
Production of Mosquito Repellent Coil ... - Entrepreneur India

Cell Biology International 1999, Vol. 23, No. 3, 219–226Article No. cbir.1998.0337, available online at http://www.idealibrary.com on
PARTIAL CHARACTERIZATION OF MOSQUITO LARVAE EXTRACTMODULATING MOUSE AND HUMAN CELL PROLIFERATION
J. R. RONDEROS1*, O. J. RIMOLDI2, M. A. SALAS3 and R. R. BRENNER2
1Catedra de Histologıa y Embriologıa ‘B’, Facultad de Ciencias Medicas—Universidad Nacional de la Plata,Calle 60 y 120, 1900 La Plata, Argentina
2INIBIOLP, UNLP-CONICET; and 3Catedra de Fisiologıa con Biofısica, FCM-UNLP, Argentina
Received 20 April 1998, accepted 9 December 1998
Mosquito larvae crude extract have been found to alter the mitotic rate of several mouseepithelial cell populations such as enterocytes and tongue keratinocytes. Also, the dialysedfraction inhibits hepatocyte proliferation in hepatectomized males. These experiments suggestedan inhibitory effect on the G1/S interphase. Consequently, we suggested the presence of somemolecule or molecules related to the TGF-� superfamily. In the present paper, we haveassayed the crude extract on human mononuclear cells and the dialysed fraction of the extracton tongue keratinocyte proliferation. Furthermore, different protein fractions obtained using amolecular exclusion chromatographic column were assayed on hepatocyte proliferation ofhepatectomized mice. Three groups of proteins have been isolated. Results show a dose-dependent effect of crude extract on mononuclear cell proliferation and the dialysed extractcaused an inhibitory effect on tongue keratinocyte proliferation. With regard to the hepatocytemitotic rate, an inhibitory effect appeared only in animals receiving the fraction with lowermolecular weight. These results suggest the presence in mosquito larvae of some peptidicmolecule or molecules resembling the activity of members of the TGF-� superfamily.
� 1999 Academic Press
K: keratinocytes; hepatocytes; mononuclear cells; mosquito; growth factors.
*To whom correspondence should be addressed. E-mail:[email protected] biological material used in the in vivo experiments performed inthis paper was collected from Catedera de Histologıa y Embriologıa‘A’, Facultad de Ciencias Medicas, Universidad Nacional de La Plata,where Dr J. R. Ronderos developed his activities until July 1996.
INTRODUCTION
Cell populations have different cycle timeswhich can vary according to distinct life times.Moreover, there exist cycling and noncycling cellsin every population, depending on the require-ments of the organism. Cell growth is regulated byseveral factors such as hormones, growth factorsand cell surface receptor synthesis. As growthcaused by cell proliferation is a fundamentalprocess in biology, some genes involved in ithave probably appeared in the genome by the timeof mitosis appearance. Several factors control-ling growth mechanisms such as growth factors
1065–6995/99/030219+08 $30.00/0
and proto-oncogenes, have been maintained innature throughout evolution. Thus, such distantorganisms as insects and mammals share some ofthese genes. Genes codifying proteins structurallyrelated to transforming growth factor-beta(TGF-�) and epidermal growth factor (EGF) havebeen found in insects, nematodes and echinoderms(Padgett et al., 1987; Massague, 1990; Muskavitchand Hoffmann, 1990; Doctor et al., 1992; Renet al., 1996). The presence of a gene in Drosophilathat encodes a receptor of the TGF-� superfamilywas also evidenced (Tin Xie et al., 1994). Further-more, the decapentaplegic gene, which codifies aprotein highly homologous with TGF-�, wasreported as an imaginal disk cell synchroniser atthe G2/M interphase during Drosophila eyedevelopment (Penton et al., 1997). In view of thesefacts, it could be expected that the mechanismcontrolling cell growth were not species specific.
