
Palaeoecological and biostratigraphical implications of the microvertebrates of Qesem Cave in Israel
14
Palaeoecological and biostratigraphical implications of the microvertebrates of Qesem Cave in Israel Lutz Christian Maul a, * , Angela A. Bruch b , Krister T. Smith c , Georgy Shenbrot d , Ran Barkai e , Avi Gopher e a Senckenberg Research Station of Quaternary Palaeontology, Weimar, Germany b ROCEEH, Senckenberg Research Institute and Natural History Museum, Frankfurt/Main, Germany c Senckenberg Research Institute and Natural History Museum, Frankfurt/Main, Germany d Ben-Gurion University of the Negev, Midreshet Ben-Gurion, Israel e Institute of Archaeology, Tel Aviv University, Israel article info Article history: Available online xxx Keywords: Qesem Cave Israel Small mammals Reptiles Biostratigraphy Palaeoecology abstract Microvertebrates are generally well suited for drawing inferences on past environmental conditions because they are closely bound to the areas in which they lived. In this paper, we discuss palaeoecological implications of two microvertebrate concentrations in the Middle Pleistocene site Qesem Cave in Israel. The ecological preferences of the nearest living relatives of the microvertebrate taxa recorded in Con- centration 1 (squares LeN/13e15) and 2 (squares G-H/16e17) at Qesem Cave allow us to infer a mosaic of open palaeoenvironment with sparse vegetation, shrubland, Mediterranean Forest, rocky areas and riverbanks. Additionally, we infer palaeoclimate from the bioclimatic distribution of extant species using the Coexistence Approach. To our knowledge, this is the first time the approach has been applied to microfaunal assemblages. These data suggest cooler and slightly drier winters and somewhat lower seasonality than at present around Qesem Cave; the differences are more pronounced for the time covered by Concentration 2 than Concentration 1. Among microvertebrates, micromammals are most suitable for biostratigraphic purposes. With the exception of Rattus cf. haasi and the Myomimus judaicus/setzeri group, the small mammal fauna of Qesem Cave comprises only taxa that live today in the Levant. R. haasi and M. judaicus have been found in Israel in the Early and Middle Pleistocene, and their latest known record was from the Acheulian site of Oumm Qatafa. Their presence in Qesem Cave extends the previously known stratigraphic range of these species. The absence of R. haasi in Mousterian sites is thus consistent with the pre-Mousterian age of Qesem Cave. Morphometric data on molars of Guenther's Vole show that the lineage segment preserved in Con- centration 2 is more primitive than in Concentration 1, indicating that the former concentration is older. The evolutionary level of the sample from Qesem Concentration 1 is similar to Tabun D (boundary of Acheulo-Yabrudian Cultural Complex and Mousterian), and that from Qesem Concentration 2 similar to Tabun Eb (Acheulo-Yabrudian). © 2015 Elsevier Ltd and INQUA. All rights reserved. 1. Introduction Qesem Cave is a very productive Middle Pleistocene archaeo- logical site near Tel Aviv in Israel dated by various methods to 420e200 ka (Barkai et al., 2003, 2010; Gopher et al., 2010; Stiner et al., 2011; Blasco et al., 2013; Mercier et al., 2013; Rosell et al., 2015; Falgu eres et al., this volume). The site is a sediment-filled karst chamber cave some 20 15 m in size, which is located ~12 km east of the present day Mediterranean coast, at an altitude of 90 m on the moderate western slopes of the Samaria hills. The cave is part of a larger karstic system within the limestone of the B'ina Formation of Turonian (Late Cretaceous) age. All layers of the more than 10 m stratigraphic sequence of Qesem Cave have been securely assigned to the Acheulo-Yabrudian Cultural Complex (AYCC) of the late Lower Palaeolithic (Barkai et al., 2003; Gopher et al., 2005; Barkai et al., 2009; Barkai and Gopher, 2011; Shimelmitz et al., 2011; Barkai et al., 2013). The huge number of artifacts, large mammal bones (Stiner et al., 2009, 2011; Blasco et al., 2013, 2014), and human teeth (Hershkovitz et al., 2011) attest to the habitual presence of humans in the cave. * Corresponding author. E-mail address: [email protected] (L.C. Maul). Contents lists available at ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint http://dx.doi.org/10.1016/j.quaint.2015.04.032 1040-6182/© 2015 Elsevier Ltd and INQUA. All rights reserved. Quaternary International xxx (2015) 1e14 Please cite this article in press as: Maul, L.C., et al., Palaeoecological and biostratigraphical implications of the microvertebrates of Qesem Cave in Israel, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint.2015.04.032
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Palaeoecological and biostratigraphical implications of the microvertebrates of Qesem Cave in Israel

lable at ScienceDirect
Quaternary International xxx (2015) 1e14
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
Palaeoecological and biostratigraphical implications of themicrovertebrates of Qesem Cave in Israel
Lutz Christian Maul a, *, Angela A. Bruch b, Krister T. Smith c, Georgy Shenbrot d,Ran Barkai e, Avi Gopher e
a Senckenberg Research Station of Quaternary Palaeontology, Weimar, Germanyb ROCEEH, Senckenberg Research Institute and Natural History Museum, Frankfurt/Main, Germanyc Senckenberg Research Institute and Natural History Museum, Frankfurt/Main, Germanyd Ben-Gurion University of the Negev, Midreshet Ben-Gurion, Israele Institute of Archaeology, Tel Aviv University, Israel
a r t i c l e i n f o
Article history:Available online xxx
Keywords:Qesem CaveIsraelSmall mammalsReptilesBiostratigraphyPalaeoecology
* Corresponding author.E-mail address: [email protected] (L.C. Maul)
http://dx.doi.org/10.1016/j.quaint.2015.04.0321040-6182/© 2015 Elsevier Ltd and INQUA. All rights
Please cite this article in press as: Maul, L.C., eIsrael, Quaternary International (2015), http
a b s t r a c t
Microvertebrates are generally well suited for drawing inferences on past environmental conditionsbecause they are closely bound to the areas in which they lived. In this paper, we discuss palaeoecologicalimplications of two microvertebrate concentrations in the Middle Pleistocene site Qesem Cave in Israel.The ecological preferences of the nearest living relatives of the microvertebrate taxa recorded in Con-centration 1 (squares LeN/13e15) and 2 (squares G-H/16e17) at Qesem Cave allow us to infer a mosaic ofopen palaeoenvironment with sparse vegetation, shrubland, Mediterranean Forest, rocky areas andriverbanks. Additionally, we infer palaeoclimate from the bioclimatic distribution of extant species usingthe Coexistence Approach. To our knowledge, this is the first time the approach has been applied tomicrofaunal assemblages. These data suggest cooler and slightly drier winters and somewhat lowerseasonality than at present around Qesem Cave; the differences are more pronounced for the timecovered by Concentration 2 than Concentration 1.
Among microvertebrates, micromammals are most suitable for biostratigraphic purposes. With theexception of Rattus cf. haasi and the Myomimus judaicus/setzeri group, the small mammal fauna of QesemCave comprises only taxa that live today in the Levant. R. haasi and M. judaicus have been found in Israelin the Early and Middle Pleistocene, and their latest known record was from the Acheulian site of OummQatafa. Their presence in Qesem Cave extends the previously known stratigraphic range of these species.The absence of R. haasi in Mousterian sites is thus consistent with the pre-Mousterian age of Qesem Cave.Morphometric data on molars of Guenther's Vole show that the lineage segment preserved in Con-centration 2 is more primitive than in Concentration 1, indicating that the former concentration is older.The evolutionary level of the sample from Qesem Concentration 1 is similar to Tabun D (boundary ofAcheulo-Yabrudian Cultural Complex and Mousterian), and that from Qesem Concentration 2 similar toTabun Eb (Acheulo-Yabrudian).
© 2015 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
Qesem Cave is a very productive Middle Pleistocene archaeo-logical site near Tel Aviv in Israel dated by various methods to420e200 ka (Barkai et al., 2003, 2010; Gopher et al., 2010; Stineret al., 2011; Blasco et al., 2013; Mercier et al., 2013; Rosell et al.,2015; Falgu�eres et al., this volume). The site is a sediment-filledkarst chamber cave some 20 � 15 m in size, which is located~12 km east of the present day Mediterranean coast, at an altitude
.
reserved.
t al., Palaeoecological and bio://dx.doi.org/10.1016/j.quaint
of 90 m on the moderate western slopes of the Samaria hills. Thecave is part of a larger karstic system within the limestone of theB'ina Formation of Turonian (Late Cretaceous) age. All layers of themore than 10 m stratigraphic sequence of Qesem Cave have beensecurely assigned to the Acheulo-Yabrudian Cultural Complex(AYCC) of the late Lower Palaeolithic (Barkai et al., 2003; Gopheret al., 2005; Barkai et al., 2009; Barkai and Gopher, 2011;Shimelmitz et al., 2011; Barkai et al., 2013).
The huge number of artifacts, large mammal bones (Stiner et al.,2009, 2011; Blasco et al., 2013, 2014), and human teeth (Hershkovitzet al., 2011) attest to the habitual presence of humans in the cave.
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

Fig. 1. Qesem Cave map of excavation squares with indication of Concentrations 1 and 2.
Table 1Microvertebrate taxa recorded from the Concentration 1 and 2 (see Fig. 1) ofQesem Cave.
Concentration 1 Concentration 2
Vertebrata indet. 72,139 153,813ActinopterygiiActinopterygii indet. 1
AmphibiaRanidae indet. 24 5
ReptiliaTestudinesTestudo (graeca) 340 252
SquamataSquamata indet. 989 546Stellagama stellio 543 883Chamaeleo chamaeleon 2,695 4,806Gekkonidae indet. 6 7Lacertidae indet. (2 species) 20 13Scincidae indet. 2 7Pseudopus sp. 432 49cf. Eryx sp. 1 1Colubroidea indet. (3þ species) 35 50
L.C. Maul et al. / Quaternary International xxx (2015) 1e142
However, the environmental conditions under which the ancienthumans lived remainpoorly known. In order to advance knowledgeofthese conditions, we focus here on small terrestrial vertebrates: smallmammals, amphibians and reptiles. These animals are generally wellsuited for drawing inferences on past environmental conditions,because theyare closely bound to the areas inwhich they lived.QesemCave is very rich in microvertebrate remains (Maul et al., 2011;Hor�a�cek et al., 2013; Smith et al., 2013; Smith et al., this volume),which were retrieved from two distinct local concentrations: con-centration area 1 in squares LeN/13e15 and concentration area 2 inGeH/16e17 (Fig. 1, for details see Smith et al., this volume).
The ecological implications of small terrestrial vertebrates are onlymeaningful insofar as they are securely dated. In addition to absolutedating methods, small mammals also provide useful information onthe biostratigraphy of the locality under consideration. Although in-dependent physical dating of the excavation areas is already available(Gopher et al., 2010; Mercier et al., 2013), biostratigraphic data frommicromammals are important since they provide a basis for corre-lation with microvertebrate samples from other localities in whichabsolute dating is lacking.Moreover, the application of various datingmethods offers the possibility to cross-check the results.
In this paper, we analyse the microvertebrate material fromQesem Cave in order to infer its age and the ecological conditionsunder which it was deposited. Initial implications on palaeoecologyand biostratigraphy of concentration area 1 were discussed in pre-vious papers (Maul et al., 2011; Hor�a�cek et al., 2013). However, thereis now much more material available e particularly from Concen-tration 2 e and we apply more refined methods here. Finally, givendiffering palaeoecological conditions and stratigraphical age of thetwo concentrations, they were not accumulated at the same timebut one after the other, resultswhichwould contribute new insightsinto intra-site processes and the history of deposition.
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
2. Material and method
2.1. Material
The microfaunal remains were collected mostly by screen-washing (mesh size 0.8 mm) and pre-sorting in the field. Somespecimens were picked directly from the fossiliferous layer duringexcavation. Some 250,000 microvertebrate skeletal and dental re-mains have been collected so far from the two concentration areas.Some 16,000 remains have been identified to genus or species level(Table 1, for details see Smith et al., this volume).
