Parental care and the transition to independence of Spanish Imperial Eagles Aquila heliaca in...
13
IBIS 129: 212-224 Parental care and the transition to independence of Spanish Imperial Eagles Aquila heliaca in Doiiana National Park, southwest Spain JUAN CARLOS ALONSO," LUIS MARIAN0 GONZALEZ,? BORJA HEREDIAf & JOSE LUIS GONZALEZt "Muse0 Nacional de Ciencias Naturales, Jf. GutiPrrez Ahascal, 28006 Madrid, f ICONA , Suhdireccidn de Recursos Naturales Renovables, Gran Via de S. Francisco 35, 28005 Madrid and $Muse0 Nacional de Ciencias Naturales, J. Gutie'rrez Abascal2, 28006 Madrid, Spain Accepted 20 June 1986 The behaviour of five broods of radio-tagged Spanish Imperial Eagles Aquila helium udalberti was studicd in Dofiana National Park, Spain during the post-fledging period. The distance between perching sites and nest, the mean flight duration and distance, the percentage of time spent flying and the home range all increased exponentially with age. As the young got older, the parents spent less time in their vicinity. Young were not seen hunting, but depended on their parents for food. They begged and chased their parents throughout the post-fledging period, with hrgher intensity at the end. Nevertheless, the adults became progressively more reluctant to feed them, as reflected in the dccrease in feeding frequency and in the number of approaching flights towards the young. At the end of the post-fledging period, adults often performed aerial displays and frequently chased their offspring. The age of independence of the different young studied varied between 123 and 145 days. The correlations between individual independence and the dates when the young were last fed by their parents, and when the highest intensity of parental aggressive behaviour occurred, were higher than correlations with the variables related to the maturation of flying. Therefore, it is suggested that parental 'meanness' and aggressive behaviour may be the factors determining the date of juvenile independence and dispersal from the home territory. Although young fledged birds depend on their parents for a period that is often longer than the nestling period (Skutch 1976), studies of parental care during post- fledging are scarce compared with those during the nestling phase. The difficulty of observing young after they have left the nest may be the main reason why few quantitative studies have been published (Davies 1976, Smith 1978, Smith & Merkt 1980, Moreno 1984). This is particularly true of raptor species (Brown 1966, Johnson 1973, Harper 1974, Kussman 1977, Sherrod 1983). Furthermore, the process of transition to independence and the factors leading to family breakup are still poorly understood. As fledglings get older they inevitably become more self- sufficient and require less parental care. However, from Trivers' (1 974) analysis of parent-offspring conflict, we would expect that the parents should initiate the process of independence by progressively reducing their effort. Working with Great Tits Parus major, Davies (1978) provided experimental evidence for the hypothesis that a reduction in the parental investment is important in forcing the young to independence. Recent field studies with Northern Wheatears Oenanthe oenanthe and Blackbirds Turdus merula support this view (Moreno 1984, Edwards 1985). Nevertheless, published information on this process in raptors is contradictory. While in some species young gradually become independent without a decrease in parental feeding, in others the period is terminated by the adults driving the young away (Brown 1976, Newton 1979). In the present paper, we describe the behaviour
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Parental care and the transition to independence of Spanish Imperial Eagles Aquila heliaca in...

I B I S 129: 212-224
Parental care and the transition to independence of Spanish Imperial Eagles Aquila heliaca in Doiiana
National Park, southwest Spain
J U A N C A R L O S A L O N S O , " L U I S M A R I A N 0 G O N Z A L E Z , ? BORJA H E R E D I A f & J O S E L U I S G O N Z A L E Z t
"Muse0 Nacional de Ciencias Naturales, J f . GutiPrrez Ahascal, 28006 Madr id , f ICONA , Suhdireccidn de Recursos Naturales Renovables, Gran V i a de
S . Francisco 35 , 28005 Madrid and $Muse0 Nacional de Ciencias Naturales, J. Gutie'rrez Abascal2 , 28006 Madrid, Spain
Accepted 20 June 1986
T h e behaviour of five broods of radio-tagged Spanish Imperial Eagles Aquila helium udalberti was studicd in Dofiana National Park, Spain during the post-fledging period. T h e distance between perching sites and nest, the mean flight duration and distance, the percentage o f time spent flying and the home range all increased exponentially with age. As the young got older, the parents spent less time in their vicinity. Young were not seen hunting, but depended on their parents for food. They begged and chased their parents throughout the post-fledging period, with hrgher intensity at the end. Nevertheless, the adults became progressively more reluctant to feed them, as reflected in the dccrease in feeding frequency and in the number of approaching flights towards the young. At the end o f the post-fledging period, adults often performed aerial displays and frequently chased their offspring. T h e age of independence of the different young studied varied between 123 and 145 days. T h e correlations between individual independence and the dates when the young were last fed by their parents, and when the highest intensity of parental aggressive behaviour occurred, were higher than correlations with the variables related to the maturation of flying. Therefore, it is suggested that parental 'meanness' and aggressive behaviour may be the factors determining the date of juvenile independence and dispersal from the home territory.