� 1999 Academic Press

220 Cell Biology International, Vol. 23, No. 3, 1999
On the other hand, it was demonstrated that inAedes aegipty larvae there exist some cell popu-lations which do not grow by hyperplasia (Tragger,1937). This fact might be related to the existence ofsome molecule or molecules inhibiting cell prolifer-ation and perhaps inducing cell differentiation.In order to find growth factors homologous ofvertebrates in insects, we have assayed a crudeextract from other mosquito larvae species (Culexpipiens L.) on the mitotic rate of several epithelialcell populations of mice in vivo. Thus, we havedemonstrated that the crude extract of mosquitolarvae has an inhibitory effect on the mitotic rate ofhepatocytes of partially hepatectomized adult mice(Ronderos and Echave Llanos, 1990). The sameeffect was seen on hepatocytes, enterocytes, sialo-cytes and tongue keratinocytes of growing mice(Ronderos et al., 1994a; Ronderos et al., 1996).Similar results were found when the dialysed frac-tion of the extract (12 kDa cut-off pore membrane)was assayed on hepatocyte proliferation of hepatec-tomized adult mice, showing an association of thebiological effect with the macromolecular fractionof the extract (Ronderos et al., 1994a). Theseresults suggested the presence of some moleculeresembling the activity of members of the TGF-beta superfamily or another unknown factor inmosquito larvae extract. In those experiments,treatments were applied before the DNA synthesiscurve increased (Echave Llanos et al., 1971) andthe sampling period was 16–24 h after treatment,when the mitotic peak occurs (Surur et al., 1985).The time elapsed between treatments and animalsacrifice suggested an inhibitory effect of themosquito extract on the cell cycle G1/S checkpoint.When the mitotic rate-sampling period coveredthe dark phase of the circadian cycle (time ofmaximum DNA synthesis) a statistically significantincrement was found in hepatocytes, nephro-cytes and tongue keratinocytes of extract recipi-ents. Taking into account the time elapsed fromtreatment administration until sacrifice (from0–12 h), these results suggested an effect of theextract at the G2/M checkpoint of the cell cycle(Ronderos, 1996). In other experiments, treatmentswere applied after the DNA synthesis peak occur-rence. The results were similar, showing decreasedmitotic rates in hepatocytes, tongue keratinocytesand enterocytes (Ronderos et al., 1994b).
The aim of the present study has been to assayboth the dialysed extract on keratinocyte and threedifferent protein fractions on hepatocyte prolifer-ation of hepatectomized adult mice. Furthermore,taking into account that members of the TGF-�superfamily have been also related to lymphocyte
function by different authors (Ahuja et al., 1993;Altiok et al., 1994; De Jong et al., 1994; Fox et al.,1992; Fox et al., 1993; Lee and Rich, 1993;Reinhold et al., 1994), we decided to assay theeffect of a crude preparation of mosquito larvae onthe proliferation of human mononuclear cells(MNC).
MATERIALS AND METHODS
Mosquito larvae extract
Third and fourth instar larvae of mosquito Culexpipiens L. (Diptera: Culicidae) were obtained froman artificial colony. Larvae were separated from thecolony, washed several times in fresh water toeliminate any breed plate residue, then filteredand weighed for extract preparation. Pooledmosquito larvae were homogenized in a Potter-Dual homogenizer in an ice bath and saline solu-tion was added (2:10) at 4�C. The homogenate wascentrifuged at 0�C at 14,500�g for 20 min in aSorvall RC5C centrifuge and then dialysed with a12,000 Dalton cut-off pore membrane in salinesolution overnight at 4�C.
Gel permeation chromatography
The dialysed homogenate was applied to a Super-ose 6HR 10/30 column, equilibrated and elutedwith buffer Tris-HCl 10 m, ClNa 0.15 , pH 8.0at a flow rate of 0.5 ml/min. The effluent wasmonitored at 280 nm. The protein-containingfractions were pooled and concentrated by ultra-filtration (Centricom-10, Amicon, Bedford). Theprotein contents were determined by the method ofLowry et al. (1951) using bovine serum albumin asstandard.
Partially hepatectomized adult male mice
C3HS (from Wilson in 1968) adult male mice(90 d old) were standardized for periodicity analy-sis (Echave Llanos and Nash, 1971; Halberg et al.,1958; Vilchez and Echave Llanos, 1971), singlecaged after weaning in a room ad hoc, lighted(fluorescent, 40 W) from 06:00–18:00 alternatingwith 12 h of darkness. Food and water were avail-able ad libitum and the temperature was 22�1�C.
