ornithomimo-saurs, emu, cassowaries, rheas, bustards, and some neo-gnaths plot together, apart from other ornithischians,most non-avian theropods, many paleognaths and neo-gnaths, and crocodile-line archosaurs. Phalangeal pro-portions therefore seem phylogenetically labile.
In terms of the parameters examined using data set 1,and across all comparisons, Poposaurus has phalangealand digital proportions generally most like those of basalornithopods (“hypsilophodontids”), basal sauropodo-morphs (“prosauropods”), non-avian theropods, and evensome galliforms (Figs. 2, 5A, 6A, 10A). Poposaurusresembles crocodylians and some other crocodile-linearchosaurs, but also basal sauropodomorphs, basal orni-thopods, and dromaeosaurids, in the length of digit III(excluding the ungual) compared with the length ofmetatarsal III (Fig. 11A). If pedal features in addition tothose treated in data set 1 are added to considerations(Fig. 2), however, they enhance the dinosaurian gestaltof the foot of Poposaurus, making it distinctly similar to
that of most non-avian theropods, ground birds (particu-larly moa), and “hypsilophodontids.” The most dinosaur-like features of the foot of Poposaurus include a rela-tively short metatarsal I plus digit I (Fig. 2), a relativelyshort aggregate length of phalanges 1–3 of digit IV (com-pared with the length of digit III excluding the ungual;Fig. 10A) and a digit IV that is not particularly slim(Fig. 2).
Using data set 2, which adds the lengths of phalanxIV4 and the unguals, as well as digit widths, to the vari-ables analyzed in data set I, allows more detailed com-parisons of Poposaurus with various dinosaurian groups.In terms of the proportions of phalangeal lengths andwidths, Poposaurus is most like non-avian theropods(other than dromaeosaurids and ornithomimosaurs),moa, kiwi, and seriemas (Figs. 5B, 7, 9). Somewhat lessclosely, the phalangeal lengths and widths of Poposaurusresemble those of Revueltosaurus, Plateosaurus, someornithopods, and basal ceratopsians. In terms of the rel-ative lengths of digits II-IV (Fig. 10B), Poposaurusresembles non-avian theropods (other than dromaeo-saurids), small to medium-sized ornithopods, Confuciu-sornis, and some moa, struthioniforms, and galliforms.Poposaurus has a digit III length to metatarsal IIIlength ratio comparable to that of ornithischians andlarge theropods other than tyrannosauroids (Fig. 11B).Comparing the length of digit I with that of digit III,Poposaurus is most like a basal ornithopod (Fig. 8B).Thus while Poposaurus has a very dinosaur-like foot—itreally was a “dinocroc”—this foot is not exactly similarto any one dinosaurian clade, but rather shows featuresreminiscent of several dinosaurian groups.
Ichnological Considerations
Interpreting pedal morphology from fossil footprints isfraught with difficulties. Footprints are not merely cop-ies of feet, but instead record the dynamics of sediment/foot interactions (Allen, 1989, 1997; Platt and Meyer,1991; P�erez-Lorente, 1993; Avanzini, 1998; Gatesy et al.,1999; Nadon, 2001; Gatesy, 2003; Manning, 2004, 2008;Farlow et al., 2006; Mil�an and Bromley 2006, 2008;Mil�an et al., 2006; Graversen et al., 2007; Marty et al.,2009; Falkingham et al., 2010; Jackson et al., 2009,2010; Huerta et al., 2012; Platt et al., 2012; Carvalhaoet al., 2013; Castanera et al., 2013). Although two-dimensional images of the surface expression of foot-prints cannot capture their complete, three-dimensionalshape, careful outline drawings of well-preserved printsmay allow judicious reconstruction of the trackmaker’sfoot skeleton (Baird, 1957). Reversing the procedure, italso may be possible to offer hypotheses about what theshape of a particularly well-preserved, fleshed-out foot-print made by a skeletal taxon would have looked like.
The dinosaur-like character of the foot of Poposaurusraises the question of whether footprints made by Popo-saurus could be mistaken for those of basal dinosauro-morphs or even true dinosaurs. Unlike many bipedaldinosaurs, in which the distal ends of metatarsals II andIV bend away from the long axis of digit III (Fig. 2), inPoposaurus metatarsals II and IV are parallel to meta-tarsal III. In our reconstructions of the foot (Fig. 12D)and footprint (Fig. 13A–C) of Poposaurus in a “neutral”configuration, with joints between metatarsals and pha-langes, and between phalanges, directly aligned, the
POPOSAURUS GRACILIS PEDAL PROPORTIONS 17

metatarsophalangeal joints of digits II, III, and IV arenearly side by side, albeit with the metatarsophalangealjoint of digit III a little forward (distal) to those of digitsII and IV. There is very little divarication of the digits.
The pedal skeleton of Poposaurus is functionally tri-dactyl, with a long and slender overall shape. Digit III isthe longest, and digit IV is longer than II. In a digiti-grade and semi-digitigrade posture (Figs. 12A,B,D,E;13A,B), digit I (hallux) might have occasionally regis-tered in the imprint as a trace of the distal end (perhapsas the claw only); the metatarsophalangeal pad of digitIV might also have touched the ground. Digit V, pre-served in the skeleton by a strongly reduced metatarsal,would not have left an impression. In the alternative
plantigrade reconstruction (Figs. 12C, 13C), there mighthave been an elongate trace of metatarsal V at the pos-terolateral margin of the footprint. This might have var-ied among footprints along trackways due to thedifferences in gait, as well as under different substrateconditions and imprint depths (cf. Gatesy et al., 1999).
The digits of Poposaurus show little or no divaricationin “neutral” pose (Fig. 12D), but interdigital angle II-IVincreases to about 25 degrees if the digits are allowedmaximum splay against the distal ends of their respec-tive metatarsals (Fig. 12E). Still more divarication wouldhave been possible if the metatarsals themselves werenot closely appressed (Fig. 2D), but rather separated dis-tally by their enveloping soft tissues, as in Alligator
Fig. 12. Alternative digital reconstructions of the ankle and pedalconfiguration of Poposaurus. Left foot shown with a digitigrade stancein anteromedial (A) and lateral (B) views, and with a plantigrade stance(C) in lateral view. (D–E) Left foot shown with digitigrade stance in ven-
tral view. (D) Digits attached to their respective metatarsals in “neutral”articulation (centered on the distal ends of the metatarsals). (E) Digitswith maximum splay (interdigital angle). Anatomical abbreviations inTable 3.
18 FARLOW ET AL.

(Fig. 3). The proximal end of the digit IV impression in aPoposaurus footprint would have been lateral andslightly posterior to that of digit III. The elongate trian-gular unguals of digits I-IV in the Poposaurus pes skele-ton suggest digit impressions with tapering distal ends.Such a reconstructed footprint of Poposaurus can becompared with known Triassic archosaur footprints.