Although young fledged birds depend on their parents for a period that is often longer than the nestling period (Skutch 1976), studies of parental care during post- fledging are scarce compared with those during the nestling phase. The difficulty of observing young after they have left the nest may be the main reason why few quantitative studies have been published (Davies 1976, Smith 1978, Smith & Merkt 1980, Moreno 1984). This is particularly true of raptor species (Brown 1966, Johnson 1973, Harper 1974, Kussman 1977, Sherrod 1983). Furthermore, the process of transition to independence and the factors leading to family breakup are still poorly understood. As fledglings get older they inevitably become more self- sufficient and require less parental care. However, from Trivers' (1 974) analysis of parent-offspring conflict, we would expect that the parents should initiate the process of independence by progressively reducing their effort. Working with Great Tits Parus major, Davies (1 978) provided experimental evidence for the hypothesis that a reduction in the parental investment is important in forcing the young to independence. Recent field studies with Northern Wheatears Oenanthe oenanthe and Blackbirds Turdus merula support this view (Moreno 1984, Edwards 1985). Nevertheless, published information on this process in raptors is contradictory. While in some species young gradually become independent without a decrease in parental feeding, in others the period is terminated by the adults driving the young away (Brown 1976, Newton 1979). In the present paper, we describe the behaviour

I987 M A T U R A T I O N O F I M P E R I A L EAGLE Y O U N G 213
of young Spanish Imperial Eagles Aguila heliaca adalberti during the period of post- fledging dependency, analyse parent-offspring interactions in an attempt to identify the factors leading to offspring independence and test the hypothesis that it is the parents who mainly determine the date of juvenile independence.
Study area
The study was carried out at Doiiana National Park, southwest Spain (37"N, 6"30'W), at 0-40 m above sea level. The area consists of three main biotopes: (a) Mediterranean scrubland of Halimium spp., Erica spp., with scattered cork oaks Quercus suber and stone pines Pinuspinea; (b) marshes covered by Scirpus spp.; and (c) coastal dunes with Corema album and Ammophila spp. The climate is mesomedi- terranean, with humid, mild winters and dry, hot summers. The average tempera- ture during the study was 23°C. No rainfall was registered during the study. For a detailed description of the area see Valverde (1958) and Allier et al. (1974).
Materials and methods The behaviour of young Spanish Imperial Eagles was observed during July to September 1984. Four broods of two (A, C, D and E) and one of one young (B) were marked individually with radio-transmitters and coloured wing-tags. The latter were yellow patagial strips of saflag attached as described by Kochert et al. (1983). The telemetry equipment consisted of solar-powered transmitters, LA 12 D S receivers, and three element hand-held yagi antennas, all from AVM-Instruments Co., Livermore, California, USA. The transmitter packages weighed 75-80 g, approximately 2-2.5% of the body weight of the bird, and had an average lifetime of 3-5 years. All operated in the 148.000-149.000 KHz range. The packages were attached to the eagles with a back-pack harness, as described by Dunstan (1977). No abnormal behaviour of the young was observed associated with the harnesses or transmitter packages. Reception ranges were about 5-8 km when the bird was perched on a tree, and 20-30 km when the bird was flying.
When the young left the nest, they were monitored by two teams each of two observers, with binoculars and telescope 80-1 30 x . Regular observations began on Day 78 (after hatch) and ended with their dispersal. If visual contact was eventually lost the young were relocated by triangulation, as described by Kenward (1 980). Once the young started to fly long distances, they were tracked from vehicles and a Dornier aircraft. The whole day was spent in the field, and a daily total of 88-905 minutes of continued observation of behaviour was recorded. The total amount of observation time for all the young together was 759 hr 31 minutes. The different family groups were observed with a variable periodicity throughout the dependency period. Additionally, every third or fourth day the location of each young was determined by radiotelemetry. Observations of behaviour were recorded on tape, including the following data: individual, time of day, location (on aerial photographs scale 1 : 5000 and 1 : 30000 of the study area), whether the bird was resting silent or calling, flying alone or with a parent or being fed by them, and interactions between young and siblings, parents or another bird. Perching sites of young and adults were individually recognized on the photographs and distances between each young and its sibling, parents and nest were recorded. Distance and duration of each flight were also measured. Home ranges were determined with a digital planimeter, following the flight lines that encompass the activity area of the birds on aerial photographs. Although it was not possible to recognize adults individually, their activities were also recorded whenever they were visible from our observation points, in the same

214 J . c . ALONSO e t al . I B I S 129
way as for young birds. As a rule, the adult closest to the young was defined as Adult 1 and the furthest as Adult 2. When only one adult was present it was considered as Adult 1 ,
Changes in the variables related to the location and flying behaviour of the young were analysed as follows. The daily means for each variable were plotted on an age- axis and the best-fitting lines for each variable and each juvenile were obtained by the least-squares method. The slopes of the resulting lines were then compared individually with the age at independence. Feeding frequency varied between days, depending on whether or not the young were fed. Due to the low frequency of feedings, the data are presented in ten-day periods for each individual. Those days in which the observation of a family group did not exceed six hours were excluded from the feeding frequency analyses.
