
Ontogenetic distribution of genetic lethality in Drosophila willistoni

Ontogenetic allometry in the thoracolumbar spine of mammal species
with differing gait use
Katrina E. Jones,a,* and Rebecca Z. Germanb,1
a Functional Anatomy and Evolution, Johns Hopkins University, Baltimore, MD 21205, USAbDepartment of Physical Medicine and Rehabilitation, Johns Hopkins University, Baltimore, MD 21287, USA*Author for correspondence (e‐mail: [email protected])1Present address: Department of Anatomy and Neurobiology, Northeast Ohio Medical University, Rootstown, OH, USA
SUMMARY Vertebrae are serially homologous structureswith tight integration through their evolution and development.However, in mammals, the thoracic and lumbar regions aremorphologically and functionally differentiated. We test thehypothesis that locomotor specialization is associated withaltered post‐natal growth patterns in vertebrae from differentvertebral regions. We use longitudinal data to examinethoracolumbar growth in two specialized half‐bounding(Oryctolagus cuniculus and Chinchilla lanigera) and twonon‐specialized (Cavia porcellus and Monodelphis domes-tica) species with similar body sizes. Lateral X‐rays of 38individuals were the source of centrum length, centrum heightand intervertebral space length measures for 19–20 thoraco-
lumbar vertebrae. The repeated measurements designincluded the same individuals soon after birth and again atadult size. Data from columns with different vertebral countswere compared by either summing (length) or averaging(height) within regions, and individual vertebrae were directlycompared at the first and last five vertebral positions.Specialized half‐bounders had longer lumbar regions thangeneralists, which was attributable to positively allometricgrowth of the lumbar centra. Lumbar centrum length wasmorevariable both ontogenetically and interspecifically than theother variables, suggesting heterochrony may be generatinglumbar variation. Craniocaudal patterns of centrum growthcorrelate with expression of regionalizing genes (i.e., Hox).
INTRODUCTION
Variation in post‐natal growth is an important mechanism forgenerating evolutionary change (Gould 1977; Alberch et al. 1979;Godfrey and Sutherland 1996), particularly in the mammalianskull and appendicular skeleton (Shea 1981, 1983; McKinney andSchoch 1985; Wayne 1986a,b; Jungers et al. 1988; Shea 1989;Ravosa 1992; Weston 2003; Bastir and Rosas 2004a,b; Schillingand Petrovitch 2006; Drake 2011). However, very little is knownabout the role of growth in the evolution of the vertebral column(O’Higgins et al. 1997; Bergmann et al. 2006). The vertebralcolumn is composed of serially homologous units that areintegrated through their evolution and development (Flower 1885;Minelli and Peruffo 1991; Buchholtz 2007). These vertebrae arefurther organized into modular regions with specific morphologiesand functions (Slijper 1946; Polly et al. 2001; Buchholtz 2007).The vertebral column also plays a vital role in locomotion, inwhichdifferent regions may be subject to divergent selection pressures(Slijper 1946; Hildebrand 1962; Fischer et al. 2002; Schilling andHackert 2006). How does vertebral growth in different anatomicalregions respond to the demands of locomotor specialization?
The influence of locomotion on limb growth has beeninvestigated using similar data to those presented here (Lammersand German 2002). The ontogenetic allometry of the limbs ofsmall mammals that were specialized for a half‐bounding gait
(O. cuniculus, rabbits, and C. lanigera, chinchillas) wascompared to that of generalist quadrupeds (M. domestica, greyshort‐tailed opossums, and Rattus norvegicus, rats). The onlysignificant difference was a narrowing of the distal hindlimbelements (tibia and metatarsus) in half‐bounders. However, theauthors proposed that the vertebral column may show morefunctional signal than the limbs due to its importance in half‐bounding. Our goal is to test this hypothesis using vertebralmeasures from the same dataset. We used a different generalizedspecies (Cavia porcellus, guinea pigs) instead of R. norvegicusbecause it was not possible to obtain vertebral data from thoseoriginal radiographs. The half‐bound is an asymmetricalmammalian gait that is characterized by simultaneous launchby both hindlimbs, followed by an aerial phase, then sequentiallanding of the forefeet (Gambaryan 1974). This gait is used bymany small mammals, however in this study we classify“specialized half‐bounders” as species that use this gait almostexclusively (Lammers and German 2002). In contrast, “general-ist quadrupeds” preferentially use a symmetrical gait (e.g., trot)and only occasionally half‐bound when at their top speeds(Pridmore 1992; Rocha‐Barbosa et al. 2005).
These species represent a broad taxonomic sampling,selected due to their availability as lab animals to collectlongitudinal data. The guinea pig (C. porcellus, generalistquadruped) and the chinchilla (C. lanigera, half‐bounder) are
EVOLUTION & DEVELOPMENT 16:2, 110–120 (2014)
DOI: 10.1111/ede.12069
110 © 2014 Wiley Periodicals, Inc.
both caviomorph rodents from South America. The Europeanrabbit (O. cuniculus, half‐bounder) is an old‐world lagomorphwhich is the sister group to rodents. In contrast the grey short‐tailed opossum (M. domestica, generalist quadruped) representsa phylogenetic outgroup to the placental mammals (Vaughanet al. 2011). It is a marsupial from South America, and thereforehas a more altricial developmental strategy than any of theplacentals (Table 1). Hence, the half‐bounding specializationwas independently evolved in these two species, thus similaritiesin morphology or growth patterns likely represent convergentevolution. Moreover, within each group both altricial andprecocial life histories are represented (Table 1). Thus, anywithin‐group similarities in growth pattern are despite consider-able contrasts in phylogenetic history and development,indicating a relatively strong influence of locomotion on post‐natal development.
These gaits are associated with different functional demandsand uses of the spine. The fastest mammalian gaits are theasymmetrical gaits such as the gallop or half‐bound, gaitsassociated with unique sagittal (dorsoventral) bending move-ments of the spine (Howell 1944; Hildebrand 1959, 1962;Gambaryan 1974; Alexander et al. 1985; Fischer andLehmann 1998; Fischer et al. 2002). This sagittal movementis manifest only in the last six to eight vertebral joints smallmammals (Schilling and Hackert 2006). Because sagittal spinalmovements are predominantly used in asymmetric gaits (i.e., thehalf‐bound) (Pridmore 1992) small mammals who use a half‐bound gait preferentially over a symmetrical gait might haveadaptations of the spine to enhance sagittal mobility. This wouldinclude elongating the thoracolumbar column to increase themoment arm of the spine and stride length (Hatt 1932;Gambaryan 1974). Since the lumbar region is primarilyresponsible for sagittal bending, and is the location of thelargest dorsoventral‐mobilizing epaxial muscles, it might also beelongated relative to the thoracic region (Hatt 1932).
Relatively few data exist on the growth of the vertebralcolumn. Bergman et al.’s work on the rat, R. norvegicus, usingcross‐sectional data (Bergmann et al. 2006), suggests thatincreased adult length of lumbar vertebrae may be caused by acombination of increasing growth rate without increasing decayrate (German and Meyers 1989). Other research indicates that
there are allometric changes, and high levels of variability, butnone of it addresses growth in a specifically locomotor context(O’Higgins et al. 1997; Valverde et al. 2010).
