
Genetic Disruption of VIP Signaling Alters Intestinal Microbial ...
Upload
independentCategory
view
3download
0
ANTIOXIDANTS amp REDOX SIGNALINGVolume 3 Number 4 2001Mary Ann Liebert Inc
Original Research Communication
v-Ha-Ras Overexpression Induces Superoxide Productionand Alters Levels of Primary Antioxidant Enzymes
JI-QIN YANG SHIJUN LI YUANHUI HUANG HANNAH J ZHANG FREDERICK E DOMANN GARRY R BUETTNER and LARRY W OBERLEY
ABSTRACT
Reactive oxygen species have been shown to play important roles in v-Ha-Ras mitogenic signaling We hypothe-sized that v-Ha-Ras overexpression would induce superoxide production and therefore modify expression of theprimary antioxidant enzyme system We have demonstrated that immortal rat kidney epithelial cells stably trans-duced with constitutively active v-Ha-ras produced significantly larger amounts of superoxide radical than wild-type or vector-transfected control cells The levels of the primary antioxidant enzymes copper- and zinc-contain-ing superoxide dismutase manganese-containing superoxide dismutase catalase and glutathione peroxidase wereincreased in the superoxide-overproducing cells DNA-binding activities of the transcription factors activator pro-tein-1 activator protein-2 and nuclear factor-kB were all enhanced in the superoxide-overproducing cells Thesev-Ha-ras transduced cells also had a shortened cell doubling time and higher plating efficiency and displayedgreater constitutive levels of phosphorylated mitogen-activated protein kinases These data demonstrate that v-Ha-Ras overexpression increases superoxide production and this apparently affects a wide variety of cell sig-naling and redox systems Antioxid Redox Signal 3 697ndash709
697
INTRODUCTION
ONCOGENIC RAS is one of the most studiedgenes in cancer 30 of all human tu-
mors contain mutated versions of Ras protein(4) The Ras family of proto-oncogenes encode21-kDa membrane-anchored GTP-bindingproteins that transduce mitogenic signals fromthe tyrosine-kinase receptors (23) Harvey virusv-Ha-Ras which is mutated in the 12th and59th amino acids is a kind of dominant acti-vated Ras (16)
Reactive oxygen species (ROS) including su-peroxide hydrogen peroxide and hydroxylradical are constantly produced by metabolicreactions in the human body (5 18) Threetypes of large-molecular-weight primary anti-
oxidant enzymes (20)mdashthe superoxide dismu-tases (SODs) catalases (CATs) and peroxi-dases [such as glutathione peroxidase (GPx)and thioredoxin peroxidase]mdashmainly controlthe intracellular level of ROS The SODs con-vert superoxide into hydrogen peroxidewhereas the CATs GPxs and thioredoxin per-oxidases convert hydrogen peroxide to harm-less water The overproduction of superoxideor hydrogen peroxide will influence activitiesof important regulatory proteins In variouscells the activation of mitogen-activated pro-tein kinase (MAPK) by the stimulation ofgrowth factors is dependent on the productionof ROS (7 14 15 30) MAPKs such as p4442MAPK p38 MAPK and c-Jun N-terminal ki-nase (JNK) MAPK are activated through phos-
Free Radical and Radiation Biology Program B180 Medical Laboratories Department of Radiology and HoldenComprehensive Cancer Center The University of Iowa Iowa City IA 52242-1181
phorylation their activation leads in turn tophosphorylation and activation of transcrip-tion factors (17 25 27) The transcription fac-tors nuclear factor-kB (NF-kB) activator pro-tein-1 (AP-1) and AP-2 (9) can bind to specificDNA sequences and induce their gene expres-sion including some of the genes involved inoxidative stress and mitogenic signaling
Generally cancer cells have low levels ofmanganese-containing SOD (MnSOD) andcopper- and zinc-containing SOD (CuZnSOD)when compared with their normal cell coun-terparts (19 20) this implies that superoxidehas a role in the cancer cell phenotype Recentstudies have connected Ras-induced mitogen-esis and superoxide production (11) increasedsuperoxide production in cancer may be a gen-eral phenomenon because tumor cells havesuch a high rate of ras gene mutation We haverecently shown in a nonmalignant but immor-tal human keratinocyte cell line that ras-trans-duced cells also had increased levels of super-oxide radicals (32) In the present work wehave extended our studies to immortal rat kid-ney epithelial cells to investigate further the ef-fects of the v-Ha-ras transfection on superoxideproduction and mitogenic signaling Using a v-Ha-ras expression vector encoding a dominantactivated Ras to transfect immortal rat kidneyepithelial cells we demonstrated that v-Ha-rasoverexpression (a) mediated an increase in cel-lular superoxide production (b) altered pri-mary antioxidant enzyme levels and (c) short-ened cell population doubling time andincreased plating efficiency In additionp44p42 and p38 MAPK were activated in Rastransduced cells as were the DNA-binding ac-tivities of AP-1 AP-2 and NF-kB Our findingssuggest that Ras might mediate mitogenic sig-naling through altering of superoxide produc-tion This work is the first to report changes inprimary antioxidant enzyme levels after rastransfection
MATERIALS AND METHODS
Cell culture
The immortal nonmalignant rat kidney ep-ithelial cell (REC) line NRK52E was a gift from
Dr Michael Robbins (Free Radical and Radia-tion Biology Program The University of Iowa)It was originally obtained from ATCC lineCRL-1571 Cells were maintained in high-glu-cose Dulbeccorsquos modified Eaglersquos medium con-taining 5 bovine calf serum 2 mM L-gluta-mine and 01 mM nonessential amino acids at37degC in 5 CO2 Cells were fed three times aweek Cells were given to us at passage 6 afterprocurement from ATCC After transfectionand selection as described below the cells wereat passage 10 The cells used in our studies werebetween 10th and 30th passages
v-Ha-ras cDNA transfer
The v-Ha-ras encoding pZIPNeoSV(X)1 plas-mid was used as the vector to generate stablytransfected cell lines This plasmid was kindlyprovided by Dr Richard C Milligan (Depart-ment of Biology and Center for Cancer Re-search Massachusetts Institute of TechnologyCambridge MA USA) The v-Ha-ras cDNAis driven by a viral LTR promoter (3) REC cells(1 3 106) were seeded in six-well plates and al-lowed to grow for 24 h The medium was re-moved and the cells were transfected with 2mg of the v-Ha-ras-pZIPNeoSV(X)1 constructand 10 ml of Lipofectamine (Life TechnologiesGaithersburg MD USA) as vehicle per dishSelection for neomycin-resistant clones wasperformed with G418 (400 mgml) and indi-vidual colonies were obtained G418 was re-moved 3 days before an experiment The clonedesignated as Neo was also transfected with v-Ha-ras-pZIPNeoSV(X)1 but did not expressv-Ha-ras
Western blotting analysis
Cells were harvested and lysed by sonica-tion and then proteins (concentrations quanti-fied using Bio-Rad protein assay kit) separatedby 125 sodium dodecyl sulfatendashpolyacry-lamide gel electrophoresis (SDS-PAGE) Theproteins were electrotransferred to nitrocellu-lose sheets After blocking in 5 milk for 30 minthe sheets were washed and treated with anti-body to pan-Ha-Ras (purchased from OncogeneScience Cambridge MA USA 11000) anti-sera to MnSOD (11000) CuZnSOD (11000)CAT (11000) antibody to phosphorylated-
YANG ET AL698
p44p42 MAPK (11000) phosphorylated-p38 MAPK (11000) or phosphorylated-JNK(11000) (New England Biolabs Inc BeverlyMA USA) for 30 min Polyclonal rabbit anti-bodies to human MnSOD CuZnSOD and CATwere prepared and characterized in our labo-ratory (21) The blot was incubated with horse-radish peroxidase-conjugated goat anti-rabbitor anti-mouse IgG (Sigma Chemical Co StLouis MO USA 110000) for 1 h at roomtemperature The washed blot was then treatedwith enhanced chemiluminescence westernblot detection solution (Amersham Life Sci-ence Buckinghamshire UK) and exposed tox-ray film
Northern blotting analysis
Total RNA was isolated by the acid guani-dinium thiocyanatendashphenolndashchloroform ex-traction method and then quantified by mi-crospectrometry RNA (18 mg) was separatedon a 15 agarose formaldehyde gel RNA wastransferred to a nylon membrane the mem-brane was then preincubated with hybridiza-tion buffer [50 formamide 53 salinendashsodiumcitrate buffer (SSC) 10 dextran sulfate 13Denhardtrsquos buffer 1 SDS and 150 mgml de-natured salmon sperm DNA] and hybridizedto a 32P-labeled probe made from v-Ha-rascDNA Following hybridization the mem-brane was washed in 23 SSC 05 SDS solu-tion twice for 15 min each at room temperatureand then washed in 013 SSC 05 SDS solu-tion twice for 15 min each at 65degC The mem-brane was wrapped in plastic wrap and ex-posed to x-ray film (Kodak Rochester NYUSA) at 280degC overnight 18S RNA was usedto normalize the amount of RNA loading
SOD-inhibitable cytochrome c assay forsuperoxide production
The production of superoxide was measuredby its reduction of cytochrome c (2) Cells werecultured in six-well plates The medium wasthen removed and cells washed twice inKrebsndashRinger buffer Cytochrome c (100 mM)in KrebsndashRinger buffer with CAT (200 Uml)was added to the wells samples were removedafter 1 h of incubation at 37degC in 5 CO2 Thespectrum of each sample was then taken be-
tween 400 and 600 nm Superoxide productionwas determined by measuring the peak heightat 550 nm over the baseline between 530 and560 nm Cytochrome c can be reduced by thesuperoxide radical anion (O2
Ccedil 2) from cyt c(Fe31) to cyt c (Fe21) with a rate constant k 515 3 105M21 s21 (pH 7ndash93) and extinction co-efficients of cyt c (Fe21)em550 5 299 3 104 M21
cm21 and cyt c (Fe31) em550 5 089 3 104 M21
cm21 (6)
SOD CAT and GPx activity gel assays
SOD CAT and GPx activities were measuredusing native gel activity stains An 8 poly-acrylamide running gel and 5 stacking gelwere used to separate the proteins For the SODactivity stain (1) the gel was stained with 243mM nitro blue tetrazolium solution for 20 minand then 28 mM riboflavin with 28 mMN N N 9 N 9 -te tramethylethylenediamine(TEMED) for 15 min The gel was placed in dis-tilled water and illuminated under a bright flu-orescent light Achromatic bands indicated thepresence of SOD For the CAT activity stain (28)the gel was first rinsed three times with distilledwater then incubated in 0003 hydrogen per-oxide for 10 min The gel was then stained with2 ferric chloride and 2 potassium ferri-cyanide when achromatic bands began to formthe stain was poured off and the gels were rinsedextensively with distilled water Achromaticbands demonstrated the presence of CAT activ-ity A similar procedure was used for GPx ac-tivity (28) Gels were first soaked in threechanges of reduced glutathione for a total of 45minutes then soaked in 0008 cumene hy-droperoxide plus reduced glutathione for 10min The gels were rinsed twice with distilledwater and then stained with ferric chloride andpotassium ferricyanide as described above
Plating efficiency