Mice experimental design
C3HS male mice (90 d old) standardized forperiodicity analysis for three weeks before surgery,

Cell Biology International, Vol. 23, No. 3, 1999 221
were partially hepatectomized (70%) at 12:00 h.After 28 h (i.e. 16/28 time of day/time posthepatectomy (TD/TPH)) the animals were intra-peritoneally injected (left side), killed by decapi-tation and exsanguination at 08/44; 12/48 and 16/52(dialysed fraction on tongue keratinocyte prolifer-ation). On the other hand, for the regenerating-liver assay fractions, mice were killed at 12/48TD/TPH (time of mitotic peak). All animals re-ceived 0.01 ml of the different solutions bygram of body weight. As regards treatments, salineand dialysed larvae extracts (2.2 mg/ml) were as-sayed for tongue keratinocyte proliferation. Forregenerating-liver experiments, the treatments as-sayed were the following: saline solution as generalcontrol of the experiment, dialysed fraction(2.0 mg/ml) as mosquito larvae control effect andchromatography eluted fractions (F1: 1.4 mg/ml;F2: 1.0 mg/ml; F3: 0.8 mg/ml). Colchicine (2.0 �g/gbody weight in 0.01 ml of distilled water) was alsoinjected intraperitoneally (left side) to all animals4 h before death.
Mitotic rate evaluation
Tongues and the triangular lobe of the livers fromexperimental mice receiving either buffer salinesolution, dialysed extract or protein fractions, wereremoved and processed for histological analysis(Echave Llanos and Sadnik, 1964). The mitotic ratewas determined in histological slides by countingthe colchicine-arrested metaphases in approxi-mately 5000 cells at a magnification of �1000.Colcichine-arrested metaphases were recorded inevery microscopic field and the total number ofnuclei was counted every ten fields (liver). In thecase of epithelial cells from the tongue, the mitoticrate was determined by counting the colchicine-arrested metaphases of dorsal and ventral surfacesseparately. Finally, mitotic rates were expressed ascolchicine metaphases/1000 nuclei.
Mononuclear cells isolation
Human heparinized blood was obtained fromhealthy adult donors. MNC were isolated bycentrifugation on a Ficoll gradient (Histopaque1077-Sigma). After washing, cells were resuspendedin RPMI medium supplemented with 2 mglutamine, 100 �g/ml streptomycin, 100 U/ml peni-cillin, 2-ME 10�5 (all from Sigma) and 10% fetalbovine serum (Gen-Argentina), and then seededinto 96 well plates at a density of 105 cells/well (3 to6 wells/treatment). MNC viability was evaluated bytrypan blue exclusion test.
MNC experimental design
MNC were cultured for 72 h at 37�C in a humidi-fied atmosphere of 5% CO2 in air. The differenttreatments and the mitogen Concanavalin A(ConA, 5 �g/ml) diluted in RPMI were addedsimultaneously at the beginning of the culture. Inorder to evaluate lymphocyte proliferation [3H]-Thymidine (1 �Ci/well) was added for the last 4 hof culture. Cells were finally recovered on filters bythe use of a cell harvester (Titerteck Cell Harvester,Flow Lab, U.S.A.). The radioactivity incorporatedwas measured in a liquid scintillation counter.
14 000
0
Doses (mg/ml)
cpm
4000
12 000
10 000
8000
6000
2000
Ctrl 1 0.5 0.1 0.05 0.01
*
*
*
Fig. 1. Dose-dependent proliferative response of ConAactivated MNC to mosquito crude extract. Bars representmean�.. Ctrl: controls without extract. *Statistically signifi-cant differences, P<0.05.
Statistical analysis
The experimental design for tongue keratinocyteassay involves variations in the mitotic rate pro-duced by both circadian rhythms and by differen-tial proliferation of the dorsal and ventral surfaces(Ronderos et al., 1994b). Consequently, the differ-ences were analysed by multifactorial analysis ofvariance, including three factors acting on the

222 Cell Biology International, Vol. 23, No. 3, 1999
mitotic rate of this cell population (i.e., treatments,sampling time and tongue surface). For hepatocyteproliferation as well as for human MNC assaystatistical analyses were performed by one wayanalysis of variance.
RESULTS
Effects of the crude extract on MNC proliferation
Doses of the crude extract ranging from 1 to0.01 mg/ml were applied to ConA activated MNCat the beginning of the culture. [3H]Thymidineincorporation showed a dual response of MNC tothe extract. Doses of 1.0 and 0.5 mg/ml caused astatistically significant decrease in proliferation,while 0.05 mg/ml of the extract led to a significant
increment of the MNC entering into the S-phase ofthe cycle. Lower doses did not cause any effect(Fig. 1).
Effect of temperature on biological activity of theextract
With the aim of knowing the effect of temperatureon the biological activity of the extract, it washeated at 90�C for 30 min and then centrifuged at1500 rpm for 5 min. The experimental design wassimilar to the one previously used. A treatmentof 0.5 mg/ml was included as control of theinhibitory effect of the extract. Results showed thatafter heating the biological effect of the extractdisappeared (Fig. 2, Table 1).