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032
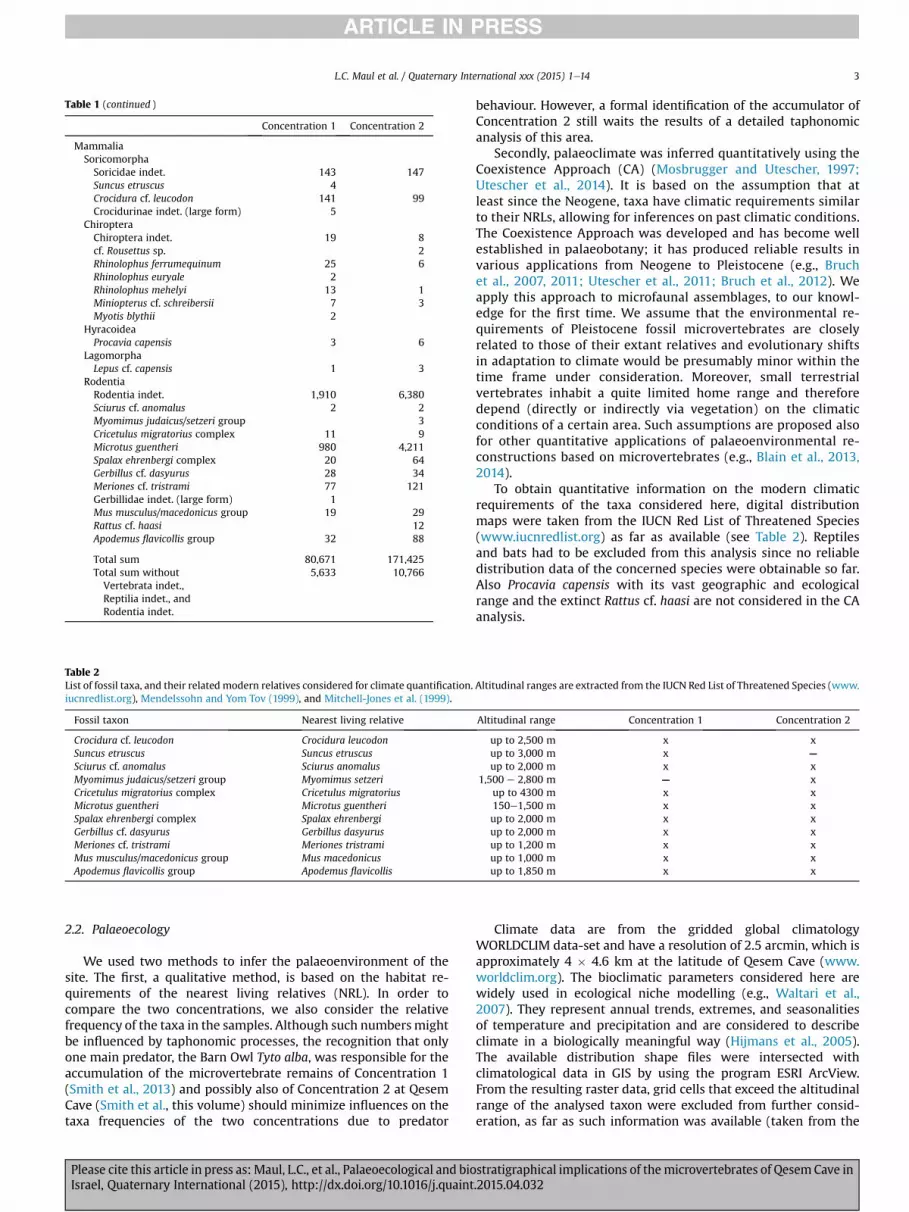
Table 1 (continued )
Concentration 1 Concentration 2
MammaliaSoricomorphaSoricidae indet. 143 147Suncus etruscus 4Crocidura cf. leucodon 141 99Crocidurinae indet. (large form) 5
ChiropteraChiroptera indet. 19 8cf. Rousettus sp. 2Rhinolophus ferrumequinum 25 6Rhinolophus euryale 2Rhinolophus mehelyi 13 1Miniopterus cf. schreibersii 7 3Myotis blythii 2
HyracoideaProcavia capensis 3 6
LagomorphaLepus cf. capensis 1 3
RodentiaRodentia indet. 1,910 6,380Sciurus cf. anomalus 2 2Myomimus judaicus/setzeri group 3Cricetulus migratorius complex 11 9Microtus guentheri 980 4,211Spalax ehrenbergi complex 20 64Gerbillus cf. dasyurus 28 34Meriones cf. tristrami 77 121Gerbillidae indet. (large form) 1Mus musculus/macedonicus group 19 29Rattus cf. haasi 12Apodemus flavicollis group 32 88
Total sum 80,671 171,425Total sum without 5,633 10,766Vertebrata indet.,Reptilia indet., andRodentia indet.
Table 2List of fossil taxa, and their related modern relatives considered for climate quantification. Altitudinal ranges are extracted from the IUCN Red List of Threatened Species (www.iucnredlist.org), Mendelssohn and Yom Tov (1999), and Mitchell-Jones et al. (1999).
Fossil taxon Nearest living relative Altitudinal range Concentration 1 Concentration 2
Crocidura cf. leucodon Crocidura leucodon up to 2,500 m x xSuncus etruscus Suncus etruscus up to 3,000 m x —
Sciurus cf. anomalus Sciurus anomalus up to 2,000 m x xMyomimus judaicus/setzeri group Myomimus setzeri 1,500 e 2,800 m — xCricetulus migratorius complex Cricetulus migratorius up to 4300 m x xMicrotus guentheri Microtus guentheri 150e1,500 m x xSpalax ehrenbergi complex Spalax ehrenbergi up to 2,000 m x xGerbillus cf. dasyurus Gerbillus dasyurus up to 2,000 m x xMeriones cf. tristrami Meriones tristrami up to 1,200 m x xMus musculus/macedonicus group Mus macedonicus up to 1,000 m x xApodemus flavicollis group Apodemus flavicollis up to 1,850 m x x
L.C. Maul et al. / Quaternary International xxx (2015) 1e14 3
2.2. Palaeoecology
We used two methods to infer the palaeoenvironment of thesite. The first, a qualitative method, is based on the habitat re-quirements of the nearest living relatives (NRL). In order tocompare the two concentrations, we also consider the relativefrequency of the taxa in the samples. Although such numbersmightbe influenced by taphonomic processes, the recognition that onlyone main predator, the Barn Owl Tyto alba, was responsible for theaccumulation of the microvertebrate remains of Concentration 1(Smith et al., 2013) and possibly also of Concentration 2 at QesemCave (Smith et al., this volume) should minimize influences on thetaxa frequencies of the two concentrations due to predator
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
behaviour. However, a formal identification of the accumulator ofConcentration 2 still waits the results of a detailed taphonomicanalysis of this area.
Secondly, palaeoclimate was inferred quantitatively using theCoexistence Approach (CA) (Mosbrugger and Utescher, 1997;Utescher et al., 2014). It is based on the assumption that atleast since the Neogene, taxa have climatic requirements similarto their NRLs, allowing for inferences on past climatic conditions.The Coexistence Approach was developed and has become wellestablished in palaeobotany; it has produced reliable results invarious applications from Neogene to Pleistocene (e.g., Bruchet al., 2007, 2011; Utescher et al., 2011; Bruch et al., 2012). Weapply this approach to microfaunal assemblages, to our knowl-edge for the first time. We assume that the environmental re-quirements of Pleistocene fossil microvertebrates are closelyrelated to those of their extant relatives and evolutionary shiftsin adaptation to climate would be presumably minor within thetime frame under consideration. Moreover, small terrestrialvertebrates inhabit a quite limited home range and thereforedepend (directly or indirectly via vegetation) on the climaticconditions of a certain area. Such assumptions are proposed alsofor other quantitative applications of palaeoenvironmental re-constructions based on microvertebrates (e.g., Blain et al., 2013,2014).
To obtain quantitative information on the modern climaticrequirements of the taxa considered here, digital distributionmaps were taken from the IUCN Red List of Threatened Species(www.iucnredlist.org) as far as available (see Table 2). Reptilesand bats had to be excluded from this analysis since no reliabledistribution data of the concerned species were obtainable so far.Also Procavia capensis with its vast geographic and ecologicalrange and the extinct Rattus cf. haasi are not considered in the CAanalysis.
Climate data are from the gridded global climatologyWORLDCLIM data-set and have a resolution of 2.5 arcmin, which isapproximately 4 � 4.6 km at the latitude of Qesem Cave (www.worldclim.org). The bioclimatic parameters considered here arewidely used in ecological niche modelling (e.g., Waltari et al.,2007). They represent annual trends, extremes, and seasonalitiesof temperature and precipitation and are considered to describeclimate in a biologically meaningful way (Hijmans et al., 2005).The available distribution shape files were intersected withclimatological data in GIS by using the program ESRI ArcView.From the resulting raster data, grid cells that exceed the altitudinalrange of the analysed taxon were excluded from further consid-eration, as far as such information was available (taken from the
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

L.C. Maul et al. / Quaternary International xxx (2015) 1e144
IUCN Red List, see Table 2). The maximum and minimum values ofthe extracted climate data were calculated for each climateparameter and taken as the climatic boundaries of the taxon. Theclimatic parameters considered here and their abbreviations aregiven in Table 3.
Table 3List of abbreviations and units of the climate parameters discussed here, and present day values for Qesem Cave area extracted from 2.5 arcminWORLDCLIM data set (values represent 30-years-means interpolated from weather station data).
Climate parameter Present day climate at Qesem
MAT Mean annual temperature [�C] 20.1Tmax Maximum temperature of warmest month [�C] 32.1Tmin Minimum temperature of coldest month [�C] 8.3TQwet Temperature of the wettest quarter [�C] 13.9TQdry Temperature of the driest quarter [�C] 25.5TQwarm Temperature of the warmest quarter [�C] 26.0TQcold Temperature of the coldest quarter [�C] 13.9MAP Mean annual precipitation [mm] 547PMwet Precipitation of the wettest month [mm] 140PMdry Precipitation of the driest month [mm] 0PQwet Precipitation of the wettest quarter [mm] 367PQdry Precipitation of the driest quarter [mm] 0PQwarm Precipitation of the warmest quarter [mm] 1PQcold Precipitation of the coldest quarter [mm] 367
With the Coexistence Approach, the climatic ranges in whichall NLRs of a given fossil assemblage can coexist (or a maximumnumber of them, in case they cannot all coexist) is determinedindependently for each climatic parameter. The resulting ‘coex-istence interval’, one for each parameter (see an examplein Fig. 2), is considered the best description of thepalaeoclimatic situation under which the given faunal assem-blage existed.
2.3. Biostratigraphy
Biostratigraphic dating uses the regional occurrence of the taxaunder consideration. In order to compare the Qesem micro-vertebrate concentrations one to the other and with those of othersites of the Levant, we consider only records of small mammals. The
Fig. 2. Example of CA analysis for one assemblage (Concentration 2) and oneparameter (Minimum temperature of the coldest month). The borders of the resultingCoexistence Interval are set by Gerbillus dasyurus and Myomimus setzeri.
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
stratigraphic range of the lower vertebrate taxa recorded in QesemCave is poorly known, so even if they are identified very precisely,present knowledge does not permit statements about the age of theassemblage.
It is possible to extract further biostratigraphic information from
the evolutionary levels of fast-evolving taxa. In Pleistocene sedi-ments, especially voles (subfamily Arvicolinae) display striking andcontinuous evolutionary changes in their dental morphology(occlusal surface pattern, enamel thickness, crown height), whichcan be expressed by morphometric ratios (van der Meulen, 1973;Heinrich, 1978; Rabeder, 1981). This, together with the fact thatthey occur in most terrestrial sites in large numbers, qualifies themas one the most frequently used biostratigraphic indicators in theQuaternary.
The occlusal surface in the first lower molar (m1) of allMicrotuslineages is characterised by continuous evolutionary change. Theanterior part of this tooth (anteroconid complex) shows anevolutionary increase in relative length, expressed by the lengthratio of the anteroconid complex and the occlusal surface of thewhole tooth, the so-called A/L Index (¼ anteroconid length � 100/tooth length; according to van der Meulen, 1973) (Fig. 3). This ratiocan be used for evolutionary and biostratigraphic assessments,however, mainly within the same lineage, and in the samegeographic region. The only arvicoline species at Qesem Cave isMicrotus guentheri.
Fig. 3. First lower molar of an arvicoline rodent with indication of the length of theocclusal surface (L) and of the anteroconid complex (A) of this tooth.
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

L.C. Maul et al. / Quaternary International xxx (2015) 1e14 5
In this paper, we use the following abbreviations: A e length ofanteroconid complex of m1, CA eCoexistence Approach, L e
occlusal length of m1, m1 e first lower molar, NLR e nearest livingrelative. For abbreviations and units of the climate parametersdiscussed, see Table 3.
3. Results and discussion
3.1. Palaeoecology
3.1.1. Habitat requirements of NLRsInformation on habitat requirements of the closest living rela-
tives of the taxa recorded from Qesem Cave are from various au-thors, including assessors of the IUCN Red List (http://www.iucnredlist.org/about/introduction).