Cells were plated in triplicate into 60-mmdishes in complete medium The number ofcells plated was 100 or 500 The dishes weremaintained in the incubator for 14 days to al-low colony formation Colonies were then fixedand stained with 01 crystal violet and 21citric acid and those colonies containing 50cells were scored
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 699
Cell population doubling time assay
Cells (2 3 104) were plated in 15 ml of com-plete medium in 24-well plates the mediumwas changed every 4 days Cells weretrypsinized and then counted on alternate daysfor 2 weeks using a hemocytometer Cell pop-ulation doubling time (DT) in hours was de-termined using the following equation (13)
DT (hours) 5 0693 (t 2 t0)ln (NtN0)
where t0 is time at which exponential growthbegan t is time in hours Nt is cell number attime t and N0 is initial cell number
Gel mobility shift assay
Nuclear protein was extracted by the fol-lowing procedure Cells were collected when80 confluent and rinsed with phosphate-buffered saline buffer Cells were then har-vested by scraping in ice-cold buffer A (10 mMHEPES 15 mM MgCl2 10 mM KCl) and incu-bated on ice for 20 min The cells were lysedwith a Dounce homogenizer (Kontes VinelandNJ USA) and the nuclei were pelleted bycentrifuging twice at 1400g for 30 s at 4degC withsubsequent removal of supernatant The nu-clear pellets were resuspended in ice-coldbuffer C (20 mM HEPES 25 glycerol 042 MNaCl 15 mM MgCl2 02 mM EDTA 05 mMphenylmethylsulfonyl fluoride) and placed onice for 15 min The suspensions were cen-trifuged (14000g for 10ndash15 min) and the su-pernatants were removed and diluted with ice-cold buffer D (20 mM HEPES 20 glycerol 01M KCl 02 mM EDTA 05 mM phenylmethyl-sulfonyl fluoride) The protein concentrationswere determined and nuclear protein extractswere stored at 280degC
DNA binding reactions were carried out with 15 mg of extracted nuclear protein anddouble-stranded synthetic oligodeoxynucleo-tide encompassing an AP-1 consensus sequence59-agcttgTGACTCAccgtag-39 AP-2 consensussequence 59-agctcaaGCCCGCGGGCtctctag-39or NF-kB consensus sequence 59-agctga-GGGGACTTTCCctag-39 Probes were 32P-la-beled using Klenow DNA polymerase and [a-32P]dCTP Nuclear protein extracts (15 mg)were incubated with 32P-labeled DNA probes
in the presence of 1 mg of poly(dIdC) (Phar-macia Biotech Inc Piscataway NJ USA) and13 gel shift buffer (10 mM Tris pH 75 50 mMNaCl 1 mM MgCl2 05 mM EDTA 05 mMdithiothreitol 4 glycerol) at room tempera-ture for 20 min The bound DNAndashprotein com-plexes were separated from free probe by gelelectrophoresis in 5 native polyacrylamidegels in 13 TBE (90 mM Tris-borate 2 mMEDTA) Electrophoresis was conducted at aconstant current of 20 mA for 1 h The gel waswrapped in a plastic wrap and exposed to x-ray film overnight at 280degC AP-1 DNA-bind-ing specificity was determined by competitionexperiments Nuclear protein extracts (15 mg)were preincubated with 1 mg of poly(dIdC) 13gel shift buffer and a 10- or 100-fold excess ofunlabeled AP-1 AP-2 or NF-kB oligonu-cleotides for 15 min at room temperature be-fore the addition of the labeled probe For su-pershift assays nuclear protein extracts (15 mg)were preincubated with 2 mg of anti-c-JunAP-1 anti-AP-2a or anti-p50 (NF-kB) antibodies(Santa Cruz Biotechnology Santa Cruz CAUSA) for 30 min at room temperature andthen incubated with 32P-labeled probes
Statistical analysis
Statistical analysis was performed using SY-STAT (Systat Evanston IL USA) A singlefactor analysis of variance followed by a post-hoc Tukey test was used to determine statisti-cal differences between means The null hy-pothesis was rejected at the 005 level ofsignificance All means were calculated fromthree experiments and error bars representstandard deviations (SD) All western blot andactivity gel assays were repeated at least twice
RESULTS
v-Ha-ras protein was overexpressed and mediatedsuperoxide production
REC cells were transfected with v-Ha-rasthrough the retrovirus plasmid pZIPNeo-SV(X)1 construct Clones were selected and theexpression of v-Ha-Ras protein was detectedby western blotting The clones with differentincreased levels of v-Ha-Ras immunoreactive
YANG ET AL700
protein (W13 U12 and U23) were used for fur-ther experiments (Fig 1A) The northern blot(Fig 1B) demonstrated two new mRNA bands(47 and 6 kb) in W13 U12 and U23 but not inREC or Neo the new bands were therefore gen-erated from the v-Ha-ras expression vector Be-cause of what has been observed earlier inother cells the likely explanations for the twonew bands are the proviral full-length mRNAand a spliced variant The intensity of the newbands suggested that v-Ha-ras was efficientlytranscribed in W13 U12 and U23 cells The 1-kb mRNA from the endogenous c-Ha-ras genewas also found its intensity was increased inW13 U12 and U23 clones There are at leasttwo possibilities for this observation besidesthe RNA loading One is that c-Ha-ras oncogeneis up regulated by v-Ha-ras overexpression theother is that some v-Ha-rasmRNAs decomposeto a 1-kb size These data suggest that the Rasprotein comes from the exogenous plasmid butdoes not exclude the possibility that some Rasprotein may come from the endogenous gene
Recent evidence has demonstrated that Rastransformed cells can stimulate cellular super-oxide production (11 22 29 32) To test if v-Ha-ras transduced REC cells produce moresuperoxide we used Fe(III) cytochrome c tomonitor the amount of superoxide productionfrom the cells Figure 2A shows that v-Ha-ras
transduced cells U12 and U23 generated largeamounts of superoxide up to fivefold morethan REC or Neo cells The addition of exoge-nous CuZnSOD protein totally inhibited the in-creased superoxide levels seen in U23 cells (Fig2B) which demonstrated that the 550-nm peakwas due to superoxide production from thecells
To test whether v-Ha-Ras protein mediatedsuperoxide production we treated U23 cellswith either lovastatin or a-hydroxyfarnesyl-phosphonic acid (a-PA) inhibitors of Ras pro-tein farnesylation (26) superoxide productionwas decreased 50 by each of these inhibi-tors (Fig 2B) and statistical analysis showedthat the difference was significant (p 005)This result demonstrated that Ras mediated thesuperoxide production from the v-Ha-ras trans-duced cells We did not see complete inhibi-tion there may be certain functional Ras pro-teins formed in spite of the drug treatment
We then studied the source of superoxideproduction from the v-Ha-ras transfected cellsNeutrophils after stimulation are well knownto produce superoxide through an NADPH ox-idase system (8) there is evidence showing thatother types of cells including epithelial cellsalso have an NADPH oxidase system in theirplasma membrane (22 29 31 32) Mitochon-dria are also an important source of superox-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 701
FIG 1 v-Ha-Ras was overexpressed in v-Ha-ras transduced REC cells (A) Western blotting analysis Cell lysates(100 mg) were run on 125 SDS-PAGE The proteins were electrotransferred to nitrocellulose sheets and treated withmouse monoclonal antibody to pan-Ras (B) Northern blotting analysis Total RNA (18 mg) was fractionated on 15agarose gel transferred onto a nylon membrane and hybridized with 32P-labeled probe made from human v-Ha-rascDNA 18S RNA was used to normalize the RNA loading amount
ide it is estimated in respiring cells that at least1ndash3 of total respiration is used to generate su-peroxide and hydrogen peroxide in elaborateelectron-transfer chains for the reduction ofmolecular oxygen to water (18) When U23 cellswere treated with diphenylene iodonium(DPI) an inhibitor of NADPH oxidase 100 mM
for 2 h before the cytochrome c assay super-oxide production was decreased to 50 of theoriginal amount and this difference was sta-tistically significant (Fig 2B p 005) How-ever when U23 cells were treated with 50 mMrotenone an inhibitor of mitochondrial respi-ratory electron-transfer chains for 2 h before
YANG ET AL702
FIG 2 Ras mediated superoxide production from the SOD-inhibitable cytochrome c assay (A) Cells were washedthen incubated with 100 mM cytochrome c in KrebsndashRinger buffer at 37degC 5 CO2 for 1 h Samples were scannedfrom 400 nm to 600 nm the 550-nm peak height was measured and adjusted for baseline and then the amount ofsuperoxide was determined Means were calculated from three experiments and error bars represent SD Signifi-cantly different from REC or Neo at the p 005 level Data indicate that Ras transduced cells had increased levelsof superoxide radical (B) U23 cells were further treated with rotenone (50 mM) DPI (100 mM) 2 h lovastatin (50 mM)48 h or a-PA (4 mM) 48 h before measurements or CuZnSOD protein (200 Uml final) was added into the cytochromec solution during the measurement Cell number was counted after the assay data were calculated and expressed asthe rate of superoxide production per hour (nmol106 cells 3 h) Means were calculated from three experiments anderror bars represent SD Significantly different from U23 at the p 005 level The data show that CuZnSOD pro-tein DPI a-PA and lovastatin all inhibited superoxide levels
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

phorylation their activation leads in turn tophosphorylation and activation of transcrip-tion factors (17 25 27) The transcription fac-tors nuclear factor-kB (NF-kB) activator pro-tein-1 (AP-1) and AP-2 (9) can bind to specificDNA sequences and induce their gene expres-sion including some of the genes involved inoxidative stress and mitogenic signaling
Generally cancer cells have low levels ofmanganese-containing SOD (MnSOD) andcopper- and zinc-containing SOD (CuZnSOD)when compared with their normal cell coun-terparts (19 20) this implies that superoxidehas a role in the cancer cell phenotype Recentstudies have connected Ras-induced mitogen-esis and superoxide production (11) increasedsuperoxide production in cancer may be a gen-eral phenomenon because tumor cells havesuch a high rate of ras gene mutation We haverecently shown in a nonmalignant but immor-tal human keratinocyte cell line that ras-trans-duced cells also had increased levels of super-oxide radicals (32) In the present work wehave extended our studies to immortal rat kid-ney epithelial cells to investigate further the ef-fects of the v-Ha-ras transfection on superoxideproduction and mitogenic signaling Using a v-Ha-ras expression vector encoding a dominantactivated Ras to transfect immortal rat kidneyepithelial cells we demonstrated that v-Ha-rasoverexpression (a) mediated an increase in cel-lular superoxide production (b) altered pri-mary antioxidant enzyme levels and (c) short-ened cell population doubling time andincreased plating efficiency In additionp44p42 and p38 MAPK were activated in Rastransduced cells as were the DNA-binding ac-tivities of AP-1 AP-2 and NF-kB Our findingssuggest that Ras might mediate mitogenic sig-naling through altering of superoxide produc-tion This work is the first to report changes inprimary antioxidant enzyme levels after rastransfection
