
Why do raptors take domestic prey? The case of Bonelli's eagles and pigeons: Eagle predation on...
12
Journal of Applied Ecology 2006 43, 1075–1086 © 2006 The Authors. Journal compilation © 2006 British Ecological Society Blackwell Publishing Ltd Why do raptors take domestic prey? The case of Bonelli’s eagles and pigeons LUIS PALMA,* PEDRO BEJA,†‡ MIGUEL PAIS* and LUIS CANCELA DA FONSECA*§ *Universidade do Algarve, FCMA, Campus de Gambelas, 8005–139 Faro, Portugal; †ERENA, Ordenamento e Gestão de Recursos Naturais, Rua Robalo Gouveia, 1–1A, 1900–392 Lisboa, Portugal; ‡CIBIO, Campus Agrário de Vairão, 4485–661 Vairão, Portugal; and § IMAR/Laboratório Marítimo da Guia, Estrada do Guincho, 2750–374 Cascais, Portugal Summary 1. Predator conservation management requires detailed understanding of the ecological circumstances associated with predation, especially that on economically valuable prey. We examined the mechanisms behind Bonelli’s eagle Hieraaetus fasciatus predation on prey of domestic origin, using dietary data from 22 pairs breeding in south-west Portugal (1992–2001) together with information on landscape composition and prey availability. 2. Numerically, 42·7% (37·7% in biomass) of eagle prey comprised domestic species, about 70% of which were rural pigeons Columba livia and the remainder were racing pigeons Columba livia and domestic fowl Gallus gallus. Rabbits Oryctolagus cuniculus, red-legged partridges Alectoris rufa and jays Garrulus glandarius were the most frequent wild prey (43·1%; 50·8% in biomass). This dietary pattern was remarkably stable over a decade, but within each year the intake of pigeons almost halved over the course of the breeding season. 3. Landscape composition significantly affected the dietary proportion of wild and domestic prey items. This was particularly evident in territories dominated by eucalyptus Eucalyptus globulus plantations, where there was reduced consumption of rural pigeons and partridges, an increased intake of minor avian prey items and greater diversity in the diet overall. 4. Bonelli’s eagles showed type II functional responses while preying on the most important wild (rabbit) and domestic (rural pigeon) prey, although the former was much stronger. Eagle predation on rabbits declined with increasing abundance of pigeons, and vice versa, but there was no switching in the traditional sense, as selection between these two species was inversely frequency dependent. 5. Synthesis and applications. Predation by Bonelli’s eagle on domestic pigeons results from a combination of high vulnerability of the pigeons to eagles and a shortage of key wild prey such as rabbits and partridges, especially during the early breeding season. Given the relatively low economic value of rural pigeons and their importance in the diet of Bonelli’s eagles, they could probably be used as a conservation tool to enhance food resources in breeding territories and to deflect predation from more valuable prey such as partridges and racing pigeons. Key-words: conservation, functional response, Hieraaetus fasciatus, Mediterranean, partridge, pigeon, predation, rabbit, switching Journal of Applied Ecology (2006) 43, 1075–1086 doi: 10.1111/j.1365-2664.2006.01213.x Correspondence: Pedro Beja, ERENA, Ordenamento e Gestão de Recursos Naturais, Rua Robalo Gouveia, 1-1A, 1900– 392 Lisboa, Portugal (fax + 351217991119; e-mail [email protected]).
Transcript of Why do raptors take domestic prey? The case of Bonelli's eagles and pigeons: Eagle predation on...

Journal of Applied Ecology
2006
43
1075ndash1086
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Blackwell Publishing Ltd
Why do raptors take domestic prey The case of Bonellirsquos eagles and pigeons
LUIS PALMA PEDRO BEJAdaggerDagger MIGUEL PAIS and LUIS CANCELA DA FONSECAsect
Universidade do Algarve FCMA Campus de Gambelas 8005ndash139 Faro Portugal
dagger
ERENA Ordenamento e Gestatildeo de Recursos Naturais Rua Robalo Gouveia 1ndash1A 1900ndash392 Lisboa Portugal
Dagger
CIBIO Campus Agraacuterio de Vairatildeo 4485ndash661 Vairatildeo Portugal and
sect
IMARLaboratoacuterio Mariacutetimo da Guia Estrada do Guincho 2750ndash374 Cascais Portugal
Summary
1
Predator conservation management requires detailed understanding of the ecologicalcircumstances associated with predation especially that on economically valuable preyWe examined the mechanisms behind Bonellirsquos eagle
Hieraaetus fasciatus
predationon prey of domestic origin using dietary data from 22 pairs breeding in south-westPortugal (1992ndash2001) together with information on landscape composition and preyavailability
2
Numerically 42middot7 (37middot7 in biomass) of eagle prey comprised domestic speciesabout 70 of which were rural pigeons
Columba livia
and the remainder were racingpigeons
Columba livia
and domestic fowl
Gallus gallus
Rabbits
Oryctolagus cuniculus
red-legged partridges
Alectoris rufa
and jays
Garrulus glandarius
were the most frequentwild prey (43middot1 50middot8 in biomass) This dietary pattern was remarkably stable over adecade but within each year the intake of pigeons almost halved over the course of thebreeding season
3
Landscape composition significantly affected the dietary proportion of wild anddomestic prey items This was particularly evident in territories dominated by eucalyptus
Eucalyptus globulus
plantations where there was reduced consumption of rural pigeonsand partridges an increased intake of minor avian prey items and greater diversity in thediet overall
4
Bonellirsquos eagles showed type II functional responses while preying on the mostimportant wild (rabbit) and domestic (rural pigeon) prey although the former wasmuch stronger Eagle predation on rabbits declined with increasing abundance ofpigeons and vice versa but there was no switching in the traditional sense as selectionbetween these two species was inversely frequency dependent
5
Synthesis and applications
Predation by Bonellirsquos eagle on domestic pigeons resultsfrom a combination of high vulnerability of the pigeons to eagles and a shortage of keywild prey such as rabbits and partridges especially during the early breeding seasonGiven the relatively low economic value of rural pigeons and their importance in thediet of Bonellirsquos eagles they could probably be used as a conservation tool to enhancefood resources in breeding territories and to deflect predation from more valuable preysuch as partridges and racing pigeons
Key-words
conservation functional response
Hieraaetus fasciatus
Mediterraneanpartridge pigeon predation rabbit switching
Journal of Applied Ecology
(2006)
43
1075ndash1086doi 101111j1365-2664200601213x
Correspondence Pedro Beja ERENA Ordenamento e Gestatildeo de Recursos Naturais Rua Robalo Gouveia 1-1A 1900ndash392 Lisboa Portugal (fax + 351217991119 e-mail pbejaerenapt)
1076
L Palma
et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
Introduction
Predators feeding on poultry livestock and game havelong been killed by humans often resulting in populationdeclines and even extermination (Thirgood
et al
2000Treves amp Karanth 2003 Graham Beckerman amp Thir-good 2005) Recently many of these predators haverecovered as a consequence of increased tolerance andprotection (Treves amp Karanth 2003 Valkama
et al
2005) However population recovery involves majorchallenges when protected predators expanding intohuman-dominated landscapes come into contact withspecies that are hunted harvested or farmed for humanconsumption or recreation (Meriggi amp Lovari 1996Landa
et al
1999 Stahl
et al
2001) Such food resourcesmay actually contribute to supporting increasedpredator densities (Kenward Marcstroumlm amp Karlbom1981 Selarings 1997 Redpath amp Thirgood 1999) therebycreating dilemmas between the protection of predatorsand the mitigation of predation damage (Thirgood
et al
2000 Woodroffe
et al
2005) These managementconflicts need to be addressed if population recoveriesof protected predators are to be sustained (GrahamBeckerman amp Thirgood 2005)
Birds of prey are often perceived as damaging byhunters gamekeepers farmers and pigeon fanciers(Shawyer Clarke amp Dixon 2000 Thirgood
et al
2000Valkama
et al
2005) Conflicts involving predation oneconomically valuable game species have been thor-oughly investigated showing that persecution remainsa threat for many predatory species (Villafuerte Vintildeuelaamp Blanco 1998 Thirgood
et al
2000 Carrete
et al
2002) Much less is known of cases involving predationon domestic prey although they may also be a significantsource of conflict (Stahl Ruette amp Gros 2002 Avery ampCummings 2004) For instance domestic pigeons
Columba livia
L are often consumed by recoveringspecies such as peregrine falcons
Falco peregrinu
s Lresulting in complaints that raptor attacks are becomingunacceptably high (Shawyer Clarke amp Dixon 2000Henderson Parrott amp Moore 2004)
High predation rates on domestic prey can usuallybe explained by three main predatory mechanisms eachof which has different implications for the mitigation ofpredator damages An alternative prey hypothesis(APH
sensu
Angelstam Lindstroumlm amp Wideacuten 1984)is assumed often implicitly when predators switch todomestic prey as wild prey becomes scarce (Meriggi ampLovari 1996) In these circumstances the restocking ofwild prey may alleviate predation on domestic speciesThis solution may be ineffective if predators take domes-tic prey primarily because of their high vulnerabilityirrespective of wild prey abundance The vulnerableprey hypothesis (VPH) calls for management strategiesreducing the exposure of domestic prey using forinstance deterrents or fencing (Shawyer Clarke ampDixon 2000 Kenward
et al
2001 Treves amp Karanth2003) Finally the problem animal hypothesis (PAH) isproposed when predation involves a behavioural feeding
specialization by a few individuals in which case theremoval of problematic animals is often advocated(Stahl
et al
2001 Treves amp Karanth 2003)Testing these hypotheses requires information on the
responses of predators to variation in the densities andrelative frequencies of wild and domestic prey Underthe APH predation on domestic prey may be low untilthe abundance of wild prey declines below a minimumthreshold Switching (
sensu
Murdoch 1969) shouldthen occur implying a frequency-dependent responseto fluctuating prey densities with the predator eatingdisproportionately more of the most abundant preyitem In this case predators are usually expected toshow a Holling type III (sigmoid) functional responseto fluctuations in prey density (Akre amp Johnson 1979)In contrast under the VPH the functional response tofluctuations in domestic prey density would show atype II (convex) curve characterized by a rapid increasein the number of prey taken per predator at low preydensities indicating the presence of highly vulnerableor rewarding prey The PAH requires that predationon domestic prey occurs only locally and largelyindependently of prey abundance In practice thesepredicted responses to fluctuations in prey abundanceare likely to be more complex as predators often feedon several different wild and domestic prey types andtheir abundance and profitability may change acrosshabitats and over time Assessing the mechanismsinvolved in a particular instance of predation on adomestic prey thus requires a good understandingof the predatorndashprey interactions and the ecologicalcontext in which they take place (Graham Beckermanamp Thirgood 2005)
Factors influencing predation on domestic prey wereexamined by analysing Bonellirsquos eagle
Hieraaetusfasciatus
Vieillot predation patterns in south-westPortugal (1992ndash2001) Bonellirsquos eagle is an endangeredspecies showing a marked decline in MediterraneanEurope since the early 1980s (Real amp Mantildeosa 1997)Bonellirsquos eagles feed on a range of vertebrates withrabbits
Oryctolagus cuniculus
L red-legged partridges
Alectoris rufa
L and pigeons
Columba
spp often beingthe main prey (Real 1991 Valkama
et al
2005) Asa consequence persecution by hunters and pigeonfanciers represents a major threat for some Bonellirsquoseagle populations (Real
et al
2001 Carrete
et al
2002)In the study area rabbits and partridges are scarcewhile rural pigeons are relatively abundant in smallvillages and isolated farmhouses Preliminary evidenceshowed that eagles consume pigeons very frequently(Palma
et al
2005) and there was the possibility thatthe pigeons acted as an alternative resource to wildprey This creates a management dilemma (Petty Lurzamp Rushton 2003) as pigeons provide an important foodresource while also representing a potential cause ofconflict with pigeon fanciers A closer examination ofthe factors affecting predation on domestic pigeons usedinformation on variation across breeding territories indiet composition land uses and the abundance of wild
1077
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
and domestic prey The aim was to assess (i) the extentof predation on domestic prey (ii) variation in dietacross years and over the breeding season (iii) theinfluence of habitat on predation patterns (iv) the func-tional responses to variation in wild and domestic preyabundance and (v) the eventual occurrence of switchingbetween wild and domestic prey This information wasthen used to infer the predatory mechanisms involved inthis predatorndashprey system and to suggest managementsolutions that might contribute to the conservation ofthese eagles while mitigating potential conflict withhumans in south-west Portugal and elsewhere
Methods
This study focused on a dense breeding nucleus of 25Bonellirsquos eagle pairs occupying about 3000 km
2
in theuplands of south-west Portugal (Fig 1) This is an areaof low mountains and rolling hills mostly under 650 masl in altitude The climate is Mediterranean with a dryand hot summer season with little precipitation Theland is predominantly covered by open to dense corkoak
Quercus suber
L woodland and extensive scrublandoften dominated by the gum cistus
Cistus ladanifer
LIn the western half of the area eucalyptus
Eucalyptusglobulus
Labill plantations for paper pulp productioncover up to 50 of the land Human settlement iscurrently low and local most of the former scattereddwellings are abandoned and agriculture is residual
The diets of 22 pairs of eagles surveyed in 1992ndash2001were described from prey identified in remains(
n
= 747) and regurgitated pellets (
n
= 1039) collectedduring the breeding season (gt 85 in MarchndashMay) atactive nests and surrounding perches together withuncollected fresh prey observed at nests (
n
= 199)Information was obtained for 9middot6
plusmn
4middot8 (2ndash17) pairs peryear with variation as a result of population growthbreeding failures unknown nest locations and reducedfield effort in 1992ndash94 and 2001 Nests were checked5middot5
plusmn
2middot2 (1ndash11) times year
minus
1
The species age and genderof each individual prey were estimated wheneverpossible using published identification keys and areference collection Metallic rings provided a minimumestimate of racing pigeons in the diet as rural pigeonswere seldom ringed within the study area