This study uses longitudinal data to assess the influence oflocomotor specialization on growth of different vertebral regionsand structures. Specifically we address five questions:
1. Do half‐bounders show elongation or increased growth of thewhole thoracolumbar column relative to generalistquadrupeds?
2. Is growth of the vertebrae or growth of the intervertebralspaces (IVSs) more important in determining overallthoracolumbar growth?
3. Is growth of the thoracic region or growth of the lumbarregion more important in determining overall thoracolumbargrowth?
4. Which specific vertebrae drive these patterns?5. Is there also variation in centrum height between locomotor
groups?
MATERIALS AND METHODS
Specimens and radiographyThe data for this study consisted of previously radiographedvertebrae of the same individuals at two points in time (Fiorelloand German 1997; Maunz and German 1997; Lammers andGerman 2002; Farmer and German 2004). The individuals werefrom four species, O. cuniculus (n¼ 8), C. lanigera (n¼ 11),C. porcellus (n¼ 10), and M. domestica (n¼ 11). The animalswere cared for in accordance with University of Cincinnatiregulations (IACUC approval numbers 91‐05‐27‐01 and 01‐02‐26‐01). The species were classified as “half‐bounders” (rabbitand chinchilla) or “generalist quadrupeds” (guinea pig andopossum) based on behavioral data (Pridmore 1992; Lammersand German 2002). The previous work documented sexualdimorphism in some of these species, so only female C. lanigeraand C. porcellus and only male M. domestica were included.These sexes were selected to minimize body size differencesbetween the species (Lammers and German 2002). However,only five female and three male O. cuniculus were available for
Table 1. Life history information and mean mass values for sampled time points (Lammers and German 2002; Farmerand German 2004)
Species n Sex Specialist? StrategyBirth
mass (g)Weaningage (days)
Mass (g),young sample
Mass (g),adult sample
Oryctolagus cuniculus 8 M/F Yes Precocial 45 28 68 1141Chinchilla lanigera 11 F Yes Altricial 50 42–56 59 571Monodelphis domestica 10 M No Altricial 0.01 48 16 106Cavia porcellus 10 F No Precocial 85 18 92 834
Jones and German Mammalian vertebral growth 111

measurement but this should introduce little variation as thisspecies is not sexually dimorphic (Silva and Downing 1995).The first time point was collected as soon as possible after birth,herein referred to as the “young” sample. For the chinchilla andguinea pig this was within 3 days of birth. For the dwarf rabbit,this was between 6 and 8 days after birth. However, the opossumM. domestica, has extremely altricial young, which are at a veryearly developmental stage when born (Smith 2002; Keyteet al. 2008). Removing their young soon after birth would resultin either neonatal death or infanticide by the mother. Hence, thejuvenile radiograph was taken 40–60 days after birth, at whichtime this marsupial has reached a similar level of behavioraldevelopment as the placental species (Maunz andGerman 1997).The second time point was the last available radiograph, taken inall cases after full adult size was reached and referred to as the“adult” sample. For O. cuniculus and C. porcellus this wasaround 28 weeks whereas for C. lanigera and M. domestica itwas around 50 weeks. For more details on animal husbandry seeLammers and German (2002) and Farmer and German (2004).Radiographic procedure was similar for each species. Animalswere anaesthetized using Isofluorane gas, which producedunconsciousness for approximately two minutes. During thistime they were positioned on radiographic film (KodakMRM‐1)and X‐rayed using a Bennett Mammography Machine. Forfurther details on radiography procedures see the original work(Fiorello and German 1997;Maunz and German 1997; Lammersand German 2002; Farmer and German 2004). Lateral radio-graphs on film were digitized using a flat‐bed, negative scanner,and imported into Image J (Rasband 2004) for measurement.
Measurements and analysis
MeasurementsCentrum length, centrum height and IVS length were calculatedfor all thoracolumbar vertebrae using between 57 and 61landmarks, depending on the vertebral count (Fig. 1). Twolandmarks were placed on the ventral‐cranial and ventral‐caudalpoints of each centrum because they weremost clearly defined onthe radiographs and hence most repeatable. The final landmarkwas placed on the ventral‐cranial portion of the first sacral inorder to include the last IVS. These were used to calculatecentrum and IVS lengths. Two more landmarks were placed onthe ventral and dorsal side of each centrum at the point at itsminimum height. Thesewere used to calculate minimum centrumheight. The presence of ribs on the radiograph was used to notethe number of thoracic and lumbar vertebrae in each specimen.
An error study was used to test the repeatability of vertebralmeasurements. One individual of each species was selected atrandom. The landmark procedure was repeated five times andrecorded (on different days)without reference to previous attempts.Centrum length, IVS length and centrum height calculated fromeach attempt and compared. The error due to landmarking wastested using a repeated‐measures ANOVAwith age and species as
crossed factors. Repeats were not significant, except in a few cases(T1 centrum length (P¼ 0.024), disc 10 (P¼ 0.017), disc 12length (P¼ 0.011)), but the value of these differences was smallcompared to differences caused by the factors, indicatingmeasurement error due to landmarking would not influence theability to distinguish growth changes or interspecific differences.
Comparing vertebral measurements between specimens andspecies is difficult because variation in vertebral count makes itdifficult to identify vertebrae that are at a homologous positionwithin the axial column. We overcame this problem in severalways. Firstly, we compared the entire length of the columnamongst species and locomotor types. Secondly, regionallengths were calculated by summing all the centrum lengthsor IVS lengths in a particular region of the column. Thirdly,comparisons of individual vertebrae were made using an anteriorand a posterior sample. The anterior sample was the first fivethoracic vertebrae and the posterior sample was the last fivelumbar vertebrae. Despite small variations in vertebral formulathese vertebrae are likely to have similar functional roles acrossspecies because they are constrained by proximity to clearlyhomologous points, that is, the boundaries between vertebralregions, such as the last lumbar vertebra articulation with thesacrum. Fourthly, centrum height was compared by calculatingthe average height for centra from a particular region.
AnalysisAnalyses were conducted in SYSTAT and MYSTAT statisticalpackages (SYSTAT Software Inc 2007). We corrected for sizedifferences among species for individual vertebrae using bodymass, collected on the same day as the radiograph at each age foreach individual (Fiorello and German 1997; Maunz andGerman 1997; Lammers and German 2002; Farmer andGerman 2004). Scaled lengths for subsequent analyses werecalculated by dividing vertebral linear measures by the cube rootof body mass for that individual at that time‐point, whichassumes geometric similarity. Differences between locomotor
Fig. 1. Landmarks shown on juvenile O. cuniculus. Dots—landmarks. Line—thoracolumbar transition. Inset shows close‐upof landmark positions.
