
Paludicola vol 7, issue 4 May 2010
70
PALUDICOLA SCIENTIFIC CONTRIBUTIONS of the ROCHESTER INSTITUTE OF VERTEBRATE PALEONTOLOGY VOLUME 7 NUMBER 4 4 MAY 2010 ISSN 1091-0263
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Paludicola vol 7, issue 4 May 2010

PALUDICOLA
SCIENTIFIC CONTRIBUTIONS of the
ROCHESTER INSTITUTE OF VERTEBRATE PALEONTOLOGY
VOLUME 7 NUMBER 4 4 MAY 2010
ISSN 1091-0263

PALUDICOLA SCIENTIFIC CONTRIBUTIONS OF THE ROCHESTER INSTITUTE OF VERTEBRATE
PALEONTOLOGY
Editors William W. Korth Rochester Institute of Vertebrate Paleontology 265 Carling Road Rochester, NY 14610 Phone (585) 482-0203 e-mail: [email protected] Judy A. Massare Department of Earth Sciences State University of New York, College at Brockport Brockport, NY 14420 Phone (585) 395-2419: Fax (585) 395-2416 e-mail: [email protected]
EDITORIAL POLICY Paludicola is published under the auspices of the Rochester Institute of Vertebrate Paleontology (RIVP). It is intended to serve as an “in-house” publication for students and professional paleontologists who are not affiliated with institutions that provide such a publication. All manuscripts considered for publication in Paludicola are expected to be original and have not been published elsewhere in any other form or are not being considered for publication anywhere else. This journal is intended as an outlet for papers on any aspect of Vertebrate Paleontology. Paludicola is published twice annually (Fall and Spring). All submitted manuscripts are subject to review by external qualified specialists and the editorial staff of Paludicola. The format of all submitted manuscripts should follow that of the Journal of Vertebrate Paleontology (printed in the first issue of each year). Questions about other editorial policies will be addressed by the editorial staff upon request. Please contact the editorial staff for prices of subscriptions and back issues (prices are subject to change). Back issues of volumes 1 – 6 can be purchased from PaleoPublications Inc. Prices and listings are available on-line at www.paleopubs.com. Copyright 2010 by the Rochester Institute of Vertebrate Paleontology

Paludicola 7(4):117-136 May 2010 © by the Rochester Institute of Vertebrate Paleontology
117
MAMMALS FROM THE BLUE ASH LOCAL FAUNA (LATE OLIGOCENE), SOUTH DAKOTA. RODENTIA PART 5: FAMILY CRICETIDAE
William W. Korth
Rochester Institute of Vertebrate Paleontology, 265 Carling Road Rochester, NY 14610 <[email protected]>
ABSTRACT Ten species of cricetid rodents are identified from the Blue Ash fauna including three new species, Eumys brachyodus, E. parvidens, Scottimus sp., cf. S. longiquus, S. kellamorum, Leidymys sp., cf. L. blacki, L. juxtaparvulus n. sp., Paciculus dakotensis n. sp., P. sp., cf. P. nebraskensis, Geringia copiosus n. sp., and an indeterminate species. The cricetids are the best represented and one of the most diverse families of rodents from the Blue Ash anthill fauna. Some of the genera of cricetids present in the fauna are more characteristic of the Arikareean land mammal age (Leidymys, Paciculus, Geringia), but the species are either smaller or morphologically more primitive than those from the Arikareean. However, three species in the fauna (E. brachyodus, E. parvidens, S. longiquus) are currently only known from the Orellan or Whitneyan. It appears that the age of the Blue Ash fauna, based on the cricetid rodents, is somewhere near the Whitneyan-Arikareean boundary.
INTRODUCTION Previously, a number of taxa from the Blue Ash fauna have been identified that are elsewhere known from the Arikareean, as well as other taxa that are more typical of earlier Orellan and Whitneyan ages (see Korth, 2009 and references therein). The cricetids are the most abundant family of rodents in the Blue Ash fauna, being represented by over 500 specimens. Distinct cricetid characterize the land mammal ages Orellan, Whitneyan, and Arikareean. The suites of genera represented from these adjacent levels are therefore diagnostic of their respective land mammal ages. The earlier horizons (Orellan and Whitneyan) are dominated by species of Eumys, Scottimus, Wilsoneumys, and Eoeumys; the Arikareean fauna is dominated by species of Leidymys, Geringia, and Paciculus with rare occurrences of Eumys (Wood, 1937, 1980; Martin, 1980; Korth, 1994). The distinctiveness of the Whitneyan and Arikareean cricetid faunas, and the great number of specimens, suggests that the cricetids should be quite valuable in determining the biostratigraphic horizon of the fauna. Dental terminology modified from that of Wood and Wilson (1936). Capital letters indicate upper teeth and lower-case letters represent lower teeth (e.g. M1 or m1). Abbreviations for institutions: AMNH, American Museum of Natural History; FAM, Frick Collections, American Museum of Natural History; CM, Carnegie Museum of Natural History; MCZ, Museum of Comparative Zoology, Harvard University; ROM,
Royal Ontario Museum; UNSM, University of Nebraska State Museum; YPM-PU, Yale Peabody Museum, Princeton Collection.
SYSTEMATIC PALEONTOLOGY
Order Rodentia Bowdich, 1821 Family Cricetidae Fischer de Waldheim, 1817
Eumys Leidy, 1856 Type Species—Eumys elegans Leidy, 1856 (=Eumys obliquidens Wood, 1937; =Cricetodon nebraskensis Wood, 1937; =Eumys pristinus Russell, 1972). Referred Species—Eumys brachyodus Wood, 1937; Eumys parvidens Wood, 1937; Eumys cricetodontoides White, 1954 (=E. latidens White, 1954; =E. spokanensis White, 1954; =E. elegans [in part] Martin, 1980); Eumys eliensis Black, 1961a (=E. brachyodus [in part] Martin, 1980; =Leidymys eliensis Lindsay, 2008). Range—Orellan to early Arikareean (early to late Oligocene) western North America. Discussion—Thirteen species of Eumys have been named from North America. Of these, five have been referred to other genera (Table 1). In his review of Oligocene and early Miocene cricetids, Martin (1980) recognized only four species, E. elegans, E. brachyodus, E. parvidens and E. pristinus. He synonymized E. obliquidens, E. cricetodontoides, E. latidens, E. spokanensis and Cricetodon nebraskensis

PALUDICOLA, VOL. 7, NO. 4, 2010 118
under E. elegans. Martin (1980) also included the only Arikareean species, E. eliensis, under E. brachyodus. Most recently, Lindsay (2008) did not recognize any of Martin’s synonymies and transferred E. eliensis to Leidymys. Probable synonymies of the species of Eumys are discussed below. __________________________________________ TABLE 1. Previously named species of Eumys from North America referred to other genera. SPECIES REFERRAL E. exiguus Wood, 1937 Scottimus exiguus (Black, 1961b) E. planidens Wilson, 1949 Wilsoneumys planidens
(Martin, 1980) E. woodi Macdonald, 1963 Paciculus woodi (Martin, 1980) E. blacki Macdonald, 1963 Leidymys blacki (Martin, 1980) E. gloveri Macdonald, 1970 Geringia gloveri (Martin, 1980) __________________________________________________
Galbreath (1953) considered Cricetodon nebraskensis as just a morphological variation of Eumys elegans. Wood (1980) agreed that it was referable to Eumys, but included it as a synonym of E. cricetodontoides. Martin (1980) followed Galbreath (1953) and included Cricetodon nebraskesnsis as a synonym of Eumys elegans. Lindsay (2008:465) recognized it as a distinct species of Eumys, E. nebraskensis stating that …”it is now considered a species of Eumys by most authorities (Wood, 1980).” However, Wood (1980) considered C. nebraskensis as a synonym of another species (E. cricetodontoides) and no references could be found to verify its recognition as a distinct species. Galbreath’s (1953) arguments are followed here and the size of specimen referred to C. nebraskensis (Wood, 1937:257) is within the range of samples of E. elegans from Nebraska (see Appendix). Therefore, it is considered here as a junior synonym of E. elegans. Russell (1972) named Eumys pristinus from the Chadronian of Saskatchewan. In his description, he misidentified two specimens: ROM 6323 was identified as a dp4 of an eomyid, but is a right m1 of Eumys; and ROM 6326, was identified as a left M3 of E. pristinus but is a lower molar of Ischyromys (Russell, 1972:fig. 10I and fig. 11A, respectively). The characters used to diagnose E. pristinus (size of the anteroconid on M1, orientation of the posterior arm of protoconid on lower molars, and relative size of the anterior cingulum on the lingual side of the lower molars) are clearly variable within populations of E. elegans from the Orellan. The size of the specimens is also well within the size range of E. elegans (Russell, 1972:41-42; Appendix, this paper). More recently, it has been demonstrated that the fauna described by Russell contained Chadronian species but also included specimens mixed from later horizons (Storer and Bryant, 1993; Storer, 1996). The most recently
published faunal list from the Chadronian of Saskatchewan by Storer (1996:252) does not include Eumys, but it does occur in later Orellan faunas of the same area. It is proposed here, that E. pristinus is a junior synonym of E. elegans and the earliest record of the genus remains the Orellan. White (1954) named three species of Eumys from the Orellan of Montana, E. cricetodontoides, E. latidens, and E. spokanensis. Martin (1980) included all of them under E. elegans, noting the morphological differences cited by White (1954) were minor variations that could be found in samples of E. elegans, and that the size of the specimens overlapped that of E. elegans as well, but provided no tables for comparison. Wood (1980) similarly noted that the morphological differences among the Montana species were not significant and retained E. cricetodontoides as the senior synonym of the other two Montana species. The greatest difference between E. cricetodontoides and E. elegans is the size of the cheek teeth. The dimensions of the lower molars are much larger in the Montana species than in specimens of E. elegans (Wood, 1937:521; Galbreath, 1953:table 14; White, 1954:412-414), but there is some overlap in size ranges. A large sample of Eumys elegans from the Orellan of Sioux County, Nebraska was also measured for comparison (see Appendix). The Student’s t-test and Mann-Whitney non-parametric test were applied to the two populations (Nebraska and Montana) and were shown to be satistically significantly different (p <0.001) for each of the dimensions of each tooth and the total length of the tooth row. This demonstrates that the Montana specimens definitely constituted a population distinct from the E. elegans sample. Therefore, Eumys cricetodontoides is recognized here as a distinct species based on its larger size. Black (1961a) named Eumys eliensis, from the Arikareean of Montana. He diagnosed the new species as: 1) having teeth proportionally larger than other species relative to the depth of the mandible; 2) teeth increase in size from m1 to m3; 3) posterior arm of the protoconid joining the metaconid on its lingual side on the molars; 4) no lingual cingulum anterior to the metaconid on m2 and m3; and 5) mental foramen near the inferior border of the mandible below anterior root of m1. Martin (1980) synonymized E. eliensis with the otherwise Whitneyan E. brachyodus because the diagnostic characters of the former all occurred in specimens of the latter. However, Martin (1980) did not provide any figures, tables of measurements, or cite any specimens of E. brachyodus that had these characters of E. eliensis. He also mistakenly listed the presence of a lingual anterior cingulum as one of Black’s (1961a) characters instead of the absence of the cingulum. The character of the relative size of the mandible and position of the mental foramen cannot be

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 119
verified in the Blue Ash sample. However the morphologies and relative sizes of the molars can be determined. The morphology of the posterior arm of the protoconid and the presence or absence of the lingual anterior cingulum does appear to be variable in the Blue Ash sample and other reported specimens of E. brachyodus (Wood, 1937; Galbreath, 1953). However, in the sample from Blue Ash and other specimens of E. brachyodus, it is evident that m1 is the largest of the lower molars. The mean length of m1 from Blue Ash is the same as the maximum length of any of the m3s (Table 2). Wood (1937:253) identified two referred specimens of E. brachyodus that had m3s longer than m1, but the difference was 3-6% the total length of m3, much less than in the type of E. eliensis (10%). The length of m2 and m3 of the holotype of E. eliensis, YPM 14022 (2.6 mm; 2.75 mm, respectively), is far larger than the length of any previously reported specimen of E. brachyodus (Wood, 1937:253; Galbreath, 1953:table 14; Table 2 and Appendix, this paper). It is evident, based on size and proportions of the lower molars that E. eliensis is distinct from E. brachyodus and should be considered a separate species. Lindsay (2008) referred E. eliensis to the genus Leidymys, because of the greater lophodonty of Leidymys compared to Eumys. However, the degree lophodonty and crown height of the molars in Eumys are greater than in species of Leidymys. E. eliensis also lacks the diagnostic doubled mesolophid on m2 and m3 of Leidymys. Therefore, E. eliensis is retained in Eumys. As a result of these synonymies, five species of Eumys are recognized: E. elegans, E. parvidens, and E. cretodontoides from the Orellan; E. brachyodus from the Whitneyan; and E. eliensis from the Arikareean.
Eumys brachyodus Wood, 1937 (Figure 1A-F; Table 2)
Type Specimen—MCZ 5069, mandible with right m1-m3. Referred Specimens—CM 73530, partial maxilla with M1-M2; CM 73560, 73576, partial maxillae with M2-M3; CM 73532, 73533, 73536, 73538, 73542, 73553, 73554, 73562, 73564, 73581-73583, 73596-73598, 76110, 83735-83740 – M1; CM 73535, 73537, 73541, 73547, 73549, 73551, 73555, 73556, 73565, 73568-73570, 73578, 73585, 73588, 73594, 73595, 76102, 76103, 76113, 76116, 76119 – M2; CM 73544, 73557-73559, 73580, 73584, 73590, 73599, 76100, 83741-83745 – M3; CM 73531, 73534, 73539, 73540, 73543, 73548, 73563, 76108, 76109, 76111, 76112, 76120-76122, 76125-76128, 76130 – m1; CM 73529, 73561, 73571, 73572, 73574, 73577, 73586, 73589, 73591, 73592, 76101, 76104-76106, 76114, 76115, 76124, 76131, 83800 – m2; CM 73545,
73546, 73550, 73552, 73566, 73567, 73573, 73575, 73579, 73587, 76117, 76118 – m3. Amended Diagnosis—Similar in size to Eumys elegans but molars wider relative to length than other species; anteroconid on m1 minute; double connection of the metalophid with the anteroconid variably present on m1 (never present in other species); lingual half of the anterior cingulum greatly reduced on m2-m3; mesolophs (-ids) minute to absent on all molars; anterocone on M1 smaller than in other species; parastyle variably present on M1 (absent in all other species); M3 slightly longer (anteroposteriorly) relative to width than in other species. Description—M1 has surprisingly little variation in the sample. The anterocone is large, but relatively smaller than in specimens of E. elegans. In a sample of 15 specimens of E. elegans (Wahlert, 2004:2), the width of the anterocone averaged 66% of the maximum width of M1. In the Blue Ash sample, the anterocone averaged 63% the maximum width of M1, slightly less. The anterocone is D-shaped in outline, the flat surface forms the posterior wall. On 70% of the Blue Ash specimens a distinct parastyle is present at the posterobuccal corner of the anterocone. In most specimens it is a small swelling, separated from the anterocone by a small, shallow valley. In a few specimens it is a distinct cusp. The four major cusps of the tooth are nearly equal in size and height. The anterior width of the tooth (protoloph) is just slightly greater than the posterior width (metaloph). The buccal cusps, paracone and metacone, are oval and anteroposteriorly compressed. The lingual cusps, protocone and hypocone, are obliquely compressed. The anterior arm of the protocone extends anteriorly to join the center of the posterior wall of the anterocone. Very short accessory lophules extend posteriorly from the anterocone on seven specimens (32%). They do not extend the entire length of the valley separating the anterocone from the protoloph. They can occur either buccal or lingual to the connection of the anterior arm of the protocone with the anterocone. The protoloph extends lingually from the paracone, attaching to the posterior arm of the protocone. Here again, on five specimens (23%) there are short lophules that run into the valley between the protocone and paracone. The endoloph runs posteriorly from the point where the protoloph connects to the posterior arm of the protocone, then bends lingually to join the hypocone. At the bend in the endoloph there is usually a short mesoloph. Only about 20% of the specimens lack a mesoloph, but it can be very small (tiny spur) in some specimens that have it. The metaloph is parallel to the protoloph and runs lingually from the metacone, joining the buccal end of the anterior arm of the hypocone. The posterior cingulum runs from the apex of the hypocone along the posterior margin of the

PALUDICOLA, VOL. 7, NO. 4, 2010 120
FIGURE 1. Molars of Eumys from the Blue Ash local fauna. A-F, Eumys brachyodus. A, CM 73533, left M1. B, CM 73547, left M2. C, CM 83743, left M3. D, CM76112, left m1. E, CM 76105, left m2. F, CM 73575, left m3. G-H, Eumys sp., cf. E. parvidens. G, CM 76123, left m1. H, CM 84645, right M3. Bar scale = 1 mm. ________________________________________________________________________________________________________________________
tooth, ending posterior and lingual to the metacone. The valley between the posterior cingulum and the metaloph and metacone is very deep. M2 has the least amount of variation of any of the cheek teeth. It is very similar in morphology to that of M1 without the anterocone. M2 is nearly equal in length and width and the anterior and posterior widths are almost exactly the same. The lingual cusps are more dramatically obliquely compressed than on M1.
The anterior arm of the protocone extends along the anterior edge of the tooth to the buccal border, forming the anterior cingulum. The protoloph runs from the paracone, always joining the endoloph posterior to the protocone. A mesoloph is present on all specimens, but does vary in length and never extends more than half the way to the buccal edge of the tooth. The configuration of the hypocone, metacone, and posterior cingulum of M2 is identical to that of M1.

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 121
TABLE 2. Dental measurements of Eumys brachyodus from the Blue Ash fauna. Abbreviations: L, length; W, width; N, number of specimens; M, mean; Min, minimum measurement; Max, maximum measurement; SD, standard deviation; CV, coefficient of variation. Measurements in mm.
M1L M1W M2L M2W M3L M3W L/W M1 L/W M2 L/W M3
N 15 23 24 24 15 16 14 22 13
M 2.91 1.98 2.05 1.90 1.69 1.83 1.45 1.08 0.94
Min 2.64 1.77 1.91 1.71 1.45 1.65 1.34 0.97 0.85
Max 3.18 2.30 2.28 2.25 1.92 1.95 1.64 1.20 1.04
SD 0.15 0.15 0.09 0.13 0.13 0.09 0.09 0.07 0.04
CV 5.09 7.54 4.20 6.59 7.49 4.72 6.07 6.18 4.47
m1L m1W m2L m2W m3L m3W L/W m1 L/W m2 L/W m3
N 21 21 18 18 11 12 21 18 11
M 2.34 1.78 2.21 1.97 2.24 1.97 1.31 1.12 1.14
Min 2.12 1.59 2.07 1.84 2.13 1.84 1.20 1.04 1.09
Max 2.53 1.98 2.35 2.09 2.33 2.13 1.41 1.25 1.24
SD 0.11 0.11 0.09 0.08 0.08 0.10 0.06 0.05 0.06
CV 4.49 5.99 3.93 3.89 3.57 4.85 4.83 4.87 4.85
_______________________________________________________________________________________________ M3 is the smallest of the upper molars. It is much narrower posteriorly than anteriorly. The length of M3 is the most variable measurement of all of the molars of the sample (Table 2). The protocone is an obliquely oriented swelling at the anterolingual corner of the tooth, continuous with the anterior cingulum that runs the entire width of the tooth. The paracone is greatly anteroposteriorly compressed, forming a transverse ridge that joins the posterior arm of the protocone. The endoloph runs posteriorly from the connection of the protoloph and the posterior arm of the protocone, curves lingually and ends posteriorly in a small hypocone. The metacone, similar to the paracone, is greatly anteroposteriorly compressed. The metaloph runs lingually from it, meeting the endoloph anterior to the hypocone. Only three of the 15 specimens have a small mesoloph. The size and position of the hypocone (more lingually or buccally placed) are variable. A posterior cingulum is usually present, but is variable in length depending on the position of the hypocone. Specimens with more lingually placed hypocones usually have a longer posterior cingulum, while others that are more buccally placed have no posterior cingulum at all. The first lower molar is longer than wide, and wider posteriorly than anteriorly. The anteroconid on all specimens is at most a small swelling at the center of the anterior cingulum, not a distinct cusp as in E. elegans. The anterior cingulum extends both buccally and lingually from the center of the anterior margin of the tooth about equal distances, and does not fuse posteriorly with the bases of the protoconid or metaconid until extremely late stages of wear. The
protoconid and metaconid are subequal in size, with the metaconid positioned slightly more anteriorly than the protoconid. The protoconid is oval in occlusal outline and slightly crescentic. The connection of the metaconid and protoconid to the anterior cingulum (or anteroconid) is variable. On 50% of the specimens, the anterior arm of the protoconid runs directly anteriorly to meet the anterior cingulum at its center. There is an anterior arm of the metaconid that extends anteriorly to meet the anterior cingulum as well, forming a double connection, on 38% of the specimens. The other specimens have differing morphologies. On two specimens, the anterior arm of the metaconid joins the anterior arm of the protoconid, forming a single loph that ultimately joins the anterior cingulum. One specimen, CM 73539, has no connection between the protoconid and anterior cingulum, but there is a connection with the anterior arm of the metaconid instead. The posterior arm of the protoconid extends lingually, joining the posterolingual corner of the metaconid on all but four of the m1s. On three of these specimens, the posterior arm of the protoconid ends just short of meeting the metaconid. On CM 76107, the posterior arm of the protoconid is a small stub extending lingually from the ectolophid. The ectolophid runs between the protoconid and hypoconid, along the center-line of the tooth. The ectolophid does not connect on CM 76130. On 63% of the specimens, a very short mesolophid is at the center of the ectolophid. It is never long. The remainder of the m1s completely lack a mesolophid. On CM 73540, a metastylid is present along the lingual margin of the tooth between the lingual cusps. It is transversely

PALUDICOLA, VOL. 7, NO. 4, 2010 122
elongated, but does not reach the ectolophid or mesolophid. On the buccal side of the ectolophid a short buccal mesolophid that is variable in length, is always present. In a little more than half of the m1s it is just a small stub. In the remainder it is slightly longer, but never reaches the buccal edge of the tooth. The posterior half of m1 has little variation. The entoconid and hypoconid are subequal in size, and the entoconid is just slightly more anterior than the hypoconid. As in the anterior cusps, the entoconid is oval in outline, whereas the hypoconid is slightly crescentic. The hypolophid runs buccally from the entoconid and joins the hypoconid at its anterolingual corner. The posterior cingulum runs lingually from the posterior margin of the hypoconid along the posterior margin of the tooth, ending just before meeting the posterior edge of the entoconid. In very late stages of wear, the lingual end of the posterior cingulum fuses with the entoconid forming an enclosed basin. The second lower molar is nearly equidimensional, being barely longer than wide. The shape of the four major cusps and ectolophid is the same as in m1: the protoconid and metaconid are spaced slightly farther apart, making the anterior width of the tooth equal to the posterior width. The anterior arm of the protoconid extends anteriorly to the anterior margin of the tooth as in m1. A buccal cingulum extends from the point where the anterior arm of the protoconid meets the anterior margin of the tooth, and reaches the buccal margin of the tooth, turning slightly posteriorly at the corner of the tooth. A lingual cingulum, extending from the same point along the anterior edge of the tooth is present on 72% of the specimens. When present, it is very weakly developed anterior to the metaconid and never reaches the lingual edge of the tooth. On the remainder of the m2s, there is no lingual extension of the anterior cingulum. The posterior arm of the protoconid extends lingually as in m1, but ends short of joining the metaconid on 68% of the specimens. On the rest of the specimens it joins the posterior side of the metaconid as in m1. The posterior arm of the protoconid joins the metaconid on its buccal side in one specimen (CM 76115) but a short lophule extends lingually from that point into the valley between the lingual cusps. A mesolophid is only pesent on a single specimen (CM 76511) as a short spur. The buccal mesolophid is present on all but one specimen and is of variable length, but never extending more than half the distance from the ectolophid to the buccal edge of the tooth. The configurations of the entoconid, hypoconid, hypolophid and posterior cingulum of m2 are the same as in m1. The m3 is much narrower posteriorly than anteriorly. It is usually longer than wide, but the length of m3 is more variable than the width, so on some specimens the width is nearly equal to the length. Both
the protoconid and metaconid are similar to those ofm1 and m2, but are anteroposteriorly compressed. The anterolingual corner of the protoconid meets the anterior margin of the tooth, rather than forming a distinct arm as in the anterior molars. The anterior cingulum varies in length, but is only anterior to the protoconid and never extends lingually. The posterior arm of the protoconid meets the metaconid on half of the specimens, and falls just short of the metaconid on the rest. A small swelling (metastylid) is at the lingual end of the posterior arm of the protoconid only if the arm reaches the edge of the tooth without meeting the metaconid. On one specimen, CM 73573, the posterior arm of the protoconid extends more posteriorly, meeting with the entoconid lingually. The ectolophid is oriented obliquely (posterolingually) rather than directly anteroposterior as in the other molars, joining the hypolophid just buccal to the entoconid, at the same point as the anterior arm of the hypoconid. None of the specimens have a mesolophid. On 10 of the 12 specimens of m3, a buccal mesolophid is present as just a small spur. On one of these specimens, CM 73579, has a second spur. The entoconid is anteroposteriorly compressed and smaller than the anterior cusps. The hypoconid forms the center of the posterior margin of the tooth and is reduced to a swelling along the posterior cingulum. Discussion—Wood (1937) diagnosed Eumys brachyodus as having molars that were broader than in other species, a greatly reduced anteroconid on m1, a double connection of the metalophid to the anteroconid on m1, small or absent mesolophids, weak lingual anterior cingulum on m2-m3, and more robust mandible and lower incisor. Martin (1980) generally followed this diagnosis, but eliminated the character of the double connection of the anteroconid because it was variable in occurrence. The specimens from Blue Ash are referable to E. brachyodus because the lower molars are proportionally wider than in other species, although not quite as proportionally wide as those of previously reported of E. brachyodus (Table 2, Appendix). The double connection of the anteroconid occurs in about 1/3 of the m1s from Blue Ash. While this character does not occur in all specimens of E. brachyodus, it does not occur in any other species. All of the other diagnostic characters of the lower molars for E. brachyodus are also present in the Blue Ash sample. The only upper molars of E. brachyodus that were reported previously consisted of a heavily-worn M1-M3 (Martin, 1980:fig. 3) for which no measurements were provided and a specimen with anomalous, heavily-worn upper teeth of uncertain identification (Korth, 1981a:fig. 2). Setoguchi (1978) referred several isolated upper molars to E. brachyodus from what he believed to be the Whitneyan of Wyoming.

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 123
However, the fauna is Orellan in age and the cited specimens are referable to Wilsoneumys planidens (Korth, 1980, 1981b, 1989). Martin (1980) added the presence of a smaller anterocone on M1 to his revised diagnosis of E. brachyodus based on his cited specimen. The upper molars from Blue Ash referred to E. brachyodus have M1s with slightly smaller anterocones than have been reported for E. elegans (Wahlert, 2004) and are relatively wider than those of other species, but only slightly so (Table2). However, the M3s from Blue Ash are relatively more elongated than those of other species. The only other feature, other than proportions, that distinguishes the Blue Ash sample from other species of Eumys is the presence of a distinct parastyle on M1. This occurs in 70% of the Blue Ash specimens, thus it is not always present, but this feature has not been reported for any other species of the genus.
Eumys parvidens Wood, 1937 (Figure 1G, H)
Type Specimen—UNSM 10036, partial skull with associated mandibles; complete dentition. Referred Specimens—CM 76123, left m1; CM 76129, right m1; and CM 84645, right M3. Measurements—CM 76123 (m1): L = 2.21 mm; W = 1.66 mm. CM 76129 (m1): L = 2.19 mm; W = 1.77 mm. CM 84645 (M3): L = 1.50 mm; W = 1.58 mm. Discussion—Morphologically, these three Blue Ash molars do not differ from those previously described for Eumys parvidens (Wood, 1937; Martin, 1980). However, the specimens are slightly larger than previously reported for E. parvidens (Wood, 1937:525) and clearly smaller than specimens of E. elegans (see Appendix). This is the latest record of E. parvidens, which is elsewhere limited to the Orellan (Wood, 1937, 1980; Martin, 1980; Korth, 1994).
Scottimus Wood, 1937 Scottimus kellamorum Black, 1961b
(Figure 2A-F; Table 3) Scottimus sp. Korth, 1981a Type Specimen—MCZ 7342, partial maxilla with left M1-M2. Referred Specimens—CM 83747-83755, 83757-83759, 83765, 83766, 83768-83771, 83882, 84655 – M1; CM 83772, 83773, 83775-83781, 83783-83786, 83822, 84656 – M2; CM 83788, 83790, 84654 – M3; CM 83791-83796, 83799, 83802, 83834, 83835 – m1; CM 83805, 83806, 83808, 83810-83815, 83817-83819, 83821, 83823, 83825-83829, 84657 – m2; CM 83830, 83831, 83833, 83837, 83838 – m3.
Emended Diagnosis—Molars similar in length to those of S. exiguus, but much narrower relative to length than any other species; molars more lophate than S. ambiguus and S. viduus; tranverse lophs (protoloph and metaloph) weak on upper molars; posterior arm of the protoconid usually joins posteriorly running loph from the metaconid on m1. Description—M1 and M2 of the holotype has been fully described by Black (1961b). The following descriptions of M1 and M2 are based on variations observed on the referred Blue Ash specimens. On the anterocone of M1, there is a small basin at the posterobuccal corner of the tooth as in S. exiguus (Wood, 1937:fig. 63; Korth, 1981a:fig. 9B). In approximately 25% of the Blue Ash M1s, there is an accessory spur extending into the valley between the anterocone and the protoloph. On two specimens there are two accessory lophules. These lophules occur randomly and can originate in the anterocone and extend posteriorly, or originate from the paracone or anterior arm of the protocone and extend anteriorly, buccally or lingually. The connection of the paracone to the protocone (protoloph) runs from the posterolingual corner of the paracone to the posterior arm of the protocone, as in the holotype. On two specimens, there is an additional connection from the paracone to the anterior arm of the protocone. On CM 83882 there is no connection between the paracone and protocone. All of the specimens of M1 have a mesoloph present, but on about 20% of the specimens the mesoloph does not extend far enough buccally to fuse with the ectoloph. Black (1961b) noted that the metaloph connected to the posterobuccal corner of hypocone. In the Blue Ash sample, the attachment of the metacone varies from the position in the holotype, to the center of the hypocone (one specimen), or directly to the posterior cingulum (72% of the specimens). On M2, the mesoloph is present on over 80% of the specimens, and on 25% the mesoloph reaches the ectoloph, whereas on the remainder it ends lingual to it. The metaloph of M2 on the holotype is weak and connects from the metacone to the center of the buccal side of the hypocone. This morphology occurs on 30% of the Blue Ash specimens. On the remainder of the specimens the metacone connects to the posterior cingulum as in M1. On one specimen, CM 83784, there is a weak connection between the paracone and protocone (protoloph) similar to the protoloph of M1. As with the anterior molars, M3 is proportionally longer relative to width than in other species of Scottimus (Table 3). The occlusal morphology of M3 is very similar to that of S. exiguus (Wood, 1937:fig. 63; Korth, 1981a:fig. 9B). It differs from S. exiguus in being greatly expanded posterolingually, and the hypocone is reduced to a small cuspule.