Dialysed fraction effect on tongue keratinocyteproliferation
The results showed, as in regenerating-liver(Ronderos et al., 1994a), a significant (P=0·001)decrease of the mitotic rate in tongue keratinocytesof dialysed extract-receiving mice when analysisincluded all factors (saline: 15.08�1.74, n=26;dialysed: 6.90�1.63, n=30) (Table 2).
When the mitotic rate was analysed discriminat-ing the effect of the sampling time, we found thatdifferences were evident only at 12/48 TD/TPH(P=0.0001) (saline: 26.34�3.41, n=10; dialysed:6.2�2.24, n=10; percentage of variation: �76.46)(Fig. 3). The analysis of treatment effects, discrimi-nating between ventral and dorsal surface response,showed that both keratinocyte subpopulationsunderwent a statistically significant decrease in themitotic rate of dialysed fraction-receiving mice(dorsal: P=0.02; ventral: P=0.005) (Fig. 4).
14 000
0
Treatments
cpm
6000
12 000
10 000
8000
4000
2000
Fresh 90°C Ctrl
*
Fig. 2. Effect of fresh (0.5 mg/ml) and heated extract (90�C for30 min) on MNC proliferation. Bars represent means�..*Statistically significant difference.
Table 1.One way analysis of variance comparisons for ConA induced MNC proliferation after treatment
with fresh (0.5 mg/ml) or 90�C heated extract against controls only receiving ConA
Source ofvariation
Sum ofsquares
DF Meansquares
F-ratio
Treatment 2.5�108 2 1.2�108 125.67 P<0.0001Residuals 8.0�106 8 1.0�106 — —Total 2.6�108 10 — — —
Analysis of chromatographic fractions
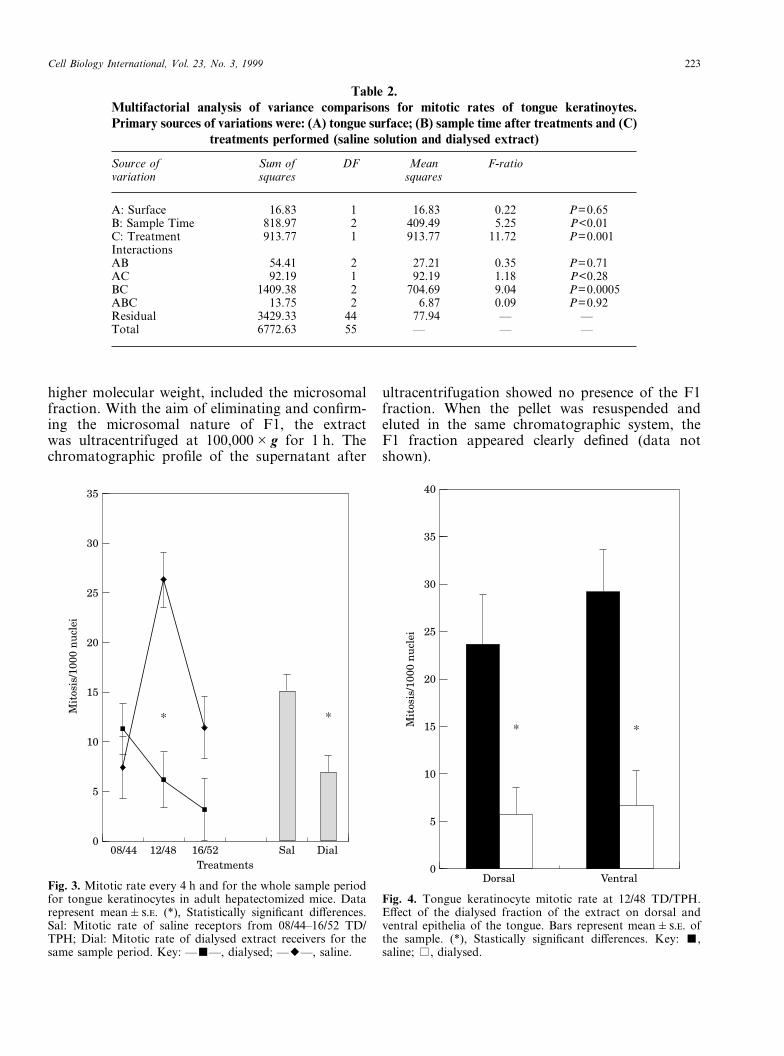
The mosquito larvae dialysed extract was appliedto a gel permeation Superose 6B column and elutedwith Tris-HCl saline buffer. Three main peakswere detected (Fig. 5). The first one (F1), with
Cell Biology International, Vol. 23, No. 3, 1999 223
Table 2.Multifactorial analysis of variance comparisons for mitotic rates of tongue keratinoytes.Primary sources of variations were: (A) tongue surface; (B) sample time after treatments and (C)
treatments performed (saline solution and dialysed extract)
Source ofvariation
Sum ofsquares
DF Meansquares
F-ratio