3.1.1.1. Fish. The presence of a large actinopterygian fish remainsprobably indicates the presence of a water body in the vicinity ofthe site, more specifically in the hunting area of the owl responsiblefor the microvertebrate accumulation.
3.1.1.2. Reptiles. The Spur-Thighed Tortoise, Testudo graeca, inhabitshumid and mesic steppe, semi-steppe and forest habitats (Disiet al., 2001).
The Hardun or Starred Agama, Stellagama stellio, is a diurnallizard, nowadays found in a variety of Mediterranean arid andsemi-arid habitats, and prefers rocky habitats in mountainous andhilly areas usually with good vegetation, mostly on vertical rockyoutcrops, cliffs, rocky hills and banks of wadis, but it occurs also intrue rocky desert and oak forests, where it climbs on trees (Disiet al., 2001; Baha El Din, 2006; Maul et al., 2011 and citationstherein).
The Common Chameleon, Chamaeleo chamaeleon, is a diurnal,arboreal animal, which lives in dry to humid habitats requiringmore or less a dense vegetation (Mediterranean forests, openpine woodland, shrubland, wadis) (Disi et al., 2001), but occursalso in semi-desert with good vegetation and some largerbushes, where it moves usually in low vegetation, but also onthe ground (Baha El Din, 2006). However, in Israel the speciesoccurs also in the true desert, in sand dunes with relativelydense shrub vegetation, e.g., in the Agur sands west of Beer-Sheva.
The Sheltopusik (European Glass Lizard), Pseudopus apodus,lives today in open habitats, such as shrubland with dense,moderately tall to tall vegetation, avoiding desert and forest (Disiet al., 2001), and prefers light woodlands with dry and warm hill-sides, often near stream banks (Disi et al., 2001).
Members of the genus Eryx, Old World sand boas, live in the SEMediterranean in arid and semi-arid habitats of sand and alluvialsoils (Disi et al., 2001; Baha El Din, 2006).
3.1.1.3. Mammals. The recent Rock hyrax Procavia capensis isspread over a huge geographic area from the Levant to South Africa.Its primary habitat requirement is rocky terrain, so it is found in awide range of habitats, extreme desert to Mediterranean forest(Mendelssohn and Yom-Tov, 1999).
The living Bicoloured White-toothed shrew Crocidura leuco-don occurs mainly in open rocky habitats, and although it is oftenfound in moist areas with thick cover, it is more tolerant of openformations (steppe and semi-desert areas) than other species ofCrocidura. This species is frequent in screes, dense tall grass, andbanks of rivers and brooks, and in central Anatolia also
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
hardwood forest (Mitchell-Jones et al., 1999; Kry�stufek andVohralík, 2001).
The recent Suncus etruscus (Etruscan shrew) is confined toareas with mean July temperature of at least 20 �C, and occurs inshrub, low maquis, and open Mediterranean oak and pine for-ests, provided that rocks are available as shelters, but it avoidsdense forests (Mitchell-Jones et al., 1999). In Lebanon it lives insemi-arid and moist habitats on the edge of pine woods(Harrison and Bates, 1991). In Israel it is found also in sanddunes, and in desert areas it is quite common in loess (forexample, around Mitzpe Ramon and Sede Boqer, see Shenbrotand Krasnov, 1997).
The living relatives of all the Qesem bat taxa (Rousettus e
Egyptian fruit bat, Rhinolophus ferrumequinum, R. euryale, R.mehelyie three Horse shoe bats, Miniopterus schreibersii e Common bent-wing bat, and Myotis blythii e Lesser mouse-eared bat) belong totypical cave-dwelling species of Eastern Mediterranean, roosting inmoist natural caverns (Hor�a�cek et al., 2013). Rousettus has a broadhabitat tolerance (if there is abundant food and an appropriateroosting site) and occurs in arid to moist tropical and subtropicalbiomes. Rhinolophus ferrumequinum, R. mehelyi, and R. euryale livein Mediterranean and sub-Mediterranean shrubland and wood-land; R. mehelyi also occurs in dry steppes. Miniopterus schreibersiioccurs in a variety of open and semi-open habitats, and Myotisblythii in scrub and grassland habitats (Mendelssohn and Yom-Tov,1999).
The recent Cape Hare, Lepus capensis, occurs in a wide range ofhabitats. In Africa it is commonly associated with open habitatsincluding desert, steppe and grassland, but on the Arabian Penin-sula it prefers shrubs, rather than grasses, to shelter under insummer (Mitchell-Jones et al., 1999).
The Persian Squirrel, Sciurus anomalus is a tree species and itsrange depends on the presence of woodland. However, it is possiblyless dependent on a close canopy forest than S. vulgaris (Kry�stufekand Vohralík, 2005), and it lives mainly in mixed deciduous forest,although it also occurs in coniferous forests (e.g., on the southerncoast of Turkey) as well as rocky outcrops (Mitchell-Jones et al.,1999).
The living species of Myomimus (Mouse-tailed dormouse) areknown from a variety of habitats with sparse trees or bushland. M.roachi occurs in dry woodland and scrub, and also at river banks(Mitchell-Jones et al., 1999), however in particular areas (e.g.,Turkish Thrace) never in forests (Kry�stufek and Vohralík, 2005). M.personatus inhabits rocky desert and occurs among shrubs, salt-warts, and almond trees.M. setzeri is an extremely rare speciese itsoriginal description is based on just three specimens ewhich werefound in a sparse forest of old pines loosely covered with grass.Pistacia savannah is considered to be a typical habitat (Kry�stufekand Vohralík, 2005).
What is named Cricetulus migratorius (Grey dwarf hamster)probably represents a complex of several cryptic species or semi-species (Lebedev, 2012), which may partly explain the broadhabitat tolerance described for the species in the literature. Itprefers arid areas, steppes and stony places covered with sparsevegetation. It avoids forests and damp habitats, but it inhabits alsodeserts and semi-deserts (Mitchell-Jones et al., 1999; Kry�stufek andVohralík, 2009). The occurrence in deserts and semi-deserts istypical for its eastern forms (in Kazakhstan, Mongolia and China). Inthe Near East it is really typical of dry mountain steppe and to alesser extent of semi-desert (e.g., in the inland of Anatolia andArmenia).
Guenther's vole, Microtus guentheri, inhabits dry grasslands andexposed hill slopes with sparse vegetation on well-drained soil,
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

L.C. Maul et al. / Quaternary International xxx (2015) 1e146
often with rocks and stones, and also along river banks; it can evenbe abundant in swamps (Mitchell-Jones et al., 1999; Kry�stufek andVohralík, 2005).
Wagner's Gerbil, Gerbillus dasyurus, lives on rocky slopes anddigs burrows under rocks. In the Mediterranean zone of Israel itlives in areas with sparse vegetation. In the Negev, the main factoraffecting the distribution is soil structure. It avoids sandy soils andextremely dense vegetation but prefers moderately coveredpatches and occurs mainly on loess hills with rocks (Shenbrot et al.,1997; Mendelssohn and Yom-Tov, 1999).
Tristram's Jird, Meriones tristrami, is found in a wide range ofhabitats, but deserts, semi-desert and dry steppes with >100 mmannual precipitation is preferred. It needs well-drained soil, andaccording to some authors (Mitchell-Jones et al., 1999; Kry�stufekand Vohralík, 2009) in some areas it avoids rocky areas and for-ests, although it was trapped by one of us (GS) in rocky habitat atforest edge.
Both Mus musculus and M. macedonicus (House mouse andMacedonian mouse, respectively) are found in a very wide range ofhabitats including dry shrubby areas, bushy banks, and wadis anddoes not strictly avoid forests and deserts, whereasM.macedonicusis limited to areas with Mediterranean shrubs (Mitchell-Jones et al.,1999).
The fossil rat, Rattus cf. haasi from Qesem is related to R. rattusand R. norvegicus (Tchernov, 1968). Non-commensal R. rattusoccur in a variety of disturbed habitats with dense woodyvegetation; humid and even damp places are preferred withinthe relatively dry Mediterranean climate. R. norvegicus, on theother hand, is more adapted to mesic habitats than R. rattus andlives along river banks and lake shores (Kry�stufek and Vohralík,2009).
Recent Apodemus flavicollis (Yellow-necked mouse) inhabits avariety of woodland habitats, primarily deciduous woodland,although it is known from other habitats including coniferous for-est and bushland. It is the dominant small mammal in all habitatsexcept open fields, which provide too little cover (Kry�stufek andVohralík, 2009). However, the morphologically similar and closelyrelated species A. witherbyi (senior synonym for A. tauricus and A.hermonensis) is more restricted to mountain oak forest and shrubs(Filippucci et al., 1989).
The Mole Rat Spalax ehrenbergi, also called ‘Spalax ehrenbergicomplex’ since it obviously represents a complex of cryptic species(Nevo et al., 1979), is strictly fossorial. It occurs in open formationsof the Mediterranean and steppic region and in semi-deserts, but italso penetrates into openings in maquis and woodland andmarginally into sandy and loess desert. Since it is limited by the100-mm isohyet, strict desert is generally avoided (Kry�stufek andVohralík, 2009) although it might occur marginally in deserthabitats.
3.1.1.4. Palaeoenvironment. Summarising the arguments above, wecan make the following qualitative assessment of the palae-oenvironment of the surrounding of Qesem Cave.
Humidity: there are species that live exclusively (Pseudopusapodus, Cricetulus migratorius, Microtus guentheri, Gerbillus dasyu-rus, Meriones tristrami) or partly (Stellagama stellio) in semi-aridhabitats and those which are less dependent on a particularminimum humidity (Chamaeleo, Procavia, cf. Rousettus). Whereas
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
Cricetulus migratorius avoids damp places, Crocidura leucodonprefers generally humid habitats, but as mentioned above, thelatter is tolerant of even steppe and semi-desert areas. This wouldimply the greatest overlap in habitat tolerances is in semi-aridareas.
Vegetation cover: Some of the species are adapted to openhabitats, such as semi-desert and desert (Gerbillus dasyurus, Crice-tulus migratorius e avoiding forest), or grassland and steppe (Mer-iones tristrami e avoids forest, Nannospalax ehrenbergi, Microtusguentheri, Crocidura leucodon). Other taxa require or tolerateMediterranean woodland (Chamaeleo, Pseudopus, Suncus etruscus,Sciurus anomalus, Apodemus flavicollis) or shrubland (Chamaeleo,Pseudopus, Myomimus setzeri, Apodemus flavicollis). Thus, we caninfer the juxtaposition of open habitats, shrubland and Mediter-ranean woodland.
Surface: Pseudopus, Crocidura leucodon, and Microtus guentheriare reported to inhabit river banks. Rocky habitats, often as shelters,are typical for Stellagama, Procavia, Crocidura leucodon, and Suncusetruscus, and occasionally for Sciurus anomalus, Cricetulus migra-torius and Gerbillus dasyurus (at forest edges even for Merionestristrami). The occurrence of rocky habitats in the surroundingseems to be evident, as well as riverbanks at least according to thepresence of fish.
Temperature: All recent relatives of the taxa recorded inQesem Cave now inhabit a Mediterranean climate, and some ofthem even occur in desert climate. No one species requiresparticularly cold or continental conditions. In sum, from the re-quirements of the recorded microvertebrate taxa we concludethe existence of a mosaic of open area with sparse vegetation butalso shrubland, Mediterranean Forest, rocky areas andriverbanks.
There are only few differences between the two concentra-tions in the presence/absence of the microvertebrate taxa. Cha-maeleo chamaeleon and Microtus guentheri are in bothconcentrations by far the dominant taxa (for details see Smithet al., this volume). Clear differences between the two concen-trations can be observed by higher relative frequency of Ranidaeand Soricidae in Concentration 1 compared to Concentration 2,where the number of Microtus guentheri is more than doubledthan in Concentration 1. However, these differences cannot beunambiguously interpreted as signals for drier conditions inConcentration 2, since all but the finds of Chamaeleo and Microtusoccur in percentages below 10%, mostly even below 1%, andchanges in one of the two extremely abundant species have amajor impact on the numbers of other species (for further detailssee Smith et al., this volume).
3.1.2. Quantitative climatic dataQuantitative climate data obtained from the Coexistence
Approach are given in Tables 4 and 5 and Fig. 4. The results for thetwo assemblages are very similar due to the nearly identical faunalcomposition. The presence of Suncus etruscus in Concentration 1(not present in Concentration 2) does not influence the results dueto the wide climatic tolerance of this species. Only the presence ofMyomimus ex. gr. setzeri/judaica in Concentration 2 leads to nar-rower coexistence intervals for some climatic parameterscompared to Concentration 1.