PALUDICOLA, VOL. 7, NO. 4, 2010 124
The lower molars referred to S. kellamorum are proportionally longer than wide, as in the upper molars (Table 3). Although there are no mandibles with complete lower dentitions, based on the measurements
FIGURE 2. Molars of Scottimus from the Blue Ash local fauna. A-F, S. kellamorum. A, CM 83788, right M3. B, CM 83775, right M2. C, CM 83749, right M1. D, CM 83837, right m3. E, CM 83806, right m2. F, CM 83795, right m1. G-K, S. sp., cf. S. longiquus. G, CM83761, left M1-M2. H, CM 83789, left M3. I, CM 83798, right m1. J, CM 83316, right m2. K, CM 83836, right m3. Bar scale = 1 mm.
___________________________________________ of the isolated referred specimens, the molars decrease in length from m1 to m3. The m1 is the largest lower molar and is approximately 15% longer than m2, and m3 is slightly shorter than m2. The first molar tapers anteriorly. The anteroconid is central, conical, and nearly has large as the other major cusps of the tooth. The anteroconid is separated from the metaconid and protoconid by a deep valley but is always connected to a loph that is continuous with the anterobuccal slope of the metaconid. On half of the m1s in the sample, there are one or two short spurs that extend into the valley between the anteroconid and protoconid, usually originating from the anteroconid, but also from the protoconid or even the loph connecting the metaconid to the anteroconid. The major cusps are obliquely compressed except the entoconid. The posterior arm of the protoconid extends lingually and is usually joined by a short loph extending posteriorly from the apex of the metaconid. On some specimens the loph continues
more lingually from the junction. On only two specimens, CM 83792, CM 83802, there is no posterior loph from the metaconid and the posterior arm of the protoconid ends in the valley between the metaconid and entoconid short of the lingual edge of the tooth. A distinct loph runs posteriorly along the lingual side of the tooth from the apex of the metaconid, ending in the valley between the metaconid and entoconid. The anterior arm of the hypoconid extands lingually, then turns anteriorly, forming the ectolophid. It joins the protoconid at its posterolingual corner. On half of the specimens, a mesolophid is present but is only a short stub extending lingually from the ectoloph. Rarely, there is a similar spur extending buccally from the ectolophid between the protoconid and hypoconid. The entoconid is slightly anteroposteriorly compressed and the anterobuccal slope of the cusp joins the lingual end of the anterior arm of the hypoconid. The posterior cingulum is high, wrapping around the posterior end of the tooth and enclosing a deep basin between the entoconid and hypoconid. There is a very short buccal extension of the posterior cingulum running from the junction of the posterolingual arm of the hypoconid and the lingual posterior cingulum. There is always a short loph (spur) extending lingually into the basin from the posterolingual corner of the hypoconid. The m2 is rectangular in occlusal shape, being longer than wide. The anterior cingulum extends the entire width of the tooth and is joined separately by anterior extensions of both the protoconid and metaconid, forming a J-shaped basin between the cusps. The posterior arm of the protoconid is the most variable character of m2. It extends posterolingually and usually joins the metaconid posterior to its apex, however, can extend to the lingual edge of the tooth and not join the metaconid at all. The edge of the tooth on the buccal side between the protoconid and hypoconid is often high and forms an anteroposterior loph. The posteror ridge of the metaconid is the same as in m1, and usually joins the lingual end of the posterior arm of the protoconid. The compression of the major cusps and their connections are the same as in m1. The only difference in the posterior part of m2 compared to m1 is the presence of a buccal spur from the ectolophid on about half the specimens, and the lack of the spur from the hypoconid into the posterior basin of the tooth on most of the specimens. The buccal extension of the posterior cingulum is more prominent on m2 than on m1. The anterior half of m3 is very similar to that of m2. The tooth tapers posteriorly. The posterior arm of the protoconid extends to the lingual edge of the tooth. The hypoconid is much smaller than in m2 and the entoconid is only a small cuspule at the lingual end of the loph extending lingually from the anterolingual corner of the hypoconid. There is no buccal extension

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 125
TABLE 3. Dental measurements of Scottimus from the Blue Ash fauna. Abbreviations as in Table 2. Measurements in mm. Measurements in mm. Scottimus kellamorum
M1L M1W M2L M2W M3L M3W L/W M1 L/W M2 L/W M3
N 14 18 14 13 2 2 13 13 2
M 2.59 1.66 1.82 1.46 1.47 1.23 1.55 1.24 1.20
Min 2.35 1.52 1.68 1.28 . 145 1.19 1.46 1.11 1.22
Max 2.81 1.78 1.95 1.60 1.48 1.26 1.66 1.40 1.17
SD 0.13 0.08 0.10 0.09 0.07 0.09
CV 5.15 4.65 5.48 6.00 4.28 6.86
m1L m1W m2L m2W m3L m3W L/W m1 L/W m2 L/W m3
N 10 10 17 19 5 5 10 17 5
M 2.27 1.40 1.94 1.48 1.83 1.35 1.62 1.31 1.36
min 2.14 1.28 1.77 1.33 1.70 1.22 1.51 1.20 1.30
max 2.52 1.51 2.09 1.64 1.92 1.42 1.76 1.38 1.43
SD 0.11 0.08 0.10 0.09 0.08 0.08 0.08 0.05 0.06
CV 4.94 5.65 5.37 6.18 4.41 5.93 4.79 3.92 4.52 Scottimus sp., cf. S. longiquus
M1L M1W M2L M2W M3L M3W L/W M1 L/W M2 L/W M3
N 6 6 1 1 1 1 6 1 1
M 2.97 2.01 2.16 1.69 1.78 1.43 1.48 1.28 1.24
Min 2.87 1.90 1.42
Max 3.07 2.16 1.52
SD 0.07 0.09 0.04
CV 2.21 4.47 2.42
m1L m1W m2L m2W m3L m3W L/W m1 L/W m2 L/W m3
N 3 3 4 4 1 1 3 4 1
M 2.68 1.73 2.18 1.60 2.17 1.58 1.55 1.37 1.37
Min 2.60 1.72 2.17 1.53 1.50 1.25
Max 2.75 1.75 2.22 1.73 1.60 1.42
SD 0.08 0.02 0.03 0.09 0.05 0.08
CV 2.82 0.88 1.15 5.75 3.14 5.59 ______________________________________________________________________________________________ of the posterior cingulum which is greatly shortened due to the reduction in the posterior width of the tooth. Discussion—The specimens from Blue Ash are referable to Scottimus kellamorum based mainly on theproportions of the molars. In his diagnosis, Black (1961b) noted that the upper molars of S. kellamorum were similar in size to those of S. exiguus, but were clearly not as wide relative to length. These same proportions are true for both the upper and lower molars referred here to S. kellamorum. The type of Scottimus kellamorum is from the Arikareean of Montana. The only other specimen referable to this species other than the Blue Ash material is a single partial maxilla with M2-M3 from the Whitneyan of South Dakota, YPM-PU 23264, originally identified as Scottimus sp. (Korth, 1981a).
The latter specimen was distinct from S. exiguus in its narrower M2 (Korth, 1981a:table 3), a character of S. kellamorum. YPM-PU 23264 should be referred to S. kellamorum and extends the range of the species to the Whitneyan.
Scottimus sp. cf. S. longiquus Korth, 1981a (Figure 2G-K; Table 3)
Scottimus longiquus Korth, 1981a (in part) Referred Specimens—CM 83761, partial maxilla with M1-M2; CM 83756, 83763, 83764, 83767– M1; CM 83789 – M3; CM 83897, 83898, 83801, 84653 – m1; CM 83809, 83816, 83820, 83824 – m2; CM 83836 – m3.

PALUDICOLA, VOL. 7, NO. 4, 2010 126
Description—The only tooth not previously described for S. longiquus is M3. All of the remaining cheek teeth in the Blue Ash fauna have the same morphology as described for the type and referred material (Korth, 1981a). M3 is very similar to that of S. lophatus (Wood, 1937:fig. 66), but is proportionally longer. The anterior cingulum extends the entire anterior width of the tooth and wraps around both the anterolingual and anterobuccal corners of the tooth. The paracone is conical and connected to the anterior cingulum anteriorly at its lingual side. The ectoloph extends posterior from the paracone, curving buccally and completely wrapping around the posterobuccal part of the tooth, forming a large loop. At the posterolingual side of the loop is a small hypocone. An anterior loph runs directly to the buccal side of the protocone, isolating an anteroposteriorly oriented basin in the center of the tooth. The protocone is obliquely compressed into a ridge that extends posteriorly from its lingual end toward the hypocone, defining another basin lingual to the large central one. Discussion—The larger specimens of Scottimus from Blue Ash are morphologically indistinguishable from S. longiquus described from the Orellan of Montana (Korth, 1981a). However, they are slightly larger in size than the latter and slightly smaller than those of S. lophatus from the Whitneyan of Nebraska (Wood, 1937). Korth (1981a) referred two specimens from the Whitneyan of South Dakota questionably to S. longiquus. These specimens are slightly larger than the type specimen (Korth, 1981a:table 4) and likely represent the same species that is present in the Blue Ash fauna. Besides the differences noted by Korth (1981a) that separate S. longiquus from S. lophatus, it appears that the relative length of M3 is also greater in the former than in the latter.
Leidymys Wood, 1936 Leidymys juxtaparvulus n. sp.
(Figure 3A-E; Table 4) Type Specimen—CM 83852, left M1. Referred Specimens—CM 83842, 83844, 83846, 83848, 83853-83858, 83971 – M1; CM 83861, 83865, 83866, 83871, 83874, 83876, 83877 – M2; CM 838883, 83884, 83888-83890, 83892, 83893, 83895, 83896, 83899, 83901, 84648 – m1; CM 83803, 83917, 83924, 83926 – m2; CM 83832, 83928, 83930 – m3. Diagnosis—Small species similar in size to L. korthi, smaller than all other species except L. parvus; differs from L. korthi in having M1 the protocone never attaching to anterocone and in possessing a small anterostyle; differs from other species in having upper molars with a long but weak mesoloph that is much lower than the other lophs; differs from L. korthi and L. alicae in having a small spur present lingual to
hypoconid on m1 within the posterior basin (as in L. nematodon and L. blacki); differs from L. nematodon and L. blacki in lacking a connection between the protoconid and anteroconid on m1. Etymology—Latin, juxta, next to; and parvulus, smallest. Description—M1 is the largest of the upper molars. The anterocone is semicircular in occlusal outline and extends anteriorly well beyond the paracone. The anterocone is limited to the buccal half of the tooth. The valley between the anterocone and protoloph is deep. Lingual to the anterocone, anterior to the protocone is a small, distinct anterostyle. On a few M1s it is modified into a high anteroposteriorly running loph between the protocone and the posterolingual corner of the anterocone. The paracone and metacone are oval (anteroposteriorly compressed), and the lingual cusps (protocone and hypocone) are slightly crescentic in outline. The anterior arm of the protocone extends anterobuccally, ending in the valley between the paracone and anterocone. On one specimen, CM 83853, the anterior arm of the protocone continues to the posterior wall of the anterocone. This connection is very weak, not strongly developed as in specimens of L. korthi (Williams and Storer, 1998). One specimen, CM 83857, has the anterior arm of the protocone running directly buccally and ending at the anterobuccal corner of the paracone. The paracone is isolated or only weakly connected to the protocone. After moderate wear, there is a thin connection between the paracone and the posterobuccal side of the protocone. On only one specimen, CM 83857, the connection is with the anterobuccal side of the protocone. The mesoloph extends to the buccal margin of the tooth, ending in a small mesostyle. The mesoloph is much lower than the metaloph and surrounding cusps. On only two of the eleven M1s, the mesoloph is extremely short, but the mesostyle is present with a short lingual extension. The metacone connects to the hypocone via the metaloph that meets the hypocone at the anterobuccal corner of the cusp. The posterior cingulum is high and extends from the apex of the hypocone around the posterior end of the tooth, ending lingually posterior to the metacone. M2 is similar in morphology to M1, but lacks the anterocone. The protocone is obliquely compressed rather than crescentic as in M1. The anterior cingulum extends from the buccal edge of the tooth to the junction with the anterior arm of the protocone. Lingual to this junction, the anterior cingulum is variable, ranging from almost nonexistent to nearly as high as the buccal half and extending nearly to the lingual edge of the tooth. The paracone connects directly lingually with the protocone on all specimens. On one specimen, CM 83877, there is a second posterior connection. The mesoloph is variable in

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 127
FIGURE 3. Molars of Leidymys from the Blue Ash local fauna. A-E, L. juxtaparvulus. A, CM 83852, left M1 (type). B, CM 83874, left M2. C, CM 83901, left m1. D, CM 83917, left m2. E, CM 83928, right m3. F-K, L. sp., cf. L. blacki. F, CM 83847, left M1. G, CM 83879, left M2. H, CM 83880, left M3. I, CM 83897, left m1. J, CM 87915, left m2. K, CM 83929, left m3. Bar scale = 1 mm. __________________________________________________
length, from very short to reaching the buccal margin of the tooth. The arrangement of the metacone, hypocone and posterior cingulum is identical to that of M1. No specimens of M3 have been referred to this species. The m1 is anteroposteriorly elongated and narrower anteriorly than posteriorly. The anteroconid is small and crescentic, wrapping around the anterior margin of the tooth. The anteroconid is isolated or weakly connected to the metaconid but never connected to the protoconid. The metaconid is circular in occlusal outline; the entoconid is anteroposteriorly compressed; and the buccal cusps (protoconid, hypoconid) are slightly crescentic. The posterior arm of the protoconid extends across the tooth, uniting with the metaconid posteriorly. The ectolophid extends directly posteriorly, from the protoconid to the hypoconid. A small mesoconid is present at the center of the ectolophid. The mesolophid is usually long, but can be variable in length. A buccal extension from the mesoconid is present on only two of the m1s. The hypolophid runs from the entoconid directly buccally, joining the hypoconid at its anterolingual corner. The posterior cingulum runs from the apex of the
hypoconid around the back end of the tooth, joining the entoconid at its posterolingual corner, isolating a posterior valley. Within the posterior valley is a short lophule (spur) that runs lingually from a point just posterior to the hypoconid on the posterior cingulum. On some specimens, this is just an enlarged cuspule rather than a distinct lophule. ___________________________________________ TABLE 4. Dental measurements of Leidymys from the Blue Ash fauna. Abbreviations as in Table 2. Measurements in mm. Leidymys juxtaparvulus
M1L M1W M2L M2W M3L M3W
N 11 12 7 7
M 1.99 1.40 1.47 1.34
Min 1.82 1.27 1.42 1.20
Max 2.11 1.51 1.54 1.41
SD 0.10 0.08 0.04 0.08
CV 4.87 5.45 2.82 5.78
m1L m1W m2L m2W m3L m3W
N 11 11 4 4 3 3
M 1.79 1.26 1.50 1.22 1.56 1.18
Min 1.73 1.18 1.44 1.16 1.49 1.13
Max 1.85 1.33 1.56 1.27 1.63 1.21
SD 0.04 0.05 0.05 0.05 0.07 0.04
CV 2.16 3.95 3.54 3.97 4.49 3.54 Leidymys sp., cf. L. blacki
M1L M1W M2L M2W M3L M3W
N 10 10 14 14 4 4
M 2.25 1.57 1.67 1.53 1.43 1.46
Min 2.08 1.45 1.58 1.44 1.38 1.41
Max 2.37 1.73 1.82 1.68 1.49 1.49
SD 0.09 0.08 0.07 0.08 0.05 0.04
CV 3.91 5.31 3.95 5.09 3.19 2.46
m1L m1W m2L m2W m3L m3W
N 9 9 24 24 8 8
M 1.98 1.45 1.80 1.45 1.78 1.42
Min 1.86 1.34 1.63 1.33 1.65 1.30
Max 2.09 1.55 1.91 1.64 1.90 1.55
SD 0.06 0.07 0.08 0.07 0.08 0.07
CV 3.19 4.59 4.19 4.75 4.44 5.09
__________________________________________ The second lower molar is rectangular in occlusal outline, longer than wide. The anterior cingulum runs the entire width of the tooth. The metaconid and entoconid are anteroposteriorly compressed; the protoconid is crescentic; and the hypoconid is obliquely compressed. The metalophid curves

PALUDICOLA, VOL. 7, NO. 4, 2010 128
anteriorly from the metaconid to join the anterior cingulum. The anterior arm of the protoconid joins the anterior cingulum at the same point. The ectolophid runs posteriorly and slightly lingually from the protoconid to the hypoconid. The posterior arm of the protoconid (=anterior mesolophid) is variable in length, usually running about half the distance from the ectolophid to the lingual edge of the tooth, but can extend longer. The second mesolophid is always short, extending to about the center of the valley separating the metaconid and entoconid. The entoconid, hypoconid, and posterior cingulum are similar to those in m1, but there is no cuspule or small spur extending into the posterior basin. The m3 is the smallest of the lower molars and narrower posteriorly than anteriorly. The anterior cingulum and cusps of the metalophid are nearly identical to that of m2. The entoconid and hypoconid are greatly reduced in size and the mesolophid (second mesolophid) is lacking. The posterior cingulum extends more posteriorly than in the anterior molars, making the tooth look more elongated. Discussion—The morphology of the molars of Leidymys juxtaparvulus is most similar to that of L. blacki from the Arikareean of South Dakota and Nebraska (Macdonald, 1963, 1970; Martin, 1980), but it is much smaller, being almost exactly the same size as L. korthi from the Arikareean of Saskatchewan (Williams and Storer, 1998:table 2). The molars of L. juxtaparvulus differ from those of L. korthi in lacking the strong connection between the anterior arm of the protocone and the anterocone on M1, possessing a short lophule (spur) extending lingually into the posterior basin of m1, and weaker development of the mesoloph on the upper molars. Leidymys juxtaparvulus is easily distinguished from L. sp., cf. L. blacki from Blue Ash by its smaller size (Table 4), weaker development of the mesoloph on the upper molars, and lack of a connection between the protoconid and anteroconid on m1.
Leidymys sp., cf. L. blacki (Macdonald, 1963) (Figure 3F-K; Table 4)
Referred Specimens—CM 83840, 83841, 83845, 83847, 83849-83851, 83859, 83870, 83972 – M1; CM 838774, 83822, 83860, 83862-83864, 83867-83869, 83873, 83875, 83878, 83879, 83969 – M2; CM 83787, 83880-83882 – M3; CM 83885-83887, 83891, 83894, 83897, 83898, 83900 – m1; CM 83804, 83904-83916, 83918-83923, 83925, 83927, 83973, 84525 – m2; CM 83929, 83931-83937, 83970 – m3. Description—The morphology of the Blue Ash sample is smaller in size but indistinguishable from teeth described and figured elsewhere for Leidymys blacki (Macdonald, 1963, 1970; Martin, 1980). There
is very little variation in the occlusal morphology of the molars. There is a small bump in the endoloph posterior to the protoloph on both M1 and M2 that is variable in size from absent to nearly forming a double connection with the paracone. On M2, the lingual half of the anterior cingulum is always present, but the height ranges from very low to nearly as high as the protoloph. M3 is the most variable of the upper molars. The size of the hypocone and metacone are the most variable characters. On m1, the anteroconid is usually connected lingualy with the metaconid, but on some specimens a short loph connects the protoconid to the anteroconid. Otherwise there is no variation m1. The relative lengths of the two mesolophids on m2 vary slightly. Generally, the anterior mesolophid (=posterior arm of the prototonid) is longer than the posterior mesolophid, but the former is variable in length and may reach the lingual border of the tooth or be shorter than the posterior mesolophid. On a few specimens of m2, the lingual ends of the two mesolophids fuse. The only other variation on m2 is the presence of a short spur into the basin formed by the posterior cingulum and hypolophid. This lophule is present on about 25% of the specimens. On a few specimens, there is a small cuspule (hypoconulid) in its place. The size of the entoconid and the posterior mesolophid are the most variable characters on m3. The entoconid may be nearly as large as the metaconid on some specimens, but may be completely absent on others. Similarly, the posterior mesolophid is never very long and may be completely absent on some specimens. Discussion—Although there is very little difference in the occlusal morphology of the molars from Blue Ash referred to Leidymys sp., cf. L. blacki and L. blacki from the Arikareean of Nebraska and South Dakota, the average dimensions of the molars from Blue Ash are consistently 9% smaller than those reported for L. blacki (Macdonald, 1963:99, 1970:table 32; Martin, 1973:table 9; Table 4, this paper). However, the size of the molars from Blue Ash are still larger than the next smallest species of Leidymys, L. alicae (Black, 1961c:74; Rasmussen, 1977:tables 60, 61). Due to the minor differences in size, the Blue Ash material cannot be definitely referred to L. blacki.
Paciculus Cope, 1879 Paciculus dakotensis n. sp.
(Figure 4A-F; Table 5) Type Specimen—CM 83942, right M1. Referred Specimens—CM 83843, 83943, 83953, 83954, 83967, 83975, 83977, 83978, 83983, 83992-83995 – M1; CM 83872, 83987, 84549, 84556, 84558, 84566, 84568 – M2; CM 84569, 84570 –M3; CM 84500, 84503, 84506, 84516-84520, 84523, 84526,

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 129
84527, 84531, 84532, 84534-84536, 84540, 84543-84545 – m1; CM 84586, 84594-84596, 84600, 84601, 84603, 84610, 84611, 84620 – m2; CM 84628, 84639 – m3. Diagnosis—Smaller than all other species except P. woodi; M3 longer than wide (wider than long in other species); upper molars not as wide relative to length as in P. insolatus; anterocone on M1 similar to that of P. insolatus, not as anteriorly expanded as in other species. Etymology—Dakota, reference to the state where the type and referred specimens were recovered (South Dakota) and the Greek suffix, -ensis, referring to location. Description—The molars are mesodont in crown height, higher crowned than the other cricetids from Blue Ash except Geringia (described below). The upper molars decrease in size from M1 to M3. M1 is longer than wide, similar to P. nebraskensis and P. montanus in proportions, not proportionally as wide as in P. insolatus. The anterocone is a large, distinct cusp that projects anteriorly as in other species. The anterior extent of the anterocone is similar to that of P. insolatus and does not extend as far anteriorly as in P. montanus or P. nebraskensis. The anterocone is attached lingually to the anterior arm of the protoconid. In approximately one-third of the M1s, there is a short lophule extending buccally or anterobuccally into the valley between the protoloph and the anterocone. This is just a small spur that is always free at the end, not fusing with any other cusp or loph. The buccal cusps (paracone and metacone) are anteroposteriorly compressed, and the protocone and hypocone are obliquely compressed. The paracone attaches via the protoloph, directly to the posterobuccal corner of the protocone. The endoloph is continuous posteriorly with the hypocone. The mesoloph is long, always reaching the lingual edge of the tooth. The metaloph is similar to the protoloph; the metacone attaching to the posterobuccal corner of the hypocone. The posterior cingulum runs from the apex of the hypocone the entire width of the tooth along its posterior margin, ending posterior to the metacone. A narrow valley separates the lingual end of the posterior cingulum and the metacone. On two specimens, CM 83995 and CM 83967, there is an anterior cingulum, anterior to the protocone, low on the crown. There is no anterior cingulum on any of the other specimens. M2 is rectangular in outline, being longer than wide. The occlusal morphology is very similar to that of M1, but lacking the large anterocone. The anterior cingulum extends the anterior width of the tooth, arising from the protocone. The lingual cusps (protocone, hypocone) are obliquely compressed to a greater degree than in M1, but the attachments of the lophs and cusps are identical to that of M1.
FIGURE 4. Molars of Paciculus from the Blue Ash local fauna. A-F, P. dakotensis. A, CM 84570, right M3. B, CM 84566, right M2. C, CM 83942, right M1 (type). D, CM 84628, right m3. E, CM 84620, right m2. F, CM 84527, right m1. G-I, P. sp., cf. P. nebraskensis. G, CM 83839, left M1. H, CM 84565, left M2. I, CM 84573, right m3. Bar scale = 1 mm.
___________________________________________ There are only two specimens referable to M3. The tooth is roughly triangular in occlusal outline, and slightly longer than wide (Table 4). M3 is smaller than the other upper molars. The occlusal morphology of CM 84570 is similar to that of M2s except that the hypocone and metacone are greatly reduced in size, making the tooth taper posteriorly. The other M3, CM 84569, has a less reduced metacone, a protocone that is not obliquely compressed, and the mesoloph connects directly with the protocone rather than arising from the endoloph. There is no endoloph on CM 84569. The m1 is similar in size to m2, but narrower anteriorly than posteriorly. The anteroconid is very small or completely absent. It is situated on the buccal side of the anterior edge of the tooth, even with or slightly anterior to the metaconid. The anteroconid is isolated from the metaconid and protoconid on 35% of the m1s, and connected lingually to the metaconid on 35% of the specimens. Only on four of the 20 specimens is there a connection with the anteroconid and both the metaconid and protoconid. Any

PALUDICOLA, VOL. 7, NO. 4, 2010 130
connection with the protoconid is a very weak, thin loph. In advanced stages of wear the anteroconid is continuous with both the metaconid and protoconid. The protoconid is circular in occlusal outline and positioned anterior to the protoconid. The protoconid is obliquely compressed. There are three transverse lophs on m1. The most anterior is the posterior arm of the protoconid that is slightly curved, and joins the posterior side of the metaconid at the lingual edge of the tooth. On 80% of the specimens this loph is complete or nearly complete. In the remainder (four specimens) the posterior arm of the protoconid is a short loph, extending only about half the width of the tooth. The second transverse loph is the mesolophid. It is usually a complete loph from the ectolophid to the lingual margin of the tooth, ending in a distinct metastylid. Commonly, the mesolophid is short, but is complemented by a buccally directed lophule from the metastylid. Only on three specimens is the mesolophid short (half the width of the tooth) without a matching metastylid. The third transverse loph is the hypolophid that runs from the anteroposteriorly compressed entoconid to a point on the ectolophid anterior to the hypoconid. The posterior cingulum extends the width of the tooth along its posterior side from the hypoconid to a point posterior to the entoconid. With wear, the posterior cingulum always joins the entoconid. The m2 is similar in dimensions to m1, but is rectangular in occlusal outline, being as wide anteriorly as posteriorly. The anterior cingulum extends the entire anterior width of the tooth, both ends being free. The metaconid and protoconid are even with one another and both join the anterior cingulum just lingual to the protoconid in a small anteroconid. The metalophid curves slightly anteriorly to meet the anterior cingulum. The posterior arm of the protoconid extends lingually to a point posterior to the metaconid, but never reaches the lingual edge of the tooth. The ectolophid runs diagonally from the protoconid to a point on the hypolophid, just buccal to the entoconid. The ectolophid bends buccally from this point to join the hypoconid. The hypolophid is angled slightly anteriorly. The hypolophid and posterior cingulum are the same as in m1. The m3 is slightly smaller than the anterior molars, and only represented by two specimens. The tooth tapers posteriorly due to the reduction of the hypoconid and entoconid. CM 84628 has an occlusal morphology very similar to that of m2, but with the reduced posterior cusps and a posteriorly extended posterior cingulum. The other specimen of m3, CM 84639, differs in the complete lack of an entoconid and hypolophid. Discussion—Paciculus dakotensis is clearly referable to this genus because of the crown-height of
the molars (mesodont), slightly reduced anterocone on M1, and generally simplified occlusal morphology of the molars. It is distinct from other species of the genus in its size. It is smaller than any other species except P. woodi, the smallest described species (Wood, 1936:table I; Black, 1961a:12; Nichols, 1979:table 8). The reduction of the anterocone on M1 in P. dakotensis is similar to the condition in the type species P. insolatus (Wood, 1936:fig. 4; Martin, 1980:fig. 20A), but P. dakotensis does not have upper molars as wide relative to the length as in the former. The small lophule in the valley between the protoloph and the anterocone on M1 of P. dakotensis occurs in only about a third of the specimens, but is unknown in any other species of the genus. Its presence is believed to be a primitive condition because this loph is commonly present and often complete to the anterocone in a number of eumyine cricetids. Paciculus dakotensis also differs from other species of the genus in that M3 is longer than wide. In all other species M3 is wider than long. Specimens referable to Geringia from the Blue Ash fauna can be separated by their smaller size from those of Paciculus dakotensis as well as other morphologies of the molars, especially m1 and M1 (see below discussion of Geringia). ___________________________________________ TABLE 5. Dental measurements of Paciculus dakotensis from the Blue Ash fauna. Abbreviations as in Table 2. Measurements in mm.
M1L M1W M2L M2W M3L M3W
N 13 13 7 7 2 2
M 2.11 1.58 1.70 1.44 1.52 1.37
Min 2.06 1.43 1.63 1.34 1.5 1.3
Max 2.18 1.75 1.77 1.51 1.53 1.43
SD 0.03 0.09 0.05 0.06
CV 1.63 5.62 2.87 4.17
m1L m1W m2L m2W m3L m3W
N 20 19 10 10 1 1
M 1.88 1.35 1.83 1.83 1.73 1.43
Min 1.78 1.24 1.76 1.46
Max 2.01 1.46 1.95 1.66
SD 0.06 0.06 0.06 0.06
CV 3.06 4.12 3.37 3.37
____________________________________________
Paciculus sp., cf. P. nebraskensis Alker, 1969 (Figure 4G-I)
Referred Specimens—CM 83839, fragmentary maxilla with left M1; CM 84565, left M2; and CM 84573, right m3.
KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 131
Measurements—CM 83839 (M1): L = 2.45 mm; W = 1.63 mm. CM 84565 (M2): L = 1.81 mm; W = 1.60 mm. CM 84573 (m3): L = 1.97 mm, W = 1.57 mm. Discussion—The morphologies of the molars referred to Paciculus cf. P. nebraskensis are similar to those of P. dakotensis but differ in being larger and slightly lower crowned than P. dakotensis, and having the molars more anteroposteriorly elongated. These specimens most closely approach P. nebraskensis from the Arikareean in size and morphology (Korth, 1992), but appear slightly lower crowned.
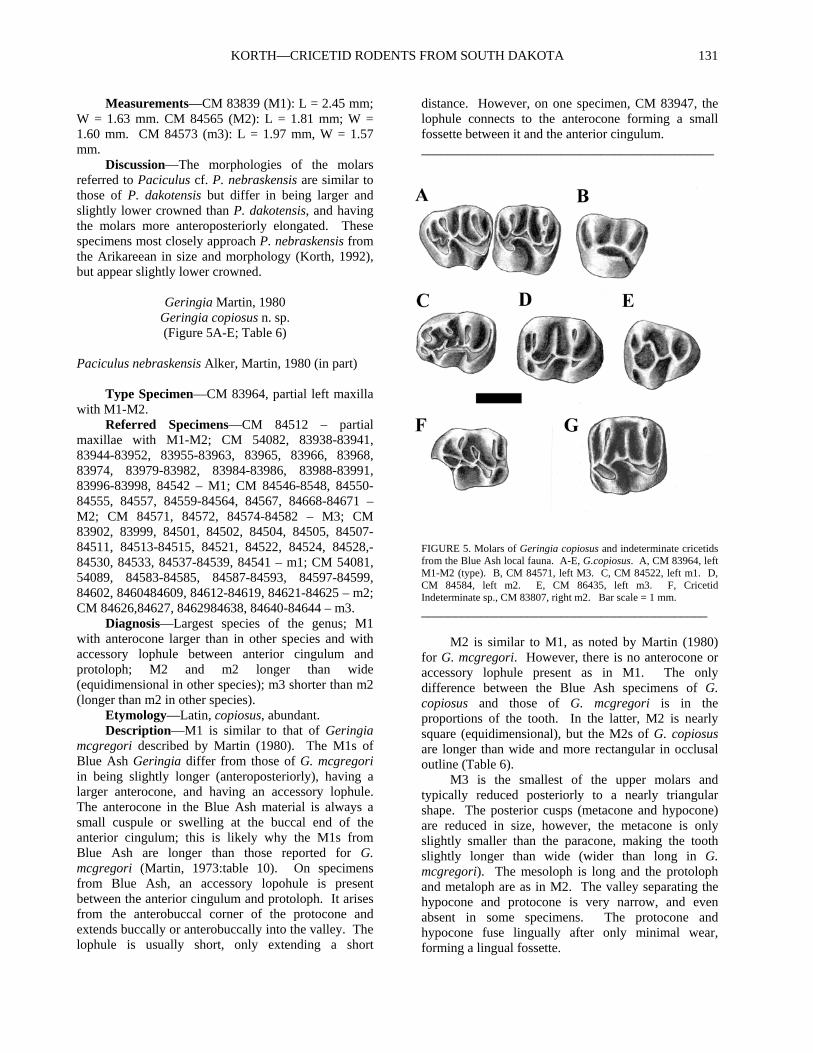
Geringia Martin, 1980 Geringia copiosus n. sp. (Figure 5A-E; Table 6)
Paciculus nebraskensis Alker, Martin, 1980 (in part) Type Specimen—CM 83964, partial left maxilla with M1-M2. Referred Specimens—CM 84512 – partial maxillae with M1-M2; CM 54082, 83938-83941, 83944-83952, 83955-83963, 83965, 83966, 83968, 83974, 83979-83982, 83984-83986, 83988-83991, 83996-83998, 84542 – M1; CM 84546-8548, 84550-84555, 84557, 84559-84564, 84567, 84668-84671 – M2; CM 84571, 84572, 84574-84582 – M3; CM 83902, 83999, 84501, 84502, 84504, 84505, 84507-84511, 84513-84515, 84521, 84522, 84524, 84528,-84530, 84533, 84537-84539, 84541 – m1; CM 54081, 54089, 84583-84585, 84587-84593, 84597-84599, 84602, 8460484609, 84612-84619, 84621-84625 – m2; CM 84626,84627, 8462984638, 84640-84644 – m3. Diagnosis—Largest species of the genus; M1 with anterocone larger than in other species and with accessory lophule between anterior cingulum and protoloph; M2 and m2 longer than wide (equidimensional in other species); m3 shorter than m2 (longer than m2 in other species). Etymology—Latin, copiosus, abundant. Description—M1 is similar to that of Geringia mcgregori described by Martin (1980). The M1s of Blue Ash Geringia differ from those of G. mcgregori in being slightly longer (anteroposteriorly), having a larger anterocone, and having an accessory lophule. The anterocone in the Blue Ash material is always a small cuspule or swelling at the buccal end of the anterior cingulum; this is likely why the M1s from Blue Ash are longer than those reported for G. mcgregori (Martin, 1973:table 10). On specimens from Blue Ash, an accessory lopohule is present between the anterior cingulum and protoloph. It arises from the anterobuccal corner of the protocone and extends buccally or anterobuccally into the valley. The lophule is usually short, only extending a short
distance. However, on one specimen, CM 83947, the lophule connects to the anterocone forming a small fossette between it and the anterior cingulum. ____________________________________________
FIGURE 5. Molars of Geringia copiosus and indeterminate cricetids from the Blue Ash local fauna. A-E, G.copiosus. A, CM 83964, left M1-M2 (type). B, CM 84571, left M3. C, CM 84522, left m1. D, CM 84584, left m2. E, CM 86435, left m3. F, Cricetid Indeterminate sp., CM 83807, right m2. Bar scale = 1 mm.
___________________________________________ M2 is similar to M1, as noted by Martin (1980) for G. mcgregori. However, there is no anterocone or accessory lophule present as in M1. The only difference between the Blue Ash specimens of G. copiosus and those of G. mcgregori is in the proportions of the tooth. In the latter, M2 is nearly square (equidimensional), but the M2s of G. copiosus are longer than wide and more rectangular in occlusal outline (Table 6). M3 is the smallest of the upper molars and typically reduced posteriorly to a nearly triangular shape. The posterior cusps (metacone and hypocone) are reduced in size, however, the metacone is only slightly smaller than the paracone, making the tooth slightly longer than wide (wider than long in G. mcgregori). The mesoloph is long and the protoloph and metaloph are as in M2. The valley separating the hypocone and protocone is very narrow, and even absent in some specimens. The protocone and hypocone fuse lingually after only minimal wear, forming a lingual fossette.