112 EVOLUTION & DEVELOPMENT Vol. 16, No. 2, March–April 2014
groups and through ontogeny were tested using a crossedANOVA model of the following design:
Scaled linear measure ¼ Constantþ Age ½young or adult�þGroup ½bounding or quadruped� ðnested : SpeciesÞþ Age� Group ðSpeciesÞ:
Species was nested within group, as a random variable, totake into account interspecific differences within locomotorgroups (German et al. 2008). A significant interaction term in thismodel (age� group) would indicate different growth patternsbetween the two locomotor groups. Post‐hoc Tukey’s HSD testswere performed when the interaction term was significant, toidentify exactly which combinations were significantly different.These analyses were performed on the following variables (Q isthe question from the introduction they are addressing): totalthoracolumbar column length (Q1), summed centrum length(Q2), summed IVS length (Q2), thoracic region length (Q3),lumber region length (Q3), and average centrum height in thethoracic and lumber regions (Q5). Results were displayedgraphically as line charts, with young and adult means for eachspecies connected. On these plots isometric growth would beindicated by a horizontal line between the scaled variables at thetwo time‐points. Positive allometry over the entire post‐natalgrowth period would result in an adult value, which was larger,whereas negative allometry would result in a young value whichwas larger. To address question four, examining individualvertebrae, two different sets of analyses were done for each agegroup. In the first, the model included two factors:
Young or adult only ¼ Constantþ Region ½anterior or posterior�þ Region � Groupþ Group ðnested : SpeciesÞ:
The second analysis examined the specific vertebrae withinsamples, only where there were significant within‐sampledifferences. It included the same locomotor variable, but nowa factor for vertebral number, with levels 1 through 5 for the fivemost anterior or posterior vertebrae in the sample. Species wasagain nested within locomotor type, and an interaction betweenregion and locomotor group was included in the model. Thereason for these separate analyses was that the complexity of themodel presented computational limitations.
RESULTS
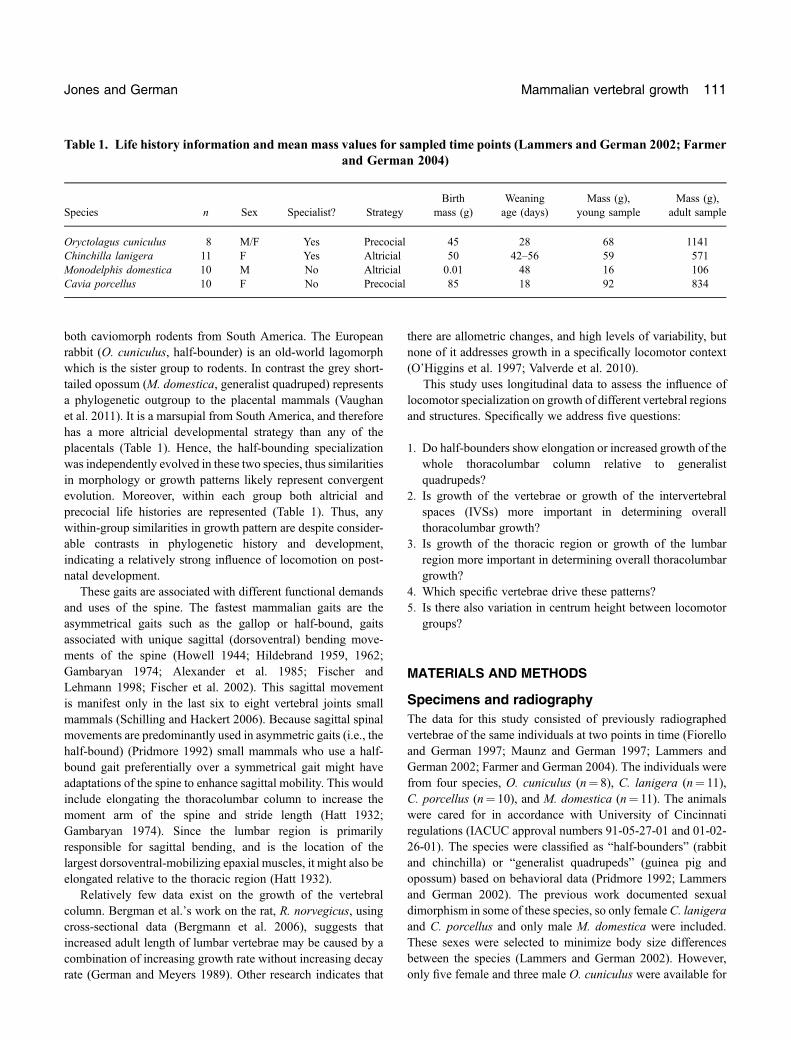
Question 1: thoracolumbar lengthThe quadrupedal species (guinea pig and opossum) exhibitedisometric growth whereas the half‐bounders (rabbit andchinchilla) grew with positive allometry. There was a significanteffect of both age and locomotor type on thoracolumbar length(Table 2), and a significant interaction between these effects.
Post‐hoc tests revealed that there was no significant differencebetween young and adult quadrupeds, indicating isometricgrowth of the column. However, there was a highly significanteffect of age on half‐bounders, suggesting their columnselongate during post‐natal growth, and reflecting the significantinteraction term in the ANOVA. Half‐bounders had significantlyshorter columns when young and significantly longer columnswhen adult than the generalists. Figure 2 shows that this patternis drivenmostly by a long adult column in the rabbitO. cuniculusand a short young column in the chinchilla C. lanigera, asreflected in the significant effect of species in the ANOVA.