A: Surface 16.83 1 16.83 0.22 P=0.65B: Sample Time 818.97 2 409.49 5.25 P<0.01C: Treatment 913.77 1 913.77 11.72 P=0.001InteractionsAB 54.41 2 27.21 0.35 P=0.71AC 92.19 1 92.19 1.18 P<0.28BC 1409.38 2 704.69 9.04 P=0.0005ABC 13.75 2 6.87 0.09 P=0.92Residual 3429.33 44 77.94 — —Total 6772.63 55 — — —
Dial
35
008/44
Treatments
Mit
osis
/100
0 n
ucl
ei
10
30
25
20
15
5
12/48 16/52 Sal
**
Fig. 3. Mitotic rate every 4 h and for the whole sample periodfor tongue keratinocytes in adult hepatectomized mice. Datarepresent mean�.. (*), Statistically significant differences.Sal: Mitotic rate of saline receptors from 08/44–16/52 TD/TPH; Dial: Mitotic rate of dialysed extract receivers for thesame sample period. Key: —�—, dialysed; —�—, saline.
35
0
Mit
osis
/100
0 n
ucl
ei
20
30
25
15
10
5
Dorsal Ventral
40
* *
Fig. 4. Tongue keratinocyte mitotic rate at 12/48 TD/TPH.Effect of the dialysed fraction of the extract on dorsal andventral epithelia of the tongue. Bars represent mean�.. ofthe sample. (*), Stastically significant differences. Key: �,saline; �, dialysed.
higher molecular weight, included the microsomalfraction. With the aim of eliminating and confirm-ing the microsomal nature of F1, the extractwas ultracentrifuged at 100,000�g for 1 h. Thechromatographic profile of the supernatant after
ultracentrifugation showed no presence of the F1fraction. When the pellet was resuspended andeluted in the same chromatographic system, theF1 fraction appeared clearly defined (data notshown).
224 Cell Biology International, Vol. 23, No. 3, 1999
Effect on hepatocyte proliferation
The first group of proteins (F1 in Fig. 5) includedthe microsomal fraction of the extract. Thisfraction showed no effect on the hepatocyteproliferation as no differences between mitotic ratesfor this group and saline solution-receiving micewere found (data not shown).
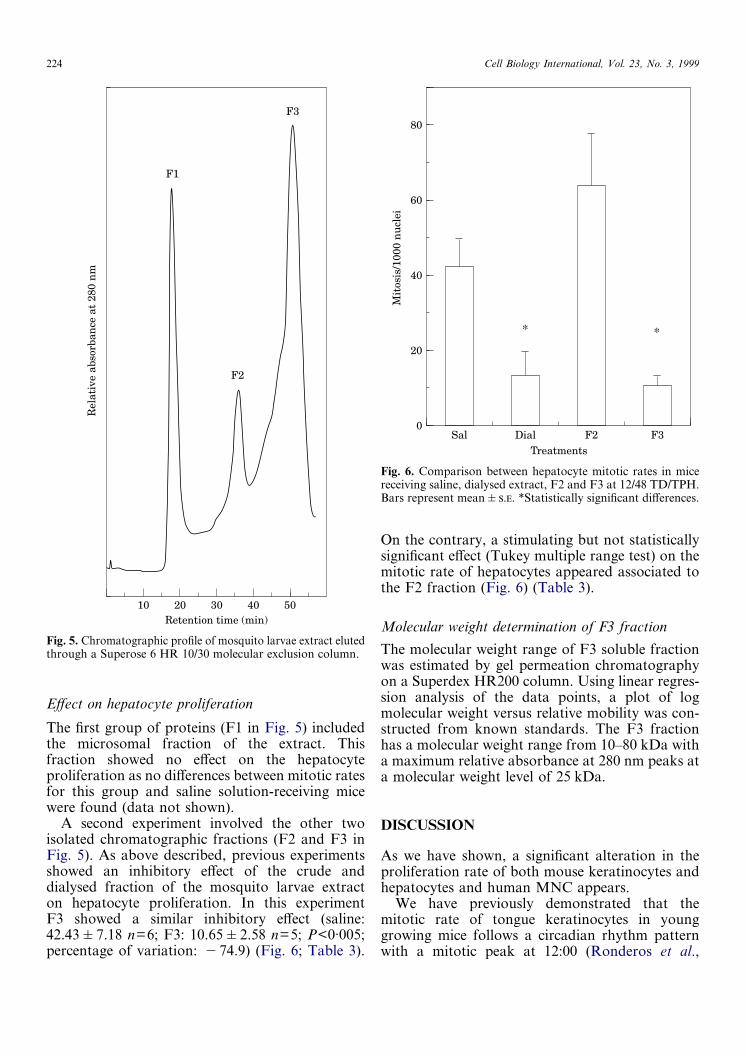
A second experiment involved the other twoisolated chromatographic fractions (F2 and F3 inFig. 5). As above described, previous experimentsshowed an inhibitory effect of the crude anddialysed fraction of the mosquito larvae extracton hepatocyte proliferation. In this experimentF3 showed a similar inhibitory effect (saline:42.43�7.18 n=6; F3: 10.65�2.58 n=5; P<0·005;percentage of variation: �74.9) (Fig. 6; Table 3).