To reduce biases normally associated with dietarystudies (Real 1996 Marchesi Pedrini amp Sergio 2002)and to increase sample sizes data from pellets remainsand direct observations were combined (Ontiverosamp Pleguezuelos 2000 Penteriani Gallardo amp Roche2002 Ontiveros Pleguezuelos amp Caro 2005) For eachpair and sampling occasion the number of individualsof a given prey species represented in the sample wastaken only from the data source yielding the largestestimate thereby avoiding duplications among datasources In the case of two siblings the estimatednumber of prey items represented in pellets was halvedto avoid duplication of the same prey in two pellets(Real 1996) The biomass of each prey species wasestimated by multiplying the number of individuals byits estimated mean weight using data from the studyarea bibliographic sources (Real 1987 Cardona ampEsteban 2002) and information from racing pigeonfanciers (D Santos personal communication) Thenumber of individuals of each prey species taken byan average eagle pair during the breeding season wasestimated following Petty Lurz amp Rushton (2003)considering one nestling per pair an overall dailyconsumption of 750 g food per pair plus one nestling(Real 1987) a mean incubation period of 39 days(Arroyo Ferreiro amp Garza 1995) and a mean nestlingperiod of 63 days (Real amp Mantildeosa 1998)
Variation in diet landscape composition and preyavailability among territories was used to estimate thefactors influencing eagle predation patterns Given thelack of detailed information on the actual breedingterritories habitat and prey data were quantified withinapproximate territory boundaries estimated usingDirichlet tessellation (Doncaster amp Woodroffe 1993)For each pair the mean geographical location ofnests was the Dirichlet centre and boundaries wereconstrained to be at a maximum of 8 km from the centre(Fig 1) These criteria were based on home range
Fig 1 Location in south-west Portugal of alternative nests of Bonellirsquos eagle pairs(1992ndash2001) and the approximate territory boundaries estimated using Dirichlettessellation
1078
L Palma
et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data from other studies (Mure 2003) and on nearly 800haphazard observations at gt 500 m from active neststhrough the 10-year study period suggesting that eachBonellirsquos eagle pair maintained an exclusive foragingterritory which only rarely extended beyond 8 kmfrom the active nest (L Palma unpublished data)Although this was a relatively crude approach eachrange boundary encompassed gt 90 of observationsof individually recognized elements of the respectivebreeding pair
Landscape composition was quantified using digitalland cover cartography from 1995 (wwwdgrfmin-agriculturaptifnmapashtm accessed 1 April 2005) con-sidering six categories (i) native Mediterranean forestsdominated by cork oaks and strawberry trees
Arbutusunedo
L (ii) eucalyptus plantations (iii) pine (mostly
Pinus pinaster
Ait) plantations (iv) scrubland (v)farmland and (vi) urban areas (Table 1) Landscapediversity was computed using the Shannon index
Rabbit abundance was estimated in SeptemberndashOctober 1997 from latrine counts in 296 250-m linetransects walked along dirt roads and firebreaks crossingeagle territories and neighbouring areas (Palma Bejaamp Rodrigues 1999) Latrine counts were used (Iborra ampLumaret 1997 Beja Palma amp Pais 2007) as directobservation of rabbits was not feasible because of densevegetation cover One transect was walked per 1-km
2
grid square with five random squares surveyed in eachof 65 5-km
2
grid squares distributed in a checkerboardpattern To increase comparability among sites eachtransect was located in the most favourable shrublandhabitats available within each square (Beja Palma ampPais 2007) The mean latrine count per breedingterritory was used to index relative rabbit abundanceOnly 14 territories for which there were at least 10line transects (15middot2
plusmn
4middot2 10ndash23) were used in furtheranalyses because information could not be obtainedfor the entire study area A comprehensive ruralpigeon survey was carried out from September 1994 toDecember 1995 encompassing the approximate breedingterritories of 18 eagle pairs known at the time Pigeonlofts were mapped by visiting all small villages andisolated farmhouses where the number of pigeonswas estimated for each loft either through enquiriesor direct counts Point estimates of pigeon abundancewere interpolated into a 250-m raster grid using kerneldensity analysis with a 5-km searching radius and
these grid values were then used to estimate averagerural pigeon densities within each breeding territoryThe abundance of racing pigeons crossing the studyarea was estimated from information provided byhoming pigeon fanciers on the periods and frequencyof races and training flights and the approximate num-bers of pigeons involved their origin and geographicaldistribution (D Santos personal communication)
The prey data only referred to a restricted periodwhereas the dietary information spanned 10 yearsThis was recognized as a shortcoming in the data butwas unavoidable It was not possible to repeat the surveysand a long diet study would be needed to characterizea large number of pairs This shortcoming was unlikely toaffect the results of this study seriously as unsystematicobservations suggested that regional trends in preyabundance remained essentially the same In particularthere were no marked changes in land cover and humanactivities that might have changed the distribution ofrabbit and pigeon abundances extensively Also norabbit crashes as a result of viral haemorrhagic diseasesimilar to that reported elsewhere (Fa Sharples amp Bell1999) or severe myxomatosis outbreaks were observedMoreover surveys were carried out approximatelyhalf-way through the study period when most dietaryinformation was collected
Preliminary to data analysis the angular transforma-tion (arcsine
radic
p
) was used to reduce the influence ofextreme values and to overcome problems associatedwith the unity sum constraint of proportional data(Zar 1996) Multicollinearity among habitat variableswas investigated using principal component analysis(PCA) thereby describing the main gradients in land-usevariability (Legendre amp Legendre 1998) A varimaxrotation was performed on the principal componentswith eigenvalues gt 1 to enhance their interpretability(Legendre amp Legendre 1998) Seasonal and annualtrends in dietary proportions were examined usinglinear regression including quadratic terms to checkfor non-linear trends (Legendre amp Legendre 1998)Relationships between spatial variation in prey con-sumption and land-use variables were examinedlikewise In temporal analyses data from all eagle pairswere pooled per time period whereas in spatial analyses
Table 1 Average (plusmn SD) landscape composition and summary results of a principal component analysis with varimax rotationdescribing the dominant land-use gradients across Bonellirsquos eagle territories in south-west Portugal (1992ndash2001) The proportionof explained variation and the scores of each variable are provided for the first two rotated axis (PC)
Land cover types Mean () plusmn SD Minimumndashmaximum PC1 (38middot1) PC2 (30middot1)
Scrubland 40middot1 14middot2 16middot8ndash62middot5 0middot84Mediterranean forests 26middot3 12middot6 6middot5ndash46middot4 minus0middot82Farmland 16middot7 15middot7 2middot7ndash55middot6 minus0middot92Eucalyptus plantations 14middot4 12middot2 0middot0ndash43middot8 0middot79Pine plantations 2middot1 2middot7 0middot0ndash7middot5 0middot63Urban 0middot4 0middot4 0middot0ndash1middot4 minus0middot78
1079
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data were pooled per breeding pair Only cases for whichthere were at least 25 prey items were used becausedietary proportions stabilize around this threshold(Ontiveros Pleguezuelos amp Caro 2005)
Functional responses were investigated by modellingthe relationships between the average number of a preytype eaten per breeding pair and the abundance of eitherpigeons or rabbits In each case functions representinglinear (type I) convex (type II) and sigmoidal (type III)curves were fitted to the data (Patterson Benjamin ampMessier 1998) and the best model was selected based onsecond-order Aikaike information criterion (AICc)because of small sample sizes (Burnham amp Anderson1998) Type II and III responses were modelled usingthe generalized MichaelisndashMenton function (Real 1977)The effects of pigeons and rabbits on the consumptionof other prey were investigated using linear regressionand partial correlation analysis (Zar 1996)
Prey switching was examined by relating the ratio ofpigeons to rabbits eaten (
e
1
e
2
) to their availability ratio(
A
1
A
2
) using the equation
e
1
e
2
= (
V
times
A
1
A
2
)
b
(Green-wood amp Elton 1979) The parameter
V
is a measure offrequency-independent selection whereas
b
is a measureof frequency dependence
b
gt 1 provides evidence forswitching with the predator eating disproportionatelymore of the most abundant prey
b
lt 1 reflects dispro-portionate predation on the less common prey and
b
= 1 indicates frequency-independent selection As
A
1
and
A
2
were expressed in distinct units and there wasno absolute estimate of rabbit densities this analysisshould be regarded as approximate and the parameter
V
should not be interpreted Nevertheless the shapeof the curve relating
e
1
e
2
to
A
1
A
2
is meaningfulproviding the best approach to evaluating the presenceof switching between the two prey species
Results
Bonellirsquos eagle territories were dominated by scrublandand oak forests together accounting for almost 70of landscape composition (Table 1) Eucalyptus plan-tations and farmland were also important whereasland cover by pine plantations and urban areas wasnegligible The PCA with varimax rotation revealeda dominant gradient largely reflecting agriculturalabandonment that accounted for 38middot1 of land covervariability ranging from territories with a significantproportion of farmland and urban areas to thosedominated by scrubland The second rotated PCAaxis (30middot8) represented the dominant forest types con-trasting territories dominated by Mediterranean oakforests with those that were predominantly eucalyptusplantations
There was strong variation among territories in theabundance of rabbits and pigeons (Fig 2) Rabbitsoccurred in only 27 of the 296 transects surveyed andthey tended to be fairly scarce where they occurred (4middot2latrines 250 m
minus
1
(4middot3 1ndash24
n
= 80) This resulted in a50-fold variation in rabbit abundance among eagleterritories with densities up to 1ndash5 latrines 250 m
minus
1
along the southern and western edges whereas rabbitswere scarce elsewhere (lt 0middot5 latrines 250 m
minus
1
) A totalof 436 pigeon lofts was identified across the regionwith on average 25middot5
plusmn
37middot7 (2ndash550) pigeons per loftPigeons were more evenly distributed than rabbitsalthough there was about a 10-fold variation amongterritories from about 0middot47 to 4middot71 pigeons km
minus
2
(2middot4
plusmn
1middot2 pigeons km
minus
2
) There was no correlation betweenrabbit and pigeon abundance (r = minus0middot37 P = 0middot262)
Fig 2 Interpolated maps of spatial variation in prey abundances across Bonellirsquos eagles territories in south-west Portugal Valuesare SD from the mean to allow comparisons between maps Dots are rabbit sampling sites and pigeon lofts Dotted lines definethe interpolation limits Hatched lines indicate territories with no information on prey abundance
1080L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
across the 11 territories for which there was informationon both prey
Dietary analysis yielded a minimum estimate of 1497individuals of 32 prey species (see Table S1 in thesupplementary material) Numerically prey of domesticorigin comprised 42middot7 of the diet (37middot7 in biomass)consumed in every year and by all eagle pairs (Table 2)About 70 of these prey items were rural pigeons butracing pigeons and domestic fowl were also consumedfrequently Considering the estimates of prey intake(Table 2) prey densities and territory sizes the averageproportion of the rural pigeons available per eaglepair that were taken through the breeding seasonwas 17middot8 plusmn 12middot6 (5middot7ndash57middot8) Rabbits red-leggedpartridges and jays Garrulus glandarius L were themost frequent wild prey numerically representing43middot1 (50middot8 biomass) of the eagle diet (Table 2)
Of the 114 racing pigeons aged through metallicrings 31middot6 were 1 year old or under 51middot8 2ndash3 yearsold and 16middot7 more than 4 years old The remains offreshly killed domestic fowl observed at nests wereall from juvenile birds (n = 12) whereas 86middot2 of therabbits were adults (n = 29) A majority of partridgeskilled by eagles were males (65middot7 n = 35)
Variation in diet between years was analysed for 1993ndash2000 ie when annual samples were sufficiently large(prey per year 185middot4 plusmn 93middot1 65ndash352) The relativeimportance of the most frequently consumed items re-mained largely the same over the study period (Kendallrsquoscoefficient of concordance W = 0middot86 P lt 0middot001) withno evidence for consistent trends in the consumption ofany prey species (Pearson correlations minus0middot41 lt r lt 0middot57P gt 0middot14 Fig 3a) The numeric proportion of ruralpigeons was fairly stable (23middot1ndash33middot8) as was that ofrabbits (15middot4ndash23middot1) and racing pigeons (6middot6ndash12middot6)The largest variation was recorded for relatively lessimportant prey such as jays (2middot0ndash10middot8) and domesticfowl (1middot2ndash7middot1)
Prey were assigned to 2-week periods from 1 Marchto 15 June (prey per 2 weeks 210middot8 plusmn 177middot6 44ndash504)to investigate variations during the breeding season(Fig 3b) Although there was concordance in diet com-position over time (Kendallrsquos W = 0middot73 P = 0middot001)most prey species showed distinctive temporal trendsThere were marked declines in the dietary importanceof rural (r = minus0middot91 P lt 0middot01) and racing pigeons (r =minus0middot96 P lt 0middot001) which halved their overall numericcontribution from a maximum of about 50 in Marchto a minimum of 23middot7 in June This was largelycompensated for by increases in jays (r = 0middot91 P lt 0middot05)and a wide group of birds represented in lsquootherrsquo prey(r = 0middot92 P lt 0middot01) from a minimum in the second half
Table 2 Diet of Bonellirsquos eagles and prey consumption per pair during the breedingseason in south-west Portugal (1992ndash2001)
Prey categories n Numbers
Biomass
Consumption per pair mean plusmn SD (minimumndashmaximum)
Domestic birdsRural pigeon 451 30middot1 26middot0 48middot0 plusmn 14middot6 (17ndash77)Racing pigeon 136 9middot1 7middot2 14middot7 plusmn 7middot7 (0ndash37)Domestic fowl 52 3middot5 4middot5 5middot4 plusmn 8middot6 (0ndash37)
Wild birdsRed-legged partridge 258 17middot2 14middot9 28middot7 plusmn 11middot9 (6ndash59)Jay 112 7middot5 2middot7 12middot0 plusmn 9middot2 (0ndash28)Other birds 198 13middot4 11middot4 24middot2 plusmn 18middot3 (2ndash68)
MammalsRabbit 276 18middot4 33middot2 30middot4 plusmn 11middot8 (16ndash58)Hare 3 0middot2 lt 0middot1 0middot2 plusmn 0middot6 (0ndash2)ReptilesOcellated lizard 11 0middot7 lt 0middot1 1middot1 plusmn 1middot9 (0ndash6)
Fig 3 Variation of Bonellirsquos eagle diet in south-west Portugal (a) over the years 1993ndash2000 and (b) across the breeding season 1 Marchminus15 June
1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1076
L Palma
et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
Introduction
Predators feeding on poultry livestock and game havelong been killed by humans often resulting in populationdeclines and even extermination (Thirgood
et al
2000Treves amp Karanth 2003 Graham Beckerman amp Thir-good 2005) Recently many of these predators haverecovered as a consequence of increased tolerance andprotection (Treves amp Karanth 2003 Valkama
et al
2005) However population recovery involves majorchallenges when protected predators expanding intohuman-dominated landscapes come into contact withspecies that are hunted harvested or farmed for humanconsumption or recreation (Meriggi amp Lovari 1996Landa
et al
1999 Stahl
et al
