
PDF - ORIGINAL UNEDITED MANUSCRIPT
30
1 The functional analysis of the dihydroflavonol 4-reductase family of Camellia sinensis: exploiting the key amino acids to reconstruct the reduction activity Haixiang Ruan 1,2# , Xingxing Shi 1,3 # , Liping Gao 2# , Arif Rashid 2 , Yan Li 2 , Ting Lei 2 , Xinlong Dai 3 , Tao Xia 1 * , and Yunsheng Wang 1, 2* 1 State Key Laboratory of Tea Plant Biology and Utilization, Anhui Agricultural University, Hefei, Anhui 230036, China; 2 School of Life Science, Anhui Agricultural University, Hefei, Anhui 230036, China; 3 College of Tea Scinence, Guizhou University, Guiyang Guizhou 550025, China; # These authors contributed equally to this work. * Correspondence: [email protected]; [email protected] Running title: Key amino acid controlling DFR reductase activity Keywords: Camellia sinensis, dihydroflavonol 4-reductase, dihydroflavonols, enzymatic characteristics, metabolic engineering, site-directed mutagenesis © The Author(s) 2022. Published by Oxford University Press. All rights reserved. This is an Open Access article distributed under the terms of the Creative Commons Attribution License http://creativecommons.org/licenses/by/4.0/, which permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited. ORIGINAL UNEDITED MANUSCRIPT Downloaded from https://academic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of PDF - ORIGINAL UNEDITED MANUSCRIPT

1
The functional analysis of the dihydroflavonol 4-reductase family of Camellia
sinensis: exploiting the key amino acids to reconstruct the reduction activity
Haixiang Ruan 1,2#
, Xingxing Shi1,3 #
, Liping Gao2#
, Arif Rashid2, Yan Li
2, Ting Lei
2, Xinlong
Dai3, Tao Xia
1 *, and Yunsheng Wang
1, 2*
1State Key Laboratory of Tea Plant Biology and Utilization, Anhui Agricultural University, Hefei,
Anhui 230036, China; 2School of Life Science, Anhui Agricultural University, Hefei, Anhui
230036, China; 3College of Tea Scinence, Guizhou University, Guiyang Guizhou 550025, China;
# These authors contributed equally to this work.
* Correspondence: [email protected]; [email protected]
Running title: Key amino acid controlling DFR reductase activity
Keywords: Camellia sinensis, dihydroflavonol 4-reductase, dihydroflavonols, enzymatic
characteristics, metabolic engineering, site-directed mutagenesis
© The Author(s) 2022. Published by Oxford University Press. All rights reserved. This is an Open
Access article distributed under the terms of the Creative Commons Attribution License
http://creativecommons.org/licenses/by/4.0/, which permits unrestricted reuse, distribution, and
reproduction in any medium, provided the original work is properly cited.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

2
Haixiang Ruan - [email protected]
Xingxing Shi - [email protected]
Liping Gao - [email protected]
Arif Rashid - [email protected]
Yan Li - [email protected]
Ting Lei - [email protected]
Xinlong Dai - [email protected]
Tao Xia - [email protected]
Yunsheng Wang - [email protected]
Highlight: We reported the first dihydroflavonol 4-reductase (CsDFRs) class II/III genes in
plants. This not only contributes to the DFR gene evolution, but provides candidate DFR gene for
flavonoid metabolic engineering.
Abstract:
Anthocyanin and proanthocyanidins (PAs) are important components of flavonoids, secondary
metabolites in plants with a wide range of industrial and pharmaceutical applications. DFR
(dihydroflavonol 4-reductase) is a pivotal enzyme which plays an important role in the flavonoid
pathway. Here, four CsDFRs genes were isolated from Camellia sinensis and their overexpression
were analyzed in vitro and in vivo. Based on transcription and metabolic analyses, the CsDFRs
expression is closely consistent with catechins and PAs accumulation. Moreover, the enzyme
activity analyses revealed that the two recombinant proteins CsDFRa and CsDFRc exhibited DFR
activity, which converts dihydroflavonols into leucocyanidin in vitro, but not CsDFRb1 and
CsDFRb3. CsDFRa and CsDFRc overexpression in AtDFR mutants (tt3) revealed that CsDFRs
are involved in the biosynthesis of anthocyanins and PAs, as CsDFRa and CsDFRc not only
restored the petiole purple phenotype, but also the seed coat color. Site-directed mutagenesis
revealed that the two amino acid residues S117 and T123 of CsDFRa play a prominent role in
controlling DFR reductase activity. Enzymatic assays indicated that CsDFRa and CsDFRc
exhibited a higher affinity for DHQ and DHK, respectively, whereas CsDFRb1N120S
and
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

3
CsDFRb1C126T
exhibited a higher affinity for DHM. Our findings present comprehensive
characterization of the DFRs from C. sinensis and shed light on their critical role in metabolic
engineering.
Introduction
Flavonoids, which include flavones, flavanols, flavonols, flavan-3-ols (catechins), anthocyanidins
and proanthocyanidins (PAs), are a class of naturally occurring secondary metabolites distributed
widely in different tissues of plants, such as leaves, flowers, stems, and fruits 1. Flavonoids are
also the most potent and health-promoting compounds. In epidemiological, clinical, and animal
studies, they fight against different diseases, such as cardiovascular disease, cancer, and other
disorders 2-4
. The metabolic and biosynthesis pathways of plant flavonoids have piqued interest in
the field of plant secondary metabolites due to their functionalities, compelling evidence of health
benefits, industrial applications, and incorporation into various food products.
The flavonoid biosynthesis pathway in numerous plant species has been extensively investigated
at the genetic, molecular and biochemical levels, with the identification of key functional genes
and transposable elements having been identified. Naringenin is an important precursor that can be
hydroxylated by flavonoid-3-hydroxylase (F3H) at the 3-position of the C ring or hydroxylated by
flavonoid-3'-hydroxylase (F3'H) or flavonoid-3'5'-hydroxylase (F3'5'H) to produce three types of
dihydroflavonols, including DHK (dihydrokaempferol), DHQ (dihydroquercetin) and DHM
(dihydromyricetin) 5. DFR (dihydroflavonol 4-reductase) is an enzyme that converts
dihydroflavonols to leucoanthocyanidins, which are precursors to anthocyanidins, flavan-3-ols,
and PAs (Figure 1A) 6.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