Chirotherians. The functionally tridactyl, mesax-onic pes imprint reconstructed for Poposaurus, withstrongly reduced and posteriorly shifted digits I and V,resembles chirotherian footprints of the ichnogeneraChirotherium, Sphingopus, and Parachirotherium fromthe Early–Middle Triassic (Fig. 13D–G). These havebeen interpreted by some authors as reflecting an evolu-tionary sequence and developments in the locomotorapparatus of crown-group archosaurs at the base of thedinosaur and bird line (Avemetatarsalia) (Haubold and
Klein, 2000, 2002). However, the makers of chirotheriantrackways from the Early-Middle Triassic were obligatequadrupeds and/or facultative bipeds, whereas the LateTriassic Poposaurus appears to have been an obligatebiped. This might indicate an evolutionary developmenttoward bipedalism in poposauroids between the Early–Middle and Late Triassic. The relatively small size ofchirotherian manus imprints relative to pes imprintshas been interpreted as an early tendency toward abipedal gait in archosaurs (Haubold, 1971) and matchesthis pattern very well.
Two ichnospecies of Chirotherium can be consideredas valid. Chirotherium barthii (Fig. 13D) has a nearlysymmetrical digit group II–IV, a reduced digit I, and athumblike and backward curved digit V with a well-defined non-ungual portion (Haubold, 1971, 2006; Kleinet al., 2011). The distinct digit V impression distin-guishes Chirotherium barthii from the reconstructedPoposaurus footprint. The smaller Chirotherium sickleri(Fig. 13E) has a less symmetrical digit group II–IV. DigitIII is longest, digit IV is distinctly longer than digit II,and digit I is strongly reduced. The posterior margin ofthe digit group I–IV is nearly straight compared with C.barthii, where it is more strongly curved (Haubold, 1971).In these features, C. sickleri approximates the recon-structed Poposaurus footprint in its semi-digitigrade con-figuration (compare Fig. 13B and E), although C. sickleridiffers from the reconstructed Poposaurus print in havinga recurved digit V impression with a non-ungual portion.Interestingly, C. sickleri has been attributed to the popo-sauroid Ctenosauriscus by Ebel et al. (1998).The ichnoge-nus Sphingopus (Fig. 13F) has a slender, elongate shape,and digits I–IV are subparallel, as in the reconstructedPoposaurus footprint, whereas in the ichnogenus Para-chirotherium (Fig. 13G) the digits are more divergent. InSphingopus the pedal digit I impression is often reducedto a trace of the claw tip (Fig. 13F). C. barthii, Sphingo-pus, and Parachirotherium show a posteriorly concaveand narrow proximal margin of the digit group I–IV. Thisindicates a narrow pes with closely appressed and over-lapping metatarsals, characteristic of Avemetatarsalia (orOrnithodira; Gauthier, 1986). In Sphingopus and C. bar-thii the proximal end of the digit IV impression is nearlyintersected by a line through the long axis of the digit IIIimpression. In contrast, the proximal end of the digit IVimpression in the reconstructed footprint of Poposaurus islateral to the long axis of digit III (Fig. 13A–C), as in Chi-rotherium sickleri.
Dinosauromorph and dinosaur footprints.Footprints of different morphologies from the Early-Middle Triassic have recently been assigned to dinosaur-omorphs (Brusatte et al., 2011b; Nied�zwiedzki et al.,2013), although this interpretation is still debated. Typi-cal footprints attributed to dinosauromorphs and truedinosaurs reflect a functionally tridactyl, symmetricalpes (Carrano and Wilson, 2001; Marsicano et al., 2007),with digit III being longest. This feature can be seen inichnotaxa such as Atreipus, Grallator, and Eubrontesfrom the Middle–Late Triassic and Early Jurassic (Hau-bold and Klein, 2002; Klein et al., 2006, 2011; Lagnaouiet al. 2012; Fig. 13H–J; Fig. 14B–K,M,N). Apart fromthe associated manus imprint of Atreipus (and thereforethe quadrupedalism of its maker), some of these
Fig. 13. Comparison of footprints reconstructed from the pes skele-ton of Poposaurus gracilis (A–C) with known archosaur footprints fromthe Early to Middle Triassic. Dotted portions at the back of the foot-print outline in panel A (digit I; metatarsophalangeal region and mostof phalanx 1 of digit IV) indicate places where whether the foot wouldroutinely have touched the ground to leave a mark is uncertain. (D)Chirotherium barthii (Middle Triassic, Anisian, Germany). (E) C. sickleri(Early Triassic, Olenekian, USA). (F) Sphingopus and (G) Parachirothe-rium (Middle Triassic, Anisian–Ladinian, France, Germany). (H–J) Atrei-pus – Grallator (“Coelurosaurichnus”, “Anchisauripus”) (Middle Triassic,Anisian–Ladinian, Germany, Morocco, France). D–F after Haubold(1971), Haubold and Klein (2002). G–H after Haubold and Klein (2000).I after Klein et al. (2011). J after Gand and Demathieu (2005).
POPOSAURUS GRACILIS PEDAL PROPORTIONS 19

ichnotaxa show similarities with the digitigrade foot-print reconstruction of Poposaurus (Fig. 14A). The pedalportion of Atreipus (Figs. 13H, 14B–D), like our recon-struction of a digitigrade footprint of Poposaurus, hasdigit impressions with a rather low divarication, and therelative positions of the proximal ends of the digitimpressions are also sometimes similar. The possibilitythat at least some stem-group crocodylians, due to con-vergent evolution in the pes, left dinosaur-like foot-prints, therefore cannot be excluded.
The chief differences in footprint shapes between ourreconstruction for Poposaurus and many ichnotaxaattributed to dinosauromorphs relate to the greater digi-tal divarication in most dinosauromorph ichnotaxa, andthe possibly related tendency for the proximal end of thedigit IV impression to be aligned with the long axis ofdigit III, as also seen in some chirotherians (Fig. 13D,F).
If the digit divarication in Poposaurus was in fact as lowas in our neutral digitigrade reconstruction, this woulddiffer from what is seen in typical dinosauromorph foot-prints (compare Fig. 14A with 14B–K,M,N).
Poposaurus and other crurotarsans have a lower ratioof metatarsal III length to femur length than do earlyMesozoic bipedal dinosaurs and other ornithodirans (Kuboand Kubo, 2012). Our data show that Poposaurus andother crocodile-line archosaurs likewise have a relativelyshort metatarsal III compared with the length of digit III(Fig. 11). Both of these relationships presumably contrib-ute to the lower stride/footprint length ratios seen intrackways attributed to non-dinosauromorph archosaursthan in trackways attributed to dinosauromorphs (Kuboand Kubo, 2013). Even if Poposaurus had a digitigradepedal carriage, and left very dinosaur-like footprints, it islikely that those prints would occur in trackways withshorter step lengths compared with footprint lengths thanseen in trackways of bipedal dinosaurs.
Evazoum. Tridactyl or tetradactyl pes imprints(Fig. 14L) of a biped or facultative biped have beendescribed from the Late Triassic of Italy and NorthAmerica under Evazoum (Nicosia and Loi, 2003; Lockleyet al., 2006), although these prints are considered to beincomplete Brachychirotherium by some authors (seeKlein et al., 2006; Lucas et al., 2010). They have beenassigned to prosauropods (Lockley et al., 2006) and popo-sauroids (D’Orazi-Porchetti et al., 2008). Compared withour Poposaurus footprint reconstruction, the digits ofEvazoum show different proportions, with digit III pro-jecting only a bit farther forward beyond digits II andIV, whereas in Poposaurus digit III projects substantiallyfarther forward. In Evazoum digit I is relatively longcompared with the condition in Poposaurus, where it isstrongly reduced.