Question 2: vertebrae versus intervertebralspaces
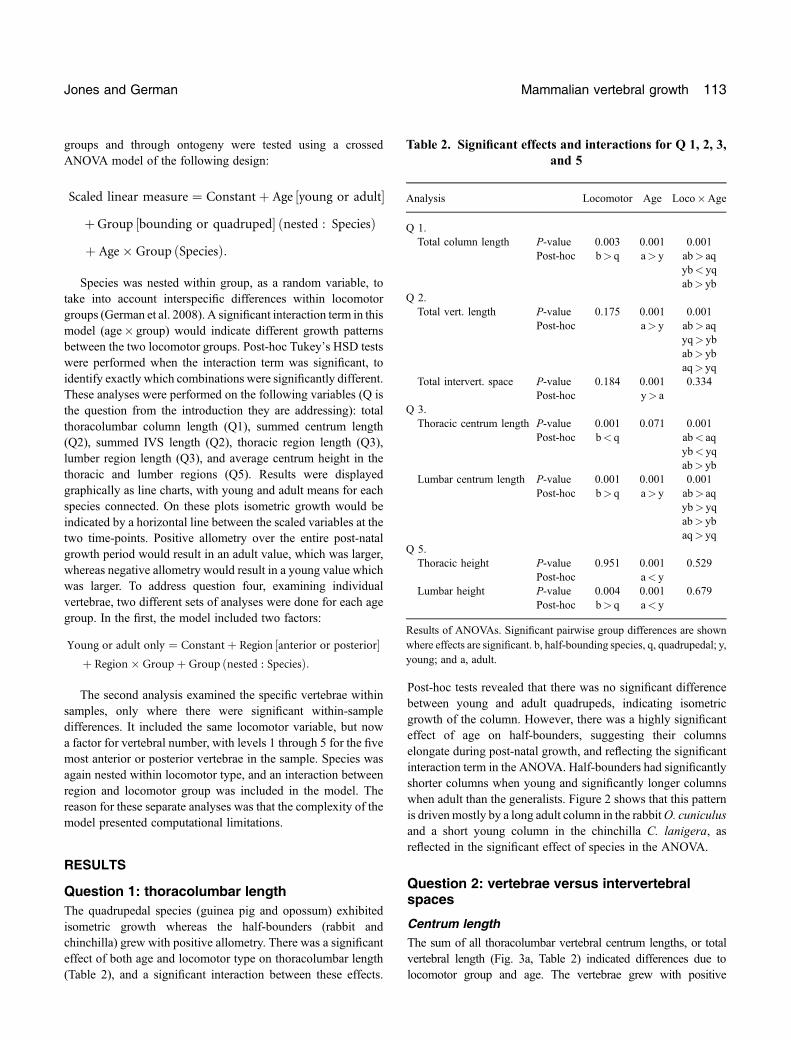
Centrum lengthThe sum of all thoracolumbar vertebral centrum lengths, or totalvertebral length (Fig. 3a, Table 2) indicated differences due tolocomotor group and age. The vertebrae grew with positive
Table 2. Significant effects and interactions for Q 1, 2, 3,and 5
Analysis Locomotor Age Loco�Age
Q 1.Total column length P‐value 0.003 0.001 0.001
Post‐hoc b> q a> y ab> aqyb< yqab> yb
Q 2.Total vert. length P‐value 0.175 0.001 0.001
Post‐hoc a> y ab> aqyq> ybab> ybaq> yq
Total intervert. space P‐value 0.184 0.001 0.334Post‐hoc y> a
Q 3.Thoracic centrum length P‐value 0.001 0.071 0.001
Post‐hoc b< q ab< aqyb< yqab> yb
Lumbar centrum length P‐value 0.001 0.001 0.001Post‐hoc b> q a> y ab> aq
yb> yqab> ybaq> yq
Q 5.Thoracic height P‐value 0.951 0.001 0.529
Post‐hoc a< yLumbar height P‐value 0.004 0.001 0.679
Post‐hoc b> q a< y
Results of ANOVAs. Significant pairwise group differences are shownwhere effects are significant. b, half‐bounding species, q, quadrupedal; y,young; and a, adult.
Jones and German Mammalian vertebral growth 113
allometry, and there was an interaction between age and locomotortype. There is no effect of locomotor group on total vertebral length,though post‐hoc tests indicate that this reflects similarity betweenthe group means (including young and adults) and masks between‐age differences. This pattern is reflected in the significant interactionterm between age and group. Post‐hoc tests reveal a very similarpattern to the whole column analysis, half‐bounders have morepost‐natal vertebral growth and shorter vertebrae when young,longer vertebrae when old, again with significant species effect.
Intervertebral spacesThe sum of all the lengths of the thoracolumbar intervertebralspaces (Fig. 3b, Table 2) exhibited a different pattern. The spacesshowed negative allometry, increasing less than body mass.ANOVAs indicate there is no significant effect of locomotor groupon the length of the IVS, nor is there a significant interaction term.However, there is an effect of age, indicating that the spacesbecome relatively smaller compared to bodymass through growth.
Question 3: Thoracic and lumbar regions
Thoracic regionThe summed lengths of the thoracic vertebrae (Fig. 4a, Table 2)grew close to isometry and so the effect of age was insignificant.There was a significant locomotion effect (P< 0.01), in whichquadrupeds tended to have longer thoracic regions than half‐bounders. There was also a significant interaction term,indicating different growth patterns among the groups. Post‐hoc tests revealed that adult half‐bounders had proportionallylonger thoracic regions than young half‐bounders whereas the
quadrupeds did not have a significant change through growth,indicating positive allometry only in the half‐bounding species.
Lumbar regionThe lumbar vertebrae had significant age and locomotion effects,as well as an interaction between them (Table 2, Fig. 4b). In allspecies the lumbar region exhibited positive allometry, becom-ing longer through post‐natal growth. However, lumbar growthwas greater in the half‐bounding group relative to the generalistgroup. Hence, the young half‐bounders had a somewhat longerlumbar region than the quadrupeds, whereas the adult half‐bounders (both species) were much longer than quadrupeds.
Question 4: Individual vertebrae
YoungThe scaled length of individual vertebrae in the young samplehad differences between the anterior and posterior regions(Fig. 5a, Table 3a; P< 0.001). In the anterior region there was a
Fig. 2. The scaled total thoracolumar column length (includingvertebrae and intervertebral spaces) for each species and time point.Bars indicate standard error. Black, half‐bounders; Grey, quad-rupeds. Species: O. cuniculus (rabbit), C. porcellus (guinea pig),C. lanigera (chinchilla) and M. domestica (opossum).
Fig. 3. (A) Scaled vertebral length and (B) scaled intervertebralspace length for each species. Species: O. cuniculus (rabbit),C. porcellus (guinea pig), C. lanigera (chinchilla) andM. domestica(opossum).
114 EVOLUTION & DEVELOPMENT Vol. 16, No. 2, March–April 2014

significant effect of locomotor group, in that the half‐bounderstended to have the shorter vertebrae (Table 3b). There was also asignificant effect of vertebral number and post‐hoc tests revealedthat T2 was shorter than T5 (Table 3c). However there was nointeraction between factors indicating the cranio‐caudal profilewas similar between groups. The results were much the same inthe posterior region with significant effects of locomotor groupand vertebral group, but no interaction. In this case vertebraeeight and nine tended to be larger than the last lumbar and six waslarger than eight (Table 3c).
AdultWithin the adults, there was also a significant difference betweenanterior and posterior regions (P< 0.001, Table 3a, Fig. 5b). Inthe anterior region there were significant effects of locomotorgroup and vertebral number (Table 3b) indicating half‐bounderstended to have shorter vertebrae and that T1, T2, and T3 wereshorter than T4 and T5. However, there was no significant
interaction. In contrast, in the posterior region all effects werehighly significant. Half‐bounders had longer posterior vertebrae(especially O. cuniculus) and all vertebrae were different exceptseven and nine (Table 3c). Post‐hoc tests revealed that the half‐bounders had significantly longer vertebrae at all positionsexcept the last lumbar, and C. lanigera had a longer penultimatelumbar vertebra than the quadruped species. The inset onFigure 5b shows the posterior region in adults expanded withbetween‐species similarities marked with boxes.