4
Figure 1. The flavonoid biosynthesis pathway and the end-product accumulation of flavonoids in C. sinensis. (A) The
flavonoid biosynthesis. DFR, dihydroflavonol 4-reductase; CHI, chalcone isomerase; ANS, anthocyanidin synthase; CHS,
chalcone synthase; F3’H, flavonoid 3’-hydroxylase; F3H, flavanone 3-hydroxylase; F3’5’H, flavonoid 3’5’-hydroxylase; FLS,
flavonol synthase; LAR, leucoanthocyanidin reductase; ANR, anthocyanidin reductase; (B) The proportion of various flavonoid
compounds. The proportion of flavonoid compounds were taken from Jiang 7.
DFR is a flavonoid biosynthesis key rate-limiting enzyme that belongs to the NADPH-dependent
epimerase/dehydratase family. The genes that code for the DFR enzyme have been characterized
and isolated from a variety of plant species, and their functions have been extensively studied 8-11
.
Several reports have shown that DFR is primarily involved in the color of the petals and seed coat.
The Arabidopsis DFR-deficient mutant (tt3), for example, has a complete lack of PAs
accumulation and the seed coat becomes transparent, and the DFR-deficient gerbera has a normal
phenotypic of white-colored petals 12,13
. Other DFR-like genes in plants, such as Oryza sativa’s
OsDFR2 and DRL1 from Arabidopsis, have been established to have virous biochemical and
physiological activities in plants, and are essential for pollen formation and male fertility.
Several studies have shown that dihydroflavonol 4-reductases from different species have distinct
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

5
substrate specificity, such as the DFRs from Cymbidium 14
and Petunia 15
, which did not
effectively reduce DHK, the precursor to orange pelargonidin-type anthocyanins. Previously, a 26
amino acid region in the middle of the sequence was identified as determining the substrate
specificity of DFR proteins 15
.
C. sinensis is a commercially significant plant that produces one of the most popular non-alcoholic
beverages on the planet. Besides its distinct flavor, tea is regarded as a health drink due to its
nutritional and therapeutic properties 16,17
. Tea has become one of the most prominent symbols of
Chinese culture. Tea health promoting properties have been mainly attributed in recent years to its
polyphenol content, particularly flavan-3-ols (catechins) and flavonols, which comprise 30% of
fresh leaf dry weight 7,18
. Catechins with gallic acid esters, such as epigallocatechin-3-gallate
(EGCG) as well as epigallocatechin (ECG), are the most abundant catechins in tea and have
gained the most attention 19
. The total amount of PAs in the root, particularly proanthocyanidins
dimers and trimers, however, significantly higher than in the leaves and stems (Figure 1B and
Table S1) 7. Overall, these findings reveal that the tea plant accumulates a high amount of
catechins, but the molecular mechanism underlying catechin metabolism and regulation in tea
leaves remains elusive.
Previous studies indicate that CsDFRa is a key gene in regulating catechin content in tea plants 20
.
Previously, five CsDFR genes have been identified using genome sequence and transcriptome
databases 21
. In this study, four CsDFR genes (CsDFRa、CsDFRb1、CsDFRb3、CsDFRc) were
identified and cloned from the bud/root of C. sinensis cv. Shuchazao, and the expression patterns
in different tissues were analyzed using qRT-PCR. A phylogenetic analysis was performed which
revealed that CsDFRa was clustered into class Ⅰ, and the other three proteins of C. sinensis were
clustered independently outside the other NADPH-dependent reductase branches (class II and
class III). The prokaryotic expression vector was constructed and the enzyme activity was
determined in vitro. Moreover, to elucidate the function of CsDFRs in vivo, CsDFRs were
overexpressed in the Arabidopsis tt3 mutant to observe phenotypic and metabolic changes. To
determine the region of the DFRa and DFRb1 enzyme responsible for the substrate specificity, we
introduced chimeric DFRs and determined the activity region. Furthermore, we confirm that the
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

6
activity of DFR can be modulated by the alteration of the amino acids in the region. This paper
reports the functional identification of DFR class II and class III genes in plants comprehensively
and systematically, and it is proposed that these findings will not only aid in the study of the DFR
gene family evolution of in shrub/vine plants, but will also provide new candidate DFR genes for
flavonoid metabolic engineering.
Results
Cloning and sequence analysis of CsDFRs
Based on the genome sequence and transcriptome information of C. sinensis, five putative
CsDFRs genes were screened and identified, including CsDFRa (GeneBank ID: KY615690),
CsDFRb1 (KY615691), CsDFRb2 (KY615692), CsDFRb3 (KY615693), and CsDFRc
(KY615694) 21
. To clone the CsDFR genes, cDNA libraries were constructed from the buds and
roots of cv. Shuchazao and screened using the gene-specific primers of CsDFRs from C. sinensis
as listed in Table S2. The other four genes, except CsDFRb2, have been successfully cloned and
sequenced (Figure S1A). The sequencing results showed that the ORF of CsDFRa, CsDFRb1,
CsDFRb3 and CsDFRc are 1044bp, 1035bp, 1023bp and 1074 bp, respectively, encoding 347, 344,
340 and 357 amino acids (Figure S1A). The characteristics of four structural genes were shown in
Figure S1B. The four DFR genes have similar genetic structures with 6 exons and 5 introns,
similar to DFR genes from other plants such as Arabidopsis and buckwheat 22,23
. To understand
further the function of the DFR genes, we predicated the major transcription factor binding motif
in the promoter region (Figure S1C), and MYB binding sites have been detected in the promoter
region of DFRa, DFRb1, and DFRc. However, no MYB binding site was detected in the promoter
region of DFRb3. In the promoter region of DFRb1, the MYB motifs MYBPLANT and
MYBPZM were not found.
Sequence alignment with several identified DFRs revealed that the N-terminus regions of the C.
sinensis DFR proteins comprised putative NADPH and substrate binding sites (Figure 2A).
Compared with other DFR proteins, the substrate binding regions of the CsDFRb3 and CsDFRc
sequences have 5 or 3 redundant residues, respectively. DFR proteins can be classified into three
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

7
types based on the conservation of the 134th residue: asparagine (Asn), aspartic acid (Asp) and
neither-Asn nor Asp 15
. CsDFRa, like most DFR proteins from other species, is an Asn-type
protein whereas the other three proteins from C. sinensis are neither Asn nor Asp-type (Figure 2).
A phylogenetic tree was constructed using the neighbor-joining method to further investigate the
homology of these four proteins with other known DFRs and NADPH-dependent reductases, such
as anthocyanidin reductase (ANR), leucoanthocyanidin reductase (LAR), and anther-specific
DFR-like enzymes. The result showed that CsDFRa was clustered into class I, with proteins from
dicotyledon, monocotyledon, and gymnosperm species. In class I, several proteins have been
found in previous literature (Figure 2B). CsDFRa was most similar to Actinidia chinensis
(identified score: 98.86%), Rhododendron simsii, and Vaccinium ashei (89.28%). Furthermore, the
other three proteins of C. sinensis were clustered independently outside the other
NADPH-dependent reductase branches (class II and III), and CsDFRb1 and b3 were slightly
closer to the core DFR branch. To our knowledge, the functions of genes in classes II and III have
not been confirmed in vivo or vitro, and most of the genes are from shrubs or vines, especially
from Actinidia chinensis, Vitis vinifera, and Manihot esculenta. Therefore, CsDFRb1, b3, and
CsDFRc might have special significance for studying the evolution of DFR gene family. The
function of these genes needs to be further elucidated.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