MATERIALS AND METHODS
Cell culture
The immortal nonmalignant rat kidney ep-ithelial cell (REC) line NRK52E was a gift from
Dr Michael Robbins (Free Radical and Radia-tion Biology Program The University of Iowa)It was originally obtained from ATCC lineCRL-1571 Cells were maintained in high-glu-cose Dulbeccorsquos modified Eaglersquos medium con-taining 5 bovine calf serum 2 mM L-gluta-mine and 01 mM nonessential amino acids at37degC in 5 CO2 Cells were fed three times aweek Cells were given to us at passage 6 afterprocurement from ATCC After transfectionand selection as described below the cells wereat passage 10 The cells used in our studies werebetween 10th and 30th passages
v-Ha-ras cDNA transfer
The v-Ha-ras encoding pZIPNeoSV(X)1 plas-mid was used as the vector to generate stablytransfected cell lines This plasmid was kindlyprovided by Dr Richard C Milligan (Depart-ment of Biology and Center for Cancer Re-search Massachusetts Institute of TechnologyCambridge MA USA) The v-Ha-ras cDNAis driven by a viral LTR promoter (3) REC cells(1 3 106) were seeded in six-well plates and al-lowed to grow for 24 h The medium was re-moved and the cells were transfected with 2mg of the v-Ha-ras-pZIPNeoSV(X)1 constructand 10 ml of Lipofectamine (Life TechnologiesGaithersburg MD USA) as vehicle per dishSelection for neomycin-resistant clones wasperformed with G418 (400 mgml) and indi-vidual colonies were obtained G418 was re-moved 3 days before an experiment The clonedesignated as Neo was also transfected with v-Ha-ras-pZIPNeoSV(X)1 but did not expressv-Ha-ras
Western blotting analysis
Cells were harvested and lysed by sonica-tion and then proteins (concentrations quanti-fied using Bio-Rad protein assay kit) separatedby 125 sodium dodecyl sulfatendashpolyacry-lamide gel electrophoresis (SDS-PAGE) Theproteins were electrotransferred to nitrocellu-lose sheets After blocking in 5 milk for 30 minthe sheets were washed and treated with anti-body to pan-Ha-Ras (purchased from OncogeneScience Cambridge MA USA 11000) anti-sera to MnSOD (11000) CuZnSOD (11000)CAT (11000) antibody to phosphorylated-
YANG ET AL698
p44p42 MAPK (11000) phosphorylated-p38 MAPK (11000) or phosphorylated-JNK(11000) (New England Biolabs Inc BeverlyMA USA) for 30 min Polyclonal rabbit anti-bodies to human MnSOD CuZnSOD and CATwere prepared and characterized in our labo-ratory (21) The blot was incubated with horse-radish peroxidase-conjugated goat anti-rabbitor anti-mouse IgG (Sigma Chemical Co StLouis MO USA 110000) for 1 h at roomtemperature The washed blot was then treatedwith enhanced chemiluminescence westernblot detection solution (Amersham Life Sci-ence Buckinghamshire UK) and exposed tox-ray film
Northern blotting analysis
Total RNA was isolated by the acid guani-dinium thiocyanatendashphenolndashchloroform ex-traction method and then quantified by mi-crospectrometry RNA (18 mg) was separatedon a 15 agarose formaldehyde gel RNA wastransferred to a nylon membrane the mem-brane was then preincubated with hybridiza-tion buffer [50 formamide 53 salinendashsodiumcitrate buffer (SSC) 10 dextran sulfate 13Denhardtrsquos buffer 1 SDS and 150 mgml de-natured salmon sperm DNA] and hybridizedto a 32P-labeled probe made from v-Ha-rascDNA Following hybridization the mem-brane was washed in 23 SSC 05 SDS solu-tion twice for 15 min each at room temperatureand then washed in 013 SSC 05 SDS solu-tion twice for 15 min each at 65degC The mem-brane was wrapped in plastic wrap and ex-posed to x-ray film (Kodak Rochester NYUSA) at 280degC overnight 18S RNA was usedto normalize the amount of RNA loading
SOD-inhibitable cytochrome c assay forsuperoxide production
The production of superoxide was measuredby its reduction of cytochrome c (2) Cells werecultured in six-well plates The medium wasthen removed and cells washed twice inKrebsndashRinger buffer Cytochrome c (100 mM)in KrebsndashRinger buffer with CAT (200 Uml)was added to the wells samples were removedafter 1 h of incubation at 37degC in 5 CO2 Thespectrum of each sample was then taken be-
tween 400 and 600 nm Superoxide productionwas determined by measuring the peak heightat 550 nm over the baseline between 530 and560 nm Cytochrome c can be reduced by thesuperoxide radical anion (O2
Ccedil 2) from cyt c(Fe31) to cyt c (Fe21) with a rate constant k 515 3 105M21 s21 (pH 7ndash93) and extinction co-efficients of cyt c (Fe21)em550 5 299 3 104 M21
cm21 and cyt c (Fe31) em550 5 089 3 104 M21
cm21 (6)
SOD CAT and GPx activity gel assays
SOD CAT and GPx activities were measuredusing native gel activity stains An 8 poly-acrylamide running gel and 5 stacking gelwere used to separate the proteins For the SODactivity stain (1) the gel was stained with 243mM nitro blue tetrazolium solution for 20 minand then 28 mM riboflavin with 28 mMN N N 9 N 9 -te tramethylethylenediamine(TEMED) for 15 min The gel was placed in dis-tilled water and illuminated under a bright flu-orescent light Achromatic bands indicated thepresence of SOD For the CAT activity stain (28)the gel was first rinsed three times with distilledwater then incubated in 0003 hydrogen per-oxide for 10 min The gel was then stained with2 ferric chloride and 2 potassium ferri-cyanide when achromatic bands began to formthe stain was poured off and the gels were rinsedextensively with distilled water Achromaticbands demonstrated the presence of CAT activ-ity A similar procedure was used for GPx ac-tivity (28) Gels were first soaked in threechanges of reduced glutathione for a total of 45minutes then soaked in 0008 cumene hy-droperoxide plus reduced glutathione for 10min The gels were rinsed twice with distilledwater and then stained with ferric chloride andpotassium ferricyanide as described above
Plating efficiency
Cells were plated in triplicate into 60-mmdishes in complete medium The number ofcells plated was 100 or 500 The dishes weremaintained in the incubator for 14 days to al-low colony formation Colonies were then fixedand stained with 01 crystal violet and 21citric acid and those colonies containing 50cells were scored
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 699
Cell population doubling time assay
Cells (2 3 104) were plated in 15 ml of com-plete medium in 24-well plates the mediumwas changed every 4 days Cells weretrypsinized and then counted on alternate daysfor 2 weeks using a hemocytometer Cell pop-ulation doubling time (DT) in hours was de-termined using the following equation (13)
DT (hours) 5 0693 (t 2 t0)ln (NtN0)
where t0 is time at which exponential growthbegan t is time in hours Nt is cell number attime t and N0 is initial cell number
Gel mobility shift assay
Nuclear protein was extracted by the fol-lowing procedure Cells were collected when80 confluent and rinsed with phosphate-buffered saline buffer Cells were then har-vested by scraping in ice-cold buffer A (10 mMHEPES 15 mM MgCl2 10 mM KCl) and incu-bated on ice for 20 min The cells were lysedwith a Dounce homogenizer (Kontes VinelandNJ USA) and the nuclei were pelleted bycentrifuging twice at 1400g for 30 s at 4degC withsubsequent removal of supernatant The nu-clear pellets were resuspended in ice-coldbuffer C (20 mM HEPES 25 glycerol 042 MNaCl 15 mM MgCl2 02 mM EDTA 05 mMphenylmethylsulfonyl fluoride) and placed onice for 15 min The suspensions were cen-trifuged (14000g for 10ndash15 min) and the su-pernatants were removed and diluted with ice-cold buffer D (20 mM HEPES 20 glycerol 01M KCl 02 mM EDTA 05 mM phenylmethyl-sulfonyl fluoride) The protein concentrationswere determined and nuclear protein extractswere stored at 280degC
DNA binding reactions were carried out with 15 mg of extracted nuclear protein anddouble-stranded synthetic oligodeoxynucleo-tide encompassing an AP-1 consensus sequence59-agcttgTGACTCAccgtag-39 AP-2 consensussequence 59-agctcaaGCCCGCGGGCtctctag-39or NF-kB consensus sequence 59-agctga-GGGGACTTTCCctag-39 Probes were 32P-la-beled using Klenow DNA polymerase and [a-32P]dCTP Nuclear protein extracts (15 mg)were incubated with 32P-labeled DNA probes
in the presence of 1 mg of poly(dIdC) (Phar-macia Biotech Inc Piscataway NJ USA) and13 gel shift buffer (10 mM Tris pH 75 50 mMNaCl 1 mM MgCl2 05 mM EDTA 05 mMdithiothreitol 4 glycerol) at room tempera-ture for 20 min The bound DNAndashprotein com-plexes were separated from free probe by gelelectrophoresis in 5 native polyacrylamidegels in 13 TBE (90 mM Tris-borate 2 mMEDTA) Electrophoresis was conducted at aconstant current of 20 mA for 1 h The gel waswrapped in a plastic wrap and exposed to x-ray film overnight at 280degC AP-1 DNA-bind-ing specificity was determined by competitionexperiments Nuclear protein extracts (15 mg)were preincubated with 1 mg of poly(dIdC) 13gel shift buffer and a 10- or 100-fold excess ofunlabeled AP-1 AP-2 or NF-kB oligonu-cleotides for 15 min at room temperature be-fore the addition of the labeled probe For su-pershift assays nuclear protein extracts (15 mg)were preincubated with 2 mg of anti-c-JunAP-1 anti-AP-2a or anti-p50 (NF-kB) antibodies(Santa Cruz Biotechnology Santa Cruz CAUSA) for 30 min at room temperature andthen incubated with 32P-labeled probes
Statistical analysis
Statistical analysis was performed using SY-STAT (Systat Evanston IL USA) A singlefactor analysis of variance followed by a post-hoc Tukey test was used to determine statisti-cal differences between means The null hy-pothesis was rejected at the 005 level ofsignificance All means were calculated fromthree experiments and error bars representstandard deviations (SD) All western blot andactivity gel assays were repeated at least twice
RESULTS
v-Ha-ras protein was overexpressed and mediatedsuperoxide production
REC cells were transfected with v-Ha-rasthrough the retrovirus plasmid pZIPNeo-SV(X)1 construct Clones were selected and theexpression of v-Ha-Ras protein was detectedby western blotting The clones with differentincreased levels of v-Ha-Ras immunoreactive
YANG ET AL700
protein (W13 U12 and U23) were used for fur-ther experiments (Fig 1A) The northern blot(Fig 1B) demonstrated two new mRNA bands(47 and 6 kb) in W13 U12 and U23 but not inREC or Neo the new bands were therefore gen-erated from the v-Ha-ras expression vector Be-cause of what has been observed earlier inother cells the likely explanations for the twonew bands are the proviral full-length mRNAand a spliced variant The intensity of the newbands suggested that v-Ha-ras was efficientlytranscribed in W13 U12 and U23 cells The 1-kb mRNA from the endogenous c-Ha-ras genewas also found its intensity was increased inW13 U12 and U23 clones There are at leasttwo possibilities for this observation besidesthe RNA loading One is that c-Ha-ras oncogeneis up regulated by v-Ha-ras overexpression theother is that some v-Ha-rasmRNAs decomposeto a 1-kb size These data suggest that the Rasprotein comes from the exogenous plasmid butdoes not exclude the possibility that some Rasprotein may come from the endogenous gene