Results
Development of flying activity and home range The young left the nest at approximately Day 65 (Nest C), 72 (Nest B), 73 (Nest A), 74 (Nest D) and 78 (Nest E). The variables related to their flying behaviour and activity (daily means for distance to nest, flight duration, percentage time spent flying, distance between siblings and home range) were strongly intercorrelated and increased exponentially with age in all the young studied (see Fig. 1). This is closely related to the appearance of soaring flights and their increasing frequency after the first 20 days of the post-fledging period (Table 1). At the end of the post-fledging period immediately prior to dispersal, the young flew long distances for several hours without their parents or siblings, and usually returned to roost in the home territory.
Interactions between parents and young
Time spent by the parents in the post-jedging area
Immediately after the young had left the nest, the adult eagles spent much time near them. The percentage of time spent by one or both adults in the vicinity of the young decreased with age (rs = - 0.96, P < 0.001, and rg = - 0.77, P < 0.05). Thus, the time the young spent alone increased significantly with age (rs = 0.89 P < 0.01; Table 2). Both adults tended to spend less time perched near the young (rg = 0.95, P < 0,001; Y S = 0.95, P < 0.001), but not to spend more time flying over the area ( 1 5 = - 0.36, n.s.; rs = 0.36, n.s.). Consequently, the general decrease observed in the time spent by the adults in the post-fledging area was due to their spending more time away from the home territory.
Distance between parents and offspring
The average distance between young and parents when they perched nearby increased to Day 115 for the closest adult and to Day 125 for the furthest. Distances then decreased progressively to similar values as those found in the beginning of the post-fledging period (F6,70 = 2.96, P < 0.05, and F6,47 = 8.02, P < 0.001, with daily weighted mean distances between each young and, respectively, the closest and furthest adult, Table 2). During the first half of the post-fledging period, adults tended to perch further from the young, spending progressively less time perching in the area. Later, the time the adults spent in the vicinity of the young was reduced to

M A T U R A T I O N O F I M P E R I A L EAGLE YOUNG
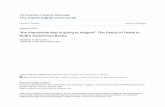
c2 A1
DI C I
D2
~, 85 95 105 115 125 I35 145
215
Age of young (days)
Figure 1. As an example to illustrate the exponential increasing trend of behavioural variables of young Imperial Eagles during the post-fledging period, we show here the logarithmic regressions with age of (a) the daily amount of time spent flying by the young ( 1 2 2 = 0.70, P i 0.001 for young Al ; ~ 1 4 = 0.69, P < 0.01 for A2; r5=0.87, P<O.Ol for B; r h = O . h S , n.s. for C1; r2=0.81, n.s. for C2; ~9=0 .73 , P<0 .02 for D1; r ~ = O . 3 7 , n. s . for D2), and (b) their home range (v22=0.62, P<O,O01 for Al ; v14=0.61, P<O.O2 for A2; r s=0 .88 ,P<0.01 forB;r~=0~52,n.s.forC1;~~=0~75,n.s.forC2;rg=0~87,P<O~OOl forDl ; r s=0 .75 , P i 0.01 for D2). Regression lines join the earliest and latest observation days. T h e curves for the other variables were also exponential and, most of them, highly significant.
little more than the time needed for feeding, and even during feedings it was the young that approached the adult. Thus, although there was a decrease in the distance between adults and offspring while perched, at the end of the period the reduction in the time they kept in visual contact was even greater, probably because the adult was already reluctant to remain near the young.