Evazoum, like some chirotherians and many dinosaur-omorph prints, also differs from our reconstructed Popo-saurus footprint (neutral configuration) by having agreater digital divarication, and alignment of an elonga-tion of the proximal end of the footprint with the longaxis of digit III, giving the print a distinct “heel.”Although it is possible that this is a real difference, keepin mind that our reconstructed Poposaurus footprint isbased on neutral matching of joint surfaces. Under load-ing of the animal’s weight, or due to the presence ofenveloping tissues, it is possible that the footprintsactually created would show more splay than in our neu-tral reconstruction (Fig. 12E).
Undetermined archosaur trackway. An archo-saur trackway of uncertain affinity was recentlydescribed from Upper Triassic (Rhaetian) deposits ofSichuan, China (Xing et al., in press). It consists of 19deeply impressed plantigrade pes imprints lacking anassociated trace of the manus. Although it cannot becompletely excluded that the manus footprints wereoverprinted by the pes, the trackway could have beenleft by a biped, such as a sauropodomorph or a stem-group crocodilian such as Poposaurus. However, thepreservation of the imprints with only two digit traces(the outermost being longest) is unusual in deep foot-prints—such as those of theropods, in which three digit
Fig. 14. Digitigrade reconstruction of Poposaurus gracilis footprint inneutral pose (A) compared with footprints attributed to Dinosauromorphaand Dinosauria (B–N). (B–D) Atreipus, including [C]”Coelurosaurichnus”(Late Triassic, Carnian–Norian, USA, France, Morocco). (E–H) Grallator(Late Triassic, Carnian–Norian, Morocco, Germany, USA). (I–J) Eubrontes(Late Triassic, Norian–Rhaetian, Sweden, southern Africa. (K) Pengxian-pus (Late Triassic, Rhaetian, China). (L) Evazoum (Late Triassic, Carnian,Italy). (M–N) Eubrontes, Grallator (Early Jurassic, USA). B–C, F, H–J, M–Nafter Haubold and Klein (2000), Klein and Haubold (2007). K after Xinget al. (2013). D–E after Lagnaoui et al. (2012). G after Lucas et al. (2010).L after Nicosia and Loi (2003).
20 FARLOW ET AL.

impressions are usually preserved (Kuban, 1989; Xinget al., in press).
Ichnological summary. Our reconstruction of thefootprint of Poposaurus gracilis shows some similaritieswith Early–Middle Triassic archosaurian ichnotaxa,such as the chirotherians Chirotherium sickleri andSphingopus. Nevertheless, such features as the distinctimpression of digit V and the quadrupedal gait of chiro-therian makers differ from our reconstruction for Popo-saurus. However, this may only reflect the differentstratigraphic ages and evolutionary developments withinPoposauroidea, between Poposaurus of the Late Triassicand Early–Middle Triassic forms like Arizonasaurus,Ctenosauriscus, and Xilousaurus (Ebel et al., 1998; Nes-bitt, 2003; Sennikov, 2012). Unfortunately, no pedal skel-etons of these taxa are preserved, making a satisfyingcorrelation with chirotherian footprints impossible fornow.
With a digit III length of about 16 cm, Poposaurusgracilis may have been comparable to a small to mid-range theropod in overall body size (somewhere betweenthe makers of Anchisauripus sillimani and A. minuscu-lus, in the terminology of Lull [1953]). The dinosaur-likepedal proportions of Poposaurus, and the similarity ofits reconstructed footprint to those of some dinosauro-morphs, suggest that some grallatorid forms could wellhave been made by Poposaurus and its close relatives.However, mistaking Poposaurus tracks for dinosaur(particularly theropod) tracks would be less likely tooccur if digit I of Poposaurus routinely touched theground. Furthermore, trackways made by Poposauruswould probably have a shorter stride/footprint lengthratio than grallatorid trackways.
Convergence in foot (and possibly footprint) shapebetween Poposaurus and bipedal dinosaurs is not aunique occurrence, judging from results of our analysesacross archosaurs more generally (Figs. 2, 9). Feet oflarge non-avian theropods seem to be very similar,whether those large theropods are abelisaurs, allosaurs,or tyrannosaurs (Farlow, 2001; Farlow et al., 2012,2013). The same is true for the feet of Plateosaurus,Tenontosaurus, Leptoceratops, and perhaps even Revuel-tosaurus, between the feet of bustards and tridactylstruthioniforms (cf. Farlow et al. 2012), and to a lesserextent among feet of kiwi, moa, and large theropods.Consequently, the broader message of our study for arch-osaur ichnology is that similarity in footprint shapesometimes may be an unreliable proxy for phylogeneticcloseness.
ACKNOWLEDEGMENTS
Marilyn Fox skilfully prepared the specimen of Poposau-rus. We thank J. Gauthier, C. Norris, W. Joyce, and D.Brinkman for access to the Poposaurus specimen. Col-leagues and institutions around the world providedaccess to herpetological, ornithological, and paleontologi-cal specimens; William Parker in particular supplied alovely cast of the foot of Revueltosaurus. Karl Bates andAndrew Heckert provided useful discussion. Farlowreceived the National Science Foundation grant(9612880-EAR) and Dinosaur Society grant, and Curriereceived the NSERC 203091-06 grant; Schachner is anAmerican Association of Anatomists Scholar, and this
research was in part funded by the American Associa-tion of Anatomists.
LITERATURE CITED
Allen JRL. 1989. Fossil vertebrate tracks and indenter mechanics. JGeol Soc London 146:600–602.
Allen JRL. 1997. Subfossil mammalian tracks (Flandrian) in theSevern Estuary, S.W. Britain: mechanics of formation, preserva-tion and distribution. Phil Trans Roy Soc London B 352:481–518.
Avanzini M. 1998. Anatomy of a footprint: bioturbation as a key tounderstanding dinosaur walking dynamics. Ichnos 6:129–139.
Baird, D. 1957. Triassic reptile footprint faunules from Milford,New Jersey. Bull Mus Comp Zool, Harvard Univ 117:449–520.
Bates KT, Schachner ER. 2012. Disparity and convergence inbipedal archosaur locomotion. J Roy Soc Interface 9:1339–1353.
B�elen Von Baczko M, Ezcurra MD. 2013. Ornithosuchidae: a group ofTriassic archosaurs with a unique ankle joint. In: Nesbitt SJ, DesojoJB, Irmis RB, editors. Phylogeny and palaeobiology of early archo-saurs and their kin. Geol Soc London Spec Pub 379. p 187–302.
Benton MJ. 1999 Scleromochlus taylori and the origin of dinosaursand pterosaurs. Phil Trans Roy Soc London B 354:1423–1446.