Recent evidence has demonstrated that Rastransformed cells can stimulate cellular super-oxide production (11 22 29 32) To test if v-Ha-ras transduced REC cells produce moresuperoxide we used Fe(III) cytochrome c tomonitor the amount of superoxide productionfrom the cells Figure 2A shows that v-Ha-ras
transduced cells U12 and U23 generated largeamounts of superoxide up to fivefold morethan REC or Neo cells The addition of exoge-nous CuZnSOD protein totally inhibited the in-creased superoxide levels seen in U23 cells (Fig2B) which demonstrated that the 550-nm peakwas due to superoxide production from thecells
To test whether v-Ha-Ras protein mediatedsuperoxide production we treated U23 cellswith either lovastatin or a-hydroxyfarnesyl-phosphonic acid (a-PA) inhibitors of Ras pro-tein farnesylation (26) superoxide productionwas decreased 50 by each of these inhibi-tors (Fig 2B) and statistical analysis showedthat the difference was significant (p 005)This result demonstrated that Ras mediated thesuperoxide production from the v-Ha-ras trans-duced cells We did not see complete inhibi-tion there may be certain functional Ras pro-teins formed in spite of the drug treatment
We then studied the source of superoxideproduction from the v-Ha-ras transfected cellsNeutrophils after stimulation are well knownto produce superoxide through an NADPH ox-idase system (8) there is evidence showing thatother types of cells including epithelial cellsalso have an NADPH oxidase system in theirplasma membrane (22 29 31 32) Mitochon-dria are also an important source of superox-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 701
FIG 1 v-Ha-Ras was overexpressed in v-Ha-ras transduced REC cells (A) Western blotting analysis Cell lysates(100 mg) were run on 125 SDS-PAGE The proteins were electrotransferred to nitrocellulose sheets and treated withmouse monoclonal antibody to pan-Ras (B) Northern blotting analysis Total RNA (18 mg) was fractionated on 15agarose gel transferred onto a nylon membrane and hybridized with 32P-labeled probe made from human v-Ha-rascDNA 18S RNA was used to normalize the RNA loading amount
ide it is estimated in respiring cells that at least1ndash3 of total respiration is used to generate su-peroxide and hydrogen peroxide in elaborateelectron-transfer chains for the reduction ofmolecular oxygen to water (18) When U23 cellswere treated with diphenylene iodonium(DPI) an inhibitor of NADPH oxidase 100 mM
for 2 h before the cytochrome c assay super-oxide production was decreased to 50 of theoriginal amount and this difference was sta-tistically significant (Fig 2B p 005) How-ever when U23 cells were treated with 50 mMrotenone an inhibitor of mitochondrial respi-ratory electron-transfer chains for 2 h before
YANG ET AL702
FIG 2 Ras mediated superoxide production from the SOD-inhibitable cytochrome c assay (A) Cells were washedthen incubated with 100 mM cytochrome c in KrebsndashRinger buffer at 37degC 5 CO2 for 1 h Samples were scannedfrom 400 nm to 600 nm the 550-nm peak height was measured and adjusted for baseline and then the amount ofsuperoxide was determined Means were calculated from three experiments and error bars represent SD Signifi-cantly different from REC or Neo at the p 005 level Data indicate that Ras transduced cells had increased levelsof superoxide radical (B) U23 cells were further treated with rotenone (50 mM) DPI (100 mM) 2 h lovastatin (50 mM)48 h or a-PA (4 mM) 48 h before measurements or CuZnSOD protein (200 Uml final) was added into the cytochromec solution during the measurement Cell number was counted after the assay data were calculated and expressed asthe rate of superoxide production per hour (nmol106 cells 3 h) Means were calculated from three experiments anderror bars represent SD Significantly different from U23 at the p 005 level The data show that CuZnSOD pro-tein DPI a-PA and lovastatin all inhibited superoxide levels
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

p44p42 MAPK (11000) phosphorylated-p38 MAPK (11000) or phosphorylated-JNK(11000) (New England Biolabs Inc BeverlyMA USA) for 30 min Polyclonal rabbit anti-bodies to human MnSOD CuZnSOD and CATwere prepared and characterized in our labo-ratory (21) The blot was incubated with horse-radish peroxidase-conjugated goat anti-rabbitor anti-mouse IgG (Sigma Chemical Co StLouis MO USA 110000) for 1 h at roomtemperature The washed blot was then treatedwith enhanced chemiluminescence westernblot detection solution (Amersham Life Sci-ence Buckinghamshire UK) and exposed tox-ray film
Northern blotting analysis
Total RNA was isolated by the acid guani-dinium thiocyanatendashphenolndashchloroform ex-traction method and then quantified by mi-crospectrometry RNA (18 mg) was separatedon a 15 agarose formaldehyde gel RNA wastransferred to a nylon membrane the mem-brane was then preincubated with hybridiza-tion buffer [50 formamide 53 salinendashsodiumcitrate buffer (SSC) 10 dextran sulfate 13Denhardtrsquos buffer 1 SDS and 150 mgml de-natured salmon sperm DNA] and hybridizedto a 32P-labeled probe made from v-Ha-rascDNA Following hybridization the mem-brane was washed in 23 SSC 05 SDS solu-tion twice for 15 min each at room temperatureand then washed in 013 SSC 05 SDS solu-tion twice for 15 min each at 65degC The mem-brane was wrapped in plastic wrap and ex-posed to x-ray film (Kodak Rochester NYUSA) at 280degC overnight 18S RNA was usedto normalize the amount of RNA loading
SOD-inhibitable cytochrome c assay forsuperoxide production
The production of superoxide was measuredby its reduction of cytochrome c (2) Cells werecultured in six-well plates The medium wasthen removed and cells washed twice inKrebsndashRinger buffer Cytochrome c (100 mM)in KrebsndashRinger buffer with CAT (200 Uml)was added to the wells samples were removedafter 1 h of incubation at 37degC in 5 CO2 Thespectrum of each sample was then taken be-
tween 400 and 600 nm Superoxide productionwas determined by measuring the peak heightat 550 nm over the baseline between 530 and560 nm Cytochrome c can be reduced by thesuperoxide radical anion (O2
Ccedil 2) from cyt c(Fe31) to cyt c (Fe21) with a rate constant k 515 3 105M21 s21 (pH 7ndash93) and extinction co-efficients of cyt c (Fe21)em550 5 299 3 104 M21
cm21 and cyt c (Fe31) em550 5 089 3 104 M21
cm21 (6)
SOD CAT and GPx activity gel assays
SOD CAT and GPx activities were measuredusing native gel activity stains An 8 poly-acrylamide running gel and 5 stacking gelwere used to separate the proteins For the SODactivity stain (1) the gel was stained with 243mM nitro blue tetrazolium solution for 20 minand then 28 mM riboflavin with 28 mMN N N 9 N 9 -te tramethylethylenediamine(TEMED) for 15 min The gel was placed in dis-tilled water and illuminated under a bright flu-orescent light Achromatic bands indicated thepresence of SOD For the CAT activity stain (28)the gel was first rinsed three times with distilledwater then incubated in 0003 hydrogen per-oxide for 10 min The gel was then stained with2 ferric chloride and 2 potassium ferri-cyanide when achromatic bands began to formthe stain was poured off and the gels were rinsedextensively with distilled water Achromaticbands demonstrated the presence of CAT activ-ity A similar procedure was used for GPx ac-tivity (28) Gels were first soaked in threechanges of reduced glutathione for a total of 45minutes then soaked in 0008 cumene hy-droperoxide plus reduced glutathione for 10min The gels were rinsed twice with distilledwater and then stained with ferric chloride andpotassium ferricyanide as described above
Plating efficiency
Cells were plated in triplicate into 60-mmdishes in complete medium The number ofcells plated was 100 or 500 The dishes weremaintained in the incubator for 14 days to al-low colony formation Colonies were then fixedand stained with 01 crystal violet and 21citric acid and those colonies containing 50cells were scored
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 699
Cell population doubling time assay
Cells (2 3 104) were plated in 15 ml of com-plete medium in 24-well plates the mediumwas changed every 4 days Cells weretrypsinized and then counted on alternate daysfor 2 weeks using a hemocytometer Cell pop-ulation doubling time (DT) in hours was de-termined using the following equation (13)
DT (hours) 5 0693 (t 2 t0)ln (NtN0)
where t0 is time at which exponential growthbegan t is time in hours Nt is cell number attime t and N0 is initial cell number
Gel mobility shift assay
Nuclear protein was extracted by the fol-lowing procedure Cells were collected when80 confluent and rinsed with phosphate-buffered saline buffer Cells were then har-vested by scraping in ice-cold buffer A (10 mMHEPES 15 mM MgCl2 10 mM KCl) and incu-bated on ice for 20 min The cells were lysedwith a Dounce homogenizer (Kontes VinelandNJ USA) and the nuclei were pelleted bycentrifuging twice at 1400g for 30 s at 4degC withsubsequent removal of supernatant The nu-clear pellets were resuspended in ice-coldbuffer C (20 mM HEPES 25 glycerol 042 MNaCl 15 mM MgCl2 02 mM EDTA 05 mMphenylmethylsulfonyl fluoride) and placed onice for 15 min The suspensions were cen-trifuged (14000g for 10ndash15 min) and the su-pernatants were removed and diluted with ice-cold buffer D (20 mM HEPES 20 glycerol 01M KCl 02 mM EDTA 05 mM phenylmethyl-sulfonyl fluoride) The protein concentrationswere determined and nuclear protein extractswere stored at 280degC
DNA binding reactions were carried out with 15 mg of extracted nuclear protein anddouble-stranded synthetic oligodeoxynucleo-tide encompassing an AP-1 consensus sequence59-agcttgTGACTCAccgtag-39 AP-2 consensussequence 59-agctcaaGCCCGCGGGCtctctag-39or NF-kB consensus sequence 59-agctga-GGGGACTTTCCctag-39 Probes were 32P-la-beled using Klenow DNA polymerase and [a-32P]dCTP Nuclear protein extracts (15 mg)were incubated with 32P-labeled DNA probes
in the presence of 1 mg of poly(dIdC) (Phar-macia Biotech Inc Piscataway NJ USA) and13 gel shift buffer (10 mM Tris pH 75 50 mMNaCl 1 mM MgCl2 05 mM EDTA 05 mMdithiothreitol 4 glycerol) at room tempera-ture for 20 min The bound DNAndashprotein com-plexes were separated from free probe by gelelectrophoresis in 5 native polyacrylamidegels in 13 TBE (90 mM Tris-borate 2 mMEDTA) Electrophoresis was conducted at aconstant current of 20 mA for 1 h The gel waswrapped in a plastic wrap and exposed to x-ray film overnight at 280degC AP-1 DNA-bind-ing specificity was determined by competitionexperiments Nuclear protein extracts (15 mg)were preincubated with 1 mg of poly(dIdC) 13gel shift buffer and a 10- or 100-fold excess ofunlabeled AP-1 AP-2 or NF-kB oligonu-cleotides for 15 min at room temperature be-fore the addition of the labeled probe For su-pershift assays nuclear protein extracts (15 mg)were preincubated with 2 mg of anti-c-JunAP-1 anti-AP-2a or anti-p50 (NF-kB) antibodies(Santa Cruz Biotechnology Santa Cruz CAUSA) for 30 min at room temperature andthen incubated with 32P-labeled probes