Question 5: Centrum height
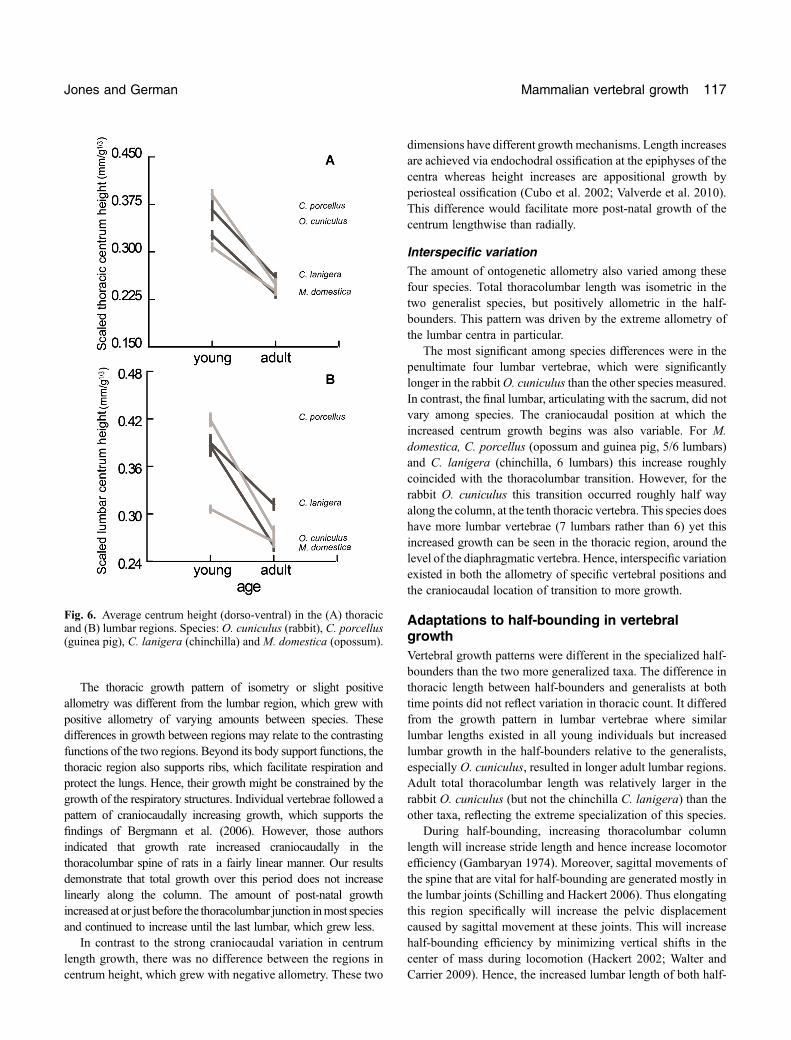
Thoracic regionThe average centrum height of a thoracic vertebra is negativelyallometric, with vertebrae becoming dorso‐ventrally shorterrelative to body mass through growth (Fig. 6a). Only age was asignificant factor (P< 0.01) for this measure; locomotor groupwas not (P¼ 0.951), with no interaction (P¼ 0.529).
Fig. 4. Summed, scaled centrum lengths from (A) the thoracic and(B) the lumbar regions. Species: O. cuniculus (rabbit), C. porcellus(guinea pig), C. lanigera (chinchilla) and M. domestica (opossum).
Fig. 5. Scaled centrum length of each individual vertebra. (A)Vertebrae are aligned with the last lumbar such that those specimenswith 19 vertebrae are not represented at the first position. The inset in(B) shows the posterior region expanded and the black boxesindicate vertebrae that were not significantly different to one another.The asterisks indicate significant differences amongst species.Species:O. cuniculus (rabbit),C. porcellus (guinea pig),C. lanigera(chinchilla) and M. domestica (opossum).
Jones and German Mammalian vertebral growth 115

Lumbar regionThe average centrum height lumbar vertebrae was alsonegatively allometric (Fig. 6b). In this case there was asignificant effect of age and locomotor group (Table 2,P< 0.01), with half‐bounders showing slightly taller centra.There was no significant interaction.
DISCUSSION
Hierarchical variation in the spineOntogenetic and interspecific variation in the thoracolumbarspine existed at multiple hierarchical levels: between columns(Q1), between structures within the column (Q2), betweenregions (Q3, Q5) and between individual vertebrae (Q4).
Ontogenetic variation
While the total thoracolumbar length was isometric or positivelyallometric, the IVSs were negatively allometric. In fact, datafrom the rat suggests that the IVS, containing the intervertebraldisk, and articular cartilage, shrinks in absolute length over thegrowth period (Bergmann et al. 2006). Our results suggest thatthe juvenile spines had a greater proportion of disk to bone thanadults. This difference between disk and bone growthmay reflectthe biomechanics the intervertebral joints, which are highlyprecise articulations and may not be amenable to significant sizechange with growth (Hukins and Meakin 2000; Urbanet al. 2000). The functioning pair of synovial joints in thisspace, the zygapophyses, may also restrict growth of the disk(Flower 1885; Slijper 1946).
Table 3. Summary of ANOVAs for Q4
Analysis
Factor
Age Region
(a)Anterior and posterior regions Young 0.001Anterior and posterior regions Adult 0.001
Analysis
Factor
Locomotor group Vertebral number Vert� loco Species
(b)Anterior young 0.001 0.001 0.596 0.002Posterior young 0.001 0.001 0.889 0.173Anterior adult 0.001 0.001 0.232 0.001Posterior adult 0.001 0.001 0.001 0.001
Post‐hoc test Locomotor group Vertebral number
(c)Anterior young b< q 2< 5Posterior young b< q 8> 10; 6> 8; 9> 10Anterior adult b< q 1< 5; 2< 4, 5; 3< 4, 5Posterior adult b> q All different except 7¼ 9
Posterior‐adult Vert� locomotion
6b> 6q P< 0.0017b> 7q P< 0.0018b> 8q P< 0.0019b> 9q P< 0.001
10b¼ 10q P¼ 0.595
(a) Difference among regions. (b) Differences within regions for a specific time‐point. (c) Post‐hoc results for significant results from 3b.
116 EVOLUTION & DEVELOPMENT Vol. 16, No. 2, March–April 2014
The thoracic growth pattern of isometry or slight positiveallometry was different from the lumbar region, which grew withpositive allometry of varying amounts between species. Thesedifferences in growth between regions may relate to the contrastingfunctions of the two regions. Beyond its body support functions, thethoracic region also supports ribs, which facilitate respiration andprotect the lungs. Hence, their growth might be constrained by thegrowth of the respiratory structures. Individual vertebrae followed apattern of craniocaudally increasing growth, which supports thefindings of Bergmann et al. (2006). However, those authorsindicated that growth rate increased craniocaudally in thethoracolumbar spine of rats in a fairly linear manner. Our resultsdemonstrate that total growth over this period does not increaselinearly along the column. The amount of post‐natal growthincreased at or just before the thoracolumbar junction inmost speciesand continued to increase until the last lumbar, which grew less.
In contrast to the strong craniocaudal variation in centrumlength growth, there was no difference between the regions incentrum height, which grew with negative allometry. These two
dimensions have different growth mechanisms. Length increasesare achieved via endochodral ossification at the epiphyses of thecentra whereas height increases are appositional growth byperiosteal ossification (Cubo et al. 2002; Valverde et al. 2010).This difference would facilitate more post‐natal growth of thecentrum lengthwise than radially.
Interspecific variationThe amount of ontogenetic allometry also varied among thesefour species. Total thoracolumbar length was isometric in thetwo generalist species, but positively allometric in the half‐bounders. This pattern was driven by the extreme allometry ofthe lumbar centra in particular.