8
Figure 2. Sequence analysis of CsDFRs. (A) Multiple sequence alignment of DFRs from various plant species. The DFR
featured motifs, including the NADPH binding region and substrate binding site are boxed in red and blue, respectively. (B)
Phylogenetic tree of CsDFRs and other reductase different plant species. The accession numbers were obtained from publically
accessible database NCBI, and are presented in the figure.
Expression pattern of CsDFRs in tea plants
Quantitative RT-PCR was used to detect the expression patterns of CsDFR genes in tea plants. The
gene-specific primers were used for the amplification of the expression tendency of four genes in
distinct tissues (buds and young leaf, mature leaf, stem and root) (Figure 3A). For normalization,
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022
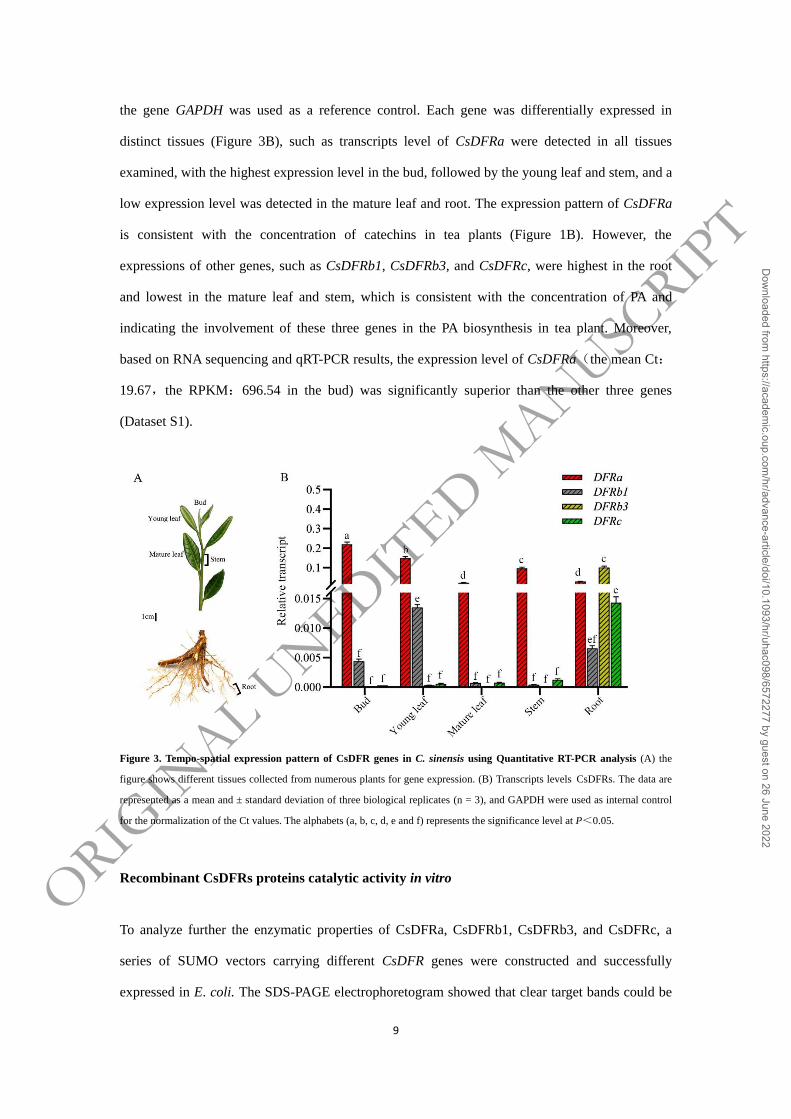
9
the gene GAPDH was used as a reference control. Each gene was differentially expressed in
distinct tissues (Figure 3B), such as transcripts level of CsDFRa were detected in all tissues
examined, with the highest expression level in the bud, followed by the young leaf and stem, and a
low expression level was detected in the mature leaf and root. The expression pattern of CsDFRa
is consistent with the concentration of catechins in tea plants (Figure 1B). However, the
expressions of other genes, such as CsDFRb1, CsDFRb3, and CsDFRc, were highest in the root
and lowest in the mature leaf and stem, which is consistent with the concentration of PA and
indicating the involvement of these three genes in the PA biosynthesis in tea plant. Moreover,
based on RNA sequencing and qRT-PCR results, the expression level of CsDFRa(the mean Ct:
19.67,the RPKM:696.54 in the bud) was significantly superior than the other three genes
(Dataset S1).
Figure 3. Tempo-spatial expression pattern of CsDFR genes in C. sinensis using Quantitative RT-PCR analysis (A) the
figure shows different tissues collected from numerous plants for gene expression. (B) Transcripts levels CsDFRs. The data are
represented as a mean and ± standard deviation of three biological replicates (n = 3), and GAPDH were used as internal control
for the normalization of the Ct values. The alphabets (a, b, c, d, e and f) represents the significance level at P<0.05.
Recombinant CsDFRs proteins catalytic activity in vitro
To analyze further the enzymatic properties of CsDFRa, CsDFRb1, CsDFRb3, and CsDFRc, a
series of SUMO vectors carrying different CsDFR genes were constructed and successfully
expressed in E. coli. The SDS-PAGE electrophoretogram showed that clear target bands could be
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

10
obtained after IPTG inducement of recombinant strains (Figure 4A).
We detect the product anthocyanin, splitted from the leucoanthocyanidin under high temperature
and acidic conditions, because the leucoanthocyanidin produced by DFR enzyme is highly
unstable and difficult to detect directly by using HPLC. The red products were observed in the
culture of the recombinant strain carrying the CsDFRa or CsDFRc gene, using different substrates,
including DHK, DHQ, or DHM. As illustrated in Figure 4B, when dihydroflavonols were used as
a substrate, no red products were observed, indicating that CsDFRb1 and b3 had little or no
reductive activity.
Figure 4. Analyses of catalytic reaction product with CsDFRs fusion proteins. (A) Schematic map of the recombinant
CsDFR construct and the profiles of expressed CsDFRs fusion protein. M: Protein molecular weight markers; 1: Before
induction; 2: The total proteins of induction by SUMO-CsDFRs; 3: Purified protein. (B) Analyze of the enzyme reaction with
SUMO-CsDFRs in vitro, and DHK、DHQ and DHM as the substrate.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