Statistical analysis
Statistical analysis was performed using SY-STAT (Systat Evanston IL USA) A singlefactor analysis of variance followed by a post-hoc Tukey test was used to determine statisti-cal differences between means The null hy-pothesis was rejected at the 005 level ofsignificance All means were calculated fromthree experiments and error bars representstandard deviations (SD) All western blot andactivity gel assays were repeated at least twice
RESULTS
v-Ha-ras protein was overexpressed and mediatedsuperoxide production
REC cells were transfected with v-Ha-rasthrough the retrovirus plasmid pZIPNeo-SV(X)1 construct Clones were selected and theexpression of v-Ha-Ras protein was detectedby western blotting The clones with differentincreased levels of v-Ha-Ras immunoreactive
YANG ET AL700
protein (W13 U12 and U23) were used for fur-ther experiments (Fig 1A) The northern blot(Fig 1B) demonstrated two new mRNA bands(47 and 6 kb) in W13 U12 and U23 but not inREC or Neo the new bands were therefore gen-erated from the v-Ha-ras expression vector Be-cause of what has been observed earlier inother cells the likely explanations for the twonew bands are the proviral full-length mRNAand a spliced variant The intensity of the newbands suggested that v-Ha-ras was efficientlytranscribed in W13 U12 and U23 cells The 1-kb mRNA from the endogenous c-Ha-ras genewas also found its intensity was increased inW13 U12 and U23 clones There are at leasttwo possibilities for this observation besidesthe RNA loading One is that c-Ha-ras oncogeneis up regulated by v-Ha-ras overexpression theother is that some v-Ha-rasmRNAs decomposeto a 1-kb size These data suggest that the Rasprotein comes from the exogenous plasmid butdoes not exclude the possibility that some Rasprotein may come from the endogenous gene
Recent evidence has demonstrated that Rastransformed cells can stimulate cellular super-oxide production (11 22 29 32) To test if v-Ha-ras transduced REC cells produce moresuperoxide we used Fe(III) cytochrome c tomonitor the amount of superoxide productionfrom the cells Figure 2A shows that v-Ha-ras
transduced cells U12 and U23 generated largeamounts of superoxide up to fivefold morethan REC or Neo cells The addition of exoge-nous CuZnSOD protein totally inhibited the in-creased superoxide levels seen in U23 cells (Fig2B) which demonstrated that the 550-nm peakwas due to superoxide production from thecells
To test whether v-Ha-Ras protein mediatedsuperoxide production we treated U23 cellswith either lovastatin or a-hydroxyfarnesyl-phosphonic acid (a-PA) inhibitors of Ras pro-tein farnesylation (26) superoxide productionwas decreased 50 by each of these inhibi-tors (Fig 2B) and statistical analysis showedthat the difference was significant (p 005)This result demonstrated that Ras mediated thesuperoxide production from the v-Ha-ras trans-duced cells We did not see complete inhibi-tion there may be certain functional Ras pro-teins formed in spite of the drug treatment
We then studied the source of superoxideproduction from the v-Ha-ras transfected cellsNeutrophils after stimulation are well knownto produce superoxide through an NADPH ox-idase system (8) there is evidence showing thatother types of cells including epithelial cellsalso have an NADPH oxidase system in theirplasma membrane (22 29 31 32) Mitochon-dria are also an important source of superox-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 701
FIG 1 v-Ha-Ras was overexpressed in v-Ha-ras transduced REC cells (A) Western blotting analysis Cell lysates(100 mg) were run on 125 SDS-PAGE The proteins were electrotransferred to nitrocellulose sheets and treated withmouse monoclonal antibody to pan-Ras (B) Northern blotting analysis Total RNA (18 mg) was fractionated on 15agarose gel transferred onto a nylon membrane and hybridized with 32P-labeled probe made from human v-Ha-rascDNA 18S RNA was used to normalize the RNA loading amount
ide it is estimated in respiring cells that at least1ndash3 of total respiration is used to generate su-peroxide and hydrogen peroxide in elaborateelectron-transfer chains for the reduction ofmolecular oxygen to water (18) When U23 cellswere treated with diphenylene iodonium(DPI) an inhibitor of NADPH oxidase 100 mM
for 2 h before the cytochrome c assay super-oxide production was decreased to 50 of theoriginal amount and this difference was sta-tistically significant (Fig 2B p 005) How-ever when U23 cells were treated with 50 mMrotenone an inhibitor of mitochondrial respi-ratory electron-transfer chains for 2 h before
YANG ET AL702
FIG 2 Ras mediated superoxide production from the SOD-inhibitable cytochrome c assay (A) Cells were washedthen incubated with 100 mM cytochrome c in KrebsndashRinger buffer at 37degC 5 CO2 for 1 h Samples were scannedfrom 400 nm to 600 nm the 550-nm peak height was measured and adjusted for baseline and then the amount ofsuperoxide was determined Means were calculated from three experiments and error bars represent SD Signifi-cantly different from REC or Neo at the p 005 level Data indicate that Ras transduced cells had increased levelsof superoxide radical (B) U23 cells were further treated with rotenone (50 mM) DPI (100 mM) 2 h lovastatin (50 mM)48 h or a-PA (4 mM) 48 h before measurements or CuZnSOD protein (200 Uml final) was added into the cytochromec solution during the measurement Cell number was counted after the assay data were calculated and expressed asthe rate of superoxide production per hour (nmol106 cells 3 h) Means were calculated from three experiments anderror bars represent SD Significantly different from U23 at the p 005 level The data show that CuZnSOD pro-tein DPI a-PA and lovastatin all inhibited superoxide levels
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

Cell population doubling time assay
Cells (2 3 104) were plated in 15 ml of com-plete medium in 24-well plates the mediumwas changed every 4 days Cells weretrypsinized and then counted on alternate daysfor 2 weeks using a hemocytometer Cell pop-ulation doubling time (DT) in hours was de-termined using the following equation (13)
DT (hours) 5 0693 (t 2 t0)ln (NtN0)
where t0 is time at which exponential growthbegan t is time in hours Nt is cell number attime t and N0 is initial cell number
Gel mobility shift assay
Nuclear protein was extracted by the fol-lowing procedure Cells were collected when80 confluent and rinsed with phosphate-buffered saline buffer Cells were then har-vested by scraping in ice-cold buffer A (10 mMHEPES 15 mM MgCl2 10 mM KCl) and incu-bated on ice for 20 min The cells were lysedwith a Dounce homogenizer (Kontes VinelandNJ USA) and the nuclei were pelleted bycentrifuging twice at 1400g for 30 s at 4degC withsubsequent removal of supernatant The nu-clear pellets were resuspended in ice-coldbuffer C (20 mM HEPES 25 glycerol 042 MNaCl 15 mM MgCl2 02 mM EDTA 05 mMphenylmethylsulfonyl fluoride) and placed onice for 15 min The suspensions were cen-trifuged (14000g for 10ndash15 min) and the su-pernatants were removed and diluted with ice-cold buffer D (20 mM HEPES 20 glycerol 01M KCl 02 mM EDTA 05 mM phenylmethyl-sulfonyl fluoride) The protein concentrationswere determined and nuclear protein extractswere stored at 280degC
DNA binding reactions were carried out with 15 mg of extracted nuclear protein anddouble-stranded synthetic oligodeoxynucleo-tide encompassing an AP-1 consensus sequence59-agcttgTGACTCAccgtag-39 AP-2 consensussequence 59-agctcaaGCCCGCGGGCtctctag-39or NF-kB consensus sequence 59-agctga-GGGGACTTTCCctag-39 Probes were 32P-la-beled using Klenow DNA polymerase and [a-32P]dCTP Nuclear protein extracts (15 mg)were incubated with 32P-labeled DNA probes
in the presence of 1 mg of poly(dIdC) (Phar-macia Biotech Inc Piscataway NJ USA) and13 gel shift buffer (10 mM Tris pH 75 50 mMNaCl 1 mM MgCl2 05 mM EDTA 05 mMdithiothreitol 4 glycerol) at room tempera-ture for 20 min The bound DNAndashprotein com-plexes were separated from free probe by gelelectrophoresis in 5 native polyacrylamidegels in 13 TBE (90 mM Tris-borate 2 mMEDTA) Electrophoresis was conducted at aconstant current of 20 mA for 1 h The gel waswrapped in a plastic wrap and exposed to x-ray film overnight at 280degC AP-1 DNA-bind-ing specificity was determined by competitionexperiments Nuclear protein extracts (15 mg)were preincubated with 1 mg of poly(dIdC) 13gel shift buffer and a 10- or 100-fold excess ofunlabeled AP-1 AP-2 or NF-kB oligonu-cleotides for 15 min at room temperature be-fore the addition of the labeled probe For su-pershift assays nuclear protein extracts (15 mg)were preincubated with 2 mg of anti-c-JunAP-1 anti-AP-2a or anti-p50 (NF-kB) antibodies(Santa Cruz Biotechnology Santa Cruz CAUSA) for 30 min at room temperature andthen incubated with 32P-labeled probes
Statistical analysis
Statistical analysis was performed using SY-STAT (Systat Evanston IL USA) A singlefactor analysis of variance followed by a post-hoc Tukey test was used to determine statisti-cal differences between means The null hy-pothesis was rejected at the 005 level ofsignificance All means were calculated fromthree experiments and error bars representstandard deviations (SD) All western blot andactivity gel assays were repeated at least twice
RESULTS
v-Ha-ras protein was overexpressed and mediatedsuperoxide production
REC cells were transfected with v-Ha-rasthrough the retrovirus plasmid pZIPNeo-SV(X)1 construct Clones were selected and theexpression of v-Ha-Ras protein was detectedby western blotting The clones with differentincreased levels of v-Ha-Ras immunoreactive
YANG ET AL700
protein (W13 U12 and U23) were used for fur-ther experiments (Fig 1A) The northern blot(Fig 1B) demonstrated two new mRNA bands(47 and 6 kb) in W13 U12 and U23 but not inREC or Neo the new bands were therefore gen-erated from the v-Ha-ras expression vector Be-cause of what has been observed earlier inother cells the likely explanations for the twonew bands are the proviral full-length mRNAand a spliced variant The intensity of the newbands suggested that v-Ha-ras was efficientlytranscribed in W13 U12 and U23 cells The 1-kb mRNA from the endogenous c-Ha-ras genewas also found its intensity was increased inW13 U12 and U23 clones There are at leasttwo possibilities for this observation besidesthe RNA loading One is that c-Ha-ras oncogeneis up regulated by v-Ha-ras overexpression theother is that some v-Ha-rasmRNAs decomposeto a 1-kb size These data suggest that the Rasprotein comes from the exogenous plasmid butdoes not exclude the possibility that some Rasprotein may come from the endogenous gene
Recent evidence has demonstrated that Rastransformed cells can stimulate cellular super-oxide production (11 22 29 32) To test if v-Ha-ras transduced REC cells produce moresuperoxide we used Fe(III) cytochrome c tomonitor the amount of superoxide productionfrom the cells Figure 2A shows that v-Ha-ras