The most significant among species differences were in thepenultimate four lumbar vertebrae, which were significantlylonger in the rabbitO. cuniculus than the other species measured.In contrast, the final lumbar, articulating with the sacrum, did notvary among species. The craniocaudal position at which theincreased centrum growth begins was also variable. For M.domestica, C. porcellus (opossum and guinea pig, 5/6 lumbars)and C. lanigera (chinchilla, 6 lumbars) this increase roughlycoincided with the thoracolumbar transition. However, for therabbit O. cuniculus this transition occurred roughly half wayalong the column, at the tenth thoracic vertebra. This species doeshave more lumbar vertebrae (7 lumbars rather than 6) yet thisincreased growth can be seen in the thoracic region, around thelevel of the diaphragmatic vertebra. Hence, interspecific variationexisted in both the allometry of specific vertebral positions andthe craniocaudal location of transition to more growth.
Adaptations to half‐bounding in vertebralgrowthVertebral growth patterns were different in the specialized half‐bounders than the two more generalized taxa. The difference inthoracic length between half‐bounders and generalists at bothtime points did not reflect variation in thoracic count. It differedfrom the growth pattern in lumbar vertebrae where similarlumbar lengths existed in all young individuals but increasedlumbar growth in the half‐bounders relative to the generalists,especially O. cuniculus, resulted in longer adult lumbar regions.Adult total thoracolumbar length was relatively larger in therabbit O. cuniculus (but not the chinchilla C. lanigera) than theother taxa, reflecting the extreme specialization of this species.
During half‐bounding, increasing thoracolumbar columnlength will increase stride length and hence increase locomotorefficiency (Gambaryan 1974). Moreover, sagittal movements ofthe spine that are vital for half‐bounding are generated mostly inthe lumbar joints (Schilling and Hackert 2006). Thus elongatingthis region specifically will increase the pelvic displacementcaused by sagittal movement at these joints. This will increasehalf‐bounding efficiency by minimizing vertical shifts in thecenter of mass during locomotion (Hackert 2002; Walter andCarrier 2009). Hence, the increased lumbar length of both half‐
Fig. 6. Average centrum height (dorso‐ventral) in the (A) thoracicand (B) lumbar regions. Species: O. cuniculus (rabbit), C. porcellus(guinea pig), C. lanigera (chinchilla) and M. domestica (opossum).
Jones and German Mammalian vertebral growth 117

bounders meets functional adaptive predictions. Kinematic dataon the limbs suggest that in small mammals young use verysimilar locomotor patterns to adults (Schilling 2005). Indeed,these species used the half‐bound from a very young age(Lammers and German 2002). Therefore, the strong ontogeneticallometry of the spine through growth likely does not reflect ashift in the types of gait used, but an increase in the efficiency oftheir use. This hypothesis could be tested by examiningkinematic data through ontogeny in these species.
There were also growth differences within each of thelocomotor groups. Most notably, elongation of the thoracolum-bar spine is much greater in O. cuniculus (rabbit) thanC. lanigera (chinchilla), which grouped more closely to thegeneralist taxa. This interspecific variation may be related tophylogeny, degree of specialization or size. O. cuniculus is amember of the family Leporidae in the order Lagomorpha, inwhich many species are highly specialized cursors, using thebound or half‐bound. In contrast, C. lanigera is a member of thefamily Chinchillidae in the order Rodentia which, though stillconsidered cursorially adapted, are less so than the rabbits(Vaughan et al. 2011).
Secondly, theOryctolagus individuals used in this study werepeadomorphic dwarfs but they are still larger than the otherspecies measured (Fiorello and German 1997, Table 1). Theenergy demands associated with bounding behavior increasewith size (Gambaryan 1974). Hence, larger mammals may showgreater specialization to increase locomotor efficiency. Finally,these time points spanned a larger change in absolute mass inO. cuniculus relative to C. lanigera, which may permit morevertebral growth, depending on the exact growth curve for thisspecies. However, this difference is unlikely to be driving ourresults because if the young sample ofO. cuniculuswere taken atan earlier developmental stage relative to the other taxon, asmight be suggested by the smaller mass of the young relative tothe adult (Table 1), then one would predict that their youngwould also have consistently smaller vertebrae. However this isnot the case. The young rabbits more often had larger vertebraethan the young chinchillas, not smaller. Instead, it is therelatively large size of the adult vertebrae that is driving theincreased growth, which was sampled well past the time eachanimal reached final adult size.
Implications for evolution of the vertebralcolumnVariation in the growth of vertebrae among species may providea heterochronic mechanism for producing differences inthe thoracolumbar column associated with locomotor type.The lumbar vertebrae which grew most were also most variablebetween species, and lumbar growth was exaggerated in the half‐bounding taxa, especially O. cuniculus. Moreover, the lumbarvertebra that grew least (the last lumbar) also showed leastinterspecific variation. Therefore, this ontogenetic allometry
may provide a source of variation for natural selection (e.g., dueto locomotor demands) to act upon. In contrast, the thoracicregion grew close to isometry and there was little interspecificvariation, which may be due to constraints imposed by theirarticulation with the ribs and consequently, other ventralstructures. Hence, evolution may influence growth differentlyin different vertebral regions.
Both vertebral count and centrum length are vital fordetermining thoracolumbar proportions (Rose 1975;Shapiro 1995; Polly et al. 2001; Narita and Kuratani 2005;Buchholtz 2007). We have shown that the growth pattern of aparticular vertebra is also a function of its position within thevertebral column. Therefore, the interaction of vertebral numberand vertebral position (i.e., the number of vertebrae in eachregion) is important in determining growth patterns. Thesefactors have independent developmental origins and geneticcontrols (Wellik 2007, 2009; Schoenwolf et al. 2008). Variationsin vertebral count (also known as meristic changes) occur whenthe somites are divided into discrete units during somitogenesis,a process controlled bymolecular oscillators (Pourquie 2003a, b;Sanger and Gibson‐Brown 2004). Conversely, variations in theposition of regional transitions along the column (homeoticchanges) are determined by expression of regionalizing genes, ofwhich the Hox genes are most well known (Wellik 2009). Theseboundaries are sometimes determined before the division of theparaxial mesoderm into somites (Carapuco et al. 2005). Finally,the morphology of segments within a particular region canchange and diversify from each other, which is known asskeletogenic variation (Polly et al. 2001).
This study illustrates the interaction of these mechanisms. Forexample, the rabbit O. cuniculus had the longest thoracolumbarcolumn of the species sampled. This extra growth was achieved byseveral means. There was skeletogenic variation, as each existinglumbar grew more than the other species, except for the last lumbar,which seems constrained. In addition, the rabbit had an extra seventhlumbar vertebra with faster lumbar‐type growth. This extra lumbarwas not noticeable in the young centrum length profile (Fig. 5a) butbecame longer than an equivalent thoracic by the time adult size wasreached (Fig. 5b). This was likely a homeotic change because therabbit also had one fewer thoracic vertebra. To truly determine themechanism (homeotic vs. meristic) behind the seventh lumbar thevertebral identities would have to be identified relative to theirsomitic level and compared to the ancestral state. Finally, theposterior thoracic vertebrae in this species also show a more“lumbar‐like” growth pattern, which is potentially a skeletogeneticor a homeotic change. Thus multiple types of mechanisms may acttogether to produce variations in adult vertebral proportions. Theseresults emphasize that variations in post‐natal growth can beimportant in generating variation in thoracolumbar length, with littlevariation in vertebral count, and that this variation tends to occur inthe posterior portion of the column.