On the contrary, a stimulating but not statisticallysignificant effect (Tukey multiple range test) on themitotic rate of hepatocytes appeared associated tothe F2 fraction (Fig. 6) (Table 3).
Molecular weight determination of F3 fraction
The molecular weight range of F3 soluble fractionwas estimated by gel permeation chromatographyon a Superdex HR200 column. Using linear regres-sion analysis of the data points, a plot of logmolecular weight versus relative mobility was con-structed from known standards. The F3 fractionhas a molecular weight range from 10–80 kDa witha maximum relative absorbance at 280 nm peaks ata molecular weight level of 25 kDa.
50Retention time (min)
Rel
ativ
e ab
sorb
ance
at
280
nm
10 20 30 40
F2
F1
F3
Fig. 5. Chromatographic profile of mosquito larvae extract elutedthrough a Superose 6 HR 10/30 molecular exclusion column.
0
Treatments
Mit
osis
/100
0 n
ucl
ei
80
60
40
20
Sal Dial F2 F3
* *
Fig. 6. Comparison between hepatocyte mitotic rates in micereceiving saline, dialysed extract, F2 and F3 at 12/48 TD/TPH.Bars represent mean�.. *Statistically significant differences.
DISCUSSION
As we have shown, a significant alteration in theproliferation rate of both mouse keratinocytes andhepatocytes and human MNC appears.
We have previously demonstrated that themitotic rate of tongue keratinocytes in younggrowing mice follows a circadian rhythm patternwith a mitotic peak at 12:00 (Ronderos et al.,

Cell Biology International, Vol. 23, No. 3, 1999 225
1996). Regarding the necessary delay existing be-tween S-phase and mitosis, the peak of prolifer-ation is coincident with the circadian variationpreviously described for the DNA synthesis in thesame cell populations of mice (Scheving et al.,1978; Scheving et al., 1979). Since the DNA syn-thesis peak occurs around 04:00, the decrease ofmitotic rate at 12/48 TD/TPH in dialysed extract-receiving mice (also significant for the wholesample period) suggests, as in hepatocytes andother cell populations previously analysed, an in-hibitory effect of the extract at G1/S restrictionpoint of the cell cycle. This effect could be relatedto some macromolecule with a molecular weighthigher than 12 kD. Since F3 also showed an inhibi-tory effect on hepatocyte proliferation, experimentstending to analyse the effect of this fraction onmouse tongue keratinocytes are now being done.
Regarding chromatographic fraction assays onliver-regenerating hepatocytes and in coincidencewith our previous reports (Ronderos et al., 1994a),an association of the extract inhibitory effect withmacromolecules was found. This effect was associ-ated to the soluble fraction with the lowest molecu-lar weight (F3) (estimated molecular weight rangefrom 10–80 kD). By taking into account the timechosen to administrate the treatments (16/28 TD/TPH, time of raising of DNA synthesis curve), theresults suggest an inhibitory effect of the extract atthe G1/S checkpoint of the cell cycle.
The experiments performed on human MNCproliferation demonstrated a biphasic effect of thecrude extract. There were both an inhibitory and astimulatory effect depending on the doses applied.No effect was seen with the lowest dose used.Furthermore, since MNC proliferation was evalu-ated by [3H]-thymidine incorporation, an alterationin the number of cells entering in the S-phase wasconfirmed. The fact that the biological effect of theextract on these cells disappeared after heating at90�C for 30 min, could suggest a protein nature ofthe molecule or molecules involved.