transduced cells U12 and U23 generated largeamounts of superoxide up to fivefold morethan REC or Neo cells The addition of exoge-nous CuZnSOD protein totally inhibited the in-creased superoxide levels seen in U23 cells (Fig2B) which demonstrated that the 550-nm peakwas due to superoxide production from thecells
To test whether v-Ha-Ras protein mediatedsuperoxide production we treated U23 cellswith either lovastatin or a-hydroxyfarnesyl-phosphonic acid (a-PA) inhibitors of Ras pro-tein farnesylation (26) superoxide productionwas decreased 50 by each of these inhibi-tors (Fig 2B) and statistical analysis showedthat the difference was significant (p 005)This result demonstrated that Ras mediated thesuperoxide production from the v-Ha-ras trans-duced cells We did not see complete inhibi-tion there may be certain functional Ras pro-teins formed in spite of the drug treatment
We then studied the source of superoxideproduction from the v-Ha-ras transfected cellsNeutrophils after stimulation are well knownto produce superoxide through an NADPH ox-idase system (8) there is evidence showing thatother types of cells including epithelial cellsalso have an NADPH oxidase system in theirplasma membrane (22 29 31 32) Mitochon-dria are also an important source of superox-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 701
FIG 1 v-Ha-Ras was overexpressed in v-Ha-ras transduced REC cells (A) Western blotting analysis Cell lysates(100 mg) were run on 125 SDS-PAGE The proteins were electrotransferred to nitrocellulose sheets and treated withmouse monoclonal antibody to pan-Ras (B) Northern blotting analysis Total RNA (18 mg) was fractionated on 15agarose gel transferred onto a nylon membrane and hybridized with 32P-labeled probe made from human v-Ha-rascDNA 18S RNA was used to normalize the RNA loading amount
ide it is estimated in respiring cells that at least1ndash3 of total respiration is used to generate su-peroxide and hydrogen peroxide in elaborateelectron-transfer chains for the reduction ofmolecular oxygen to water (18) When U23 cellswere treated with diphenylene iodonium(DPI) an inhibitor of NADPH oxidase 100 mM
for 2 h before the cytochrome c assay super-oxide production was decreased to 50 of theoriginal amount and this difference was sta-tistically significant (Fig 2B p 005) How-ever when U23 cells were treated with 50 mMrotenone an inhibitor of mitochondrial respi-ratory electron-transfer chains for 2 h before
YANG ET AL702
FIG 2 Ras mediated superoxide production from the SOD-inhibitable cytochrome c assay (A) Cells were washedthen incubated with 100 mM cytochrome c in KrebsndashRinger buffer at 37degC 5 CO2 for 1 h Samples were scannedfrom 400 nm to 600 nm the 550-nm peak height was measured and adjusted for baseline and then the amount ofsuperoxide was determined Means were calculated from three experiments and error bars represent SD Signifi-cantly different from REC or Neo at the p 005 level Data indicate that Ras transduced cells had increased levelsof superoxide radical (B) U23 cells were further treated with rotenone (50 mM) DPI (100 mM) 2 h lovastatin (50 mM)48 h or a-PA (4 mM) 48 h before measurements or CuZnSOD protein (200 Uml final) was added into the cytochromec solution during the measurement Cell number was counted after the assay data were calculated and expressed asthe rate of superoxide production per hour (nmol106 cells 3 h) Means were calculated from three experiments anderror bars represent SD Significantly different from U23 at the p 005 level The data show that CuZnSOD pro-tein DPI a-PA and lovastatin all inhibited superoxide levels
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

protein (W13 U12 and U23) were used for fur-ther experiments (Fig 1A) The northern blot(Fig 1B) demonstrated two new mRNA bands(47 and 6 kb) in W13 U12 and U23 but not inREC or Neo the new bands were therefore gen-erated from the v-Ha-ras expression vector Be-cause of what has been observed earlier inother cells the likely explanations for the twonew bands are the proviral full-length mRNAand a spliced variant The intensity of the newbands suggested that v-Ha-ras was efficientlytranscribed in W13 U12 and U23 cells The 1-kb mRNA from the endogenous c-Ha-ras genewas also found its intensity was increased inW13 U12 and U23 clones There are at leasttwo possibilities for this observation besidesthe RNA loading One is that c-Ha-ras oncogeneis up regulated by v-Ha-ras overexpression theother is that some v-Ha-rasmRNAs decomposeto a 1-kb size These data suggest that the Rasprotein comes from the exogenous plasmid butdoes not exclude the possibility that some Rasprotein may come from the endogenous gene
Recent evidence has demonstrated that Rastransformed cells can stimulate cellular super-oxide production (11 22 29 32) To test if v-Ha-ras transduced REC cells produce moresuperoxide we used Fe(III) cytochrome c tomonitor the amount of superoxide productionfrom the cells Figure 2A shows that v-Ha-ras
transduced cells U12 and U23 generated largeamounts of superoxide up to fivefold morethan REC or Neo cells The addition of exoge-nous CuZnSOD protein totally inhibited the in-creased superoxide levels seen in U23 cells (Fig2B) which demonstrated that the 550-nm peakwas due to superoxide production from thecells
To test whether v-Ha-Ras protein mediatedsuperoxide production we treated U23 cellswith either lovastatin or a-hydroxyfarnesyl-phosphonic acid (a-PA) inhibitors of Ras pro-tein farnesylation (26) superoxide productionwas decreased 50 by each of these inhibi-tors (Fig 2B) and statistical analysis showedthat the difference was significant (p 005)This result demonstrated that Ras mediated thesuperoxide production from the v-Ha-ras trans-duced cells We did not see complete inhibi-tion there may be certain functional Ras pro-teins formed in spite of the drug treatment
We then studied the source of superoxideproduction from the v-Ha-ras transfected cellsNeutrophils after stimulation are well knownto produce superoxide through an NADPH ox-idase system (8) there is evidence showing thatother types of cells including epithelial cellsalso have an NADPH oxidase system in theirplasma membrane (22 29 31 32) Mitochon-dria are also an important source of superox-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 701
FIG 1 v-Ha-Ras was overexpressed in v-Ha-ras transduced REC cells (A) Western blotting analysis Cell lysates(100 mg) were run on 125 SDS-PAGE The proteins were electrotransferred to nitrocellulose sheets and treated withmouse monoclonal antibody to pan-Ras (B) Northern blotting analysis Total RNA (18 mg) was fractionated on 15agarose gel transferred onto a nylon membrane and hybridized with 32P-labeled probe made from human v-Ha-rascDNA 18S RNA was used to normalize the RNA loading amount
ide it is estimated in respiring cells that at least1ndash3 of total respiration is used to generate su-peroxide and hydrogen peroxide in elaborateelectron-transfer chains for the reduction ofmolecular oxygen to water (18) When U23 cellswere treated with diphenylene iodonium(DPI) an inhibitor of NADPH oxidase 100 mM
for 2 h before the cytochrome c assay super-oxide production was decreased to 50 of theoriginal amount and this difference was sta-tistically significant (Fig 2B p 005) How-ever when U23 cells were treated with 50 mMrotenone an inhibitor of mitochondrial respi-ratory electron-transfer chains for 2 h before
YANG ET AL702
FIG 2 Ras mediated superoxide production from the SOD-inhibitable cytochrome c assay (A) Cells were washedthen incubated with 100 mM cytochrome c in KrebsndashRinger buffer at 37degC 5 CO2 for 1 h Samples were scannedfrom 400 nm to 600 nm the 550-nm peak height was measured and adjusted for baseline and then the amount ofsuperoxide was determined Means were calculated from three experiments and error bars represent SD Signifi-cantly different from REC or Neo at the p 005 level Data indicate that Ras transduced cells had increased levelsof superoxide radical (B) U23 cells were further treated with rotenone (50 mM) DPI (100 mM) 2 h lovastatin (50 mM)48 h or a-PA (4 mM) 48 h before measurements or CuZnSOD protein (200 Uml final) was added into the cytochromec solution during the measurement Cell number was counted after the assay data were calculated and expressed asthe rate of superoxide production per hour (nmol106 cells 3 h) Means were calculated from three experiments anderror bars represent SD Significantly different from U23 at the p 005 level The data show that CuZnSOD pro-tein DPI a-PA and lovastatin all inhibited superoxide levels
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

ide it is estimated in respiring cells that at least1ndash3 of total respiration is used to generate su-peroxide and hydrogen peroxide in elaborateelectron-transfer chains for the reduction ofmolecular oxygen to water (18) When U23 cellswere treated with diphenylene iodonium(DPI) an inhibitor of NADPH oxidase 100 mM
for 2 h before the cytochrome c assay super-oxide production was decreased to 50 of theoriginal amount and this difference was sta-tistically significant (Fig 2B p 005) How-ever when U23 cells were treated with 50 mMrotenone an inhibitor of mitochondrial respi-ratory electron-transfer chains for 2 h before
YANG ET AL702
FIG 2 Ras mediated superoxide production from the SOD-inhibitable cytochrome c assay (A) Cells were washedthen incubated with 100 mM cytochrome c in KrebsndashRinger buffer at 37degC 5 CO2 for 1 h Samples were scannedfrom 400 nm to 600 nm the 550-nm peak height was measured and adjusted for baseline and then the amount ofsuperoxide was determined Means were calculated from three experiments and error bars represent SD Signifi-cantly different from REC or Neo at the p 005 level Data indicate that Ras transduced cells had increased levelsof superoxide radical (B) U23 cells were further treated with rotenone (50 mM) DPI (100 mM) 2 h lovastatin (50 mM)48 h or a-PA (4 mM) 48 h before measurements or CuZnSOD protein (200 Uml final) was added into the cytochromec solution during the measurement Cell number was counted after the assay data were calculated and expressed asthe rate of superoxide production per hour (nmol106 cells 3 h) Means were calculated from three experiments anderror bars represent SD Significantly different from U23 at the p 005 level The data show that CuZnSOD pro-tein DPI a-PA and lovastatin all inhibited superoxide levels
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709
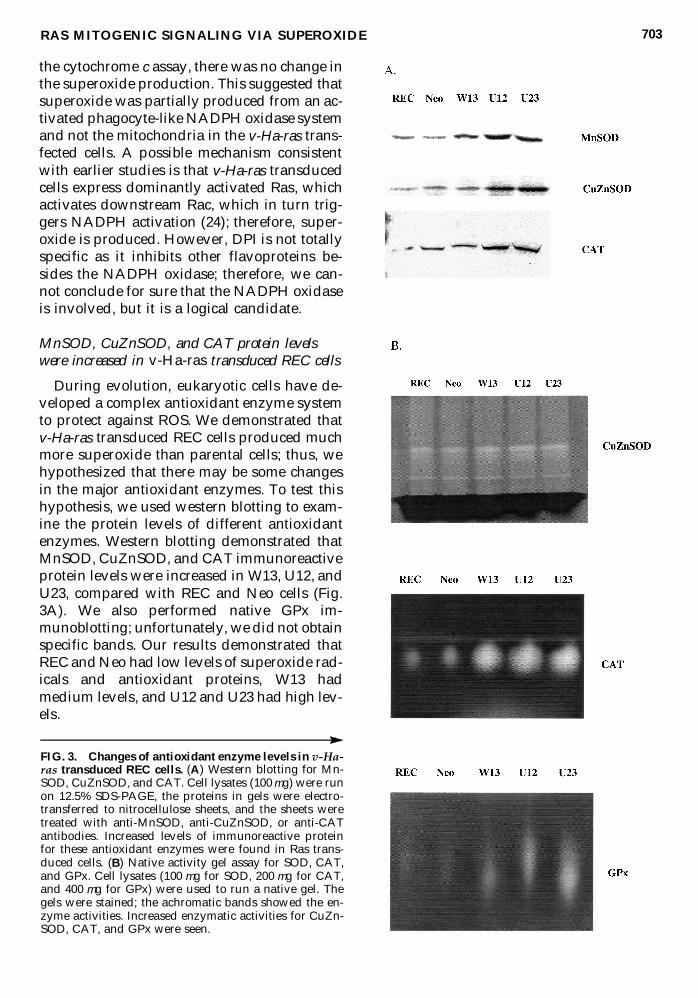
the cytochrome c assay there was no change inthe superoxide production This suggested thatsuperoxide was partially produced from an ac-tivated phagocyte-like NADPH oxidase systemand not the mitochondria in the v-Ha-ras trans-fected cells A possible mechanism consistentwith earlier studies is that v-Ha-ras transducedcells express dominantly activated Ras whichactivates downstream Rac which in turn trig-gers NADPH activation (24) therefore super-oxide is produced However DPI is not totallyspecific as it inhibits other flavoproteins be-sides the NADPH oxidase therefore we can-not conclude for sure that the NADPH oxidaseis involved but it is a logical candidate