11
Functional confirmation of the CsDFRs in Arabidopsis
In A. thaliana, tt3 was the DFR genes mutant, which not only showed a deficiency in anthocyanin
synthesis but also exhibited an absence of the seed coat pigment. Therefore, the tt3 mutant is a
suitable model plant for determining whether CsDFR is associate with the biosynthesis of
anthocyanin and PAs.
The ORFs of three CsDFRs (including CsDFRa, CsDFRb1, and CsDFRc) were transformed into
tt3 mutant driven by 35S promoter, to elucidate whether CsDFRs have a functional tt3 ortholog
that can restore the above-mentioned deficient phenotypes of Arabidopsis tt3 mutant. After
overexpression of CsDFRs in Arabidopsis tt3 mutant, perceptible phenotypic vary were observed
in the CsDFRa and CsDFRc transgenic seedlings and/or seed coats relative to the tt3 mutant
(Figure 5A). The CsDFRa and CsDFRc not only restored the petiole purple phenotype but also
recovered the seed coats color of the tt3 mutant, as compared to the tt3 mutant and CsDFRb1
transgenic lines (Figure 5A).
DFR converts dihydroflavonols into leucoanthocyanins in the flavonoid biosynthesis pathway.
Following that, leucoanthocyanins were further converted into anthocyanins by ANS. These
aforementioned compounds mentioned will play a role in the synthesis of PAs in plants 24,25
.
Polyphenols were extracted from wild type (Ler), tt3 mutant, and CsDFRs transgenic lines to
detect the contents of anthocyanins and soluble PAs in the seedlings and seeds. CsDFRa and
CsDFRc transgenic Arabidopsis had significantly higher anthocyanins content in the seedlings
than CsDFRb1 and tt3 mutant lines (Figure 5B). Furthermore, DMACA staining revealed that
bluish compounds were formed in the seeds extract of the CsDFRa and CsDFRc transgenic lines,
indicating that the PAs were increased in the seeds compared to the control tt3 mutant, along with
wild type as a positive control (Figure 5B). These findings suggested that CsDFRa and CsDFRc
were involved in the biosynthesis of anthocyanin and PAs, and they are consistent with the
catalytic function of CsDFRa and CsDFRc, which can catalyze the dihydroflavonols into
leucoanthocyanidins, but CsDFRb1 cannot in vivo.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

12
Figure 5. Overexpression CsDFRs in Arabidopsis mutant tt3. (A) The phenotype of the Ler, tt3, CsDFRa/tt3, CsDFRb1/tt3
and CsDFRc/tt3 in seeding or seeds. (B) The absorbance values of the anthocyanin extracts and of bluish compounds from
DMACA reactive compounds at 530 nm and 640 nm, respectively. All the values are the means of three biological replicates, and
the different letters (A, B, C and a, b, c) indicated the significant level at P<0.05.
Identification of a region that determines the activity deference between CsDFRa and
CsDFRb1
Although the amino acid sequences of the CsDFRb1 and CsDFRa are quite similar, the CsDFRa
can reduce dihydroflavonol, while the CsDFRb1 cannot. We used cDNA sequences from these
two genes to create five chimeric DFR genes in order to find the area that controls the activity
difference between CsDFRa and CsDFRb1. (Figure 6A and Dataset S2). Prior to catalytic analysis,
the purified proteins were prepared and purified. The reducing activity of DHQ was used as the
substrate to test the reducing activity of these chimeric DFR proteins (Figure 6B).
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

13
Figure 6. Truncation and reorganization that determines the putative activity switch region between CsDFRa and
CsDFRb1. (A) The model of amino acid position by truncation. FS1, FS2, FS3, FS4 and FS5 stands for five different
reorganization strategies, amino acid residues with red represent fusion between CsDFRa and CsDFRb1, substrate-binding
region and putative activity switch region are boxed in black and blue, respectively. (B) Analyze of the enzyme reaction with
CsDFRa, FS1, FS2, FS3, FS4, FS5 and CsDFRb1 in vitro and DHQ as the substrate.
The chimeric gene FS1 was constructed with 5’-CsDFRb1 and 3’-CsDFRa at the start of the
CsDFRa substrate-binding region, while FS2 was created with 58 amino acids before the
substrate-binding region. The results of activity tests revealed that cyanidin formation could be
observed in the reaction system with FS1 protein in vitro, but not in the reaction system with FS2
protein. We speculate that the DFR protein has an activity switch region prior to the
substrate-binding region. Based on this speculation, three chimeric genes were further constructed,
and the sequence of CsDFRb1 was gradually being replaced by CsDFRa before the
substrate-binding region. The activity test results showed that the FS3 and FS4 recombinant
proteins exhibited reductase activity, whereas the FS5 recombinant enzyme has lost its reductase
activity. Therefore, it was predicted that the amino acid residues between FS4 and FS5
recombination points might have significant impact on DFR enzyme activity.
Site-directed mutagenesis of the activity switch region
The amino acid sequence alignment demonstrated that the amino acids in the putative activity
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

14
switch (PAS) region of CsDFRa and CSDFRb1 are relatively conserved, with only four amino
acids difference (Figure 7A). We conducted a series of gene mutation studies on different residues
to further confirm that these distinct amino acids alter the DFR activity, and DHQ was used as a
substrate to evaluate the reducing activity of the mutated proteins (Figure 7B). These results
showed that the CsDFRa mutated proteins, CsDFRaS117N
and CsDFRaT123C
, lost the DHQ reducing
activity. Meanwhile, the DFR activity was restored by CsDFRb1 mutations, CsDFRb1N120S
and
CsDFRb1C126T
. The mutations in the other two different residues, such as DFRaI118V
and
DFRaI119M
, had no effect on the activity of the non-mutated protein (data no shown). The results of
site-directed mutagenesis showed that the serine at 117 and threonine at 123 of CsDFRa are
critical for modulating DFR reducing activity. Interestingly, these key amino acids are both belong
to the hydroxyl amino acid family.
CsDFRa and CsDFRb1 homologous modeling was carried out using dihydroquercetin (DHQ) and
NADPH as substrates, and splicing experiments were conducted using grape DFR crystals (PDB
code 3BXX) 26
. There was no significant difference in the binding of DFR residues to the substrate
and NADPH in CsDFRs and site-directed mutated proteins (Figure 7B). Furthermore, the PAS
region was found in helix α4, and the X-ray 3D structure revealed that this helix promotes water
molecule binding to the carbonyl group of the asparagine main chain and the amino group of the
catalytic lysine 27
. This is the first of a cluster of water molecules that transfers a proton from the
bulk solvent to the catalytic lysine 28,29
.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

15
Figure 7. Identification of key amino acids that is critical for modulating DFR reducing activity. (A) Analysis of enzymes
activity with different site-directed mutagenesis of the activity switch region. The red asterisk indicates the different amino acids
of DFRa and DFRb1 at the putative activity switch region, DHQ was used as a substrate to evaluate the reducing activity of the
mutated proteins. (B) Homology model of DFRa, DFRb1 and site-directed mutagenesis with docked dihydroquercetin (DHQ)
and NADPH ligand, the PAS region was located in the helix α4 (red band). The model of homology was built using grape DFR
crystals as a template (PDB code 3BXX).
Catalytic activities of recombinant CsDFR proteins in vitro
The dihydroflavonol substrate kinetic parameters of recombinant CsDFR proteins, including
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