Begging and feeding behaviour
Begging consisted of calls lasting from a few seconds to a few minutes, alternating

216 J . c. ALONSO et a l . I B I S 129
Table 1. Changes wi th age of young Imperial Eagles in the proportion of main fl ight types,figures are weighted means f o r all young birds
Flights Age, days considered Wing-flapping Gliding Soaring
after hatching n ( O " ) ("") ( " 0 )
78 -85 69 95.7 4.3 0.0 86-95 86 91.9 8.1 0.0 96-105 102 84.3 3.9 11.8
106-115 63 57.1 9.5 33.3 116-125 40 62.5 2.5 35.0 126-135 29 72.4 0.0 27.6 136 150 36 47.2 5.7 47.1
Notes: T h e trends for wing-flapping and soaring are significant ( r ~ = 0-86, P < 0.02 and r5 = 0.92, f i 0.01).
with resting periods of similar duration. When one of the adults approached, especially if carrying prey, the young called excitedly and showed the typical begging attitude, with fallen wings and neck stretched towards the adult. We classed as one begging activity each behavioural sequence comprising various consecutive begging and silent periods. If different behaviour (e.g., preening, flying) was performed between two such begging sequences, these were considered as two different begging activities.
The frequency of begging by the young while perched tended to be higher at the beginning, remained constant through most of the post-fledging period, and slightly increased at the end, although these trends were not significant (Fh,s() = 0.27, n.s., Fig. 2 ) . The frequency of pursuit flights by young birds toward perched or flying adults followed a similar pattern, with significant variations throughout the dependence period ( F h , 8 0 = 2.69, P < 0.05). In contrast, the frequency with which parents approached their young, whether they brought food or not, decreased steadily through the dependency period (F~,Ro= 2.27, P < 0.05).
Both male and female adults supplied prey. Parental feeding frequency decreased with age in all five birds for which enough data were obtained (Fig. 3). The decreasing trend was statistically significant in four (r2 = - 0.97, P < 0.05 for A2; r3 = - 0.97, P < 0.01 for C1; r3 = - 0.95, P < 0.02 for C2; r2 = - 0.98, P < 0.05 for B). Furthermore, food transfers varied with age. Initially a parent always carried prey to the place where the young were perched, tearing and offering them pieces of meat, as during the nestling period (Meyburg 1975). The frequency of this kind of food transfer decreased with age. Soon, the adults began to leave food at a distance from the young who would fly to the prey. Both types of food transfer may be considered as parent-initiated and were gradually substituted by a third type (rg = 0.95, P < 0.001) in which the young bird would fly to the place where the parent had landed with food. The adult would then fly away immediately, leaving the young to feed alone. On two occasions a juvenile was even attacked when approaching a feeding adult, although the latter finally let it feed alone on the remains of its meal.
Both siblings generally ate together from the same prey while they were under 95 days of age (75% of the cases). Thereafter they usually ate separately (75n/b of the cases), either from different prey or from the same prey but one after the other. On one occasion, on Day 80, young A1 offered prey and fed his sibling, A2, several times

M A T U R A T I O N OF I M P E R I A L EAGLE Y O U N G

218
0.3
c -
a,
i
a
- 2 0.2-
5 z 0.1 t
J . c . A L O N S O e t a l . -
P /
1 ,n , 4’
.O‘ / 2’ /
j ’ / . ,. ‘0- . .. .. :\ , -0.
‘ \ +- 0- : - -o- - 1.8- - -. 0 -
a- 0
I I I y o - * 4 0 I
I B I S 129
I00 c
$ - L
c W
VL
e ;; 50 0 - L 0
aJ ). c a
85 95 105 115 125 135 150
Age of young (days)
Figure 3 . Changes with age of Imperial Eagles in (a) the average number of feedings per young and per day of observation; two symbols of the same kind (open and black) represent siblings: .=young A t , O=youngA2,*=youngB, r = y o u n g C l , ~ = y o u n g C 2 , m = y o u n g D l , O=youngD2, r = y o u n g E l , V =young E2; data for nest D during the first part of the dependency period were not considered, due to the short observation periods; and (b) the type of food transfer: the adult carrying the prey to the place where the young is perched (white bars); the adult leaving prey at a certain distance from the young (striped bars); and young flying towards the adult just arrived with prey (black bars). Here data for all the young are pooled. T h e values represented are ten-day means. Total observation time, see Table 2.
Figure 2. Changes with age in the frequency of approaching flights of adult Imperial Eagles towards their offspring (black dots), frequency of pursuit flights of young towards adults (open dots), and frequency of begging activities of young while perched (open squares). Data for all the young pooled and weighted as ten-day means. Total observation time, see Table 2.

rabl
e 3.