TGF-�, which inhibits cell proliferation actingon late G1-phase cyclin-dependent kinases (Reddyet al., 1994) and other members of the family, havebeen suggested as mediators for the regulationof DNA synthesis and differentiation in bothhepatocytes (Kay and Fausto, 1997; Russell et al.,1988; Michalopoulos, 1992; Michalopoulos andDeFrances, 1997) and lymphocytes (Ahuja et al.,1993; Altiok et al., 1994; De Jong et al., 1994; Foxet al., 1992; Fox et al., 1993; Lee and Rich, 1993;Reinhold et al., 1994). Members of this growthfactor superfamily have been also related to theregulation of the endocrine function, particularlywith respect to pituitary hormones (De et al., 1996;Jin et al., 1997; Murata and Ying, 1991; Pastorcicet al., 1995). We have shown that the dialysedextract from mosquito larvae also has a specificmodulatory effect on the rat pituitary hormonesrelease. It inhibits both prolactin and GH and stimu-lates LH and TSH release (unpublished results).
Finally, our results suggest the existence of someknown or unknown peptidic molecule modulatingcell behaviour in mammals.
ACKNOWLEDGEMENTS
Thanks are due to Mrs Rubini Crabouras and MrsGabriela Simonetto for her skilful technical assist-ance and to the members of the Insect PathologyLaboratory (CEPAVE-UNLP) for the mosquitolarvae supply.
Table 3.One way analysis of variance comparisons for mitotic rates of hepatocytes between saline,dialysed, F2 and F3 receiving mice showing general statistically significant difference between
treatments
Source ofvariation
Sum ofsquares
DF Meansquares
F-ratio
Treatment 10631.03 3 3543.68 7.56 P=0.02Residuals 8438.49 18 468.81 — —Total 19069.52 21 — — —
REFERENCES
A SS, P F, Y H, B JE, BDT, 1993. Effect of transforming growth factor-beta onearly and late activation events in human T-cells. J Immunol150(8 part 1): 3109–3118.
A A, J M, M T, K E, 1994.Biphasic effect of transforming growth factor-beta on

226 Cell Biology International, Vol. 23, No. 3, 1999
Epstein-Barr virus-induced activation of human tonsillar Bcells. Immunol Lett 40: 111–115.
D A, M TE, S RC, B N, S DK,1996. Pituitary lactotrope express transforming growth fac-tor � (TGF�) type II receptor mRNA and protein andcontains 125I-TGF� binding sites. J Endocrinol 149: 19–27.
D J R, V L RAW, R FW, S C,D P, M MD, 1994. Differential effect of trans-forming growth factor-beta1 on the activation of humannaive and memory CD4+ T lymphocytes. Int Immunol 6:631–638.
D JS, J PD, R KE, V M, HFM, 1992. Sequence, biochemical characterisation, and devel-opmental expression of a new member of the TGF-beta super-family in Drosophila melanogaster. Dev Biol 151: 491–505.
E L JM, S IL, 1964. Preparation of livertissue for the evaluation of mitotic activity. Stain Technol 39:289–294.
E L JM, D’A M, S M, B R,S JM, 1971. Circadian variations in DNA synthesis,mitotic activity and cell size of hepatocyte population ofyoung immature male mice growing liver. Virchows Archiv,Abt B Zellpath 8: 309–317.
E L JM, N RE, 1971. Circadian rhythm inthe gastric content of normal and hepatectomized mice.J Interdiscipl Cycle Res 2: 445–454.
F FE, C RJ, F HC, L RJ, M JS,N PC, 1992. Transforming growth factor-betainhibits human T-cell proliferation through multiple targets.Lymphokine Cytokine Res 11: 299–305.
F FE, F HC, D R, C S, N PC,1993. Evidence that TGF-beta can inhibit humanT-lymphocyte proliferation through paracrine and autocrinemechanisms. Cell Immunol 150: 45–58.
H F, B CP, S RH, B J, 1958. 24-hourrhythms at several levels of integration in mice on differentlighting regimens. Proc Soc Exp Biol 97: 897–900.
J L, Q X, K E, S N, S BW, KK, Y WF, J, L RV, 1997. Transforming GrowthFactor-� receptor II and p27Kip1 expression in nontumorousand neoplastic human pituitaries. Amer J Pathol 151: 509–519.
K MA, F N, 1997. Liver regeneration: prospects fortherapy based on new technologies. Mol Med Today 3:108–115.