16
CsDFRa, CsDFRc, CsDFRb1N120S
and CsDFRb1C1267T
, were determined in potassium phosphate
buffer at pH 7. CsDFRa Km values with DHK, DHQ, and DHM were 145.10, 41.80, and 58.44
μM, respectively. Meanwhile, for CsDFRc, the Km values for DHK, DHQ, and DHM were 42.31,
81.80, and 105.56 μM, respectively. The results indicated that CsDFRa may have a higher affinity
for DHQ, whereas CsDFRc has a higher affinity for DHK (Figure 8 and Table S3).
Enzymatic assays were used to evaluate the activities of the site-directed mutated proteins of
CsDFRb1N120S
and CsDFRb1C126T
. CsDFRb1N120S
Km values for DHK, DHQ, and DHM were
148.96, 108.66, and 96.58 μM, respectively, while CsDFRb1C126T
Km values were 131.09, 102.57,
and 47.75 μM. These findings revealed that the N120S and C126T substitution for CsDFRb1
exhibit a higher affinity for the substrate DHM (Figure 8 and Table S3).
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

17
Figure 8. Kinetic parameters for DFRa, DFRc, DFRb1N120S and DFRb1C126T. In each enzyme reaction assay Two two
micrograms with purified recombinant proteins were used.. A uniform concentration of 0.05 mg dihydroflavonol (DHK, DHQ
and DHM) was used as substrate. The data are expressed as means ± SD (n=3).
Discussion
CsDFR involvement in the biosynthesis of flavonoids in C. sinensis
DFR catalyzes the NADPH-dependent reduction of three different dihydroflavones to form the
corresponding leucoanthocyanins 10
. Leucoanthocyanins are the precursors for the synthesis of
anthocyanins and flavan-3-ols. Therefore, DFR is the pivotal enzyme responsible for anthocyanins,
catechins, and proanthocyanidins accumulation in plants. Catechins, procyanidins, and flavonols
are the most abundant flavonoids in C. sinensis, with catechins mainly accumulated in buds and
young leaves, while procyanidins are mainly accumulated in roots 7,30
.
Numerous studies suggest that DFR expression is positively correlated with anthocyanin and PAs
accumulation. In Saussurea medusa, for example, the SmDFR gene was expressed highly in
flowers but significantly low in leaves and roots 31
. In Lilium asiatica, two DFR genes were highly
expressed in the colored parts of the perianth, anther, filament, stigma, and scale 32
. Shimada
investigated the tissue expression specificity of DFR using Bermuda fruit, flower, leaf, stem, and
root as materials, and demonstrated that the tissue expression specificity of DFR gene was
consistent with its catalytic activity 33
. Tang LK demonstrated that AtDFR was highly expressed
during pollen tube development. The results suggested that AtDFR may play an important role in
pollen maturation and growth 34
. In tea plants, Mamati showed that the DFR gene was often highly
expressed in tissues with high catechin content 35
.
Four DFR genes were identified and described from C. sinensis in this work. Among those genes,
CsDFRa is strongly expressed in the bud and young leaf of C. sinensis, implying that CsDFRa is
the major gene determining the accumulation of catechins in tea plant. While the other genes, on
the other hand, are significantly expressed in the young root of tea plant, indicating that the three
genes are linked to PAs buildup in the root.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

18
Catalytic specificity analysis of the DFR family
Dihydroflavonol is classified into three types based on the number hydroxyls groups on the B-ring,
they include DHK (mono-hydroxyl in B-ring), DHQ (di-hydroxyl in B-ring) and DHM
(tri-hydroxyl in B-ring). In this study, enzyme activity analysis showed that four DFR enzyme
exhibited different selectivities to three substrates in vitro. According to the structure analysis of
proteins, DFR contains two important domains: the N-terminal highly conserved “NADPH”
binding site and the substrate “dihydroflavones” binding site 36
.
Previous studies has shown that the amino acid 134 is important for the catalytic activity of DFR
protein 15
. Therefore, based on the type of amino acid 134, DFR can be divided into three
categories. Type 1, when position 134 is Aspartic acid (Asp), it is called the Asp-type, which can
catalyze DHK to form the leucoanthocyanins. This type of DFR protein has been reported in
cranberry 37
. Type 2, when the position of amino acid 134 is asparagine (Asn), it is called the
Asn-type, which can't catalyze the DHK to form leucoanthocyanins. Type 3, when position of
amino acid 134 is neither Asp nor Asn, it is called non-Asp or Asn-type.
With the development of bioinformatics, DFRs have been identified in numerous plant species.
Meanwhile, the previously identified genes of DFR family can also be divided into different types.
For instance, in the Lotus corniculatus Linn, five DFR genes can be divided into three types.
Among them, LcDFR1 belongs to the non-Asp or Asn-type, LcDFR2 and LcDFR3 belong to the
Asp-type, the other two, LcDFR4 and LcDFR5, belong to the Asn-type 38
.
In this study, we found that the understanding of the basic profile of DFR involvement in the
catechin biosynthesis in C. sinensis is still being unraveled. The evolutionary analysis shows that
CsDFRa belongs to Class I, and the function of most genes in this subfamily has been confirmed
to have dihydroflavonol reductase activity (Figure 2B). Members of Class I are classified as Asn
or Asp types based on the difference in the key amino acid sites in the substrate binding region.
The kinetic analysis of recombinant enzyme revealed that CsDFRa had a high DHQ conversion
efficiency.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

19
The rooted phylogenetic tree analysis shows that CsDFRb1, CsDFRb3 are grouped into the Class
II and CsDFRc are grouped into class III, which belongs to the non-Asp or Asn-type. The
enzymatic activities of members of this subfamily have not been reported before. In the present
study, we demonstrate that CsDFRc has the catalytic activity of dihydroflavonol 4-reductase by
the functional analysis of prokaryotic recombinant protein and Arabidopsis transgenic in vitro and
in vivo, respectively. The kinetic analysis of the recombinant enzyme showed that CsDFRc
displayed highest conversion efficiency for DHK. However, CsDFRb1 and CsDFRb3 did not
detect the catalytic activity of Dihydroflavonol 4-reductase.
Crucial amino acid residues or regions for DFR protein
Several studies have indicated that the DFR substrate specificity is determined by a 26 amino acid
region 15
. CsDFRb1 without catalytic activity and CsDFRa with catalytic activity were truncated
and recombined for each other, and the recombinant protein was expressed in E. coli to further
investigate the key amino acid residues that determine the function of dihydroflavonol reductase.
Analysis of enzyme activity showed that the twenty amino acid residues before the substrate
binding region might have important effects on DFR enzyme activity. Furthermore, site-directed
mutation confirmed that N120 and T126 are the key amino acid sites determining the catalytic
activity of CsDFRb. Interestingly, among them, CsDFRb1N120S
and CsDFRb1T126C
have catalytic
activity, and their affinity for DHM is significantly higher than that of DHQ and DHK. These
results can be used for metabolic engineering of B-ring trihydroxyflavonoid.
Homologous modeling and substrate docking showed that the site-directed mutation had no effect
on the three-dimensional structure of the DFR protein, and there was no significant difference in
the spatial conformation between the substrate and NADPH. The putative activity switch (PAS)
region is located in helix α4 region (Figure 7B), which mainly determines the second H+ supply in
the reduction reaction 27
. Our findings confirmed that the key amino acid residues, serine and
cysteine in PAS region (S117 and C123 of CsDFRa), are mercapto amino acid or hydroxyl amino
acid, which may participate in the transfer of H+. The hydroxyl group established H-bond with
water molecule or protein is critical for maintaining the reductase activity. The establishment of
the H-bond with water molecule and proteins can’t be refuted, because the binding of a water
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