Benton MJ. 2012. Origin and early evolution of dinosaurs. In:Brett-Surman M, Holtz, TR, Jr., Farlow JO, editors. The completedinosaur (second edition). Bloomington: Indiana University Press.p 331–345.
Bourdon E, de Ricqles A, Cubo J. 2009. A new transantarctic rela-tionship: morphological evidence for a Rheidae-Dromaiidae-Casuariidae clade (Aves, Palaeognathae, Ratitae). Zool J Linn Soc156:641–663.
Boyd CA, Brown CM, Scheetz RD, Clarke JA. 2009. Taxonomic revi-sion of the basal neornithischian taxa Thescelosaurus and Buge-nasaura. J Vert Paleontol 29:758–770.
Brusatte SL. 2012. Dinosaur paleobiology. West Sussex: Wiley-Blackwell.
Brusatte SL, Benton MJ, Lloyd GT, Ruta M, Wange SC. 2011a.Macroevolutionary patterns in the evolutionary radiation of arch-osaurs (Tetrapoda: Diapsida). Earth Environ Sci Trans Roy SocEdinburgh 101:367–382.
Brusatte SL, Nied�zwiedzki G, Butler RJ. 2011b. Footprints pull ori-gin and diversification of dinosaur stem-lineage deep into EarlyTriassic. Proc Roy Soc London B 278:1107–1113.
Butler RJ. 2010. The anatomy of the basal ornithischian dinosaurEocursor parvus from the lower Elliot Formation (Late Triassic)of South Africa. Zool J Linn Soc 160: 648–684.
Butler RJ, Smith RMH, Norman DB. 2007. A primitive ornithi-schian dinosaur from the Late Triassic of South Africa, and theearly evolution and diversification of Ornithischia. Proc Roy SocB 274: 2041–2046.
Butler RJ, Brusatte SL, Reich M, Nesbitt SJ, Schoch RR, HornungJJ. 2011. The sail-backed reptile Ctenosauriscus from the latestEarly Triassic of Germany and the timing and biogeography ofthe early archosaur radiation. PLoS One 6:e25693. doi: 10.1371/journal.pone.0025693.
Carrano MT, Wilson JA. 2001. Taxon distributions and the tetrapodtrack record. Paleobiology 27:564–582.
Carvalhao I de S, Borghi L, Leonardi G. 2013. Preservation of dino-saur tracks induced by microbial mats in the Sousa Basin (LowerCretaceous), Brazil. Cret Res 44:112–121.
Castanera D, Vila B, Razzolini NL, Falkingham PL, Canudo JI,Manning PL, Galobart A. 2013. Manus track preservation bias asa key factor for assessing trackmaker identity and quadrupedal-ism in basal ornithopods. PLoS One 8:e54177. doi: 10.1371/journal.pone.0054177.
Clements JF. 2007. The Clements checklist of birds of the world,sixth edition. Ithaca, New York: Clements Publishing Associates.
Colbert EH, Mook CC. 1951. The ancestral crocodile Protosuchus.Bull Amer Mus Nat Hist 97:143–182.
Cruickshank ARI. 1972. The proterosuchian thecodonts. In: JoyseyKA, Kemp TS, editors. Studies in vertebrate evolution. Edin-burgh: Oliver and Boyd. p 89–119.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 21

Crush PJ. 1984. A late Triassic sphenosuchid crocodilian fromWales. Palaeontology 27:131–157.
Cullen TM, Ryan MJ, Schr€oder-Adams C, Currie PJ, Kobayashi Y.2013. An ornithomimid (Dinosauria) bonebed from the Late Cre-taceous of Alberta, with implications for the behavior, classifica-tion, and stratigraphy of North American ornithomimids. PLoSOne 8:e58853.doi:10.1371/journal.pone.0058853.
Desojo JB, B�aez AM. 2005. El esqueleto postcraneano de Neoaeto-sauroides (Archosauria: Aetosauria) del Tri�asico Superior delcentro-oeste de Argentina. Ameghiniana 42:115–126.
Desojo JB, Ezcurra MD, Schultz CL. 2011. An unusual new archo-sauriform from the Middle-Late Triassic of southern Brazil andthe monophyly of Doswelliidae. Zool J Linn Soc 161:839–871.
Desojo JB, Heckert AB, Martz JW, Parker WG, Schoch RR, SmallBJ, Sulej T. 2013. Aetosauria: a clade of armoured pseudosu-chians from the Upper Triassic continental beds. In: Nesbitt SJ,Desojo JB, Irmis RB, editors. Phylogeny and palaeobiology ofearly archosaurs and their kin. Geol Soc London Spec Pub 379. p203–239.
D’Orazi-Porchetti S, Nicosia U, Mietto P, Petti FM, Avanzini M.2008. Atreipus-like footprints and their co-occurrence with Eva-zoum from the upper Carnian (Tuvalian) of Trentino-Alto Adige.Studi Trentini Scien Naturali, Acta Geol 83:277–287.
Dzik J. 2003. A beaked herbivorous archosaur with dinosaur affin-ities from the early Late Triassic of Poland. J Vert Paleontol 23:556–574.
Ebel K, Falkenstein F, Haderer, F-O, Wild R. 1998. Ctenosauriscuskoeneni (v. Huene) und der Rauisuchier von Waldshut – Biome-chanische Deutung der Wirbels€aule und Beziehungen zu Chiro-therium sickleri Kaup. Stuttr Beitr Naturk B 261:1–18.
Ewer RF. 1965. The anatomy of the thecodont reptile Euparkeriacapensis Broom. Phil Trans Roy Soc London B 248:379–435; 8:371–425.
Ezcurra MD. 2010. A new early dinosaur (Saurischia: Sauropodo-morpha) from the Late Triassic of Argentina: a reassessment ofdinosaur origins and phylogeny. J Syst Paleontol 8:371–425.
Ezcurra MD, Butler RJ, Gower DJ. 2013. ‘Proterosuchia’: the ori-gin and early history of Archosauriformes. In: Nesbitt SJ,Desojo JB, Irmis RB, editors. Phylogeny and palaeobiology ofearly archosaurs and their kin. Geol Soc London Spec Pub.379. p 9–33.
Falkingham PL, Margetts L, Manning PL. 2010. Fossil vertebratetracks as paleopenetrometers: confounding effects of foot morphol-ogy. Palaios 25:356–360.
Farlow JO. 2001. Acrocanthosaurus and the maker of Comancheanlarge-theropod footprints. In: Tanke DH, Carpenter C, editors.Mesozoic vertebrate life: new research inspired by the paleontol-ogy of Philip J. Currie. Bloomington: Indiana University Press. p408–427.
Farlow JO, Chapman RE, Breithaupt B, Matthews N. 2012. Thescientific study of dinosaur footprints. In: Brett-Surman M, Holtz,TR, Jr., Farlow JO, editors. The complete dinosaur (second edi-tion). Bloomington: Indiana University Press. p 713–759.
Farlow JO, Holtz, TR, Jr., Worthy TH, Chapman RE, 2013. Feet ofthe fierce (and not so fierce): pedal proportions in large theropods,other non-avian dinosaurs, and large ground birds. In: ParrishJM, Molnar RE, Currie PJ, Koppelhus EB, editors. Tyrannosauridpaleobiology. Bloomington: Indiana University Press. p 88–132.