PALUDICOLA, VOL. 7, NO. 4, 2010 132
The m1 is tapered anteriorly and is similar in size to m2. The anteroconid is very small and connects to both the metaconid and protoconid on 73% of the specimens. The few without the connection have very little wear. This connection isolates a small anterior fossettid as described by Martin (1980) for the genus. ____________________________________________ TABLE 6. Dental measurements of Geringia copiosus from the Blue Ash fauna and comparative measurements of G. mcgregori. G. mcgregori based on measurements listed in Martin (1973:table 10) from the Gering Formation of Nebraska. Abbreviations as in Table 2. Measurements in mm. Geringia copiosus
M1L M1W M2L M2W M3L M3W
N 45 45 18 18 11 11
M 1.78 1.45 1.48 1.30 1.32 1.22
Min 1.55 1.16 1.37 1.17 1.27 1.19
Max 2.00 1.74 1.55 1.42 1.40 1.26
SD 0.11 0.12 0.05 0.08 0.04 0.02
CV 6.42 8.03 3.60 6.17 3.28 1.87
m1L m1W m2L m2W m3L m3W
N 25 25 34 35 17 16
M 1.65 1.20 1.66 1.37 1.59 1.32
Min 1.5 0.98 1.55 1.17 1.44 1.20
Max 1.77 1.37 1.75 1.48 1.68 1.43
SD 0.07 0.09 0.06 0.07 0.06 0.06
CV 4.10 7.19 3.40 5.15 4.07 4.77 Geringia mcgregori
M1L M1W M2L M2W M3L M3W
N 4 4 4 4 3 3
M 1.53 1.33 1.42 1.41 1.15 1.21
Min 1.47 1.20 1.33 1.36 1.12 1.18
Max 1.57 1.39 1.46 1.48 1.18 1.26
SD 0.05 0.09 0.06 0.05 0.03 0.04
CV 3.20 6.75 4.10 3.88 2.65 3.43
m1L m1W m2L m2W m3L m3W
N 4 4 4 4 3 3
M 1.71 1.28 1.56 1.49 1.71 1.36
Min 1.63 1.22 1.49 1.43 1.60 1.11
Max 1.75 1.41 1.63 1.61 1.79 1.56
SD 0.06 0.09 0.06 0.08 0.10 0.23
CV 3.37 7.09 3.84 5.56 5.69 16.85
__________________________________________ The posterior arm of the protoconid is almost always complete, or nearly complete to the lingual margin of the tooth, usually fusing with the posterior side of the metaconid, and rarely with the metastylid. A
mesolophid is usually present but is almost always very short or even absent. On three of 26 specimens, the mesolophid extends the entire width of the tooth. Martin (1980) diagnosed the genus as having the mesolophid and hypolophid of m1 forming a V-shape. However, it appears that what he was referring to as a mesolophid is the posterior arm of the protoconid. The ectolophid connects the protoconid to the hypoconid and the entoconid is attached to the ectolophid anterior to the hypoconid via the hypolophid. The posterior cingulum encircles a posterior basin as described by Martin (1980) for G. mcgregori. The m2 is rectangular in occlusal outline and nearly identical in size to m1 (Table 6). In other species of Geringia, the m2 is nearly square in outline and shorter than m1. The morphology of m2 of G. copiosus differs little from that described for G. mcgregori (Martin, 1980). The anterior cingulum has a short lingual extension anterior to the protoconid, and both the metaconid (via the metalophid) and protoconid join the anterior cingulum buccal to its center. The mesolophid (which consists of the posterior arm of the protoconid) is always long, but does not always extend the entire width of the tooth. The remainder of the morphology of m2 is the same as in m1. Unlike the m3 of G. mcgregori, the m3 of G. copiosus is shorter than m2. The morphology of the anterior cingulum, protoconid, and metaconid are similar to that of m2. The m3 is tapered and expanded posteriorly. The hypoconid is greatly reduced and the entoconid variable reduced. There are always two transverse lophs: the posterior arm of the protoconid and the hypolophid. The hypolophid is always complete but the posterior arm of the protoconid (mesolophid) does not always reach the lingual edge of the tooth as in m2. Discussion—Geringia copiosus is the best represented rodent in the Blue Ash fauna, being known from more than 150 specimens. It is referable to Geringia mainly based on its mesodont molars and the marked reduction of the anterocone on M1. It is easily distinguishable from the other species of Geringia by the lesser reduction of the anterocone on M1, its larger size, presence of a short mesolophid on m1, and the proportions of the teeth (M2, m2 longer than wide; m3 shorter than m2). All specimens of M1 of G. copiosus have a short lophule that extends buccally from the protocone into the valley separating the protoloph and anterior cingulum that is unknown in other species. Specimens of Geringia copiosus are easily separable from those of Paciculus dakotensis by size (Tables 5, 6) and molar morphology, especially M1 and m1. The M1s of G. copiosus have a much smaller anterocone and always have the distinct accessory lophule between the protoloph and anterior cingulum.

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 133
On P. dakotensis, the anterocone is larger and the accessory lophule is present on only some of the specimens. The m1 of G. copiosus usually has two transverse lophs (posterior arm of the protoconid and hypolophid), whereas m1 of P. dakotensis has three (posterior arm of protoconid, mesolophid, hypolophid). There is usually a mesolophid on m1 of G. copiosus, but it is very short and does not reach the lingual side of the tooth as in P. dakotensis. The anteroconid is almost always connected to the protoconid on specimens of G. copiosus, but in P. dakotensis, this connection is never until the tooth is in a late stage of wear. These differences between Geringia and Paciculus from Blue Ash also appear to be distinctions between other species of these genera (see Black, 1961a:fig. 4; Nichols, 1979:pl. 6, figs. 7, 8; Martin, 1980:figs. 16-18; Korth, 1992:fig. 16). Martin (1980:fig. 21A) figured a partial maxilla with M1-M2 that he identified as Paciculus nebraskensis. Later, Korth (1992) described a large sample of P. nebraskensis and noted that the specimen figured by Martin did not belong to that species and was likely a specimen of Geringia. The specimen figured by Martin, UNSM 11527, is of appropriate size to belong to G. copiosus, and has the accessory lophule on M1 diagnostic of the species. It is very likely that UNSM 11527 is referable to G. copiosus.
Cricetid indeterminate sp. (Figure 5F)
Referred Specimens—CM 83807, right m2; and CM 86472, CM 84661, left M2s. Measurements—CM 83807 (m2): L = 1.61 mm, W = 1.38 mm. CM 86472 (M2): L = 1.66, W = 1.60 mm. CM 84661 (M2): L = 1.70; W = 1.61. Description—The m2 (CM 83807) is rectangular, longer than wide. It is very low crowned, much lower than the other cricetids reported from Blue Ash. The anterior cingulum extends the entire width of the anterior margin of the tooth, joining the metaconid linugally, and wrapping around the anterobuccal corner of the tooth. The lingual cusps are anteroposteriorly compressed and the buccal cusps are obliquely compressed. The metalophid curves anteriorly from the metaconid and is joined by the anterior arm of the protoconid just before it joins the anterior cingulum just lingual to the protoconid. The connection of the metalophid to the anterior cingulum is wide, but there is no distinct cuspule. The ectolophid runs only slightly diagonally from the protoconid to the hypoconid. There is a small mesoconid along it, just anterior to the hypoconid. The mesolophid is a continuation of the posterior arm of the protoconid and ends in the center of the valley separating the metaconid and entoconid. The lingual end of the
mesolophid is a triangular cusp. A minute metastylid is at the base of the posterior slope of the metaconid along the lingual edge of the tooth. A low ridge runs along the lingual edge of the tooth between the metaconid and entoconid. The hypolophid runs buccally from the entoconid, joining the ectolophid just posterior to the mesoconid. A small, thin lophule runs directly posteriorly from the junction of the ectolophid and the hypolophid and joins the posterior cingulum, forming a triangular fossette lingual to the hypoconid and ending posteriorly at a hypoconulid. The posterobuccal corner of the hypoconid is broken away, but the posterior cingulum runs from the hypoconid along the posterior margin of the tooth and is continuous with the posterior wall of the entoconid, enclosing another triangular basin lingual to the first. The M2 is nearly square in occlusal outline, just slightly longer than wide. It is brachydont, but slightly higher-crowned lingually than buccally. The anterior cingulum extends the entire width of the tooth. The protoloph joins the anterior cingulum lingual to the center of the tooth via the anterior arm of the protocone. The buccal cusps are anteroposteriorly compressed and the protoloph and metaloph are parallel and directed transversely across the tooth. The lingual cusps (protocone and hypocone) are obliquely compressed. There is a high anteroposteriorly directed ridge along the buccal border of the tooth, connecting the paracone and metacone, and enclosing a basin between the protoloph and metaloph. The enoloph runs atnerobuccally from the hypocone and joins the buccal side fo the protocone by a narrow connetion. Just posterior to the junction of the entoloph with the protocone is a small spur that extends buccally. The mesolophis lang and parallel to the metaloph and protoloph, but ends just before meeting the buccal anteroposterior ridge. The posterior cingulum extends the entire width of the tooth from the apex of the hypocone along the posterior margin of the tooth, ending at the buccal edge of the tooth, just posterior to the metacone Discussion—The occlusal morphology of the molars referred to an indeterminate cricetid most closely approaches those of Geringia and Paciculus but are much lower crowned. On m2, the distinctive accessory loph dividing the posterior basin of the in two and the triangular cusp at the lingual end of the mesolophid are unique among the Blue Ash cricetids.
The M2s are referred to the same species as the lower molar due to their similar size and crown-height. Also, both molars have a distinct anteroposteriorly directed loph isolating a central basin. On the M2 it is along the buccal margin and on the m2 it is along the lingual margin. At present, these specimens cannot be referred to any known genus or species of cricetid from North America.

PALUDICOLA, VOL. 7, NO. 4, 2010 134
CONCLUSIONS The age of the Blue Ash anthill fauna has been questioned because of the combination of Arikareean, Whitneyan, and earlier taxa (see Korth, 2009). The cricetid fauna is similarly ambiguous. At the level of genus, the Blue Ash fauna appears to be Arikareean with the presence of otherwise Arikareean and younger genera such as Paciculus, Geringia, and Leidymys (Martin, 1980; Korth, 1994; Lindsay, 2008). However, the species from Blue Ash referable to these genera are either smaller or morphologically more primitive. G. copiosus has a less reduced anterocone on M1 than Arikareean species, P. dakotensis and L. juxtaparvulus are both smaller than nearly all of the Arikareean species. The species from Blue Ash that occur in the Arikareean cannot be definitely referred to these species, Leidymys sp., cf. L. blacki and Paciculus sp., cf. P. nebraskensis, because they are either smaller (as in the former) or lower crowned (as in the latter). Similarly, Scottimus kellamorum was originally reported from the Arikreean (Black, 1961b). However, it appears that it is also from the Whitneyan (= Scottimus sp., Korth, 1981a). Two species, Eumys parvidens and Scottimus longiquus, were previously known from the Orellan (Wood, 1937, Korth, 1981a, 1994). However, S. longiquus also has been reported from the Whitneyan (Korth, 1981a). Eumys brachyodus is restricted to the Whitneyan (excluding synonymy of E. eliensis: see above discussion of Eumys). The cricetid fauna from Blue Ash is similar to that of other groups described in that the provincial ages Whitneyan and Arikareean cannot be definitely assigned. The cricetids suggest that this fauna is either the earliest Arikareean (marked the first occurrence of Leidymys, Geringia, and Paciculus) or the latest Whitneyan (presence of relict species Eumys brachyodus, E. parvidens and Scottimus longiquus).
ACKNOWLEDGMENTS The specimens described above were graciously loaned by Dr. M. R. Dawson of CM, with curatorial assistance from A. Henrici. AMNH specimens of E. brachyodus were loaned with permission of Dr. J. Meng with assistance from I. Rutzky. Measurement of UNSM specimens of Eumys elegans was permitted by Dr. M. R. Voorhies of that institution. Statistical analyses were performed by Dr. J. A. Massare of SUNY Brockport. Camera-lucida for figures was provided by Dr. G. McIntosh of the Rochester Museum and Science Center. Earlier versions of this paper were critically reviewed by Drs. J. Storer and J. Baskin.
LITERATURE CITED Black, C. C. 1961a. Rodents and lagomorphs form the
Miocene Fort Logan and Deep River Formations of Montana. Postilla 48:1-20.
Black, C. C. 1961b. New rodents from the early Miocene deposits of Sixty-Six Mountain, Wyoming. Breviora 146:1-7.
Black, C. C. 1961c. Fossil mammals from Montana. Pt. 1. Additions to the late Miocene Flint Creek local fauna. Annals of Carnegie Museum 36:69-76.
Galbreath, E. C. 1953. A contribution to the Tertiary geology and paleontology of northeastern Colorado. University of Kansas Paleontological Contributions, Vertebrata 13:1-120.
Korth, W. W. 1980. Paradjidaumo (Eomyidae, Rodentia) from the Brule Formation, Nebraska. Journal of Paleontology, 54:943-951.
Korth, W. W. 1981a. New Oligocene rodents from western North America. Annals of Carnegie Museum, 50:289-318.
Korth, W. W. 1981b. Metadjidaumo (Eomyidae, Rodentia) from Colorado and Wyoming. Journal of Paleontology, 55:598-602.
Korth, W. W. 1989. Stratigraphic occurrence of rodents and lagomorphs in the Orella Member, Brule Formation, Sioux County, Nebraska. University of Wyoming Contributions to Geology, 28:15-20.
Korth, W. W. 1992. Small mammals from the Harrison Formation (late Arikareean, early Miocene), Cherry County, Nebraska. Annals of Carnegie Museum, 61:69-131.
Korth, W. W. 1994. The Tertiary Record of Rodents in North America. Plenum Press, New York, 319 pp.
Korth, W. W. 2009. Mammals from the Blue Ash local fauna (late Oligocene), South Dakota. Rodentia. Part 3: Family Sciuridae. Paludicola 7:47-60.
Leidy, J. 1856. Notices of remains of extinct Mammalia discovered by Dr. F. V. Hayden in Nebraska Territory. Procedings of the Academy of Natural Sciences, Philadelphia 8:88-90.
Lindsay, E. H. 2008. Cricetidae. Pp. 456-476 in C. M. Janis, G. F. Gunnell and M. D. Uhen (eds.), Evolution of the Teritary Mammals of North America, Volume 2: Small Mammals, Xenarthrans, and Marine Mammals. Cambridge University Press, New York.
Macdonald, J. R. 1963. The Miocene Faunas from the Wounded Knee Area of Western South Dakota. Bulletin of the American Museum of Natural History 25:139–238.

KORTH—CRICETID RODENTS FROM SOUTH DAKOTA 135
Macdonald, J. R. 1970. Review of the Miocene Wounded Knee faunas of southwestern South Dakota. Bulletin of the Los Angeles County Museum of Natural History 8:1–82.
Martin, L. D. 1973. The mammalian fauna of the Lower Miocene Gering Formation of western Nebraska and the early evolution of the North American Cricetidae. Unpublished PhD dissertation, University of Kansas, 264 pp.
Martin, L. D. 1980. The early evolution of the Cricetidae in North America. Univeristy of Kansas Paleontological Contributions 102:1-42.
Nichols, R. 1976. Early Miocene mammals from the Lemhi Valley of Idaho. Tebiwa 18:9-47.
Rasmussen, D. L. 1977. Geology and mammalian paleontology of the Oligocene-Miocene Cabbage Patch Formation, central-western Montana. Unpublished PhD dissertation, University of Kansas, 775 pp.
Russell, L. S. 1972. Tertiary mammals of Saskatchewan, Part II, the Oligocene fauna, non-ungulate orders. Life Sciences Contribution, Royal Ontario Museum 84:1-63.
Setoguchi, T. 1978. Paleontology and geology of the Badwater Creek area, central Wyoming. Part 16. The Cedar Ridge local fauna (late Oligocene).
Storer, J. E. 1996. Eocene-Oligocene faunas from the Cypress Hills Formation, Saskatchewan. Pp. 278-311, in D. R. Prothero and R. J. Emry (eds.), The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, New York.
Storer, J. E. and H. N. Bryant. 1993. Biostratigraphy of the Cypress Hills Formation (Eocene to Miocene), Saskatchewan: equid types (Mammalia, Perissodactyla) and associated faunal assemblages. Journal of Paleontology 67:660-669.
Wahlert, J. H. 2004. The skull and dentition of Eumys elegans from the Oligocene of North Dakota. Bulletin of the Carnegie Museum of Natural History 36:335-348.
White, T. E. 1954. Preliminary analysis of the fossil vertebrates of the Canyon Ferry Reservoir area. Proceedings of the U. S. National Museum 103;305-438.
Williams, M. R. and J. E. Storer. 1998. Cricetid rodents fo the Kealy Springs local fauna (early Arikareean; late Oligocene) of Saskatchewan. Paludicola 1:143-149.
Wood, A. E. 1936. The cricetid rodents described by Leidy and Cope from the Tertiary of North America. American Museum Novitates 822:1-8.
Wood, A. E. 1937. The mammalian fauna of the White River Oligocene, Pt. 2, Rodentia. Transactions of the American Philosophical Society 28:155-269.
Wood, A. E. 1980. The Oligocene rodents of North America. Transactions of the American Philosophical Society 70:1-68.
Wood, A. E. and R. W. Wilson. 1936. A suggested nomenclature for the cusps of the cheek teeth of rodents. Journal of Paleontology 10:388-391.

PALUDICOLA, VOL. 7, NO. 4, 2010 136
APPENDIX Comparative dental measurements of Eumys. E. elegans uppers from AMNH collections (FAM) from the Orellan of North Dakota (Wahlert, 2004:table 1). E. elegans lowers from two samples of UNSM specimens from the Orellan of Sioux County Nebraska (UNSM localities Sx – 6 and Sx - 14). E. brachyodus from Wood (1937:253), Galbreath (1953:table 14), and AMNH and CM collections from the Whitneyan of North Dakota and Nebraska, respectively. Abbreviations as in Table 2. Measurements in mm. Eumys elegans
M1L M1W M2L M2W M3L M3W L/W M1 L/W M2 L/W M3
N 15 14 15 15 14 13 14 15 13
M 2.99 1.97 2.04 1.97 1.56 1.74 1.51 1.04 0.90
Min 2.80 1.80 1.90 1.80 1.30 1.60 1.45 0.95 0.81
Max 3.20 2.10 2.20 2.10 1.80 1.90 1.60 1.17 1.06
SD 0.15 0.08 0.11 0.10 0.14 0.10 0.05 0.06 0.06
CV 4.88 4.19 5.17 4.96 8.90 5.53 3.19 5.54 6.76
m1L m1W m2L m2W m3L m3W L/W m1 L/W m2 L/W m3
N 34 33 34 33 34 34 33 32 34
M 2.34 1.72 2.11 1.93 2.25 1.86 1.36 1.13 1.21
Min 2.10 1.60 1.90 1.80 1.95 1.65 1.24 1.02 1.03
Max 2.60 1.90 2.40 2.20 2.50 2.15 1.56 1.24 1.37
SD 0.12 0.08 0.10 0.09 0.12 0.11 0.07 0.06 0.08
CV 5.06 4.59 4.77 4.83 5.49 5.96 5.19 5.28 6.64
m1L m1W m2L m2W m3L m3W L/W m1 L/W m2 L/W m3
N 32 35 44 42 41 40 32 42 40
M 2.42 1.75 2.17 1.93 2.17 1.85 1.38 1.13 1.18
Min 2.10 1.55 1.90 1.70 1.95 1.65 1.21 1.02 1.08
Max 2.75 2.10 2.55 2.25 2.50 2.10 1.48 1.24 1.32
SD 0.16 0.12 0.14 0.13 0.12 0.10 0.06 0.06 0.05
CV 6.80 6.82 6.31 6.84 5.50 5.22 4.22 5.18 4.48 Eumys brachyodus
m1L m1W m2L m2W m3L m3W L/W m1 L/W m2 L/W m3
N 9 10 12 12 10 10 9 12 10
M 2.36 1.89 2.27 2.13 2.28 2.00 1.26 1.07 1.14
Min 2.10 1.71 2.02 1.90 1.92 1.72 1.18 0.97 1.08
Max 2.62 2.11 2.47 2.28 2.50 2.10 1.32 1.17 1.21
SD 0.14 0.13 0.14 0.11 0.17 0.12 0.05 0.06 0.05
CV 6.06 6.88 6.05 4.98 7.44 6.21 3.91 5.18 4.25

Paludicola 7(4):137-157 May 2010 © by the Rochester Institute of Vertebrate Paleontology
137
A NEW CHUB (ACTINOPTERYGII, CYPRINIFORMES, CYPRINIDAE) FROM THE MIDDLE MIOCENE (EARLY CLARENDONIAN) ALDRICH STATION FORMATION, LYON COUNTY,
NEVADA
Thomas S. Kelly
Research Associate, Vertebrate Paleontology Section, Natural History Museum of Los Angeles County 900 Exposition Boulevard, Los Angeles, California 90007
ABSTRACT A new chub, Lavinia lugaskii, is described from the middle Miocene (early Clarendonian) Aldrich Station Formation of Lyon County, Nevada. Lavinia lugaskii represents a basal member of the Lavinia-Hesperoleucus lineage, indicating that this lineage diverged from a common ancestor with Mylopharodon before 12.5 – 12.0 million years before present. This is the oldest recognized species of Lavinia and the first new chub species to be documented from the Miocene of Nevada in over 30 years.
INTRODUCTION A sample of fish fossils is now known from localities that occur in an outlier of the Aldrich Station Formation, exposed just west of Mickey Canyon on the northwest flank of the Pine Groove Hills, Lyon County, Nevada. All of the fish remains were recovered from a single stratigraphic level represented by a thin (~0.06 m) shale bed. This level can be traced laterally for about 0.5 km and yielded fossil fish remains at several points along its exposure. The fish producing shale bed is separated from a second slightly thicker, overlying shale bed by a thin sandstone bed. The two shale beds contain leaf impressions and yielded the Mickey Canyon Spring Flora from the Museum of Paleontology, University of California, Berkeley (UCMP), plant locality PA624 (Schorn and Shelton, 1991; Schorn et al., 1994). Previously, only one partial fish skeleton was recorded from PA624, which was collected by Howard Schorn and identified by Howard Hutchison as Cyprinidae (unpublished locality data, UCMP). An additional 70 specimens in the vertebrate paleontology collection of the Natural History Museum of Los Angeles County (LACM) from the lower fish producing shale bed, including three complete skeletons and 65 partial skeletons, allow a detailed analysis of this cyprinid fish and indicate it represents a new species. The purpose of this report is to describe this new cyprinid and compare it with other known fossil and extant chubs from the Great Basin and adjacent areas.
METHODS Measurements of the skeletons and individual
bones were made to the nearest 0.1 mm with a vernier caliper. Measurements of the pharyngeal teeth were made with an optical micrometer to the nearest 0.01 mm. Estimated standard lengths for partial skeletons were extrapolated using the mean ratios of the standard length to landmark measurements (e.g., ratios of the SL to head length, pectoral fin origin to pelvic fin origin length, the posterior anal and dorsal fin bases to caudal base lengths) for the three complete skeletons. Detailed locality data are on file at the Vertebrate Paleontology Section of the Natural History Museum of Los Angeles County. All taxa identifications were determined by the author using published accounts and comparative material in the vertebrate paleontology collections of the Natural History Museum of Los Angeles, the Museum of Paleontology, University of California, Berkeley, and the W. M. Keck Earth Science and Mineral Engineering Museum, University of Nevada, Reno. Additional comparative specimens of Lavinia (L. hibbardi, L. condonianus, and L. exilicauda) and Hesperoleucus were provided by Gerald R. Smith and Douglas W. Nelson of the University of Michigan. Older published Potassium-Argon radioisotopic ages presented herein were recalibrated using the IUGS constants following the method of Dalrymple (1979). Older published 40Ar/39Ar ages were recalibrated relative to the Fish Canyon Tuff sanidine interlaboratory standard at 28.02 Ma. Divisions and

PALUDICOLA, VOL. 7, NO. 4, 2010 138
boundaries of the North American Land Mammal ages follow Tedford et al. (2004).
Abbreviations are as follows: ap, greatest anteroposterior length; trgs, greatest oblique transverse width of pharyngeal tooth grinding surface; HL, head length; ht, greatest height of pharyngeal tooth above pharyngeal bone; Ma, millions of years before present; N, number of specimens; OR, observed range; SL, standard length; TL, total length; tr, greatest transverse width.
GEOLOGIC SETTING
The Wassuk Group is comprised of, from oldest to youngest, the Aldrich Station Formation, the Coal Valley Formation, and the Morgan Ranch Formation (Axelrod, 1956; Goliad and Stewart, 1984). The group has been studied extensively because it has yielded superposed faunas and floras of middle to late Miocene age (Axelrod, 1956; Evernden and James, 1964; Evernden et al., 1964; Gilbert and Reynolds, 1973; Golia and Stewart, 1984; Stewart and Reynolds, 1987; Schorn and Shelton, 1991; Wolfe et al., 1997; Kelly, 1998; Perkins et al., 1998; Kelly and Secord, 2009). Axelrod (1956) first described the group and Gilbert and Reynolds (1973) mapped the group and further defined the formational boundaries. The group was deposited in a large, northwest-trending basin that extended from about 5 km south of Coal Valley northwestward to the Mount Wilson area in the southern Singatze Range, or about 55 km north of Coal Valley (Gilbert and Reynolds, 1973; Golia and Stewart, 1984). During the deposition of the Aldrich Station Formation, the basin contained a large circumneutral, eutrophic lake with paludal conditions at the margin (Golia and Stewart, 1984). Wolf et al. (1997) provided evidence that the paleoaltitude during the deposition of the Aldrich Station Formation was at about its present altitude, or about 1.5 – 2 km above sea level. Gilbert and Reynolds (1973) reassigned the uppermost portion of the Aldrich Station Formation of Axelrod (1956) to the base of the Coal Valley Formation. The Aldrich Station Formation is comprised of carbonaceous mudstone and siltstone, diatomaceous shale, sandstone, and pebble conglomerate (Axelrod, 1956; Gilbert and Reynolds, 1973). The type section of the Aldrich Station Formation occurs in Coal Valley, about 45 km southeast of the Mickey Canyon section, where it is about 755 m thick (Gilbert and Reynolds, 1973). In the Mickey Canyon section, the Aldrich Station Formation thins to about 370 m, mainly at the expense of the diatomaceous claystones and mudstones of middle portion of the formation (Gilbert and Reynolds, 1973). The age of the Aldrich Station Formation has been determined by biostratigraphic correlation of the flora
and a series of K-Ar dates on tuffs above and within the formation (Evernden and James, 1964; Evernden et al., 1964; Gilbert and Reynolds, 1973). Radiometric dates of tuffs within the Aldrich Station Formation range from about 11.5 to 12.8 Ma and tuffs within the overlying Coal Valley Formation range from about 11.5 to 6.98 Ma (Gilbert and Reynolds, 1973; Tedford et al., 2004; Kelly and Secord, 2009). In the vicinity of Mickey Canyon, a K-Ar radiometric date of 11.45 ± .25 Ma (corrected) was recorded by Gilbert and Reynolds (1973) from a tuff within the upper portion of the Aldrich Station Formation section. Based on the tephrochronologic correlation of silicic fallout tuffs, Perkins et al. (1998) provided ages for the Aldrich Hill 1 ash and Aldrich Hill 2 ash within the Aldrich Station Formation in the type section at Coal Valley of 12.07 ± 0.10 and 12.01 ± 0.10 Ma, respectively. The fish level in the Mickey Canyon section occurs near the base of the formation, well below the dated tuff. The shale of the fish level in the Mickey Canyon section contains large amounts of charcoal, similar to that found in the shales of the lower levels of the type section of the Aldrich Station Formation in Coal Valley. Also, the lower levels in the Mickey Canyon section contain coarse sandstone beds, similar to those in the lower levels of the type section in Coal Valley. Schorn and Shelton (1991) regard the Mickey Canyon Spring Flora as a correlative of the early Clarendonian Aldrich Station Flora because they contain many of the same species. Considering that the middle portion of the Aldrich Station Formation in the Mickey Canyon section is much reduced and similar lithologies are observed in the lower levels of the Mickey Canyon section and the type section in Coal Valley (Axelrod, 1956; Gilbert and Reynolds, 1973) along with the floristic correlations, it is probable that the lower half of the Mickey Canyon section is a correlative of the lower levels in the type section. Thus, the fish level in the Mickey Canyon section is most likely about 12.5 to 12.0 Ma in age, equivalent to the early Clarendonian North American Land Mammal age (Cl1 of Tedford et al., 2004). The stratigraphy of the beds at the fish localities include the following, from highest to lowest: 1) a conspicuous ledge forming, yellowish-buff medium-grained sandstone (0.3 – 0.5 m thick); 2) a thin, light gray shale containing leaf impressions (0.1 – 0.2 m thick); 3) a thin, dark brown mudstone (0.05 m thick); 4) a greenish-buff, fine to pebbly sandstone (0.04 – 0.06 m thick); 5) a thin, light gray to greenish-brown laminated shale containing several bedding planes with numerous articulated fish and occasional leaf impressions (0.05 – 0.07 m thick); 6) a thin, dark brown, mudstone (0.05 m thick); 7) a resistant, dark reddish-brown, pebbly conglomerate (0.03 – 0.05 m thick); and 8) a thick sequence of yellowish-buff to