20
molecule to the carbonyl group of the main-chain of the asparagine (N120 of CsDFRb1) and the
amino group of the catalytic lysine (T126 of CsDFRb1), they either break H-bond or induce steric
hindrance for substrate binding, which coincides with previous findings 15
. Taken together, these
studies provide a theoretical foundation for learing more about DFR’s catalytic.
Conclusion
In conclusion, we isolated and characterized the CsDFR genes (CsDFRa, CsDFRc, CsDFRb1 and
CsDFRb3) in C. sinensis and demonstrated their roles in vitro and in vivo. We found that its
function is closely associated with catechins and PAs accumulation. Its physiological role was
studied in the Arabidopsis tt3 mutant, and it was found that CsDFRa and CsDFRc recovered tt3
mutant phenotypes. Moreover, the enzyme activity assay was determined in vitro for the DFR
activity of the CsDFRs and found that CsDFRa and CsDFRc have the DFR activity which
converts dihydroflavonols into leucocyanidin. Through site-directed mutations, it was found that
two amino acid residues of CsDFRs play an important role in enzyme activity. The enzymes
kinetic revealed that the recombinant CsDFR enzyme showed that the optimal substrates for
CsDFRa and CsDFRc were DHQ and DHK, and the optimal substrates for CsDFRb1N120S
and
CsDFRb1C126T
were DHM, respectively. Thus, our data suggests that DFRs evolutionarily
conserved in many plant species, which helps to provide a new candidate gene for the metabolic
engineering of the flavonoids.
Methods
Plant materials
Different tissues bud, young leaf, mature leaf, stem and root of C. sinensis cv. Shuchazao were
collected in early autumn from a five years old plant at Anhui Agricultural University in Hefei,
China (north latitude 31.86, east longitude 117.27, altitude 20 m above mean sea level)
experimental garden. The samples were frozen and stored at -80°C for future use.
The Landsberg erecta (Ler, wild-type) and AtDFR mutant (tt3, CS84) of Ler Arabidopsis were
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

21
obtained from the Arabidopsis Biological Resource Center (abrc.osu.edu) and grown in the growth
chamber of the College of Life Sciences, Anhui Agricultural University with a 16 h light/8 h dark
cycle at 22±2°C with a light intensity of 200 μmol m-2
s-1
.
Isolation and cloning of CsDFR genes
RNAiso Plus for plant tissue kits (Takara, Dalian, China) was used to isolate total RNA from the
bud and root. Standard end-to-end PCR reaction were performed on CsDFRs genes from the
NCBI database, with gene specific primers designed (Table S2) according to the cDNA sequence.
End-to-end PCR was carried out at 98°C for 30 s, 32 cycles at 98°C for 10 s, 58°C for 15 s, 72°C
for 30 s, and a final extension at 72°C for 10 min. Subsequently, the PCR amplification products
were purified using a Gel Extraction Kit (Aidlab, Beijing, China) and cloned into an Easy-Blunt
vector (TransGen Biotech). The resulting vector was introduced into DH5α competent cells and
sequenced.
Phylogenetic analysis
The unabridged length amino acid sequences of DFR proteins from C. sinensis and several other
plants were acquired from the NCBI database (https://blast.ncbi.nlm.nih.gov/Blast.cgi) for
Phylogenetic analysis. ClustalW version 2.1 was used to align all of the sequences and deduced
amino acid sequences of CsDFRs, and the alignment was then submitted to MEGA7 software
(https://www.megasoftware.net) to generate a tree with neighbor-joining and bootstrapping (1000
replicates) analysis, as well as handling gaps with complete deletion.
Quantitative real-time PCR analysis
The gene-specific qRT-PCR primers were designed to study the tempo-spatial expression patterns
of CsDFR genes in C. sinensis (Table S2). On a Peltier Thermal Cycler PTC200 (Bio-Rad, USA),
the transcript level was determined using the SYBR premix Ex Taq II kit (Takara, Japan)
Triplicate of all biological samples were analyzed. The Ct values of the housekeeping gene
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) from C. sinensis were used to normalize
the reaction.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

22
Heterologous expression of CsDFRs in A. thaliana
Using Gateway technology (Invitrogen, USA) the full-length CsDFRs cDNA was shuttled into the
pCB2004 binary vector. Subsequently, the positive recombinant pCB2004-CsDFRs vector was
Agrobacterium tumefaciens (GV3101) using electroporation, and the Agrobacterium was then
used for floral dip transformation of Arabidopsis mutant tt3 39
. The transformed plants were grown
in the glasshouse under carefully controlled conditions (16 h light/8 h dark cycle at 22±2°C with a
relative humidity of 40%).
Extraction and quantification analysis of anthocyanins and PAs from plant materials
For the extraction of anthocyanins and PAs, briefly, freeze-dried samples (including seedlings and
seeds) of 0.05 g of CsDFRs transgenic lines, WT and tt3 mutant were ground by grinders.
Subsequently, the total polyphenols were extracted at a low temperature with 1.2 mL of extraction
solution A (HCl 0.5%: methanol 80%: water 19.5%, v/v) to the samples and vortexed and
sonicated for 30 min at 4°C 7. The samples were centrifuged at 13000g for 15 minutes before
being re-extracted twice more as indicated above until the final volume of supernatants was 2mL.
Finally, the combined solution was centrifuged for 10 minutes at 13 000g and the supernatants
were stored at −20°C prior to total content analysis of anthocyanins and PAs. The contents of total
anthocyanins in the supernatants were measured with UV spectrometer at 530 nm. After a reaction
with the DMACA reagent (0.2% DMACA, w/v, HCl/methanol 1:7, v/v), the total soluble PAs
content in the seeds was determined by a UV spectrometer at 640 nm. Briefly, the reaction mixture
(1.3 mL total volume) contains 0.3 mL of supernatants and 1 mL of DMACA reagent.
Subsequently, the reaction mixture was incubated at 25°C for 5 min, and the absorbent values of
the reaction mixture were measured at 640 nm.
Heterologous expression of CsDFRs in Escherichia coli
The CsDFRs ORF was subcloned into the pET-SUMO vector (Invitrogen, California, USA) in its
entirety. The identity of the cloned gene was validated using T7 primers and sequencing analysis.
The resulting vector SUMO-CsDFRs were transformed into BL21 (DE3) E. coli competent cells,
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