2001) Such food resourcesmay actually contribute to supporting increasedpredator densities (Kenward Marcstroumlm amp Karlbom1981 Selarings 1997 Redpath amp Thirgood 1999) therebycreating dilemmas between the protection of predatorsand the mitigation of predation damage (Thirgood
et al
2000 Woodroffe
et al
2005) These managementconflicts need to be addressed if population recoveriesof protected predators are to be sustained (GrahamBeckerman amp Thirgood 2005)
Birds of prey are often perceived as damaging byhunters gamekeepers farmers and pigeon fanciers(Shawyer Clarke amp Dixon 2000 Thirgood
et al
2000Valkama
et al
2005) Conflicts involving predation oneconomically valuable game species have been thor-oughly investigated showing that persecution remainsa threat for many predatory species (Villafuerte Vintildeuelaamp Blanco 1998 Thirgood
et al
2000 Carrete
et al
2002) Much less is known of cases involving predationon domestic prey although they may also be a significantsource of conflict (Stahl Ruette amp Gros 2002 Avery ampCummings 2004) For instance domestic pigeons
Columba livia
L are often consumed by recoveringspecies such as peregrine falcons
Falco peregrinu
s Lresulting in complaints that raptor attacks are becomingunacceptably high (Shawyer Clarke amp Dixon 2000Henderson Parrott amp Moore 2004)
High predation rates on domestic prey can usuallybe explained by three main predatory mechanisms eachof which has different implications for the mitigation ofpredator damages An alternative prey hypothesis(APH
sensu
Angelstam Lindstroumlm amp Wideacuten 1984)is assumed often implicitly when predators switch todomestic prey as wild prey becomes scarce (Meriggi ampLovari 1996) In these circumstances the restocking ofwild prey may alleviate predation on domestic speciesThis solution may be ineffective if predators take domes-tic prey primarily because of their high vulnerabilityirrespective of wild prey abundance The vulnerableprey hypothesis (VPH) calls for management strategiesreducing the exposure of domestic prey using forinstance deterrents or fencing (Shawyer Clarke ampDixon 2000 Kenward
et al
2001 Treves amp Karanth2003) Finally the problem animal hypothesis (PAH) isproposed when predation involves a behavioural feeding
specialization by a few individuals in which case theremoval of problematic animals is often advocated(Stahl
et al
2001 Treves amp Karanth 2003)Testing these hypotheses requires information on the
responses of predators to variation in the densities andrelative frequencies of wild and domestic prey Underthe APH predation on domestic prey may be low untilthe abundance of wild prey declines below a minimumthreshold Switching (
sensu
Murdoch 1969) shouldthen occur implying a frequency-dependent responseto fluctuating prey densities with the predator eatingdisproportionately more of the most abundant preyitem In this case predators are usually expected toshow a Holling type III (sigmoid) functional responseto fluctuations in prey density (Akre amp Johnson 1979)In contrast under the VPH the functional response tofluctuations in domestic prey density would show atype II (convex) curve characterized by a rapid increasein the number of prey taken per predator at low preydensities indicating the presence of highly vulnerableor rewarding prey The PAH requires that predationon domestic prey occurs only locally and largelyindependently of prey abundance In practice thesepredicted responses to fluctuations in prey abundanceare likely to be more complex as predators often feedon several different wild and domestic prey types andtheir abundance and profitability may change acrosshabitats and over time Assessing the mechanismsinvolved in a particular instance of predation on adomestic prey thus requires a good understandingof the predatorndashprey interactions and the ecologicalcontext in which they take place (Graham Beckermanamp Thirgood 2005)
Factors influencing predation on domestic prey wereexamined by analysing Bonellirsquos eagle
Hieraaetusfasciatus
Vieillot predation patterns in south-westPortugal (1992ndash2001) Bonellirsquos eagle is an endangeredspecies showing a marked decline in MediterraneanEurope since the early 1980s (Real amp Mantildeosa 1997)Bonellirsquos eagles feed on a range of vertebrates withrabbits
Oryctolagus cuniculus
L red-legged partridges
Alectoris rufa
L and pigeons
Columba
spp often beingthe main prey (Real 1991 Valkama
et al
2005) Asa consequence persecution by hunters and pigeonfanciers represents a major threat for some Bonellirsquoseagle populations (Real
et al
2001 Carrete
et al
2002)In the study area rabbits and partridges are scarcewhile rural pigeons are relatively abundant in smallvillages and isolated farmhouses Preliminary evidenceshowed that eagles consume pigeons very frequently(Palma
et al
2005) and there was the possibility thatthe pigeons acted as an alternative resource to wildprey This creates a management dilemma (Petty Lurzamp Rushton 2003) as pigeons provide an important foodresource while also representing a potential cause ofconflict with pigeon fanciers A closer examination ofthe factors affecting predation on domestic pigeons usedinformation on variation across breeding territories indiet composition land uses and the abundance of wild
1077
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
and domestic prey The aim was to assess (i) the extentof predation on domestic prey (ii) variation in dietacross years and over the breeding season (iii) theinfluence of habitat on predation patterns (iv) the func-tional responses to variation in wild and domestic preyabundance and (v) the eventual occurrence of switchingbetween wild and domestic prey This information wasthen used to infer the predatory mechanisms involved inthis predatorndashprey system and to suggest managementsolutions that might contribute to the conservation ofthese eagles while mitigating potential conflict withhumans in south-west Portugal and elsewhere
Methods
This study focused on a dense breeding nucleus of 25Bonellirsquos eagle pairs occupying about 3000 km
2
in theuplands of south-west Portugal (Fig 1) This is an areaof low mountains and rolling hills mostly under 650 masl in altitude The climate is Mediterranean with a dryand hot summer season with little precipitation Theland is predominantly covered by open to dense corkoak
Quercus suber
L woodland and extensive scrublandoften dominated by the gum cistus
Cistus ladanifer
LIn the western half of the area eucalyptus
Eucalyptusglobulus
Labill plantations for paper pulp productioncover up to 50 of the land Human settlement iscurrently low and local most of the former scattereddwellings are abandoned and agriculture is residual
The diets of 22 pairs of eagles surveyed in 1992ndash2001were described from prey identified in remains(
n
= 747) and regurgitated pellets (
n
= 1039) collectedduring the breeding season (gt 85 in MarchndashMay) atactive nests and surrounding perches together withuncollected fresh prey observed at nests (
n
= 199)Information was obtained for 9middot6
plusmn
4middot8 (2ndash17) pairs peryear with variation as a result of population growthbreeding failures unknown nest locations and reducedfield effort in 1992ndash94 and 2001 Nests were checked5middot5
plusmn
2middot2 (1ndash11) times year
minus
1
The species age and genderof each individual prey were estimated wheneverpossible using published identification keys and areference collection Metallic rings provided a minimumestimate of racing pigeons in the diet as rural pigeonswere seldom ringed within the study area
To reduce biases normally associated with dietarystudies (Real 1996 Marchesi Pedrini amp Sergio 2002)and to increase sample sizes data from pellets remainsand direct observations were combined (Ontiverosamp Pleguezuelos 2000 Penteriani Gallardo amp Roche2002 Ontiveros Pleguezuelos amp Caro 2005) For eachpair and sampling occasion the number of individualsof a given prey species represented in the sample wastaken only from the data source yielding the largestestimate thereby avoiding duplications among datasources In the case of two siblings the estimatednumber of prey items represented in pellets was halvedto avoid duplication of the same prey in two pellets(Real 1996) The biomass of each prey species wasestimated by multiplying the number of individuals byits estimated mean weight using data from the studyarea bibliographic sources (Real 1987 Cardona ampEsteban 2002) and information from racing pigeonfanciers (D Santos personal communication) Thenumber of individuals of each prey species taken byan average eagle pair during the breeding season wasestimated following Petty Lurz amp Rushton (2003)considering one nestling per pair an overall dailyconsumption of 750 g food per pair plus one nestling(Real 1987) a mean incubation period of 39 days(Arroyo Ferreiro amp Garza 1995) and a mean nestlingperiod of 63 days (Real amp Mantildeosa 1998)
Variation in diet landscape composition and preyavailability among territories was used to estimate thefactors influencing eagle predation patterns Given thelack of detailed information on the actual breedingterritories habitat and prey data were quantified withinapproximate territory boundaries estimated usingDirichlet tessellation (Doncaster amp Woodroffe 1993)For each pair the mean geographical location ofnests was the Dirichlet centre and boundaries wereconstrained to be at a maximum of 8 km from the centre(Fig 1) These criteria were based on home range
Fig 1 Location in south-west Portugal of alternative nests of Bonellirsquos eagle pairs(1992ndash2001) and the approximate territory boundaries estimated using Dirichlettessellation
1078
L Palma
et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data from other studies (Mure 2003) and on nearly 800haphazard observations at gt 500 m from active neststhrough the 10-year study period suggesting that eachBonellirsquos eagle pair maintained an exclusive foragingterritory which only rarely extended beyond 8 kmfrom the active nest (L Palma unpublished data)Although this was a relatively crude approach eachrange boundary encompassed gt 90 of observationsof individually recognized elements of the respectivebreeding pair
Landscape composition was quantified using digitalland cover cartography from 1995 (wwwdgrfmin-agriculturaptifnmapashtm accessed 1 April 2005) con-sidering six categories (i) native Mediterranean forestsdominated by cork oaks and strawberry trees
Arbutusunedo
L (ii) eucalyptus plantations (iii) pine (mostly
Pinus pinaster
Ait) plantations (iv) scrubland (v)farmland and (vi) urban areas (Table 1) Landscapediversity was computed using the Shannon index
Rabbit abundance was estimated in SeptemberndashOctober 1997 from latrine counts in 296 250-m linetransects walked along dirt roads and firebreaks crossingeagle territories and neighbouring areas (Palma Bejaamp Rodrigues 1999) Latrine counts were used (Iborra ampLumaret 1997 Beja Palma amp Pais 2007) as directobservation of rabbits was not feasible because of densevegetation cover One transect was walked per 1-km
2
grid square with five random squares surveyed in eachof 65 5-km
2
grid squares distributed in a checkerboardpattern To increase comparability among sites eachtransect was located in the most favourable shrublandhabitats available within each square (Beja Palma ampPais 2007) The mean latrine count per breedingterritory was used to index relative rabbit abundanceOnly 14 territories for which there were at least 10line transects (15middot2
plusmn
4middot2 10ndash23) were used in furtheranalyses because information could not be obtainedfor the entire study area A comprehensive ruralpigeon survey was carried out from September 1994 toDecember 1995 encompassing the approximate breedingterritories of 18 eagle pairs known at the time Pigeonlofts were mapped by visiting all small villages andisolated farmhouses where the number of pigeonswas estimated for each loft either through enquiriesor direct counts Point estimates of pigeon abundancewere interpolated into a 250-m raster grid using kerneldensity analysis with a 5-km searching radius and
these grid values were then used to estimate averagerural pigeon densities within each breeding territoryThe abundance of racing pigeons crossing the studyarea was estimated from information provided byhoming pigeon fanciers on the periods and frequencyof races and training flights and the approximate num-bers of pigeons involved their origin and geographicaldistribution (D Santos personal communication)
The prey data only referred to a restricted periodwhereas the dietary information spanned 10 yearsThis was recognized as a shortcoming in the data butwas unavoidable It was not possible to repeat the surveysand a long diet study would be needed to characterizea large number of pairs This shortcoming was unlikely toaffect the results of this study seriously as unsystematicobservations suggested that regional trends in preyabundance remained essentially the same In particularthere were no marked changes in land cover and humanactivities that might have changed the distribution ofrabbit and pigeon abundances extensively Also norabbit crashes as a result of viral haemorrhagic diseasesimilar to that reported elsewhere (Fa Sharples amp Bell1999) or severe myxomatosis outbreaks were observedMoreover surveys were carried out approximatelyhalf-way through the study period when most dietaryinformation was collected
Preliminary to data analysis the angular transforma-tion (arcsine
radic
p
) was used to reduce the influence ofextreme values and to overcome problems associatedwith the unity sum constraint of proportional data(Zar 1996) Multicollinearity among habitat variableswas investigated using principal component analysis(PCA) thereby describing the main gradients in land-usevariability (Legendre amp Legendre 1998) A varimaxrotation was performed on the principal componentswith eigenvalues gt 1 to enhance their interpretability(Legendre amp Legendre 1998) Seasonal and annualtrends in dietary proportions were examined usinglinear regression including quadratic terms to checkfor non-linear trends (Legendre amp Legendre 1998)Relationships between spatial variation in prey con-sumption and land-use variables were examinedlikewise In temporal analyses data from all eagle pairswere pooled per time period whereas in spatial analyses
Table 1 Average (plusmn SD) landscape composition and summary results of a principal component analysis with varimax rotationdescribing the dominant land-use gradients across Bonellirsquos eagle territories in south-west Portugal (1992ndash2001) The proportionof explained variation and the scores of each variable are provided for the first two rotated axis (PC)
Land cover types Mean () plusmn SD Minimumndashmaximum PC1 (38middot1) PC2 (30middot1)
Scrubland 40middot1 14middot2 16middot8ndash62middot5 0middot84Mediterranean forests 26middot3 12middot6 6middot5ndash46middot4 minus0middot82Farmland 16middot7 15middot7 2middot7ndash55middot6 minus0middot92Eucalyptus plantations 14middot4 12middot2 0middot0ndash43middot8 0middot79Pine plantations 2middot1 2middot7 0middot0ndash7middot5 0middot63Urban 0middot4 0middot4 0middot0ndash1middot4 minus0middot78
1079
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data were pooled per breeding pair Only cases for whichthere were at least 25 prey items were used becausedietary proportions stabilize around this threshold(Ontiveros Pleguezuelos amp Caro 2005)