KELLY—MICOENE CHUB FROM NEVADA 139
greenish-brown sandstones with minor shale beds (43 – 45 m thick). The fish producing shale bed may represent mass-mortality episodes of monospecific schools over a short geologic time scale because it is very thin, extends laterally for some distance with the fish fossils abundantly represented only on a few individual bedding planes within the shale bed, has only yielded a single species of fish with the majority of the individuals having a similar SL, and most of the fish fossils are articulated skeletons. The taphonomy of the fish producing shale bed of the Aldrich Station Formation appears to be very similar to that described by Bell et al. (1989) for the mass-mortality layers of fossil Stickleback fish of the Miocene Truckee Formation. However, further investigation is required to confidently determine whether this hypothesis is correct. The second overlying shale bed yielded the majority of the plant specimens comprising the Mickey Canyon Spring Flora with fish remains generally absent or rarely represented by disassociated bone fragments.
SYSTEMATIC PALEONTOLOGY
Class Actinopterygii Order Cypriniformes Family Cyprinidae
Lavinia Baird and Girard, 1854 Lavinia lugaskii new species
Figures 1-6, Tables 1-2
Holotype―LACM 154390, complete skeleton (part and counterpart) on shale matrix.
Referred Specimens―From locality LACM 7743: complete skeleton, LACM 154333; partial skeletons, LACM 154334, 154335, 154336, 154337, 154338, 154339, 154340, 154341, 154342, 154343, 154344, 154345, 154346, 154347, 154348, 154349, 154350, 154351, 154352, 154353, 154354, 154355, 154356, 154357, 154358, 154359, 154360, 154361, 154362, 154363, 154364, 154365, 154366, 154367, 154368, 154369, 154370, 154371, 154372, 154373, 154374, 154375, 154376, 154377, 154378, 154379, 154380, 154381, 154382, 154383, 154384, 154385, 154397, 154459, 154461, 154462; partial skull, LACM 154460. From locality LACM 7744: partial skeletons, LACM 154386, 154387. From locality LACM 7745: complete skeleton, LACM 154389; partial skeletons, LACM 154388, 154391, 154392, 154393, 154394, 154395, 154396; opercle, LACM 154463. From locality UCMP PA624: partial skeleton, UCMP 141400.
Diagnosis―Lavinia lugaskii differs from Lavinia hibbardi (Smith, 1975) by having the following: 1) most anterior pharyngeal tooth of major row better developed (relatively larger) and always with well-
developed rugose grinding surface; 2) pharyngeal teeth of major row relatively more expanded transversely (deeper) and terminal tips more rounded (not sharply pointed) and more hooked (especially posterior two teeth); 3) pharyngeal tooth of minor row larger, longer, and with better developed, slightly striated grinding surface; 4) maxilla with much less developed mid-dorsal process (relatively much shorter) and ventral margin slightly convex (not nearly straight); and 5) lateral process of frontal positioned more posteriorly. It differs from Lavinia condonianus (Smith, 1975) by having the following: 1) most anterior pharyngeal tooth of major row much better developed (not peg-like); 2) all pharyngeal teeth of the major row notably compressed anteroposteriorly, moderately hooked at their tips, and have well-developed rugose grinding surfaces; 3) pharyngeal tooth of minor row relatively much larger and better developed (not rudimentary) with a slightly striated grinding surface; and 4) length of anterior arm of pharyngeal arch (portion anterior to first tooth of major row) significantly longer than length of major tooth row. It differs from extant Lavinia exilicauda Baird and Girard, 1854, by having the following: 1) pharyngeal tooth formula 1/5-5/1; 2) grinding surfaces of pharyngeal teeth relatively wider with much better developed, wavy linear, rugose ridges; 3) terminal tips of pharyngeal teeth of major row more rounded (not as sharply pointed) and less acutely hooked; 4) length of anterior arm of pharyngeal arch (portion anterior to first tooth of major row) significantly longer than major tooth row; 5) anterolateral foramen positioned more anteriorly on gnathal ramus; 6) mid-dorsal process of dentary relatively wider; 7) lateral process of frontal positioned more posteriorly; 8) relative head length larger (HL divisible into TL about 4.3-4.5 times); 9) dorsal fin origin not positioned as far posteriorly of pelvic fin origin; 10) anal fin origin positioned posterior to posterior margin of dorsal fin base; and 11) lower dorsal and anal fin ray counts. It differs from Hesperoleucus (= ?Lavinia) symmetricus (Baird and Girard, 1854) by having the following: 1) pharyngeal tooth formula 1/5-5/1; 2) pharyngeal teeth with much better developed rugose grinding surfaces and less sharply pointed terminal tips (more rounded); 3) length of anterior arm of pharyngeal arch (portion anterior to first tooth of major row) significantly longer than major tooth row; 4) relative head length slightly larger (HL divisible into TL about 4.3-4.5 times); 5) dorsal fin origin positioned slightly less posterior of pelvic fin origin; 6) anal fin origin positioned posterior to posterior edge of dorsal fin base; 7) pectoral fin ray count lower; and 8) much larger body size.
Type Locality―LACM 7745, Aldrich Station Formation, Mickey Canyon area, northwest flank of the Pine Groove Hills, Lyon County, Nevada.

PALUDICOLA, VOL. 7, NO. 4, 2010 140
Age―Middle Miocene (early Clarendonian North American Land Mammal age), about 12.5 – 12.0 Ma.
Etymology―Named in honor of the late Thomas P. Lugaski of the W. M. Keck Earth Science and Mineral Engineering Museum, University of Nevada, Reno, in recognition of his contributions to our knowledge of the Miocene chubs of Nevada.
Description―All of the specimens are preserved on the shale matrix bedding planes, resulting in most being compressed laterally and a few being compressed dorsoventrally. Most of the specimens are partial skeletons, but three specimens (holotype, LACM 154390, and referred specimens, LACM 154333 and 154389) are represented by nearly complete skeletons (Figures 1A-C, 2A-B). The holotype is particularly well preserved, even showing the scale impressions along the dorsal and ventral aspects of the body outline. The general morphology of the head, body and fins is typically chub-like (Figures 1-2). The head length is divisible into the TL about 4.3 to 4.5 times. As compared with other middle Miocene chubs from Nevada, the body depth is moderately deep (maximum body depth divisible into TL about 4.2 – 4.3 times). The largest specimen (LACM 154341) is incomplete, missing the anterior part of the head, posterior end of the caudal peduncle, and most of the caudal fin, but is estimated to have had a TL of about 290 mm. Based on comparative ratios of the three complete skeletons, the size range of the TLs of the sample appears to be from about 140 to 290 mm, with the majority being between about 155 - 180 mm. The caudal peduncle is moderately robust with the least depth divisible into the TL about 13.7 - 14.2 times. The vertebrae count, not including the weberian apparatus, is 36. Morphometric and meristic data for the holotype and referred skeletons are presented in Table 1. Pharyngeal teeth are present in 29 specimens, including the three most complete skeletons. The most complete pharyngeal in the sample is present on the partial, disarticulated skeleton of LACM 154397 (estimated SL undeterminable). In this specimen, the tooth bearing portion of the left pharyngeal was split away from the rest of the pharyngeal arch, but after preparation was precisely placed back into position. There are five teeth in the major row and one tooth in the minor row (Figure 3A-B). The first (most anterior) tooth of the major row exhibits the following characters: the smallest and shortest tooth of the major row; the tooth is moderately compressed anteroposteriorly; the grinding surface is elongated and contains rugose, wavy linear ridges; and the terminal tip is moderately pointed and slightly hooked. It is basically a slightly smaller version of the second tooth of the major row. The second tooth of the major row exhibits the following characters: the tooth base is
ovoid and moderately short; the grinding surface is moderately compressed anteroposteriorly and elongated transversely with rugose ridges and two small cuspules along each margin near the tip; and the tip is rounded and only slightly hooked. The third tooth of the major row differs from the second tooth by having the following: taller and larger; the terminal tip is slightly more rounded; and the grinding surface is slightly more elongated transversely and wider anteroposteriorly with extensive rugose ridges, but lacks the two small cuspules along the anterior margins. The fourth tooth of the major row differs from the third tooth by having the following: slightly greater in height; the grinding surface is slightly more elongated and slightly more compressed anteroposteriorly; and the terminal tip is slightly less rounded and more hooked. The fifth tooth of the major row is very similar to the forth tooth, but is slightly smaller and shorter, and is on an elevated platform of the pharyngeal. The single tooth of the minor row is tall and slightly compressed anteroposteriorly with a weakly developed grinding surface exhibiting slight striations and a slightly hooked tip. The measurements (in mm) for the teeth of the major row are (from anterior to posterior): first tooth, ap = 0.49, tr = 0.75, trgs = 0.70, ht = 1.51; second tooth, ap = 0.54, tr = 0.87, trgs = 1.08, ht = 1.90; third tooth, ap = 0.59, tr = 1.00, trgs = 1.20 , ht = 2.25; forth tooth, ap = 0.49, tr = 0.90, trgs = 1.13, ht = 2.54; and fifth tooth, ap = 0.44, tr = 0.59, trgs = 0.87, ht = 2.28. The measurements (in mm) for the minor row tooth are; ap = 0.21, tr = 0.26, trgs = 0.24, ht = 1.02. The length of the major tooth row is 4.4 mm. The anterior limb of the pharyngeal arch is elongated with a greatest length of 10.7 mm along the axis of the major tooth row and a length anterior to the first tooth of the major row of 7.0 mm. The dorsal limb is recurved posterolaterally with a total length of about 7.5 mm. The length from the tip of the dorsal limb to the tip of the anterior limb is 9.5 mm. The tooth formula for the right pharyngeal is 5/1. On LACM 154388 (estimated SL ~136 mm), a partial left pharyngeal is present with six teeth, but the dorsal and anterior limbs are missing (Figure 4A-C). One smaller tooth is positioned near and posterior to the base of the second tooth of the major row and probably represents a replacement tooth for the major row. A single tooth is also present slightly ventral to and between the teeth of the major row, but the position of its base and its morphology clearly indicate that it belongs to a minor row. Apparently, there were five teeth in the major row and one tooth in the minor row. The grinding surfaces of all of the teeth of the major row exhibit well-developed, rugose, linear ridges, whereas the grinding surface of the minor tooth is less developed with slight striations. The measurements (in mm) for the teeth of the major row

KELLY—MICOENE CHUB FROM NEVADA 141
FIGURE 1. Lavinia lugaskii. A-B, skeleton of holotype (LACM 154390) showing part and counter part. C, skeleton of referred specimen (LACM 154389). All lateral views. Scale = 10 mm.

PALUDICOLA, VOL. 7, NO. 4, 2010 142
FIGURE 2. Lavinia lugaskii. A-B, skeleton showing part and counterpart, LACM 154333. C, partial skeleton showing detail of well forked tail, LACM 154363. All lateral views. Scales = 10 mm.

KELLY—MICOENE CHUB FROM NEVADA 143
are (from anterior to posterior): first tooth, ap = 0.55, tr = 0.75, trgs = 1.05, ht = 1.80; second tooth, ap = 0.59, tr = 1.02, trgs = 1.18 , ht = 2.20; third tooth (?replacement tooth), ap = 0.50, tr = 0.74, trgs = 1.03, ht = 1.59; forth tooth, ap = 0.50, tr = 0.89, trgs = 1.09, ht = 2.49; and fifth tooth, ap = 0.44, tr = 0.55, trgs = 0.80, ht = 0.99. The measurements (in mm) of the minor row tooth are; ap = 0.19, tr = 0.25, trgs = 0.11, ht = 1.00. Little can be said of the morphology of the pharyngeal bone of LACM 154388 because it is compressed and damaged, with the anterior and dorsal limbs, and ventral-lateral portions missing. On LACM 154393 (estimated SL ~132 mm), some of the bones of the head are disarticulated from the skeleton and imbedded in the surrounding matrix, including the pharyngeal arches (Figure 4D-E). The partial right pharyngeal of LACM 154393 has the mesial-ventral aspect exposed (Figure 4D). It possesses the two most anterior teeth of the major row, the bases and impressions in the matrix of the crowns of the third and forth teeth of the major row, the base of the fifth tooth of the major row, and the minor row tooth. These teeth exhibit the same morphology of those of LACM 154388 and 154397, and confirm that the right pharyngeal tooth formula is 5/1. The posterior teeth of the major row are on a moderately elevated platform. The anterior limb tapers anteriorly, is slightly downturned ventrally, and exhibits a marked narrowing below the anterior base of the most anterior tooth of the major row. The length of the anterior arm (8.0 mm, portion anterior to first tooth of major row) is significantly longer than the major tooth row base length (4.5 mm). The dorsal limb of the right pharyngeal is not visible because it extends into the matrix. No further attempt to remove the matrix from this specimen was done because the matrix is much harder than the bone and it would result in destruction of the bone. The alar margin is smoothly rounded except for a noticeable indentation anteriorly due to the narrowing at the base of the anterior limb. The greatest length of the right pharyngeal in the axis of the tooth row is 12.5 mm. The partial left pharyngeal of LACM 154393 has the mesial aspect exposed (Figure 4E). It possesses the bases of the five pharyngeal teeth of the major row and impressions in the matrix of the two most anterior teeth of the major row. The minor row tooth is not visible on the left pharyngeal due to the matrix and no attempt to remove the matrix was made because of the fragile condition of the bone. However, it is assumed that it was present since one occurs on the corresponding right pharyngeal and a single minor tooth is present on both left pharyngeals of LACM 154388 and 154397. The major tooth row length measured at the base of the teeth is 4.5 mm. The tip of the anterior limb of the left pharyngeal is missing with
a broken length of 4.6 mm. The dorsal limb is missing a portion of the bone in the center, but the outline of the missing portion is clearly visible because of the impression left in the matrix. The dorsal limb is recurved posterolaterally with a total length of 6.5 mm and a length from the middle of the major tooth row to the tip of 5.3 mm. The length from the tip of the dorsal limb to the broken tip of the anterior limb is 10.1 mm. On LACM 154395 (estimated SL ~121 mm), a partial right pharyngeal and an isolated tooth are present. The right pharyngeal is split in half exhibiting the hollow bases of the teeth in the part and counter part. There are five teeth in the major row and one tooth in the minor row (Figure 5D). The length of the major tooth row is 3.8 mm. On LACM 154385 (estimated SL ~136 mm), a partial right pharyngeal is present with five teeth present in the major row. On LACM 154396 (estimated SL ~125 mm), partial left and right pharyngeal arches are visible. The left pharyngeal is missing the anterior end of the anterior limb and has the following present; the first (most anterior) tooth of the major row, the base and impression of the second tooth of the major row, the third tooth of the major row, and the base and impression of the fourth tooth of the major row, the base of the fifth tooth of the major row, and the base and impression of a single minor row tooth. The right pharyngeal is also missing most of the anterior limb and exhibits the most anterior tooth of the major row, the base and impression of the second tooth of the major row, and the bases of the third through fifth teeth of major row. LACM 154395, 154385, and 154396 confirm the pharyngeal tooth formula at 1/5-5/1. The teeth of these specimens exhibit the same proportions and morphology as the corresponding teeth in those of LACM 154388, 154393 and 154397, including the rugose, wavy linear ridges on the grinding surfaces. On LACM 154379 (estimated SL ~114 mm), a partial left pharyngeal is present (Figure 5E). The left pharyngeal is damaged and missing a portion of the alar margin and the dorsal limb. This specimen exhibits the elongated anterior limb and the five teeth of the major row. The first two teeth of the major row are slightly broken away from their bases. The length of the major tooth row is about 4.0 mm, the greatest length of the anterior limb in the axis of the tooth row is 10.8 mm, and the length of the anterior limb anterior to the first tooth of the major row is 6.8 mm. In addition to those described above, pharyngeal teeth are also present on the compressed bones of the head or in the matrix near the head on 22 other specimens (LACM 154333, 154335, 154340, 154343, 154344, 154350, 154355, 154358, 154361, 154373, 154374, 154375, 154376, 154378, 154380, 154381, 154382, 154383, 154384, 154389, 154390, 154394)

PALUDICOLA, VOL. 7, NO. 4, 2010 144
TABLE 1. Morphometric and meristic data for the holotype (LACM 154390) and referred specimens (LACM 154333, 154389) of Lavinia lugaskii. Thousandths of SL are included in parentheses. Holotype Character/measurement LACM 154390 LACM 154333 LACM 154389 Fin ray count
Dorsal 8 8 8 Pectoral 9 10 9 Pelvic 9 10 9 Anal 8 8 8
Caudal 19 19 19 Vertebrae count 36 36 36 SL 132.0 mm 115.5 mm 138.0 mm TL 177.2 mm 153.0 mm ~181 mm Head length 41.1 mm (311) 33.8 mm (285) 42.0 mm (302) Head depth 33.6 mm (255) 25.7 mm (225) ~30 mm (~217) Maximum body depth 41.5 mm (314) 36.0 mm (311) 42.0 mm (304) Fins
Dorsal fin length 30.0 mm (227) ~26 mm (~225) 30.5 mm (221) Dorsal fin base length 15.3 mm (116) - 16.3 mm (118) Pectoral fin length ~29 mm (~220) 23.0 mm (199) ~25 mm (~181) Pectoral fin base length 8.0 mm (60) 5.0 mm (43) ~5.6 mm (~41) Pelvic fin length ~25 mm (~189) 21.7mm (188) ~23.5 mm (~170) Pelvic fin base length 5.8 mm (44) 4.0 mm (35) ~5.5 mm (~40) Anal fin length 26.6 mm (202) 22.5 mm (195) ~26 mm (~188) Anal fin base length 15.3 mm (116) 12.6 mm (109) 16.9 mm (122) Caudal fin length 45.2 mm (342) 37.5 mm (325) ~43 mm (~316)
Minimum caudal fin depth ~21.2 mm (~161) ~19.8 mm (~142) ~24 mm (~174) Widest caudal fin depth 44.2 mm (335) 40.6 mm (356) -
Snout to dorsal origin length 76.0 mm (576) 60.6 mm (525) 75.6 mm (548) Snout to occupit length 40.5 mm (307) 30.2 mm (261) 36.6 mm (265) Snout to pectoral origin length 41.0 mm (311) 31.5 mm (273) 39.7 mm (288) Snout to pelvic origin length 72.0 mm (545) 63.6 mm (551) 73.5 mm (533) Snout to anal origin length 95.0 mm (720) 85.2 mm (738) 101.6 mm (736) Pectoral origin to pelvic origin length 34.0 mm (258) 32.5 mm (281) 35.5 mm (257) Pectoral origin to anal origin length 58.0 mm (439) 58.5 mm (506) 63.1 mm (457) Pectoral origin to dorsal origin length 45.7 mm (346) 33.1 mm (287) 41.6 mm (301) Pectoral origin to caudal base length 98.0 mm (742) 91.5 mm (792) 102.1 mm (740) Dorsal origin to occupit length 39.2 mm (297) 33.8 mm (292) 43.0 mm (312) Dorsal origin to anal origin length 23.5 mm (178) ~26 mm (~225) 24.3 mm (176) Pelvic origin to dorsal posterior base length 21.2 mm (161) 18.8 mm (142) 20.5 mm (148) Pelvic origin to anal origin length 28.0 mm (212) ~31.3 mm (~271) 30.0 mm (217) Pelvic origin to caudal base length 62.0 mm (470) 66.0 mm (571) 65.5 mm (475) Pelvic origin to greatest caudal length 105.0 mm (795) 104.0 mm (900) 111.0 mm (804) Anal origin to caudal base length 39.5 mm (299) 37.0 mm (320) 40.5 mm (293) Length of caudal pedicle
Dorsal fin posterior base to caudal base 43.0 mm (326) 42.0 mm (364) 42.9 mm (311) Anal posterior base to caudal base 24.0 mm (182) 21.8 mm (189) 24.9 mm (180)
Caudal peduncle least depth 12.5 mm (71) ~11 mm (~95) 13.2 mm (96) Relative positions Dorsal fin origin to pelvic fin origin 8 mm posterior (60) 5.5 mm posterior (50) 6.0 mm posterior (43) Anal fin origin to posterior dorsal fin base 3 mm posterior (20) 2 mm posterior (20) 3.7 mm posterior (27) TL/ head length 4.31 4.53 4.31 TL/caudal peduncle least depth 14.2 ~13.9 ~13.7

KELLY—MICOENE CHUB FROM NEVADA 145
FIGURE 3. Lavinia lugaskii. Left pharyngeal teeth of LACM 154397. A, anterodorsal view: 1, first (most anterior) tooth of the major row; 2, second tooth of major row; 3, third tooth of major row; 4, fourth tooth of major row; 5, fifth tooth of major row; 6, minor row tooth. B, mesial view, tooth numbers correspond to positions of those in 3A. Scales = 0.5 mm. ____________________________________________________________________________________________________________
including the holotype (Figure 5A-C). The teeth of these specimens agree in morphology with the corresponding teeth of those described above, including the rugose grinding surfaces. Due to compression and some probable postmortem deterioration, the cranial and jaw bones are often broken apart, overlapping or fused together in the specimens, making it difficult to distinguish the morphology of individual bones. In addition, because the shale matrix was often split along the bedding plane to expose the fossil, many of the specimens have the bones split apart with portions on each half of the resulting part and counterpart. However in some cases, certain bones are disarticulated from the skeleton or complete enough to allow a description of their morphology.
The premaxillae are best preserved in LACM 154350 (estimated SL ~123mm) and LACM 154344 (estimated SL ~ 96 mm), wherein only their lateral aspects are exposed. The premaxilla extends anteriorly from the anteroventral margin of the maxilla to the tip of the snout and then turns posterodorsally at an acute angle as a smaller dorsal projection, giving the tip of the snout a somewhat pointed appearance. Measurements of the premaxillae of LACM 154350 and 154344 are, respectively: lengths from the anterior tip to the border of the maxilla = 8.4 and 5.8 mm; lengths of the dorsal posterior projection = 3.8 and 2.5 mm; and the greatest depths along the portion bordering the orifice = 2.4 and 2.1 mm. The maxillae are best preserved in LACM 154462 (estimated SL = 137 mm), 154459 (estimated SL = 134 mm), and 154350. The maxilla of LACM 154462 is

PALUDICOLA, VOL. 7, NO. 4, 2010 146
almost complete with its lateral aspect exposed, only missing very small portions along the posterior edges of the posterior process. It is elongated anteroposteriorly (9.7 mm), with the greatest depth occurring across the body and dorsal process (4.2 mm), and gradually tapers anteriorly to a narrow (~ 1 mm) neck. The ventral margin is slightly convex. The dorsal process has a relatively wide base with a moderate height of about 2.5 mm. A relatively well-developed ridge for the insertion of the palatine ligament is present. The posterior process is moderately well developed and flairs out from a relatively narrow pedicel. The head of the maxilla is not visible because it extends mesiodorsally into the matrix. The partial maxilla of LACM 154350 has its lateral aspect exposed and exhibits the same morphology as that of LACM 154462. The measurements of LACM 154350 are total length ~8-9 mm, greatest depth across the body and dorsal process = 3.2 mm, the width of anterior neck between the dorsal process and head = 0.07 mm, and height of the base of the dorsal process = 2.5 mm. The partial maxilla of LACM 154459 has its mesial aspect exposed, including the head of the maxilla and part of the anteromesial (premaxillary) process, but the dorsal process and part of the posterior process are broken away. The head has a rounded outline with the anteromesial process extending ventrally from the head. The neck connecting the head to the anterior body of the maxilla is very short, so that the head is positioned close to the anterodorsal aspect of the maxilla. Composite lateral and mesial views of the maxilla outline based on LACM 154462, 154459, and 154350 are presented in Figure 6A-B. The dentaries are best preserved in LACM 154390 (holotype), 154350, 154391 (estimated SL ~145 mm), and LACM 154462. The coronoid process of the dentary is centrally positioned and well developed with a rounded dorsal arch. The gnathal ridge is flaired laterally and the anterior end is mesially curved. The ventral edge is relatively flat with only a slight ventral deviation near the anterior tip. The anterolateral foramen is positioned well anterior to the midpoint of the gnathal ramus. Seven lateral pores are present along the lateroventral aspect of the dentary. The mechelian groove terminates anteriorly on the gnathal ramus at a level that is just posterior to the caudal end of the gnathal ridge. The measurements (in mm) of the dentaries of LACM 154390, 154350, 154391, and LACM 154462 are, respectively: lengths from the anterior tip to posterior tip, 12.7, ~12.9, 13.3, and 13.0; lengths from the anterior tip to the top of the coronoid process, 9.3, 8.0, 9.9, and 8.8; lengths from the anterior tip to the anterior base of coronoid process, 6.3, 6.0, 6.9, and 6.7; lengths from the top of the coronoid process to the posterior tip, ~7.4, 7.6, ~8.3, and 8.1; depths at the anterior tips (symphyses), 1.5,
1.4, 1.6, and 1.4; depths just anterior to the anterior base of coronoid process, 2.5, 2.3, 3.1, and 2.6; widths of the base of the coronoid process, 2.7, 2.8, 3.2, and 3.5; and depths from the highest point on the coronoid process to the ventral edge of dentary, 5.9, 6.0, 6.0, and 5.7. When the mouth is closed it forms an oblique angle of about 27 degrees relative to the horizontal axis of the head and the maxilla extends posteriorly to the level of the anterior edge of the orbit. A composite lateral view of the dentary outline based on LACM 154390, 154350, 154391, and 154462 is presented in Figure 6C. The preopercle is complete and fairly well preserved in one specimen, LACM 154355 (estimated SL ~94 mm), wherein its mesial surface is exposed. It is shaped in a wide concave arc formed with an angle of 108 degrees and has the following measurements: width from the dorsal tip to the anteroventral tip, 13.0 mm; dorsal limb length from the apex, 7.6 mm; anteroventral limb length from the apex, 7.3 mm; and greatest depth at the apex of the angle between the dorsal and ventral limbs, 13.0 mm. Although many specimens exhibit a partial opercle or partial outline of it in the matrix, the best preserved opercle is a disarticulated specimen (LACM 154463), which consists of almost the entire bone laterally exposed with only a few small areas along the ventral and posterior borders missing. However, the entire outline of the bone can be discerned because there are impressions left in the matrix where the bone is missing. The lateral aspect of the opercle outline is triangular in shape with the anterior articular process extending anterodorsally from the body as a narrow, elongated projection (Figure 6D). Overall, it is very similar in shape to those of other species of Pliocene Lavinia (see Smith, 1975). Measurements (in mm) for LACM 154463 are as follows (opercle measurements and terminology follow Miranda and Escala, 2005); distance from the tip of the articular process to the articular facet = 4.0, distance from the tip of the articular process to the inferior angle = 12.2, distance from the tip of the articular process to the superior angle = 8.7, distance from superior angle to inferior angle = 18.0, greatest distance between posterior angle and anterior edge = 11.8, and posterior angle height = 7.3. Partial frontals are common on many of the specimens, but they are usually incomplete or badly crushed. The best preserved frontals are on LACM 154459 and 154460 (a dorsoventrally preserved partial cranium, estimated SL undeterminable). On LACM 154459, the dorsal aspects of the frontals are exposed, whereas on LACM 154460 the ventral aspects are exposed. The dorsal and ventral outlines of the left frontals are presented in Figure 6E-F. The frontal of L. lugaskii is morphologically similar to those of L.

KELLY—MICOENE CHUB FROM NEVADA 147
FIGURE 4. Lavinia lugaskii. A, partial skeleton showing location of partial pharyngeal arch, lateral view, LACM 154388, scale = 5 mm. B, enlarged area of A showing partial pharyngeal arch and teeth, anterodorsal view, scale = 0.5mm: 1, replacement tooth of major row; 2-5, major row teeth; 6, minor row tooth. C, close up of rugrose linear grinding surface of fourth tooth of major row, scale = 0.5 mm. D, partial right pharyngeal with first and second teeth of major row, bases and impressions in matrix of third and forth teeth of major row, base of fifth tooth of major row, and minor row tooth, mesial-ventral view, LACM 154393. E, partial left pharyngeal with impressions in matrix of two most anterior teeth of major row and bases of all five teeth of major row, mesial view, LACM 154393. Scales for D and E = 5 mm.