Inte
rind
ivid
ual
vari
atio
ns in
the
rate
s of
incr
ease
(1
) of
vari
ous
para
met
ers
rela
ted
to l
ocat
ion
and
flyi
ng b
ehao
iour
of
youn
g Im
peri
al E
agle
s, a
ge of
term
inat
ion of
pare
ntal
fe
edin
g, o
ccur
renc
e of
pare
ntal
cou
rtsh
ip a
nd a
ggre
ssiv
e be
havi
our
and
age of j
uven
ile i
ndep
ende
nce
Rat
e of
inc
reas
e (1
) wit
h ag
e of
the:
A
ge o
f
You
ng
.4 1 .4 2 B c1
c
2
D1
D2
El
E2
Dis
tanc
e to
nes
t
0.01
47
0.00
96
0.02
68
0-01
10
0.01
13
0.01
42
0.01
71
0.01
46
0.01
42
Dis
tanc
e be
twee
n si
blin
gs
0.04
09
0.04
09
0.02
68
0,02
68
0.02
06
0,02
06
0.02
32
0,02
32
-
Mea
n fl
ight
dur
atio
n
0.04
56
0.04
24
0.04
18
0.03
26
0.03
10
0.03
57
0.02
27
Per
cent
age
tim
e fl
ying
0.05
36
0.05
82
0.04
54
0.02
34
0.02
8 1
0.02
26
0.01
30
Hom
e-ra
nge
0.06
93
0.07
50
0.05
51
0.05
09
0.04
65
0.05
68
0.04
1 5
Age
of
firs
t E
nd o
f H
ighe
st c
ourt
ship
so
arin
g fl
ight
pa
rent
al f
eedi
ng
beha
viou
r of
adul
ts
Fir
st a
ttac
ks
pare
nt-o
ffsp
ring
97
97
107
116
122
115
138
115-
125
105-
115
135-
145
125-
1 35
125-
135
145-
1 50
14
5-15
0 14
5-15
0 14
5-15
0
too
100
113
136
136
138
138
147
147
~~
98
100
113
135
135
138
138
137
150
123
143
136
137
144
144
142
145 -
'oot
e: (1
) As e
stim
ator
of t
he ra
te o
f inc
reas
e of e
ach
para
met
er, w
e us
ed th
e re
gres
sion
coef
fici
ent
-log
b - o
f the
exp
onen
tial
line
s lo
gy =
log
a +
(1og
b)x o
btai
ned
by lo
gari
thm
ic
tran
sfor
mat
ion
from
the
exp
onen
tial
cur
ves
y=ab
" (
see
text
and
Fig
. I)
0 z 4
0
C z 0

2 20 J . c . A L O N S O e t a2. I B I S 1 2 9
between 0813 and 1025 hr. A1 tore pieces of meat, which were transferred beak to beak to A2. This behaviour has also been observed in the young of other raptors (Brown 1976, Sherrod 1983). Aggression between siblings was not seen, apart from a few instances during feedings.
Parental aggression towards young
Towards the end of the post-fledging period, adult aggression was observed on several occasions and studied in all five family groups. It took the form of chasing flights and stoops toward the young, sometimes when the latter were perched, but mostly when they were flying. Often the adults hit the young, with their talons interlocked, making them lose their balance and lose height.
This behaviour has been observed in other raptor species (Rowe 1947, Cade 1960, Gargett 1972,1975, Steyn 1973, Sherrod 1983, Robertson 1984) and is usually interpreted as aggression from adults to young. The first signs of aggression were highly correlated in time with the highest intensity of autumn aerial display in the different pairs studied (r7 = 0.98, P < 0.01), as well as with the termination of parental feedings (r7 = 0.90, P < 0.01 ; Table 3). The aerial displays and aggression from adult to young took place more or less regularly from the date they first occurred until the end of the post-fledging period.
Hunting behaviour of the young During our observations, none of the young was seen clearly to be hunting. We observed only twice what could be interpreted as hunting attempts by the young, both in the absence of the parents:
On 29 July, at 0624 h, young A1 at Day 89 flew straight from tree to ground, where we saw him moving and flapping two or three times, jumping and then staying on the ground until 0645 h, when he flew to the tree, apparently with a full crop.
On 18 August at Day 83, young B also flew several times to the ground, at 0925, 0945, 1126, 1134 and 1200 h, staying on the ground for 180, 45, 60 and 30 s, respectively. He was not seen flapping his wings or jumping, so he could have been eating food remains dropped at a previous feeding. Furthermore, between the fourth and the fifth time he was fed by one of the adults at 1145 h, so probably he was not hungry enough to try hunting for himself.
We consider that these observations, given the large number of hours in the field, are not enough to conclude that the young hunt for themselves during the post- fledging period.