Functional responses were investigated by modellingthe relationships between the average number of a preytype eaten per breeding pair and the abundance of eitherpigeons or rabbits In each case functions representinglinear (type I) convex (type II) and sigmoidal (type III)curves were fitted to the data (Patterson Benjamin ampMessier 1998) and the best model was selected based onsecond-order Aikaike information criterion (AICc)because of small sample sizes (Burnham amp Anderson1998) Type II and III responses were modelled usingthe generalized MichaelisndashMenton function (Real 1977)The effects of pigeons and rabbits on the consumptionof other prey were investigated using linear regressionand partial correlation analysis (Zar 1996)
Prey switching was examined by relating the ratio ofpigeons to rabbits eaten (
e
1
e
2
) to their availability ratio(
A
1
A
2
) using the equation
e
1
e
2
= (
V
times
A
1
A
2
)
b
(Green-wood amp Elton 1979) The parameter
V
is a measure offrequency-independent selection whereas
b
is a measureof frequency dependence
b
gt 1 provides evidence forswitching with the predator eating disproportionatelymore of the most abundant prey
b
lt 1 reflects dispro-portionate predation on the less common prey and
b
= 1 indicates frequency-independent selection As
A
1
and
A
2
were expressed in distinct units and there wasno absolute estimate of rabbit densities this analysisshould be regarded as approximate and the parameter
V
should not be interpreted Nevertheless the shapeof the curve relating
e
1
e
2
to
A
1
A
2
is meaningfulproviding the best approach to evaluating the presenceof switching between the two prey species
Results
Bonellirsquos eagle territories were dominated by scrublandand oak forests together accounting for almost 70of landscape composition (Table 1) Eucalyptus plan-tations and farmland were also important whereasland cover by pine plantations and urban areas wasnegligible The PCA with varimax rotation revealeda dominant gradient largely reflecting agriculturalabandonment that accounted for 38middot1 of land covervariability ranging from territories with a significantproportion of farmland and urban areas to thosedominated by scrubland The second rotated PCAaxis (30middot8) represented the dominant forest types con-trasting territories dominated by Mediterranean oakforests with those that were predominantly eucalyptusplantations
There was strong variation among territories in theabundance of rabbits and pigeons (Fig 2) Rabbitsoccurred in only 27 of the 296 transects surveyed andthey tended to be fairly scarce where they occurred (4middot2latrines 250 m
minus
1
(4middot3 1ndash24
n
= 80) This resulted in a50-fold variation in rabbit abundance among eagleterritories with densities up to 1ndash5 latrines 250 m
minus
1
along the southern and western edges whereas rabbitswere scarce elsewhere (lt 0middot5 latrines 250 m
minus
1
) A totalof 436 pigeon lofts was identified across the regionwith on average 25middot5
plusmn
37middot7 (2ndash550) pigeons per loftPigeons were more evenly distributed than rabbitsalthough there was about a 10-fold variation amongterritories from about 0middot47 to 4middot71 pigeons km
minus
2
(2middot4
plusmn
1middot2 pigeons km
minus
2
) There was no correlation betweenrabbit and pigeon abundance (r = minus0middot37 P = 0middot262)
Fig 2 Interpolated maps of spatial variation in prey abundances across Bonellirsquos eagles territories in south-west Portugal Valuesare SD from the mean to allow comparisons between maps Dots are rabbit sampling sites and pigeon lofts Dotted lines definethe interpolation limits Hatched lines indicate territories with no information on prey abundance
1080L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
across the 11 territories for which there was informationon both prey
Dietary analysis yielded a minimum estimate of 1497individuals of 32 prey species (see Table S1 in thesupplementary material) Numerically prey of domesticorigin comprised 42middot7 of the diet (37middot7 in biomass)consumed in every year and by all eagle pairs (Table 2)About 70 of these prey items were rural pigeons butracing pigeons and domestic fowl were also consumedfrequently Considering the estimates of prey intake(Table 2) prey densities and territory sizes the averageproportion of the rural pigeons available per eaglepair that were taken through the breeding seasonwas 17middot8 plusmn 12middot6 (5middot7ndash57middot8) Rabbits red-leggedpartridges and jays Garrulus glandarius L were themost frequent wild prey numerically representing43middot1 (50middot8 biomass) of the eagle diet (Table 2)
Of the 114 racing pigeons aged through metallicrings 31middot6 were 1 year old or under 51middot8 2ndash3 yearsold and 16middot7 more than 4 years old The remains offreshly killed domestic fowl observed at nests wereall from juvenile birds (n = 12) whereas 86middot2 of therabbits were adults (n = 29) A majority of partridgeskilled by eagles were males (65middot7 n = 35)
Variation in diet between years was analysed for 1993ndash2000 ie when annual samples were sufficiently large(prey per year 185middot4 plusmn 93middot1 65ndash352) The relativeimportance of the most frequently consumed items re-mained largely the same over the study period (Kendallrsquoscoefficient of concordance W = 0middot86 P lt 0middot001) withno evidence for consistent trends in the consumption ofany prey species (Pearson correlations minus0middot41 lt r lt 0middot57P gt 0middot14 Fig 3a) The numeric proportion of ruralpigeons was fairly stable (23middot1ndash33middot8) as was that ofrabbits (15middot4ndash23middot1) and racing pigeons (6middot6ndash12middot6)The largest variation was recorded for relatively lessimportant prey such as jays (2middot0ndash10middot8) and domesticfowl (1middot2ndash7middot1)
Prey were assigned to 2-week periods from 1 Marchto 15 June (prey per 2 weeks 210middot8 plusmn 177middot6 44ndash504)to investigate variations during the breeding season(Fig 3b) Although there was concordance in diet com-position over time (Kendallrsquos W = 0middot73 P = 0middot001)most prey species showed distinctive temporal trendsThere were marked declines in the dietary importanceof rural (r = minus0middot91 P lt 0middot01) and racing pigeons (r =minus0middot96 P lt 0middot001) which halved their overall numericcontribution from a maximum of about 50 in Marchto a minimum of 23middot7 in June This was largelycompensated for by increases in jays (r = 0middot91 P lt 0middot05)and a wide group of birds represented in lsquootherrsquo prey(r = 0middot92 P lt 0middot01) from a minimum in the second half
Table 2 Diet of Bonellirsquos eagles and prey consumption per pair during the breedingseason in south-west Portugal (1992ndash2001)
Prey categories n Numbers
Biomass
Consumption per pair mean plusmn SD (minimumndashmaximum)
Domestic birdsRural pigeon 451 30middot1 26middot0 48middot0 plusmn 14middot6 (17ndash77)Racing pigeon 136 9middot1 7middot2 14middot7 plusmn 7middot7 (0ndash37)Domestic fowl 52 3middot5 4middot5 5middot4 plusmn 8middot6 (0ndash37)
Wild birdsRed-legged partridge 258 17middot2 14middot9 28middot7 plusmn 11middot9 (6ndash59)Jay 112 7middot5 2middot7 12middot0 plusmn 9middot2 (0ndash28)Other birds 198 13middot4 11middot4 24middot2 plusmn 18middot3 (2ndash68)
MammalsRabbit 276 18middot4 33middot2 30middot4 plusmn 11middot8 (16ndash58)Hare 3 0middot2 lt 0middot1 0middot2 plusmn 0middot6 (0ndash2)ReptilesOcellated lizard 11 0middot7 lt 0middot1 1middot1 plusmn 1middot9 (0ndash6)
Fig 3 Variation of Bonellirsquos eagle diet in south-west Portugal (a) over the years 1993ndash2000 and (b) across the breeding season 1 Marchminus15 June
1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis
1077
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
and domestic prey The aim was to assess (i) the extentof predation on domestic prey (ii) variation in dietacross years and over the breeding season (iii) theinfluence of habitat on predation patterns (iv) the func-tional responses to variation in wild and domestic preyabundance and (v) the eventual occurrence of switchingbetween wild and domestic prey This information wasthen used to infer the predatory mechanisms involved inthis predatorndashprey system and to suggest managementsolutions that might contribute to the conservation ofthese eagles while mitigating potential conflict withhumans in south-west Portugal and elsewhere
Methods
This study focused on a dense breeding nucleus of 25Bonellirsquos eagle pairs occupying about 3000 km
2
in theuplands of south-west Portugal (Fig 1) This is an areaof low mountains and rolling hills mostly under 650 masl in altitude The climate is Mediterranean with a dryand hot summer season with little precipitation Theland is predominantly covered by open to dense corkoak
Quercus suber
L woodland and extensive scrublandoften dominated by the gum cistus
Cistus ladanifer
LIn the western half of the area eucalyptus
Eucalyptusglobulus
Labill plantations for paper pulp productioncover up to 50 of the land Human settlement iscurrently low and local most of the former scattereddwellings are abandoned and agriculture is residual
The diets of 22 pairs of eagles surveyed in 1992ndash2001were described from prey identified in remains(
n
= 747) and regurgitated pellets (
n
= 1039) collectedduring the breeding season (gt 85 in MarchndashMay) atactive nests and surrounding perches together withuncollected fresh prey observed at nests (
n
= 199)Information was obtained for 9middot6
plusmn
4middot8 (2ndash17) pairs peryear with variation as a result of population growthbreeding failures unknown nest locations and reducedfield effort in 1992ndash94 and 2001 Nests were checked5middot5
plusmn
2middot2 (1ndash11) times year
minus
1
The species age and genderof each individual prey were estimated wheneverpossible using published identification keys and areference collection Metallic rings provided a minimumestimate of racing pigeons in the diet as rural pigeonswere seldom ringed within the study area
To reduce biases normally associated with dietarystudies (Real 1996 Marchesi Pedrini amp Sergio 2002)and to increase sample sizes data from pellets remainsand direct observations were combined (Ontiverosamp Pleguezuelos 2000 Penteriani Gallardo amp Roche2002 Ontiveros Pleguezuelos amp Caro 2005) For eachpair and sampling occasion the number of individualsof a given prey species represented in the sample wastaken only from the data source yielding the largestestimate thereby avoiding duplications among datasources In the case of two siblings the estimatednumber of prey items represented in pellets was halvedto avoid duplication of the same prey in two pellets(Real 1996) The biomass of each prey species wasestimated by multiplying the number of individuals byits estimated mean weight using data from the studyarea bibliographic sources (Real 1987 Cardona ampEsteban 2002) and information from racing pigeonfanciers (D Santos personal communication) Thenumber of individuals of each prey species taken byan average eagle pair during the breeding season wasestimated following Petty Lurz amp Rushton (2003)considering one nestling per pair an overall dailyconsumption of 750 g food per pair plus one nestling(Real 1987) a mean incubation period of 39 days(Arroyo Ferreiro amp Garza 1995) and a mean nestlingperiod of 63 days (Real amp Mantildeosa 1998)
Variation in diet landscape composition and preyavailability among territories was used to estimate thefactors influencing eagle predation patterns Given thelack of detailed information on the actual breedingterritories habitat and prey data were quantified withinapproximate territory boundaries estimated usingDirichlet tessellation (Doncaster amp Woodroffe 1993)For each pair the mean geographical location ofnests was the Dirichlet centre and boundaries wereconstrained to be at a maximum of 8 km from the centre(Fig 1) These criteria were based on home range
Fig 1 Location in south-west Portugal of alternative nests of Bonellirsquos eagle pairs(1992ndash2001) and the approximate territory boundaries estimated using Dirichlettessellation
1078
L Palma
et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data from other studies (Mure 2003) and on nearly 800haphazard observations at gt 500 m from active neststhrough the 10-year study period suggesting that eachBonellirsquos eagle pair maintained an exclusive foragingterritory which only rarely extended beyond 8 kmfrom the active nest (L Palma unpublished data)Although this was a relatively crude approach eachrange boundary encompassed gt 90 of observationsof individually recognized elements of the respectivebreeding pair
Landscape composition was quantified using digitalland cover cartography from 1995 (wwwdgrfmin-agriculturaptifnmapashtm accessed 1 April 2005) con-sidering six categories (i) native Mediterranean forestsdominated by cork oaks and strawberry trees
Arbutusunedo
L (ii) eucalyptus plantations (iii) pine (mostly
Pinus pinaster
Ait) plantations (iv) scrubland (v)farmland and (vi) urban areas (Table 1) Landscapediversity was computed using the Shannon index
Rabbit abundance was estimated in SeptemberndashOctober 1997 from latrine counts in 296 250-m linetransects walked along dirt roads and firebreaks crossingeagle territories and neighbouring areas (Palma Bejaamp Rodrigues 1999) Latrine counts were used (Iborra ampLumaret 1997 Beja Palma amp Pais 2007) as directobservation of rabbits was not feasible because of densevegetation cover One transect was walked per 1-km
2
grid square with five random squares surveyed in eachof 65 5-km
2
grid squares distributed in a checkerboardpattern To increase comparability among sites eachtransect was located in the most favourable shrublandhabitats available within each square (Beja Palma ampPais 2007) The mean latrine count per breedingterritory was used to index relative rabbit abundanceOnly 14 territories for which there were at least 10line transects (15middot2
plusmn
4middot2 10ndash23) were used in furtheranalyses because information could not be obtainedfor the entire study area A comprehensive ruralpigeon survey was carried out from September 1994 toDecember 1995 encompassing the approximate breedingterritories of 18 eagle pairs known at the time Pigeonlofts were mapped by visiting all small villages andisolated farmhouses where the number of pigeonswas estimated for each loft either through enquiriesor direct counts Point estimates of pigeon abundancewere interpolated into a 250-m raster grid using kerneldensity analysis with a 5-km searching radius and
these grid values were then used to estimate averagerural pigeon densities within each breeding territoryThe abundance of racing pigeons crossing the studyarea was estimated from information provided byhoming pigeon fanciers on the periods and frequencyof races and training flights and the approximate num-bers of pigeons involved their origin and geographicaldistribution (D Santos personal communication)
The prey data only referred to a restricted periodwhereas the dietary information spanned 10 yearsThis was recognized as a shortcoming in the data butwas unavoidable It was not possible to repeat the surveysand a long diet study would be needed to characterizea large number of pairs This shortcoming was unlikely toaffect the results of this study seriously as unsystematicobservations suggested that regional trends in preyabundance remained essentially the same In particularthere were no marked changes in land cover and humanactivities that might have changed the distribution ofrabbit and pigeon abundances extensively Also norabbit crashes as a result of viral haemorrhagic diseasesimilar to that reported elsewhere (Fa Sharples amp Bell1999) or severe myxomatosis outbreaks were observedMoreover surveys were carried out approximatelyhalf-way through the study period when most dietaryinformation was collected
Preliminary to data analysis the angular transforma-tion (arcsine
radic
p