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032
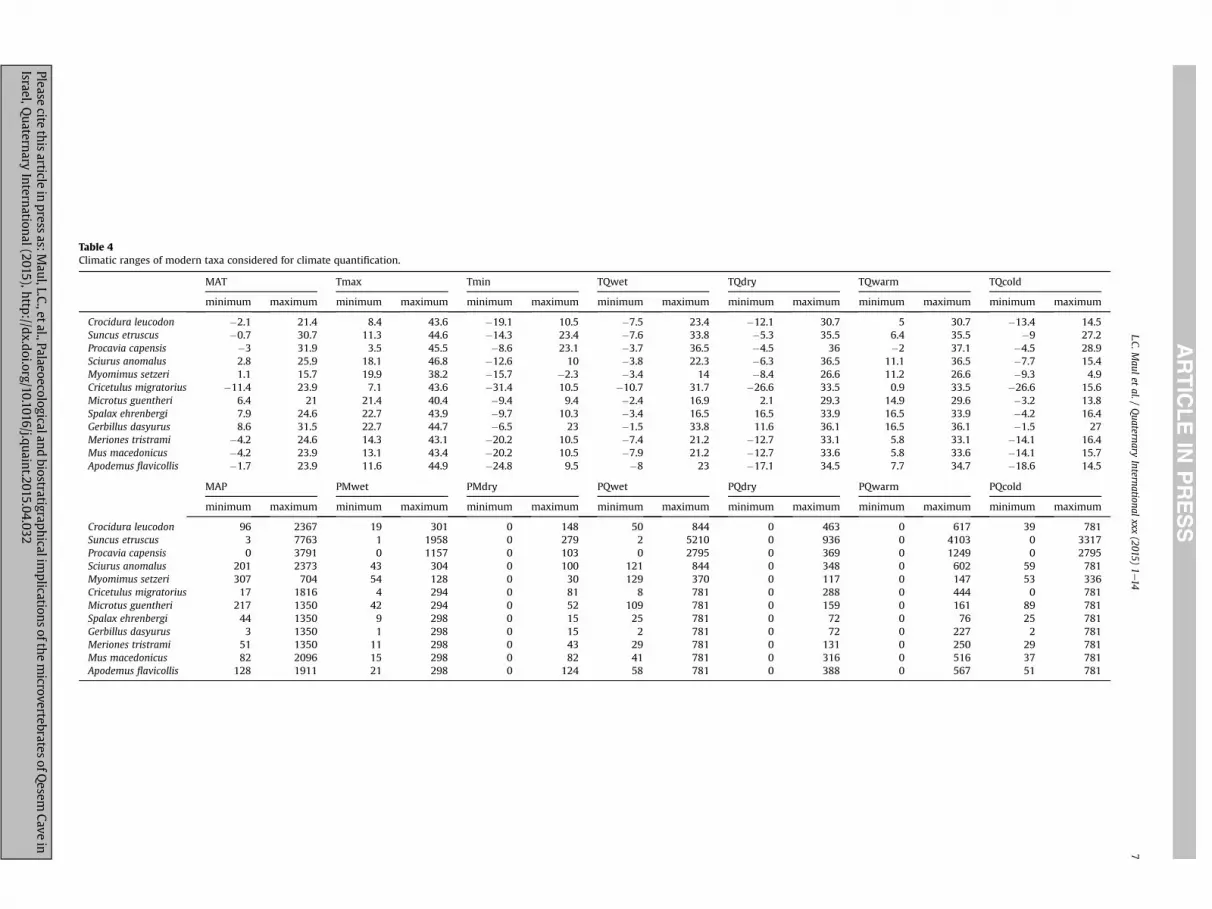
Table 4Climatic ranges of modern taxa considered for climate quantification.
MAT Tmax Tmin TQwet TQdry TQwarm TQcold
minimum maximum minimum maximum minimum maximum minimum maximum minimum maximum minimum maximum minimum maximum
Crocidura leucodon �2.1 21.4 8.4 43.6 �19.1 10.5 �7.5 23.4 �12.1 30.7 5 30.7 �13.4 14.5Suncus etruscus �0.7 30.7 11.3 44.6 �14.3 23.4 �7.6 33.8 �5.3 35.5 6.4 35.5 �9 27.2Procavia capensis �3 31.9 3.5 45.5 �8.6 23.1 �3.7 36.5 �4.5 36 �2 37.1 �4.5 28.9Sciurus anomalus 2.8 25.9 18.1 46.8 �12.6 10 �3.8 22.3 �6.3 36.5 11.1 36.5 �7.7 15.4Myomimus setzeri 1.1 15.7 19.9 38.2 �15.7 �2.3 �3.4 14 �8.4 26.6 11.2 26.6 �9.3 4.9Cricetulus migratorius �11.4 23.9 7.1 43.6 �31.4 10.5 �10.7 31.7 �26.6 33.5 0.9 33.5 �26.6 15.6Microtus guentheri 6.4 21 21.4 40.4 �9.4 9.4 �2.4 16.9 2.1 29.3 14.9 29.6 �3.2 13.8Spalax ehrenbergi 7.9 24.6 22.7 43.9 �9.7 10.3 �3.4 16.5 16.5 33.9 16.5 33.9 �4.2 16.4Gerbillus dasyurus 8.6 31.5 22.7 44.7 �6.5 23 �1.5 33.8 11.6 36.1 16.5 36.1 �1.5 27Meriones tristrami �4.2 24.6 14.3 43.1 �20.2 10.5 �7.4 21.2 �12.7 33.1 5.8 33.1 �14.1 16.4Mus macedonicus �4.2 23.9 13.1 43.4 �20.2 10.5 �7.9 21.2 �12.7 33.6 5.8 33.6 �14.1 15.7Apodemus flavicollis �1.7 23.9 11.6 44.9 �24.8 9.5 �8 23 �17.1 34.5 7.7 34.7 �18.6 14.5
MAP PMwet PMdry PQwet PQdry PQwarm PQcold
minimum maximum minimum maximum minimum maximum minimum maximum minimum maximum minimum maximum minimum maximum
Crocidura leucodon 96 2367 19 301 0 148 50 844 0 463 0 617 39 781Suncus etruscus 3 7763 1 1958 0 279 2 5210 0 936 0 4103 0 3317Procavia capensis 0 3791 0 1157 0 103 0 2795 0 369 0 1249 0 2795Sciurus anomalus 201 2373 43 304 0 100 121 844 0 348 0 602 59 781Myomimus setzeri 307 704 54 128 0 30 129 370 0 117 0 147 53 336Cricetulus migratorius 17 1816 4 294 0 81 8 781 0 288 0 444 0 781Microtus guentheri 217 1350 42 294 0 52 109 781 0 159 0 161 89 781Spalax ehrenbergi 44 1350 9 298 0 15 25 781 0 72 0 76 25 781Gerbillus dasyurus 3 1350 1 298 0 15 2 781 0 72 0 227 2 781Meriones tristrami 51 1350 11 298 0 43 29 781 0 131 0 250 29 781Mus macedonicus 82 2096 15 298 0 82 41 781 0 316 0 516 37 781Apodemus flavicollis 128 1911 21 298 0 124 58 781 0 388 0 567 51 781
L.C.Maul
etal./
Quaternary
Internationalxxx
(2015)1e14
7
Pleasecite
thisarticle
inpress
as:Maul,L.C.,etal.,Palaeoecologicaland
biostratigraphicalimplications
ofthemicrovertebrates
ofQesem
Cavein
Israel,Quaternary
International(2015),http://dx.doi.org/10.1016/j.quaint.2015.04.032
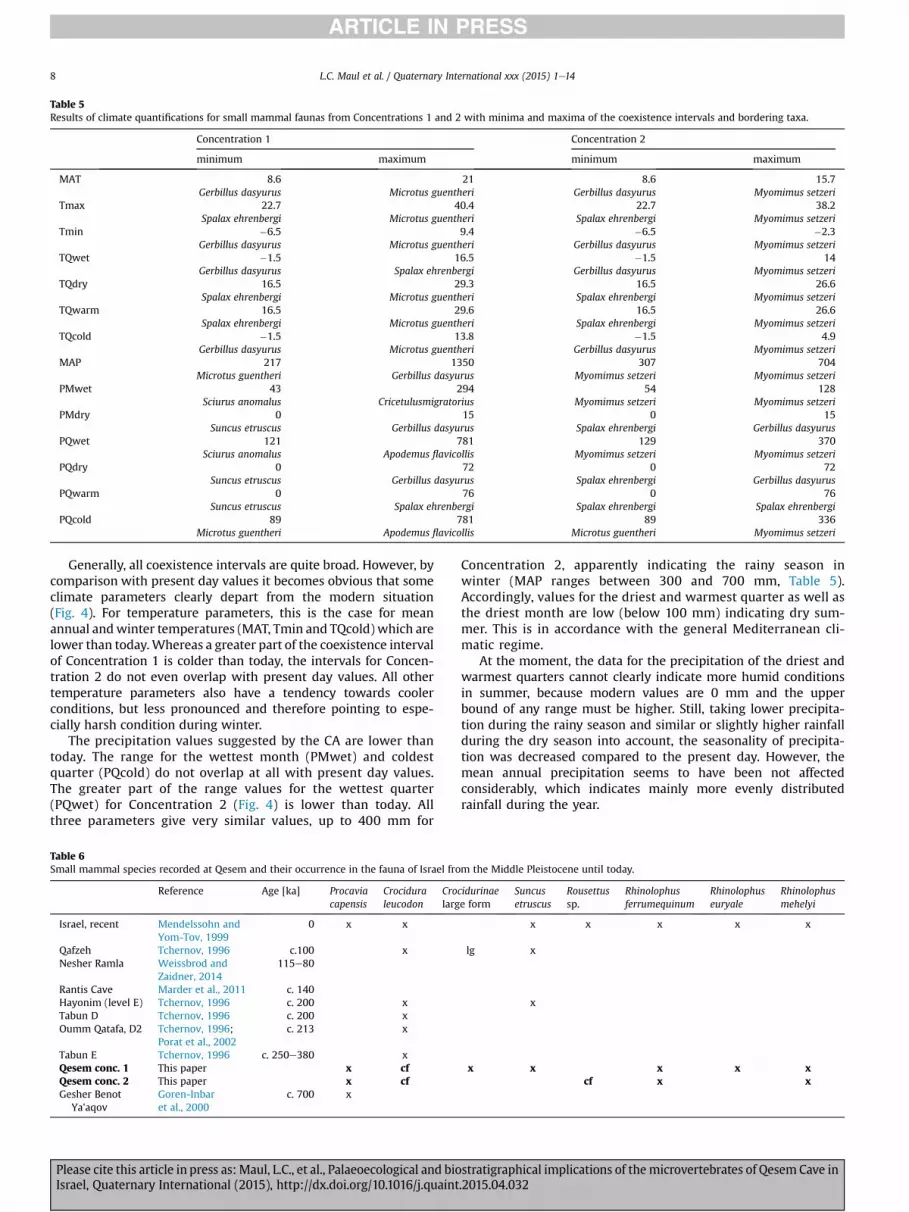
Table 5Results of climate quantifications for small mammal faunas from Concentrations 1 and 2 with minima and maxima of the coexistence intervals and bordering taxa.