Farlow JO, Langston W Jr., Deschner EE, Solis R, Ward W,Kirkland BL, Hovorka S, Reece TL, Whitcraft J. 2006. Texasgiants: dinosaurs of the Heritage Museum of the Texas Hill Coun-try. Canyon Lake, Texas: Heritage Museum of the Texas HillCountry.
Farlow JO, Lockley MG. 1993. An osteometric approach to the iden-tification of the makers of early Mesozoic tridactyl dinosaur foot-prints. In: Lucas SG, Morales M, editors. The nonmarine Triassic.Albuquerque: New Mexico Mus Nat Hist Sci Bull 3. p 123–131.
Franca MAG, Ferigolo J, Langer MC. 2011. Associated skeletons ofa new middle Triassic “Rauisuchia” from Brazil. Naturwissen-schaften 98:389–395.
Fraser N, Henderson D. 2006. Dawn of the dinosaurs: life in theTriassic. Bloomington: Indiana University Press.
Galton PM. 1974. The ornithischian dinosaur Hypsilophodon fromthe Wealden of the Isle of Wight. Bull Brit Mus (Nat Hist) Geol25: 1–152c.
Galton PM. 2012. Notes on bones and teeth of Bromsgroveia andother archosaurian reptiles from the lower Middle Triassic (Ani-sian) of the English Midlands. Rev Pal�eobiol, Geneve 31:171–204.
Gand G, Demathieu G. 2005. Les pistes dinosauroides du Triasmoyen francais: interpr�etation et r�e�evaluation de la nomencla-ture. Geobios 38:725–749.
Gatesy SM. 2003. Direct and indirect features: what sediment did adinosaur touch? Ichnos 10:91–98.
Gatesy SM, Middleton KM, Jenkins FA Jr., Shubin NH. 1999.Three-dimensional preservation of foot movements in Triassictheropod dinosaurs. Nature 399:141–144.
Gauthier JA. 1986. Saurischian monophyly and the origin of birds.Mem Calif Acad Sci 8:1–55.
Gauthier JA, Nesbitt SJ, Schachner ER, Bever GS, Joyce WG. 2011.The bipedal stem crocodilian Poposaurus gracilis: inferring func-tion in fossils and innovation in archosaur locomotion. Bull Pea-body Mus Nat Hist 52:107–126.
Gilmore CW. 1915. Osteology of Thescelosaurus, an ornithopodousdinosaur from the Lance Formation of Wyoming. Proc USNM 49:591–616.
Graversen O, Mil�an J, Loope DB. 2007. Dinosaur tectonics: a struc-tural analysis of theropod undertracks with a reconstruction oftheropod walking dynamics. J Geol 115:641–654.
Harshman J, Braun EL, Braun MJ, Huddleston CJ, Bowie RCK,Chojnowski JL, Hackett SJ, Han K-L, Kimball RT, Marks BD,Miglia KJ, Moore WS, Reddy S, Sheldon FH, Steadman DW,Steppan SJ, Witt CC, Yuri T. 2008. Phylogenomic evidence formultiple losses of flight in ratite birds. Proc Nat Acad Sci 105:13462–13467.
Haubold H. 1971. Die Tetrapodenf€ahrten des Buntsandsteins.Pal€aontol Abh A 4:395–548.
Haubold H. 2006. Die Saurierf€ahrten Chirotherium barthii Kaup,1835 – das Typusmaterial aus dem Buntsandstein bei Hildbur-ghausen/Th€uringen und das Chirotherium-Monument. Ver€offNaturhist Mus Schleusingen 21:3–31.
Haubold H, Klein H. 2000. Die dinosauroiden F€ahrten Parachiro-therium – Atreipus – Grallator aus dem unteren Mittelkeuper(Obere Trias: Ladin, Karn, ?Nor) in Franken. Hall Jahrb GeowissB 22:59–85.
Haubold H, Klein, H. 2002. Chirotherien und Grallatoriden aus derUnteren bis Oberen Trias Mitteleuropas und die Entstehung derDinosauria. Hall Jahrb Geowiss B 24:1–22.
Heckert AB, Lucas SG (eds). 2002. Upper Triassic stratigraphy andpaleontology. Albuquerque: New Mexico Museum of Natural His-tory and Science Bulletin, Publication 21.
Heckert AB, Lucas SG, Rinehart LF, Celeskey MD, Spielmann JA,Hunt AP. 2010. Articulated skeletons of the aetosaur Typothoraxcoccinarum Cope (Archosauria: Stagonolepididae) from the UpperTriassic Bull Canyon Formation (Revueltian: early-mid Norian),eastern New Mexico, USA. J Vert Paleontol 30:619–642.
Huerta P, Fern�andez-Baldor FT, Farlow JO, Montero D. 2012.Exceptional preservation processes of 3D dinosaur footprint castsin Costalomo (Lower Cretaceous, Cameros Basin, Spain). TerraNova 24:136–141.
Irmis RB. 2011. Evaluating hypotheses for the early diversificationof dinosaurs. Earth Environ Sci Trans Roy Soc Edinburgh 101:397–426.
Irmis RB, Nesbitt SJ, Sues H.-D. 2013. Early Crocodylomorpha. In:Nesbitt SJ, Desojo JB, Irmis RB, editors. Phylogeny and palaeobi-ology of early archosaurs and their kin. Geol Soc London SpecPub 379. p. 275–302.
Jackson SJ, Whyte MA, Romano M. 2009. Laboratory-controlledsimulations of dinosaur footprints in sand: a key to understand-ing vertebrate track formation and preservation. Palaios 24:222–238.
Jackson SJ, Whyte MA, Romano M. 2010. Range of experimentaldinosaur (Hypsilophodon foxii) footprints due to variation in sandconsistency: how wet was the track? Ichnos 17:197–214.
22 FARLOW ET AL.

Jalil N-E, Peyer K. 2007. A new rauisuchian (Archosauria, Suchia)from the Upper Triassic of the Argana Basin, Morocco. Palaeontol50:417–430.
Kammerer CF, Nesbitt SJ, Shubin NH. 2012. The first silesauriddinosauriform from the Late Triassic of Morocco. Acta PalaeontolPol 57:277–284.
Klein H, Haubold H. 2007. Archosaur footprints—potential for bio-chronology of Triassic continental sequences. In: Lucas SG, Spiel-mann JA, editors. The Global Triassic. Albuquerque: New MexicoMuseum Natural History and Science Bulletin, Publication 41. p120–130.
Klein H, Lucas SG, Haubold H. 2006. Tetrapod track assemblage ofthe Redonda Formation (Upper Triassic, Chinle Group) in east-central New Mexico – re-evaluation of ichnofaunal diversity fromstudies of new material. In: Harris JD, Lucas SG, Spielmann JA,Lockley MG, Milner AC, Kirkland JI, editors. The Triassic-Jurassic terrestrial transition. Albuquerque: New MexicoMuseum Natural History and Science Bulletin, Publication 37. p.241–250.