MnSOD CuZnSOD and CAT protein levelswere increased in v-Ha-ras transduced REC cells
During evolution eukaryotic cells have de-veloped a complex antioxidant enzyme systemto protect against ROS We demonstrated thatv-Ha-ras transduced REC cells produced muchmore superoxide than parental cells thus wehypothesized that there may be some changesin the major antioxidant enzymes To test thishypothesis we used western blotting to exam-ine the protein levels of different antioxidantenzymes Western blotting demonstrated thatMnSOD CuZnSOD and CAT immunoreactiveprotein levels were increased in W13 U12 andU23 compared with REC and Neo cells (Fig3A) We also performed native GPx im-munoblotting unfortunately we did not obtainspecific bands Our results demonstrated thatREC and Neo had low levels of superoxide rad-icals and antioxidant proteins W13 hadmedium levels and U12 and U23 had high lev-els
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 703
FIG 3 Changes of antioxidant enzyme levels in v-Ha-ras transduced REC cells (A) Western blotting for Mn-SOD CuZnSOD and CAT Cell lysates (100 mg) were runon 125 SDS-PAGE the proteins in gels were electro-transferred to nitrocellulose sheets and the sheets weretreated with anti-MnSOD anti-CuZnSOD or anti-CATantibodies Increased levels of immunoreactive proteinfor these antioxidant enzymes were found in Ras trans-duced cells (B) Native activity gel assay for SOD CATand GPx Cell lysates (100 mg for SOD 200 mg for CATand 400 mg for GPx) were used to run a native gel Thegels were stained the achromatic bands showed the en-zyme activities Increased enzymatic activities for CuZn-SOD CAT and GPx were seen
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709
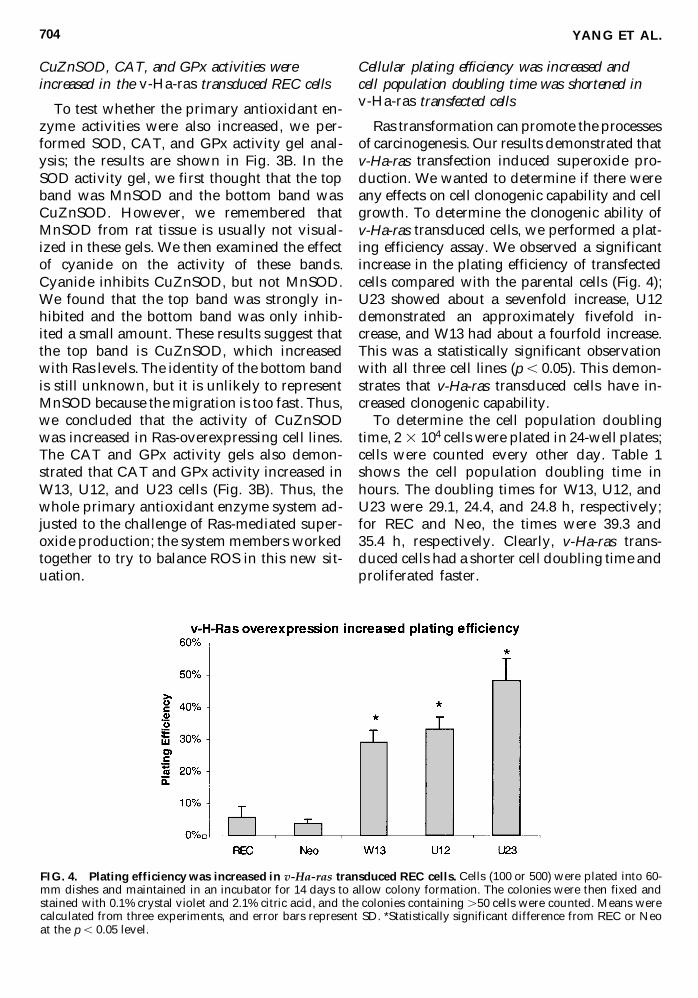
CuZnSOD CAT and GPx activities wereincreased in the v-Ha-ras transduced REC cells
To test whether the primary antioxidant en-zyme activities were also increased we per-formed SOD CAT and GPx activity gel anal-ysis the results are shown in Fig 3B In theSOD activity gel we first thought that the topband was MnSOD and the bottom band wasCuZnSOD However we remembered thatMnSOD from rat tissue is usually not visual-ized in these gels We then examined the effectof cyanide on the activity of these bandsCyanide inhibits CuZnSOD but not MnSODWe found that the top band was strongly in-hibited and the bottom band was only inhib-ited a small amount These results suggest thatthe top band is CuZnSOD which increasedwith Ras levels The identity of the bottom bandis still unknown but it is unlikely to representMnSOD because the migration is too fast Thuswe concluded that the activity of CuZnSODwas increased in Ras-overexpressing cell linesThe CAT and GPx activity gels also demon-strated that CAT and GPx activity increased inW13 U12 and U23 cells (Fig 3B) Thus thewhole primary antioxidant enzyme system ad-justed to the challenge of Ras-mediated super-oxide production the system members workedtogether to try to balance ROS in this new sit-uation
Cellular plating efficiency was increased and cell population doubling time was shortened in v-Ha-ras transfected cells
Ras transformation can promote the processesof carcinogenesis Our results demonstrated thatv-Ha-ras transfection induced superoxide pro-duction We wanted to determine if there wereany effects on cell clonogenic capability and cellgrowth To determine the clonogenic ability ofv-Ha-ras transduced cells we performed a plat-ing efficiency assay We observed a significantincrease in the plating efficiency of transfectedcells compared with the parental cells (Fig 4)U23 showed about a sevenfold increase U12demonstrated an approximately fivefold in-crease and W13 had about a fourfold increaseThis was a statistically significant observationwith all three cell lines (p 005) This demon-strates that v-Ha-ras transduced cells have in-creased clonogenic capability
To determine the cell population doublingtime 2 3 104 cells were plated in 24-well platescells were counted every other day Table 1shows the cell population doubling time inhours The doubling times for W13 U12 andU23 were 291 244 and 248 h respectivelyfor REC and Neo the times were 393 and 354 h respectively Clearly v-Ha-ras trans-duced cells had a shorter cell doubling time andproliferated faster
YANG ET AL704
FIG 4 Plating efficiency was increased in v-Ha-ras transduced REC cells Cells (100 or 500) were plated into 60-mm dishes and maintained in an incubator for 14 days to allow colony formation The colonies were then fixed andstained with 01 crystal violet and 21 citric acid and the colonies containing 50 cells were counted Means werecalculated from three experiments and error bars represent SD Statistically significant difference from REC or Neoat the p 005 level
v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

v-Ha-ras transfection enhanced AP-1 AP-2 andNF-kB DNA binding
The effects of v-Ha-ras transfection on tran-scription factor DNA-binding activity was in-vestigated by electrophoretic gel mobility shiftassays AP-1 AP-2 and NF-kB DNA-bindingactivities were significantly increased in v-Ha-ras transduced cells compared with parentalREC cells and vector controls Fig 5A showsthat AP-1 DNA binding was greatly enhancedin the Ras transfectants W13 U12 and U23when compared with that in parental and Neocells However when U23 nuclear protein ex-tracts were preincubated with 2 mg of anti-c-JunAP-1 antibody and then incubated withAP-1 32P-labeled probe the DNA-binding ac-tivity was greatly inhibited (Fig 5A lane 6)The specificity of AP-1 binding to the DNAprobe was further demonstrated by competi-tion experiments with unlabeled oligos TheDNAndashprotein complex was partially or com-pletely abolished in the presence of a 10- or 100-fold excess of unlabeled AP-1 probe (Fig 5Blanes 3 and 4) whereas 10- or 100-fold excessof unlabeled AP-2 and NF-kB oligos did not ef-fectively compete for protein bound to the la-beled AP-1 probe (Fig 5B lanes 5ndash8) These re-sults demonstrate that the bands in W13 U12and U23 lanes were due specifically to AP-1DNA binding
AP-2 DNA binding was also enhanced inW13 U12 and U23 cells compared with that inREC and Neo (Fig 5C) The binding activitywas supershifted when W13 nuclear proteinextracts were preincubated with 2 mg of anti-AP-2a antibody (Fig 5C lane 6) which dem-onstrates that the bands in the W13 U12 andU23 lanes were due specifically to AP-2 DNAbinding Interestingly AP-2 DNA-binding ac-
tivities appeared to decrease with increasing v-Ha-ras expression NF-kB DNA-binding activ-ity was also higher in W13 U12 and U23 (Fig5D) and supershift assays demonstrated thatthe binding was due specifically to the NF-kBndashprobe complex formation (Fig 5D lane 6)SP-1 was not activated in any of the cell linesstudied (data not shown)
v-Ha-ras transduction increased p44p42 andp38 kinase activities
In many cells MAPK activation is depen-dent on the production of ROS after the stim-ulation of growth factors (7 14 15 30) MAPKsin turn phosphorylate and activate certain tran-scription factors We have demonstrated thatDNA-binding activities of AP-1 AP-2 and NF-kB were increased in v-Ha-ras transduced RECcells Therefore we investigated whether cer-tain MAPKs were turned on in these cells Wefocused on these kinases including p4442stress-activated protein kinaseJNK and p38MAPK because they can phosphorylate andactivate transcription factors However the ac-tivities of MAPKs are also dependent on theirremaining in the phosphorylated form as onlythe phosphorylated kinases are active Thustheir activities could be inferred by westernblotting using antibodies against the phospho-rylated form of these kinases Fig 6 suggeststhat p4442 MAPK and p38 MAPK activitieswere increased in W13 U12 and U23 but notin REC and Neo cells because both the phos-pho-p4442 MAPK and phospho-p38 MAPKprotein levels were increased in W13 U12 andU23 cells Western blotting for phospho-JNKwas also performed but the result showed nodifference in the protein level (data not shown)We cannot actually conclude in these studies
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 705
TABLE 1 V-HA-RAS EXPRESSION SHORTENED CELL POPULATION DOUBLING TIME
Cells REC Neo W13 U12 U23
Cell population doubling time (h) 393 6 29 354 6 48 291 6 28 244 6 23 248 6 19
Cells (2 3 104) were plated in 15 ml of complete medium in 24-well plates Cells were trypsinized and then countedon alternate days for 2 weeks using a hemocytometer and cell population doubling time was calculated Data aremeans 6 SDU12 and U23 are significantly different from REC or Neo (p 005)
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709
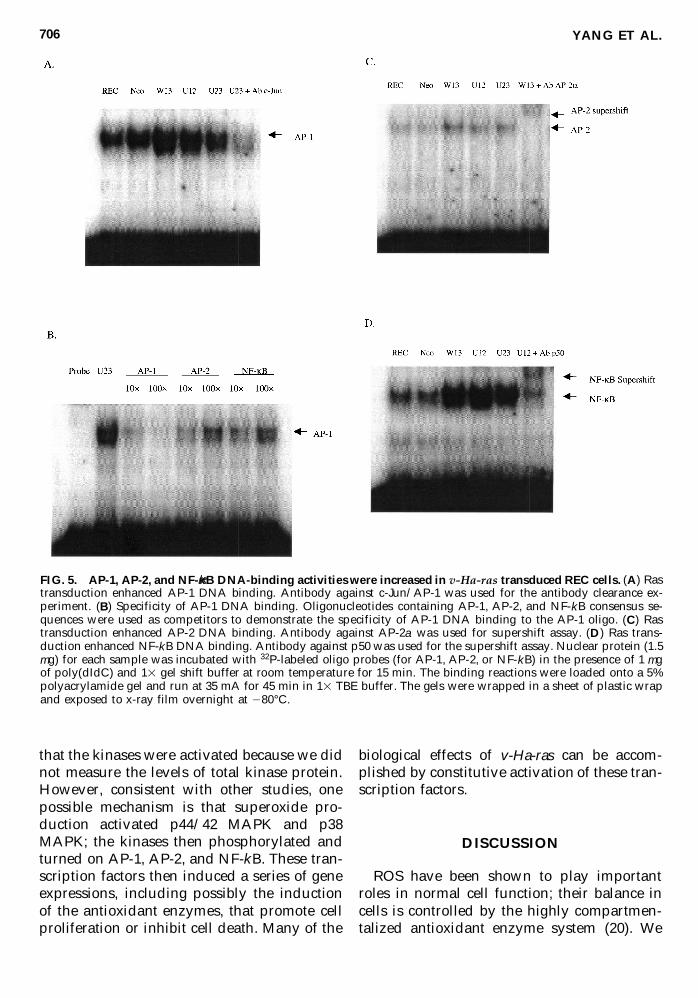
that the kinases were activated because we didnot measure the levels of total kinase proteinHowever consistent with other studies onepossible mechanism is that superoxide pro-duction activated p4442 MAPK and p38MAPK the kinases then phosphorylated andturned on AP-1 AP-2 and NF-kB These tran-scription factors then induced a series of geneexpressions including possibly the inductionof the antioxidant enzymes that promote cellproliferation or inhibit cell death Many of the
biological effects of v-Ha-ras can be accom-plished by constitutive activation of these tran-scription factors
DISCUSSION
ROS have been shown to play importantroles in normal cell function their balance incells is controlled by the highly compartmen-talized antioxidant enzyme system (20) We
YANG ET AL706
FIG 5 AP-1 AP-2 and NF-kB DNA-binding activities were increased in v-Ha-ras transduced REC cells (A) Rastransduction enhanced AP-1 DNA binding Antibody against c-JunAP-1 was used for the antibody clearance ex-periment (B) Specificity of AP-1 DNA binding Oligonucleotides containing AP-1 AP-2 and NF-kB consensus se-quences were used as competitors to demonstrate the specificity of AP-1 DNA binding to the AP-1 oligo (C) Rastransduction enhanced AP-2 DNA binding Antibody against AP-2a was used for supershift assay (D) Ras trans-duction enhanced NF-kB DNA binding Antibody against p50 was used for the supershift assay Nuclear protein (15mg) for each sample was incubated with 32P-labeled oligo probes (for AP-1 AP-2 or NF-kB) in the presence of 1 mgof poly(dIdC) and 13 gel shift buffer at room temperature for 15 min The binding reactions were loaded onto a 5polyacrylamide gel and run at 35 mA for 45 min in 13 TBE buffer The gels were wrapped in a sheet of plastic wrapand exposed to x-ray film overnight at 280degC
have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