Concentration 1 Concentration 2
minimum maximum minimum maximum
MAT 8.6 21 8.6 15.7Gerbillus dasyurus Microtus guentheri Gerbillus dasyurus Myomimus setzeri
Tmax 22.7 40.4 22.7 38.2Spalax ehrenbergi Microtus guentheri Spalax ehrenbergi Myomimus setzeri
Tmin �6.5 9.4 �6.5 �2.3Gerbillus dasyurus Microtus guentheri Gerbillus dasyurus Myomimus setzeri
TQwet �1.5 16.5 �1.5 14Gerbillus dasyurus Spalax ehrenbergi Gerbillus dasyurus Myomimus setzeri
TQdry 16.5 29.3 16.5 26.6Spalax ehrenbergi Microtus guentheri Spalax ehrenbergi Myomimus setzeri
TQwarm 16.5 29.6 16.5 26.6Spalax ehrenbergi Microtus guentheri Spalax ehrenbergi Myomimus setzeri
TQcold �1.5 13.8 �1.5 4.9Gerbillus dasyurus Microtus guentheri Gerbillus dasyurus Myomimus setzeri
MAP 217 1350 307 704Microtus guentheri Gerbillus dasyurus Myomimus setzeri Myomimus setzeri
PMwet 43 294 54 128Sciurus anomalus Cricetulusmigratorius Myomimus setzeri Myomimus setzeri
PMdry 0 15 0 15Suncus etruscus Gerbillus dasyurus Spalax ehrenbergi Gerbillus dasyurus
PQwet 121 781 129 370Sciurus anomalus Apodemus flavicollis Myomimus setzeri Myomimus setzeri
PQdry 0 72 0 72Suncus etruscus Gerbillus dasyurus Spalax ehrenbergi Gerbillus dasyurus
PQwarm 0 76 0 76Suncus etruscus Spalax ehrenbergi Spalax ehrenbergi Spalax ehrenbergi
PQcold 89 781 89 336Microtus guentheri Apodemus flavicollis Microtus guentheri Myomimus setzeri
L.C. Maul et al. / Quaternary International xxx (2015) 1e148
Generally, all coexistence intervals are quite broad. However, bycomparison with present day values it becomes obvious that someclimate parameters clearly depart from the modern situation(Fig. 4). For temperature parameters, this is the case for meanannual andwinter temperatures (MAT, Tmin and TQcold) which arelower than today.Whereas a greater part of the coexistence intervalof Concentration 1 is colder than today, the intervals for Concen-tration 2 do not even overlap with present day values. All othertemperature parameters also have a tendency towards coolerconditions, but less pronounced and therefore pointing to espe-cially harsh condition during winter.
The precipitation values suggested by the CA are lower thantoday. The range for the wettest month (PMwet) and coldestquarter (PQcold) do not overlap at all with present day values.The greater part of the range values for the wettest quarter(PQwet) for Concentration 2 (Fig. 4) is lower than today. Allthree parameters give very similar values, up to 400 mm for
Table 6Small mammal species recorded at Qesem and their occurrence in the fauna of Israel fro
Reference Age [ka] Procaviacapensis
Crociduraleucodon
Crolarg
Israel, recent Mendelssohn andYom-Tov, 1999
0 x x
Qafzeh Tchernov, 1996 c.100 xNesher Ramla Weissbrod and
Zaidner, 2014115e80
Rantis Cave Marder et al., 2011 c. 140Hayonim (level E) Tchernov, 1996 c. 200 xTabun D Tchernov, 1996 c. 200 xOumm Qatafa, D2 Tchernov, 1996;
Porat et al., 2002c. 213 x
Tabun E Tchernov, 1996 c. 250e380 xQesem conc. 1 This paper x cfQesem conc. 2 This paper x cfGesher Benot
Ya'aqovGoren-Inbaret al., 2000
c. 700 x
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
Concentration 2, apparently indicating the rainy season inwinter (MAP ranges between 300 and 700 mm, Table 5).Accordingly, values for the driest and warmest quarter as well asthe driest month are low (below 100 mm) indicating dry sum-mer. This is in accordance with the general Mediterranean cli-matic regime.
At the moment, the data for the precipitation of the driest andwarmest quarters cannot clearly indicate more humid conditionsin summer, because modern values are 0 mm and the upperbound of any range must be higher. Still, taking lower precipita-tion during the rainy season and similar or slightly higher rainfallduring the dry season into account, the seasonality of precipita-tion was decreased compared to the present day. However, themean annual precipitation seems to have been not affectedconsiderably, which indicates mainly more evenly distributedrainfall during the year.
m the Middle Pleistocene until today.
cidurinaee form
Suncusetruscus
Rousettussp.
Rhinolophusferrumequinum
Rhinolophuseuryale
Rhinolophusmehelyi
x x x x x
lg x
x
x x x x xcf x x
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

L.C. Maul et al. / Quaternary International xxx (2015) 1e14 9
3.2. Biostratigraphy
To the best of our knowledge, no reliable species-level dataexist on the distribution of reptiles in the Pleistocene Levant.Procavia capensis, Suncus etruscus and Crocidura leucodon occurredat least from the Middle Pleistocene onwards and still live in Israeltoday. Rousettus was previously thought to occur in the Levantonly since the very end of the Pleistocene or the Holocene.However, Qesem Cave provided the first Pleistocene record of thisgenus in this region (Hor�a�cek et al., 2013). Rhinolophus ferrume-quinum, R. euryale, R. mehelyi, Miniopterus schreibersii, Myotisblythii and Lepus capensis are supposed to occur in the area notearlier than the Middle Pleistocene and do not indicate a partic-ular geological age. Sciurus anomalus is known from severalPleistocene sites in Israel, but always in low numbers. Cricetulusmigratorius, Spalax ehrenbergi, Gerbillus dasyurus, Meriones tris-trami and Mus macedonicus are also known since the MiddlePleistocene (see Table 6).
Biostratigraphic information can be obtained from Myomimus,which is not known from the recent fauna in Israel. However, M.judaicus is recorded from Lower Palaeolithic Acheulian Ubeidiyaand from the Upper Acheulian layer D2 of Oumm Qatafa (Haas,1951; Tchernov, 1996). The latter was dated by ESR to 215 ka(Porat et al., 2002) which, however, presumably underestimates itstrue age (Gopher et al., 2010). M. qafzehensis and M. roachi arerecorded after the Middle Palaeolithic (Mousterian) of Qafzeh,species which differ slightly from our finds (Smith et al., this vol-ume). If the close taxonomic relation between Myomimus judaicusfrom Oumm Qatafa and Myomimus sp. from the M. judaicus/setzerigroup from Qesem Cave should be confirmed, a minimum age of215 kawould be implied for Concentration 2. However, we considerthe dating of Porat et al. (2002) as much too young and it is morethan plausible following the Qesem stratigraphy and radiometricdating (Mercier et al., 2013; Falgueres et al. this volume) to assignthis concentration a date no younger than 300 ka and possiblysomewhat older (see also below).
Rattus haasi is also known from the Upper Acheulian layer D2 ofOummQatafa (Haas, 1951; Tchernov, 1968) but not in younger sites.The absence of R. haasi in Mousterian sites thus corroborates thepre-Mousterian age for Concentration 2 at Qesem.
Morphometric data of Microtus finds provide further strati-graphic information. The general evolutionary trend of an increaseof the A/L ratio inMicrotus starts with values of approximately 40 inthe oldest (c. 1.8e2.0 Ma), most primitive members of the genusfrom Europe and Asia, referred to as Microtus (Allophaiomys)deucalion (e.g., Vill�any 5 in Hungary e van der Meulen, 1974; Tizdar
Miniopterusschreibersii
Myotisblythii
Lepuscapensis
Sciurusanomalus
Myomimusjudaicus
Cricetulusmigratorius
Microtusguentheri
Gd
x x x x x x
x x xx
x xx x xx x x
x x x
x xcf x cf cf gr xcf cf cf gr gr x
x
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
in Ukrainee Pevzner et al., 1998; Tesakov,1998), and reaches valuesup to 56 in recent samples of various Microtus species(Nadachowski, 1982). Fig. 5 shows the general trend, which israther clearly visible over thewhole stratigraphic range ofMicrotus.Beginning at c. 1.0 Ma, the most primitive forms of Microtus(commonly grouped in the subgenus Allophaiomys) branch intovarious lineages, which are often referred to distinct subgenera:Pallasiinus, Stenocranius, Terricola, Iberomys, and Microtus (seeFig. 5). In Middle and Late Pleistocene sites of the Near East,Microtus guentheri is the only Microtus species. Morphologicallyand genetically it is closely related toM. arvalis (subgenusMicrotus,indicated with a ‘þ’ in Fig. 5) (Jaarola et al., 2004).
In Fig. 6, A/L mean values (rhombs), the 95% confidence intervals(rectangles) and the potential maximal age range (vertical lines) ofthe samples are plotted (mean values and confidence interval arealways drawn in the middle of the age line); the individual data aregiven in Table 7. The A/L of the Microtus samples follow a visibledirection, beginning with the data from Gesher Benot Ya'aqov(Acheulian), through Tabun E (Acheulo-Yabrudian), Tabun D, C andB (Mousterian) up to Recent samples from north and central Israel.Data fromQesem Cave and Tabun Eb, are all between 54 and 55 andfit well in the interval of the Acheulo-Yabrudian Cultural Complexbetween 400 ka and 200 ka.
However, A/L values of around 54 occur also in samples from thesemi-desert at the northern margin of the Negev, showing theexistence of regional differences within Israel with the lowest meanvalues in the Negev, and the highest mean values in the Hula Basin.It is therefore important to compare only regional values, particu-larly if we deal with extremely different habitats, such as wetland(Hula valley), shrubland (Jezreel valley) and semi-desert (Negev).
The data of the fossil samples displaymore progressive values inQesem Concentration 1, which is similar to those from Tabun D (c.200 ka), than in Concentration 2. These results unambiguouslyimply that the two concentrations were not deposited at the sametime. The trend documented above suggests a greater age forConcentration 2, which is also in agreement with the older ageindicated by the occurrence of Rattus haasi andMyomimus judaicus/setzeri in this sample (see above and Table 6). Concentration 2 haseven lower values than Tabun Eb. An age of c. 250e380 ka has beenestimated for a lower level of Tabun, Ed, (discussion in Gopher et al.,2010), so the age of Eb is probably younger than the lowerboundary of this range.
Concentration 2 can be related to the so called Deep Pit Area (or‘The Southern Area’) of Qesem Cave. The radiometric dating for theupper and lower part of this area lies mostly in a 200e350 ka timeinterval, the TL and ESR/U-dating of the upper part can be mainly
erbillusasyurus
Merionestristrami
Gerbillidaelarge form
Musmacedonicus
Rattushaasi
Apodemusflavicollis
Spalaxehrenbergi
x x x x x
x x x xx x
x x cf xx x xx x x
x x x x X
x x xcf cf x gr gr grcf cf gr cf gr grx x x
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032
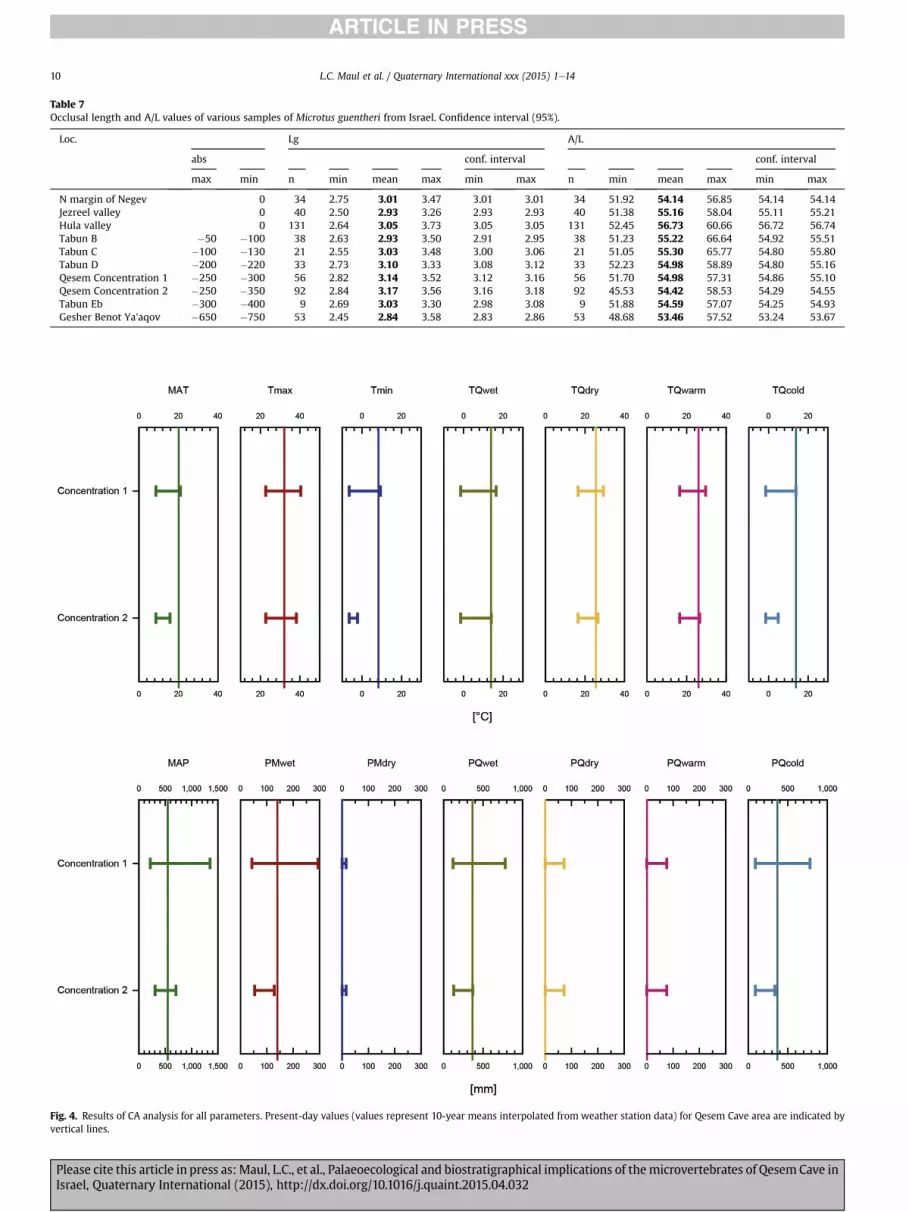
Fig. 4. Results of CA analysis for all parameters. Present-day values (values represent 10-year means interpolated from weather station data) for Qesem Cave area are indicated byvertical lines.