Klein H, Voigt S, Saber H, Schneider JW, Hminna A, Fischer J,Lagnaoui A, Brosig A. 2011. First occurrence of a Middle Triassictetrapod ichnofauna from the Argana Basin (Western High Atlas,Morocco). Palaeogeogr Palaeoclimatol Palaeoecol 307:218–231.
Krebs B. 1965. Die Triasfauna der Tessiner Kalkalpen. XIX. Ticino-suchus ferox, nov. gen. nov. sp. Ein neuer Pseudosuchier aus derTrias des Monte San Georgio. Schweiz Pal€aontol, Abh 81:1–140.
Kuban G. 1989. Elongate dinosaur tracks. In: Gillette DD, LockleyMG, editors. Dinosaur tracks and traces. Cambridge: CambridgeUniversity Press. p. 57–92.
Kubo T, Benton MJ. 2007. Evolution of hindlimb posture in archo-saurs: Limb stresses in extinct vertebrates. Palaeontology 50:1519–1529.
Kubo T, Benton MJ. 2009. Tetrapod postural shift estimated fromPermian and Triassic trackways. Palaeontology 52:1029–1037.
Kubo T, Kubo M. 2012. Associated evolution of bipedality and cur-soriality among Triassic archosaurs: a phylogenetically controlledevaluation. Paleobiol 38:474–485.
Kubo T, Kubo M. 2013. Analysis of Triassic archosauriform track-ways: difference in stride/footprint ratio between dinosauro-morphs and other archosauriforms. Palaios 28:259–265.
Lagnaoui A, Klein H, Voigt S, Hminna A, Saber H, Schneider JW,Werneburg R. 2012. Late Triassic tetrapod-dominated ichnoas-semblages from the Argana Basin (Western High Atlas, Morocco).Ichnos 19:238–253.
Langer, M. 2003. The pelvic and hind limb anatomy of the stem-sauropodomorph Saturnalia tupiniquim (Late Triassic, Brazil).PaleoBios 23:1–40.
Langer MC, Nesbitt SJ, Bittencourt JS, Irmis RB. 2013. Non-dino-saurian dinosauromorphs. In: Nesbitt SJ, Desojo JB, Irmis RB,editors. Phylogeny and palaeobiology of early archosaurs andtheir kin. Geological Society of London Special Publication 379. p157–186.
Lecuona A, Desojo JB. 2011. Hind limb osteology of Gracilisuchusstipanicicorum (Archosauria: Pseudosuchia). Earth Environ SciTrans Roy Soc Edinburgh 102:105–128.
Lee MSJ, Worthy TH. 2012. Likelihood reinstates Archaeopteryx asa primitive bird. Biol Letters 8:295–303.
Lee Y-N, Ryan MJ, Kobayashi Y. 2011. The first ceratopsian dino-saur from South Korea. Naturwissenschaften 98:39–49.
Li C, Wu X-C, Cheng Y-N, Sato T, Wang L. 2006. An unusual archo-saurian from the marine Triassic of China. Naturwissenschaften93:200–206.
Li C, Wu X-C, Zhao L-J, Sato T, Wang L-T. 2012. A new archosaur(Diapsida, Archosauriformes) from the marine Triassic of China.J Vert Paleo 32:1064–1081.
Livezey BC, Zusi R. 2007. Higher-order phylogeny of modern birds(Theropods, Aves: Neornithes) based on comparative anatomy. II.Analysis and discussion. Zool J Linn Soc 149:1–95.
Lockley MG, Lucas SG, Hunt AP. 2006. Evazoum and the renamingof northern hemisphere “Pseudotetrasauropus”: implications fortetrapod ichnotaxonomy at the Triassic-Jurassic boundary. In:Harris JD, Lucas SG, Spielmann JA, Lockley MG, Milner AC,
Kirkland JI, editors. The Triassic-Jurassic terrestrial transition.New Mexico Museum Natural History and Science Bulletin, Pub-lication 37. p 199–206.
Longrich N. 2008. A new, large ornithomimid from the CretaceousDinosaur Park Formation of Alberta, Canada: implications forthe study of dissociated dinosaur remains. Palaeontology 51:983–997.
Lucas SG, Spielmann JA, Klein H, Lerner AJ. 2010. Ichnology ofthe Upper Triassic Redonda Formation (Apachean) in east-centralNew Mexico. Albuquerque: New Mexico Museum Natural Historyand Science Bulletin, Publication 47.
Lull RS. 1953. Triassic life of the Connecticut Valley. Hartford: Con-necticut State Geological and Natural History Survey Bulletin,Publication 81.
Makovicky PJ, Kobayashi Y, Currie PJ. 2004. Ornithomimosauria.In: Weishampel DB, Dodson P, Osm�olska H. The Dinosauria.Berkeley: University of California Press. p 137–150.
Makovicky PJ, Li D, Gao K-Q, Lewin M, Erickson GM, Norell MA.2010. A giant ornithomimosaur from the Early Cretaceous ofChina. Proc Royal Soc B 277:191–198.
Mallison H. 2011. Plateosaurus in 3D: how CAD models andkinetic-dynamic modelling bring an extinct animal to life. In:Klein N, Remes K, Gee CT, Sander PM, editors. Biology of thesauropod dinosaurs: understanding the life of giants. Blooming-ton: Indiana University Press. p 219–236.
Manning PL. 2004. A new approach to the analysis and interpreta-tion of tracks: examples from the dinosauria. Geol Soc LondonSpec Pub 228:93–123.
Manning PL. 2008. T. rex speed trap. In: Larson P, Carpenter K,editors. Tyrannosaurus rex, the tyrant king. Bloomington: IndianaUniversity Press. p 204–231.
Marsicano CA, Domnanovich NS, Mancuso AC. 2007. Dinosaur ori-gins: evidence from the footprint record. Hist Biol 19:83–91.
Martinez RN, Alcober OA. 2009. A basal sauropodomorph (Dinosau-ria: Saurischia) from the Ischigualasto Formation (Triassic, Car-nian) and the early evolution of sauropodomorpha. PLoS One4(2):e4397.doi:10.1371/journal.pone.0004397.
Marty D, Strasser A, Meyer CA. 2009. Formation and taphonomy ofhuman footprints in microbial mats of present-day tidal-flat envi-ronments: implications for the study of fossil footprints. Ichnos16:127–142.
Mil�an J, Avanzini M, Clemmensen B, Garc�ıa-Ramos, Pi~nuela L.2006. Theropod foot movement recorded by Late Triassic, EarlyJurassic and Late Jurassic fossil footprints. In: Harris JD, LucasSG, Spielmann JA, Lockley MG, Milner AC, Kirkland JI, editors.The Triassic-Jurassic terrestrial transition. Albuquerque: NewMexico Museum of Natural History and Science Bulletin, Publica-tion 37. p 352–364.
Mil�an J, Bromley RG. 2006. True tracks, undertracks anderoded tracks, experimental work with tetrapod tracks in fieldand laboratory. Palaeogeogr Palaeoclimatol Palaeoecol 231:253–264.