) was used to reduce the influence ofextreme values and to overcome problems associatedwith the unity sum constraint of proportional data(Zar 1996) Multicollinearity among habitat variableswas investigated using principal component analysis(PCA) thereby describing the main gradients in land-usevariability (Legendre amp Legendre 1998) A varimaxrotation was performed on the principal componentswith eigenvalues gt 1 to enhance their interpretability(Legendre amp Legendre 1998) Seasonal and annualtrends in dietary proportions were examined usinglinear regression including quadratic terms to checkfor non-linear trends (Legendre amp Legendre 1998)Relationships between spatial variation in prey con-sumption and land-use variables were examinedlikewise In temporal analyses data from all eagle pairswere pooled per time period whereas in spatial analyses
Table 1 Average (plusmn SD) landscape composition and summary results of a principal component analysis with varimax rotationdescribing the dominant land-use gradients across Bonellirsquos eagle territories in south-west Portugal (1992ndash2001) The proportionof explained variation and the scores of each variable are provided for the first two rotated axis (PC)
Land cover types Mean () plusmn SD Minimumndashmaximum PC1 (38middot1) PC2 (30middot1)
Scrubland 40middot1 14middot2 16middot8ndash62middot5 0middot84Mediterranean forests 26middot3 12middot6 6middot5ndash46middot4 minus0middot82Farmland 16middot7 15middot7 2middot7ndash55middot6 minus0middot92Eucalyptus plantations 14middot4 12middot2 0middot0ndash43middot8 0middot79Pine plantations 2middot1 2middot7 0middot0ndash7middot5 0middot63Urban 0middot4 0middot4 0middot0ndash1middot4 minus0middot78
1079
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data were pooled per breeding pair Only cases for whichthere were at least 25 prey items were used becausedietary proportions stabilize around this threshold(Ontiveros Pleguezuelos amp Caro 2005)
Functional responses were investigated by modellingthe relationships between the average number of a preytype eaten per breeding pair and the abundance of eitherpigeons or rabbits In each case functions representinglinear (type I) convex (type II) and sigmoidal (type III)curves were fitted to the data (Patterson Benjamin ampMessier 1998) and the best model was selected based onsecond-order Aikaike information criterion (AICc)because of small sample sizes (Burnham amp Anderson1998) Type II and III responses were modelled usingthe generalized MichaelisndashMenton function (Real 1977)The effects of pigeons and rabbits on the consumptionof other prey were investigated using linear regressionand partial correlation analysis (Zar 1996)
Prey switching was examined by relating the ratio ofpigeons to rabbits eaten (
e
1
e
2
) to their availability ratio(
A
1
A
2
) using the equation
e
1
e
2
= (
V
times
A
1
A
2
)
b
(Green-wood amp Elton 1979) The parameter
V
is a measure offrequency-independent selection whereas
b
is a measureof frequency dependence
b
gt 1 provides evidence forswitching with the predator eating disproportionatelymore of the most abundant prey
b
lt 1 reflects dispro-portionate predation on the less common prey and
b
= 1 indicates frequency-independent selection As
A
1
and
A
2
were expressed in distinct units and there wasno absolute estimate of rabbit densities this analysisshould be regarded as approximate and the parameter
V
should not be interpreted Nevertheless the shapeof the curve relating
e
1
e
2
to
A
1
A
2
is meaningfulproviding the best approach to evaluating the presenceof switching between the two prey species
Results
Bonellirsquos eagle territories were dominated by scrublandand oak forests together accounting for almost 70of landscape composition (Table 1) Eucalyptus plan-tations and farmland were also important whereasland cover by pine plantations and urban areas wasnegligible The PCA with varimax rotation revealeda dominant gradient largely reflecting agriculturalabandonment that accounted for 38middot1 of land covervariability ranging from territories with a significantproportion of farmland and urban areas to thosedominated by scrubland The second rotated PCAaxis (30middot8) represented the dominant forest types con-trasting territories dominated by Mediterranean oakforests with those that were predominantly eucalyptusplantations
There was strong variation among territories in theabundance of rabbits and pigeons (Fig 2) Rabbitsoccurred in only 27 of the 296 transects surveyed andthey tended to be fairly scarce where they occurred (4middot2latrines 250 m
minus
1
(4middot3 1ndash24
n
= 80) This resulted in a50-fold variation in rabbit abundance among eagleterritories with densities up to 1ndash5 latrines 250 m
minus
1
along the southern and western edges whereas rabbitswere scarce elsewhere (lt 0middot5 latrines 250 m
minus
1
) A totalof 436 pigeon lofts was identified across the regionwith on average 25middot5
plusmn
37middot7 (2ndash550) pigeons per loftPigeons were more evenly distributed than rabbitsalthough there was about a 10-fold variation amongterritories from about 0middot47 to 4middot71 pigeons km
minus
2
(2middot4
plusmn
1middot2 pigeons km
minus
2
) There was no correlation betweenrabbit and pigeon abundance (r = minus0middot37 P = 0middot262)
Fig 2 Interpolated maps of spatial variation in prey abundances across Bonellirsquos eagles territories in south-west Portugal Valuesare SD from the mean to allow comparisons between maps Dots are rabbit sampling sites and pigeon lofts Dotted lines definethe interpolation limits Hatched lines indicate territories with no information on prey abundance
1080L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
across the 11 territories for which there was informationon both prey
Dietary analysis yielded a minimum estimate of 1497individuals of 32 prey species (see Table S1 in thesupplementary material) Numerically prey of domesticorigin comprised 42middot7 of the diet (37middot7 in biomass)consumed in every year and by all eagle pairs (Table 2)About 70 of these prey items were rural pigeons butracing pigeons and domestic fowl were also consumedfrequently Considering the estimates of prey intake(Table 2) prey densities and territory sizes the averageproportion of the rural pigeons available per eaglepair that were taken through the breeding seasonwas 17middot8 plusmn 12middot6 (5middot7ndash57middot8) Rabbits red-leggedpartridges and jays Garrulus glandarius L were themost frequent wild prey numerically representing43middot1 (50middot8 biomass) of the eagle diet (Table 2)
Of the 114 racing pigeons aged through metallicrings 31middot6 were 1 year old or under 51middot8 2ndash3 yearsold and 16middot7 more than 4 years old The remains offreshly killed domestic fowl observed at nests wereall from juvenile birds (n = 12) whereas 86middot2 of therabbits were adults (n = 29) A majority of partridgeskilled by eagles were males (65middot7 n = 35)
Variation in diet between years was analysed for 1993ndash2000 ie when annual samples were sufficiently large(prey per year 185middot4 plusmn 93middot1 65ndash352) The relativeimportance of the most frequently consumed items re-mained largely the same over the study period (Kendallrsquoscoefficient of concordance W = 0middot86 P lt 0middot001) withno evidence for consistent trends in the consumption ofany prey species (Pearson correlations minus0middot41 lt r lt 0middot57P gt 0middot14 Fig 3a) The numeric proportion of ruralpigeons was fairly stable (23middot1ndash33middot8) as was that ofrabbits (15middot4ndash23middot1) and racing pigeons (6middot6ndash12middot6)The largest variation was recorded for relatively lessimportant prey such as jays (2middot0ndash10middot8) and domesticfowl (1middot2ndash7middot1)
Prey were assigned to 2-week periods from 1 Marchto 15 June (prey per 2 weeks 210middot8 plusmn 177middot6 44ndash504)to investigate variations during the breeding season(Fig 3b) Although there was concordance in diet com-position over time (Kendallrsquos W = 0middot73 P = 0middot001)most prey species showed distinctive temporal trendsThere were marked declines in the dietary importanceof rural (r = minus0middot91 P lt 0middot01) and racing pigeons (r =minus0middot96 P lt 0middot001) which halved their overall numericcontribution from a maximum of about 50 in Marchto a minimum of 23middot7 in June This was largelycompensated for by increases in jays (r = 0middot91 P lt 0middot05)and a wide group of birds represented in lsquootherrsquo prey(r = 0middot92 P lt 0middot01) from a minimum in the second half
Table 2 Diet of Bonellirsquos eagles and prey consumption per pair during the breedingseason in south-west Portugal (1992ndash2001)
Prey categories n Numbers
Biomass
Consumption per pair mean plusmn SD (minimumndashmaximum)
Domestic birdsRural pigeon 451 30middot1 26middot0 48middot0 plusmn 14middot6 (17ndash77)Racing pigeon 136 9middot1 7middot2 14middot7 plusmn 7middot7 (0ndash37)Domestic fowl 52 3middot5 4middot5 5middot4 plusmn 8middot6 (0ndash37)
Wild birdsRed-legged partridge 258 17middot2 14middot9 28middot7 plusmn 11middot9 (6ndash59)Jay 112 7middot5 2middot7 12middot0 plusmn 9middot2 (0ndash28)Other birds 198 13middot4 11middot4 24middot2 plusmn 18middot3 (2ndash68)
MammalsRabbit 276 18middot4 33middot2 30middot4 plusmn 11middot8 (16ndash58)Hare 3 0middot2 lt 0middot1 0middot2 plusmn 0middot6 (0ndash2)ReptilesOcellated lizard 11 0middot7 lt 0middot1 1middot1 plusmn 1middot9 (0ndash6)
Fig 3 Variation of Bonellirsquos eagle diet in south-west Portugal (a) over the years 1993ndash2000 and (b) across the breeding season 1 Marchminus15 June
1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1078
L Palma
et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data from other studies (Mure 2003) and on nearly 800haphazard observations at gt 500 m from active neststhrough the 10-year study period suggesting that eachBonellirsquos eagle pair maintained an exclusive foragingterritory which only rarely extended beyond 8 kmfrom the active nest (L Palma unpublished data)Although this was a relatively crude approach eachrange boundary encompassed gt 90 of observationsof individually recognized elements of the respectivebreeding pair
Landscape composition was quantified using digitalland cover cartography from 1995 (wwwdgrfmin-agriculturaptifnmapashtm accessed 1 April 2005) con-sidering six categories (i) native Mediterranean forestsdominated by cork oaks and strawberry trees
Arbutusunedo
L (ii) eucalyptus plantations (iii) pine (mostly
Pinus pinaster
Ait) plantations (iv) scrubland (v)farmland and (vi) urban areas (Table 1) Landscapediversity was computed using the Shannon index
Rabbit abundance was estimated in SeptemberndashOctober 1997 from latrine counts in 296 250-m linetransects walked along dirt roads and firebreaks crossingeagle territories and neighbouring areas (Palma Bejaamp Rodrigues 1999) Latrine counts were used (Iborra ampLumaret 1997 Beja Palma amp Pais 2007) as directobservation of rabbits was not feasible because of densevegetation cover One transect was walked per 1-km
2
grid square with five random squares surveyed in eachof 65 5-km
2
grid squares distributed in a checkerboardpattern To increase comparability among sites eachtransect was located in the most favourable shrublandhabitats available within each square (Beja Palma ampPais 2007) The mean latrine count per breedingterritory was used to index relative rabbit abundanceOnly 14 territories for which there were at least 10line transects (15middot2
plusmn
4middot2 10ndash23) were used in furtheranalyses because information could not be obtainedfor the entire study area A comprehensive ruralpigeon survey was carried out from September 1994 toDecember 1995 encompassing the approximate breedingterritories of 18 eagle pairs known at the time Pigeonlofts were mapped by visiting all small villages andisolated farmhouses where the number of pigeonswas estimated for each loft either through enquiriesor direct counts Point estimates of pigeon abundancewere interpolated into a 250-m raster grid using kerneldensity analysis with a 5-km searching radius and
these grid values were then used to estimate averagerural pigeon densities within each breeding territoryThe abundance of racing pigeons crossing the studyarea was estimated from information provided byhoming pigeon fanciers on the periods and frequencyof races and training flights and the approximate num-bers of pigeons involved their origin and geographicaldistribution (D Santos personal communication)
The prey data only referred to a restricted periodwhereas the dietary information spanned 10 yearsThis was recognized as a shortcoming in the data butwas unavoidable It was not possible to repeat the surveysand a long diet study would be needed to characterizea large number of pairs This shortcoming was unlikely toaffect the results of this study seriously as unsystematicobservations suggested that regional trends in preyabundance remained essentially the same In particularthere were no marked changes in land cover and humanactivities that might have changed the distribution ofrabbit and pigeon abundances extensively Also norabbit crashes as a result of viral haemorrhagic diseasesimilar to that reported elsewhere (Fa Sharples amp Bell1999) or severe myxomatosis outbreaks were observedMoreover surveys were carried out approximatelyhalf-way through the study period when most dietaryinformation was collected
Preliminary to data analysis the angular transforma-tion (arcsine
radic
p
) was used to reduce the influence ofextreme values and to overcome problems associatedwith the unity sum constraint of proportional data(Zar 1996) Multicollinearity among habitat variableswas investigated using principal component analysis(PCA) thereby describing the main gradients in land-usevariability (Legendre amp Legendre 1998) A varimaxrotation was performed on the principal componentswith eigenvalues gt 1 to enhance their interpretability(Legendre amp Legendre 1998) Seasonal and annualtrends in dietary proportions were examined usinglinear regression including quadratic terms to checkfor non-linear trends (Legendre amp Legendre 1998)Relationships between spatial variation in prey con-sumption and land-use variables were examinedlikewise In temporal analyses data from all eagle pairswere pooled per time period whereas in spatial analyses
Table 1 Average (plusmn SD) landscape composition and summary results of a principal component analysis with varimax rotationdescribing the dominant land-use gradients across Bonellirsquos eagle territories in south-west Portugal (1992ndash2001) The proportionof explained variation and the scores of each variable are provided for the first two rotated axis (PC)
Land cover types Mean () plusmn SD Minimumndashmaximum PC1 (38middot1) PC2 (30middot1)
Scrubland 40middot1 14middot2 16middot8ndash62middot5 0middot84Mediterranean forests 26middot3 12middot6 6middot5ndash46middot4 minus0middot82Farmland 16middot7 15middot7 2middot7ndash55middot6 minus0middot92Eucalyptus plantations 14middot4 12middot2 0middot0ndash43middot8 0middot79Pine plantations 2middot1 2middot7 0middot0ndash7middot5 0middot63Urban 0middot4 0middot4 0middot0ndash1middot4 minus0middot78
1079
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data were pooled per breeding pair Only cases for whichthere were at least 25 prey items were used becausedietary proportions stabilize around this threshold(Ontiveros Pleguezuelos amp Caro 2005)