Table 7Occlusal length and A/L values of various samples of Microtus guentheri from Israel. Confidence interval (95%).
Loc. Lg A/L
abs conf. interval conf. interval
max min n min mean max min max n min mean max min max
N margin of Negev 0 34 2.75 3.01 3.47 3.01 3.01 34 51.92 54.14 56.85 54.14 54.14Jezreel valley 0 40 2.50 2.93 3.26 2.93 2.93 40 51.38 55.16 58.04 55.11 55.21Hula valley 0 131 2.64 3.05 3.73 3.05 3.05 131 52.45 56.73 60.66 56.72 56.74Tabun B �50 �100 38 2.63 2.93 3.50 2.91 2.95 38 51.23 55.22 66.64 54.92 55.51Tabun C �100 �130 21 2.55 3.03 3.48 3.00 3.06 21 51.05 55.30 65.77 54.80 55.80Tabun D �200 �220 33 2.73 3.10 3.33 3.08 3.12 33 52.23 54.98 58.89 54.80 55.16Qesem Concentration 1 �250 �300 56 2.82 3.14 3.52 3.12 3.16 56 51.70 54.98 57.31 54.86 55.10Qesem Concentration 2 �250 �350 92 2.84 3.17 3.56 3.16 3.18 92 45.53 54.42 58.53 54.29 54.55Tabun Eb �300 �400 9 2.69 3.03 3.30 2.98 3.08 9 51.88 54.59 57.07 54.25 54.93Gesher Benot Ya'aqov �650 �750 53 2.45 2.84 3.58 2.83 2.86 53 48.68 53.46 57.52 53.24 53.67
L.C. Maul et al. / Quaternary International xxx (2015) 1e1410
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and biostratigraphical implications of themicrovertebrates of Qesem Cave inIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint.2015.04.032
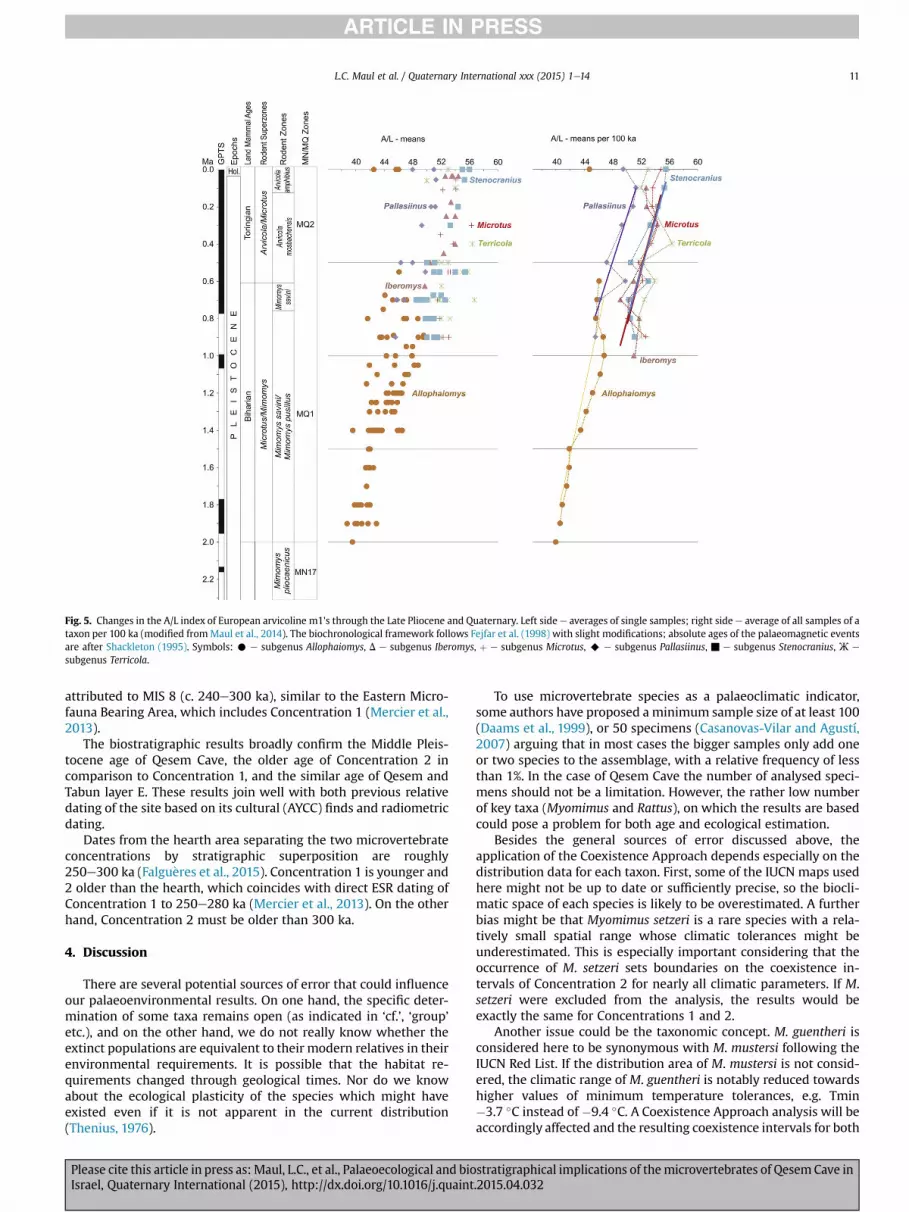
Fig. 5. Changes in the A/L index of European arvicoline m1's through the Late Pliocene and Quaternary. Left side e averages of single samples; right side e average of all samples of ataxon per 100 ka (modified fromMaul et al., 2014). The biochronological framework follows Fejfar et al. (1998) with slight modifications; absolute ages of the palaeomagnetic eventsare after Shackleton (1995). Symbols: C e subgenus Allophaiomys, D e subgenus Iberomys, þ e subgenus Microtus, A e subgenus Pallasiinus, ▪ e subgenus Stenocranius, Ж e
subgenus Terricola.
L.C. Maul et al. / Quaternary International xxx (2015) 1e14 11
attributed to MIS 8 (c. 240e300 ka), similar to the Eastern Micro-fauna Bearing Area, which includes Concentration 1 (Mercier et al.,2013).
The biostratigraphic results broadly confirm the Middle Pleis-tocene age of Qesem Cave, the older age of Concentration 2 incomparison to Concentration 1, and the similar age of Qesem andTabun layer E. These results join well with both previous relativedating of the site based on its cultural (AYCC) finds and radiometricdating.
Dates from the hearth area separating the two microvertebrateconcentrations by stratigraphic superposition are roughly250e300 ka (Falgu�eres et al., 2015). Concentration 1 is younger and2 older than the hearth, which coincides with direct ESR dating ofConcentration 1 to 250e280 ka (Mercier et al., 2013). On the otherhand, Concentration 2 must be older than 300 ka.
4. Discussion
There are several potential sources of error that could influenceour palaeoenvironmental results. On one hand, the specific deter-mination of some taxa remains open (as indicated in ‘cf.’, ‘group’etc.), and on the other hand, we do not really know whether theextinct populations are equivalent to their modern relatives in theirenvironmental requirements. It is possible that the habitat re-quirements changed through geological times. Nor do we knowabout the ecological plasticity of the species which might haveexisted even if it is not apparent in the current distribution(Thenius, 1976).
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
To use microvertebrate species as a palaeoclimatic indicator,some authors have proposed a minimum sample size of at least 100(Daams et al., 1999), or 50 specimens (Casanovas-Vilar and Agustí,2007) arguing that in most cases the bigger samples only add oneor two species to the assemblage, with a relative frequency of lessthan 1%. In the case of Qesem Cave the number of analysed speci-mens should not be a limitation. However, the rather low numberof key taxa (Myomimus and Rattus), on which the results are basedcould pose a problem for both age and ecological estimation.
Besides the general sources of error discussed above, theapplication of the Coexistence Approach depends especially on thedistribution data for each taxon. First, some of the IUCN maps usedhere might not be up to date or sufficiently precise, so the biocli-matic space of each species is likely to be overestimated. A furtherbias might be that Myomimus setzeri is a rare species with a rela-tively small spatial range whose climatic tolerances might beunderestimated. This is especially important considering that theoccurrence of M. setzeri sets boundaries on the coexistence in-tervals of Concentration 2 for nearly all climatic parameters. If M.setzeri were excluded from the analysis, the results would beexactly the same for Concentrations 1 and 2.
Another issue could be the taxonomic concept. M. guentheri isconsidered here to be synonymous with M. mustersi following theIUCN Red List. If the distribution area of M. mustersi is not consid-ered, the climatic range of M. guentheri is notably reduced towardshigher values of minimum temperature tolerances, e.g. Tmin�3.7 �C instead of �9.4 �C. A Coexistence Approach analysis will beaccordingly affected and the resulting coexistence intervals for both
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032
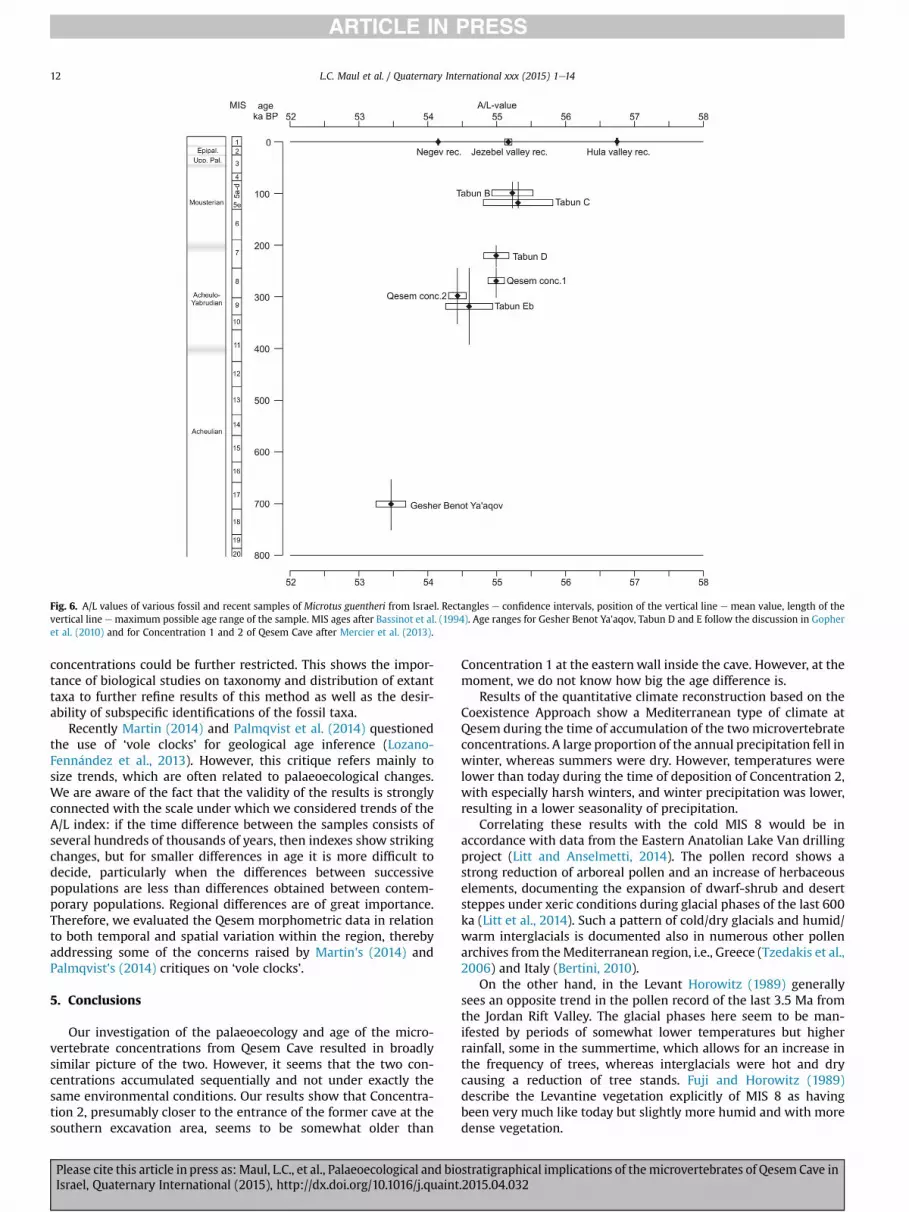
Fig. 6. A/L values of various fossil and recent samples of Microtus guentheri from Israel. Rectangles e confidence intervals, position of the vertical line e mean value, length of thevertical line e maximum possible age range of the sample. MIS ages after Bassinot et al. (1994). Age ranges for Gesher Benot Ya'aqov, Tabun D and E follow the discussion in Gopheret al. (2010) and for Concentration 1 and 2 of Qesem Cave after Mercier et al. (2013).