Mil�an J, Bromley RG. 2008. The impact of sediment consistency ontrack and undertrack morphology: experiments with emu tracksin layered cement. Ichnos 15:18–24.
Nadon GC. 2001. The impact of sedimentology on vertebrate trackstudies. In: Tanke DH, Carpenter K, editors. Mesozoic vertebratelife. Bloomington: Indiana University Press. p 395–407.
Moreno K, Carrano MT, Snyder R. 2007. Morphological changes inpedal phalanges through ornithopod dinosaur evolution: a biome-chanical approach. J Morph 268:50–63.
Nesbitt SJ. 2003. Arizonasaurus and its implications for archosaurdivergence. Proc Roy Soc B 70:S234–S237.
Nesbitt S. 2007. The anatomy of Effigia okeeffeae (Archosauria:Suchia), theropod-like convergence, and the distribution of relatedtaxa. Bull Amer Mus Nat Hist 302:84.
Nesbitt S. 2011. The early evolution of archosaurs: relationshipsand the origin of major clades. Bull Amer Mus Nat Hist 352:1–292.
Nesbitt SJ, Norell MA. 2006. Extreme convergence in the bodyplans of an early suchian (Archosauria) and ornithomimid dino-saurs (Theropoda). Proc Roy Soc London B 273:1045–1048.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 23

Nesbitt SJ, Irmis RB, Parker WG. 2007. A critical re-evaluation ofthe Late Triassic dinosaur taxa of North America. J Syst Paleon-tol 5:209–243.
Nesbitt SJ, Barrett PM, Werning S, Sidor CA, Charig AJ. 2013a. Theoldest dinosaur? A Middle Triassic dinosauriform from Tanzania.Biol Letters 9:20120949. http://dx.doi.org/10.1098/rsbl.2012.0949.
Nesbitt SJ, Irmis RB, Parker WG, Smith ND, Turner AH, Rowe T.2009a. Hindlimb osteology and distribution of basal dinosauro-morphs from the Late Triassic of North America. J Vert Paleo 29:498–516.
Nesbitt SJ, Sidor CA, Irmis RB, Angelczyk KD, Smith RMH, TsujiLA. 2010. Ecologically distinct dinosaurian sister group showsearly diversification of Ornithodira. Nature 464:95–98.
Nesbitt SJ, Smith ND, Irmis RB, Turner AH, Downs A., Norell MA.2009b. A complete skeleton of a Triassic saurischian and the earlyevolution of dinosaurs. Science 326:1530–1533.
Nesbitt SJ, Brusatte SL, Desojo JB, Liparini A, De Franca MAG,Weinbaum JC, Gower DJ. 2013b. Rauisuchia. In: Nesbitt SJ, Des-ojo JB, Irmis RB, editors. Phylogeny and palaeobiology of earlyarchosaurs and their kin. Geol Soc London Spec Pub 379:241–274.
Nicosia U, Loi M. 2003. Triassic footprints from Lerici (La Spezia,northern Italy). Ichnos 10:127–140.
Nied�zwiedzki G, Brusatte SL, Butler RJ. 2013. Prorotodactylus andRotodactylus tracks: an ichnological record of dinosauromorphsfrom the Early-Middle Triassic of Poland. In: Nesbitt SJ, DesojoJB, Irmis RB (editors). Phylogeny and palaeobiology of early arch-osaurs and their kin. Geol Soc London Spec Pub 379:319–351.
Nied�zwiedzki G, Sulej T, Dzik J. 2012. A large predatory archosaurfrom the Late Triassic of Poland. Acta Palaeontol Polon 57:267–276.
Osborn HF. 1916. Skeletal adaptations of Ornitholestes, Struthiomi-mus, and Tyrannosaurus. Bull Amer Mus Nat Hist 35:733–771.
Padian K. 2008. Were pterosaur ancestors bipedal or quadrupedal?:morphometric, functional and phylogenetic considerations. Zitteli-ana Reihe B 28:21–33.
Padian K, Li C, Pchelnikova J. 2010. The trackmaker of Apatopus(Late Triassic, North America): implications for the evolution ofarchosaur stance and gait. Palaeontology 53:175–189.
Parker WG, Nesbitt SJ. 2013. Cranial remains of Poposaurus graci-lis (Pseudosuchia: Poposauroidea) from the Upper Triassic, thedistribution of the taxon, and its implications for poposauroid evo-lution. In: Nesbitt SJ, Desojo JB, Irmis RB, editors. Phylogenyand palaeobiology of early archosaurs and their kin. Geol SocLondon Spec Pub 379:503–523.
Parker WG, Irmis RB, Nesbitt SJ, Martz JW, Browne LS. 2005. TheLate Triassic pseudosuchian Revueltosaurus callenderi and itsimplications for the diversity of early ornithischian dinosaurs.Proc Roy Soc B 272:963–969.
Parrish JM. 2012. Evolution of the archosaurs. In: Brett-SurmanM, Holtz, TR, Jr., Farlow JO, editors. The complete dinosaur (sec-ond edition). Bloomington: Indiana University Press. p 317–329.
Peecock BR, Sidor CA, Nesbitt SJ, Smith RMH, Steyer JS,Angielczyk KD. 2013. A new silesaurid from the Upper NtawereFormation of Zambia (Middle Triassic) demonstrates the rapiddiversification of Silesauridae (Avemetatarsalia, Dinosauri-formes). J Vert Paleontol 33:1127–1137.
P�erez-Lorente F. 1993. Dinosaurios plant�ıgrados en La Rioja. Zub�ıa5:189–228.
Peyer K, Carter JG, Sues H-D, Novak SE, Olsen PE. 2008. A newsuchian archosaur from the Upper Triassic of North Carolina. JVert Paleontol 28:363–381.
Phillips MJ, Gibb GC, Crimp EA, Penny D. 2010. Tinamous andmoa flock together: mitochondrial genome sequence analysisreveals independent losses of flight among ratites. Syst Biol 59:90–107.
Piechowski R, Dzik J. 2010. The axial skeleton of Silesaurus opolen-sis. J Vert Paleontol 30:1127–1141.
Platt BF, Hasiotis ST, Hirmas DR. 2012. Empirical determina-tion of physical controls on megafaunal footprint formationthrough neoichnological experiments with elephants. Palaios27:725–737.
Platt N, Meyer CA. 1991. Dinosaur footprints from the Lower Cre-taceous of Spain: their sedimentological and palaeontological con-text. Palaeogeogr Palaeoclimatol Palaoecol 85:321–333.
Rinehart LF, Lucas SG, Heckert AB, Spielmann JA, Celeskey MD.2009. The paleobiology of Coelophysis bauri (Cope) from theUpper Triassic (Apachean) Whitaker Quarry, New Mexico, withdetailed analysis of a single quarry block. Albuquerque: NewMexico Museum of Natural History and Science Bulletin, Publica-tion 45.
Romer AS. 1972. The Cha~nares (Argentina) Triassic reptile fauna.XII. The postcranial skeleton of the thecodont Chanaresuchus.Breviora 385:1–21.