23
according to the manufacturer’s instructions. All of the chimeric and point mutation genes were
cloned into the pET-SUMO vector with the same method as above. Positive colony, expression
strains were cultivated in 200 mL of LB medium with 50 μg·mL−1
kanamycin at 37°C. To
promote recombinant protein expression, 1 mM isopropyl-β-D-thiogalactoside (IPTG) was added
when the E.coil OD600 reached 0.6. The cells were collected by centrifugation after 8 hours of
induction at 28°C, and the fusion protein was purified using His-tag affinity chromatography
according to the manufacturer’s instructions. Twelve percent SDS polyacrylamide gel staining and
Coomassie brilliant blue staining were used to validate the isolated protein fraction. The activity
assay in vitro was performed using pure recombinant proteins. All specific primers for
SUMO-CsDFRs, chimeric DFRb1/DFRa, and amino acid point mutant genes were listed in Table
S2.
Enzyme activity assay and analysis of kinetics parameters
The enzymatic reaction consisted of 0.1M pH 7.0 potassium phosphate buffer, 2 mM NADPH, 0.5
mM dihydroflavonols (DHK, DHQ and DHM) and 0.05 mg total soluble proteins content. The
reaction mixtures were incubated at 30°C for 30 min, and the total volume was 200 μl. Because
leucoanthocyanidin is unstable in solution and difficult to detect directly using HPLC, a three
times volume of butanol–HCl reagent (95:5, v/v) was added, and the reaction was splitted at 95°C
for 1h to form anthocyanidin and was centrifuged at 4000g for 15min. The absorption of the
supernatants was measured at OD530 nm by UV spectrophotometer. According to the absorption
value of anthocyanin standard at different concentrations, the standard curve was established to
quantify the content of products in the sample. To determine the apparent Km value of the reaction
mixture of different substrates, 0.05 mg purified recombinant enzyme and three dihydroflavonols
at a series of concentrations. The enzymatic products were quantified as the standard curve of
anthocyanins standard. Apparent Km data were calculated using Hanes plots.
Molecular Docking statistic and site-directed mutagenesis
The homology model of CsDFRa and CsDFRb1 were constructed with the crystal structure of
grape DFR (PDB code 3BXX) as a template using the online server SWISS-MODEL
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

24
(http://swissmodel.expasy.org/interactive). Furthermore, dihydroquercetin and NADPH were
taken as the substrates, and they were docked into the model of CsDFRa and CsDFRb1 with the
lowest energy conformation, using the software AutodockTools-1.5.6
(http://mgltools.scripps.edu/downloads). The resulting spatial model structure of the CsDFRa and
CsDFRb1 protein, including binding sites and substrate linkage pockets, were analyzed and
visualized using Python 27 software. The site-directed mutation plasmids CsDFRaS117N
,
CsDFRaT123C
, CsDFRb1N120S
, CsDFRb1C127T
, DFRaI178V
and DFRaI179M
were generated using an
overlap extension PCR technology with CsDFRa and CsDFRb1 plasmids as the templates,
respectively. All of the specific oligonucleotide primers used to construct the mutations plasmids
is listed in Table S2.
Statistical methods
Three separate experiments or three biological replicates were used in each of the aforementioned
test. Using SPSS software, mean values were generated and then statistically analyzed using a
Student’s test. The two-tailed test of significance was used. Statistical significance was determined
by P-values of less than 0.05.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (31770729,
31870676); the National Key R&D Program of China (2018YFD1000601); Collegiate
Collaborative Innovation Foundation of Anhui Province (GXXT-2020-081) and Joint Funds of the
National Natural Science Foundation of China (U21A20232).
Author contributions
RHX, SXX and WYS conceived and designed the research. WYS, GLP and XT conducted
experiments. SXX, RHX, LY, LT and DXL performed the experiments. RHX, RA and WYS
analyzed the data. RHX and DXL wrote the manuscript, RA and WYS edited and contributed to
the content.
Conflict of Interest
There are no conflicts of interest declared by any of the writers.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

25
Data availability statement
All data supporting the conclusions of this study may be found in the publication and its
supplemental materials, which are available online. Any additional relevant information can be
obtained from the corresponding author upon request (Yunsheng Wang).
Supporting information
Figure S1: Schematic representation of CsDFRs gene structures and promoter elements.
Table S1: Relative quantity of different flavonoid compounds in different tissues.
Table S2: List primers used in this study.
Table S3: Kinetic parameters for DFRa, DFRc, DFRb1N120S
and DFRb1C126T
.
References
1. Winkel-Shirley B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry,
cell biology, and biotechnology. Plant Physiol. 2001;126:485-93.
2. Cabrera C, Artacho R, Gimenez R. Beneficial effects of green tea--a review. J Am Coll
Nutr. 2006;25:79-99.
3. Chen A Y, Chen Y C. A review of the dietary flavonoid, kaempferol on human health
and cancer chemoprevention. Food Chem. 2013;138:2099-107.
4. Dalgaard F, Bondonno N P, Murray K et al. Associations between habitual flavonoid
intake and hospital admissions for atherosclerotic cardiovascular disease: a
prospective cohort study. Lancet Planet Health. 2019;3:e450-e59.
5. Tanaka Y, Brugliera F. Flower colour and cytochromes P450. Philosophical
transactions of the Royal Society of London. Series B, Biological sciences.
2013;368:20120432.
6. Hua C, Linling L, Shuiyuan C et al. Molecular cloning and characterization of three
genes encoding dihydroflavonol-4-reductase from Ginkgo biloba in anthocyanin
biosynthetic pathway. PLoS One. 2013;8:e72017.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