L HM, R S, 1993. Differential activation of CD8 positiveT cells by transforming growth factor-beta 1. J Immunol 151:668–677.
L OH, R NJ, F AL, R RJ, 1951.Protein measurement with the folin phenol reagent. J BiolChem 193: 265–275.
M J, 1990. The Transforming Growth Factor-�family. Annu Rev Cell Biol 6: 597–641.
M GK, 1992. Editorial: Liver regeneration andgrowth factors: old puzzles and new perspectives. Lab Invest67: 413–415.
M GK, DF MC, 1997. LiverRegeneration. Science 276: 60–66.
M T, Y S, 1991. Transforming Growth Factor-� andactivin inhibit basal secretion of prolactin in a pituitary mono-layer culture system. Proc Soc Exp Biol Med 198: 599–605.
M AT, H FM, 1990. Homologous ofvertebrate growth factors in Drosophila melanogaster andother invertebrates. Curr Top Dev Biol 24: 289–328.
P RW, S J RD, G WM, 1987. Atranscript from Drosophila pattern gene predicts a protein
homologous to the transforming growth factor-beta family.Nature 325: 81–84.
P M, D A, B N, V W, S DK,1995. Reduction in the expression and action of transform-ing growth factor beta 1 on lactotropes during estrogen-induced tumorigenesis in the anterior pituitary. Cancer Res55: 4892–4898.
P A, S SB, H FM, 1997. Regulationof cell cycle synchronisation by decapentaplegic duringDrosophila eye development. Science 275: 203–206.
R B, H BA, H PH, 1994. Inhibition of G1phase cyclin dependent kinases by Transforming GrowthFactor-� 1. J Cell Biochem 56: 418–425.
R D, B U, B F, L U, UAJ, F HD, A S, 1994. Transforming growthfactor-� 1 (TGF-�1) inhibits DNA synthesis of PWM-stimulated PBMC via suppression of IL-2 and IL-6production. Cytokine 6: 382–388.
R P, L C, J R, A PS, P D, RDL, 1996. Control of C. elegans larval development byneuronal expression of a TGF-� homologue. Science 274:1389–1391.
R JR, E L JM, 1990. Inhibitory effect ofmosquito larvae extract on the mitotic rate of hepatocytesof young growing and adult partially hepatectomized mice.Com Biol 9: 133–142.
R JR, S AO, A CA, E LJM, 1994a. Inhibition of mouse parenchimal cells prolifer-ation by mosquito larvae extract. Cell Mol Biol 40: 1183–1188.
R JR, O A, S AO, E L JM,1994b. Further action of mosquito larvae extract on themitotic rate of mouse cell populations. Bio Cell 18: 51–57.
R JR, 1996. Early effect of mosquito larvae extract onmouse cells proliferation In Vivo. Cell Biol Int 20: 249–254.
R JR, R FL, O A, 1996. Mosquitolarvae extract induces mitotic rate alterations of mice tonguekeratinocytes along a circadian rhythm period. Cell Biol Int20: 567–572.
R WE, C RS, O AJ, M HL, 1988.Type � transforming growth factor reversibily the earlyproliferation response to partial hepatectomy in the rat.Proc Natl Acad Sci USA 85: 5126–5130.
S LE, B ER, P JE, T-H T, 1978.Circadian variation in cell division of the mouse alimentarytract, bone marrow and corneal epithelium. Anat Rec 191:479–486.
S LA, Y YC, T TH, S LE, 1979.Circadian phase-dependent stimulatory effects of EpidermalGrowth Factor on deoxyribonucleic acid synthesis in thetongue, esophagus and stomach of the adult male mouse.Endocrinol 105: 1475–1480.
S JM, M FR, B AF, E L JM,1985. Variations in DNA synthesis and mitotic indices inhepatocytes and sinusoid litoral cells of adult intact malemouse along a circadian time span. Chronobiol Int 2: 161–168.
T X, F AL, P RW, 1994. The Drosophilasaxophone gene: A serine-Threonine kinase receptor of theTGF-beta superfamily. Science 263: 1756–1759.
T W, 1937. Cell size in relation to the growth andmetamorphosis of the mosquito, Aedes aegypti. J Exp Zool76: 467–489.
V CA, E L JM, 1971. Circadian rhythm inthe feeding activity of Oxymycterus rutilans. Role played bylight and food availability. J Interdiscipl Cycle Res 2: 73–77.
Copyright © 2022 FDOKUMEN




