Schachner ER, Manning PL, Dodson P. 2011. Pelvic and hindlimbmyology of the basal archosaur Poposaurus gracilis (Archosauria:Poposauroidea). J Morphol 272:1464–1491.
Schoch, R. 2007. Osteology of the small aetosaur Aetosaurus fromthe Upper Triassic of Germany. N Jahrb Geol Pal€aontol Abh 246:1–35.
Sen K. 2005. A new rauisuchian archosaur from the Middle Triassicof India. Palaeontol 48:185–196.
Sennikov AG. 2012. The first ctenosauriscid (Reptilia: Archosauro-morpha) from the Lower Triassic of eastern Europe. Paleontol J46:499–511.
Senter P. 2007. Analysis of forelimb function in basal ceratopsians.J Zool 273:305–314.
Sereno PC. 1991. Basal archosaurs: phylogenetic relationships andfunctional implications. J Vert Paleontol 11 (Suppl to Number 4):1–53.
Sereno PC, Arcucci. 1993. Dinosaurian precursors from the MiddleTriassic of Argentina: Lagerpeton chanarensis. J Vert Paleontol13:385–399.
Sereno PC, Arcucci AB. 1994. Dinosaurian precursors from the Mid-dle Triassic of Argentina: Marasuchus lilloensis gen. nov. J VertPaleontol 14:53–73.
Sereno PC, Mart�ınez RN, Alcober OA. 2013. Osteology of Eoraptorlunensis (Dinosauria, Sauropodomorpha). J Vert Paleontol 32(Supplement to Number 6): 83–179.
Shapiro MD, You H, Shubin NH, Luo Z, Downs JP. 2003. A largeornithomimid pes from the Lower Cretaceous of the Mazongshanarea, northern Gansu Province, People’s Republic of China. J VertPaleontol 23:695–698.
Smith JB, Farlow JO. 2003. Osteometric approaches to trackmakerassignment for the Newark Supergroup ichnogenera Grallator,Anchisauripus, and Eubrontes. In: Letourneau PM, Olsen PE, edi-tors. The great rift valleys of Pangea in eastern North America,Part Two: sedimentology, stratigraphy, and paleontology. NewYork: Columbia University Press. p 273–292.
Smith JV, Braun EL, Kimball RT. 2013. Ratite nonmonophyly: inde-pendent evidence from 40 novel loci. Syst Biol 62:35–49.
Sookias RB, Butler RJ. 2013. Euparkeriidae. In: Nesbitt SJ, DesojoJB, Irmis RB, editors. Phylogeny and palaeobiology of early arch-osaurs and their kin. Geol Soc London Spec Pub 379:35–48.
Stocker MR, Butler RJ. 2013. Phytosauria. In: Nesbitt SJ, DesojoJB, Irmis RB, editors. Phylogeny and palaeobiology of early arch-osaurs and their kin. Geol Soc London Spec Pub 379:91–117.
Sues H-D, Fraser NC. 2010. Triassic life on land (the great transi-tion). New York: Columbia University Press.
Trotteyn MJ, Arcucci AB, Raugust T. 2013. Proterochampsia: anendemic archosauriform clade from South America. In: Nesbitt SJ,Desojo JB, Irmis RB, editors. Phylogeny and palaeobiology of earlyarchosaurs and their kin. Geol Soc London Spec Pub 379:59–90.
Trotteyn MJ, Mart�ınez RN, Alcober OA. 2012. A new proterochamp-sid Chanaresuchus ischigualastensis (Diapsida, Archosauri-formes) in the early Late Triassic Ischigualasto Formation,Argentina. J Vert Paleo 32:485–489.
Turner AH, Nesbitt SJ. 2013. Body size evolution during the Trias-sic archosauriform radiation. In: Nesbitt SJ, Desojo JB, Irmis RB,editors. Phylogeny and palaeobiology of early archosaurs andtheir kin. Geol Soc London Spec Pub 379:573–597.
Walker AD. 1961. Triassic reptiles from the Elgin Area: Stagonole-pis, Dasygnathus and their allies. Phil Trans Roy Soc London244:103–204.
24 FARLOW ET AL.

Weinbaum JC. 2013. Postcranial skeleton of Postosuchus kirkpa-tricki (Archosauria: Paracrocodylomorpha), from the Upper Trias-sic of the United States. In: Nesbitt SJ, Desojo JB, Irmis RB,editors. Phylogeny and palaeobiology of early archosaurs andtheir kin. Geol Soc London Spec Pub 379:525–553.
Weinbaum JC, Hungerb€uhler A. 2007. A revision of Poposaurusgracilis (Archosauria: Suchia) based on two new specimens fromthe Late Triassic of the southwestern U.S.A. Pal€aontol Zeit 81:131–145.
Weishampel DB, Jianu C-M, Csiki Z, Norman DB. 2003. Osteologyand phylogeny of Zalmoxes (N.G.), an unusual euornithopod dino-saur from the latest Cretaceous of Romania. J Syst Paleontol 1:65–123.
White MA, Benson RBJ, Tischler TR, Hocknull SA, Cook AG,Barnes DG, Poropat SF, Wooldridge SJ, Sloan T, Sinapius GHK,Elliott DA. 2013. New Australovenator hind limb elements per-taining to the holotype reveal the most complete neovenatoridleg. PLoS One 8:e68649. doi: 10.1371/journal.pone.0068649.
Xing L, Peng G, Marty D, Ye Y, Klein H, Gierli�nski GD, Shu C, inpress. An unusual trackway of a possibly bipedal archosaur from
the Late Triassic of the Sichuan Basin, China. Acta PalaeontolPolon: http://dx.doi.org/10.4202/app.2012.0087.
Xing LD, Klein H, Lockley MG, Wang SL, Chen W, Ye Y,Matsukawa M, Zhang JP. 2013. Earliest records of theropod andmammal-like tetrapod footprints in the Upper Triassic of SichuanBasin, China. Vert PalAsiat 51:184–198.
Xu L, Kobayashi Y, L€u J, Lee Y-N, Liu Y, Tanaka K, Zhang X, LiaS, Zhang J. 2011a. A new ornithomimid dinosaur with NorthAmerican affinities from the Late Cretaceous Qiupa Formation inHenan Province of China. Cret Res 32:213–222.
Xu X, You H, Du K, Han F. 2011b. An Archaeopteryx-like theropodfrom China and the origin of Avialae. Nature 475:465–470.
Zanno LE, Varricchio DJ, O’Connor PM, Titus AL, Knell MJ. 2011.A new troodontid theropod, Talos sampsoni gen. et sp. nov., fromthe Upper Cretaceous Western Interior Basin of North America.PLoS One 6:e24487.doi:10.1371/journal.pone.0024487.
Ziegler KE, Heckert AB, Lucas SG (eds.). 2003. Paleontology andgeology of the Upper Triassic (Revueltian) Snyder Quarry, NewMexico. Albquerque: New Mexico Museum of Natural History andScience Bulletin, Publication 24.
POPOSAURUS GRACILIS PEDAL PROPORTIONS 25
Copyright © 2022 FDOKUMEN




