Functional responses were investigated by modellingthe relationships between the average number of a preytype eaten per breeding pair and the abundance of eitherpigeons or rabbits In each case functions representinglinear (type I) convex (type II) and sigmoidal (type III)curves were fitted to the data (Patterson Benjamin ampMessier 1998) and the best model was selected based onsecond-order Aikaike information criterion (AICc)because of small sample sizes (Burnham amp Anderson1998) Type II and III responses were modelled usingthe generalized MichaelisndashMenton function (Real 1977)The effects of pigeons and rabbits on the consumptionof other prey were investigated using linear regressionand partial correlation analysis (Zar 1996)
Prey switching was examined by relating the ratio ofpigeons to rabbits eaten (
e
1
e
2
) to their availability ratio(
A
1
A
2
) using the equation
e
1
e
2
= (
V
times
A
1
A
2
)
b
(Green-wood amp Elton 1979) The parameter
V
is a measure offrequency-independent selection whereas
b
is a measureof frequency dependence
b
gt 1 provides evidence forswitching with the predator eating disproportionatelymore of the most abundant prey
b
lt 1 reflects dispro-portionate predation on the less common prey and
b
= 1 indicates frequency-independent selection As
A
1
and
A
2
were expressed in distinct units and there wasno absolute estimate of rabbit densities this analysisshould be regarded as approximate and the parameter
V
should not be interpreted Nevertheless the shapeof the curve relating
e
1
e
2
to
A
1
A
2
is meaningfulproviding the best approach to evaluating the presenceof switching between the two prey species
Results
Bonellirsquos eagle territories were dominated by scrublandand oak forests together accounting for almost 70of landscape composition (Table 1) Eucalyptus plan-tations and farmland were also important whereasland cover by pine plantations and urban areas wasnegligible The PCA with varimax rotation revealeda dominant gradient largely reflecting agriculturalabandonment that accounted for 38middot1 of land covervariability ranging from territories with a significantproportion of farmland and urban areas to thosedominated by scrubland The second rotated PCAaxis (30middot8) represented the dominant forest types con-trasting territories dominated by Mediterranean oakforests with those that were predominantly eucalyptusplantations
There was strong variation among territories in theabundance of rabbits and pigeons (Fig 2) Rabbitsoccurred in only 27 of the 296 transects surveyed andthey tended to be fairly scarce where they occurred (4middot2latrines 250 m
minus
1
(4middot3 1ndash24
n
= 80) This resulted in a50-fold variation in rabbit abundance among eagleterritories with densities up to 1ndash5 latrines 250 m
minus
1
along the southern and western edges whereas rabbitswere scarce elsewhere (lt 0middot5 latrines 250 m
minus
1
) A totalof 436 pigeon lofts was identified across the regionwith on average 25middot5
plusmn
37middot7 (2ndash550) pigeons per loftPigeons were more evenly distributed than rabbitsalthough there was about a 10-fold variation amongterritories from about 0middot47 to 4middot71 pigeons km
minus
2
(2middot4
plusmn
1middot2 pigeons km
minus
2
) There was no correlation betweenrabbit and pigeon abundance (r = minus0middot37 P = 0middot262)
Fig 2 Interpolated maps of spatial variation in prey abundances across Bonellirsquos eagles territories in south-west Portugal Valuesare SD from the mean to allow comparisons between maps Dots are rabbit sampling sites and pigeon lofts Dotted lines definethe interpolation limits Hatched lines indicate territories with no information on prey abundance
1080L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
across the 11 territories for which there was informationon both prey
Dietary analysis yielded a minimum estimate of 1497individuals of 32 prey species (see Table S1 in thesupplementary material) Numerically prey of domesticorigin comprised 42middot7 of the diet (37middot7 in biomass)consumed in every year and by all eagle pairs (Table 2)About 70 of these prey items were rural pigeons butracing pigeons and domestic fowl were also consumedfrequently Considering the estimates of prey intake(Table 2) prey densities and territory sizes the averageproportion of the rural pigeons available per eaglepair that were taken through the breeding seasonwas 17middot8 plusmn 12middot6 (5middot7ndash57middot8) Rabbits red-leggedpartridges and jays Garrulus glandarius L were themost frequent wild prey numerically representing43middot1 (50middot8 biomass) of the eagle diet (Table 2)
Of the 114 racing pigeons aged through metallicrings 31middot6 were 1 year old or under 51middot8 2ndash3 yearsold and 16middot7 more than 4 years old The remains offreshly killed domestic fowl observed at nests wereall from juvenile birds (n = 12) whereas 86middot2 of therabbits were adults (n = 29) A majority of partridgeskilled by eagles were males (65middot7 n = 35)
Variation in diet between years was analysed for 1993ndash2000 ie when annual samples were sufficiently large(prey per year 185middot4 plusmn 93middot1 65ndash352) The relativeimportance of the most frequently consumed items re-mained largely the same over the study period (Kendallrsquoscoefficient of concordance W = 0middot86 P lt 0middot001) withno evidence for consistent trends in the consumption ofany prey species (Pearson correlations minus0middot41 lt r lt 0middot57P gt 0middot14 Fig 3a) The numeric proportion of ruralpigeons was fairly stable (23middot1ndash33middot8) as was that ofrabbits (15middot4ndash23middot1) and racing pigeons (6middot6ndash12middot6)The largest variation was recorded for relatively lessimportant prey such as jays (2middot0ndash10middot8) and domesticfowl (1middot2ndash7middot1)
Prey were assigned to 2-week periods from 1 Marchto 15 June (prey per 2 weeks 210middot8 plusmn 177middot6 44ndash504)to investigate variations during the breeding season(Fig 3b) Although there was concordance in diet com-position over time (Kendallrsquos W = 0middot73 P = 0middot001)most prey species showed distinctive temporal trendsThere were marked declines in the dietary importanceof rural (r = minus0middot91 P lt 0middot01) and racing pigeons (r =minus0middot96 P lt 0middot001) which halved their overall numericcontribution from a maximum of about 50 in Marchto a minimum of 23middot7 in June This was largelycompensated for by increases in jays (r = 0middot91 P lt 0middot05)and a wide group of birds represented in lsquootherrsquo prey(r = 0middot92 P lt 0middot01) from a minimum in the second half
Table 2 Diet of Bonellirsquos eagles and prey consumption per pair during the breedingseason in south-west Portugal (1992ndash2001)
Prey categories n Numbers
Biomass
Consumption per pair mean plusmn SD (minimumndashmaximum)
Domestic birdsRural pigeon 451 30middot1 26middot0 48middot0 plusmn 14middot6 (17ndash77)Racing pigeon 136 9middot1 7middot2 14middot7 plusmn 7middot7 (0ndash37)Domestic fowl 52 3middot5 4middot5 5middot4 plusmn 8middot6 (0ndash37)
Wild birdsRed-legged partridge 258 17middot2 14middot9 28middot7 plusmn 11middot9 (6ndash59)Jay 112 7middot5 2middot7 12middot0 plusmn 9middot2 (0ndash28)Other birds 198 13middot4 11middot4 24middot2 plusmn 18middot3 (2ndash68)
MammalsRabbit 276 18middot4 33middot2 30middot4 plusmn 11middot8 (16ndash58)Hare 3 0middot2 lt 0middot1 0middot2 plusmn 0middot6 (0ndash2)ReptilesOcellated lizard 11 0middot7 lt 0middot1 1middot1 plusmn 1middot9 (0ndash6)
Fig 3 Variation of Bonellirsquos eagle diet in south-west Portugal (a) over the years 1993ndash2000 and (b) across the breeding season 1 Marchminus15 June
1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1079
Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society
Journal of Applied Ecology
43
1075ndash1086
data were pooled per breeding pair Only cases for whichthere were at least 25 prey items were used becausedietary proportions stabilize around this threshold(Ontiveros Pleguezuelos amp Caro 2005)
Functional responses were investigated by modellingthe relationships between the average number of a preytype eaten per breeding pair and the abundance of eitherpigeons or rabbits In each case functions representinglinear (type I) convex (type II) and sigmoidal (type III)curves were fitted to the data (Patterson Benjamin ampMessier 1998) and the best model was selected based onsecond-order Aikaike information criterion (AICc)because of small sample sizes (Burnham amp Anderson1998) Type II and III responses were modelled usingthe generalized MichaelisndashMenton function (Real 1977)The effects of pigeons and rabbits on the consumptionof other prey were investigated using linear regressionand partial correlation analysis (Zar 1996)
Prey switching was examined by relating the ratio ofpigeons to rabbits eaten (
e
1
e
2
) to their availability ratio(
A
1
A
2
) using the equation
e
1
e
2
= (
V
times
A
1
A
2
)
b
(Green-wood amp Elton 1979) The parameter
V
is a measure offrequency-independent selection whereas
b
is a measureof frequency dependence
b
gt 1 provides evidence forswitching with the predator eating disproportionatelymore of the most abundant prey
b
lt 1 reflects dispro-portionate predation on the less common prey and
b
= 1 indicates frequency-independent selection As
A
1
and
A
2
were expressed in distinct units and there wasno absolute estimate of rabbit densities this analysisshould be regarded as approximate and the parameter
V
should not be interpreted Nevertheless the shapeof the curve relating
e
1
e
2
to
A
1
A
2
is meaningfulproviding the best approach to evaluating the presenceof switching between the two prey species
Results
Bonellirsquos eagle territories were dominated by scrublandand oak forests together accounting for almost 70of landscape composition (Table 1) Eucalyptus plan-tations and farmland were also important whereasland cover by pine plantations and urban areas wasnegligible The PCA with varimax rotation revealeda dominant gradient largely reflecting agriculturalabandonment that accounted for 38middot1 of land covervariability ranging from territories with a significantproportion of farmland and urban areas to thosedominated by scrubland The second rotated PCAaxis (30middot8) represented the dominant forest types con-trasting territories dominated by Mediterranean oakforests with those that were predominantly eucalyptusplantations
There was strong variation among territories in theabundance of rabbits and pigeons (Fig 2) Rabbitsoccurred in only 27 of the 296 transects surveyed andthey tended to be fairly scarce where they occurred (4middot2latrines 250 m
minus
1
(4middot3 1ndash24
n
= 80) This resulted in a50-fold variation in rabbit abundance among eagleterritories with densities up to 1ndash5 latrines 250 m
minus
1
along the southern and western edges whereas rabbitswere scarce elsewhere (lt 0middot5 latrines 250 m
minus
1
) A totalof 436 pigeon lofts was identified across the regionwith on average 25middot5
plusmn
37middot7 (2ndash550) pigeons per loftPigeons were more evenly distributed than rabbitsalthough there was about a 10-fold variation amongterritories from about 0middot47 to 4middot71 pigeons km
minus
2
(2middot4
plusmn
1middot2 pigeons km
minus
2
) There was no correlation betweenrabbit and pigeon abundance (r = minus0middot37 P = 0middot262)
Fig 2 Interpolated maps of spatial variation in prey abundances across Bonellirsquos eagles territories in south-west Portugal Valuesare SD from the mean to allow comparisons between maps Dots are rabbit sampling sites and pigeon lofts Dotted lines definethe interpolation limits Hatched lines indicate territories with no information on prey abundance
1080L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
across the 11 territories for which there was informationon both prey
Dietary analysis yielded a minimum estimate of 1497individuals of 32 prey species (see Table S1 in thesupplementary material) Numerically prey of domesticorigin comprised 42middot7 of the diet (37middot7 in biomass)consumed in every year and by all eagle pairs (Table 2)About 70 of these prey items were rural pigeons butracing pigeons and domestic fowl were also consumedfrequently Considering the estimates of prey intake(Table 2) prey densities and territory sizes the averageproportion of the rural pigeons available per eaglepair that were taken through the breeding seasonwas 17middot8 plusmn 12middot6 (5middot7ndash57middot8) Rabbits red-leggedpartridges and jays Garrulus glandarius L were themost frequent wild prey numerically representing43middot1 (50middot8 biomass) of the eagle diet (Table 2)
Of the 114 racing pigeons aged through metallicrings 31middot6 were 1 year old or under 51middot8 2ndash3 yearsold and 16middot7 more than 4 years old The remains offreshly killed domestic fowl observed at nests wereall from juvenile birds (n = 12) whereas 86middot2 of therabbits were adults (n = 29) A majority of partridgeskilled by eagles were males (65middot7 n = 35)
Variation in diet between years was analysed for 1993ndash2000 ie when annual samples were sufficiently large(prey per year 185middot4 plusmn 93middot1 65ndash352) The relativeimportance of the most frequently consumed items re-mained largely the same over the study period (Kendallrsquoscoefficient of concordance W = 0middot86 P lt 0middot001) withno evidence for consistent trends in the consumption ofany prey species (Pearson correlations minus0middot41 lt r lt 0middot57P gt 0middot14 Fig 3a) The numeric proportion of ruralpigeons was fairly stable (23middot1ndash33middot8) as was that ofrabbits (15middot4ndash23middot1) and racing pigeons (6middot6ndash12middot6)The largest variation was recorded for relatively lessimportant prey such as jays (2middot0ndash10middot8) and domesticfowl (1middot2ndash7middot1)
Prey were assigned to 2-week periods from 1 Marchto 15 June (prey per 2 weeks 210middot8 plusmn 177middot6 44ndash504)to investigate variations during the breeding season(Fig 3b) Although there was concordance in diet com-position over time (Kendallrsquos W = 0middot73 P = 0middot001)most prey species showed distinctive temporal trendsThere were marked declines in the dietary importanceof rural (r = minus0middot91 P lt 0middot01) and racing pigeons (r =minus0middot96 P lt 0middot001) which halved their overall numericcontribution from a maximum of about 50 in Marchto a minimum of 23middot7 in June This was largelycompensated for by increases in jays (r = 0middot91 P lt 0middot05)and a wide group of birds represented in lsquootherrsquo prey(r = 0middot92 P lt 0middot01) from a minimum in the second half
Table 2 Diet of Bonellirsquos eagles and prey consumption per pair during the breedingseason in south-west Portugal (1992ndash2001)
Prey categories n Numbers
Biomass
Consumption per pair mean plusmn SD (minimumndashmaximum)
Domestic birdsRural pigeon 451 30middot1 26middot0 48middot0 plusmn 14middot6 (17ndash77)Racing pigeon 136 9middot1 7middot2 14middot7 plusmn 7middot7 (0ndash37)Domestic fowl 52 3middot5 4middot5 5middot4 plusmn 8middot6 (0ndash37)
Wild birdsRed-legged partridge 258 17middot2 14middot9 28middot7 plusmn 11middot9 (6ndash59)Jay 112 7middot5 2middot7 12middot0 plusmn 9middot2 (0ndash28)Other birds 198 13middot4 11middot4 24middot2 plusmn 18middot3 (2ndash68)
MammalsRabbit 276 18middot4 33middot2 30middot4 plusmn 11middot8 (16ndash58)Hare 3 0middot2 lt 0middot1 0middot2 plusmn 0middot6 (0ndash2)ReptilesOcellated lizard 11 0middot7 lt 0middot1 1middot1 plusmn 1middot9 (0ndash6)
Fig 3 Variation of Bonellirsquos eagle diet in south-west Portugal (a) over the years 1993ndash2000 and (b) across the breeding season 1 Marchminus15 June
1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1080L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
across the 11 territories for which there was informationon both prey