L.C. Maul et al. / Quaternary International xxx (2015) 1e1412
concentrations could be further restricted. This shows the impor-tance of biological studies on taxonomy and distribution of extanttaxa to further refine results of this method as well as the desir-ability of subspecific identifications of the fossil taxa.
Recently Martin (2014) and Palmqvist et al. (2014) questionedthe use of ‘vole clocks’ for geological age inference (Lozano-Fenn�andez et al., 2013). However, this critique refers mainly tosize trends, which are often related to palaeoecological changes.We are aware of the fact that the validity of the results is stronglyconnected with the scale under which we considered trends of theA/L index: if the time difference between the samples consists ofseveral hundreds of thousands of years, then indexes show strikingchanges, but for smaller differences in age it is more difficult todecide, particularly when the differences between successivepopulations are less than differences obtained between contem-porary populations. Regional differences are of great importance.Therefore, we evaluated the Qesem morphometric data in relationto both temporal and spatial variation within the region, therebyaddressing some of the concerns raised by Martin's (2014) andPalmqvist's (2014) critiques on ‘vole clocks’.
5. Conclusions
Our investigation of the palaeoecology and age of the micro-vertebrate concentrations from Qesem Cave resulted in broadlysimilar picture of the two. However, it seems that the two con-centrations accumulated sequentially and not under exactly thesame environmental conditions. Our results show that Concentra-tion 2, presumably closer to the entrance of the former cave at thesouthern excavation area, seems to be somewhat older than
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
Concentration 1 at the eastern wall inside the cave. However, at themoment, we do not know how big the age difference is.
Results of the quantitative climate reconstruction based on theCoexistence Approach show a Mediterranean type of climate atQesem during the time of accumulation of the two microvertebrateconcentrations. A large proportion of the annual precipitation fell inwinter, whereas summers were dry. However, temperatures werelower than today during the time of deposition of Concentration 2,with especially harsh winters, and winter precipitation was lower,resulting in a lower seasonality of precipitation.
Correlating these results with the cold MIS 8 would be inaccordance with data from the Eastern Anatolian Lake Van drillingproject (Litt and Anselmetti, 2014). The pollen record shows astrong reduction of arboreal pollen and an increase of herbaceouselements, documenting the expansion of dwarf-shrub and desertsteppes under xeric conditions during glacial phases of the last 600ka (Litt et al., 2014). Such a pattern of cold/dry glacials and humid/warm interglacials is documented also in numerous other pollenarchives from theMediterranean region, i.e., Greece (Tzedakis et al.,2006) and Italy (Bertini, 2010).
On the other hand, in the Levant Horowitz (1989) generallysees an opposite trend in the pollen record of the last 3.5 Ma fromthe Jordan Rift Valley. The glacial phases here seem to be man-ifested by periods of somewhat lower temperatures but higherrainfall, some in the summertime, which allows for an increase inthe frequency of trees, whereas interglacials were hot and drycausing a reduction of tree stands. Fuji and Horowitz (1989)describe the Levantine vegetation explicitly of MIS 8 as havingbeen very much like today but slightly more humid and with moredense vegetation.
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

L.C. Maul et al. / Quaternary International xxx (2015) 1e14 13
At first glance, the observed reduction of winter precipitation inour climate reconstruction based on the micromammal assem-blages from Qesem seems to be inconsistent with the results ofHorowitz (Fuji and Horowitz, 1989; Horowitz, 1989). However, asdiscussed above (Fig. 4) our data do not rule out slightly highersummer precipitation. It may well be that summer precipitation ismore important for sustaining tree stands in the Levant than annualor winter precipitation. It is very likely that the more even distri-bution of rainfall over the year as documented in our data (Fig 3) iskey for supporting higher vegetation density. Finally, the mosaic ofhabitats seems to be not uncommon for the environment of theregion under consideration, as north- and south-facing slopes ofeast-west trending valleys e including the Wadi Rabah in whichQesem Cave is situated e show considerable differences in micro-habitat characteristics (Nevo, 1995).
Acknowledgements
We are indebted to the QesemCave excavation team for the timeconsuming picking of microvertebrate samples during the fieldseasons, and to the students at Tel Aviv University and ArlettUlbricht (Jena) for picking and sorting works in the lab. We alsowould like to thank Prof. Tamar Dayan, Dr. Motti Charter and AriehLandsman of the Tel Aviv Zoological Museum for access tocomparative material of extant Microtus guentheri. For financialsupport we thank the Fritz Thyssen Foundation, the Israel ScienceFoundation, the Leakey Foundation, the Wenner Gren Foundation,and the CARE Archaeological Foundation.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.quaint.2015.04.032.
References
Baha El Din, S., 2006. A Guide to the Reptiles and Amphibians of Egypt. TheAmerican University in Cairo Press, Cairo.
Barkai, R., Gopher, A., Lauritzen, S.E., Frumkin, A., 2003. Uranium series dates fromQesem Cave, Israel, and the end of the Lower Palaeolithic. Nature 423,977e979.
Barkai, R., Lemorini, C., Shimelmitz, R., Lev, Z., Stiner, M.C., Gopher, A., 2009. A bladefor all seasons? Making and using Amudian blades at Qesem Cave, Israel. Hu-man Evolution 24, 57e75.
Barkai, R., Lemorini, C., Gopher, A., 2010. Palaeolithic cutlery 400 000e200 000years ago: tiny meat-cutting tools from Qesem Cave, Israel. Antiquity 84.
Barkai, R., Gopher, A., 2011. Innovative human behavior between Acheulian andMousterian: a view from Qesem Cave, Israel. In: Le Tensorer, J.-M., Jagher, R.,Otte, M. (Eds.), The Lower and Middle Palaeolithic in the Middle East andNeighbouring Regions. Universit�e de Liege, Liege.
Barkai, R., Gopher, A., Solodenko, N., Lemorini, C., 2013. An Amudian Oddity: a giantbiface from Late Lower Palaeolithic Qesem Cave. Tel Aviv 40, 176e186.
Bassinot, F.C., Labeyrie, L.D., Vincent, E., Quidelleur, X., Shackleton, N.J., Lancelot, Y.,1994. The astronomical theory of climate and the age of the Brunhes-Matuyamamagnetic reversal. Earth and Planetary Science Letters 126, 91e108.
Bertini, A., 2010. Pliocene to Pleistocene palynoflora and vegetation in Italy: state ofthe art. Quaternary International 225, 5e24.
Blain, H.-A., Gleed-Owen, C.P., L�opez-García, J.M., Carri�on, J.S., Jennings, R.,Finlayson, G., Finlayson, C., Giles-Pacheco, F., 2013. Climatic conditions for thelast Neanderthals: herpetofaunal record of Gorham's Cave, Gibraltar. Journal ofHuman Evolution 64, 289e299.
Blain, H.-A., Agustí, J., Lordkipanidze, D., Rook, L., Delfino, M., 2014. Paleoclimaticand paleoenvironmental context of the Early Pleistocene hominins fromDmanisi (Georgia, Lesser Caucasus) inferred from the herpetofaunal assem-blage. Quaternary Science Reviews 105, 136e150.
Blasco, R., Rosell, J., Cuartero, F., Fern�andez Peris, J., Gopher, A., Barkai, R., 2013.Using bones to shape stones: MIS 9 bone retouchers at both edges of theMediterranean Sea. PLoS One 8, e76780.
Blasco, R., Rosell, J., Gopher, A., Barkai, R., 2014. Subsistence economy and social life:a zooarchaeological view from the 300 kya central hearth at Qesem Cave, Israel.Journal of Anthropological Archaeology 35, 248e268.
Bruch, A.A., Uhl, D., Mosbrugger, V., 2007. Miocene climate in Europe d Patternsand evolution: a first synthesis of NECLIME. Palaeogeography PalaeoclimtologyPalaeoecology 253, 1e7.
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
Bruch, A.A., Utescher, T., Mosbrugger, V., 2011. Precipitation patterns in the Mioceneof Central Europe and the development of continentality. PalaeogeographyPalaeoclimtology Palaeoecology 304, 202e211.
Bruch, A.A., Sievers, C., Wadley, L., 2012. Quantification of climate and vegetationfrom southern African Middle Stone Age sites e an application using LatePleistocene plant material from Sibudu, South Africa. Quaternary Science Re-views 45, 7e17.
Casanovas-Vilar, I., Agustí, J., 2007. Ecogeographical stability and climate forcing inthe Late Miocene (Vallesian) rodent record of Spain. Palaeogeography Palae-oclimtology Palaeoecology 248, 169e189.
Daams, R., Meulen, A.J.v.d., Pel�aez-Campomanes, P., �Alvarez Sierra, �A., 1999. Trendsin rodent assemblages from the Aragonian (earlyemiddle Miocene) of theCalatayud-Daroca Basin, Arag�on, Spain, 127e139. In: Agustí, J., Rook, L.,Andrews, P. (Eds.), Hominoid Evolution and Climatic Change in Europe. TheEvolution of Terrestrial Ecosystems in Europe. Cambridge University Press,Cambridge.
Disi, A.M., Modrý, D., Ne�cas, P., Rifai, L., 2001. Amphibians and Reptiles of theHashemite Kingdom of Jordan. Edition Chimaira, Frankfurt/Main.
Falgu�eres, C., Richard, M., Tombret, O., Shao, Q., Bahain, J.J., Gopher, A., Barkai, R.,2015. New ESR/U-series dates in Yabrudian and Amudian layers at Qesem Cave,Israel. Quaternary International. http://dx.doi.org/10.1016/j.quaint.2015.02.006(in press).
Fejfar, O., Heinrich, W.-D., Lindsay, E.H., 1998. Updating the neogene rodent bio-chronology in Europe. Mededelingen Nederlands Instituut voor Geo-wetenschappen TNO 60, 533e554.
Filippucci, M.G., Simson, S., Nevo, E., 1989. Evolutionary biology of the genus Apo-demus Kaup, 1829 in Israel. Allozymic and biometric analyses with descriptionof a new species: Apodemus hermonensis (Rodentia, Muridae). Bolletino dizoologia 56, 361e376.
Fuji, N., Horowitz, A., 1989. Brunhes Epoch paleoclimates of Japan and Israel.Palaeogeogr Palaeocl 72, 79e88.
Gopher, A., Barkai, R., Shimelmitz, R., Khalaly, M., Lemorini, C., Hershkovitz, I.,Stiner, M.C., 2005. Qesem Cave: an Amudian site in Central Israel. Journal of theIsrael Prehistoric Society 35, 69e92.
Gopher, A., Ayalon, A., Bar-Matthews, M., Barkai, R., Frumkin, A., Karkanas, P.,Shahack-Gross, R., 2010. The chronology of the Late Lower Paleolithic in theLevant: U series dates of speleothems from Middle Pleistocene Qesem cave,Israel. Quaternary Geochronology 5, 644e656.
Goren-Inbar, N., Feibel, C.S., Verosub, K.L., Melamed, Y., Kislev, M.E., Tchernov, E.,Saragusti, I., 2000. Pleistocene Milestones on the Out-of-Africa Corridor atGesher Benot Ya'aqov. Israel. Science 289, 944e947.
Haas, G., 1951. II. Remarques sur la microfaune de mammif�eres de la Grotte d'Oum-Qatafa. In: Neuville, R. (Ed.), Le Pal�eolithique et le M�esolithique du d�esert deJud�ee. Masson et Cie Editeurs, Paris, pp. 218e233.
Harrison, D.L., Bates, P.J.J., 1991. The Mammals of Arabia. Harrison ZoologicalMuseum Publication, Sevenoaks.