The craniocaudal variation in vertebral growth documentedhere correlates well with expression patterns of Hox genes
118 EVOLUTION & DEVELOPMENT Vol. 16, No. 2, March–April 2014

established in mice (Schoenwolf et al. 2008). In particular, Hox‐10 paralogues suppress rib expression and pattern thethoracolumbar transition (Wellik and Capecchi 2003;Wellik 2009). This regional transition was marked by a strongincrease in vertebral growth in three of the four species,supporting data from the rat indicating a link between theHox‐10domain and post‐natal growth (Bergmann et al. 2006). Thelumbosacral transition is thought to be determined by the anteriorexpression ofHox‐11 (Wellik and Capecchi 2003). These resultsshow drastically less ontogenetic and interspecific variation inthe length of the last lumbar relative to the others. Doesproximity to the lumbosacral transition and Hox‐11 expressionreduce centrum growth in this vertebra?Whether the influence ofthese regionalizing genes on post‐natal growth is direct or via themechanical influences created by surrounding tissues remainsunclear. Mechanical loading environment (compression vs.distraction/tension) does influences the growth of caudalvertebrae in experimental rats (Stokes et al. 1996). Thereforeregional variation in the other axial structures may also indirectlycontrol craniocaudal centrum growth patterns.
CONCLUSION
This study demonstrated that ontogenetic allometry differs amongthe structures and regions of the thoracolumbar spine. Weidentified the lumbar centra (with the exception of the last lumbar)as the greatest source of ontogenetic allometry, and also the greatestsource of interspecific variation in these species. Speciesspecialized for half‐bounding, especially the rabbit O. cuniculus,had elongated lumbar regions via increased post‐natal growth oflumbar vertebrae. This is in agreement with biomechanicalpredictions that increased lumbar length will increase half‐bounding efficiency. In contrast, the thoracic region grew closerto isometry and varied less among species. Therefore, locomotorspecialization was associated with different post‐natal growthpatterns in vertebrae from different vertebral regions. Further,heterochrony of lumbar epiphyseal growth may be important forgenerating variation and increasing evolvability of this region.Future work should characterize the nature of the heterochroniesresulting in the differences between these locomotor groups andtest for allometry in locomotor performance.
AcknowledgmentsWewould like to thank Andy Lammers for providing bodymass data forthe animals and original X‐rays and Chris Ruff for use of his flatbedscanner. Valerie DeLeon had many insightful thoughts and commentsduring the design and implementation of this study. We would also liketo thank Karen Sears and one anonymous reviewer for their helpfulcomments on the manuscript.
REFERENCES
Alberch, P., Gould, S. J., Oster, G. F., andWake, D. B. 1979. Size and shape inontogeny and phylogeny. Paleobiology 5: 296–317.
Alexander, R. M., Dimery, N. J., and Ker, R. F. 1985. Elastic structures in theback and their role in galloping in some mammals. J. Zool. 207: 467–482.
Bastir, M., and Rosas, A. 2004a. Comparative ontogeny in humans andchimpanzees: similarities, differences and paradoxes in postnatal growthand development of the skull. Ann. Anat. 186: 503–509.
Bastir, M., and Rosas, A. 2004b. Facial heights: implications of postnatalontogeny and facial orientation for skull morphology in humans andchimpanzees. Am. J. Phys. Anthropol 47(5): 60–61.
Bergmann, P. J., Melin, A. D., and Russell, A. P. 2006. Differential segmentalgrowth of the vertebral column of the rat (Rattus norvegicus). Zoology109: 54–65.
Buchholtz, E. A. 2007. Modular evolution of the cetacean vertebral column.Evol. Dev. 9: 278–289.
Carapuco, M., Novoa, A., Bobola, N., and Mallo, M. 2005. Hox genesspecify vertebral types in the presomitic mesoderm.Genes Dev. 19: 2116–2121.
Cubo, J., Azagra, D., Casinos, A., and Castanet, J. 2002. Heterochronicdetection through a function for the ontogenetic variation of bone shape. J.Theor. Biol. 215: 57–66.
Drake, A. G. 2011. Dispelling dog dogma: an investigation of heterochronyin dogs using 3D geometric morphometric analysis of skull shape. Evol.Dev. 13: 204–213.
Farmer, M. A., and German, R. Z. 2004. Sexual dimorphism in thecraniofacial growth of the guinea pig (Cavia porcellus). J. Morphol. 259:172–181.
Fiorello, C. V., and German, R. Z. 1997. Heterochrony within species:craniofacial growth in giant, standard, and dwarf rabbits. Evolution 51:250–261.
Fischer,M. S., and Lehmann, R. 1998.Application of cineradiography for themetric and kinematic study of in‐phase gaits during locomotion of the pika(Ochotona rufescens, Mammalia: Lagomorpha). Zool. Anal. ComplexSyst. 101: 148–173.
Fischer, M. S., Schilling, N., Schmidt, M., Haarhaus, D., andWitte, H. 2002.Basic limb kinematics of small therian mammals. J. Exp. Biol. 205: 1315–1338.
Flower, W. H. 1885. An Introduction to the Osteology of the Mammalia.Macmillan, London.
Gambaryan, P. P. 1974. How Mammals Run. Halsted Press, New York.German, R. Z., Crompton, A.W., and Thexton, A. J. 2008. Variation in EMG
activity: a hierarchical approach. Integr. Comp. Biol. 48: 283–293.German, R. Z., and Meyers, L. L. 1989. The role of time and size in
ontogenetic allometry 1: review. Growth Dev. Aging 53: 101–106.Godfrey, L. R., and Sutherland, M. R. 1996. Paradox of peramorphic
paedomorphosis: heterochrony and human evolution. Am. J. Phys.Anthropol. 99: 17–42.
Gould, S. J. 1977. Ontogeny and Phylogeny. Harvard University Press,Cambridge, MA.
Hackert, R. 2002. Aspects of Sagittal Bending of the Spine in SmallMammals. PhD Thesis, Biological‐Pharmaceutical Faculty, Friedrich‐Schiller‐University of Jena, Jena, Germany.
Hatt, R. T. 1932. The vertebral columns of ricochetal rodents. Bull. Am. Mus.Nat. Hist. 63: 599–738.
Hildebrand, M. 1959. Motions of the running cheetah and horse. J. Mammal.40: 481–495.
Hildebrand, M. 1962.Walking, running, and jumping.Am. Zool. 2: 151–155.Howell, A. B. 1944. Speed in Mammals. University of Chicago Press,
Chicago.Hukins, D.W. L., andMeakin, J. R. 2000. Relationship between structure and
mechanical function of the tissues of the intervertebral joint. Am. Zool. 40:42–52.
Jungers,W. L., Cole, T.M., and Owsley, D.W. 1988.Multivariate analysis ofrelative growth in the limb bones of Arikara Indians. Growth Dev. Aging52: 103–107.