Dietary analysis yielded a minimum estimate of 1497individuals of 32 prey species (see Table S1 in thesupplementary material) Numerically prey of domesticorigin comprised 42middot7 of the diet (37middot7 in biomass)consumed in every year and by all eagle pairs (Table 2)About 70 of these prey items were rural pigeons butracing pigeons and domestic fowl were also consumedfrequently Considering the estimates of prey intake(Table 2) prey densities and territory sizes the averageproportion of the rural pigeons available per eaglepair that were taken through the breeding seasonwas 17middot8 plusmn 12middot6 (5middot7ndash57middot8) Rabbits red-leggedpartridges and jays Garrulus glandarius L were themost frequent wild prey numerically representing43middot1 (50middot8 biomass) of the eagle diet (Table 2)
Of the 114 racing pigeons aged through metallicrings 31middot6 were 1 year old or under 51middot8 2ndash3 yearsold and 16middot7 more than 4 years old The remains offreshly killed domestic fowl observed at nests wereall from juvenile birds (n = 12) whereas 86middot2 of therabbits were adults (n = 29) A majority of partridgeskilled by eagles were males (65middot7 n = 35)
Variation in diet between years was analysed for 1993ndash2000 ie when annual samples were sufficiently large(prey per year 185middot4 plusmn 93middot1 65ndash352) The relativeimportance of the most frequently consumed items re-mained largely the same over the study period (Kendallrsquoscoefficient of concordance W = 0middot86 P lt 0middot001) withno evidence for consistent trends in the consumption ofany prey species (Pearson correlations minus0middot41 lt r lt 0middot57P gt 0middot14 Fig 3a) The numeric proportion of ruralpigeons was fairly stable (23middot1ndash33middot8) as was that ofrabbits (15middot4ndash23middot1) and racing pigeons (6middot6ndash12middot6)The largest variation was recorded for relatively lessimportant prey such as jays (2middot0ndash10middot8) and domesticfowl (1middot2ndash7middot1)
Prey were assigned to 2-week periods from 1 Marchto 15 June (prey per 2 weeks 210middot8 plusmn 177middot6 44ndash504)to investigate variations during the breeding season(Fig 3b) Although there was concordance in diet com-position over time (Kendallrsquos W = 0middot73 P = 0middot001)most prey species showed distinctive temporal trendsThere were marked declines in the dietary importanceof rural (r = minus0middot91 P lt 0middot01) and racing pigeons (r =minus0middot96 P lt 0middot001) which halved their overall numericcontribution from a maximum of about 50 in Marchto a minimum of 23middot7 in June This was largelycompensated for by increases in jays (r = 0middot91 P lt 0middot05)and a wide group of birds represented in lsquootherrsquo prey(r = 0middot92 P lt 0middot01) from a minimum in the second half
Table 2 Diet of Bonellirsquos eagles and prey consumption per pair during the breedingseason in south-west Portugal (1992ndash2001)
Prey categories n Numbers
Biomass
Consumption per pair mean plusmn SD (minimumndashmaximum)
Domestic birdsRural pigeon 451 30middot1 26middot0 48middot0 plusmn 14middot6 (17ndash77)Racing pigeon 136 9middot1 7middot2 14middot7 plusmn 7middot7 (0ndash37)Domestic fowl 52 3middot5 4middot5 5middot4 plusmn 8middot6 (0ndash37)
Wild birdsRed-legged partridge 258 17middot2 14middot9 28middot7 plusmn 11middot9 (6ndash59)Jay 112 7middot5 2middot7 12middot0 plusmn 9middot2 (0ndash28)Other birds 198 13middot4 11middot4 24middot2 plusmn 18middot3 (2ndash68)
MammalsRabbit 276 18middot4 33middot2 30middot4 plusmn 11middot8 (16ndash58)Hare 3 0middot2 lt 0middot1 0middot2 plusmn 0middot6 (0ndash2)ReptilesOcellated lizard 11 0middot7 lt 0middot1 1middot1 plusmn 1middot9 (0ndash6)
Fig 3 Variation of Bonellirsquos eagle diet in south-west Portugal (a) over the years 1993ndash2000 and (b) across the breeding season 1 Marchminus15 June
1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1081Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
of March (0middot8 and 8middot9) up to a maximum in June(10middot2 and 30middot5) Patterns for other prey species weremore complex A unimodal pattern was found forpartridges (multiple r = 0middot96 P = 0middot01) that increasedfrom about 2 in early March up to a maximum ataround 20 in AprilndashMay and declined thereafter Theconsumption of rabbits did not show any significanttrend through the breeding season
Land use explained 22ndash61 of variation in the dietaryproportions of both domestic and wild prey (Table 3)The consumption of rural pigeons declined steadilywith increasing eucalyptus cover while racing pigeonsshowed unimodal responses to Mediterranean oakforests The intake of domestic fowl was apparentlynot affected by landscape composition Partridgeconsumption increased with oak forests declinedwith eucalyptus cover farmland and urban areas andresponded unimodally to scrubland The other preycategory and diet diversity increased markedly witheucalyptus plantation cover Diet diversity and theintake of jays showed unimodal responses to farmlandThe unimodal response to urban areas was the onlylandscape effect recorded for rabbits
A type II functional curve significantly explained halfthe variation in rabbit consumption (Fig 4a) whilethere was no support for alternative response types(∆AICc gt 2middot5) The abundance of rabbits also affectedthe consumption of other prey with less rural andracing pigeons eaten in territories with more rabbits(Fig 5a) The inverse relationship between rabbitabundances and partridge consumption was marginallysignificant (P lt 0middot051) When controlling for the effectsof rural pigeon densities using partial correlationsrabbit abundances still showed significant inverserelationships with rural (r = minus0middot63 P = 0middot050) and
racing pigeons (r = minus0middot75 P = 0middot013) and approachedsignificance with partridges (r = minus0middot57 P = 0middot085)Although the functional response curve accounted forlt 25 in the consumption of rural pigeons (Fig 4b)the type II model still performed better than the twoalternative models (∆AICc gt 2middot5) The number ofrabbits eaten was inversely related to pigeon densities(Fig 5b) even after controlling for rabbit abundancesusing partial correlations (r = minus0middot81 P = 0middot004) Shannondiet diversity computed at the species level wasunrelated to the abundance of both rabbits (r = 0middot33P = 0middot317) and pigeons (r = minus0middot27 P = 0middot280)
The ratio of pigeons to rabbits consumed per eaglepair increased significantly with the ratio of pigeon torabbit abundances (Fig 6) The frequency-dependentparameter b was significantly smaller than unity(95 confidence interval 0middot09ndash0middot42) suggesting thatprey selection was inversely frequency dependentThis indicated that at low pigeon to rabbit ratios eaglescaptured more pigeons than might be expected whereasmore rabbits than expected were taken in breedingterritories with high pigeon to rabbit ratios There wastherefore no evidence of Bonellirsquos eagles switchingto pigeons when the abundance of rabbits declined inrelation to that of pigeons Instead the less abundantprey always tended to be consumed more than expected
Discussion
Domestic prey played a major role in the diet ofBonellirsquos eagles in south-west Portugal This resultedprimarily from the high consumption of rural pigeonswhich was the single most important prey itemalthough racing pigeons and domestic fowl were alsotaken frequently These were major dietary componentsfor a very large proportion of this eagle populationand their importance was consistently high over adecade Domestic prey was particularly important earlyin the breeding season when it accounted for more thanhalf the prey eaten Overall however wild prey such asrabbits partridges and jays were a major dietary com-ponent of most eagle pairs These species together withseveral minor avian prey such as cattle egrets Bubulcusibis L thrushes Turdus spp and woodpeckers Picusviridis L became increasingly important later in thebreeding season
The importance of pigeons in south-west Portugalparticularly that of rural pigeons was higher than thatreported for most Bonellirsquos eagle populations (39middot2vs 19middot2 plusmn 12middot8 1middot5ndash39middot0 n = 14 Valkama et al2005) Although some studies found a relatively highconsumption of woodpigeons Columba palumbus L(Real 1991 Ontiveros Pleguezuelos amp Caro 2005) theoverall dietary contribution of pigeons was still lowerthan that observed in south-west Portugal Only in thediet of peregrine falcons and sometimes in goshawks
Table 3 Explained variance (R2) significance levels and directions of association (ndashnegative + positive cap unimodal) for significant relationships between Bonellirsquos eaglediet and land uses across 20 breeding territories in south-west Portugal
Prey categories Oak forest Eucalyptus forest Scrubland Farmland Urban
Domestic preyRural pigeon ndasha0middot40Racing pigeon cap0middot41Wild preyRabbit cap0middot45Partridge +a0middot38 ndasha0middot33 cap0middot35 minus0middot22 ndasha0middot22Jay cap0middot45Other birds +a0middot61Diversity +a0middot33 cap0middot37
P lt 0middot05 P lt 0middot01 P lt 0middot001 a Quadratic term
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis
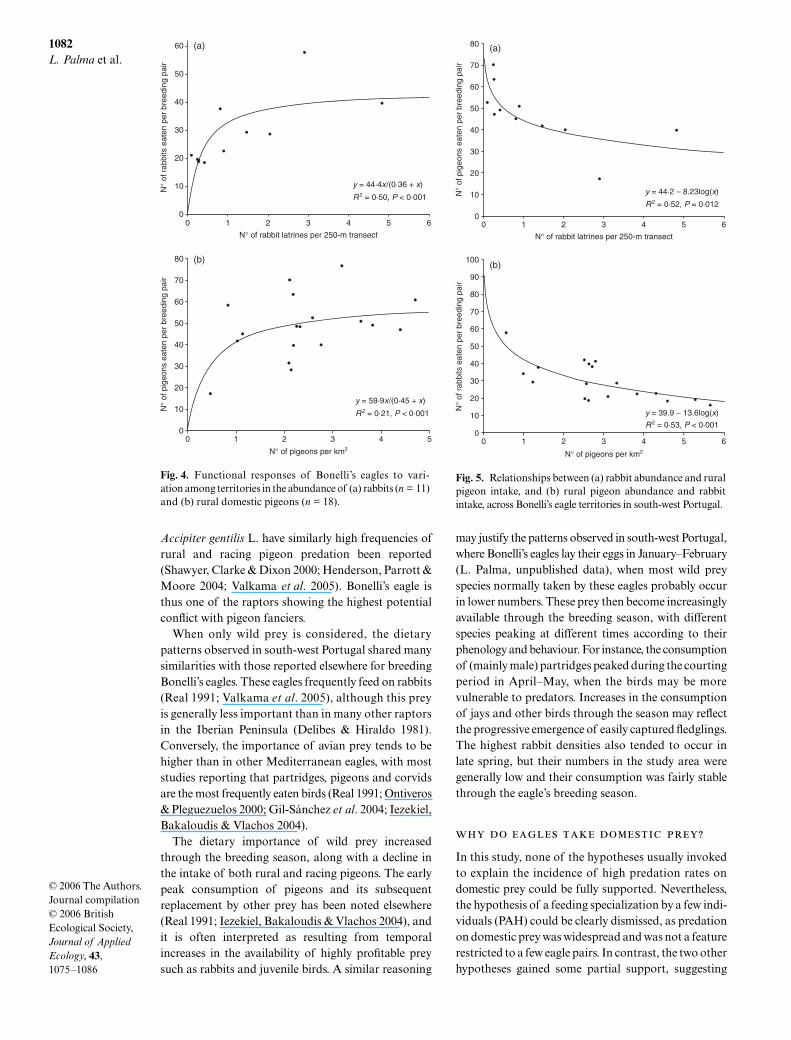
1082L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Accipiter gentilis L have similarly high frequencies ofrural and racing pigeon predation been reported(Shawyer Clarke amp Dixon 2000 Henderson Parrott ampMoore 2004 Valkama et al 2005) Bonellirsquos eagle isthus one of the raptors showing the highest potentialconflict with pigeon fanciers
When only wild prey is considered the dietarypatterns observed in south-west Portugal shared manysimilarities with those reported elsewhere for breedingBonellirsquos eagles These eagles frequently feed on rabbits(Real 1991 Valkama et al 2005) although this preyis generally less important than in many other raptorsin the Iberian Peninsula (Delibes amp Hiraldo 1981)Conversely the importance of avian prey tends to behigher than in other Mediterranean eagles with moststudies reporting that partridges pigeons and corvidsare the most frequently eaten birds (Real 1991 Ontiverosamp Pleguezuelos 2000 Gil-Saacutenchez et al 2004 IezekielBakaloudis amp Vlachos 2004)
The dietary importance of wild prey increasedthrough the breeding season along with a decline inthe intake of both rural and racing pigeons The earlypeak consumption of pigeons and its subsequentreplacement by other prey has been noted elsewhere(Real 1991 Iezekiel Bakaloudis amp Vlachos 2004) andit is often interpreted as resulting from temporalincreases in the availability of highly profitable preysuch as rabbits and juvenile birds A similar reasoning
may justify the patterns observed in south-west Portugalwhere Bonellirsquos eagles lay their eggs in JanuaryndashFebruary(L Palma unpublished data) when most wild preyspecies normally taken by these eagles probably occurin lower numbers These prey then become increasinglyavailable through the breeding season with differentspecies peaking at different times according to theirphenology and behaviour For instance the consumptionof (mainly male) partridges peaked during the courtingperiod in AprilndashMay when the birds may be morevulnerable to predators Increases in the consumptionof jays and other birds through the season may reflectthe progressive emergence of easily captured fledglingsThe highest rabbit densities also tended to occur inlate spring but their numbers in the study area weregenerally low and their consumption was fairly stablethrough the eaglersquos breeding season
In this study none of the hypotheses usually invokedto explain the incidence of high predation rates ondomestic prey could be fully supported Neverthelessthe hypothesis of a feeding specialization by a few indi-viduals (PAH) could be clearly dismissed as predationon domestic prey was widespread and was not a featurerestricted to a few eagle pairs In contrast the two otherhypotheses gained some partial support suggesting
Fig 4 Functional responses of Bonellirsquos eagles to vari-ation among territories in the abundance of (a) rabbits (n = 11)and (b) rural domestic pigeons (n = 18)
Fig 5 Relationships between (a) rabbit abundance and ruralpigeon intake and (b) rural pigeon abundance and rabbitintake across Bonellirsquos eagle territories in south-west Portugal
1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1083Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
that the heavy predation by Bonellirsquos eagles on domesticprey recorded in south-west Portugal might result fromboth a shortage of wild prey in accordance with APHand a positive selection of domestic prey following theVPH
Compared with other areas there was a shortagein south-west Portugal of those wild prey speciescommonly favoured by the Bonellirsquos eagle such asrabbits and partridges (Gil-Saacutenchez 1998) Rabbits wereparticularly scarce as they were largely absent frommost of the region and their abundance was reducedwhere they occurred (Palma Beja amp Rodrigues 1999Beja Palma amp Pais 2007 this study) Unsystematicobservations suggested that partridges also occurred atlow densities across the region (L Palma unpublisheddata) In contrast rural pigeons were very abundantthrough the year Information from homing pigeonassociations suggested that racing pigeons were alsovery abundant with at least 10ndash15 thousand pigeonscrossing the study area each week in JanuaryndashAprilStrayed or lost pigeons could thus be commonly foundwithin the study area (L Palma unpublished data)presumably providing easy and rewarding prey forraptors (Shawyer Clarke amp Dixon 2000 HendersonParrott amp Moore 2004)
Breeding territories with the lowest abundance ofboth wild and domestic prey were probably thosewith high eucalyptus cover where there was reducedconsumption of rural pigeons and partridges alongwith increases in both the intake of minor prey itemsand diet diversity Considering the predictions ofoptimal foraging theory (Stephens amp Krebs 1986) andthe findings of empirical studies on other birds of prey(Lindeacuten amp Wikman 1983 Marchesi Sergio amp Pedrini2002) these dietary responses are consistent with ashift to low-ranked prey and an associated widening ofniche breadth in a situation where there is reducedavailability of the most profitable prey Previous studiesand personal observations suggest that both rabbitsand partridges avoid eucalyptus landscapes (PalmaBeja amp Rodrigues 1999 L Palma unpublished data)
whereas rural pigeons were scarce because of the effectsof marked rural depopulation A less obvious wideningof niche breadth may have occurred in territorieswith intermediate farmland cover where diet diversityincreased along with a high consumption of jaysThis pattern might also be explained by the lowerabundance of preferred wild and domestic prey Indeedpartridges were consumed most frequently in breedingterritories dominated by native oak forest and inter-mediate scrubland cover where they appeared mostabundant (L Palma unpublished data) and less withincreasing cover by farmland and urban areas Incontrast rural pigeons were presumably most abundantin farmland although this was not evident in terms ofdietary responses Rabbits were not responsive to thisgradient probably because land-use influences wereconfounded by climatic gradients affecting rabbitabundances across the region (L Palma P Beja ampM Pais unpublished data)