Heinrich, W.-D., 1978. Zur biometrischen Erfassung eines Evolutionstrends beiArvicola (Rodentia, Mammalia) aus dem Pleistoz€an Thüringens.S€augetierkundliche Informationen 2, 3e21.
Hershkovitz, I., Smith, P., Sarig, R., Quam, R., Rodríguez, L., García, R., Arsuaga, J.L.,Barkai, R., Gopher, A., 2011. Middle Pleistocene dental remains from Qesem Cave(Israel). American Journal of Physical Anthropology 144, 575e592.
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G., Jarvis, A., 2005. Very high reso-lution interpolated climate surfaces for global land areas. International Journalof Climatology 25, 1965e1978.
Hor�a�cek, I., Maul, L.C., Smith, K.T., Gopher, A., Barkai, R., 2013. Bat remains (Mam-malia, Chiroptera) from the Middle Pleistocene site of Qesem Cave, Israel, withthe first Pleistocene record of fruit bats in the Mediterranean region. Palae-ontologia Electronica 16. Article number 16.13.23 A.
Horowitz, A., 1989. Continuous pollen diagrams for the last 3.5 m.y. from Israel:Vegetation, climate and correlation with the oxygen isotope record. Palae-ogeography Palaeoclimtology Palaeoecology 72, 63e78.
Jaarola, M., Martínkov�a, N., Gündüz, I., Brunhoff, C., Zima, J., Nadachowski, A.,Amori, G., Bulatova, N.S., Chondropoulos, B., Fraguedakis-Tsolis, S.,Gonz�alez-Esteban, J., Jos�e L�opez-Fuster, M., Kandaurov, A.S., Kefelioglu, H.,da Luz Mathias, M., Villate, I., Searle, J.B., 2004. Molecular phylogeny of thespeciose vole genus Microtus (Arvicolinae, Rodentia) inferred from mito-chondrial DNA sequences. Molecular Phylogenetics and Evolution 33,647e663.
Kry�stufek, B., Vohralík, V., 2001. Mammals of Turkey and Cyprus. Science andResearch Centre of the Republic of Slovenia, Koper.
Kry�stufek, B., Vohralík, V., 2005. Mammals of Turkey and Cyprus. Rodentia I:Sciuridae, Dipodidae, Gliridae, Arvicolinae. Science and Research Centre of theRepublic of Slovenia, Koper.
Kry�stufek, B., Vohralík, V., 2009. Mammals of Turkey and Cyprus. Rodentia II: Cri-cetinae, Muridae, Spalacidae, Calomyscidae, Capromyidae, Hystricidae, Castor-idae. Science and Research Centre of the Republic of Slovenia, Koper.
Lebedev, V.S., 2012. Subfamily Cricetinae s. str. In: Pavlinov, I.Y., Lissovsky, A.A.(Eds.), The Mammals of Russia. A Taxonomic and Geographic Reference. KMKScientific Press Ltd., Moscow, pp. 211e220.
Litt, T., Anselmetti, F.S., 2014. Lake Van deep drilling project PALEOVAN. QuaternaryScience Reviews 104, 1e7.
Litt, T., Pickarski, N., Heumann, G., Stockhecke, M., Tzedakis, P.C., 2014. A 600,000year long continental pollen record from Lake Van, eastern Anatolia (Turkey).Quaternary Science Reviews 104, 30e41.
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032

L.C. Maul et al. / Quaternary International xxx (2015) 1e1414
Lozano-Fenn�andez, I., Agustí, J., Cuenca-Besc�os, G., Blain, H.A., L�opez-García, J.M.,Vallverdú, J., 2013. Pleistocene evolutionary trends in dental morphology ofMimomys savini (Rodentia, Mammalia) from Iberian peninsula and discussionabout the origin of the genus Arvicola. Quaternaire 24, 179e190.
Marder, O., Yeshurun, R., Lupu, R., Bar-Oz, G., Belmaker, M., Porat, N., Ron, H.,Frumkin, A., 2011. Mammal remains at Rantis Cave, Israel, and MiddleeLatePleistocene human subsistence and ecology in the Southern Levant. Journal ofQuaternary Science n/aen/a.
Martin, R.A., 2014. A Critique of Vole Clocks Quaternary Science Reviews 94.Maul, L.C., Smith, K.T., Barkai, R., Barash, A., Karkanas, P., Shahack-Gross, R.,
Gopher, A., 2011. Microfaunal remains at Middle Pleistocene Qesem Cave, Israel:preliminary results on small vertebrates, environment and biostratigraphy.Journal of Human Evolution 60, 464e480.
Maul, L.C., Masini, F., Parfitt, S.A., Rekovets, L.I., Savorelli, A., 2014. Evolutionarytrends in arvicolids and the endemic murid Mikrotia e new data and a criticaloverview. Quaternary Science Reviews 96, 240e258.
Mendelssohn, H., Yom-Tov, Y., 1999. Fauna Palaestina. Mammalia of Israel. The IsraelAcademy of Sciences and Humanities, Jerusalem.
Mercier, N., Valladas, H., Falgu�eres, C., Shao, Q., Gopher, A., Barkai, R., Bahain, J.-J.,Vialettes, L., Joron, J.-L., Reyss, J.-L., 2013. New datings of Amudian layers atQesem Cave (Israel): results of TL applied to burnt flints and ESR/U-series toteeth. Journal of Archaeological Science 40, 3011e3020.
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., Kry�stufek, B., Reijnders, P.J.H.,Spitzenberger, F., Stubbe, M., Thissen, J.B.M., Vohralík, V., Zima, J., 1999. TheAtlas of European Mammals. Academic Press, London.
Mosbrugger, V., Utescher, T., 1997. The coexistence approach - a method for quan-titative reconstructions of tertiary terrestrial palaeoclimate data using plantfossils. Palaeogeography Palaeoclimtology Palaeoecology 134, 61e86.
Nadachowski, A., 1982. Late Quaternary Rodents of Poland with Special Reference toMorphotype Dentition Analysis of Voles. Polska Akademia Nauk, Zaklad ZoologiSyst. Dosw., Warszawa, Krak�ow.
Nevo, E., Guttman, R., Haber, M., Erez, E., 1979. Habitat selection in evolving molerats. Oecologia 43, 125e138.
Nevo, E., 1995. Asian, African and European biota meet at 'Evolution Canyon' Israel:local test of global diversity and genetic diversity patterns. Proceedings of theRoyal Society London (B) 262, 149e155.
Palmqvist, P., Gonz�alez-Donoso, J.M., De Renzi, M., 2014. Rectilinear evolution inarvicoline rodents and numerical dating of Iberian Early Pleistocene sites.Quaternary Science Reviews 98, 100e109.
Pevzner, M., Tesakov, A.S., Vangengeim, E.A., 1998. The position of the Tizdar locality(Taman Peninsula, Russia) in the magnetochronological scale. Paludicola 2,95e97.
Porat, N., Chazan, M., Schwarcz, H.P., Kolska Horwitz, L., 2002. Timing of the Lowerto Middle Paleolithic boundary: new dates from the Levant. Journal of HumanEvolution 43, 107e122.
Rabeder, G., 1981. Die Arvicoliden (Rodentia, Mammalia) aus dem Plioz€an und dem€alteren Pleistoz€an von Nieder€osterreich. Beitr€age zur Pal€aontologie von€Osterreich 8, 1e373.
Rosell, J., Blasco, R., Fern�andez Peris, J., Carbonell, E., Barkai, R., Gopher, A., 2015.Recycling bones in the Middle Pleistocene: some reflections from Gran DolinaTD10-1 (Spain), Bolomor Cave (Spain) and Qesem Cave (Israel). QuaternaryInternational 361, 297e312.
Shackleton, N.J., 1995. New data on the evolution of Pliocene climatic variability. In:Vrba, E.S., Denton, G.H., Partridge, T.C., Burckle, L.H. (Eds.), Paleoclimate andEvolution, with Emphasis on Human Origins. Yale University Press, New Haven,London, pp. 249e261.
Please cite this article in press as: Maul, L.C., et al., Palaeoecological and bioIsrael, Quaternary International (2015), http://dx.doi.org/10.1016/j.quaint
Shenbrot, G.I., Krasnov, B.R., 1997. Additional records of two small mammalianspecies in the Central Negev. Israel Journal of Zoology 43, 299e300.
Shenbrot, G.I., Krasnov, B.R., Khokhlova, I.S., 1997. Biology of Wagner's gerbil Ger-billus dasyurus (Wagner, 1842) (Rodentia: Gerbillidae) in the Negev Highlands,Israel. Mammalia 61, 467e486.
Shimelmitz, R., Barkai, R., Gopher, A., 2011. Systematic blade production at lateLower Paleolithic (400e200 kyr) Qesem Cave, Israel. Journal of Human Evolu-tion 61, 458e479.
Smith, K.T., Maul, L.C., Barkai, R., Gopher, A., 2013. To catch a chameleon, or actu-alism vs. natural history in the taphonomy of the microvertebrate fraction atQesem Cave, Israel. Journal of Archaeological Science 40, 3326e3339.
Smith, K.T., Maul, L.C., Barkai, R., Gopher, A., this volume. The microvertebrates ofQesem Cave: A comparison of the two main concentrations. QuaternaryInternational.
Stiner, M.C., Gopher, A., Barkai, R., 2009. Cooperative hunting and meat sharing400-200 kya at Qesem Cave, Israel. Proceedings of the National Academy ofSciences U.S.A 106, 13207e13212.
Stiner, M.C., Gopher, A., Barkai, R., 2011. Hearth-side socioeconomics, hunting andpaleoecology during the late Lower Paleolithic at Qesem Cave, Israel. Journal ofHuman Evolution 60, 213e233.
Tchernov, E., 1968. Succession of rodent faunas during the Upper Pleistocene ofIsrael. In: Herre, W., R€ohrs, M. (Eds.), Mammalia Depicta. Paul Parey, Hamburg,Berlin, pp. 1e152.
Tchernov, E., 1996. Rodent faunas, chronstratigraphy and paleobiogeography of thesouthern Levant during the Quaternary. Acta zoologica cracoviensia 39,513e530.
Tesakov, A.S., 1998. Early stage of Allophaiomys evolution in Eastern Europe. Pal-udicola 2, 98e105.
Thenius, E., 1976. Pleistoz€ane S€augetiere als Klima-Indikatoren. Archaeologia Aus-triaca Beiheft 13, 91e112.
Tzedakis, P.C., Hooghiemstra, H., P€alike, H., 2006. The last 1.35 million years atTenaghi Philippon: revised chronostratigraphy and long-term vegetationtrends. Quaternary Science Reviews 25, 3416e3430.
Utescher, T., B€ohme, M., Mosbrugger, V., 2011. The Neogene of Eurasia: spatialgradients and temporal trends d The second synthesis of NECLIME. Palae-ogeography Palaeoclimtology Palaeoecology 304, 196e201.
Utescher, T., Bruch, A.A., Erdei, B., François, L., Ivanov, D., Jacques, F.M.B., Kern, A.K.,Liu, Y.S., Mosbrugger, V., Spicer, R.A., 2014. The coexistence approachdTheor-etical background and practical considerations of using plant fossils for climatequantification. Palaeogeography Palaeoclimtology Palaeoecology 410, 58e73.
van der Meulen, A.J., 1973. Middle Pleistocene smaller mammals from the MontePeglia (Orvieto, Italy) with special reference to the phylogeny of Microtus(Arvicolidae, Rodentia). Quaternaria 17, 1e144.
van der Meulen, A.J., 1974. On Microtus (Allophaiomys) deucalion (Kretzoi 1969)(Arvicolidae, Rodentia) from the upper Vill�anyian (Lower Pleistocene) ofVill�any-5, S. Hungary. Koniklijke Nederlandse Akadamie Wetenschapen. Pro-ceedings, Series B 77, 259e266.
Waltari, E., Hijmans, R.J., Peterson, A.T., Ny�ari, �A.S., Perkins, S.L., Guralnick, R.P., 2007.Locating Pleistocene Refugia: comparing Phylogeographic and ecological NicheModel Predictions. PLoS One 2 (7), e563. http://dx.doi.org/10.1371/journal.pone.0000563.
Weissbrod, L., Zaidner, Y., 2014. Taphonomy and paleoecological implications offossorial microvertebrates at the Middle Paleolithic open-air site of NesherRamla, Israel. Quaternary International 331, 115e127.
stratigraphical implications of themicrovertebrates of Qesem Cave in.2015.04.032




