PALUDICOLA, VOL. 7, NO. 4, 2010 148
hibbardi (see Smith, 1975) and extant Lavinia, but the lateral process is positioned more posteriorly. For example, in L. hibbardi and extant Lavinia the tip of the lateral process is positioned posteriorly from the anterior edge of the frontal at about 55-58% of the total length, whereas in L. lugaskii it is positioned posteriorly at about 68% of the total length. Measurements (in mm) of LACM 154459 and 154460, respectively, are as follows; ap = 15.4 and 16.2, tr = 6.9 and 7.0, and distance between the anterior edge to tip of the lateral process = 10.4 and 11.0. In one specimen (LACM 154361, estimated SL ~133 mm), the right cleithrum is disarticulated from the body and positioned on the matrix near the pelvic fin, exposing its mesial aspect. A distinct ridge is present that extends down from the dorsal tip of the vertical wing to the base of the horizontal wing. The measurements of LACM 154361 are follows (using those defined by Miranda and Escala, 2005): the vertical wing length = 11.5 mm; the horizontal wing length = 11.7 mm; the chord length (the widest length between the dorsal tip and the mesocoracoid articulation crest of the horizontal wing) = 19.1 mm; the external margin length (chord length minus the mesocoracoid articulation crest of the horizontal wing) = 18.0 mm; and the posterior angle within the wings = 97 degrees. The pectoral and pelvic fins are of moderate size, while the dorsal, anal and caudal fins are relatively large (Table 2). The fin ray counts are as follows: dorsal, 8 (N = 30); pectoral, usually 9, with one at 10 (N = 18); pelvic, usually 9, with one at 10 (N = 27); anal 8 (N = 27); and caudal, usually 19, with one at 20 (N = 19). The dorsal fin is positioned posteriorly on the body with its origin posterior to the pelvic fin origin. The angle formed from the origin of the pelvic fin to the origin of the dorsal fin is 83 - 85 degrees posterior. The posterior portion of the dorsal fin extends beyond the origin of the anal fin (Figure 1). However, the anal fin origin is posterior to the dorsal fin posterior base. The caudal fin is rather deeply forked (Figure 2C). There are eight dorsal and eight ventral procurrent rays in the caudal fin. Coburn and Cavender (1992) regarded an equal number of dorsal and ventral procurrent caudal fin rays to be a derived condition, often seen in members of the Western North American chub group.
Discussion―The Aldrich Station chub exhibits the following suite of characters: 1) moderate size (TL ~140 – 290 mm); 2) the HL is divisible into the TL about 4.31 – 4.53 times; 3) the body is relatively deep (body depth 304 - 314 thousandths of SL); 4) the pharyngeal tooth formula 1/5 - 5/1; 5) all of the pharyngeal teeth of the major row are compressed anteroposteriorly and exhibit moderately rounded and
hooked tips, and transversely elongated grinding surfaces with distinctive rugose, wavy linear ridges; 6) the minor row tooth is moderately tall and anteroposteriorly compressed, the grinding surface is slightly developed with faint striations, and the tip is slightly hooked; 7) the anterior arm of the pharyngeal arch (portion anterior to first tooth of major row) is significantly longer than the major tooth row length; 8) the dorsal process of the maxilla is moderately developed and centrally positioned; 9) the dentary has a mesially curved anterior tip, centrally positioned dorsal process, anteriorly positioned anterolateral foramen, laterally flaired gnathal ridge, and eight lateral pores; 10) the lateral process of the frontal is positioned posteriorly from the anterior margin at about 68% of the total length of the frontal; 11) the fin ray counts are dorsal 8, pectoral usually 9 (rarely 10), pelvic usually 9 (rarely 10), anal 8, and caudal usually 19 (rarely 20); 12) the dorsal, anal and caudal fins are relatively large; 13) the caudal fin is deeply forked; 14) the dorsal fin origin is positioned posterior to the pelvic fin origin; 15) the anal fin origin is positioned posterior to the dorsal fin posterior base; and 16) the caudal peduncle is moderate in depth (TL/caudal peduncle least depth = 13.7 - 14.2). The Aldrich Station chub is morphologically most similar to Pliocene Lavinia hibbardi, including the synapomorphic shared character state of well-developed rugose grinding surfaces on the pharyngeal teeth (see Smith et al., 1982, 2000, 2002) along with its dentary morphology that includes a mesially curved anterior tip of the gnathal ramus, a centrally positioned dorsal process, laterally flaired gnathal ridge, and eight lateral pores. These shared characters indicate that the Aldrich Station chub is referable of Lavinia and it is herein assigned to a new species, L. lugaskii. Smith (1975) described the genus Idadon from the Glenns Ferry Formation, Idaho, and assigned two species to it, the type species I. condonianus (Cope, 1883) and I. hibbardi. Subsequent investigators (e.g., Smith et al., 1982; Smith and Miller, 1985) noted that Idadon exhibited similarities, especially the rugose or striated grinding surfaces of the pharyngeal teeth, to extant Lavinia and other Pliocene representatives of Lavinia. Smith et al. (2000) provided evidence that Idadon should be regarded as a junior synonym of Lavinia and referred I. hibbardi to Lavinia (Idadon) hibbardi, and Smith et al. (2002) provided further evidence in support of this synonymy. Smith et al. (2002) regard Mylopharodon Ayres, 1855, as the closest sister taxon to Lavinia and Hesperoleucus Snyder, 1913. Lavinia lugaskii is easily distinguished from all species of Mylopharodon by the morphology of its pharyngeal teeth (not molariform). Undetermined species of Lavinia have also been recorded from

KELLY—MICOENE CHUB FROM NEVADA 149
earliest Pliocene (latest Hemphillian) sediments in the Madeline Plains, California, earliest Pliocene (latest Hemphillian) sediments in Cache Valley, Utah, the early to late Pliocene (early to late Blancan) Sunrise Pass Formation, Nevada, the late Pliocene (late Blancan) Magazine Road Local Fauna, Honey Lake, California, and late Pliocene (late Blancan) sediments in the south end of Secret Valley, California (McClellan, 1977; Kelly, 1994; Wagner et al., 1997; Smith et al., 2002; K. Gobalet, per. communication, 2009). Lavinia hibbardi and L. condonianus from the Glenns Ferry Formation are known from large samples of isolated pharyngeal arches and other bones (Smith, 1975). Lavinia hibbardi is characterized by having the following (Smith, 1975; Smith et al., 2000): 1) five pharyngeal teeth in major row and one or two rudimentary teeth in the minor row; 2) all or nearly all of the pharyngeal teeth of the major row exhibit rugose grinding surfaces; 3) the length of the anterior arm of the pharyngeal (portion anterior to the first tooth of the major row) is longer than the major tooth row; 4) the maxilla has a high mid-dorsal process whose depth is more than one-half of the maxilla length, a relatively straight ventral margin, and a posterior (dentary) process that is large and flairs out from a slender pedicel; and 5) the dentary has a gnathal ramus with flaired lateral edges and a large coronoid process that is positioned near the midpoint, and is mesially curved anteriorly. Lavinia condonianus differs from L. hibbardi by having the following (Smith, 1975): 1) the anterior most tooth of the major row of the pharyngeal teeth is peg-like, the second tooth is conic-molariform, and the subsequent teeth are less compressed anteroposteriorly (more round in cross section) and less sharply pointed; 2) the rugose grinding surfaces are less developed and usually restricted to the posterior teeth of the major row; and 3) the length of the anterior arm of the pharyngeal (portion anterior to the first tooth of the major row) is relatively shorter and subequal to the length of the major tooth row. Other samples of fossil Lavinia sp. from the Pliocene have not been formally described or assigned specific status, but Smith et al. (2002) provided a list of the following shared characters that support their assignment to the genus: 1) the pharyngeal teeth have slightly to moderately corrugated grinding surfaces; 2) the dentary has a laterally flaired gnathal ramus, medially positioned coronoid process, mesially curved anterior end, and eight lateral pores; and 3) in the Cache Valley sample, high fin ray counts, like extant Lavinia exilicauda. Extant Lavinia has been well described in the literature (e.g., Baird and Girard, 1854; Snyder, 1913; Fowler, 1924; Murphy, 1945, 1948; Coburn and Cavender, 1992; Smith et al., 1982, 2000, 2002; Moyle, et al., 1995; Moyle, 2002). It is
characterized by having the following: 1) pharyngeal teeth usually 0/5-5/0 (sometimes 0/4-4/0) with the grinding surfaces striated; 2) the anterolateral foramen of the dentary is positioned on the caudal portion of the gnathal ridge (at least halfway back or more) and eight lateral pores are usually present; 3) the dentary has a gnathal ramus with flaired lateral edges and a large coronoid process that is positioned near the midpoint, and is mesially curved anteriorly; 4) the head is relatively small (HL is divisible into TL about 5.3 times); 5) the dorsal fin is positioned posteriorly on the body with its origin well posterior to the pelvic fin origin; 6) the anal fin is long with its origin anterior to or even with the posterior edge of the dorsal fin base; 7) high fin ray counts (dorsal usually 10-13, pectoral 17, pelvic 19, anal usually 11 – 14); 8) a large forked caudal fin; 9) a narrow caudal peduncle; and 10) moderate to large size (maximum SL >350 mm). Hesperoleucus, regarded by some investigators as a junior synonym of Lavinia (e.g., Moyle, 2002; Aguilar and Jones, 2009) and represented by extant H. symmetricus, has been well described in the literature (e.g., Barid and Girard, 1854; Snyder, 1913; Murphy, 1945, 1948; Hopkirk, 1973; Brown et al., 1992; Moyle, et al., 1995; Moyle, 2002; Smith et al., 2002). It is characterized by having the following: 1) pharyngeal teeth 0/5 - 4/0 with striated grinding surfaces; 2) the anterolateral foramen is positioned anteriorly on the gnathal ramus; 3) the dentary has a gnathal ramus with flaired lateral edges and a large coronoid process that is positioned near the midpoint, and is mesially curved anteriorly; 4) the relative size of the head is moderate (HL divisible into TL about 4.8 times); 5) the dorsal fin origin is positioned well posterior to the pelvic fin origin; 6) the dorsal and anal fins are relatively large; 7) the anal fin origin is slightly anterior to the posterior dorsal fin base; 8) the fin ray counts are dorsal usually 7-9, pectoral usually 12-13, pelvic usually 8-9, and anal usually 6-8; 9) a moderately forked caudal fin; 10) a narrow caudal peduncle; and 11) small size (SL = 60.4 mm, OR = 33.1 – 85.5 mm, TL usually <100mm). Lavinia lugaskii can be easily distinguished from L. hibbardi and all other recognized species of Lavinia by the differences listed in the diagnosis above. Lavinia lugaskii can also be easily distinguished from Hesperoleucus symmetricus, the putative closest sister taxon to Lavinia (Smith et al., 2002), by the differences listed in the diagnosis above. Although the samples of Lavinia sp. from the Pliocene have not yet been formally described, L. lugaskii differs from the Cache Valley Lavinia sample from Utah by having lower fin ray counts and better developed rugose grinding surfaces on the pharyngeal teeth. Three other species belonging to the western chub group have been previously described from the

PALUDICOLA, VOL. 7, NO. 4, 2010 150
FIGURE 5. Lavinia lugaskii. A-C, pharyngeal teeth of holotype, LACM 154390, showing morphology and rugose grinding surfaces: A, partial left forth and fifth teeth of major row (removed from matrix along side of B to expose grinding surfaces, tips are missing), dorsal view; B, left second and third teeth of major row, mesial view; and C, three partial teeth of right pharyngeal, posterior dorsal view. D, cross section of pharyngeal teeth, LACM 154395, showing five teeth of major row (numbers 1 – 5) and minor row tooth (number 6). E, partial right pharyngeal, LACM 154379, showing elongated anterior limb and five teeth of major row, mesial view. Numbered tooth positions correspond to those in Figure 3B. Scales for A-D = 0.5 mm, scale for E = 1 mm.

KELLY—MICOENE CHUB FROM NEVADA 151
TABLE 2. Comparative morphometric and meristic data for Lavinia lugaskii and three other Miocene chub species from Nevada. All measurements in parentheses are in thousandths of standard length. Lavinia Gila? Gila Siphateles lugaskii esmeralda 3,4,5 turneri 1,2,3,4,5 traini 5
Total length 140 - ~290 mm 126 mm 142 mm 65.2-89.5 mm Vertebrae count 36 36 39 39 Fin ray count
Dorsal 8 8 9 7-8 Pectoral 9-10 14 11 13 Pelvic 9-10 8 9 8 Anal 8 8 10 8 Caudal 19 -20 19-21 23 18-20
Head length (285-311) (240) (291) (255-316) Head depth (217-255) (253) (180) (174-221) Maximum body depth (304-314) (290) (189) (166-246) Fins
Dorsal fin length (221 -227) (161) (157) (134-178) Dorsal fin base length (116-118)) (103) (111) (106) Pectoral fin length (181- ~220) - (122) (88-157) Pelvic fin length (170 – 189) - (144) (91-145) Anal fin length (188 – 202) (142) (163) (93-163) Caudal fin length (304 – 352) (337) (283) (216-330)
Min. caudal depth (142 - 174) (84) (72) (110) Snout to dorsal origin length (525-576) (547) (577) (573-592) Snout to occupit length (261-307) - (254) (286-338) Snout to pelvic origin length (533-551) (640) (567) (501-571) Snout to anal origin length (720-738) (753) (747) (734-823) Pectoral origin to anal origin length (439-506) - (433) (413) Pelvic origin to dorsal origin length (287 - 346) (258) (236) (191-226) Pelvic origin to anal origin length (186-271) (128) (193) (151-288) Pelvic origin to caudal base length (470-571) (452) (444) (463-537) Anal origin to caudal base length (293-320) (316) (272) (288) Position of dorsal origin to pelvic origin moderately posterior well anterior slightly posterior even Position of anal fin origin to dorsal posterior fin base slightly posterior moderately posterior well posterior even Forked caudal fin deeply moderately moderately moderately TL/head length 4.31-4.53 5.25 3.50 3.99-4.76 TL/caudal peduncle least depth 13.7 - 14.2 - 16.1 - 1, Lucas (1900). 2, La Rivers (1962). 3, La Rivers (1966). 4, Lugaski (1977). 5, Lugaski (1979).

PALUDICOLA, VOL. 7, NO. 4, 2010 152
middle to late Miocene of Nevada. These include two species of Gila Baird and Girard, 1853, and one species of Siphateles Cope, 1883 (Gila turneri [Lucas, 1900], Gila? esmeralda [La Rivers, 1966], and Siphateles traini [Lugaski, 1979]). However, the generic assignments of some of these species have varied or been questioned since they were first described (e.g., La Rivers, 1962, 1966; Lugaski, 1977; Smith et al., 2002). For completeness, comparison of L. lugaskii to these species is warranted (Table 2). The holotype of Gila turneri is from the valley of the Silver Peak Range at the southwest end of the Big Smokey Valley, Silver Peak Quadrangle, Esmeralda County, Nevada, but the exact locality has never been relocated (Lugaski, 1977). The age of the type locality is undetermined, but likely to be middle Miocene (late Barstovian to middle Clarendonian) based on the age of the “Esmeralda Formation” in the Silver Peak Quadrangle (Robinson, 1966; Abers and Stewart, 1972; Diamond and Ingersoll, 2002). Gila turneri has also been recorded from the middle part of the “Esmeralda Formation” of Stewart Valley, Mineral County, Nevada (Smith et al. 2002). The middle part of the formation in Stewart Valley has been radioisometrically dated (Ar40-Ar39) at 11.74 ± 0.03 and 11.58 ± 0.05, indicating a middle Miocene (Clarendonian) age (Swisher, 1992). Lucas (1900) originally referred this species to Leuciscus Cuvier, 1817. La Rivers (1962) referred this species to Richardsonius Girard, 1856, which was followed by Uyeno and Miller (1963) who questionably referred this species to Gila. Lavinia lugaskii differs from G. turneri by having the following (Table 2): 1) larger body size; 2) the maximum head and body depths are relatively larger; 3) the relative head length is less (HL is divisible into TL about 4.3 - 4.6 times); 4) the total vertebrae count is lower; 5) the dorsal, pectoral, anal, and caudal fin ray counts are lower; 6) the dorsal, pectoral, pelvic, anal, and caudal fins are relatively larger; 7) the dorsal fin origin is positioned slightly more posterior to the pelvic fin origin; 8) the caudal peduncle is more robust; and 9) the caudal fin is more deeply forked. The morphology of the pharyngeal arches and teeth of G. turneri have not been reported. Gila? esmeralda is from sedimentary unit 2 of the Esmeralda Formation (Albers and Stewart, 1972) exposed seven miles southeast of Coaldale Junction, Esmeralda County, Nevada (Lugaski, 1977). Evernden and James (1964) dated this unit by K-Ar at 13.0 Ma (corrected), indicating a probable late Barstovian age for the fish fauna. Lugaski (1977) noted that the lacustrine deposits that yielded G.? esmeralda represent a different depositional basin from the lacustrine deposits that yielded G. turneri. Smith (1975) regarded G.? esmeralda as a probable junior
synonym of G. turneri, but Lugaski (1977) provided convincing evidence that G.? esmeralda is distinct from G. turneri. Dowling et al. (2002) and Smith et al. (2002) only questionably regarded G. esmeralda as belonging to Gila because it exhibits certain similarities to Siphateles. Lavinia lugaskii differs from G.? esmeralda by having the following (Table 2): 1) larger body size; 2) ____________________________________________
FIGURE 6. Lavinia lugaskii. A-B, composite right maxilla outline based on LACM 154462, 154459, and154350, A, lateral view, anterior right, B, mesial view, anterior left. C, composite left dentary outline based on LACM 154390, 154350, 154391, and 154462, lateral view, anterior left. D, left opercle, LACM 154463, lateral view. E, left frontal, LACM 154459, dorsal view, anterior up. F. left frontal, LACM 154460, ventral view, anterior up. Scales = 5 mm.
__________________________________________ the relative head length is greater (HL is divisible into TL about 4.3 – 4.6 times); 3) the pectoral fin ray count is lower and pelvic fin ray count is higher; 4) the dorsal and anal fins are relatively larger; 5) the snout to pelvic fin origin length is relatively shorter; 6) the dorsal fin origin is positioned posterior to the pelvic fin origin, whereas in G.? esmeralda it is positioned well anterior to the pelvic fin; 7) the anal fin origin is positioned slightly less posterior to the dorsal fin posterior base; and 8) the caudal fin is more deeply forked. The morphology of the pharyngeal arches and teeth of G.? esmeralda have not been reported. Siphateles traini is from the zeolite beds exposed on the west side of Jersey Valley, Pershing County,

KELLY—MICOENE CHUB FROM NEVADA 153
Nevada (Lugaski, 1979). Deffeyes (1959) and Papke (1972) divide the lacustrine deposits into three units in the Jersey Valley area as follows; basal unit at least 427 m (1700 ft) thick, a middle unit 314 m (1,030 ft) thick, and an upper unit of unknown thickness. The uppermost portion of the upper unit contains the zeolite beds that yielded S. traini (Deffeyes, 1959; Johnson, 1977). Mammalian fossils of middle Miocene (Clarendonian) age occur 213 m (700 ft) below the fish locality level (Deffeyes, 1959; Johnson, 1977), indicating a possible late Clarendonian or early Hemphillian age for S. traini. However, Dowling et al. (2002) regard the type locality to be earliest Pliocene (latest Hemphillian) in age, or about 6 Ma. Lavinia lugaskii differs from S. traini by having the following (Table 2): 1) larger body size; 2) the maximum head and body depths are relatively larger; 3) the total vertebrae count is lower; 4) the pectoral fin ray count is lower and the pelvic fin ray count is higher; 5) the dorsal, pectoral, pelvic, and anal fins are relatively larger; 6) the dorsal fin origin is positioned posterior to the pelvic fin origin, whereas in S. traini they are even; 7) the anal fin origin is positioned posterior to the dorsal posterior fin base, whereas in S. traini they are even; 8) the pectoral fin origin to anal fin origin length is relatively longer; and 9) the caudal fin is more deeply forked. The morphology of the pharyngeal arches and teeth of S. traini have not been reported. An additional Miocene record of Gila from Nevada is Gila sp. from the late Miocene (Hemphillian) Truckee Formation exposed near the communities of Fernley and Hazen, Nevada, about 90 km northeast of the Mickey Canyon section (Smith, 1981; Baumgartner, 1982; Smith et al., 2002). Lavinia lugaskii is easily distinguished from G. sp. by having pharyngeal teeth with rugose, wavy ridged grinding surfaces and a relatively shorter dentary. Lavinia lugaskii can also be distinguished from all other species of Gila and Siphateles by having rugose grinding surfaces on the pharyngeal teeth. The distinguishing characters of L. lugaskii clearly indicate that it is distinct from the three previously named Miocene chub species and G. sp. from Nevada. In addition, L. lugaskii was recovered from lacustrine sediments that were deposited in a large depositional basin that was separated from those that yielded G. turneri, G.? esmeralda, S. traini, and G. sp. Whether this separate depositional basin was connected by any drainage system to the other basins is currently unknown (Gilbert and Reynolds, 1973; Golia and Stewart, 1984). As noted above, Smith et al. (2000, 2002) provided convincing evidence that Idadon is junior synonym of Lavinia based on their shared dentary morphology and the presence of striated, rugose
grinding surfaces on the pharyngeal teeth. However, the degree of the development of the rugose grinding surfaces is not equal in the four species of Lavinia and Hesperoleucus. It is well-developed in L. lugaskii and L. hibbardi, moderately to well-developed on some of the pharyngeal teeth of L. condonianus, and weakly developed in extant Lavinia and Hesperoleucus (Smith, 1975; Smith et al., 2000). Similarly, the minor row of pharyngeal teeth varies with the L. lugaskii having one moderately well-developed tooth, L. hibbardi and L. condonianus having one or two rudimentary teeth, and extant Lavinia and Hesperoleucus having none. The number of teeth in the major row also differs with species of Lavinia usually having five (sometimes four in L. exilicauda) and Hesperoleucus usually having five on the left pharyngeal and four on the right pharyngeal. The fin ray counts also vary between these taxa, with L. lugaskii having relatively low counts (e.g., dorsal and anal = 8), extant Lavinia having high counts (e.g., dorsal = 10-13, anal = 11-14), and Hesperoleucus having low counts (e.g., dorsal = 7-9, anal = 6-8). The fin ray counts for L. condonianus and I. hibbardi are unknown (Smith, 1975; Smith et al., 2000). Lavinia hibbardi and L. condonianus further differ from extant Lavinia by having pharyngeal teeth that are more rounded in cross-section (relatively wider) and less hooked terminally (Smith et al., 2000). The earliest Pliocene (latest Hemphillian or ~6 Ma) sample of Lavinia sp. from Cache Valley, Utah, has not been formally described or compared with other members of the Lavinia-Hesperoleucus lineage, but Smith et al. (2002) note that it exhibits slightly corrugated grinding surfaces on the pharyngeal teeth, flaired dentaries, and high fin ray counts. These character states, especially the high fin ray counts and weakly corrugated grinding surfaces of the teeth, are most similar to those of extant Lavinia. Thus, middle Miocene Lavinia lugaskii, certain Pliocene representatives of Lavinia, extant Lavinia, and Hesperoleucus seem to form a morphocline with each taxon exhibiting morphologically different character states for the fin ray counts (unknown for L. hibbardi and L. condonianus), rows and numbers of pharyngeal teeth, and development of rugose grinding surfaces on the teeth. However, it is difficult to determine with certainty the polarities of these character states based on an incomplete fossil record with significant chronological gaps. Moreover, the polarities of some of the character states of the dentary have not been adequately determined and some of them (mesially curved anterior end and flaired lateral edges of the gnathal ramus) are also shared in some degree with Orthodon Girard, 1856, or Mylopharodon (Smith, 1975; Smith et al., 2000), which would indicate that these character states may be sympleisomorphic for Lavinia and Hesperoleucus. Although, it appears that

PALUDICOLA, VOL. 7, NO. 4, 2010 154
the weakly corrugated grinding surfaces on the pharyngeal teeth and high fin ray counts seen in extant Lavinia and L. sp. from Cache Valley are probably derived character states because, in much older L. lugaskii, the rugose grinding surfaces are much better developed and the fin ray counts are lower.
CONCLUSIONS In series of papers analyzing morphological and molecular data, a general consensus is emerging in support a monophyletic Western North American chub group consisting of three ancient lineages (Coburn and Cavender, 1992; Simons and Mayden, 1998, 1999; Dowling et al., 2002; Smith et al., 2002). One of these lineages is the western chub group that is further divided in four clades: the Mylopharodon-Lavinia-Hesperoleucus and Siphateles clade; the Gila-Acrocheilus-Klamathella clade; the Relictus-Eremichthys clade; and the Ptychocheilus clade (Smith et al., 2002). The Mylopharodon-Lavinia-Hesperoleucus and Siphateles clade is further divided into two subgroups, the Mylopharodon-Lavinia-Hesperoleucus subgroup and the Siphateles subgroup. The occurrence of Lavinia lugaskii in the middle Miocene indicates that the Lavinia-Hesperoleucus lineage is older than previously thought. The Mylopharodon-Lavinia-Hesperoleucus subgroup is regarded as the closest sister group to the Siphateles subgroup with Mylopharodon and Lavinia diverging from a common ancestor after the divergence of Mylopharodon and Siphateles from their common ancestor (Dowling et al, 2002; Smith et al., 2002). Moyle (2002) regards Hesperoleucus as a junior synonym of Lavinia based on protein and molecular similarities and the fact they are interfertile (e.g., Miller, 1945; Hopkirk, 1973; Avise et al., 1975; Avise and Ayala, 1976; Moyle and Massingill, 1981), whereas Smith et al. (2002) regard Hesperoleucus as generically distinct from Lavinia based on the fossil record and morphological differences. Smith et al. (2002) considered the molecular similarity of these two taxa as probably a result of introgression (see also Smith, 1992). Based on nuclear and mitochondrial DNA similarities, Aguilar and Jones (2009) recently provided additional evidence of a close relationship between these two taxa. The oldest record of Mylopharodon is M. doliolus Smith and Kimmel, 1982 (in Smith et al., 1982) from the middle Miocene (early Clarendonian, about 12 Ma) Esmeralda Formation of Nevada (Smith et al., 1982, 2002; Smith and Cossel, 2002). Lavinia was previously known from the earliest Pliocene (latest Hemphillian) to Recent (Smith, 1975; McClellan, 1977; Wagner et al., 1997; Smith et al., 2002) and L. lugaskii extends this lineage at least six million years to the middle
Miocene (early Clarendonian), or about 12.5 – 12.0 Ma. Based on molecular sequences, Dowling et al. (2002) estimated the time of divergence of Lavinia and Mylopharodon from a common ancestor to be about 6 Ma and that of Siphateles and Mylopharodon to be about 12 Ma. Also based on molecular sequences, Smith et al. (2002) regarded the divergence of Siphateles and Mylopharodon from a common ancestor, estimated at more than 8 Ma, to have occurred before that of Lavinia and Mylopharodon from their common ancestor. If one accepts the previous hypothesis that Mylopharodon is the closest sister taxon to the Lavinia-Hesperoleucus lineage (Dowling et al., 2002; Smith et al., 2002), then the occurrence of L. lugaskii at 12.5-12.0 Ma would indicate that this lineage diverged from a common ancestor with Mylopharodon earlier than the early Clarendonian. Although resolving the generic status of Hesperoleucus is beyond the scope of this paper, certain implications arise with the occurrence of L. lugaskii in the middle Miocene. The fossil record of Hesperoleucus is limited, wherein it has only been recorded from the late Pleistocene to Holocene. If Hesperoleucus diverged from a common ancestor with an earliest Pliocene or later representative of Lavinia, which appears to be supported by the protein and molecular evidence and the putative shared derived character state of weakly corrugated grinding surfaces on the pharyngeal teeth, then Lavinia becomes paraphyletic unless Hesperoleucus is regarded as a synonym of Lavinia. Conversely, if Hesperoleucus diverged from a common ancestor with L. lugaskii prior to the middle Miocene (before 12.5-12.0 Ma), the shared character state of weakly corrugated grinding surfaces observed in L. exilicauda and H. symmetricus is due to convergence, and the protein and molecular similarities are due to introgression, then Hesperoleucus could be retained at generic rank and Lavinia would not be a paraphyletic taxon, but this would necessitate a long ghost lineage for Hesperoleucus. However, determining which of these phylogenetic scenarios is most likely correct must await more complete fossil records for both taxa.
AKNOWLEDGEMENTS I am grateful to Samuel A. McLeod of the LACM for his help in the curation of the specimens and for providing access to the vertebrate paleontology collection at the LACM. Patricia Holroyd of the UCMP and Rachel Dolbier of the W. M. Keck Earth Science and Mineral Engineering Museum, University of Nevada, Reno, provided access to the paleontological collections at their respective institutions. Vanessa Rhue of the LACM provided additional curation data

KELLY—MICOENE CHUB FROM NEVADA 155
for the specimens. I am indebted to Gerald R. Smith of the Museum of Paleontology and the Museum of Zoology, University of Michigan (UMMZ), Kenneth W. Gobalet of the California State University, Bakersfield, William W. Korth of the Rochester Institute of Vertebrae Paleontology, and Samuel A. McLeod of the LACM for their advice and constructive comments on the original draft of this report. Also, Gerald R. Smith and Douglas W. Nelson of the UMMZ graciously provided comparative material of Lavinia and Hesperoleucus.
LITERATURE CITED Abers, J. P. and J. H. Stewart. 1972. Geology and
mineral deposits of Esmeralda County, Nevada. Nevada Bureau of Mines and Geology Bulletin 78:1-80.
Aguilar, A and W. J. Jones. 2009. Nuclear and mitochondrial diversification in two native California minnows: insights into taxonomic identity and regional phylogeography. Molecular Phylogenetics and Evolution 51:373-381.
Avise, J. C. and F. J. Ayala. 1976. Genetic differentiation in speciose versus depauperate phylads: evidence from the California minnows. Evolution 30:46-58.
Avise, J. C., J. J. Smith, and f. J. Ayala. 1975. Adaptive differentiation with little genic change between two native California minnows. Evolution 29:411-426.
Axelrod, D. I. 1956. Mio-Pliocene floras from west-central Nevada. University of California Publications in Geological Sciences 33:1-322.
Ayres, W. O. 1855. Description of new fishes from California. Proceedings of the California Academy of Natural Science 1:23-77.
Baird, S. F. and C. Girard. 1853. Descriptions of some new fishes from the River Zuni. Proceedings of the Academy of Natural Science, Philadelphia 6:368-369.
Baird, S. F. and C. F. Girard. 1854. Description of new fishes, collected by Dr. A. L. Heermann, naturalist attached to the survey of the Pacific Railroad route, under Lt. R. S. Willaimson, U.S. Army. Academy of Natural Sciences Philadelphia 2:409-411.
Baumgartner, J. V. 1982. A new fossil ictalurid catfish from the Miocene middle member of the Truckee Formation, Nevada. Copeia 1982(1):38-46.
Bell, M. A., C. E. Wells, and J. A. Marshall. 1989. Mass-mortality layers of fossil Stickleback fish: catastrophic kills of polymorphic schools. Evolution 43:607-619.
Brown, L. R., P. B. Moyle, W. A. Bennett, and B. D. Quelvog. 1992. Implications of morphological variation among populations of California roach Lavinia symmetricus (Cyprinidae) for conservation policy. Biological Conservation 62:1-10.
Coburn, M. M. and T. M. Cavender. 1992. Interrelationships of North American cyprinid fishes, in Mayden, R. L. (ed.), Systematics, Historical Ecology, and North American Freshwater Fishes. Stanford University Press, Stanford, California, p. 328-373.
Cope, E. D. 1883. On the fishes of the Recent and Pliocene lakes of the western part of the Great Basin, and of the Idaho Pliocene lake. Proceedings of the National Academy of Sciences Philadelphia :134-166.
Cuvier, G. 1817. Le règne animal distribué d'après son organisation, pour servir de base à l'histoire naturelle des animaux et d'introduction à l'anatomie comparée. Avec figures, dessinées d'après nature. Tome II, contenant les reptiles, les poissons, les mollusques et les annélides. Paris, p. 532.
Dalrymple, G. B. 1979. Critical tables for conversion of K-Ar ages from old to new constants. Geology 7:558-560.
Diamond, D. S. and R. V. Ingersoll. 2002. Structural and sedimentologic evolution of a Miocene supradetachment basin, Silver Peak Range and adjacent areas, west-central Nevada. International Geology Review 44:588-623.
Deffeyes, K. S. 1959. Erionite from Cenozoic tuffaceous sediments, central Nevada. The American Mineralogist 44:501-509.
Dowling, T. E., C. A. Tibbets, W. L. Minckley, and G. R. Smith. 2002. Evolutionary relationships of the Plagopterins (Teleostei: Cyprinidae) from cytochrome b sequences. Copeia 2002(3):665-678.
Evernden, J. F. and G. T. James. 1964. Potassium-Argon dates and the Tertiary floras of North America. American Journal of Science 262:945-974.
Evernden, J. F., D. E. Savage, G. H. Curtis, and G. T. James. 1964. Potassium-Argon dates and the Cenozoic mammalian chronology of North America. American Journal of Science 262:145-198.
Fowler, H. W. 1924. Notes on North American cyprinoid fishes. Proceedings of the Academy of Natural Sciences, Philadelphia 76:389-416.
Girard, C. F. 1856. Researches upon the cyprinoid fishes inhabiting the fresh waters of the United States, west of the Mississippi Valley, from specimens in the museum of the Smithsonian