The dietary responses of eagles to variation in theabundance of the most important prey of wild (rabbit)and domestic (rural pigeon) origin also suggested thatdomestic prey could compensate for the shortage ofwild prey although there was no switching in thetraditional sense (Murdoch 1969) Rabbits were clearlythe primary prey item with their consumption followinga pronounced type II response curve and the highestintake of both rural and racing pigeons occurring at thelowest rabbit densities However eagles also showed atype II functional response to rural pigeons althoughmuch weaker than in the case of rabbits and not the typeIII sigmoid curve that might be expected if there wasswitching (Akre amp Johnson 1979 Patterson Benjaminamp Messier 1998) Furthermore there was a negativerelationship between the consumption of rabbits andthe abundance of rural pigeons as well as evidence thatselection between these two prey was inversely frequencydependent with pigeons consumed more often thanexpected at high rabbit densities and the same occurringfor rabbits at high pigeon densities This may tentativelybe related to the critical dietary role of pigeons earlyin the breeding season when there seems to be lowabundance of most wild prey including rabbits Inthese circumstances many rural pigeons may be takenin areas where the abundance of rabbits will increasefollowing their spring reproductive peak Later in theseason rabbits may be taken more often than expectedas they could be more profitable to Bonellirsquos eagles thanpigeons Testing this hypothesis would require moredetailed information on the seasonal variation in theabundance of wild and domestic prey and their relativeprofitability to the predator
Taken together our results suggest that rural pigeonsshould be regarded as complementary prey items torabbits and partridges rather than an alternative preythat is consumed only where there is a shortage of theseprimary wild prey Conversely racing pigeons wereprobably taken opportunistically because of their highvulnerability Where the most profitable wild and
Fig 6 Frequency-dependent selection curve for Bonellirsquoseagles preying on rural pigeons (1) and rabbits (2) relating theconsumption (e1e2) and availability (A1A2) ratios
1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1084L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
domestic prey were scarce the eagles widened theirniche breadth and consumed a range of relatively minoravian prey This is therefore a complex multispeciessystem that supports the view that predator conflictscannot be addressed as a single pairwise predatorndashprey interaction (Graham Beckerman amp Thirgood2005)
Conflicts with pigeon fanciers are considered a majorthreat for the conservation of some Bonellirsquos eaglepopulations (Real et al 2001 Carrete et al 2002) Thesituation is probably not yet a serious one in south-west Portugal despite eagles generally showing highpredation rates on domestic pigeons Enquiries to ruralpigeon breeders (n = 326) revealed that they generallyattributed a low value to pigeons regarding themsimply as pets and home-grown food (93middot3) withonly 6middot7 of those censused making use of pigeons fortrade racing or recreational shooting (L Palmaunpublished data) Most rural pigeon breeders dis-regarded eagles as a significant threat as they weregenerally unaware of their presence except possiblywhere predation was concentrated at particular loftsIn the case of racing pigeons Bonellirsquos eagles killed avery small proportion of the huge numbers releasedregularly indicating that they do not represent a realthreat to pigeon races Overall there is therefore arelatively benign situation justifying the lack of activepersecution that is apparent elsewhere (Real et al 2001)However given the dietary importance of domesticpigeons for these eagles management options reducingthe potential conflicts with pigeon breeders should notbe disregarded as it is possible that less-tolerant atti-tudes may develop in the near future
Rearing pigeons in specially designed lofts might bean effective means of reducing potential conflicts withpigeon breeders while maintaining an adequate foodsupply for Bonellirsquos eagles Lofts could be establishedby conservation organizations in territories withdepleted food resources and in areas where persecutionmight be a problem Pigeons provided at these lsquocon-servationrsquo lofts might act as diversionary food to reducepredatory pressure upon both game and racing pigeons(Carrete et al 2002) similar to the lofts with low-valuepigeons that have been used elsewhere to deflectpredation by peregrine falcons (Shawyer Clarke ampDixon 2000) In south-west Portugal this managementapproach could help to offset a possible medium-termdecline in traditional pigeon rearing associated withrural abandonment with negative effects on the eaglersquosfood resources It might also help to reduce predationon valuable game species such as partridges given thedevelopment of small game hunting in this regionProviding health-screened pigeons could also reducethe infection of eagles by the protozoan Trichomonasgallinae Rivolta which is a significant cause of nestlingmortality in this and other Bonellirsquos eagle populations
(Houmlfle et al 2000 Real Mantildeosa amp Muntildeoz 2000)These applications of rural pigeons as a conservationtool require testing in south-west Portugal and elsewhereto evaluate their potential benefit to eagle breedingsuccess disease prevalence and conflicts with people
Acknowledgements
We are grateful to Joseacute Raiado Carlos Noivo PedroPortela Rita Alcazar Francisco Faria Paulo Cabritaand Joatildeo Ferreira for field assistance Rita Inaacutecio forcollaboration in dietary analysis Daniel Santos forinformation on pigeon racing and Fernando Gradefor the census of rural pigeons We thank Simon Thir-good and two anonymous referees for critical reviewof the manuscript This study was supported by thePortuguese Institute for Nature Conservation and thePortuguese Science Foundation (PRAXISBIA13296)
References
Akre BG amp Johnson DM (1979) Switching and sigmoidfunctional response curves by damselfly naiads withalternative prey available Journal of Animal Ecology 48703ndash720
Angelstam P Lindstroumlm E amp Wideacuten P (1984) Role ofpredation in short-term population fluctuations of somebirds and mammals in Fennoscandia Oecologia 62 199ndash208
Arroyo B Ferreiro E amp Garza V (1995) El Aacuteguila Perdicera(Hieraaetus fasciatus) en Espantildea Censo Reproduccioacuten yConservacioacuten ICONA Madrid Spain
Avery ML amp Cummings JL (2004) Livestock depredationsby black vultures and golden eagles Sheep and GoatResearch Journal 19 58ndash63
Beja P Palma L amp Pais M (2007) Rabbit habitats inMediterranean scrubland the role of scrub structure andcomposition Wildlife Biology 13(1) March 2007
Burnham KP amp Anderson DP (1998) Model Selection andInference A Practical Information-Theoretic ApproachSpringer-Verlag New York NY
Cardona DB amp Esteban DR (2002) Guiacutea de aves del estrechode Gibraltar Parque Natural lsquoLos Alcornocalesrsquo y Comarcade lsquola Jandarsquo Orni Tour Cadiz Spain
Carrete M Saacutenchez-Zapata JA Martiacutenez J SaacutenchezMA amp Calvo JF (2002) Factors influencing the decline ofa Bonellirsquos eagle Hieraaetus fasciatus population insoutheastern Spain demography habitat or competitionBiodiversity and Conservation 11 975ndash985
Delibes M amp Hiraldo F (1981) The rabbit as prey in theIberian Mediterranean ecosystems Proceedings of theWorld Lagomorph Conference (eds K Myers amp CDMacinnes) pp 614ndash622 University of Guelph Guelph
Doncaster CP amp Woodroffe R (1993) Den site candetermine shape and size of badger territories implicationsfor group living Oikos 66 88ndash93
Fa JE Sharples CM amp Bell DJ (1999) Habitat correlatesof European rabbit (Oryctolagus cuniculus) distributionafter the spread of RVHD in Cadiz Province Spain Journalof Zoology 249 83ndash96
Gil-Saacutenchez JM (1998) Seleccioacuten de presa por el aacuteguila-azorperdicera (Hieraaetus fasciatus) durante el periacuteodo denidificacioacuten en la Provincia de Granada (SE de Espantildea)Ardeola 45 151ndash160
Gil-Saacutenchez JM Moleoacuten M Otero M amp Bautista J(2004) A nine-year study of successful breeding in a
1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1085Eagle predation on domestic pigeons
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Bonellirsquos eagle population in southeast Spain a basis forconservation Biological Conservation 118 685ndash694
Graham K Beckerman AP amp Thirgood S (2005) Humanndashpredatorndashprey conflicts ecological correlates prey lossesand patterns of management Biological Conservation 122159ndash171
Greenwood JJD amp Elton RA (1979) Analysing experimentson frequency-dependant selection by predators Journal ofAnimal Ecology 48 721ndash737
Henderson I Parrott D amp Moore N (2004) RacingPigeons Impact of Raptor Predation Scottish NaturalHeritage Edinburgh UK
Houmlfle U Blanco JM Palma L amp Melo P (2000) Tricho-moniasis in Bonellirsquos Eagle (Hieraaetus fasciatus) nestlingsin south-west Portugal Raptor Biomedicine III (eds JTLumeij JD Remple PT Redig M Lierz amp JE Cooper)pp 45ndash51 Zoological Education Network Lake WorthFL
Iborra O amp Lumaret JP (1997) Validity limits of pellet-group counts in the wild rabbit (Oryctolagus cuniculus)Mammalia 61 205ndash218
Iezekiel S Bakaloudis DE amp Vlachos CG (2004) The dietof Bonellirsquos eagle Hieraaetus fasciatus in Cyprus RaptorsWorldwide (eds RD Chancellor amp B-U Meyburg)pp 581ndash587 WWGBPMME Budapest Hungary
Kenward RE Halls DG Walls SS amp Hodder KH(2001) Factors affecting predation by buzzards Buteo buteoon released pheasants Phasianus colchicus Journal ofApplied Ecology 38 813ndash822
Kenward RE Marcstroumlm V amp Karlbom M (1981)Goshawk winter ecology in Swedish pheasant habitatsJournal of Wildlife Management 45 397ndash408
Landa A Gudvangen K Swenson JE amp Roslashskaft E(1999) Factors associated with wolverine Gulo gulo predationon domestic sheep Journal of Applied Ecology 36 963ndash973
Legendre P amp Legendre L (1998) Numerical Ecology 2ndedn Elsevier Amsterdam the Netherlands
Lindeacuten H amp Wikman M (1983) Goshawk predation ontetraonids availability of prey and diet of the predator in thebreeding season Journal of Animal Ecology 52 953ndash968
Marchesi L Pedrini P amp Sergio F (2002) Biases associatedwith diet study methods in the Eurasian eagle-owl Journalof Raptor Research 36 11ndash16
Marchesi L Sergio F amp Pedrini P (2002) Costs and benefitsof breeding in human-altered landscapes for the eagle owlBubo bubo Ibis 144 164ndash177
Meriggi A amp Lovari S (1996) A review of wolf predation insouthern Europe does the wolf prefer wild prey to livestockJournal of Applied Ecology 33 1561ndash1571
Murdoch WW (1969) Switching in general predatorsexperiments on predator specificity and stability of preypopulations Ecological Monographs 39 335ndash354
Mure M (2003) Approche par suivi visual du domaine vital delrsquoAigle de Bonelli (Hieraaetus fasciatus) en Ardegraveche PhDThesis Eacutecole Pratique des Hautes Eacutetudes Paris France
Ontiveros D amp Pleguezuelos JM (2000) Influence of preydensities in the distribution and breeding success ofBonellirsquos eagle (Hieraaetus fasciatus) managementimplications Biological Conservation 93 19ndash25
Ontiveros D Pleguezuelos JM amp Caro J (2005) Preydensity prey detectability and food habits the case ofBonellirsquos eagle and the conservation measures BiologicalConservation 123 19ndash25
Palma L Beja P amp Rodrigues M (1999) The use of sightingdata to analyse Iberian lynx habitat and distributionJournal of Applied Ecology 36 812ndash824
Palma L Beja P Tavares PC amp Monteiro LR (2005)Spatial variation of mercury levels in nesting Bonellirsquoseagles from southwest Portugal effects of diet compositionand prey contamination Environmental Pollution 134549ndash557
Patterson BR Benjamin LK amp Messier F (1998) Preyswitching and feeding habits of eastern coyotes in relationto snowshoe hare and white-tailed deer densities CanadianJournal of Zoology 76 1885ndash1897
Penteriani V Gallardo M amp Roche P (2002) Landscapestructure and food supply affect eagle owl (Bubo bubo) densityand breeding performance A case of intra-populationalheterogeneity Journal of Zoology 257 365ndash372
Petty SJ Lurz PWW amp Rushton SP (2003) Predation ofred squirrels by northern goshawks in a conifer forest innorthern England can this limit squirrel numbers andcreate a conservation dilemma Biological Conservation111 105ndash114
Real L (1977) The kinetics of functional response AmericanNaturalist 111 289ndash300
Real J (1987) Evolucioacuten cronoloacutegica del reacutegimen alimenticiode una poblacioacuten de Hieraaetus fasciatus en Catalunyafactores causantes adaptacioacuten y efectos SupplementoRicerca Biologica Selvaggina 12 185ndash205
Real J (1991) Lrsquoagraveliga perdiguera Hieraaetus fasciatus a Catalunyastatus ecologia trogravefica biologia reproductora i demografiaPhD Thesis University of Barcelona Barcelona Spain
Real J (1996) Biases in diet study methods in the Bonellirsquoseagle Journal of Wildlife Management 60 632ndash638
Real J amp Mantildeosa S (1997) Demography and conservationof western European Bonellirsquos eagle Hieraaetus fasciatuspopulations Biological Conservation 79 59ndash66
Real J amp Mantildeosa S (1998) Post-nestling dependence periodin the Bonellirsquos eagle Hieraaetus fasciatus Ornis Fennica75 129ndash137
Real J Grande JM Mantildeosa S amp Saacutenchez-Zapata JA(2001) Causes of death in different areas for Bonellirsquos eaglesin Spain Bird Study 48 221ndash228
Real J Mantildeosa S amp Muntildeoz E (2000) Trichomoniasis in aBonellirsquos eagle population in Spain Journal of WildlifeDiseases 36 64ndash70
Redpath SM amp Thirgood SJ (1999) Numerical and func-tional responses in generalist predators hen harriers andperegrines on Scottish grouse moors Journal of AnimalEcology 68 879ndash892
Selarings V (1997) Influence of prey availability on re-establishmentof goshawk Accipiter gentilis nesting territories OrnisFennica 74 113ndash120
Shawyer C Clarke R amp Dixon N (2000) A Study into theRaptor Predation of Domestic Pigeons Department of theEnvironment Transport and Regions London UK
Stahl P Ruette S amp Gros L (2002) Predation on free-rangingpoultry by mammalian and avian predators field lossestimates in a French rural area Mammalian Review 32227ndash234
Stahl P Vandel JM Herrenschmidt V amp Migot P (2001)Predation on livestock by an expanding reintroduced lynxpopulation long-term trends and spatial variabilityJournal of Applied Ecology 38 674ndash687
Stephens DW amp Krebs JR (1986) Foraging Theory PrincetonUniversity Press Princeton NJ
Thirgood SJ Redpath SM Newton I amp Hudson P(2000) Raptors and red grouse conservation conflicts andmanagement solutions Conservation Biology 14 95ndash104
Treves A amp Karanth KU (2003) Humanndashcarnivore conflictand perspectives on carnivore management worldwideConservation Biology 17 1491ndash1499
Valkama J Korpimaumlki E Arroyo B Beja P Bretagnolle VBro E Kenward R Mantildeosa S Redpath SM ThirgoodS amp Vintildeuela J (2005) Birds of prey as limiting factors ofgamebird populations in Europe a review BiologicalReviews 80 171ndash203
Villafuerte R Vintildeuela J amp Blanco JC (1998) Extensivepredator persecution caused by population crash in a gamespecies the case of red kites and rabbits in Spain BiologicalConservation 84 181ndash188
1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis

1086L Palma et al
copy 2006 The Authors Journal compilation copy 2006 British Ecological Society Journal of Applied Ecology 43 1075ndash1086
Woodroffe R Lindsey P Romantildeach S Stein A amp oleRanah SMK (2005) Livestock predation by endangeredAfrican wild dogs (Lycaon pictus) in northern KenyaBiological Conservation 124 225ndash234
Zar JH (1996) Biostatistical Analysis 3rd edn Prentice HallInternational London UK
Received 17 November 2005 final copy received 24 May 2006Editor Simon Thirgood
Supplementary materialThe following supplementary material is availableas part of the online article (full text) from httpwwwblackwell-synergycom
Table S1 Detailed results of Bonellirsquos eagle dietanalysis




