Keyte, A. L., Imam, T., Alonzo, M., Halbert, T., and Smith, K. K. 2008.Building a marsupial neonate: evolution of the limb development programin opossum. Dev. Biol. 319: 498–498.
Lammers, A. R., and German, R. Z. 2002. Ontogenetic allometry in thelocomotor skeleton of specialized half‐bounding mammals. J. Zool. Soc.258: 485–495.
Jones and German Mammalian vertebral growth 119

Maunz, M., and German, R. Z. 1997. Ontogeny and limb bone scaling in twonew world marsupials,Monodelphis domestica and Didelphis virginiana.J. Morphol. 231: 117–130.
McKinney, M. L., and Schoch, R. M. 1985. Titanothere allometry,heterochrony, and biomechanics: revising an evolutionary classic.Evolution 39: 1352–1363.
Minelli, A., and Peruffo, B. 1991. Developmental pathways, homology andhomonomy in metameric animals. J. Evol. Biol. 4: 429–445.
Narita, Y., and Kuratani, S. 2005. Evolution of the vertebral formulae inmammals: a perspective on developmental constraints. J. Exp. Zool. BMol. Dev. Evol. 304B: 91–106.
O’Higgins, P., Milne, N., Johnson, D. R., Runnion, C. K., and Oxnard, C. E.1997. Adaptation in the vertebral column: a comparative study of patternsof metameric variation in mice and men. J. Anat. 190: 105–113.
Polly, P. D., Head, J. J., and Cohn, M. J. 2001. Testing modularity anddissociation: the evolution of regional proportions in snakes. In M.Zelditch (ed.). Beyond Heterochrony: The Evolution of Development.Wiley‐Liss, New York.
Pourquié, O. 2003a. A biochemical oscillator linked to vertebratesegmentation. In G. B. Müller and and S. A. Newman (eds.). Originationof Organismal Form: Beyond the Gene in Developmental andEvolutionary Biology. MIT Press, Cambridge, MA. pp. 183–194.
Pourquié, O. 2003b. The segmentation clock: converting embryonic timeinto spatial pattern. Science 301: 328–330.
Pridmore, P. A. 1992. Trunk movements during locomotion in the marsupialMonodelphis domestica (Didelphidae). J. Morphol. 211: 137–146.
Rasband, W. S. 2004. ImageJ. National Institutes of Health, Bethesda, MD.Ravosa, M. J. 1992. Allometry and heterochrony in extant and extinct
malagasy primates. J. Hum. Evol. 23: 197–217.Rocha‐Barbosa, O., Loguercio, M. F. D., Renous, S., and Gasc, J. P. 2005.
Limb joints kinematics and their relation to increasing speed in the guineapig Cavia porcellus (Mammalia: Rodentia). J. Zool. 266: 293–305.
Rose,M.D. 1975. Functional proportions of primate lumbar vertebral bodies.J. Hum. Evol. 4: 21–38.
Sanger, T. J., and Gibson‐Brown, J. J. 2004. The developmental bases of limbreduction and body elongation in squamates. Evolution 58: 2103–2106.
Schilling, N. 2005. Ontogenetic development of locomotion in smallmammals—a kinematic study. J. Exp. Biol. 208: 4013–4034.
Schilling, N., and Hackert, R. 2006. Sagittal spine movements of small therianmammals during asymmetrical gaits. J. Exp. Biol. 209: 3925–3939.
Schilling, N., and Petrovitch, A. 2006. Postnatal allometry of the skeleton inTupaia glis (Scandentia: Tupaiidae) and Galea musteloides (Rodentia:Caviidae): a test of the three‐segment limb hypothesis. Zoology 109: 148–163.
Schoenwolf, G. C., Bleyl, S. B., Brauer, P. R., and Francis‐West, P. H. 2008.Larsen’s Human Embryology. 4th Ed. Churchill‐Livingstone, Philadel-phia, PA.
Shapiro, L. 1995. Functional morphology of idrid lumbar vertebrae. Am. J.Phys. Anthropol. 98: 323–342.
Shea, B. T. 1981. Relative Growth of the Limbs and Trunk in the AfricanApes. Am. J. Phys. Anthropol. 56: 179–201.
Shea, B. T. 1983. Allometry and heterochrony in the African apes. Am. J.Phys. Anthropol. 62: 275–289.
Shea, B. T. 1989. Heterochrony in human evolution: the case for neotenyreconsidered. Yearb. Phys. Anthropol. 32: 69–101.
Silva, M., and Downing, J. A. 1995. CRC Handbook of Mammalian BodyMasses. CRC Press, Boca Raton.
Slijper, E. J. 1946. Comparative biologic‐anatomical investigations on thevertebral column and spinal musculature of mammals. Verh. Kon. Ned.Akad. Wet. 42: 1–128.
Smith, K. K. 2002. Sequence heterochrony and the evolution ofdevelopment. J. Morphol. 252: 82–97.
Stokes, I. A. F., Spence, H., Aronsson, D. D., and Kilmer, N. 1996.Mechanical modulation of vertebral body growth—implications forscoliosis progression. Spine 21: 1162–1167.
SYSTAT Software, Inc. 2007. MYSTAT 12.Urban, J. P. G., Roberts, S., and Ralphs, J. R. 2000. The nucleus of the
intervertebral disc from development to degeneration. Am. Zool. 40: 53–61.
Valverde, E., Galvez‐Lopez, E., Alba‐Fernandez, C., Del Rio, L., andCasinos, A. 2010. Lumbar ontogenetic growth and sexual dimorphism inmodern humans. Am. J. Hum. Biol. 22: 596–603.
Vaughan, T. A., Ryan, J. M., and Czaplewski, N. J. 2011. Mammalogy. 5thEd. Jones and Bartlett Publishers, PLC, Sudbury, MA.
Walter, R. M., and Carrier, D. R. 2009. Rapid acceleration in dogs:ground forces and body posture dynamics. J. Exp. Biol. 212: 1930–1939.
Wayne, R. K. 1986. Cranial morphology of domestic and wild canids: theinfluence of development on morphological change. Evolution 40: 243–261.
Wayne, R. K. 1986. Limb morphology of domestic and wild canids: theinfluence of development on morphological change. J. Morphol. 187:301–319.
Wellik, D. M. 2007. Hox patterning of the vertebrate axial skeleton. Dev.Dynam. 236: 2454–2463.
Wellik, D. M. 2009. Hox genes and vertebrate axial pattern. Curr. Top. Dev.Biol. 88: 257–278.
Wellik, D. M., and Capecchi, M. R. 2003. Hox10 and Hox11 genes arerequired to globally pattern the mammalian skeleton. Science 301: 363–367.
Weston, E. M. 2003. Evolution of ontogeny in the hippopotamus skull: usingallometry to dissect developmental change. Biol. J. Linn. Soc. 80: 625–638.
120 EVOLUTION & DEVELOPMENT Vol. 16, No. 2, March–April 2014
Copyright © 2022 FDOKUMEN




