PALUDICOLA, VOL. 7, NO. 4, 2010 156
Institution. Proceedings of the Academy of Natural Sciences of Philadelphia 8: 165-213.
Gilbert, C. M. and M. W. Reynolds. 1973. Character and chronology of basin development, western margin of the Basin and Range Province. Geological Society of America Bulletin 84:2489-2510.
Golia, R. T. and J. H. Stewart. 1984. Depositional environments and paleogeography of the upper Miocene Wassuk Group, west-central Nevada. Sedimentary Geology 38:159-180.
Hopkirk, J. D. 1973. Endemism in fishes of the Clear Lake region. University of California, Publications in Zoology 96:1-160.
Johnson, M. G. 1977. Geology and mineral deposits of Pershing County, Nevada. Nevada Bureau of Mines and Geology Bulletin 89:1-115.
Kelly, T. S. 1994. Two Pliocene (Blancan) vertebrate faunas from Douglas County, Nevada. PaleoBios 16:1-23.
Kelly, T. S. 1998. New Miocene mammalian faunas from west central Nevada. Journal of Paleontology 72:137-149.
Kelly, T. S. and R. Secord. 2009. Biostratigraphy of the Hunter Creek Sandstone, Verdi Basin, Washoe County, Nevada, in Oldow, J. S. and P. H. Cashman (eds.), Late Cenozoic structure and evolution of the Great Basin-Sierra Nevada transition. Geological Society of America Special Paper 447, p. 133-146.
La Rivers, I. 1962. Fishes and fisheries of Nevada. University of Nevada Press, Reno, Nevada, p. 782.
La Rivers, I. 1966. Paleontological Miscellanei I: A new cyprinid fish from the Esmeralda Formation (Pliocene) of southeastern Nevada. Biological Society of Nevada, Occasional Papers 11:1-4.
Lucas, F. A. 1900. A new fossil cyprinoid, Leuciscus turneri, from the Miocene of Nevada. Proceedings of the U.S. National Museum 23:333-334.
Lugaski, T. 1977. Additional notes and discussion of the relationship of Gila esmeralda La Rivers 1966 from the “Esmeralda” Formation, Nevada. Biological Society of Nevada, Occasional Papers 43:1-4.
Lugaski, T. 1979. Gila traini, a new Pliocene cyprinid fish from Jersey Valley, Nevada. Journal of Paleontology 53:1160-1164.
McClellan, P. H. 1977. Paleontology and Paleoecology of Neogene freshwater fishes from the Salt Lake Beds, northern Utah. M.S. thesis, University of California, Berkeley, 243 p.
Miller, R. R. 1945. The status of Lavinia ardesiaca, a cyprinid fish from the Pajaro-Salinas Basin, California. Copeia 1945:197-204.
Miranda, R. and M. C. Escala. 2005. Morphometrical comparison of cleithra, opercular, and pharyngeal bones of autochthonous Leuciscinae (Cyprinidae) of Spain. Folia Zoologica 54:173-188.
Moyle, P. B. 2002. Inland fishes of California. University of California Press, Berkeley, p. 502.
Moyle, P. B. and M. Massingill. 1981. Hybridization between hitch, Lavinia exilicauda, and Sacramento Blackfish, Orthodon microlepidotus, in San Luis Reservoir, California. California Fish and Game 67:196-198.
Moyle, P. B., R. M. Yoshiyama, J. E. Williams, and E. D. Wikramanayake. 1995. Fish species of special concern in California. Report prepared for State of California, the Resources Agency, Department of Fish and Game, Inland Fisheries Division, Rancho Cordova, p. 153-172.
Murphy, R. R. 1945. The status of Lavinia ardesiaca, a cyprinid fish from the Pajaro-Salinas River basin, California. Copeia 1945(4):197-204.
Murphy, R. R. 1948. Distribution and variation of the roach (Hesperoleucus symmetricus) in the coastal region of California. M.S. thesis, University of California, Berkeley, 55 p.
Papke, K. 1972. Erionite and other associated zeolites in Nevada. Nevada Bureau of Mines and Geology Bulletin 79:1-32.
Perkins, M. E., F. H. Brown, W. P. Nash, W. McIntosh, and S. K. Williams. 1998. Sequence, age, and source of silicic fallout tuffs in middle to late Miocene basins of the northern Basin and Range province. Geological Society of America Bulletin 110:344-360.
Robinson, P. T. 1966. Zeolitic diagenesis of Mio-Pliocene rocks of the Silver Peak Ragne, Esmeralda County, Nevada. Journal of Sedimentary Petrology 36:1007-1015.
Schorn, H. E. and W. H. Shelton. 1991. A seed cone of Pinus sp. (subsect. Oocarpae) from the late Miocene of the Mickey Wash area, Lyon County, Nevada. PaleoBios 13:1-2.
Schorn, H. E., C. J. Bell, S. W. Starratt, and D. W. Wheeler. 1994. A computer-assisted annotated bibliography and preliminary survey of Nevada Paleobotany. U.S. Geological Survey Open-File Report 94-441:1-180.
Simons, A. M. and R. L. Mayden. 1998. Phylogenetic relationships of the western North American phoxinins (Actinopterygii: Cyprinidae) as inferred from mitochondrial 12S and 16S ribosomal RNA sequences. Molecular Phylogenetics and Evolution 9:308-329.
Simons, A. M. and R. L. Mayden. 1999. Phylogenetic relationships of North American cyprinids and

KELLY—MICOENE CHUB FROM NEVADA 157
assessment of homology of the open posterior myodome. Copeia 1999(1):13-21.
Smith, G. R. 1975. Fishes of the Pliocene Glenns Ferry Formation, southwest Idaho. University of Michigan, Papers on Paleontology 14:1-68.
Smith, G. R. 1981. Late Cenozoic freshwater fishes of North America. Annual Review of Ecology and Systematics 12:163-193.
Smith, G. R. 1992. Introgression in fishes: significance for paleontology, cladistics, and evolutionary rates. Systematic Biology 41:41-57.
Smith, G. R. and J. Cossel, Jr. 2002. Fishes from the late Miocene Poison Creek and Chalk Hills Formations, Owyhee County, Idaho. Idaho Museum of Natural History, Occasional Papers 37:23-35.
Smith, G. R. and R. R. Miller, 1985. Taxonomy of fishes from Miocene Clarkia lake beds, Idaho, in C. J. Smiley, ed., Late Cenozoic History of the Pacific North West. AAAS Pacific Division, San Francisco, pp. 75-84.
Smith, G. R., K. Swirydezuk, P. G. Kimmel, and B. H. Wilkinson. 1982. Fish biostratigraphy of late Miocene to Pleistocene sediments of the western Snake River plain, Idaho, in B. Bonnichsen and R. M. Breckenridge (eds.), Cenozoic Geology of Idaho. Idaho Bureau of Mines and Geology Bulletin 26, p. 519-541.
Smith, G. R., N. Morgan, and E. Gustafson. 2000. Fishes of the Mio-Pliocene Ringold Formation, Washington: Pliocene capture of the Snake River by the Columbia River. University of Michigan, Papers on Paleontology 32:1-47.
Smith, G. R., T. E. Dowling, K. W. Gobalet, T. Lugaski, D. K. Shiozawa, and R. P. Evans. 2002. Biogeography and timing of evolutionary events among Great Basin fishes, in Hershler and Madsen, and Curry (eds.), Great Basin Aquatic Systems History. Smithsonian Contributions to Earth Sciences 33, p. 1-104.
Snyder, J. O. 1913. Fishes of streams tributary to Monterey Bay, California. Bulletin of the U.S. Bureau of Fisheries 32:47-72.
Stewart, J. H. and M. W. Reynolds. 1987. Geologic map of the Pine Groove Hills quadrangle, Lyon County, Nevada. U.S. Geological Survey Open File Report 87-658:1-9.
Swisher, C. C. 1992. 40Argon/39Ar dating and its application to the calibration of the North American Land Mammal Ages. PhD dissertation, University of California, Berkeley, p. 239.
Tedford, T. H., Albright III, L. B., Barnosky, A. D., Ferrusquia-Villafranca, I., Hunt Jr., R. M., Storer, J. E., Swisher III, C. C., Voorhies, M. R., Webb, S. D., and Whistler, D. P. 2004. Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene epochs), in Woodburne, M. O., ed., Late Cretaceous and Cenozoic mammals of North America: Columbia University Press, New York, p. 169-231.
Uyeno, T. and R. R. Miller. 1963. Summary of late Cenozoic freshwater fish records for North America. Occasional Papers of the Museum of Zoology, University of Michigan 631:1-34.
Wagner, H. M., C. B. Hanson, E. P. Gustafson, K. W. Gobalet, and D. P. Whistler. 1997 . Biogeography of the Pliocene and Pleistocene vertebrate faunas of northeastern California and their temporal significance to the development of the Modoc Plateau and Klamath Mountain Region. San Bernardino County Museum Association Quarterly 44:13-21.
Wolfe, J. A., H. E. Schorn, C. E. Forest, and P. Molnar. 1997. Paleobotanical evidence for high altitudes in Nevada during the Miocene. Science 276:1672-1675.

Paludicola 7(4):158-169 May 2010 © by the Rochester Institute of Vertebrate Paleontology
158
NEW RECORDS OF MARSUPIALIA, LIPOTYPHLA, AND PRIMATES FROM THE DUCHESNEAN (MIDDLE EOCENE) SIMI VALLEY LANDFILL LOCAL FAUNA, SESPE
FORMATION, CALIFORNIA
Thomas S. Kelly
Research Associate, Vertebrate Paleontology Section, Natural History Museum of Los Angeles County
900 Exposition Blvd, Los Angeles, California 90007
ABSTRACT A paleontologic mitigation program at the Simi Valley Landfill and Recycling Center, Ventura County, California, has
yielded significant new samples of isolated small mammal teeth from the Duchesnean (middle Eocene) Simi Valley Landfill Local Fauna of the Sespe Formation. New records of a marsupial, lipotyphlans, and a microsyopid primate for this local fauna include: Herpetotherium sp., Batodonoides walshi n. sp., cf. Oligoryctes (genus and species undetermined), and cf. Uintasorex sp. In addition, significantly larger samples of the erinaceomorph lipotyphlan Sespedectes singularis and the marsupial Peradectes californicus are now recorded from the Simi Valley Landfill Local Fauna.
INTRODUCTION The Sespe Formation of Simi Valley, Ventura County, California, has long been recognized for its abundant fossil mammals of middle Eocene to upper Oligocene (Uintan-Arikarrean) age (e.g., Stock, 1932, 1935, 1936; Golz, 1976; Golz and Lillegraven, 1977; Mason, 1988; Kelly, 1990, 1992, 2009; Kelly et al., 1991; Kelly and Whistler, 1994, 1998). Five superposed local faunas of middle Eocene (Uintan-Duchesnean) age have been recognized from the middle member of the Sespe Formation (Kelly, 1990, 1992; Kelly et al., 1991). The youngest of these faunas is the Duchesnean Simi Valley Landfill Local Fauna from bed 30A (Natural History Museum of Los Angeles County locality 5876) of the middle member of the Sespe Formation (Kelly et al., 1991). Prothero et al. (1996) documented the paleomagnetism of the middle member and placed bed 30A within Chron 17r of the geomagnetic polarity time scale, or about 38.0 – 37.8 million years before present (Luterbacher et al., 2004). Recently, 345 isolated teeth of fossil small mammals were recovered by wet screen sieving of bulk matrix from bed 30A during a paleontologic mitigation program at the Simi Valley Landfill and Recycling Center (Lander, 2008). These teeth represent a significant increase in the samples of fossil mammals of the Simi Valley Landfill Local Fauna.
Within these new samples are new occurrences of marsupials, lipotyphlans, and a microsyopid primate from the Simi Valley Landfill Local Fauna (Walsh, in Lander, 2008, Appendix C), including a new species of geolabidid. The purpose of this report is to document these new occurrences.
METHODS Measurements of teeth were made with an optical micrometer to the nearest 0.01 mm. Eutherian cheek teeth cusp and dental terminology follow standard usage, while those of marsupials follows Marshall et al. (1990) and Korth (1994). Upper and lower teeth are designated by uppercase and lowercase letters, respectively. All specimens were recovered by wet screen sieving of bulk matrix from bed 30A (LACM locality 5876) of the middle member of the Sespe Formation at the Simi Valley Landfill and Recycling Center during a mitigation program directed by Paleo Environmental Associates, Inc., for Waste Management of California, Inc. All specimens are deposited in the Vertebrate Paleontology Section of the Natural History Museum of Los Angles County. Detailed locality data are available at this institution and also see Lander (2008). Abbreviations and acronyms are as follows: ap, greatest anteroposterior length; L, left; ht-tal, height of talonid; ht-tri, height of trigonid; LACM, Natural

159 KELLY—DUCHESNEAN MAMMALS FROM SIMI VALLEY
History Museum of Los Angeles County; N, number of specimens; Ma, million years before present; OR, observed range; R, right; tr, greatest transverse width; tra, anterior transverse width; trp, posterior transverse width; tr-talb, greatest transverse width of talonid basin.
SYSTEMATIC PALEONTOLOGY
Cohort Marsupialia Illiger, 1811 Order Didelphimorpha Gill, 1872
Family Herpetotheriidae Trouessart, 1879 Genus Herpetotherium Cope, 1873
Herpetotherium sp.
Referred Specimen―Partial LM1 or 2, LACM 153689.
Description―The partial upper molar is missing the posterior labial corner, including a large portion of the stylar shelf (Figure 1A). It is moderately worn and the enamel surface is somewhat abraded, which probably occurred during deposition in a fluvial environment. The tooth is dilambdodont with a V-shaped centrocrista. Stylar cusp A is small and positioned near the anterolabial corner of the tooth, close to the posterolingual terminus of the preparacrista and just anterior and lingual to stylar cusp B. It is connected anterolabially to the anterior cingulum. Stylar cusp B is worn down (or abraded), but its occlusal outline clearly indicates it is well developed and positioned along the labial border of the tooth, slightly more anteriorly than the paracone. The occurrence and morphology of other stylar cusps cannot be determined because of the missing portion of the stylar shelf. The paracone and metacone are well developed with triangular occlusal outlines. The metacone is larger and slightly taller than the paracone. The protocone is large with a triangular occlusal outline and slightly rounded along its lingual border. The preprotocrista extends anterolabially from the protocone to a weak, but distinct protoconule and then continues to join the anterior cingulum. The postprotocrista extends posterolabially from the protocone to a small, distinct metaconule, which is larger than the protoconule. The anterior cingulum is moderately developed and extends lingually from stylar cusp A to join the preprotocrista. The enamel along the labial border of the stylar shelf is slightly crenulated. Measurements of partial upper molar are ap = 1.90 mm and tra = 2.33 mm (ap and tr dimensions of the whole tooth would be larger).
Discussion―Many species that were formerly assigned to Peratherium Aymard, 1846, from the early to middle Eocene of North America are now regarded as belonging to Herpetotherium with Peratherium restricted to European species (Rothecker and Storer,
1996; Korth, 2008). Herpetotherium was historically assigned to a subfamily (Herpetotheriinae Trouessart, 1879) of Didelphidae Gray, 1921 (e.g, Korth, 1994; McKenna and Bell, 1997), but recent investigations indicate that Herpetotherium is the sister group to Didelphidae and the subfamily has been elevated to familial rank, Herpetotheriidae (Kirsch et al., 1997; Case et al., 2005; Sánchez-Villagra et al., 2007; Becker et al., 2008). Species of Herpetotherium are primarily distinguished by size and the relative size and positions of the stylar cusps (e.g., McGrew, 1959; Fox, 1983; Krishtalka and Stucky, 1983a,b, 1984; Korth, 1994, 2008; Eberle and Storer, 1995; Rothecker and Storer, 1996). ___________________________________________
FIGURE 1. Herpetotherium sp. and Peradectes californicus from Simi Valley Landfill Local Fauna. A, Herpetotherium sp., partial LM1 or 2, LACM 153689. B-F, Peradectes californicus: B, RM4, LACM 153695; C-D, Lm1, LACM 153691; E-F, Rm3, LACM 153694. All occlusal views except D (labial view) and F (lingual view). Scale = 1 mm. __________________________________________________
Herpetotherium is a relatively rare taxon in the Sespe Formation with H. cf. H. knighti being previously recorded from the Uintan Tapo Canyon and

PALUDICOLA, VOL. 7, NO. 4, 2010 160
Brea Canyon local faunas (Stock, 1936; Kelly, 1990). The Sespe Uintan sample of Herpetotherium is only known from lower dentitions. Except for its larger size, LACM 153689 is very similar in occlusal morphology to those in the Uintan sample of H. cf. H. knighti from San Diego (Lillegraven, 1976). It differs from Bridgerian H. knighti (McGrew, 1959) by having a slightly weaker protoconule and larger size. Troxell (1923) described H. marsupium from the Bridger Formation, Wyoming. This species is now known from Texas, Utah, Wyoming, and Canada, ranging from the Wasatchian to Duchesnean (Kishtalka and Stucky, 1983b, 1984; Korth, 2008). Rothecker and Storer (1996) described H. sp., cf. H. marsupium from the Duchesnean Lac Pelletier Lower Fauna of Saskatchewan. The Sespe upper molar is similar in size to those of H. marsupium and slightly larger than those of H. sp., cf. H. marsupium. It is larger than those of H. fugax Cope, 1873, H. edwardi (Gazin, 1952), H. youngi (McGrew, 1937), H. valens (Lambe, 1908), and H. merriami (Stock and Furlong, 1922), and much smaller than those of H. comstocki (Cope, 1884). A specific assignment for the Sespe specimen cannot be made until better material is available from the Simi Valley Landfill Local Fauna that includes complete upper molars with stylar cusps C and D present. Therefore, LACM 153689 is referred to an indeterminate species of Herpetotherium. This is the first record of the genus from the Duchesnean of the Sespe Formation.
Family Peradectidae Crochet, 1979 Genus Peradectes Matthew and Granger, 1921
Peradectes californicus (Stock, 1936)
Referred Specimens―RM4, LACM 153695; Partial Rdp3, LACM 153690; Lm1, LACM 153691; Lm3, LACM 153692, 153693; Rm3, LACM 153694.
Discussion―The teeth of Peradectes from the Simi Valley Landfill Local Fauna (Figure 1B-F) are indistinguishable in size and occlusal morphology from those of the Uintan samples of P. californicus from San Diego and the Sespe Formation (Stock, 1936; Lillegraven, 1976), and are referred to this species. Measurements of the referred teeth are presented in Table 1. Previously, only one lower molar of P. californicus had been reported from the Simi Valley Landfill Local Fauna (Kelly and Whistler, 1994).
Grandorder Lipotyphla Haeckel, 1866 Order Erinaceomorpha Gregory, 1910 Family Sespedectidae Novacek, 1985
Genus Sespedectes Stock, 1935 Sespedectes singularis Stock, 1935
Referred Specimens―Partial RM1, LACM 153697; RM2, LACM 153698; LM3, LACM 153699, 153701; partial LM3, LACM 153700, 153702, 153703, 153704, 153705; RM3, LACM 153707; partial RM3, LACM 153706; Lm1, LACM 153696.
Discussion―Novacek (1985) provided a detailed description of Sespedectes singularis. Only two teeth of S. singularis were previously known from the Simi Valley Landfill Local Fauna (Kelly and Whistler, 1994). The new sample of teeth from the Simi Valley Landfill Local Fauna are indistinguishable in size and occlusal morphology from those of the Uintan-Duchesnean sample of S. singularis from the Sespe Formation (Novacek, 1976, 1985) and are referred to this species. Measurements of the new sample are provided in Table 2. ___________________________________________ TABLE 1. Measurements (in mm) of Peradectes californicus from Simi Valley Landfill Local Fauna. LACM # Position/Dimension Measurement 153695 RM4 ap 1.02 tr 1.71 153690 partial Rdp3 ap – tra – trp 0.72 153691 Lm1 ap 1.39 tra 0.92 trp 0.72 153692 Lm3 ap 1.30 tra 0.93 trp 0.76 153693 Lm3 ap 1.44 tra 0.80 trp 0.84 153694 Rm3 ap 1.46 tra 0.75 trp 0.79 ____________________________________________
Order Soricomorpha Gregory, 1910 Family Geolabididae McKenna, 1960a Genus Batodonoides Novacek, 1976
Batodonoides walshi new species
Holotype―Lm2, LACM 153709. Type Locality―LACM 5876, bed 30A of the
middle member, Sespe Formation, Simi Valley Landfill and Recycling Center, Ventura County, California.
Referred Specimens―RP4, LACM 153708; Lm3, LACM 153710; Lm3, LACM 153711.
Fauna and Age―Simi Valley Landfill Local Fauna, middle Eocene (Duchesnean), or about 38.0-37.8 Ma.

161 KELLY—DUCHESNEAN MAMMALS FROM SIMI VALLEY
Etymology―Named in honor of the late Steven L. Walsh of the Department of Paleontology, San
FIGURE 2. Batodonoides walshi n. sp. from Simi Valley Landfill Local Fauna. A-B, RP4, LACM 153708, A-occlusal view, B-lingual view. C-E, Lm2, holotype, LACM 153709, C-occlusal view, D-labial view, E-lingual view. F-H, Lm3, LACM 153710, F-occlusal view, G-labial view, H-lingual view. Scale = 1 mm. __________________________________________________
Diego Natural History Museum, California, for his extensive contributions to our understanding of Eocene mammals of the Pacific Coast.
Diagnosis―Differs from Batodonoides powayensis Novacek, 1976, and B. vanhouteni Bloch et al., 1998, by a distinctive talonid morphology including the following: 1) relatively much narrower and more anteroposteriorly orientated m2-3 talonid basins; 2) single, tall talonid cusp (hypoconulid or labially displaced hypoconid) positioned at posterior most point of talonid basin; and 3) lacking a notch at the posterior aspect of talonid. Further differs from B. powayensis
by having the m3 paraconid slightly taller relative to the protoconid height. Further differs from B. vanouteni by having the following: 1) P4 with distinct, small protoconule and incipient metaconule present; 2) m2 metaconid slightly taller relative to protoconid height; and 3) larger size.
Description―The P4 (LACM 153708) has a triangular occlusal outline and is in early wear (Figures 2A-B). The paracone is very tall, sharply pointed, and the dominant cusp. There is no metacone present. A very steep parastylar crest (precrista) descends sharply from the anterior apex of the paracone to join with the preprotocrista. A small, but distinctive, protoconule is present along the preprotocrista, just lingual to the junction of the preprotocrista and parastylar crest. A distinctive, large, long, blade-like postcrista extends posterolabially from the apex of the paracone to a small metastyle. A small, crescentric parastylar lobe is present that is low and rather flat with a raised external rim. Along this rim or cingulum, at the anterolabial corner of the tooth, is a small parastyle. A narrow, shelf-like, labial cingulum, which is continuous with the rim or cingulum along the parastylar lobe, extends posteriorly from the parastylar lobe to the metastyle. There are no stylar cusps present. The preprotocrista and postprotocrista form a continuous rim along the lingual aspect of the tooth from the posterolingual base of the paracone to a small, distinct protoconule, completely enclosing the trigon. Although a distinct, elevated protocone is not present, a thickening of this rim occurs at its most lingual aspect, which appears to be an incipient protocone. Also, on this rim at the junction of the posterolingual base of the paracone and the postprotocrista is a distinct, raised, circular swelling that appears to be a small metaconule. The postmetacrista is a distinct crest that ascends posterolabially from the metaconule to the metastyle. The talon is relatively flat, but curves rapidly upward at the junction with the lingual base of the paracone. Anterolingual and posterolingual cingula are lacking. The m2 is in early wear (Figures 2C-E, 3A). It is characterized by a well-developed trigonid with the apexes of the protoconid and metaconid significantly higher than the talonid. The talonid is transversely narrower than the trigonid. The protoconid is the largest and tallest cusp (height = 1.01 mm from labial base to apex) with the metaconid only slightly lower in height. The paraconid is the lowest cusp of the trigonid with its apex 0.29 mm lower than that of the protoconid. The bases of the protoconid and metaconid are fused for just over half their height, forming a steep wall along the anterior aspect of the talonid basin, and above which they are separated by a deep V-shaped protocristid notch. The protoconid and metaconid are slightly compressed anteroposteriorly, so that the protocristid (= metalophid) forms a continuous, sharp,

PALUDICOLA, VOL. 7, NO. 4, 2010 162
blade-like edge from the apex of the protoconid to the bottom of the protocristid notch and back up to the apex of the metaconid. The paraconid is positioned TABLE 2. Measurements (in mm) of Sespedectes singularis from Simi Valley Landfill Local Fauna. LACM # Position/Dimension Measurement 153698 RM2 ap 1.52 tra 2.25 trp 2.03 153699 LM3 ap 1.15 tra 1.69 trp 1.12 153701 LM3 ap 1.18 tra 1.87 trp 1.15 153707 RM3 ap 1.12 tra 1.82 trp 1.13 153700 partial LM3 ap – tra – trp 1.14 153702 partial LM3 ap – tra – trp 1.14 153703 partial LM3 ap 1.21 tra – trp – 153704 partial LM3 ap – tra – trp 1.14 153705 partial LM3 ap – tra – trp 1.14 153706 partial RM3 ap – tra – trp 1.13 153696 Lm1 ap 1.65 tra 1.35 trp 1.47 __________________________________________ anterolingually with its apex in line with that of the metaconid. A weakly developed paracristid (preprotocristid plus parastylar cristid = protolophid) extends from the protoconid to the anterolabial apex of the paraconid. A well-developed anterior cingulid is present that extends upward from the anterolabial base of the protoconid to the anterolabial base of the paraconid. The talonid is distinctive, in that it is narrower than the trigonid with a moderately deep talonid basin that is much narrower than the overall width of talonid. The talonid basin is rimmed by a distinct, continuous cristid (= entoconid cristid plus cristid obliqua) This talonid cristid ascends sharply
from the posterolingual base of the metaconid to the posterior most point of the talonid basin and then descends sharply as a cristid obliqua, and terminates very close to and just lingual of the middle of the fused bases of the protoconid and metaconid, wherein a small notch separates the cristid obliqua from the wall. This small notch is a continuation of a transversely orientated, small, shallow valley that lies along the anterior aspect of the talonid basin. The cristid obliqua portion of the continuous talonid cristid is lower than the lingual portion. No indication of an entoconid is present along the lingual aspect of the talonid cristid. A single, distinct, elevated cusp is present at the posterior most point of the talonid basin along the talonid cristid with its position central to the long axis of the talonid. The position of this cusp would indicate that it is a hypoconulid. There is no indication of a hypoconid along the posterolabial aspect of the talonid basin or talonid cristid. Whether this cusp is homologous with a hypoconulid or a lingually displaced hypoconid cannot be determined. No matter which cusp it represents, the morphology of the talonid is very distinctive with the combination of a very, narrow talonid basin and a single, tall, posteriorly positioned cusp, which gives the talonid a sectorial appearance. ___________________________________________
FIGURE 3. Line drawings Lm2-3 of Batodonoides walshi n. sp. from Simi Valley Landfill Local Fauna showing occlusal outlines of trigonid and talonid basins, A, Lm2, holotype, LACM 153709. B, Lm3, LACM 153710. Occlusal views, scale = 1 mm. ___________________________________________________
Two m3s are represented; one is barely worn (LACM 153710) and the other is in moderate wear (LACM 153711). The m3 is smaller than the m2 with the trigonid not as tall (Figures 2F-H, 3B). The morphology of the m3 trigonid and anterior cingulid are very similar to those of the m2, but the paraconid is positioned slightly closer to the metaconid. The m3 talonid exhibits a similar morphology with that of the m2 talonid, but differs by being even more compressed transversely and the single, posteriorly positioned hypoconulid (or displaced hypoconid) is relatively taller (m3 talonid height = 66% of trigonid height, whereas m2 talonid height = 56% of trigonid height). The talonid basin has also a slight constriction labially

163 KELLY—DUCHESNEAN MAMMALS FROM SIMI VALLEY
and lingually just anterior to the hypoconulid (or displaced hypoconid), making the cusp even more distinct and the talonid basin more sectorial in appearance. Measurements of the teeth are presented in Table 3. ____________________________________________ TABLE 3. Measurements (in mm) of Batodonoides walshi n. sp. from Simi Valley Landfill Local Fauna. See methods for abbreviations for dimensions. LACM # Position/Dimension Measurement 153708 RP4 ap 0.91 tr 0.93 ht 0.87 153709 Lm2 ap 1.01 tra 0.63 trp 0.47 ht-tri 1.01 ht-tal 0.62 tr-talb 0.21 153710 Lm3 ap 0.91 tra 0.62 trp 0.42 ht-tri 0.82 ht-tal 0.54 tr-talb 0.18 153711 Lm3 ap 0.87 tra 0.53 trp 0.39 ht-tri 0.80 ht-tal 0.53 tr-talb 0.17 ____________________________________________
Discussion―Four genera are usually assigned to the Geolabididae (McKenna, 1960a; Novacek, 1976; Lillegraven et al., 1981, Bloch, et al., 1998; Gunnell et al., 2008); the late Cretaceous Batodon Marsh, 1892, the Wasatchian to Uintan Batodonoides, the Wasatchian to Arikareean Centetodon Marsh, 1872, and the late Wasatchian to Bridgerian Marsholestes McKenna and Haase, 1992 (= Myolestes of Matthew, 1909). However, Wood and Clemens (2001) did not assign Batodon to any family and referred it to Placentalia, incertae sedis, whereas Kielan-Jaworowska et al. (2004) assigned Batodon to the family Cimolestidae Marsh, 1889, order Cimolesta McKenna, 1975. Batodon and Marsholestes are monotypic, represented by B. tenuis Marsh, 1892, and M. dasypelix (Matthew, 1909), respectively. Batodonoides is a relatively rare taxon with two species previously recognized; B. powayensis from the Uintan of southern California and B. vanhouteni from the Wasatchian of Wyoming (Novacek, 1976; Kelly, et al., 1991; Walsh, 1991, 1996; Bloch et al., 1998). This
rarity is probably due its very small size, wherein most all specimens have been recovered by wet screen sieving of matrix or acid preparation of freshwater limestones (Block et al., 1998; Bloch and Bowen, 2001; Silcox and Rose, 2001). The Sespe geolabidid can be confidently assigned to Batodonoides because it exhibits the following suite of characters (Novacek, 1976; Bloch et al., 1998; Gunnell et al., 2008): 1) the P4 is lacking a metacone and stylar cusps; 2) the lower molar trigonids are very tall, with the protoconids and metaconids fused for much of their height, and the talonids are narrower than the trigonids; 3) m3 smaller than m2; and 4) very small size. Except for the distinctive lower molar talonid occlusal morphology, the lower molars of the Sespe Batodonoides are overall very similar morphologically to those of B. powayensis and B. vanhouteni. The P4 occlusal morphology of the Sespe Batodonoides is also very similar to that of B. vanhouteni, differing primarily by having a small protoconule and incipient metaconule present. ____________________________________________
FIGURE 4. cf. Oligoryctes, genus and species undetermined, from Simi Valley Landfill Local Fauna, partial LM1 or 2, LACM 153688, occlusal view. Scale = 1 mm. __________________________________________________
The m2-3 talonids of the Sespe Batodonoides have taken the trend seen in B. powayensis and B. vanhouteni of a transverse narrowing of the talonid basins and reduction of the talonid cusps to an extreme, giving the talonids an almost sectorial appearance, especially m3. The m2-3 greatest talonid basin transverse widths of the Sespe Batodonoides are 43-44% of the m2-3 trps, whereas those of B. powayensis and B. vanhouteni are 60-78% of the trps. The occlusal outlines of the talonid basins of the Sespe Batodonoides are > -shaped with a single, tall cusp at the posterior most point of the trigonid basin, which is also centrally positioned relative to the long axis of the

PALUDICOLA, VOL. 7, NO. 4, 2010 164
tooth. In B. powayensis, weak lower molar entoconids are usually present and positioned near (or sometimes fused with) a distinct hypoconulid, whereas entoconids are lacking in the Sespe Batodonoides. Lower molar entoconids are also lacking in B. vanhouteni. In B. powayensis and B. vanhouteni a moderately to well-developed hypoconid is present that is positioned posterolabially along the rim of the talonid basin and separated from the hypoconulid by a distinct notch at posterior aspect of the basin. In the Sespe Batodonoides, this notch is completely lacking and there is no cusp present along the entire lingual rim of the talonid basin. Although the sample of Batodonoides from the Simi Valley Landfill Local Fauna is extremely small, the very distinctive talonid morphology clearly separates it from all other species of Batodonoides and justifies recognizing a new species, B. walshi. This is the first record of the genus from the Duchesnean and, thus, the youngest known occurrence of the genus.
Family Oligoryctidae Asher et al., 2002 Genus Oligoryctes Hough, 1956
cf. Oligoryctes, genus and species undetermined
Referred Specimens―partial LM1 or 2, LACM 153688.
Description―LACM 153688 is broken with a partial occlusal outline that is anteroposteriorly narrow and most similar to those of the M1-2s of Oligoryctes Hough, 1956 (Figure 4). The tooth is zalambdodont (no metacone present) with a single, tall paracone whose apex is sharp and centrally positioned. The parastyle and metastyle are well-developed with a moderately deep ectoflexus occurring between them. Distinct anterior and posterior cingula are present. The talon is broken away, but the tooth exhibits a distinct widening and thickening of the enamel just labial to the broken edge, especially posteriorly, suggesting that a protocone was present. Measurements of the partial upper molar are ap = 1.52 mm and tr = 0.76 mm.
Discussion―Five genera of soricomorph lipotyphlans that have or are presumed to have zalambdodont upper molars (lacking metacones) are currently recognized (Asher et al., 2002; Lopatin, 2006; Gunnell et al., 2008) as follows: from North America, Wasatchian Parapternodus Bown and Schankler, 1982, and Koniaryctes Robinson and Kron, 1998; Uintan through Whitneyan Oligoryctes; Duchesnean through Orellan Apternodus Matthew, 1903; and from Mongolia, middle Eocene Asiapternodus Lopatin, 2006. Koniaryctes is the only genus whose upper molars are unknown, but they are presumed to be zalambdodont based on its lower dentition (Robinson and Kron, 1998; Asher et al., 2002). Although most of these genera were previously
assigned to the Apternodontidae (e.g., Matthew, 1910; Hough, 1956; Bown and Schankler, 1982; Robinson and Kron, 1998), Asher et al. (2002) recognized three separate families for these taxa: Apternodontidae (including Apternodus); Parapternodontidae (including Parapternodus and Koniaryctes); and Oligoryctidae (including Oligoryctes). Subsequently, Lopatin (2006) added Asiapternodus to the Apternodontidae. A third unnamed oligoryctid was also recognized from the Bridgerian of Colorado, Nevada, Utah, and Wyoming (Asher et al., 2002). Walsh (1996) included Oligoryctes in a faunal list from the early Uintan Poway Fauna of California, but this material has never been formally described. Asher et al. (2002) provided evidence that these genera form a monophyletic clade wherein Apternodus is the sister taxon to Oligoryctes, which is the sister taxon to a soricid-parapternodontid clade. The Sespe partial upper molar is most similar to those of Oligoryctes, including a distinct anterior cingulum, small size, possibly a small protocone present, and lacking a metacone (zalambdodont). However, because of the broken condition of the specimen, generic assignment is unwarranted, so it is referred to an undetermined oligoryctid close to Oligoryctes. This is the first record of Oligoryctidae from the Sespe Formation and Simi Valley Landfill Local Fauna.
Order Primates Linnaeus, 1758 Family Microsyopidae Osborn and Wortman, 1892
Uintasoricinae Szalay, 1969 Genus Uintasorex Matthew, 1909
cf. Uintasorex sp.
Referred Specimen―LM3, LACM 153954. Description―The LM3 is moderately well-worn
resulting in the three primary cusps being of about equal height (Figure 5). The paracone, metacone and protocone are well developed with the paracone larger than the metacone and the protocone about equal in size to the paracone. The metacone is slightly compressed laterally. The protocone is positioned slightly anterior of the center of the tooth so that its apex is more aligned with that of the paracone than with that of the metacone. The trigon basin is relatively wide and shallow. The preprotocrista is a low crest that extends anterolabially to a small, but distinct protoconule, which is positioned anteriorly, near the anterolingual base of the paracone. The postprotocrista is distinct and extends labially to about the middle of the lingual aspect of the metacone. Possibly due the worn condition of the tooth, a distinct metaconule is not visible on the tooth, so whether or not it was present cannot be determined. Similarly, a distinct posterolingual cingulum cannot be discerned, but a