have demonstrated that v-Ha-ras mediated su-peroxide production in Ras transfected RECcells Moreover Ras overexpression also mod-ulated antioxidant enzyme levels increasedDNA-binding activity of transcription factorsAP-1 AP-2 and NF-kB and increased proteinlevels of phosphorylated MAPK p44p42 andp38 in these cells The transfected cells pos-sessed typical Ras phenotypes including short-ened cell doubling time and increased cellularplating efficiency These results suggest thatRas mitogenic signaling may be mediatedpartly though redox regulation by superoxide
Oncogenic ras can transform many immortalcell lines to the tumorigenic state and plays acentral role in cellular signal transductionThere are at least two possible signaling path-ways for Ras to pass mitogenic signaling oneis the well defined RasndashRaf pathway and an-other is the RasndashRacndashsuperoxide pathway Inthe second pathway Ras activates Rac whichcan promote the NADPH oxidase to generatesuperoxide superoxide can activate MAPKsand transcription factors to induce mitogeniceffects Previous work has shown that Ras canmediate superoxide overproduction When fi-broblasts were transformed by mutant onco-genic Ras superoxide production was detectedin the transformed cells but not in parental andvector control cells and the Ras-transformedcells displayed a greater rate of DNA synthe-sis (11) Dr Bar-Sagirsquos research suggests that
Rac-induced superoxide production is a criti-cal mediator of mitogenic signaling (12) Thisevidence suggests that Ras proteins could in-duce mitogenic signaling partially throughmodification of the superoxide production inthese cells
Our studies have demonstrated for the firsttime that v-Ha-ras transduced REC cells in-creased primary antioxidant enzyme levels including MnSOD and CuZnSOD levels How-ever cancer cells generally express low con-stitutive levels of MnSOD and CuZnSOD whencompared with their cells of origin (19 20) Onepossible reason for this discrepancy in antiox-idant proteins is that cancer development is along and multiple-step process during thisprocess MnSOD and CuZnSOD gene tran-scription becomes less active than their cells oforigin We have found that DNA methylationin the MnSOD promoter region could inhibittranscription factor binding (10) which pro-vides one mechanism to explain this phenom-enon Although the cell line we used in thepresent studies is immortal it is still in an earlystage of the carcinogenic process (less than 30thpassage) and thus the primary antioxidant en-zyme genes could still show normal transcrip-tion activity Another paradox is that increasedSOD activity has usually been shown to slowcell proliferation in either normal or malignantcells (13 20) in the present work increasedSOD is associated with increased cell prolifer-
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 707
FIG 6 The protein levels of phospho-p44p42 and phospho-p38 MAPK were increased in the Ras-transducedREC cells Cell lysates (200 mg) were used to run on 125 SDS-PAGE then the proteins in gels were electrotrans-ferred to nitrocellulose sheets and the sheets treated with anti-phospho-p44p42 MAPK or anti-phospho-p38 MAPKantibodies Western blotting demonstrated that both phospho-ERK and phospho-p38 MAPK protein levels were in-creased Coomassie Blue staining of transferred gel indicated equal protein loading
ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

ation One possible explanation for this is thatthe cells with higher SOD activity may be pro-tected against cell death due to apoptosis or ne-crosis
Approximately 30 of all human tumorshave ras mutations which implies that in-creased superoxide may be produced in thesecells However cancer cells usually have lessMnSOD and CuZnSOD activity this suggeststhat cancer cells may take advantage of super-oxide overproduction to promote cell prolifer-ation As less superoxide is scavenged by Mn-SOD or CuZnSOD tumor cells can maintain arelatively higher steady-state level of superox-ide which may act as a mediator of mitogenicsignaling In the present study it appeared thatthe hydrogen peroxide-scavenging enzymeswere increased more by Ras overexpressionthan the superoxide-scavenging enzymes Thiswould suggest that Ras overexpression mightlead to larger increases in superoxide radicalthan hydrogen peroxide Studies of the effectsof modulating superoxide production on Rasmitogenic signaling will provide insights intothe mechanisms linking superoxide andgrowth control
ACKNOWLEDGMENTS
This work was supported by NIH grantsP50-DE10758 and P01-CA66081
ABBREVIATIONS
AP-1 and AP-2 activator protein-1 and -2CAT catalase CuZnSOD copper- and zinc-containing superoxide dismutase DPIdiphenylene iodonium GPx glutathione per-oxidase JNK c-jun N-terminal kinase MAPKmitogen-activated protein kinase MnSODmanganese-containing superoxide dismutaseNF-kB nuclear factor-kB a-PA a-hydroxyfar-nesylphosphonic acid PAGE polyacrylamidegel electrophoresis Ras Ras protein ras rasgene or mRNA REC rat kidney epithelial cellROS reactive oxygen species SD standard de-viation SDS sodium dodecyl sulfate SOD su-peroxide dismutase SSC salinendashsodium citratebuffer TBE TrisndashboratendashEDTA
REFERENCES
1 Beauchamp C and Fridovich I Superoxide dismutaseimproved assays and an assay applicable to acry-lamide gels Anal Biochem 44 276ndash287 1971
2 Butler J Koppenol WH and Margoliash E Kineticsand mechanism of the reduction of ferricytochrome cby the superoxide anion J Biol Chem 257 10747ndash107501982
3 Cepko CL Roberts BE and Mulligan RC Construc-tion and applications of a highly transmissible murineretrovirus shuttle vector Cell 37 1053ndash1062 1984
4 Cotran RS Kumar V and Robbins SL Pathologic Ba-sic of Disease 5th edit Philadelphia PA WB Saun-ders Company 1994
5 Freeman BA Biological sites and mechanisms of freeradical production In Free Radicals in Molecular Biol-ogy Aging and Disease edited by Armstrong D SohalRS Cutler RG and Slater TF New York Raven Press1984 pp 43ndash52
6 Fridovich I Cytochrome c In CRC Handbook of Meth-ods for Oxygen Radical Research edited by GreenwaldRA Boca Raton FL CRC Press Inc 1985 pp121ndash122
7 Guyton KZ Liu Y Gorospe M Xu Q and HolbrookN Activation of mitogen-activated protein kinase byH2O2 Role in cell survival following oxidant injuryJ Biol Chem 271 4138ndash4142 1996
8 Henderson LM and Chappell JB NADPH oxidase ofneutrophils Biochim Biophys Acta 1273 87ndash107 1996
9 Huang Y and Domann FE Redox modulation of AP-2 DNA binding activity in vitro Biochem Biophys ResCommun 249 307ndash312 1998
10 Huang Y Peng J Oberley LW and Domann FETranscriptional inhibition of manganese superoxidedismutase (SOD2) gene expression by DNA methy-lation of 59 CpG island Free Radic Biol Med 23314ndash320 1997
11 Irani K Xia Y Zweier JL Sollott SJ Der CJ FearonER Sundaresan M Finkel T and Goldschmidt-Cler-mont PJ Mitogenic signaling mediated by oxidants inRas-transformed fibroblasts Science 275 1649ndash16521997
12 Joneson T and Bar-Sagi D A Rac1 effector site con-trolling mitogenesis through superoxide productionJ Biol Chem 273 17991ndash17994 1998
13 Lam E Zwacka R Engelhardt JF Davidson BL Do-mann FE Yan T and Oberley LW Adenovirus-me-diated manganese superoxide dismutase gene trans-fer to hamster cheek pouch carcinoma cells CancerRes 57 5550ndash5556 1997
14 Levine A Tenhaken R Dixon R and Lamb C H2O2from the oxidative burst orchestrates the plant hy-persensitive disease resistance response Cell 79583ndash593 1994
15 Liu Y Guyton KZ Gorospe M Xu Q Lee J and Hol-brook N Differential activation of ERK JNKSAPKand p38CSBPRK MAP kinase family members dur-ing the cellular response to arsenite Free Radic BiolMed 21 771ndash781 1996
YANG ET AL708
16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709

16 Lowy DR and Willumsen BM Function and regula-tion of ras Annu Rev Biochem 62 851ndash891 1993
17 Muller JM Cahill MA Rupec RA Baeuerle PA andNordheim A Antioxidants as well as oxidants acti-vate c-fos via Ras-dependent activation of extracellu-lar-signal-regulated kinase 2 and Elk-1 Eur J Biochem244 45ndash52 1997
18 Nohl H and Jordan W The mitochondrial site of su-peroxide formation Biochem Biophys Res Commun 138533ndash539 1986
19 Oberley LW and Buettner GR Role of superoxide dis-mutase in cancer a review Cancer Res 39 1131ndash11491979
20 Oberley LW and Oberley TD Role of antioxidant en-zymes in the cancer phenotype In Oxygen Gene Ex-pression and Cellular Function edited by Clerch L andMassaro D New York NY Marcel Dekker Inc 1997pp 279ndash307
21 Oberley TD Oberley LW Slattery AF Lauchner LJand Elwell JH Immunohistochemical location of anti-oxidant enzymes in adult Syrian hamster tissues andduring kidney development Am J Pathol 137 199ndash2141990
22 Pagano PJ Clark JK Cifuentes-Pagano ME Clark SMCallis GM and Quinn MT Localization of a consti-tutively active phagocyte-like NADPH oxidase in rab-bit aortic adventitia enhancement by angiotensin IIProc Natl Acad Sci U S A 94 14483ndash14488 1997
23 Pai EF Kabsch W Krengel U Holmes KC John J andWittinghofer A Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21in the triphosphate conformation Nature 341 209ndash214 1989
24 Prigmore E Ahmed S Best A Kozma R Manser ESegal AW and Lim L A 68-kDa kinase and NADPHoxidase component p67phox are targets for Cdc42Hsand Rac1 in neutrophils J Biol Chem 270 10717ndash107221995
25 Pulverer BJ Kyriakis JM Avruch J Nikolakaki E andWoodgett JR Phosphorylation of c-jun mediated byMAP kinases Nature 353 670ndash674 1991
26 Santillo M Mondola P Gioielli A Seru R Iossa S An-nella T Vitale M and Bifulco M Inhibitors of Ras far-nesylation revert the increased resistance to oxidativestress in K-Ras transformed NIH 3T3 cells BiochemBiophys Res Commun 229 739ndash745 1996
27 Sen CK and Packer L Antioxidant and redox regula-tion of gene transcription FASEB J 414 709ndash720 1996
28 Sun Y Elwell JH and Oberley LW A simultaneousvisualization of the antioxidant enzymes glutathioneperoxidase and catalase on polyacrylamide gels FreeRadic Res Commun 5 67ndash75 1988
29 Sundaresan M Yu ZX Ferrans VJ Irani K and FinkelT Requirement for generation of H2O2 for platelet-derived growth factor signal transduction Science270 296ndash299 1995
30 Sundaresan M Yu Z Ferrans VJ Sulciner DJ GutkindJS Irani K Goldschmidt-Clermont PJ and Finkel TRegulation of reactive-oxygen-species generation infibroblasts by Rac1 Biochem J 318 379ndash382 1996
31 Turner CP Toye AM and Jones OTG Keratinocytesuperoxide generation Free Radic Biol Med 24 401ndash407 1997
32 Yang J-Q Li J Domann FE Buettner GR and Ober-ley LW Superoxide generation in v-Ha-ras-trans-duced human keratinocyte HaCaT cells Mol Carcinog26 180ndash199 1999
Address reprint requests toDr Larry W Oberley
Free Radical and Radiation Biology ProgramB180 Medical LaboratoriesDepartment of RadiologyThe University of Iowa
Iowa City IA 52242-1181
E-mail larry-oberleyuiowaedu
Received for publication January 8 2001 ac-cepted March 16 2001
RAS MITOGENIC SIGNALING VIA SUPEROXIDE 709
Copyright © 2022 FDOKUMEN




