Dates of independence of the young and relation to other variables The ages in which different young dispersed varied between 123 and 145 days (Table 3). This was considered as the date of independence when the young moved permanently from the home range or stayed several days up to 135 km away from it. Normally, siblings reached independence on similar dates.
The dates on which dispersal of the different young took place were correlated significantly with the dates on which they were last fed by the adults (rg=0.80, P < 0*05), as well as with the dates on which first aggressions from adults were seen (rg=O*75, P < 0.05). The correlation between date of dispersal and rate of increase (see Table 3, for definition) of the distance between siblings was also significant (r5 = - 0.98, P< 0.01). However, no significant correlations occurred between the date of dispersal and the rates of increase of distance to the nest (r4 = 0.28, n.s.), mean

I987 M A T U R A T I O N O F I M P E R I A L EAGLE Y O U N G 221
flight duration (Y+ = - 0.56, n.s.), onset of soaring flight (rq= 0.64, n.s.), time spent flying (yq = - 0.66, n.s.) and home range size ( ~ 4 = - 0.69, n.s.). Therefore, it seems that the rate of increase of variables relating to the location of the young with respect to the nest, or those that reflect the improvement of flying ability, are less correlated with the date of independence than are the variables that reflect interactions between parents and offspring, i.e., feeding frequency and aggressive behaviour.
Discussion The results described in this paper support the view that the young birds ‘want’ more parental investment than the adults are ready to give (Trivers 1974). Parental ‘meanness’ increases through the period of dependency and adults eventually decide to end the parent-offspring conflict and behave aggressively towards their young. This behaviour finally causes the young, who are already skilled, to leave. A conflict of interests is clear, since the general investment of the parents decreases with the age of young, while the young keep seeking the same or more parental investment. The adults progressively decrease the time spent near the young, the frequency of feeding and the proportion of parent-initiated feeds. Additionally, the adults progressively increase the distance between them and the young. Simultaneously, an increase is observed at the end of the dependency period in the begging activity of the young while perched, the frequency of pursuit flights toward their parents and the proportion of young-initiated feeds, i.e., flights toward the adults and solicitation. However, it seems that the young do not hunt for themselves during the post- fledging period, as observed in some raptor species (Gargett 1972, Newton 1979, Beske 1982) but not in others (Brown 1966, Kussmann 1977, Newton 1979, Cramp & Simmons 1980, Ratcliffe 1980, Sherrod 1983). Consequently, as the frequency of parental feeds decreases, the stress on the young increases. Greater competition between siblings is seen in the gradual increase in the distances between them, as well as by a higher proportion of feeds where only one of the siblings eats or one eats after the other. A decrease in feeding frequency has been observed in other species (Morehouse & Brewer 1968, Davies 1976, Skutch 1976, Moreno 1984, Edwards 1985). At the end of the dependence period, the adults become aggressive towards their young, until after several days the latter finally disperse. Aggression from adults has been observed in several other species towards the end of the post-fledging period (Gargett 1972, 1975, Steyn 1973, Holleback 1974, Sherrod 1983, Robertson 1984) although in yet other species independence happens without such aggression (Lind 1965, Ashmole & Tovar 1968, Skutch 1976, Newton 1979, Scott 1980, Moreno 1984).
It seems, therefore, that in the Spanish Imperial Eagle it is the adults who finally impose independence on their offspring, by cutting their rations and behaving aggressively towards them. The alternative process, in which the young decide the date of independence, seems to be the rule in certain large tropical eagles (Brown 1976). If this were true in our case, one might expect that aggression would not occur, the young would show more indifference towards the adults at the end of the post-fledging period instead of continuing to solicit food, the young would know how to hunt and the profitability of self-foraging should exceed that of begging. The first three assumptions are fulfilled by some tropical raptors who have a very long post- fledging period (Brown & Amadon 1968, Brown 1976, Mader 1981, 1982, Vernon 1984), and all of them by some other bird species (Davies 1976, Skutch 1976, Sauey 1979, Moreno 1984).
The coincidence in time of adult aerial displays and aggression to the young suggests that autumnal sexual recrudescence could be the factor leading to

222 J . c . ALONSO e t a l . I B I S 129
aggressive behaviour. There is also a clear correspondence between the date of juvenile independence, the cessation of feeding and the aggressive behaviour of the parents. However, the correlations are not so clear between the date of independence and the variables relating to the maturation of flying. This provides further support for the view that it is the behaviour of the adults that proximately determines the independence of the young. By the time the adults stop feeding their offspring and start chasing them, the latter have acquired enough experience and knowledge of flying.