165 KELLY—DUCHESNEAN MAMMALS FROM SIMI VALLEY
very slight shelf along the posterior aspects of the protocone and postprotocrista suggests that a weak posterolingual cingulum was present. The anterolingual cingulum is weakly developed and extends lingually from the anterior base of the protocone to the anterolabial base of the paracone. Measurements of LACM 153954 are ap = 0.73 and tr = 0.75 mm. ___________________________________________
FIGURE 5. cf. Uintasorex sp. from Simi Valley Landfill Local Fauna, LM3, LACM 153954, occlusal view, scale = 0.5 mm. ____________________________________________________
Discussion―Two species of Uintasorex are
currently recognized, U. parvulus Matthew, 1909, from the Bridgerian of Wyoming (Matthew, 1909; Szalay, 1969; Rudman, 1981), and U. montezumicus Lillegraven, 1976, from the Uintan of the San Diego area, California (Matthew, 1909; Szalay, 1969; Lillegraven, 1976; Golz and Lillegraven, 1977; Walsh, 1991, 1996; Silcox and Gunnell, 2008). The only previous record of Uintasorex from the Sespe Formation was a single lower molar from the Uintan Tapo Canyon Local Fauna referred to U. sp., cf. U. montezumicus by Kelly and Whistler (1994). Uintasorex parvulus and U. montezumicus are similar in size and overall dental morphology, differing primarily in relative cusp positions and degree of development of cingula/cingulids and lophs/ lophids (Lillegraven, 1976). The Tapo Canyon Uintasorex sp., cf. U. montezumicus differs from U. montezumicus by its smaller size and having the widths of the m1 trigonid and talonid slightly smaller relative to the corresponding ap length (Kelly and Whistler, 1994). The M3 from the Sespe Formation at the Simi Valley Landfill differs from those of U. montezumicus by having the following: 1) the posterolingual cingulum is less developed or lacking; 2) the transverse width is slightly larger relative to the ap length (ap/tr = 0.973 versus 0.883 for U. montezumicus); 3) a distinct metconule lacking; and 4) smaller in size. Storer (1995) described an isolated M3 of Uintasorex sp. from the
late Duchesnean Lac Pelletier Upper Fauna of Saskatchewan. The Sespe M3 is much smaller than the Lac Pelletier M3. Gazin (1958) referred a sample of isolated teeth to U. parvulus from the Bridgerian Green River Formation, but Szalay (1969) assigned this sample to Uintasorex sp. because the teeth are smaller in size than those of U. parvulus from the Bridger Formation. The Sespe M3 differs from those of this taxon by having the following: 1) the protoconule slightly less developed; 2) a distinct metaconule is lacking; 3) the posterolingual cingulum is less developed or lacking; 4) the transverse width is slightly larger relative to the ap length; 5) and slightly smaller size. The Sespe M3 can be easily distinguished from those of Wasatchian Niptomomys McKenna, 1960b, the only other genus of the Uintasoricinae, by having the following: 1) the paracone and metacone are positioned relatively further apart resulting in the occlusal outline being more triangular (less ovoid) and the greatest ap width occurs across the labial aspect of the tooth; 2) a more weakly developed protoconule; 3) lacking a incipient metastyle; and 4) much smaller size. At least two million years separate the Uintasorex sp., cf. U. montezumicus specimen (Lm1) of the Tapo Canyon Local Fauna from the uintasoricine specimen (LM3) of the Simi Valley Landfill Local Fauna (Kelly et al., 1991; Prothero et al., 1996). Whether the two uintasoricine teeth from the Sespe Formation represent the same or different species cannot be determined, but they are both characterized by their small size. Until a much larger sample from the Sespe Formation is available, LACM 153954 is herein referred to cf. Uintasorex sp. This is the first record of the Uintasoricinae from the Duchesnean portion of the Sespe Formation.
AKNOWLEDGEMENTS
I am indebted to Samuel A. McLeod of the LACM for his support and providing access to the new sample of small mammal teeth from Sespe Formation. I am grateful to Rachel Dolbier of the W. M. Keck Earth Science and Mineral Engineering Museum, University of Nevada, Reno, for providing help in securing an inter-institutional loan of the specimens reported on herein. Special thanks are given to E. Bruce Lander of Paleo Environmental Associates, Inc. (PEA) and the LACM, and Mark A. Roeder of the San Diego Natural History Museum (SDMNH) and PEA for their support and advice during the preparation of this paper. I am thankful to William W. Korth of the Rochester Institute of Vertebrae Paleontology and Gregg Gunnell of the University of Michigan for their constructive comments and advice on the original draft of this report. My appreciation and admiration goes out to the late Steven L. Walsh of the SDMNH for his many

PALUDICOLA, VOL. 7, NO. 4, 2010 166
insightful comments and conversations regarding Eocene small mammals that he provided me over a 20 year period. Waste Management of California, Inc., provided funding for the Paleontologic Resource Impact Mitigation Program at the Simi Valley Landfill and Recycling Center.
LITERATURE CITED
Asher, R. J., M. C. McKenna, R. J. Emry, A. R.
Tabrum, and D. G. Kron. 2002. Morphology and relationships of Apternodus and other extinct, zalambdodont, placental mammals. Bulletin of the American Museum of Natural History 273:1-117.
Aymard, A. 1846. Essai monographique sur un genre noveau de mammifére fossile trouvé dans la Houte-Loire et nommé Enteldon. Annes de la Société d’Agriculture, Sciences, Arts, et Commerce du Puy 12:227-267.
Becker, R. M. D., M. Archer, H. Godthelp, B. S. Mackness, S. J. Hand, and J. Muirhead. 2008. A bizarre new family of Marsupialia (Incertae sedis) from the early Pliocene of northeastern Australia: implications for the phylogeny of bunodont marsupials. Journal of Paleontology 82:749-762.
Bloch, J. I. and G. J. Bowen. 2001. Paleocene-Eocene microvertebrates in Freshwater Limestones of the Willwood Formation, Clarks Fork Basin, Wyoming, in Gunnell, G. F., ed., Eocene Biodiversity Unusual Occurrences and Rarely Sampled Habitats, New York, Kluwer Academic/Plenum Publishers, p. 95-129.
Bloch, J. I., K. D. Rose, and P. D. Gingerich. 1998. New species of Batodonoides (Lipotyphla, Geolabididae) from the early Eocene of Wyoming: smallest known mammal? Journal of Mammalogy 79:804-827.
Bown, T. M. and D. M. Schankler. 1982. A review of the Proteutheria and Insectivora of the Willwood Formation (lower Eocene), Bighorn Basin, Wyoming. Bulletin of the U.S. Geological Survey 1523:1-79.
Case, J. A., F. J. Goin, and M. O. Woodburne. 2005. “South American” marsupials from the late Cretaceous of North America and the origin of marsupial cohorts. Journal of Mammalian Evolution 12:461-494.
Cope, E. D. 1873. Third notice of extinct Vertebrata from the Tertiary of the Plains. Palaeontological Bulletin 16:1-8.
Cope, E. D. 1884. The Vertebrata of the Tertiary formations of the West. Report of the U.S. Geological Survey of the Territories. F. V.
Hayden, geologist in charge, Washington, D.C., 1,009 p.
Crochet, J-Y. 1979. Diversité systématique des Didelphidae (Marsupialia) européens Tertiaires. GéoBios 12:365-378.
Eberle, J. and J. E. Storer. 1995. Herpetotherium valens (Lambe), a didelphid marsupial from the Calf Creek Local Fauna (Chadronian), Saskatchewan. Journal of Vertebrate Paleontology 15:785-794.
Fox, R. C. 1983. Notes on the North American Tertiary marsupials Herpetotherium and Peradectes. Canadian Journal of Earth Sciences 20:1565-1578.
Gazin, C. L. 1952. The lower Eocene Knight Formation of western Wyoming and its mammalian faunas. Smithsonian Miscellaneous Collections 177:1-82.
Gazin, C. L. 1958. A review of the middle and upper Eocene primates of North America. Smithsonian Miscellaneous Collections 136:1-112.
Gray, J. E. 1821. On the natural arrangement of vertebrose animals. The London Medical Repository Monthly Journal and Review 15:296-310.
Golz, D. J. 1976. Eocene Artiodactyla of southern California. Natural History Museum of Los Angeles County, Science Bulletin 26:1-85.
Golz, D. J. and J. A. Lillegraven, 1977. Summary of known occurrences of terrestrial vertebrates from Eocene strata of southern California. University of Wyoming Contributions in Geology 15:43-65.
Gunnell, G. F., T. M. Bown, J. H. Hutchison, and J. I. Bloch. 2008. Lipotyphla, in Janis, C. M, G. F. Gunnell, and M. E. Uhen, eds., Evolution of Tertiary Mammals of North America, Cambridge, Cambridge University Press, p. 89-125.
Hough, M. J. 1956. A new insectivore from the Oligocene of the Wind River Basin, Wyoming, with notes on the taxonomy of the Oligocene Tenrecoidea. Journal of Paleontology 30:531-541.
Kelly, T. S. 1990. Biostratigraphy of Uintan and Duchesnean land mammal assemblages from the middle member of the Sespe Formation, Simi Valley, California. Natural History Museum of Los Angeles County, Contributions in Science 419:1-42.
Kelly, T. S. 1992. New Uintan and Duchesnean (middle and late Eocene) rodents from the Sespe Formation, Simi Valley, California. Southern California Academy of Sciences Bulletin 91:97-120.
Kelly, T. S. 2009. A new species of Heliscomys (Rodentia, Heliscomyidae) from the Duchesnean (middle Eocene) Simi Valley Landfill Local

167 KELLY—DUCHESNEAN MAMMALS FROM SIMI VALLEY
Fauna, Sespe Formation, California. Paludicola 7:67-77.
Kelly, T. S. and D. P. Whistler. 1994. Additional Uintan and Duchesnean (middle and late Eocene) mammals from the Sespe Formation, Simi Valley, California. Natural History Museum of Los Angeles County, Contributions in Science 439:1-29.
Kelly, T. S. and D. P. Whistler. 1998. A new eomyid rodent from the Sespe Formation of southern California. Journal of Vertebrate Paleontology 18:440-443.
Kelly, T. S., E. B. Lander, D. P. Whistler, M. A. Roeder, and R. E. Reynolds. 1991. Preliminary report on a paleontologic investigation of the lower and middle members, Sespe Formation, Simi Valley Landfill, Ventura County, California. PaleoBios 13:1-13.
Kielan-Jaworowska, Z., R. L. Cifelli, and Z. Luo. 2004. Mammals from the age of dinosaurs. New York, Columbia University Press, p. 630.
Kirsch, J. A., F. J. Lapointe, and M. S. Springer. 1997. DNA-hybridization studies of marsupials and their implications for metatherian classification. Australian Journal of Zoology 45:211-280.
Korth, W. W. 1994. Middle Tertiary marsupials (Mammalia) from North America. Journal of Paleontology 68:376-397.
Korth, W. W. 2008. Marsupialia, in Janis, C. M, G. F. Gunnell, and M. E. Uhen, eds., Evolution of Tertiary Mammals of North America, Cambridge, Cambridge University Press, p. 39-47.
Krishtalka, L. and R. K. Stucky. 1983a. Revision of the Wind River faunas, early Eocene of central Wyoming, Part 3, Marsupialia. Annals of Carnegie Museum 52:205-227.
Krishtalka, L. and R. K. Stucky. 1983b. Paleocene and Eocene marsupials of North America. Annals of Carnegie Museum 52:229-263.
Krishtalka, L. and R. K. Stucky. 1984. Middle Eocene marsupials (Mammalia) from northeastern Utah and the mammalian fauna from Powder Wash. Annals of Carnegie Museum 53:31-45.
Lambe, L. M. 1908. The Vertebrata of the Oligocene of the Cypress Hills, Saskatchewan. Contributions to Canadian Palaeontology 3:1-65.
Lander, E. B. 2008. Simi Valley Landfill and Recycling Center landfill expansion, Ventura County, California, paleontologic resource impact mitigation program, fourteenth progress report for period January 1, 2003 to December 31, 2007, project 2006-8. Prepared for Waste Management of California, Inc., Simi Valley Landfill and Recycling Center, 35 p.
Lillegraven, J. A. 1976. Didelphids (Marsupialia) and Uintasorex (?Primates) from later Eocene
sediments of San Diego County, California. Transactions of the San Diego Society of Natural History 18:85-112.
Lillegraven, J. A., M. C. McKenna, and L. Krishtalka. 1981. Evolutionary relationships of middle Eocene and younger species of Centetodon (Mammalia, Insectivora, Geolabididae) with a description of the dentition of Ankylodon (Adapisoricidae). University of Wyoming Publications 45:1-115.
Lopatin, A. V. 2006. Early Paleogene insectivore mammals of Asia. Paleontological Journal 40 (Supplement 3):S205-S405.
Luterbacher,H. P., J. R. Ali, H. Brinkhuis, F. M. Gradstein, J. J. Hooker, S. Monechi, J. G. Ogg, J. Powell, U. Röhl, A. Sanfilippo, and B. Schmitz. 2004. The Paleogene period, in Gradstein, F. M., J. G. Ogg, and A. G. Smith, eds., A geologic time scale 2004, Cambrige, Cambridge University Press, p. 384-408.
Marsh, O. C. 1872. Preliminary description of new Tertiary mammals. American Journal of Science 38:81-92.
Marsh, O. C. 1889. Discovery of Cretaceous Mammalia. American Journal of Science 38:81-92.
Marsh, O. C. 1892. Discovery of Cretaceous Mammalia, Part III. American Journal of Science 3:249-262.
Marshall, L. G., J. A. Case, and M. O. Woodburne. 1990. Phylogenetic relationships of the families of marsupials, in Genoways, H. H., ed., Current Mammalogy, Volume 2, New York, Plenum Press, p. 433-505.
Mason, M. A. 1988. Mammalian paleontology and stratigraphy of the early to middle Tertiary Sespe and Titus Canyon Formations, southern California. Ph.D. dissertation, University of California, Berkeley, 257 p.
Matthew, W. D. 1903. The fauna of Titanotherium beds of Pipestone Springs, Montana. Bulletin of the American Museum of Natural History 19:197-226.
Matthew, W. D. 1909. Carnivora and Insectivora of the Bridger Basin, Middle Eocene. Memoirs of the American Museum of Natural History 9:289-567.
Matthew, W. D. 1910. On the skull of Apternodus and the skeleton of a new artiodactyl. Bulletin of American Museum of Natural History 28:33-42.
Matthew, W. D. and W. Granger. 1921. New genera of Paleocene mammals. American Museum Novitates 13:1-7.
McGrew, P. O. 1937. New marsupials of the Tertiary of Nebraska. Journal of Geology 45:448-455.
McGrew, P. O. 1959. The geology and paleontology of the Elk Mountain and Tabernacle Butte area,

PALUDICOLA, VOL. 7, NO. 4, 2010 168
Wyoming. Bulletin of the American Museum of Natural History 117:117-176.
McKenna, M. C. 1960a. The Geolabidinae, a new subfamily of early Cenozoic erinaceoid insectivores. University of California Publications in Geological Sciences 37:131-164.
McKenna, M. C. 1960b. Fossil Mammalia from the early Wasatchian Four Mile Fauna, Eocene of northwest Colorado. University of California, Publications in Geological Sciences 37:1-130.
McKenna, M. C. 1975. Toward a phylogenetic classification of the Mammalia, in W. P. Luckett and F. S. Szalay, eds., Phylogeny of the Primates, New York, Plenum Press, p. 21-46.
McKenna, M. C. and S. K. Bell. 1997. Classification of mammals above the species level. Columbia University Press, New York, p. 631.
McKenna, M. C. and F. Haase. 1992. Marsholestes, a new name for the Eocene insectivoran Myolestes Matthew, not Myolestes Brèthes, 1904. Journal of Vertebrate Paleontology 12:256.
Novacek, M. J. 1976. Insectivora and Proteutheria of the late Eocene (Uintan) of San Diego County, California. Contributions in Science 283:1-51.
Novacek, M. J. 1985. The Sespedectinae, a new subfamily of Hedgehog-like insectivores. American Museum Novitates 2822:1-24.
Osborn, H. F. and J. L. Wortman. 1892. Fossil mammals of the Wasatch and Wind River beds. Collection of 1891. Bulletin of the American Museum of Natural History 4:1-147.
Prothero, D. R., J. L. Howard, and T. H. Huxley Dozier. 1996. Stratigraphy and paleomagnetism of the upper middle Eocene to lower Miocene (Uintan to Arikareean) Sespe Formation, Ventura County, California, in Prothero, D. R. and R. J. Emry, eds., The Terrestrial Eocene-Oligocene Transition in North America, Cambridge, Cambridge University Press, p. 171-188.
Robinson, P. and D. G. Kron. 1998. Koniaryctes, a new genus of apternodontid insectivore from lower Eocene rock of the Powder River Basin, Wyoming. Contributions to Geology, University of Wyoming 32:187-190.
Rothecker, J. and J. E. Storer. 1996. The marsupials of the Lac Pelletier Lower Fauna, middle Eocene (Duchesnean) of Saskatchewan. Journal of Vertebrate Paleontology 16:770-774.
Rudman, S. 1981. Observations on Uintasorex parvulus (Mammalia, Primates) from The Bridger Formation, Southwestern Wyoming. MS thesis, University of Wisconsin, Milwaukee.
Sánchez-Villagra, M., S. Ladevèze, I. Horovitz, C. Argot, J. J. Hooker, T. E. Macrini, T. Martin, S. Moore-Fay, C. de Muizon, T. Schmelzle, and R. J. Asher. 2007. Exceptionally preserved North
American Paleogene metatherians: adaptations and discovery of a major gap in the opossum fossil record. Biology Letters 3:318-322).
Silcox, M. T. and K. D. Rose. 2001. Unusual vertebrate microfaunas from the Willwood Formation, early Eocene of the Bighorn Basin, Wyoming, in Gunnell, G. F., ed., Eocene Biodiversity Unusual Occurrences and Rarely Sampled Habitats, New York, Kluwer Academic/Plenum Publishers, p. 131-164.
Silcox, M. T. and G. F. Gunnell. 2008. Pleisadapiformes, in Janis, C. M, G. F. Gunnell, and M. E. Uhen, eds., Evolution of Tertiary Mammals of North America, Cambridge, Cambridge University Press, p. 89-125.
Stock, C. 1932. Eocene land mammals on the Pacific Coast. Proceedings of the National Academy of Sciences 18:518-523.
Stock, C. 1935. Insectivora from the Sespe uppermost Eocene, California. Proceedings of the National Academy of Sciences 21:214-219.
Stock, C. 1936. Sespe Eocene didelphids. Proceedings of the National Academy of Sciences 22:122-124.
Stock, C. and E. L. Furlong. 1922. A marsupial from the John Day Oligocene of Logan Butte, eastern Oregon. University of California Publications in Geological Sciences 13:311-317.
Storer, J. E. 1995. Small mammals of the Lac Pelletier Lower Fauna, Duchesnean of Saskatchewan, Canada: insectivores and insectivore-like groups, a plagiomenid, a microsyopid, and Chiroptera, in Sarjeant, W. A. S., ed., Vertebrate Fossils and the Evolution of Scientific Concepts, Amsterdam, Gordon and Breach Publishers, p. 595-615.
Szalay, F. S. 1969. Uintasoricinae, a new subfamily of early Tertiary mammals (?Primates). American Museum Novitates 2363:1-36.
Trouessart, E. L. 1879. Cataloque des mammiféres vivants et fossils, Revue et Magazine de Zoologie, Pure et appliqué (Paris), Marsupialia, Series 3, VII:223, 279 footnote.
Troxell, E. L. 1923. A new marsupial. American Journal of Science 5:507-510.
Walsh, S. L. 1991. Eocene mammal faunas of San Diego County, in Abbott, P. L. and J. A. May, eds., Eocene Geologic History San Diego Region. Pacific Section, Society of Economic Paleontologists and Mineralogists Book 68, p. 161-177.
Walsh, S. L. 1996. Middle Eocene mammalian faunas of San Diego County, California, in Prothero, D. R. and R. J. Emry, eds., The Terrestrial Eocene-Oligocene Transition in North America, New York, Cambridge University Press, p. 75-119.

169 KELLY—DUCHESNEAN MAMMALS FROM SIMI VALLEY
Wood, C. B. and W. A. Clemens. 2001. A new
specimen and a functional reassociation of the molar dentition of Batodon tenuis (Placentalia, incertae sedis), latest Cretaceous (Lancian), North America. Bulletin of the Museum of Comparative Zoology 156:99-118.

Paludicola 7(4):170-180 May 2010 © by the Rochester Institute of Vertebrate Paleontology
170
NEW MATERIAL FROM THE TYPE SPECIMEN OF MEGALNEUSAURUS REX (REPTILIA: SAUROPTERYGIA) FROM THE JURASSIC SUNDANCE FORMATION, WYOMING
William R. Wahl1, Judy A. Massare2, and Mike Ross3
1Wyoming Dinosaur Center, 110 Carter Ranch Road, Thermopolis, Wyoming 82243, [email protected] 2Earth Sciences Dept, SUNY College at Brockport, Brockport, New York 14420, [email protected]
3 2614 Navarre Street, Casper, Wyoming 82601, [email protected]
ABSTRACT In 2008, an articulated distal forelimb of the type specimen of the large pliosauromorph Megalneusaurus rex (UW 4602)
was discovered adjacent to the original excavation pit from which two hindlimbs had been collected in 1895. The new material includes six complete or partial carpals, four metacarpals, and nearly all of the phalanges. Although the new bones were damaged by weathering and gypsum crystal growth, the articulated arrangement of the bones is preserved. Important features include (1) broadly flared metacarpals that articulate proximally and distally with the adjacent metacarpals; (2) curved facets on phalanges that are concave proximally and convex distally; (3) laterally interlocking phalanges between digits I and II and between digits IV and V for the entire length of the outer digits; and (4) tightly articulated, interlocking phalanges among all digits distal to the 3th phalanx. Examination of the material collected in 1895 indicates that a similar structure occurred on the hindlimb as well. The results of this arrangement are rigid, reinforced leading and trailing edges of the flipper, as well as a stiff distal end. During swimming, the limb moved as a rigid unit, with no flexibility at any articulation distal to the head of the propodial. The stiff flipper generated thrust by pushing backward and downward against the water during power stroke; and generated lift when the limb was rotated and moved forward and upward during the recovery stroke.
INTRODUCTION In 1895, Wilbur Knight discovered the type specimen of Megalneusaurus rex (UW 4602; for institutional abbreviations see below), originally described as Cimoliosaurus rex, the only Jurassic pliosauromorph known from North America (Knight, 1895; Wahl et al, 2007). Knight collected a nearly complete, articulated limb including the propodial, most of which is presently on display at the Geological Museum, University of Wyoming. Material including cervical, dorsal, and caudal vertebrae, neural arches, a large portion of the limb girdle, and ribs were also reported in the paper which erected a new genus for the species (Knight, 1898). This material has since been lost. Knight (1898) did not mention nor figure the disarticulated second limb of the same specimen, which includes a complete set of epipodials, mesopodials, metapodials, and phalanges of the same individual, which is presently housed in the UW collections. Only two other specimens from Wyoming can be referred to M. rex: a weathered neural arch, collected as float from Natrona County (UW 24238), and a very weathered propodial fragment from Hot Springs County (WDC SS019). Neither specimen has been described in publications. Another specimen of M. rex comprising a partial propodial has been reported
from the Upper Jurassic Naknek Formation of Alaska (Weems and Blodgett, 1994). In 1996, the excavation site of the type specimen of Megalneusaurus rex was relocated in the Gas Hills area of eastern Fremont County, WY (Wahl, et al., 2007). The discovery revealed that UW 4602 is from the upper part of the Redwater Shale Member of the Sundance Formation, about 10 m below the Windy Hill Sandstone. The specimen is thus of Oxfordian age (Kvale, et al., 2001). We report here on the discovery in 2008 of a third limb from the type specimen of Megalneusaurus rex (UW 4602).
Institutional Abbreviations—AMNH American Museum of Natural History, NY, U.S.A.; BMNH, British Museum of Natural History, London, U.K.; UW, University of Wyoming, Laramie, WY, U.S.A.; WDC, Wyoming Dinosaur Center, Big Horn Basin Foundation, Thermopolis, WY, U.S.A.;
MATERIAL The new limb, which is missing the propodial and epipodials, is 106 cm long from the proximal edge of the intermedium to the distal end. The limb has five digits, with the first considerably shorter than the others. It is missing the metapodial and half of the first phalanx of digit V, but is otherwise fairly complete, but

WAHL, ET AL.—TYPE SPECIMEN OF MEGALNEUSAURUS REX 171
poorly preserved. The most distal phalanges of digits II, III, IV, and V are not discernable as individual bones. Their proximal and distal ends may be inferred from slight ‘bumps’ in the brown, iron-rich matrix at the distal end of the limb. This matrix is distinct from that surrounding the limb, and we infer that it is derived from weathering of bone. The history of the collection of UW 4602 is unusual (Wahl, et al., 2007). Wilbur Knight (UW) had been in the field with a man named Stewart, who worked for E. D. Cope at the AMNH. Knight had left Stewart and was returning home when he discovered the specimen. He borrowed a wagon from a nearby rancher to take the specimen back to Laramie. He intended to return with more packing material and remove the rest of the bones, but before he could return, Stewart removed material that Knight had left behind and sent it to Cope at the AMNH (Wahl, et al., 2007). We suspect that the third limb reported here had been exposed and reburied by either Knight or Stewart because we removed soil rather than in situ shale from the upper part of the specimen. Adjacent to the limb, shale was encountered at a shallower depth. Furthermore, the proximal limb elements were iron-stained and somewhat weathered, suggesting partial exposure, perhaps during the initial exploration of the site in 1895. In addition, after removing some of the soil, we found a broken phalanx that was missing its proximal end. The missing piece may have been collected by Knight or Stewart. At least one phalanx of UW 4602 in the UW collections was incorrectly assembled from pieces of phalanges of two different digits; thus some partial phalanges were collected in 1895. Likewise, an old nail, confidently dated to the late 1800’s, was found lying under the soil, directly on top and in contact with the elements of the new limb. This may have been accidentally dropped or intentionally left as a marker by someone expecting to return to the site. The spoils pile from the original excavation, now reduced to a low mound, ringed the old pit. The new limb was found less than 2 m SE of the original excavation pit, with the proximal row of bones exposed at the surface. The limb was lying on its ventral surface along a bedding plane, with the long axis oriented almost N-S. It dipped into the ground at an angle of 10- 15. A hard, irregularly cross-bedding sandstone was under and around the limb. It was about 8 cm thick under the limb and extended eastward beyond the limb, thickening to more than 20 cm. The sandstone did not occur on the west side of the limb. A similar sandstone lens is exposed on the side of the 1895 pit. We attribute its presence to the carcass acting as a baffle, causing eddies and scouring on the leeward side and collecting coarser sediments (Schafer, 1972). Stratigraphically above the limb was a dark gray shale, which was overlain by a thinly bedded, cross-bedded,
friable sandstone. A similar stratigraphy was exposed on the side of the old excavation pit. The shallow depth of the in situ matrix was such that some bone surfaces were damaged by plant roots. Calcareous concretion filled some of the spaces between the limb elements. This matrix was free of cephalopod hooklets that were present in large amounts at the site (Wahl, et al., 2007), possibly suggesting that the limb material had undergone some early diagenesis before the complete breakdown of the carcass. The underside of the flipper was covered with a layer of gypsum, and the articular facets of many phalanges and some of the bone surfaces have been destroyed by this crystal growth. Furthermore, space that had developed in the matrix during diagenesis was also filled with gypsum crystals. Several phalanges were stained a patchy, yellow-orange color on their surfaces because of contact with the iron-rich shale above. __________________________________________ TABLE 1. Measurements of individual bones of Megalneusaurus rex (UW 4602). All measurements in cm. Measurements and bone names of the 1895 specimens are those published by Knight (1898), although our assessment is that these specimens are hindlimbs. Note that the measurements of Knight (1898) are higher than those of the 2008 specimen described herein.
cm 1895
specimen 2008
specimen"ulnare" length 9.5 5.0+? width 13.0 broken "intermedium" length 10.0 7.4 width 14.0 11.5 "radiale" length 8.5 7.5 width 13.5 10.9 The in situ limb had a slight curvature to it, which we interpret as a posterior curvature, curving towards the old excavation pit where the two limbs were removed in 1895. Between the new limb and the old excavation pit is an area from which we have recovered numerous chunks of concretion containing cephalopod hooklets, including some very densely packed pieces. We interpret these pieces as stomach contents of Megalneusaurus rex (Wahl, et al., 2007), as the preservation is similar to what has been reported for an ichthyosaur and other plesiosaurs from the Redwater Shale (Massare and Young, 2005; Wahl, 2006). The relative positions of the excavation pits and presumed stomach contents would suggest that the rear of the animal was towards the 1895 excavation pit. Thus based on the field relationships, the new discovery is most likely a forelimb. Andrews (1913) and Tarlo (1960) indicated that the the hindlimb is larger than the forelimb for Upper Jurassic pliosaurids. More recently, O’Keefe and