26
7. Jiang X, Liu Y, Li W et al. Tissue-specific, development-dependent phenolic
compounds accumulation profile and gene expression pattern in tea plant [Camellia
sinensis]. PLoS One. 2013;8:e62315.
8. Bowers J E, Dangl G S, Vignani R et al. Isolation and characterization of new
polymorphic simple sequence repeat loci in grape (Vitis vinifera L.). Genome.
1996;39:628-33.
9. Helariutta Y, Elomaa P, Kotilainen M et al. Cloning of cDNA coding for
dihydroflavonol-4-reductase (DFR) and characterization of dfr expression in the
corollas of Gerbera hybrida var. Regina (Compositae). Plant Mol Biol.
1993;22:183-93.
10. Wang J, Wang Y, Wang S et al. Cloning and temporal-spatial expression analysis of
dfr gene from Scutellaria baicalensis with different colors. Sheng Wu Gong Cheng Xue
Bao. 2021;37:1312-23.
11. Wang L, Zhu Y, Wang P et al. Functional Characterization of a Dihydroflavanol
4-Reductase from the Fiber of Upland Cotton (Gossypium hirsutum). Molecules.
2016;21:32.
12. Bashandy H, Pietiainen M, Carvalho E et al. Anthocyanin biosynthesis in gerbera
cultivar 'Estelle' and its acyanic sport 'Ivory'. Planta. 2015;242:601-11.
13. Kubasek W L, Shirley B W, Mckillop A et al. Regulation of Flavonoid Biosynthetic
Genes in Germinating Arabidopsis Seedlings. Plant Cell. 1992;4:1229-36. ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

27
14. Johnson E T, Yi H, Shin B et al. Cymbidium hybrida dihydroflavonol 4-reductase does
not efficiently reduce dihydrokaempferol to produce orange pelargonidin-type
anthocyanins. Plant J. 1999;19:81-5.
15. Johnson E T, Ryu S, Yi H et al. Alteration of a single amino acid changes the substrate
specificity of dihydroflavonol 4-reductase. Plant J. 2001;25:325-33.
16. Alagawany M, Abd El-Hack M E, Saeed M et al. Nutritional applications and beneficial
health applications of green tea and l-theanine in some animal species: A review. J
Anim Physiol Anim Nutr (Berl). 2020;104:245-56.
17. Yao S S, Guo W F, Lu Y et al. Flavor characteristics of lapsang souchong and smoked
lapsang souchong, a special Chinese black tea with pine smoking process. J Agric
Food Chem. 2005;53:8688-93.
18. Asakawa T, Hamashima Y, Kan T. Chemical synthesis of tea polyphenols and related
compounds. Curr Pharm Des. 2013;19:6207-17.
19. Hu J, Zhou D, Chen Y. Preparation and antioxidant activity of green tea extract
enriched in epigallocatechin (EGC) and epigallocatechin gallate (EGCG). J Agric Food
Chem. 2009;57:1349-53.
20. Rani A, Singh K, Ahuja P S et al. Molecular regulation of catechins biosynthesis in tea
[Camellia sinensis (L.) O. Kuntze]. Gene. 2012;495:205-10.
21. Wang W, Zhou Y, Wu Y et al. Insight into Catechins Metabolic Pathways of Camellia
sinensis Based on Genome and Transcriptome Analysis. J Agric Food Chem.
2018;66:4281-93.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

28
22. Katsu K, Suzuki R, Tsuchiya W et al. A new buckwheat dihydroflavonol 4-reductase
(DFR), with a unique substrate binding structure, has altered substrate specificity.
Bmc Plant Biol. 2017;17:239.
23. Shirley B W, Hanley S, Goodman H M. Effects of ionizing radiation on a plant genome:
analysis of two Arabidopsis transparent testa mutations. Plant Cell. 1992;4:333-47.
24. Pang Y, Abeysinghe I S, He J et al. Functional characterization of proanthocyanidin
pathway enzymes from tea and their application for metabolic engineering. Plant
Physiol. 2013;161:1103-16.
25. Xie D Y, Sharma S B, Paiva N L et al. Role of anthocyanidin reductase, encoded by
BANYULS in plant flavonoid biosynthesis. Science. 2003;299:396-9.
26. Trabelsi N, Petit P, Manigand C et al. Structural evidence for the inhibition of grape
dihydroflavonol 4-reductase by flavonols. Acta Crystallogr D Biol Crystallogr.
2008;D64:883-91.
27. Petit P, Granier T, D'estaintot B L et al. Crystal structure of grape dihydroflavonol
4-reductase, a key enzyme in flavonoid biosynthesis. J Mol Biol. 2007;368:1345-57.
28. Benach J, Filling C, Oppermann U C et al. Structure of bacterial
3beta/17beta-hydroxysteroid dehydrogenase at 1.2 A resolution: a model for multiple
steroid recognition. Biochemistry. 2002;41:14659-68.
29. Bottoms C A, Smith P E, Tanner J J. A structurally conserved water molecule in
Rossmann dinucleotide-binding domains. Protein science : a publication of the Protein
Society. 2002;11:2125-37.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

29
30. Zhao J, Li P, Xia T et al. Exploring plant metabolic genomics: chemical diversity,
metabolic complexity in the biosynthesis and transport of specialized metabolites with
the tea plant as a model. Crit Rev Biotechnol. 2020;40:667-88.
31. Li H, Qiu J, Chen F et al. Molecular characterization and expression analysis of
dihydroflavonol 4-reductase (DFR) gene in Saussurea medusa. Mol Biol Rep.
2012;39:2991-9.
32. Nakatsuka A, Izumi Y, Yamagishi M. Spatial and temporal expression of chalcone
synthase and dihydroflavonol 4-reductase genes in the Asiatic hybrid lily. Plant
Science. 2003;165:759-67.
33. Shimada N, Sasaki R, Sato S et al. A comprehensive analysis of six dihydroflavonol
4-reductases encoded by a gene cluster of the Lotus japonicus genome. Journal Of
Experimental Botany. 2005;56:2573-85.
34. Tang L K, Chu H, Yip W K et al. An anther-specific dihydroflavonol 4-reductase-like
gene (DRL1) is essential for male fertility in Arabidopsis. New Phytol.
2009;181:576-87.
35. Mamati G E, Liang Y R, Lu J L. Expression of basic genes involved in tea polyphenol
synthesis in relation to accumulation of catechins and total tea polyphenols. J Sci
Food Agr. 2006;86:459-64.
36. Martens S, Teeri T, Forkmann G. Heterologous expression of dihydroflavonol
4-reductases from various plants. FEBS Lett. 2002;531:453-8. ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022

30
37. Polashock J J, Griesbach R J, Sullivan R F et al. Cloning of a cDNA encoding the
cranberry dihydroflavonol-4-reductase (DFR) and expression in transgenic tobacco.
Plant Science. 2002;163:241-51.
38. Shimada N, Sasaki R, Sato S et al. A comprehensive analysis of six dihydroflavonol
4-reductases encoded by a gene cluster of the Lotus japonicus genome. J Exp Bot.
2005;56:2573-85.
39. Clough S J, Bent A F. Floral dip: a simplified method for Agrobacterium-mediated
transformation of Arabidopsis thaliana. Plant J. 1998;16:735-43.
ORIGIN
AL UNEDIT
ED MANUSC
RIPT
Dow
nloaded from https://academ
ic.oup.com/hr/advance-article/doi/10.1093/hr/uhac098/6572277 by guest on 26 June 2022




