Final independence in the young Spanish Imperial Eagle takes place relatively early in comparison with other large sedentary eagles for which comparable data exist (Newton 1979). The ultimate causes that have favoured this early break-up must be mainly ecological (Trivers 1974, Wilson 1975, Krebs & Davies 1981) and are probably related to the necessity of the adults to devote their whole time to foraging and recovering before starting a new reproductive cycle. It is a fact that in many passerine birds that breed more than once a year, last broods tend to receive more prolonged parental care than first broods, which are often neglected as the parents turn their attention to another set of eggs (Skutch 1976). Our data support the hypothesis that high breeding rates are correlated with short post-fledging periods (Newton 1979), as the productivity of the Imperial Eagle in Spain is about 2.1 young per successful pair per year (own data), which is high compared with other large eagles, even of temperate areas (Neston 1979, Cramp & Simmons 1980). In species inhabiting more stable or predictable environments and/or those where predation is more pronounced, as in the tropics or some marine ecosystems, the reproductive rate is lower (Cody 1966) and the post-fledging period longer, both in raptors (Brown 1976, Newton 1979, Mader 1981,1982) and in other bird species (Ashmole & Tovar 1968, Dunn 1972, Fogden 1972, Feare 1975).
We are indebted to J. Alonso, B. Arroyo, L . Bergrem, J . Bustamante, M. Caballero, R. Cadenas, L. Dominguez, L. Fernandez, M. Fernandez, M. Ferrer, R. Heredia, E. Johnson, J . Lahoz, A. Ortega, E. Pullman, and M. de la Riva for their help in the field, and to L. Garcia, R. Coronado, L. Berbiela, J . de Benito, P. Pereira, A. Ramhn, F. Robles and the rest of the members of the conservation staff of the Doiiana National Park for their help and facilities provided. Our thanks are also due to F. Hiraldo, F. Gonzalez-Bernaldez, J. Castroviejo and C. Morillo for their encouragement and support, and to F. Hiraldo, I. Newton, and two anonymous referees for reading and criticizing the manuscript. We specially appreciate the invaluable help provided by F. Palacios. The 407 squadron of the Spanish Air Force and its Captain General provided the aircraft for aerial control. M. Merino drew the figures and B. Thomas revised the English text. The material and the field work were supported financially by the Direccion General del Medio Ambiente, the Instituto Nacional para la Conservaci6n de la Naturaleza, and C.S.1.C.- C.A.I.C.Y.T. Project No. 944. During the study the authors benefited from fellowships of the Instituto Nacional de Investigaciones Agrarias and Consejo Superior de Investigaciones Cientificas.
References
ASHMOLE, N.P. & TOVAR, H. 1968. Prolonged parental care in Royal Terns and other birds. Auk 85: 9&
ALLIER, C., GONZALEZ-BERNALDEZ, F. & RAMIREZ-DIAZ, L. 1974. Mapa ecoliigico de la Reserva Biologica 100.
de Doiiana. Division de Ciencias. Sevilla: CSIC. BESKE, A.E. 1982. Local and migratory movements of radio-tagged juvenile Harriers. Raptor Res. 16(2):
39-53, BROWN, L. 1966. Observations on some Kenya eagles. Ibis 108: 531-572. BROWN, L. 1976. Eagles of the World. London: David and Charles. BROWN, L. & AMADON, D. 1968. Eagles, Hawks and Falcons of the World. London: Hamlyn

I987 M A T U R A T I O N O F I M P E R I A L EAGLE Y O U N G 223
CADE, T.J. 1960. Ecology of the Peregrine and Gyrfalcon populations in Alaska. Univ. of Calif. Publ.
CODY, M.L. 1966. A general theory of clutch size. Evolution 20: 174-184. CRAMP, S. & SIMMONS, K.L. 1980. The Handbook of the Birds of the Western Paleartic. Vol. 11. Oxford:
Oxford University Press. DAVIES, N. B. 1976. Parental care and the transition to independent feeding in the young Spotted
Flycatcher (Muscicapa striata). Behaviour 59: 280-295. DAVIES, N.B. 1978. Parental meanness and offspring independence: an experiment with hand-reared
Great Tits Parus major. Ibis 120: 509-514. DUNN, E.K. 1972. Effects of age on the fishing ability of Sandwich Terns Sterna sandwicensis. Ibis 114:
360-366. DUNSTAN, T.C. 1977. Types and uses of radio-packages for the North American Falconiform and
Strigiform birds. In Long, E.M. (ed) Proc. I . Int. Conf. Wildl. Biotelemetry. pp. 30-39. Wyoming: Laramie.