172 PALUDICOLA, VOL. 7, NO. 4, 2010
Carrano (2005) suggested that all large pliosauromorphs probably had larger hindlimbs than forelimbs. Table 1 presents a comparison of individual bones, using measurements from Knight (1898) and our measurements of the new material. The new material is consistently smaller. We estimate that the new limb is approximately 15% smaller than Knight’s material, further suggesting that the 2008 material is a forelimb. Knight originally described the 1895 material as a hindlimb in a brief preliminary report (Knight, 1895), but in a second, more detailed paper, he identified it as a forelimb (Knight, 1898). It now appears that his first identification was correct.
DESCRIPTION Material Collected in 2008: Forelimb—The new limb is missing the humerus, radius, and ulna (Figure 1). From the shape of the proximal edges of the carpals, it appears that the radius articulated with both the radiale and the intermedium, whereas the ulna articulated mainly with the long, straight proximal end of the intermedium. The ulnare is broken, but was likely only half the width of the intermedium. A second row of carpals consists of three closely spaced polygonal bones (Figure 2). Metacarpals II, III, and IV are broadly flared proximally and articulate with adjacent metacarpals (Figure 3). Although damaged, metacarpal I shows a short, shelf-like projection in contact with a projection on metacarpal II. This feature is preserved better in the 1895 hindlimb material and is figured below. Metacarpal V and the proximal end of the first phalanx of digit V are missing. Metacarpal V probably articulated with the ulnare as in other pliosauromorphs (Andrews, 1913, figs. 3, 6 ). Digit I is the shortest, digits II and V are of nearly equal length, digit III is longer, and digit IV is the longest. The difference in lengths of digits II-V appears due to a variation in the size of the phalanges as well as in the number of phalanges (Table 2). The phalange count as preserved is I-5, II-8, III-9, IV-8, and V-8. Digit I is complete, although the distal phalanx is very poorly preserved. Only the proximal end of the eighth phalanx is present on digit II. The distal phalanges have been weathered to a dark brown powder possibly due to the accretion of iron-rich minerals or dissolution by plant roots. Irregularities in the brown matrix at the distal end suggest one additional phalanx in digits II, III and V, and two additional phalanges in digit IV. The 1895 hindlimbs may also be missing distal phalanges, but the relative lengths of the digits in the 2008 limb are consistent with the 1895 articulated hindlimb. The phalanges have an hour-glass shape, and all are broken in the middle, narrowest portion of the shaft.
The articular facet between successive phalanges is a curved, sinuous surface (Figure 4), although gypsum crystal growth has damaged the ends of most phalanges of the new specimen. In three dimensions, the phalanges are irregularly concave proximally and irregularly convex distally. Thus it is possible to orient an isolated phalanx if the proximal and distal ends are well-preserved. These are not, however, ball and socket articulations; the facets are complexly curved and do not allow rotation between the bones. In addition, the cross-sectional shapes of the metacarpals and proximal phalanges vary from one digit to the next. These features are better preserved in the 1895 material and are discussed further below. Metacarpal I is shorter than metacarpal II (Figure 3; Table 2). This causes an offset of the phalanges of digit I relative to digit II, such that the concave middle of the shaft on a phalanx fits around the contact between two phalanges on the adjacent digit (Figure 5). A similar offset occurs between digits IV and V. This produces tight, laterally interlocked phalanges on the leading and trailing edges of the flipper for the entire length of digits I and V. This is in contrast to the condition of the inner digits, where gaps occur between the shafts of the first two or three phalanges (Figure 1). The pattern of offset and interlocking phalanges is similar to the articulated hindlimb from 1895 on display at UW, but inexplicably not what is shown in Knight’s (1898) reconstruction. At the third phalanx of the three inner digits, differences in the accumulated length of the phalanges cause an offset such that the hour-glass shapes of phalanges on adjacent digits interlock (Figure 6). This is approximately the position on the limb where we infer that digit I ends. Distally the interlocking is tighter and separation between successive phalanges is reduced to no more than a 1 mm. The phalanges are all dorso-ventrally compressed and take on an oval cross-section.
Material Collected in 1895: Hindlimbs—An articulated hindlimb of UW 4602 collected in 1895 is on display at the Geological Museum at UW. It includes the tibia and fibula, but not the femur, and a nearly complete distal portion of the limb. The phalange count as mounted is I-5, II-9, III-9, IV-9, and V-8. The distal phalanges of digits II and IV are fragments only. Three phalanges on digit I have been reconstructed, but we were unable to examine the mount closely to determine if any other elements were casts or reconstructed. The mount shows the same interlocking pattern of phalanges that is preserved in the forelimb, in contrast to the published reconstruction (Knight, 1898). Because of this, we suspect that the mount is in its original articulated state rather than reconstructed, but we did not have access to the

WAHL, ET AL.—TYPE SPECIMEN OF MEGALNEUSAURUS REX 173
FIGURE 1. UW 4602 Megalneusaurus rex. Forelimb of type specimen discovered in 2008, shown in ventral view. Scale bar = 30 cm. ________________________________________________________________________________________________________________________
specimen to confirm this. The second hindlimb of UW 4602 from 1895 is disarticulated and stored in the UW collections. Knight (1898) indicated that the propodial was 99.1 cm long and figured it as a complete bone. No complete propodials are in the UW collections. Presently there are three pieces of propodial that do not now fit together. One piece is a trilobed head of a femur (Brown, 1981, fig. 176), with numerous holes on the surface, perhaps indicating a cartilagenous cap (Figure 7A, B). The propodial shaft cross-section is quadrate, although Knight (1898) described it as ‘prismatic’. A second piece is the well-preserved distal end of a propodial, which we also identify as a femur (Figure 8). The facets are concave and articulate with the convex surfaces of the tibia and fibula in the UW collections. A third fragment of propodial is from the distal end of the shaft, but is broken at both its distal and proximal ends (Figure 9). It is a smaller, more slender bone than the distal femur, and we suggest that it may be a portion of a humerus. Based on the orientation of the forelimb excavated in 2008, it is possible that the humerus, or a portion of it, was lying on or near the surface and could have been collected in 1895 by Knight or Stewart. A feature noted by Knight (1898) but not described in detail, are interlocking projections on the tibia and fibula. The tibia has a bony projection on its medial margin that fits into a depression on the fibula at the proximal end of the medial margin (Figure 10; also see articulation in Figure 8). Distally, the fibula has a projection that fits into a depression on the tibia. Similar interlocking articulations occur between the tibia and fibula of Peloneustes philarchus (BMNH 2440) from the Oxford Clay of England (WRW pers. obs.). Andrews (1913, fig. 23) shows the articulation
FIGURE 2. UW 4602 Megalneusaurus rex 2008 forelimb. Proximal end in ventral view, showing the articulated radiale (top right), intermedium, and partial ulnare (top left), as well as second row of three carpals (below), the first of which (lower right) is a fragment. Scale bar = 10 cm. ___________________________________________________
in a diagram, but he does not describe the projections and depressions. Based on the articulated forelimb, we reconstructed the disarticulated hindlimb in the UW collection (Figure 11). The difference in size and cross-sectional shape of the metatarsals and phalanges allows the isolated bones to be put together in articulated positions with considerable confidence. Some phalanges had been broken at the narrowest portion of the shaft and reassembled. One was assembled from separate pieces of phalanges from different digits, as the proximal and distal ends have different cross-sectional shapes. When the bones are articulated, two proximal phalanges are “left over”. In addition, we have collected four badly weathered fragments of phalanges from the spoils pile

174 PALUDICOLA, VOL. 7, NO. 4, 2010
TABLE 2. Measurements of metacarpals and phalanges of 2008 material of Megalneusaurus rex (UW 4602). All measurements in cm. frag: only a fragment is preserved; pres: presence inferred from irregularities in the iron-rich brown matrix distal to previous element.
Digit I Digit II Digit III Digit IV Digit V
element length
cm width
cm length
cm width
cm length
cm width
cm length
cm width
cm length
cm width
cm metacarpal 9.0 5.6 9.7 4.8 9.8 4.3 10.3 3.8 missing phalanx 1 7.7 3.5 9.0 3.4 10.1 3.3 10.2 3.3 frag phalanx 2 7.7 1.9 8.2 2.7 8.3 3.2 8.5 3.1 9.6 2.5 phalanx 3 6.9 1.7 8.0 2.6 8.3 2.6 8.8 2.7 9.0 2.0 phalanx 4 8.4 1.7 7.3 2.3 8.0 2.2 8.0 2.2 8.5 2.0 phalanx 5 frag 7.0 1.9 6.9 1.5 7.3 2.1 7.7 1.9 phalanx 6 6.3 1.9 5.6? 6.5 1.4 7.0 1.5 phalanx 7 5.6 5.0? 6.0 6.5 6.0? phalanx 8 frag frag frag frag phalanx 9 pres ? pres pres pres ? phalanx 10 pres ? pres ?
surrounding the old pit. As neither hindlimb at UW appears to be missing proximal phalanges, this may be evidence that elements from more than two limbs were exposed and collected in 1895. Our reconstruction shows a gentle posterior curvature to the hindlimb, similar to that preserved in the forelimb and shown in the hindlimb on display at UW. The maximum width of the limb is located near the distal end of the femur (Figure 11). Some distal phalanges are missing. ____________________________________________
FIGURE 3. UW 4602 Megalneusaurus rex 2008 forelimb. Metacarpals I (left), II, III, and IV (left), illustrating the flared proximal ends that articulate with the adjacent metacarpal. Scale bar = 10cm. ____________________________________________________
The tarsals consist of six polygonal bones arranged in two rows, comparable to the forelimb. The tibiale and intermedium are wider than long, and articulate along a straight contact with the tibia and fibula. The
FIGURE 4. UW 4602 Megalneusaurus rex 2008 forelimb. Curved contact between successive phalanges, showing the convex distal end of one phalange (right) and concave distal end of the successive one (left). The distal ends of the right phalanges have been damaged by gypsum crystal growth. The proximal end of the flipper is to the right. Scale bar = 10 cm.
_____________________________________ fibulare is longer than wide and articulates with a small facet on the trailing edge of the fibula. The second row of tarsals articulate tightly with the first. Their distal surfaces are convex and articulate with concave surfaces of metatarsals II, III, and IV. Metatarsal V

WAHL, ET AL.—TYPE SPECIMEN OF MEGALNEUSAURUS REX 175
articulates with the fibulare, but the convex articular surface is on the metatarsal. Metatarsal I is smaller than the others and has a narrow, shelf-like projection that contacts the expanded proximal end of the metatarsal II (Figure 12). Peloneustes philarchus (BMNH 2440) also shows this feature (WRW pers. obs.), although it was not noted in Andrews (1913). ____________________________________________
FIGURE 5. UW 4602 Megalneusaurus rex 2008 forelimb. Leading edge of forelimb, showing portion of digits I (bottom), II, and III (top). Note the tightly articulated phalanges of digits I and II, and the asymmetric shape of the phalanges of digit I. The proximal end of the flipper is to the right. Scale bar = 10 cm __________________________________________________
Most of the phalanges have an hour-glass shape, and a few had been broken in the middle and reassembled. The pattern of an irregularly concave proximal facet and an irregularly convex distal facet continues for the at least the first three phalanges on digit II, III, and IV, similar to what is preserved on the new forelimb. Because metatarsal I is much shorter than metatarsal II, the phalanges of digits I and II are offset and interlock along their lateral margin as in the new forelimb. The phalanges of digit IV and digit V similarly interlock laterally because of the offset produced by the proximal shift of metatarsal V to articulate with the fibulare. Phalanges of digit V of the hindlimb are narrower and longer than those of the other digits, and some lack the pronounced hour-glass shape. They are also longer and narrower than those of digit V of the forelimb. The metatarsals and phalanges vary in the cross-sectional shape at their proximal and distal ends (Figure 13). Digit I metatarsal and phalanges are triangular in cross-section, with the tapered end of the triangle towards the leading edge of the flipper. The cross-section is quadrate in digits II and III, triangular in digit IV, and oval in digit V, with the short axis oriented dorso-ventrally. When articulated as they
would have been in life, the resulting dorso-ventral cross-section of the flipper is a streamlined, hydrofoil shape (Figure 13). Similar differences in cross-sectional shapes of the digits occur on the forelimb, but the preservation is not as good. The articulation of the phalanges on adjacent digits form a convex upper (dorsal) surface and a slightly concave lower (ventral) surface to the flipper starting at the metapodial row. The ventral concavity becomes more pronounced distally, beginning at the 3th phalanx, where the hour-glass shaped phalanges of adjacent digits tightly interlock. Thus the limb as a whole had a slight ventral curvature and a hydrofoil shape in cross-section. ____________________________________________
FIGURE 6. UW 4602 Megalneusaurus rex 2008 forelimb. Laterally interlocking hour-glass shaped phalanges of digit II (bottom), III, IV, and V (top). The proximal end of the flipper is to the right. The arrow indicates the proximal end of the fifth phalanx of digit II; above is the distal end of the fourth phalanx of digit III, above again is the fifth phalanx of digit IV, and above again is the distal end of the fourth phalanx of digit V. Scale bar = 10 cm
_____________________________________
FUNCTIONAL SIGNIFICANCE Based on the material collected in 2008, a new reconstruction of the distal forelimb of Megalneusaurus rex is presented here (Figure 14). The morphology of both the forelimbs and hindlimbs of M. rex show features that restrict movement between elements and reinforce the flipper as a whole. One such feature is the concavo-convex articulation of the tibia and fibula with the femur. Another is the proximal bony projection on the tibia that fit into a depression on the fibula (Figure 8). At their distal contact, a less prominent projection on the fibula fits into a slight depression in the tibia. Together these features reinforced and immobilize the

176 PALUDICOLA, VOL. 7, NO. 4, 2010
A B
FIGURE 7. UW 4602 Megalneusaurus rex 1895 material. A. Proximal end of the head of the femur, collected by Knight. An outline is figured in Knight, 1895. B. Femur head in lateral view. Scale bar = 10cm. ________________________________________________________________________________________________________________
articulation, preventing movement of elements in both the paraxial (forward and backward) and dorso-ventral (up and down) planes. The proximally flared, tightly articulated metapodials similarly prevent paraxial and dorso-ventral flexion (Figure 3). The sinuous articular facets of the phalanges (Figure 4) most likely prevented torsional rotation between successive phalanges and reduced flexion in the longitudinal plane. The laterally interlocking phalanges distal to the 3rd phalanx prevented paraxial movement of individual elements at the distal end of the limb. The streamlined hydrofoil or blade shape of the limb was formed by the bony elements themselves, without the need of soft tissue to mold the skeletion into a hydrofoil shape. The tight packing of limb elements left little room for soft tissue between elements (Figs. 1-5). The wedge shaped cross-section of digits I and II, coupled with their tightly interlocking phalanges would preventing flexion in the dorso-ventral, paraxial, and longitudinal planes. This strengthened and stiffened the leading edge of the flipper. Similarly, the offset of phalanges and subsequent lateral interlocking of digits IV and V stiffened and strengthened the trailing edge. Distally, tightly interlocking, dorso-ventrally flattened phalanges formed a flat, rigid flipper. Thus every edge of the flipper meeting the resistance of the water was wedge shaped or flattened, with interlocking contacts of successive repeated shapes. The cross-sectional shape is a hydrofoil, but from the first row of phalanges to the distal narrowing tip, the shape is also bow-like, with a slightly concave ventral surface. This concavity from the wedge-shaped
leading edge to the trailing edge is a by- product of the close packing of the elements (Figure 13). Such a slight ‘cupping’ or ‘sculling’ shape would reinforce the flipper against dorsal-ventral stresses. ___________________________________________
FIGURE 8. UW 4602 Megalneusaurus rex 1895 disarticulated material. Distal end of the femur shown articulated with tibia (right) and fibula (left) in ventral view. Scale bar = 10 cm.

WAHL, ET AL.—TYPE SPECIMEN OF MEGALNEUSAURUS REX 177
FIGURE 9. UW 4602 Megalneusaurus rex 1895 material. Fragment of (?)humerus. The bone is dorso-ventrally flattened, so is from the distal portion of the shaft. Scale bar = 10 cm. _____________________________________________________
Whether this flipper structure is typical of Jurassic pliosauromorphs remains to be seen, as articulated limbs preserved in situ are unusual. Tightly packed digits with phalanges in lateral contact has been reported in the Cretaceous pliosauromorph Edgarosarus (Druckenmiller, 2008, fig. 16). Two other Creteceous pliosauromorphs, Trinacromerum and Nichollssaura, have tightly interlocking phalanges between adjacent digits, similar to Megalneusaurus (Adams, 1997; Druckenmiller and Russell, 2008; Druckenmiller, 2009). In addition, Trinacromerum has tongue and groove articulations that prevented movement at articular surfaces and stiffened the limb (Adams, 1997). Adams (1997) suggested that these features of Trinacromerum were adaptations for high speed swimming because they allowed the fin to lengthen (relative to body size) still maintain rigidity. In Megalneusaurus, the rigid flipper may likewise have been an adaptation for high speed swimming (see discussion of aspect ratio below), but it may have also been a scaling effect related to its large size. The limbs of plesiosaurs have long been interpreted as functioning as ‘wings’ for underwater flight (Robinson, 1975; Godfrey, 1984; Riess and Frey, 1991). Underwater flight sensu stricto, however, requires a figure-eight motion in a vertical plane, which was impossible in plesiosaurs because the limb could not be elevated above the horizontal (Tarsitano and Reiss, 1982; Godfrey, 1984). Instead, plesiosaur ‘flight’ comprised a backward and downward power
stroke that generated thrust, coupled with a forward, and presumably upward, recovery stroke that generated lift (Godfrey, 1984; Reiss and Frey, 1991). The construction of the limb of Megalneusaurus rex suggests that the limb was inflexible at every joint, and that it moved as a rigid unit from the articulation with the limb girdles. The stiffening of both the leading and trailing edges and the slight ventral curvature suggest that the power stroke of Megalneusaurus rex was drag-based, generating thrust by pushing against the water, consistent with earlier interpretations (Godfrey, 1984; Reiss and Frey, 1991). The streamlined cross-section and overall hydrofoil shape of the limb would have generated lift when the limb was rotated and moved forward in the water (Godfrey, 1984; Reiss and Frey, 1991). Movement of the neck and large head would have been necessary to keep the animal on a level course in the water. The locomotion of plesiosaurs was neither underwater flight nor rowing, but the limbs of M. rex show features advantageous to both modes. Plesiosaur locomotion, however, was unique among extant and extinct tetrapods, and a comprehensive analysis of the hydrodynamics has yet to be done. ____________________________________________
FIGURE 10. UW 4602 Megalneusaurus rex 1895 material. Medial edge of the fibula. (ventral view) with proximal end to the right, shows a depression (right arrow) into which a projection on the tibia fits, and a projection (left arrow) that fits into a depression on the tibia. Also note the articulation between the tibia and fibula in Fig. 7. Scale bar = 5 cm
_____________________________________
The details of propulsion with two sets of identical limbs is not understood. Brown (1981) suggested that only the front flippers were used for propulsion, while the hind flippers were extended backward and held against the body. Recent experimental work by Long, et al. (2006), demonstrated that for flapping underwater flight at a steady cruising speed, using one pair of limbs is more efficient than two. Reiss and Frey

178 PALUDICOLA, VOL. 7, NO. 4, 2010
FIGURE 11. UW 4602 Megalneusaurus rex 1895 material. Individual flipper bones of the hindlimb in ventral view, shown in articulation, based on the morphology of the forelimb. Note the backward curvature and the interlocking phalanges on the leading and trailing edges of the limb. Scale = 30 cm. _____________________________________________________________________________________________________________
FIGURE 12. UW 4602 Megalneusaurus rex 1895 material. Disarticulated metatarsals of digits II (left) and I (right) show bony processes that articulated as show. Note that the shorter metatarsal of digit I would result in the phalanges of digit I being offset with respect to those of digit II. Scale bar = 5 cm. ____________________________________________________
(1991) argued that for a large animal that is not a true underwater flier, a rapid loss of momentum would occur during the recovery stroke. They suggested (Frey and Reiss, 1982; Frey and Reiss, 1991) that plesiosaurs moved their front and back pairs of limbs alternately, such that when one pair was in the power stroke the second pair was in the recovery stroke. Thus thrust generation would be continuous and the speed would not drop during the recovery phase (Frey and Reiss, 1982; Reiss and Frey, 1991). Fur seals and sea lions add a glide phase between the power stroke and recovery stroke in routine swimming (JM pers. obs.),
and this may have been another option for plesiosaurs. Any one of these gaits may have been employed for steady cruising. Long, et al. (2006) found that two pairs of limbs, moving in phase, were superior to one pair for rapid acceleration and quick stopping. Plesiosaurs likely employed different gaits for different swimming activities. Knight (1898) measured the length of propodial as 99.1 cm and its distal width as 36.5 cm. The entire hindlimb of Megalneusaurus rex is 220.9 cm long (Knight, 1898). Using these measurements and the equation developed by O’Keefe (2001) would give an aspect ratio of 8.0 for the hindlimb. The aspect ratio of the forelimb is more difficult to estimate. From the piece of presumed humerus in the UW collection, we estimate that the distal humerus width was 85% that of the femur. The 2008 limb material is 106 cm long, but does not include the epipodials. Using these two measurements, the forelimb aspect ratio would be at least 8.2. If the length of an epipodial was included, this figure would be higher. These aspect ratios are within the typical pliosauromorph range (O’Keefe, 2001; O’Keefe and Carrano, 2005). For comparison, Liopleurodon, a large pliosauromorph from the Callovian stage Oxford Clay Formation, has an aspect ratio of 7.83 for the forelimb and 7.85 for the hindlimb (O’Keefe, 2001). O’Keefe (2001) interpreted this flipper shape as consistent with a high speed, maneuverable pursuit predator. Cephalopod hooklets thought to be from UW 4602 suggest that belemnites were part of the diet of Megalneusaurus rex. Belemnites are by far the most common possible prey item in the Sundance

WAHL, ET AL.—TYPE SPECIMEN OF MEGALNEUSAURUS REX 179
FIGURE 13. UW 4602 Megalneusaurus rex 1895 material. View of distal ends of the metatarsals in articulation positions, resting on their dorsal surfaces. Digit I, on the leading edge of the flipper, is to the right. The difference in cross-sectional shape also occurs in the proximal phalanges. The digit V metapodial (left) is offset proximally (into the page), as it would have been during life. Note the hydrofoil shaped cross-section. Scale bar = 5 cm. _____________________________________________________________________________________________________________
F179ormation. Long distance cruising to locate shoals of belemnites may have been the hunting style of M. rex, and might explain why such a large predator was in the relatively shallow Sundance Sea. M. rex , however, was not as maneuverable as the cryptoclidid plesiosaurs (O’Keefe, 2001) or ophthalmosaurid ichthyosaurs (Massare et al., 2006) with whom it shared the sea. It may therefore have had to rely on an ambush attack to capture these larger prey items, as suggested by the results of Long et al. (2006).
ACKNOWLEDGEMENTS The specimen was collected under permit #PA05-WY-131 from the Bureau of Land Management, Wyoming State Office, and we thank Dale Hanson. Craig Bromley, and the BLM for the opportunity to reopen Wilbur Knight’s excavation site. We appreciate the thoughtful reviews by F. R. O’Keefe, L. F. Noè, and P. S. Druckenmiller, which greatly improved the clarity of this paper. Thanks also to D. Lomax, C. Racay, D. Roth, and J. Turnbull for help in the excavation and removal of the specimen. The Wyoming Dinosaur Center generously provided space and equipment for the preparation of the specimen. Thanks also to M. Cassiliano for access to material in the UW collections.
LITERATURE CITED Adams, D. A. 1997. Trinacromerum bonneri, new
species, last and fastest pliosaur of the Western Interior Seaway. Texas Journal of Science, 49: 179-198.
Andrews, C. W. 1913. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay, Part II.
British Museum (Natural History), London. 206 pp., 13 pl.
Brown, D. 1981. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bulletin of the British Museum of Natural History 35: 1-347.
Druckenmiller, P. S. 2009. The new plesiosaurian genus Nichollssaura from Alberta: Canada, replacement name for the preoccupied genus Nichollsia. Journal of Vertebrate Paleontology, 29: 276.
Druckenmiller, P. S. and A. P. Russell. 2008. Skeletal anatomy of an exceptionally complete specimen of a new genus of plesiosaur from the Early Cretaceous (Early Albian) of north-eastern Alberta, Canada. Palaeontographica Abteilung A 283: 1-33.
Frey, E. and J. Riess. 1982. Considerations concerning plesiosaur locomotion. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 164: 193-194.
Godfrey, S. J. 1984. Plesiosaur subaqueous locomotion: a reappraisal. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 11: 661-672.
Knight, W. C. 1895. A new Jurassic plesiosaur from Wyoming. Science, new series, 2: 449.
Knight, W. C. 1898. Some new Jurassic vertebrates from Wyoming. American Journal of Science, 4: 378-381.
Kvale, E. P., G. D. Johnson, D. L. Mickelson, K. Keller, L. Furer, and A. W. Archer. 2001. Middle Jurassic (Bajocian and Bathonian)

180 PALUDICOLA, VOL. 7, NO. 4, 2010
dinosaur megatracksites, Bighorn Basin, Wyoming, U.S.A. Palios 16: 233-254.
Long, J. H., J. Schumacher, N. Livingston, and M. Kemp. 2006. Four flippers or two? Tetrapodal swimming with an aquatic robot. Bioinspiration and Biomimetics 1: 20-29.
___________________________________________________
FIGURE 14: Reconstruction of distal forelimb, from carpals to distal phalanges, of Megalneusaurus rex (Knight, 1895), based on 2008 forelimb of the type specimen UW 4602. Shaded bones are conjectural. Scale bar = 30 cm
Massare, J. A., E. A. Buchholtz, J. Kenney, and A.-M. Chomat. 2006. Vertebral morphology of Ophthalmosaurus natans (Reptilia: Ichthyosauria) from the Jurassic Sundance Formation of central Wyoming, Paludicola 5: 242-254.
Massare, J. A. and H. A. Young. 2005. Gastric contents of an ichthyosaur from the Sundance Formation (Jurassic) of central Wyoming. Paludicola 5: 20-27.
O’Keefe, F. R. 2001. Ecomorphology of plesiosaur flipper geometry. Journal of Evolutionary Biology 14: 987-991.
O’Keefe, F. R. and M. T. Carrano. 2005. Correlated trends in the evolution of the plesiosaur locomotor system. Paleobiology, 31: 656-675.
Reiss, J. and E. Frey. 1991. The evolution of underwater flight and the evolution of plesiosaurs, p. 131-144 in J. M. V. Rayner and R. J. Wooten (eds.) Biomechanics in Evolution, Society for Experimental Biology Seminar Series #36, Cambridge University Press.
Robinson, J. A. 1975. The locomotion of plesiosaurs. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 149: 286-332.
Schafer, W. 1972. Part C. Death, disintegration and burial: I. Vertebrata, p. 10-90 in G. Y. Craig, (ed.), Ecology and Paleontology of Marine Environments, University of Chicago Press, Chicago.
Tarlo, L. B. 1960. A review of the Upper Jurassic pliosaurs. Bulletin of the British Museum of Natural History 4: 1-189.
Tarsitano, S. and J. Reiss. 1982. Plesiosaur locomotion-underwater flight versus rowing. Neues Jahrbuch für Geologie und Paläeontologie, Abhandlungen, 164: 188-192.
Wahl, W. R. 2006. A juvenile plesiosaur (Reptilia: Sauropterygia) assemblage from the Sundance Formation (Jurassic), Natrona County, Wyoming. Paludicola 5: 255-261.
Wahl, W. R., M. Ross, and J. A. Massare. 2007. Rediscovery of Wilbur Knight’s Megalneusaurus rex site: new material from an old pit. Paludicola 6: 194-204.
Weems, R. E and R. B Blodgett. 1994. The pliosaurid Megalneusaurus: A newly recognized occurrence in the Upper Jurassic Naknek Formation of the Alaska peninsula. U.S. Geological Survey Bulletin 2152: 169-175.

181
AUTHOR INDEX TO VOLUME 7 Brinkman, Donald B. A sea turtle skull (Cheloniidae: Carettini) from the Lower Miocene
Nye Formation of Oregon, USA 2 39-46 Evander, Robert L. Craniometry of the Equidae part 1: two dimensional shape analysis 1 1-13 Grady, Frederick See Martin, Marcolini, and Grady Kelly, Thomas S. A new species of Heliscomys (Rodentia, Heliscomyidae) from the
Duchesnean (middle Eocene) Simi Valley landfill local fauna, Sespe Formation, California 3 67-77
A new chub (Actinopterygii, Cypriniformes, Cyprinidae) from the middle Miocene (early Clarendonian) Aldrich Station Formation, Lyon County, Nevada 4 137-157
New records of Marsupialia, Lipotyphla, and Primates from the Duchesnean (middle Eocene) Simi Valley Landfill Local Fauna, Sespe Formation, California 4 158-169
Kihm, Allen J. and Karew K. Schumaker Domnina (Mammalia, Socricomorpha) from the latest Eocene (Chadronian)
Medicine Pole Hills local fauna of North Dakota 1 26-36 Korth, William W. Mammals from the Blue Ash local fauna (late Oligocene), South Dakota.
Rodentia, Part 2: Families Florentiamyidae and Geomyidae 1 14-25 Mammals form the blue Ash local fauna (late Oligocene), South Dakota. Rodentia, Part 3: Family Sciuridae 2 47-60 Mammals from the Blue Ash local fauan (late Oligocene), South Dakota.
Lipotyphla and additional Marsupialia 3 78-88 Mammals form the blue Ash local fauna (late Oligocene), South Dakota. Rodentia, Part 4: Family Aplodontia 3 89-106
Mammals form the blue Ash local fauna (late Oligocene), South Dakota. Rodentia, Part 5: Cricetidae 4 117-136 Marcolini, Federica See Martin, Marcolini, and Grady Martin, James E. Taxonomic note: Parapliosaccomys witteorum (Rodentia, Geomyidae),
a replacement name for Parapliosaccomys wittei Martin, 2008 1 37 Martin, Robert A. Federica Marcolini, and Frederick Grady The early Pleistocene Hamilton Cave muskrats and a review of muskrat size
change through the late Neogene 2 61-66 Massare, Judy A. See Wahl, Massare and Ross Ross, Mike See Wahl, Massare and Ross

182
Schumaker, Karew K. See Kihm and Schumaker Wahl, William R. Taphonomy of a nose dive; bone and tooth displacement and mineral
accretion in an ichthyosaur skull 3 107-116 Wahl, William R., Judy A. Massare, and Mike Ross New material from the type specimens of Megalneusaurus rex (Reptilia:
Sauropterygia) from the Jurassic Sundance Formation, Wyoming. 4 170-180

ROCHESTER INSTITUTE OF VERTEBRATE PALEONTOLOGY The Rochester Institute of Vertebrate Paleontology (RIVP) was founded in December of 1992 as a nonprofit corporation for the purpose of furthering scientific research in Vertebrate Paleontology. Officers President: William W. Korth Provost: Judy A. Massare Research Associates Jon A. Baskin Department of Biological and Health Sciences, Texas A&M University-Kingsville. Kingsville, TX 78363 Jeffrey Chiarenzelli Department of Geology St. Lawrence University 23 Romoda Drive Canton, NY 13617 Robert L. Evander Department of Vertebrate Paleontology American Museum of Natural History Central Park West at 79th St. New York, NY 10024 Margaret Diamond 5215 Ledge Lane Williamsville, NY 14221
Board of Directors William W. Korth, Rochester, NY Judy A. Massare, Rochester, NY George McIntosh, Rochester, NY Mark C. Lawler 33 Wigwam Road Pennellville, NY 13132 Robert A. Martin Department of Biological Sciences Murray State University Murray, KY 42071 Allen R. Tedrow Idaho Museum of Natural History Idaho State University Pocatello, ID 83209 William R. Wahl Wyoming Dinosaur Center 110 Carter Ranch Rd. Thermopolis, WY 82443

CONTENTS Mammals from the Blue Ash local fauna (late Oligocene), South Dakota. Rodentia, Part 4: Family Cricetidae WILLIAM W. KORTH 117-136 A new chub (Actinopterygii, Cypriniformes, Cyprinidae) from the
middle Miocene (early Clarendonian) Aldrich Station Formation, Lyon County, Nevada THOMAS S. KELLY 137-157
New records of Marsupialia, Lipotyphla, and Primates from the
Duchesnean (middle Eocene) Simi Valley Landfill Local Fauna, Sespe Formation, California THOMAS S. KELLY 158-169
New material from the type specimen of Megalneusaurus rex (Reptilia: Sauropterygia)
from the Jurassic Sundance Formation, Wyoming WILLIAM R. WAHL, JUDY A. MASSARE, AND MIKE ROSS 170-180
Author index to Volume 7 181-182




