EDWARDS, P.J. 1985. Brood division and transition to independence in Blackbirds Turdusmevula. Ibis 127:
FEARE, C.J. 1975. Post-fledging parental care in Crested and Sooty Terns. Condor 77: 368-370. FOGDEN, M.P. 1972. The seasonality and population dynamics of equatorial forest birds in Sarawak. Ibis
GARGETT, V. 1972. Observations at a Black Eagle nest in the Matopos, Rhodesia. Ostrich 43: 77-108. GARGETT, V. 1975. The spacing of Black Eagles in the Matopos, Rhodesia. Ostrich 46: 1 4 3 . HARPER, J.F. 1974. Activities of fledging Bald Eagles in North Central Minnesota. M.S. thesis. Illinois:
HOLLEBACK, M. 1974. Behavioral interactions and the dispersal of the family in Black-capped Chickadees.
JOHNSON, S.J. 1973. Post-fledging activity of the Red-tailed Hawk. Raptor Res. 7: 4 3 4 8 . KENWARD, R.E. 1980. Radio monitoring birds of prey. In Amlaner, C. & MacDonald, D. W. (eds), A
Handbook on Biotelemetry and Radio-tracking, pp. 97-104. Oxford: Pergamon Press. KOCHERT, M.N., STEENHOF, K. & MORITSCH, M.Q. 1983. Evaluation of patagial markers for raptors and
ravens. Wild. SOC. Bull. 1 1 : 271-281. KREBS, J.R. & DAVIES N.B. 1981. Behavioral Ecology: an evolutionary approach. Oxford: Blackwell
Scientific Publications. KUSSMAN, J.V. 1977. Post-fledging behavior of the northern Bald Eagle (Haliaetus leucocephalus
alascanus) in the Chipewa National Forest. M. Sc. thesis. Minnesota: University of St. Paul. LIND, H. 1965. Parental feeding in the Oystercatcher (Haematopus ostralegus). Dansk Om. Foren.
Tidsskr. 59: 1-31. MADER, W.J. 1981. Notes on nesting raptors in the llanos of Venezuela. Condor 83: 48-51. MADER, W.J. 1982. Ecology and breeding habits of the Savanna hawk in the llanos of Venezuela. Condor
MEYBURG, B.U. 1975. On the biology of the Spanish Imperial Eagle (Aquila heliaca adalberti). Ardeola 21:
MOREHOUSE, E.L. & BREWER, R. 1968. Feeding of nestling and fledging kingbirds. Auk 85: 44-54. MORENO, J . 1984. Parental care of fledged young, division of labor and the development of foraging
NEWTON, I. 1979. Population ecology of raptors. Berkhamsted: T & A D Poyser. RATCLIFFE, D. 1980. The Peregrine Falcon. Berkharnsted: T & A D Poyser. ROBERTSON, A. 1984. Post-fledging dependence periods of Cape Vultures at the Potberg colony. Proc. 11.
ROWE, E.G. 1947. The breeding biology of Aquila verreauxi. Ibis 89: 576-606. SAUEY, R.T. 1979. Feeding biology of juvenile Siberian Crane in India. In Lewis, J. C. (ed), Proc. Int.
SCOTT, D.K. 1980. Functional aspects of prolonged parental care in Bewick’s Swan. Behaviour 28: 938-
SHERROD, S.K. 1983. Behavior of fledgling Peregrines. New York: The Peregrine Fund, Ithaca. SKUTCH, A.F. 1976. Parent birds and their young. Austin, Texas: University of Texas Press. SMITH, J.N. 1978. Division of labour by Song Sparrows feeding fledged young. Can. J . Zool. 56: 187-191. SMITH, J.N. & MERKT, J.R. 1980. Development and stability of single parent family units in the Song
STEYN, P. 1973. Eagle days. London: Purnell & Sons. TRIVERS, B.L. 1974. Parent-offspring conflict. Am. 2001. 14: 249-264.
ZOO^. 63(3): 151-290.
42-59.
114: 307-343.
Western Illinois University.
Wilson Bull. 86: 4 6 6 4 6 8 .
84: 261-271.
241-283.
techniques in the Northern Wheatear (Oenanthe oenanthe). Auk 101: 741-752.
Symp. on Afr. Predatory Birds. Natal Bird Club.
Crane Workshop 1978, pp. 205-21 0. Colorado State University.
952.
Sparrow. Can. J . Zool. 58: 1869-1875.

J . c . ALONSO et al . I B I S 129 224
VALVERDE, J.A. 1958. An ecological sketch of the Cot0 Doiiana. Br. Birds 51: 1-23. VERNON, C.J. 1984. The breeding periodicity of the Crowned Eagle. Proc. 11. Symp. Afr. Predatory
WILSON, E.O. 1975. Sociobiology. Harvard: Belknapp Press. Birds. Natal Bird Club.